
Submitted:
29 January 2024
Posted:
30 January 2024
You are already at the latest version
Abstract
Daptomycin is one of the last therapeutic resources for multidrug-resistant gram- positive bacteria. Despite its structural similarities with glycopeptides, its mechanisms of action and resistance are different, and in some respects are not completely understood. Mutations in several genes have been associated to daptomycin resistance, especially in mprF, walkR, rpoB and rpoC, but their role and importance remain to be elucidated. We have studied mutations in 11 genes, which have been previously associated to daptomycin non- susceptibility, in 9 daptomycin non-susceptible Staphylococcus aureus clinical isolates (daptomycin MIC: > 1mg/L). Susceptibility to daptomycin, vancomycin, linezolid, oxacillin, telavancin and dalbavancin were studied. walkR, agrA, cls1, cls2, fakA, pnpA, clpP, prs, rpoB, rpoC and mprF were amplified by PCR and sequenced. The sequences were compared with the S. aureus ATCC 25923 complete genome (GenBank gi: 685631213) by using BLAST® software. We did not find any changes in walkR, pnpA, prs and clpP. All isolates excepting isolate 5 showed a high number of significant mutations (between 13 and 25 amino acid changes) in mprF. Most isolates also showed mutations in rpo genes, cls genes and fakA. Daptomycin non-susceptibility in S. aureus clinical isolates seems to be reached through different mutations combinations when compared to S. aureus ATCC 25293. Especially mprF and cls1 showed a very high polymorphism in most isolates. Meanwhile, one isolate, St5, showed only single mutation in mprF (P314T).
Keywords:
daptomicina
; mutaciones
; resistencia
; Staphylococcus aureus
1. Introduction
Staphylococcus aureus has shown a high capacity for developing resistance to each newer anti- staphylococcal antibiotic developed. Antibiotic resistance in S. aureus results in higher morbidity, mortality, length of hospitalization and health expenditure [1]. The cyclic anionic lipopeptide antibiotic daptomycin (DAP) [2] is becoming a main resource in therapy against multidrug-resistant S. aureus, especially methicillin-resistant S. aureus (MRSA), due to its activity against glycopeptide and linezolid-resistant MRSA [3,4]. The mechanism of action of DAP differs from conventional glycopeptides (vancomycin, teicoplanin), and is more like cationic antimicrobial peptides produced by the immune system. Vancomycin and teicoplanin impair the bacterial cell wall synthesis. They bind to the terminal dipeptide D-alanyl-D-alanine (D-Ala-D-Ala), within the disaccharide pentapeptide attached to lipid II and the forming PGN chain. This way, transglycosylation and transpeptidation are prevented, thus hindering proper wall synthesis [5,6]. In addition, vancomycin has other secondary actions, probably less important in the lysis of the microorganism, such as inhibition of RNA synthesis and alteration of wall permeability.
The action of DAP is based on binding to the bacterial cell membrane, through its hydrophobic end, in the presence of calcium ions [7,8,9]. This binding occurs both in exponential growth phase and stationary phase, causing membrane depolarization due to the loss of potassium ions from the cytoplasm [10]. This process leads to the disruption of multiple functions of the bacterial cell membrane, such as synthesis of proteins, DNA and RNA without the need for the antimicrobial to penetrate the cytoplasm. Unlike other antimicrobials, the mechanism of action does not involve lysis of bacterial cells [11] but rather bacterial cell death due to alterations in cellular homeostasis [7,12].
Intrinsic resistance to DAP by hydrolytic cleavage of ester bonds inside the DAP molecule has been described, in absence of contact with this or other antibiotics, among low percentage of GC gram-positive bacteria, such as staphylococci [13].
Streptomyces strains highly resistant to daptomycin (MIC≥256 µg/ml) have been found to inactivate the antibiotic through two primary modes of daptomycin inactivation: ring hydrolysis, leading to linearization of the cyclic compound and deacylation of the lipid tail [14]. Additionally, Paenibacillus lautus has been shown to inactivate daptomycin. The production of the inactivating activity in P. lautus was inducible by exposure to daptomycin and some data suggest might be caused by metalloesterases (14).
This “natural” resistance mechanism has been suggested to be associated with natural production of DAP-like molecules by different microorganisms. Though this type of resistance has not been shown in clinically important bacteria yet, resistance genes coding for these DAP inactivating enzymes might be a potential source of resistance if they could be captured by pathogenic bacteria, as has happened before for other resistance determinants, such as CTX-M enzymes, originated in Kluyvera spp. and now present in most clinically important enterobacteria [15].
Nevertheless, DAP-non susceptibility observed in isolates obtained both in vitro and from therapeutic failures in S. aureus, seems to be mainly associated with electrostatic repulsion of the DAP–calcium complex from the cell surface, due to an increase in the positive charge of the bacterial surface [16].
DAP-non susceptible isolates obtained both in vitro and from therapeutic failures, usually harbor mutations in genes associated with cytoplasmic membrane (mprF) and cell wall homeostasis (yycFG, also known as walK), and mutations in RNA polymerase subunits rpoB/rpoC [7,8]. DAP-non susceptible isolates have been described, both in vitro and in vivo, lacking mutations in some of these genes, but not in all of them [7,9,10,11]. Other authors have shown that DAP-non susceptibility may also involve the upregulation of genes associated with cell wall synthesis and turnover, such as the two-component regulator vraSR [12]. Otherwise, whole genome sequencing studies have shown that DAP-non susceptible isolates may harbor mutations in other genes, such as agrA, pnpA, pgsA, prs, cls2 and clpP, some of them associated with the cytoplasmic membrane and cell wall metabolism [10,11].
Therefore, the present study had the following aims: identification of S. aureus strains (MRSA and MSSA) non-susceptible to DAP isolated from 3 different hospitals in Spain; test sensitivity to different drugs such as oxacillin, vancomycin, linezolid, dalbavancin and telavancin; amplification and sequencing of the main genes associated with DAP resistance.
2. Materials and Methods
2.1. Selection of bacterial strains
Nine S. aureus clinical isolates identified as DAP non-susceptible S. aureus (MIC >1 mg/L) by microdilution conventional susceptibility systems (WalkAway, Perkin Elmer, USA) were included in this study. Seven of them had been isolated in hospitals in the province of Badajoz (Southwest of Spain), and two in the Hospital Universitario de Salamanca (Middle West of Spain). All isolates except MSa9 were from patients who had received daptomycin treatment, although data on duration of treatment and dosage were not accessible. To our knowledge, the patient from whom the MSa9 isolate was obtained did not receive treatment with daptomycin at any time.
Microorganisms were identified by MALDI-TOF mass spectrometry (MicroflexTM, Bruker Daltonics, Germany). All the isolates used were preserved and stored at -80°C in glycerol broth with 10% skimmed milk.
2.2. Antibiotic susceptibility
Susceptibility to DAP, vancomycin (VAN), linezolid (LZD) and oxacillin (OXA) were tested by using a broth microdilution method (MicroScan WalkAway, Perkin Elmer, USA), according manufacturer instructions. Telavancin (TLV) and dalbavancin (DLV) were studied by E-test (Liofilchem, Italy). The microdilution sensitivity study was carried out using an automated commercial method (MicroScan WalkA-way, Perkin Elmer, USA), in which the antimicrobials are lyophilised so that a predetermined concentration is reached by adding the set amount of culture medium. The microorganism is inoculated in cation-adjusted Mueller Hinton broth, supplemented with 2% NaCl for oxacillin sensitivity and 50 microg/ml calcium for DAP. An inoculum of the microorganism equivalent to 0.5 of the McFarland (McF) standard is inoculated and incubated at 37°C for 20-24 hours. The reading is done in an automated way in a MicroScan equipment.
For the E-test study, a Mueller Hinton agar plate was inoculated with an inoculum equivalent to 0.5 McF, strips of the different antimicrobials are deposited on the inoculated agar, incubated at 37°C for 24 hours and read visually. For the DAP E-tests it is not necessary to supplement the agar with Ca2+, as the DAP strips have it incorporated.
Methicillin resistance, in strains with e-test sensitive to oxacillin, was verified by PCR specific for mecA and mecC genes (the primers used appear in Table 1). The isolates were subjected to the E-test according to the manufacturer's instructions. Susceptibility criteria for VAN, LZD, DLV, TLV and OXA were those proposed by the CLSI (VAN: < 2 mg/L, susceptible; 4-8 mg/L, intermediate; > 16 mg/L, resistant. LZD: < 4 mg/L, susceptible; > 8 mg/L, resistant. DLV: < 0.25 mg/L, susceptible; intermediate and resistant categories not defined. TLV: < 0.12 mg/L, susceptible; intermediate and resistant categories not defined. OXA: < 0.5 mg/L, susceptible; > 1 mg/L, resistant) [13]. The only categories considered for DAP were "susceptible" and "non-susceptible", as CLSI reports only the sensitivity criterion (MIC < 1 mg/L).
2.3. DNA extraction and amplification of genes associated with DAP non-susceptibility in S. aureus
A suspension with turbidity equivalent to 1 McF was obtained from colonies grown for 24 hours, at 37 ºC on blood agar. Then, 500 μl of the suspension were transferred to the automated nucleic acid extraction system NucliSENS® easyMAG (bioMérieux, France). Nucleic acids extraction was performed according to manufacturer instructions. DNA was preserved at -80°C.A series of genes previously associated with DAP non-susceptibility [7,14] were amplified and sequenced. Amplification was carried out in an Eppendorf Master Cycler® thermal cycler (Eppendorf, Germany). All primers used in the study were designed specifically (Table 1), based on the S. aureus ATCC 25923 complete genome sequence deposited in GenBank (gi: 685631213) using the Primer Blast® application (NCBI, USA). The primers were manufactured by Sigma-Genosys (USA), and the MasterMix® PCR amplification mix (Promega, USA9, a premixed 2X solution of Taq DNA Polymerase, dNTPs and reaction buffer, was used for PCR reaction. PCR conditions were as follows: 95°C x 5 minutes (denaturation); 35 cycles of: 95°C x 30 seconds, 1 minute at the temperature reported in supplementary section (Table 1S) specific for each primer (hybridization), 72°C x 1 minute (elongation); and finally, 72°C x 5 minutes (elongation).
2.4. Visualization of amplicons and purification of PCR product
The products were subjected to electrophoresis in 2% agarose gel together with a size control (DNA Ladder 100 bp, Promega, USA). Blue/Orange 6x Loading Dye (Promega, USA) was used as loading buffer, and RedSafe® Nucleic Acid Staining Solution (Invitrogen, USA) was used as a fluorescent dye for DNA detection.
As a general rule, amplified DNA was cleaned for sequencing by using Exo-SAP It® (Chang Biosciences, China). When DNA was purified from the gel, the MEGAquick-Spin Total Fragment DNA Purification kit (Intron Biotechnology, South Korea) was used.
Amplicons were sequenced by the Sanger method in a 3500 Genetic Analyzer sequencer (Applied Biosystems, USA) and visualized by using the software Chromas 2.5.1.0. Sequences were compared with the S. aureus ATCC 25923 complete genome sequence deposited in GenBank (gi: 685631213). S. aureus ATCC 25923 is commonly used as a control in standard laboratory tests. Comparison and analysis were carried out by using the BLAST® software, freely available in the NIH website.
3. Results
3.1. Antibiotic susceptibility
DAP MICs ranged between 1 - 4 mg/L (Table 1). All the isolates were susceptible to VAN, LZD, TLV and DLV (Table 1). The MIC of DLV for MSa3 isolate was higher (0.1 mg/L), though still into the susceptible range (break point: 0.25 mg/L). Three out of 9 isolates (33.3%) were phenotypically MRSA (OXA MIC ≥ 4 mg/L), and 4 MSSA (44.44%) (OXA MIC < 2 mg/L). Two isolates (MSa4 and MSa7) were OS-MRSA (OXA MICs=2 mg/L, PCR positive for mecA) (primers on supplementary section Table S1).
3.2. Mutations profiles associated with non-susceptibility
All the isolates, excepting MSa5, showed significant mutations in the genes rpoB, mprF, rpoC, cls1, cls2, and fakA, while MSa5 showed significant mutations only in rpoB and mprF genes (Figure 1) (Table 2, Table 3 and Table 4). We did not detect any significant mutations in walk, agrA, pnpA, clpP and prS genes. The genes with the highest number of mutations were mprF (31-35 amino acid changes) and cls1 (6-16 amino acid changes) (Figure 1).
mprF gene presented a high number of mutations in all isolates (Table 2), excepting MSa5, which only had one mutation in this region, P314T. Mutations on this position also appeared in other clinical isolates, such as MSa1 (P314T) and MSa6 (P314L). Forty-three different mutations were identified, in all, in mprF genes. Most isolates showed different combinations of these mutations. Most changes appeared in the central area of the gene, although we find changes throughout the gene. MSa6 had the highest number of mutations in mprF, exactly 35, followed by strains MSa1, MSa4 and MSa8 with 34 changes.
As in mprF, we found a high number of changes in the genes encoding for the cardiolipin synthase, especially in cls1. Only MSa5 showed no mutations in cls1 (Table 3). Though a number of mutations appeared in more than one isolate, no isolates showed the same mutations profile. Only one mutation (I238V) appeared in all the isolates showing mutations in this gene. MSa6 was the isolate with the highest number of mutations in this gene (16 mutations). cls2 only showed 4 different mutations (Table 3), half of which were shared by all the isolates, excepting MSa5, which did not show any mutation in this gene.
All the isolates, including MSa5, showed mutations in rpoB, however, all the isolates excepting MSa5 had mutations in rpoC (Table 4).
fakA is a protein also referred to as mw1109 in previous studies a fatty acid kinase that has been associated with unsensitivity to DAP in mutants selected in vitro [10]. All the isolates tested in this study, excepting the clinical isolate MSa5, showed mutations in this gene (Table 4). Mutation profiles in fakA were more homogeneous than in mprF and cls1. Only MSa6 showed a mutations profile neatly different.
We did not identify changes in walkR in any of our 9 clinical isolates. Other genes also associated occasionally to reduced susceptibility to DAP, such as agrA, clpP, pnpA and prs, did not show any significant mutations in this study.
In summary, we found two different mutation profiles. A group of 8 isolates had changes in 6 genes (mprF, rpoB, rpoC, cls1, cls2 and fakA), while one isolate had changes in only 2 genes (mprF and rpoB). mprF and cls1 showed a high number of mutations/isolate in most isolates, while for the other genes the number of mutations/isolate was much shorter. In the absence of the MLST analysis, we could not rule out that the identified substitutions could potentially be associated with a specific genetic lineage rather than daptomycin resistance.
4. Discussion
DAP resistance seems to be based on complex mechanisms and has been associated with the presence of mutations in different genes, most of them involved in the metabolism and homeostasis of the cell membrane, and, in some cases, of the bacterial wall [7,14,15]. A cause-effect relationship between specific mutations in specific genes and a specific increase in the MICs of DAP has hardly been described, and the repulsion hypothesis does not explain resistance to DAP in all S. aureus isolates. Only some studies have shown that the replacement of the mutated mprF gene with a wild one reverses, at least partially, the MIC of DAP [16,17]. In the present study, the MIC interval of DAP ranged between 2 and 4 mg/L in all cases, except MSa2, MIC of 1 mg/L. Although non-sensitivity to daptomycin in S. aureus is MIC > 1 mg/L, due to its proximity to the range, MSa2 was not ruled out from the study.
Mutations in rpoB and mprF genes have been reported in most staphylococcal clinical isolates showing DAP MICs increase [7,10,11]. mprF codes for the bifunctional enzyme MprF, that contributes to the positive charge of the cell membrane surface trough the lysinylation of phosphatidylglycerol (PG) and the translocation of lysinylated PG (L-PG) from the inner to the outer leaflet of the cell membrane. The positively charged cell membrane surface resulting from this process, helps repel the DAP molecule from the surface [18,19,20]. The association between mprF mutations and DAP non-susceptibility is supported by the finding that the inactivation of the mutated mprF can reverse, at least partially, DAP non-susceptibility [21].
MprF protein is composed of 14 transmembrane domains and a cytosolic C-terminal domain [11]. The first eight N-terminal transmembrane domains are involved in translocation of L-PG to the outer CM. The next four transmembrane “central” are involved both in L-PG synthesis and flipping, while the cytosolic C-terminal domain is involved only in L-PG synthesis. Single nucleotide polymorphism (SNP) in mprF genes have been reported in most staphylococcal clinical isolates showing DAP MICs increase [7,10,11], and are associated with a gain of function. Nevertheless, some authors have shown that SNPs can also be found in 30% of DAP- susceptible isolates, and that some DAP-susceptible isolates with MICs of 1 mg/L can show up to 30 amino acid changes in mprF [22], suggesting a high level of heterogeneity within mprF in some staphylococcal isolates. Nevertheless, the DAP-non susceptible isolates studied by these authors [22], showed only one amino acid substitution in MprF (L341S or L826F). Changes in these or in very near positions have been previously reported associated with DAP-non susceptibility (S829L, S295L, P314L, S337L, T345I, T345A, I420N, [7,8,11,23,24].
Unlike results reported by other authors, we found one SNPs in mprF only in one isolate (MSa5). Mutations at the 314 position had been reported previously associated with DAP-non susceptibility, but as P314L [7] instead of P314T, as appears in MSa5. P314L was found in other strain (MSa6, Table 2). The other isolates had a high number of mutations affecting the three areas. They have between 6 and 8 mutations in the N-terminal “flippase” domains (mode: 6 mutations), 17-20 mutations (mode: 18 mutations) in the central, bifunctional domains, and 8- 9 mutations (mode: 8) in the phosphatidylglycerol lysinylation domain. Six isolates (MSa1, MSa4, MSa5, MSa6, MSa7 and MSa8) included in this wide group of mutations some amino acid changes in positions previously associated with DAP-non susceptibility (S295, P314, T345) [7,11,16], but other isolates (MSa2, MSa3 and MSa9,) show mutations previously not reported in association with DAP- non susceptibility. Moreover, they show no mutations in the area comprised between S295 and I420, where most DAP-non susceptibility associated mutations reported are located, though they show more than 30 amino acid changes throughout the gene. The study reflects a large genetic variability between our clinical isolates, and the strain used as reference strain (S. aureus ATCC 25923), especially in some of the genes studied. Some of the mutations observed in the different genes have been previously described associated with daptomycin resistance in previous studies [7,8,11,16,23], but others have not. As already mentioned in the results section, the absence of MLST typing forces us to consider the possibility that, at least some of these previously undescribed changes are not associated with daptomycin resistance, but with the genetic variability existing between the different MLST types.
All the isolates showed rpoB mutations. The rpoB and rpoC genes encode for bacterial RNA polymerase β and β’ subunits [7,8]. Mutations described in rpoB and rpoC associated with DAP- non susceptibility are different from those affecting to other antibiotics, such as rifampicin [25]. Unlike mprF mutations that, at least in vitro, usually emerge at relatively early times during the selection process, rpoB and rpoC mutations emerge later [7]. Some studies have reported rpoB changes in mutants selected in vitro, but not in clinical isolates [7], while other studies [11] did not detect rpoB changes in strains selected in vitro and detected them only in clinical isolates. Some authors [25,26] have shown that single point mutations in rpoB, such as A477D or A621E, can reduce the susceptibility both to daptomycin and vancomycin. Such mutations have been shown to cause cell wall thickening and reduction of the negative charge of the outer layer in S. aureus. All the isolates tested in this study showed a rpoB mutation (F737Y) that had not been previously described in non-susceptible isolates, whether clinical or obtained in vitro. Changes in the rpoC gene were also detected in all isolates, excepting MSa5. Single rpoC mutations (N341D) have also associated with treatment failures in staphylococcal infections [27]. Mutations in the walkR sequence, a regulator of cell wall metabolism and virulence in S. aureus, B. subtilis and S. pneumoniae [28] have been associated with vancomycin resistance [29], and also to reduced DAP susceptibility, in association or not with mprF and agrA mutations [10,29].
fakA had only been associated with DAP-non susceptibility in one study, and only in mutants selected in vitro [10]. In that study, authors find a L133H mutations associated with DAP-non susceptibility. In our study, this protein showed different changes in 8 out of 9 clinical isolates. Since the function of this protein is unknown, it is not possible to know its role in DAP susceptibility, or even if there is really a cause-effect relationship.
cls genes encode for a cardiolipin synthase, and mutations in these genes have been associated with DAP low susceptibility in staphylococci [11]. Cls proteins are membrane-bound enzymes that synthesize cardiolipin, an important anionic membrane phospholipid, from the phosphatidyl moiety of two PG molecules [30]. While in logarithmic growth PG is the main membrane phospholipid, in stationary growth conditions, and under conditions of stress, such as unfavorable growth conditions or cell-wall acting antibiotics, cardiolipin can accumulate up to 25%–30% of membrane phospholipid. cls2 has been the most frequently reported cls gene in this aspect. In this study, all isolates excepting MSa5 showed mutations in this gene. Most isolates shared two mutations (V135I, H205R), and two of them added a third mutation (A459L, A471E). None of these mutations had been described previously. Mutations previously reported appear between the amino acids 20 and 60 (A23V, T33N, L52F, F60S) [11], in the two transmembrane domains described in this protein, while our mutations appear outside these domains and the cardiolipin synthase active domains.
Mutations in cls1 have been described with a much lower frequency in association to DAP reduced susceptibility. Nevertheless, we found mutations in this gene in the same isolates in which we found mutations in cls2, and the number of mutations in this gene/isolate ranged from 6 to 16. Most mutations found in the isolates tested in this study appear outside the transmembrane and cardiolipin synthase active domains, but V18A (in cls1), that appears in all isolates excepting MSa5, and G20A, that appears in MSa6 and MSa9 (Table 3), are in the first transmembrane domain, and I238V, that also appears in all isolates excepting MSa5, and I421M (isolates MSa1, MSa4 and MSa9) appear in cardiolipin synthase domains. Results concerning cls1 and cls2 suggest that, though DAP-non susceptibility associated mutations are more frequently described in cls2, in the isolates tested in this study it seems more likely that the most transcendent mutations are those observed in cls1 since, unlike cls2, in almost all isolates we found mutations that specifically affect the transmembrane and cardiolipin synthase functional domains. Mutations in these areas may have a summation effect with those found in mprF, since the negative charge provided by cardiolipin would be reduced or even suppressed, reducing the attraction between DAP and the bacterial membrane [31].
5. Conclusions
The present study confirms that the acquisition of daptomycin resistance may be associated with different mutational profiles. Therefore, various combinations of mutations in different genes may end up leading to the same result of low susceptibility to daptomycin. However, resistance is mainly associated with mutations in the mprF gene, involved in amount and situation on the membrane of lysyl phosphatidyl glycerol molecules, rpoB, which encodes an RNA polymerase, and cls genes, which encode a cardiolipin synthase, involved in the metabolism of membrane phospholipids. In the isolates analyzed in this study, mprF, cls1 and cls2 show a much higher number of mutations than reported in other studies, many of which had not been described so far. There is no relationship between the number of changes in the genes studied and the degree of sensitivity (MICs). That is, we can find strains with the same MICs and different genetic profiles. Future studies should clarify the real role of each of these mutations.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, JLMB, NGC. Study direction: JLMB. Study development: NG-C, MNGZ, AMBC. Conclusions and writing: NGC, JLMB. All authors have read and agreed to the published version of the manuscript.
Funding
This study was partially financed by Grant GRS 1831/A/18, Gerencia Regional de Salud, Consejería de Sanidad, Junta de Castilla y León.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Institutional Review Board (or Ethics Committee) of Hospital Universitario de Salamanca.
Data Availability Statement
The authors declare that data related to this research are available from the authors upon reasonable request.
Acknowledgments
We thank Dr. Miguel Fajardo (Hospital General Universitario de Badajoz) for providing us with part of the isolates studied.
Conflicts of Interest
The authors declare they have no conflicts of interest.
References
- Eisenstein, B.I.; Oleson, F.B. Jr.; Baltz, R.H. Daptomycin: from the mountain to the clinic, with essential help from Francis Tally, MD. Clin Infect Dis 2010, 50 Suppl 1, S10–S15. [Google Scholar] [CrossRef]
- Steenbergen, J.N.; Alder, J.; Thorne, G.M.; Tally, F.P. Daptomycin: a lipopeptide antibiotic for the treatment of serious Gram-positive infections. J Antimicrob Chemother 2005, 55, 283–8. [Google Scholar] [CrossRef]
- Anastasiou, D.M.; Thorne, G. M.; Luperechio, S.A.; Alder, J.D. In vitro activity of daptomycin against clinical isolates with reduced susceptibilities to linezolid and quinupristin/ dalfopristin. Int J Antimicrob Agents 2006, 28, 385–8. [Google Scholar] [CrossRef]
- Cotroneo, N.; Harris, R.; Perlmutter, N.; Beveridge, T.; Silverman, J.A. Daptomycin exerts bactericidal activity without lysis of Staphylococcus aureus. Antimicrob Agents Chemother 2008, 52, 2223–2225. [Google Scholar] [CrossRef]
- Bhullar, K.; Waglechner, N.; Pawlowski, A.; et al. Antibiotic resistance is prevalent in an isolated cave microbiome. PloS One 2012, 7, 1–11. e34953. [Google Scholar] [CrossRef]
- Bayer, A.S.; Schneider, T.; Sahl, H.G. Mechanisms of daptomycin resistance in Staphylococcus aureus: role of the cell membrane and cell wall. Ann N Y Acad Sci 2013, 1277, 139–58. [Google Scholar] [CrossRef]
- Friedman, L.; Alder, J.D.; Silverman, J.A. Genetic changes that correlate with reduced susceptibility to daptomycin in Staphylococcus aureus. Antimicrob Agents Chemother 2006, 50, 2137–2145. [Google Scholar] [CrossRef]
- Mishra, N.N.; Rubio, A.; Nast, C.C.; Bayer, A.S. Differential Adaptations of Methicillin-Resistant Staphylococcus aureus to Serial In Vitro Passage in Daptomycin: Evolution of Daptomycin Resistance and Role of Membrane Carotenoid Content and Fluidity. Int J Microbiol 2012, 2012, 683450. [Google Scholar] [CrossRef]
- Pillai, S.K.; Gold, H.S.; Sakoulas, G.; Wennersten, C.; Moellering, R.C. Jr.; Eliopoulos, G.M. Daptomycin nonsusceptibility in Staphylococcus aureus with reduced vancomycin susceptibility is independent of alterations in MprF. Antimicrob. Agents Chemother 2007, 51, 2223–2225. [Google Scholar] [CrossRef]
- Song, Y.; Rubio, A.; Jayaswal, R.K.; Silverman, J.A.; Wilkinson, B.J. Additional routes to Staphylococcus aureus daptomycin resistance as revealed by comparative genome sequencing, transcriptional profiling, and phenotypic studies. PLoS One 2013, 8, e58469. [Google Scholar] [CrossRef]
- Peleg, A.Y.; Miyakis, S.; Ward, D.V.; et al. Whole genome characterization of the mechanisms of daptomycin resistance in clinical and laboratory derived isolates of Staphylococcus aureus. PLoS One 2012, 7, e28316. [Google Scholar] [CrossRef]
- Mehta, S.; Cuirolo, A.X.; Plata, K.B.; et al. VraSR two-component regulatory system contributes to mprF-mediated decreased susceptibility to daptomycin in in vivo-selected clinical strains of methicillin-resistant Staphylococcus aureus. Antimicrob Agents Chemother 2012, 56, 92–102. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 27th ed.; CLSI Supplement M100S; Clinical Laboratory Standards Institute: Wayne, Philadelphia (USA), 2017. [Google Scholar]
- Fujimura, S.; Nakano, Y.; Watanabe, A. A correlation between reduced susceptibilities to vancomycin and daptomycin among the MRSA isolates selected in mutant selection window of both vancomycin and daptomycin. J Infect Chemother 2014, 20, 752–756. [Google Scholar] [CrossRef]
- Humphries, R. M, Pollett, S.; Sakoulas, G. A current perspective on daptomycin for the clinical microbiologist. Clin Microbiol Rev 2013, 26, 759–780. [Google Scholar] [CrossRef]
- Cameron, D.R.; Mortin, L.I.; Rubio, A.; et al. Impact of daptomycin resistance on Staphylococcus aureus virulence. Virulence. 2015, 6, 127–131. [Google Scholar] [CrossRef]
- Gómez-Casanova, N.; Siller Ruiz, M.; Muñoz Bellido, J.L. Mechanisms of resistance to daptomycin in Staphylococcus aureus. Rev Esp Quimioter 2017, 30, 391–396. [Google Scholar]
- Ernst, C.M.; Peschel, A. Broad spectrum antimicrobial peptide resistance by MprF-mediated aminoacylation and flipping of phospholipids. Mol Microbiol 2011, 80, 290–299. [Google Scholar] [CrossRef]
- Ernst, C.M.; Staubitz, P.; Mishra, N.; Yang, S.J.; Hornig, G.; Kalbacher, H.; Bayer, A.S.; Kraus, D.; Peschel, A. The bacterial defensin resistance protein MprF consists of separable domains for lipid lysinilation and antimicrobial peptide repulsion. PLOS Pathogens 2009, 5, e1000660. [Google Scholar] [CrossRef]
- Kristian, S.A.; Dürr, M.; Van Strijp, J.A.; Neumeister, B.; Peschel, A. MprF-mediated lysinylation of phospholipids in Staphylococcus aureus leads to protection against oxygen-independent neutrophil killing. Infect Immun 2003, 71, 546–549. [Google Scholar] [CrossRef]
- Yang, S.J.; Nast, C.C.; Mishra, N.N.; Yeaman, M.R.; Fey, P.D.; Bayer, A.S. Cell wall thickening is not a universal accompaniment of the daptomycin nonsusceptibility phenotype in Staphylococcus aureus: evidence for multiple resistance mechanisms. Antimicrob Agents Chemother 2010, 54, 3079–3085. [Google Scholar] [CrossRef]
- Bayer, A.S.; Mishra, N.N.; Sakoulas, G.; Nonejuie, P.; Nast, C.C.; Pogliano, J.; Chen, K.T.; Ellison, S.N.; Yeaman, M.R.; Yang, S.J. Heterogeneity of mprF sequences in methicillin-resistant Staphylococcus aureus clinical isolates: role in cross-resistance between daptomycin and host defense antimicrobial peptides. Antimicrob Agents Chemother 2014, 58, 7462–7. [Google Scholar] [CrossRef]
- Murthy, M.H.; Olson, M.E.; Wickert, R.W.; Fey, P.D.; Jalali, Z. Daptomycin non-susceptible methicillin-resistant Staphylococcus aureus USA 300 isolate. J Med Microbiol 2008, 57, 1036–1038. [Google Scholar] [CrossRef]
- Sabat, A.J.; Tinelli, M.; Grundmann, H.; Akkerboom, V.; Monaco, M.; del Grosso, M.; Errico, G.; Pantosti, A.; Friedrich, A.W. Daptomycin resistant Staphylococcus aureus clinical strain with novel non-synonymous mutations in the mprF and vraS genes: a new insight into daptomycin resistance. Front Microbiol 2018, 9, 2705. [Google Scholar] [CrossRef]
- Cui, L.; Isii, T.; Fukuda, M.; Ochiai, T.; Neoh, H.; Camargo, I.L.B.D.C.; Watanabe, Y.; Shoji, M.; Hiramatsu, K. An rpoB mutation confers dual heteroresistance to daptomycin and vancomycin in S. aureus. Antimicrob Agents Chemother 2010, 54, 5222–33. [Google Scholar] [CrossRef]
- Bæk, K.T.; Thøgersen, L.; Mogenssen, R.G.; Mellergaard, M.; Thomsen, L.E.; Petersen, A.; Skov, S.; Cameron, D.R.; Peleg, A.Y.; Frees, D. Stepwise decrease in daptomycin susceptibility in clinical S. aureus isolates associated with an initial mutation in rpoB and a compensatory inactivation of the clpX gene. Antimicrob Agents Chemother 2015, 59, 6983–91. [Google Scholar] [CrossRef]
- Hagiya, H.; Sugawara, Y.; Kimura, K.; Hamaguchi, S.; Nishi, I.; Hayashi, M.; Akeda, Y.; Tomono, K. Emergence of daptomycin non-susceptible coagulase-negative Staphylococci in patients with cardiovascular device infections: Two cases report investigated by whole genome analysis. Medicine (Baltimore) 2018, 97, e13487. [Google Scholar] [CrossRef]
- Dubrac, S.; Bisicchia, P.; Devine, K.M.; et al. A matter of life and death: cell wall homeostasis and the WalKR (YycGF) essential signal transduction pathway. Mol Microbiol 2008, 7, 1307–1322. [Google Scholar] [CrossRef]
- Howden, B.P.; McEvoy, C.R.; Allen, D.L.; Chua, K.; Gao, W.; Harrison, P.F.; Bell, J.; Coombs, G.; Bennett-Wood, V.; Porter, J.L.; Robins-Browne, R.; Davies, J.K.; Seemann, T.; Stinear, T.P. Evolution of multidrug resistance during Staphylococcus aureus infection involves mutation of the essential two component regulator WalKR. PLoS Pathog 2011, 7, e1002359. [Google Scholar] [CrossRef]
- Koprivnjak, T.; Zhang, D.; Ernst, C.M.; Peschel, A.; Nauseef, W.M.; Weiss, J.P. Characterization of Staphylococcus aureus cardiolipin synthases 1 and 2 and their contribution to accumulation of cardiolipin in stationary phase and within phagocytes. J Bacteriol 2011, 193, 4134–4142. [Google Scholar] [CrossRef]
- Camargo, I. L.; Neoh, H. M.; Cui, L.; Hiramatsu, K. Serial daptomycin selection generates daptomycin-nonsusceptible Staphylococcus aureus strains with a heterogeneous vancomycin-intermediate phenotype. Antimicrob. Agents Chemother 2008, 52, 4289–4299. [Google Scholar] [CrossRef]
Figure 1.
Number of mutations of each clinical isolate for each gene studied.
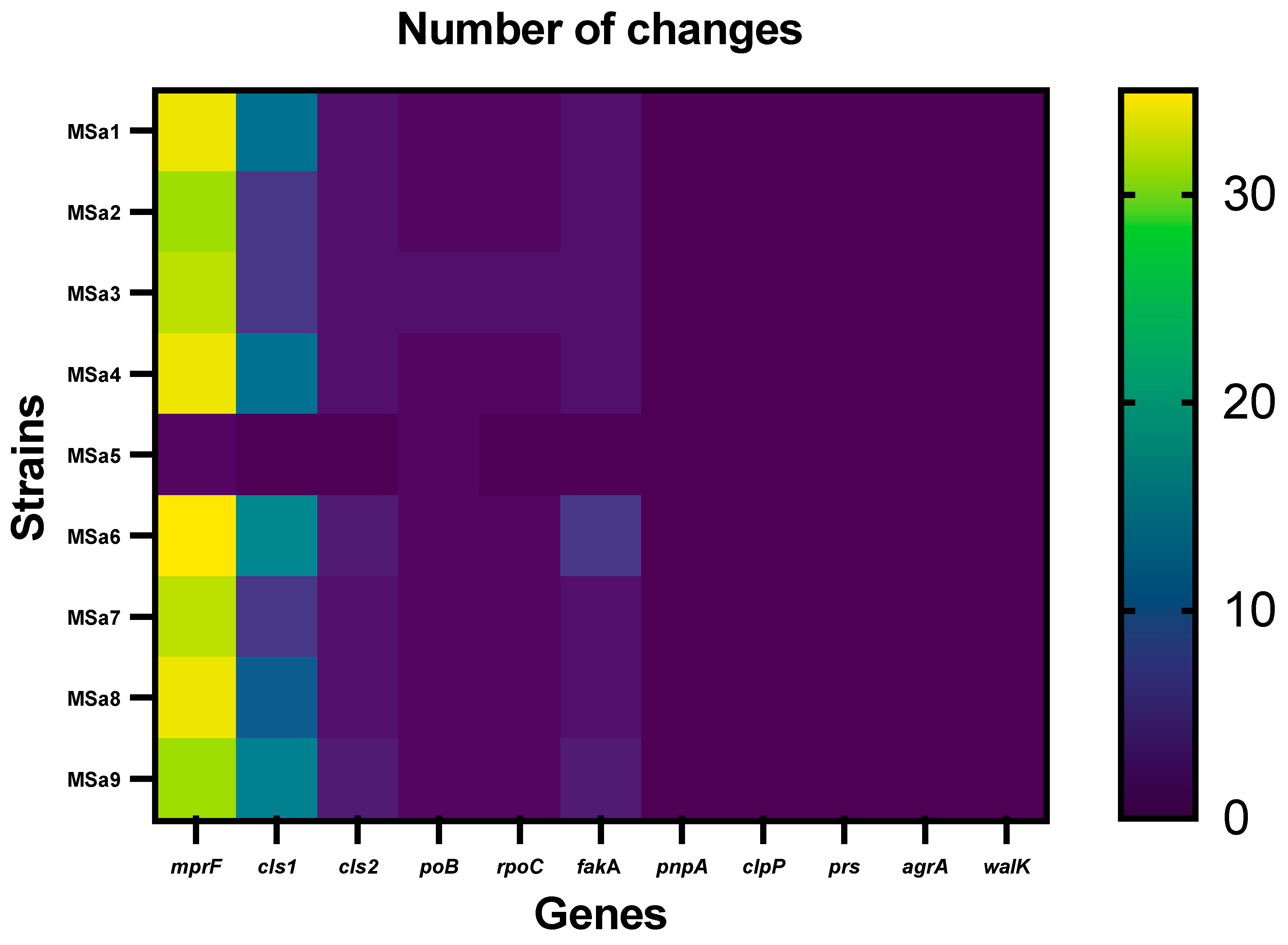

Table 1.
Antibiotic susceptibility of the isolates studied (MIC, mg/L). DAP: Daptomycin; VAN: Vacomycin; OXA: Oxacillin; LZD: Linezolid; DLV: Dalbavancin; TLV: Telavancin. S: susceptible; R: resistant.
Table 1.
Antibiotic susceptibility of the isolates studied (MIC, mg/L). DAP: Daptomycin; VAN: Vacomycin; OXA: Oxacillin; LZD: Linezolid; DLV: Dalbavancin; TLV: Telavancin. S: susceptible; R: resistant.
| Strains | DAP | VAN | OXA | LZD | DLV | TLV |
| MSa1 | 4.0 (R) | 1.0 (S) | 16 (R) | 2.0 (S) | 0.064 (S) | 0.125 (S) |
| MSa2 | 1.0 (S) | 1.0 (S) | 256 (R) | 2.0 (S) | 0.047 (S) | 0.047 (S) |
| MSa3 | 4.0 (R) | 2.0 (S) | 0.125 (S) | 2.0 (S) | 0.100 (S) | 0.125 (S) |
| MSa4* | 2.0 (R) | 2.0 (S) | 2.0 (S) | 2.0 (S) | 0.064 (S) | 0.125 (S) |
| MSa5 | 4.0 (R) | 1.0 (S) | 0.064 (S) | 2.0 (S) | 0.047 (S) | 0.125 (S) |
| MSa6 | 4.0 (R) | 1.0 (S) | 4.0 (R) | 2.0 (S) | 0.032 (S) | 0.125 (S) |
| MSa7* | 2.0 (R) | 1.0 (S) | 2.0 (S) | 2.0 (S) | 0.064 (S) | 0.094 (S) |
| MSa8 | 4.0 (R) | 1.0 (S) | 0.125(S) | 2.0 (S) | 0.047 (S) | 0.125 (S) |
| MSa9 | 2.0 (R) | 1.0 (S) | 0.5 (S) | 2.0 (S) | 0.032 (S) | 0.094 (S) |
*MRSA (mecA +)
Table 2.
Mutations in mprF gene.
| Nº of isolate (MSax) | Amino acid mutations1 | Base changes* |
| 1, 2, 3, 4, 6, 7, 8, 9 |
V26A; N160D; A171V; L174F; Y194F; V223A; L371I; Y400F; I406L; I409T; L413F; V426A; A430V; I446V; L451I; I459L; I464V; F473L; V478I; K489R; I494L; V503I; A505M; N522K; E525D; D531N; D554N; N556T; I575L; T696E; N710E |
T1353395C; A1353696G; C1353730T-T1353731A; G1353740T; A1353799T; T1353886C-C1353887T; C1354329A; A1354417T; A1354434T-C1354436A; T1354444C-T1354445A; A1354457C; T1354495C; C1354507T; A1354554G-T1354556A; C1354569A; A1354593T; A1354608G-C1354610T; T1354637A; G1354650A; A1354683C-A1354684G-A1354685C; A1354698C; G1354725A-T1354727A; G1354731A-C1354732T-A1354733G; T1354784A; G1354793T; G1354809A; G1354878A; A1354885C-T1354886A; A1354941T- C1354943A; A1355305G-C1355304A; A1355346G-T1355348A |
| 1, 4 |
I375M; I461T |
A1354343G; T1354600C-A1354601T |
| 1, 5 |
P314T |
C1354158A-G1354160T |
| 6 |
I9V; G105A; P314L2; P721T |
A1353243G; G1353532C; C1354159T-G1354160T; C1355379 |
| 3 |
L291I |
T1354089A |
| 4, 7 |
T345I2,4,5,6 |
C1354252T |
| 8 |
V287I; S295P3; A500S |
G1354077A-G1354079A; T1354101C; G1354716T- A1354718T |
1 Amino acid numbering corresponds to their position within the mprF gene. * Bases numbering correspond to the complete genome of S. aureus subsp. aureus strain ATCC 25923, (GenBank: CP009361.1). 2 Friedman [7]; 3 Friedman [7] and Peleg [11] S295L; 4 Mishra [8]; 5 Friedman [7], Mishra [8], Murthy [23] and Cameron [16]: T345A.
Table 3.
Mutations in cardiolipin synthase genes (cls1 and cls2).
| Genes | Nº of strain (MSax) | Amino acid mutations1 | Base changes* |
| cls1 | 1, 4, 6, 8, 9 |
K147Q; K170Q; H174K; N197K |
A1306452C; A1306521C; C1306533A-T1306535A; T1306604A |
| 1, 4, 9 | F389Y; I421M; N448K | T1307179A; C1307276G; T1307357G | |
| 1, 2, 3, 4, 6, 7, 8, 9 | I238V | A1306725G-T1306727A | |
| 1, 2, 3, 4, 7, 8 | E469K | G1307418A | |
| 1, 2, 3, 4, 7, 8 | Q2R; F3Y; S4T | A1306018G; T1306021A; T1306023A | |
| 1, 2, 3, 4, 7, 8 | V18A | T1306166C | |
|
6, 9 |
V18A; G87A; V132I; A175V |
T1306166C-C1306167A; G1306273C-A1306274G; G1306407A; C1306537T-T1306538G |
|
| 9 | S43A; T44K | T1306140G; C1306144A-T1306145A | |
| 6, 9 | G20A | G1306072C-A1306073C | |
| 6 |
V300E; G308K; P309S; L310F; S313A; V455A |
T1306912A-T1306913A; G1306935A-G1306936A; C1306938T; G1306943C; T1306950G-A1306952G; T1307377C | |
| 5 | NC | NC | |
|
cls2 |
1, 2, 3, 4, 6, 7, 8, 9 |
V135I; H205R |
G2125012A; A2125223G |
| 6 | A471E | C2126021A | |
| 9 | I459L | A2125984T | |
| 5 | NC | NC |
1Amino acid numbering corresponds to their position within the cls1 and cls2 genes. *Bases numbering correspond to the complete genome of S. aureus subsp. aureus strain ATCC 25923, (GenBank: CP009361.1). NC: No change.
Table 4.
Mutations in rpoB, rpoC and fakA genes.
| Genes | Nº of strain (MSax) | Amino acid mutations1 | Base changes* |
| rpoB | 1, 2, 3, 4, 5, 6, 7, 8, 9 | F737Y | T545943A |
| 3 | M513I | G545272A | |
| rpoC | 1, 2, 3, 4, 6, 7, 8, 9 | V864I | G550011A-T550013A |
| 3 | P100L | C547720T | |
| 5 | NC | NC | |
| fakA | 1, 2, 3, 4, 6, 7, 8, 9 | L214I; D497E | C1191817A; T1192668A |
| 9 | E431D | A1192470C | |
| 6 |
I144V; E277K; Y287H; A513E |
A1191607G; G1192006A; T1192036C; C1192715A | |
| 5 | NC | NC |
1Amino acid numbering corresponds to their position within the rpoB, rpoC and fakA (mw1109) genes. *Bases numbering correspond to the complete genome of S. aureus subsp. aureus strain ATCC 25923, (GenBank: CP009361.1). NC: No change
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



