
Submitted:
29 January 2024
Posted:
30 January 2024
You are already at the latest version
Abstract
The global increase in obesity has significant health implications, with an increased BMI as a known risk factor for diseases such as type 2 diabetes, cardiovascular diseases, or different cancer types. Long-term effectiveness of diet therapy in addressing morbid obesity is extremely limited with no adequate pharmaceutical agents available as treatment option resulting in bariatric surgery as being the only viable option to achieve and maintain significant long-term weight loss. One important role in the overall human health plays the gut microbiome and its complex composition, which is usually altered and reduced in complexity/diversity in severely obese patients. In this study, the influence of bariatric surgery and the resulting weight-loss on the gut microbiome composition of twelve morbid obese (BMI ≥ 40) female adult central European patients was investigated by comparing the relative abundances of the major microbial phyla Actinobacteria, Bacteroidetes, Firmicutes, Proteobacteria prior- and post-surgery. We also aimed to give insight into the major changes of individual prominent and promising future probiotic bacteria characterized by an overall increase in abundance accompanied by a switch of enterotypes. Identifying specific microbial alterations associated with successful weight-loss outcomes may contribute to the development of future therapeutic interventions by supplementation with next-generation probiotics.
Keywords:
gut microbiome
; 16S rRNA sequencing
; obesity
; bariatric surgery
; enterotypes
1. Introduction
Obesity poses a significant public health challenge, resulting from changes in eating habits and disruptions in the body's regulation of energy intake, expenditure, and storage [1,2]. It has become a worldwide epidemic and a global public health crisis, with its prevalence tripling since 1975 in western countries [3,4]. Furthermore, it was first classified as a disease by the World Health Organization in the year 2000 [5]. Unhealthy diets that are high in calories and sedentary lifestyles have been identified as key factors contributing to this widespread problem. However, the exact role of genetic, social, and environmental factors in the development of obesity remains not fully understood [3,6]. First, in 1963, the most severe form of obesity was described with the term “morbid obesity” [7], which is defined by a body mass index (BMI) of ≥ 35 kg/m2 concomitant with significant comorbidities or a BMI of ≥ 40 kg/m2 (Figure S1) [8,9]. The global rise in obesity has eminent health implications, as elevated BMI is a well-established risk factor for diseases such as type 2 diabetes [10], cardiovascular diseases [11] and various cancers like pancreas, colon or exclusively in women endometrial cancer [12]. Moreover, there is a concerning surge in the prevalence of metabolic disturbances associated with obesity in contemporary society and this cluster of risk factors, referred to as metabolic syndrome, affects approximately one in four adults globally [6]. The projected increases in overweight and obesity among adults suggest that the burden of obesity-related morbidity and mortality will continue to rise in the coming decades, especially in combination with a decreased physical activity.
Long-term effectiveness of diet therapy in addressing obesity is extremely limited and currently there are no truly adequate pharmaceutical agents available for the treatment of obesity, particularly in cases of morbid obesity [13]. Bariatric surgery represents the sole viable option for the most severe form of obesity, consistently achieving and maintaining significant weight-loss. Since 1991, the National Institutes of Health (NIH) established guidelines for surgical therapy of morbid obesity, now recognized as bariatric surgery [14]. There are several bariatric surgery methods, the most abundant and established techniques in Germany are gastric bypass as well as sleeve gastrectomy (Figure 1A). These standard techniques primarily perform through a decreased stomach volume and malabsorption to delay the mixing of digestive juice with food [15].
The contribution of the gut microbiota in the development of obesity has gained significant attention in the last years. Research has demonstrated that the microbiomes of obese individuals exhibit structural and functional characteristics, in contrast to their lean counterparts [16,17]. Preliminary evidence indicates that disruptions to the microbiome in obesity promote enhanced extraction of energy from food, leading to disturbances in nutrient distribution and the onset of obesity [18], which strongly suggests the microbiome is a potential target for obesity dedicated therapeutics. The human digestive system harbors trillions of microorganisms, making the gut microbiota highly diverse [19,20] and is recognized as one of the most densely inhabited microbial environments on earth [21,22,23]. While the exact taxonomic composition that defines a "healthy" gut microbiome remains unclear, it is evident that microbial diversity is essential for maintaining the host's physical health. Obese individuals exhibit significantly lower bacterial diversity in comparison to their lean counterparts, and a reduction in fecal microbial gene richness is linked to various physiological indicators of obesity and metabolic syndrome [24,25,26]. Moreover, the aging-process plays a crucial role in metabolism accompanied by changes in diet, medication, physical activity, and lifestyle, which in turn affect the gut microbiome, typically, with a decrease in microbiome diversity over the years [27,28].
Elucidating the functional properties of the complex gut microbiome, often referred to as the black box of the human body, is still in its infancy. Ongoing metagenomic studies have revealed that most of the human gut microbiota consist of two predominant bacterial phyla, Firmicutes and Bacteroidetes, which together account for over 90 % of the total community. Additionally, there are other less dominant phyla present, such as Proteobacteria and Actinobacteria [29,30,31]. It became clear that bariatric surgery and the resulting weight-loss alters both the diversity and composition of the relative abundances of the colon phyla (Expected changes during weight-loss after bariatric surgery are displayed in Figure 1B). This includes a decrease in Firmicutes as well as an increase in Bacteroidetes [32,33]. Several studies discussed the evidence of an increased Firmicutes/Bacteroidetes (F-/B)-ratio as a marker of obesity. In support of this assumption, it was shown that a low-caloric diet and the resulting weight loss result in the approximation of a normal state through a decrease in the F/B-ratio [23,34,35,36].
The less dominant phylum Actinobacteria provides energy through their production of short-chain fatty acids (SCFA), like acetate, propionate, and butyrate, from carbohydrate fermentation. Also, they are crucial for maintaining gut barrier homeostasis and reaching higher abundances in healthy gut microbiome profiles [37,38,39]. In contrast, members belonging to the phylum Proteobacteria are less abundant in the gut of healthy persons and increase while feeding a high-caloric diet [26,40]. In terms of determining of diversity in the human gut microbial ecosystem, one possible parameter is the Shannon index [41]. Several studies observed a noteworthy negative association between the Shannon diversity index and obese people in contrast to lean counterparts [25,42,43] as well as an increase, and therefore a normalization, of alpha-diversity after bariatric surgery [44,45,46].
Contrary to the distribution at the phylum level, there it has been suggested that variation at the genus level is discontinuous but forming three clusters known as enterotypes exhibit varying proportional compositions. As humans in general like to cluster similar aspects into categories, which is also valid for gut microbiomes, and hence were first namely introduced as enterotypes by Arumugam et al. in 2011 [47]. Diets rich in animal protein and fats, resembling a "westernized" diet, were found to be associated with elevated levels of Bacteroides (enterotype 1), while diets high in simple carbohydrates, as commonly observed in agrarian societies, were linked to higher levels of Prevotella (enterotype 2). Whereas the drivers of the enterotype 3 enriched in Ruminococcus are able to degrade mucins [8,47]. Additionally, it was discovered that the gut microbiota responded to short-term dietary interventions. However, it did not result in a shift in their respective enterotype tending to be highly stable [48,49].
The consumption of selected microbes, marketed as probiotics, has been well-documented as a potential method to influence gut microbiota. Probiotics are defined as live microorganisms that, when administered in sufficient quantities, confer a beneficial effect on the host's health [50]. It is important to note that the majority of probiotics available in the market primarily consist of microorganisms from the genera Lactobacillus and Bifidobacterium [51]. However, during the investigation of further potent probiotics, high abundances of Akkermanisa muciniphila were mentioned several times in connection with an improved metabolic status [52,53,54]. It also gained significant attention due to its verified positive correlation to health after bariatric surgery [55].
This study aims to investigate the impact of bariatric surgery and subsequent weight-loss on the gut microbiome of severely obese (BMI 40 <) female patients with various comorbidities. Stool samples were collected from twelve patients both pre- and post-bariatric surgery, which involved either gastric sleeve or gastric bypass procedures. The patients' gut microbiome composition was analyzed using 16S rRNA next-generation sequencing and compared to a healthy and young lean female, as well as to a male control volunteer. By examining changes in the gut microbiota following surgery, this study seeks to shed light on the potential microbial contributions to the therapeutic effects of bariatric surgery and weight-loss. Previous studies, only focused on comparing weight-loss after sleeve gastrectomy with the outcome after a very low-calorie diet (VLCD) over six months [56]. Aiming that both approaches, each involving five patients, lead to significant changes in the gut microbiome composition and nutrient absorption. However, these studies had limitations, including the integration of patients with only the Bacteroides enterotype, the lack of comparison to control groups or a healthy cohort (except for SCFAs), and the limited number of patients included in the intestinal microbiome sequencing analysis. In contrast, our study addressed these limitations by including twelve female morbidly obese patients, as well as two lean controls (a young and healthy female and male). We collected stool samples from the patients one day prior to surgery and at various time points thereafter, with the longest follow-up being 559 days. Additionally, we included patients with different enterotypes to provide a more comprehensive analysis. The overall outcome of our study suggests the need for new and more widespread studies in this field. By overcoming the limitations of previous research and providing a broader scope of investigation, our study serves as a valuable addition to the existing knowledge. Understanding how bariatric surgery influences the human intestinal microbiota can provide valuable insights into the complex interplay between the microbiome and obesity also resulting in changes in immune cell populations of obese patients after bariatric surgery [57]. Furthermore, it may help identify specific microbial markers or alterations associated with successful weight-loss outcomes, ultimately contributing to the development of future therapeutic interventions through the supplementation of probiotics for obesity and related comorbidities.
2. Results
In 1991, the National Institutes of Health established guidelines for the surgical therapy of morbid obesity (BMI ≥40 or BMI ≥35 in the presence of significant comorbidities), now referred to as bariatric surgery, criteria which are also valid at the University Hospital Ulm [13,14]. Here we present results on the analysis of gut microbiota from twelve female severe obese patients with comorbidities like prediabetes, arterial hypertonia, depressive disorder, smoking, arthrosis and/or hyperthyroidism (Table 1). Stool samples were taken one day before surgery and after certain timepoints after the bariatric surgery which was either gastric sleeve or gastric bypass (11 and 1 person, respectively; Table 1). The shortest sampling interval after surgery was 122 days, the longest 559 days and for three patients’ samples at intermediate timepoints were collected, however, the results presented here exclusively refer to the latest timepoint for each patient (Figure 2). Age spanned from 27 to 65 years with most patients being between 40 to 50 years in age. The highest individual body weight before the surgery was 159 kg at a height of 162 cm resulting in also the highest starting BMI of 61 kg/m2 within this cohort (Table 1).
Age influences the composition of the gut microbiome, as age-related processes may influence the gut microbiota and its related metabolic alterations [58]. Similarly, obesity itself severely influences microbiome compositions in comparison to healthy persons [26,59]. In order to eliminate potential differences in microbiome composition due to sex [60], we opted to include a healthy lean male volunteer, along with a respective female person, as controls. Both individuals are young, aged 27 and 32 years, with a BMI of 22 kg/m2 (fitting in the “normal weight” criteria, as shown in Figure S1) and no comorbidities. Thus, they represent “the extreme” for the cohort of this study thereby labeling the starting point of a microbiome on the way to its age-dependent alterations during the lifespan of an individuum.
The expectation following bariatric surgery and success of the therapy is repression of obesity hallmarked by significant reduction of body weight and subsequent normalization of the BMI, assuming that possible severe changes in individual dietary and sportive habits do not nullify the positive therapeutic effects (Figure 1B). In fact, all patients of this study experienced a decrease in bodyweight and BMI after surgery, as measured by the rate of BMI loss per week (“Delta BMI/week” in Figure 3). Although the range of BMI reduction was broad, ranging from 0.2 to 0.6 BMI/week, with a calculated average value of actually 0.4 BMI/week across all patients. The BMI reductions defined as slow (∆BMI < 0.3/week) and fast (∆BMI > 0.3/week) cluster in two groups of patients according to ages higher or below 50 years with the tendency for elder patients to exhibit slower weight loss compared to younger patients. Interestingly, exceptions existed in both groups with patient 022 being 55 years old showing the fastest BMI reduction (-0.6 BMI/week) and patient 023 being the youngest with 27 years, achieved a BMI reduction close to the minimal change (∆BMI = 0.2 BMI/week). Whereas patient 022 is characterized by overall healthy constitutions such as a non-smoker with no arthrosis. Patient 023 exhibited the extreme opposite, having both properties present.
The intestinal microbiota is composed of more than 1,500 species, distributed across more than 50 different phyla [61]. Among these, Bacteroidetes and Firmicutes have been reported as the most dominant phyla, followed by Proteobacteria, Fusobacteria, Tenericutes, Actinobacteria and Verrucomicrobia, constituting up to 90 % of the total microbial population in humans [20,62]. Notably, Firmicutes, Bacteroidetes, Actinobacteria, and Proteobacteria are the most predominant bacterial phyla in the human gut [20].
The microbiome analyses presented here were conducted using 16S rRNA next-generation Illumina sequencing [63,64] of the respective stool samples collected both pre- and post-bariatric surgery, alongside control samples derived from lean volunteers. High diversity, at least in the case of the gut microbiome, has generally been linked to overall health [65]. This association may be due to the enhanced functional redundancy, which can be achieved with a more diverse set of microbial community, and consequently, leading to an increased metabolic flexibility and adaptability [66]. A relative lack of diversity has been discussed as apparent in various diseases, including obesity [24,26,66].
The Shannon diversity index has been widely used in population genetic studies and has thus been proposed as a measure of diversity across various levels, ranging from genes and populations to whole species and even ecosystems and there both richness (number of different taxa) and evenness (equal distribution of taxa) are taken into account [41,67]. Comparing the Shannon diversity indices before and after bariatric surgery revealed a notable increase in diversity for nine out of eleven patients, extending from 3 to 42 % (Figure 4). On average, the index experienced a 10 % increase, rising from 6.7 to 7.4 after the surgery. Interestingly, a decrease of the Shannon diversity index was only observed in patients 012 and 013, who had the highest initial indices before surgery.
Major bacterial phyla in the human gut, namely Actinobacteria, Bacteroidetes, Firmicutes and Proteobacteria, constituted the dominant groups, each accounting for more than 2 % of the total relative abundance. Before the surgery, their relative abundances ranged from 7 to 49 %, following the order Actinobacteria < Proteobacteria < Bacteroidetes < Firmicutes. Together, these phyla accounted for over 76 % in total (Figure 5A and Figure S2A-D). The remaining thirteen phyla, with abundances of each (drastically) lower than 2 %, contributed to not more than 7.3 % in total, and were collectively grouped under the label “Phylum < 2 %” (Figure S2F,G.). Bacteria that could not distinctly be identified below their genus level as a consequence of the sequencing/analysis method constituted “the Unspecific” in our study (Figure S2E).
A slight decrease of 0.95 % in the median of the largest main phyla Firmicutes, as well as a significant increase of 14.75 % in Bacteroidetes abundancy was observed in the patients after surgery during their weight reduction (Figure 5A and Figure S2B,C). These findings align with a previous study investigating microbiome alterations upon changes in dietary behavior towards a low-calorie regimen [22,34].
The Firmicutes/Bacteroidetes-ratio has frequently been discussed in scientific literature as a hallmark of obesity [36]. Increases in Bacteroidetes abundancy accompanied with the resulting decrease of the F/B-ratio, has been positively associated with weight loss in connection with a caloric restricted diet [22,23,34,35,65]. Upon surgery, seven of twelve patients exhibited reductions in the F/B-ratio between 4 and 36 %. Interestingly, on the contrary, the F/B-ratio increased by an average of 37.5 % in five patients, with a maximum difference of 65 % (Figure 5B), indicating an ambiguous pattern of these major bacterial groups after surgery within the cohort of individuals in our study.
In addition, Actinobacteria and of the group labeled as “Phylum < 2 %” gained higher abundances after surgery, with median values increasing by 15 % and 29 %, respectively. Conversely, Proteobacteria and “Unspecific” both decreased after surgery with Proteobacteria assigning the most prominent difference (32 %) of all phyla and “Unspecific” representing the lower end of alterations with only 1 %. Generally, the scatter of values in the Box-Whisker-plots was characterized by a broad distribution, probably indicating a considerable influence of individual parameters like (changes in) dietary composition, activity levels, development of comorbidities or personal lifestyle choices (e.g., smoking cessation, alcohol abstinence) after surgery.
In the year 2011, Arumugam et al. identified so called enterotypes in human gut microbiomes, thereby introducing a system attempting to not only classify these types but also to describe the composition of microbiota with respect to their metabolic capabilities in turn taking also personal preferences in the dietary composition into account for a more holistic view of nutrition, health, and the microbiome [47]. With seven individuals belonging to type 1, the Bacteroides enterotype was dominant before surgery in our cohort (Figure 6). The remaining individuals were distributed among enterotypes 2 and 3, with four members belonging to enterotype 3 and only one grouping into enterotype 2 (Figure 6, Table 1). This distribution demonstrates that even before surgery, the Bacteroides enterotype is dominant, while both others representing deviations from this standard. Microbiome enterotypes per se can be considered as major systematic entities or biological markers not only for the classification of microbiomes but also for the lifestyle and dietary behavior. Significant alterations in these parameters leading to the change from one enterotype to another may probably be the most significant hallmark for success in anti-obesity therapy in terms of microbiome integrity and health. Notably, after surgery, the dominance of type 1 enterotype has extended almost to exclusiveness with only one patient left behind in her original type 3, whereas four patients switched their enterotypes, leading to the complete elimination of enterotype 2 in this cohort of patients. This remarkable major change also led to an approximation of the cohort to the lean control microbiomes which both belonging to type 1 (Figure 6, Table 2).
In addition to major taxa of relevance and higher systematic orders like the enterotypes, a vast amount of literature has accumulated over decades on individual genera of microorganisms with a suspected or proved significant impact on gut related human health. This includes already known prominent probiotic bacteria or strains currently emerging as promising candidates for future probiotics, as well as potent human pathogens (Figure 7A,B). We here have defined a group of health-related bacterial genera that exhibited changes in relative abundance in the patients’ microbiomes. The members of this group showed alterations of at least 3 % in both directions, either gain or loss, in the mean relative abundance within the respective microbiomes. This selected group accounts for approximately 10 % of the mean relative abundance of all genera observed in the microbiomes of patients before and after surgery, as well as in the lean control volunteers.
The largest effect (Delta (∆) relative abundance) of 66 % was observed for Bifidobacteria which are widely recognized and extensively discussed probiotic bacteria. They drastically underrepresented before bariatric surgery but showed a substantial increase after surgery not only approximating but even slightly surpassing the mean relative abundance within the healthy lean controls (Figure 7A). Compared to this, the second group of well-established probiotics, the Lactobacilli, only exhibited a marginal rise of 4 % (Figure 7B). The positive development of the microbiomes, indicated by the increase of Bifidobacteria, appeared to be consolidated by a simultaneous increase of the genus Akkermansia. This includes A. muciniphila, which is at the moment frequently proclaimed as the upcoming next relevant probiotic [68] due to its association with a healthier metabolic status and better clinical outcomes following calorie restriction in overweight/obese adults [52]. The opposite of the scale of alterations (i.e. decline) in relative abundance was marked by the genus Pseudomonas, which can include notable opportunistic human pathogens (e.g. P. aeruginosa), even completely disappeared from the microbiomes of four patients. A similar effect was observed for Escherichia, which were reduced after surgery then precisely meeting the mean value of this genus in the lean control healthy volunteers. Interestingly, the genus Roseburia was considerably declined after surgery and weight loss by 17 % also strongly narrowing the abundance observed in the healthy control group (Figure 7A). Members of the genus Roseburia have been demonstrated to play a role in regulating gut barrier homeostasis [69], a function that may lose importance within the physiological background of a “healing” microbiome following bariatric surgery and weight reduction.
3. Discussion
Obesity has been recognized as one of the most important health threads of so-called developed societies with a western lifestyle including all problematic dietary behavior and reduced physical activity resulting in a high risk to develop obesity associated comorbidities like diabetes and/or cardiovascular diseases. Changing nutrition to low calorie diet concepts can reduce body weight although these attempts are not sufficient for patients suffering from severe obesity and/or metabolic syndrome, since severely obese patients typically fail to persevere long term dietary changes and the resulting productive and efficient calorie reduction. Ultima ratio for these individuals is bariatric surgery nowadays widely accepted as most efficacious and enduring treatment to enforce calorie reduction by limiting food ingestion [70].
In this study, the influence of bariatric surgery and the resulting weight loss on the gut microbiome of morbid obese (BMI >40) female adult patients was investigated prior and post-surgery by comparing the relative abundances of the major microbial phyla Actinobacteria, Bacteroidetes, Firmicutes, Proteobacteria, low abundance phyla of each lower than 2 %, as well as genera without further specification below the genus level which were grouped together in the category “Unspecific”.
Two groups of beneficial bacteria are dominant in the human gut, the Bacteroidetes and the Firmicutes. The relative proportion of Bacteroidetes has been shown to be decreased in obese people in comparison with lean control volunteers, and that this proportion can increase up on weight loss as a consequence of low-calorie diet [22,34]. In the patients of our study cohort this Firmicutes and Bacteroidetes dominance was already present in the microbiomes before bariatric surgery and was kept constant for the median relative abundance of Firmicutes also after surgery with a decrease of only < 1 %. Bacteroidetes in comparison gained abundance after surgery with a drastic increase of 15 %.
Alterations affecting the dominant phyla Firmicutes and Bacteroidetes were first described in obese animals and humans with increased abundances of Firmicutes at the expense of Bacteroidetes [22,34,36]. When these individuals were submitted to a calorie-restricted diet, an increase of their Bacteroidetes abundance was observed as well as the normalization of their Firmicutes/Bacteroidetes ratio, in parallel with weight loss [22,34]. Support came from studies in animals, kept at high-fat or high-fiber diets showing higher Firmicutes and Bacteroidetes abundances [23,35]. Similar findings were reported in children living in rural African areas, who consumed a traditional diet rich in fiber and showed higher proportions of Bacteroidetes and lower of Firmicutes, compared to children from western countries whose diet included large amounts of protein, fat, sugar, and starch [71,72]. These findings and results obtained with obese animals and humans [16,24,73,74,75,76,77], led to the suggestion that the Firmicutes were probably more effective in energy extraction from food than Bacteroidetes, thereby promoting a more efficient absorption of calories and the subsequent gain of body weight and an increase of BMI [36,76].
A high F-/B-ratio has been discussed already in 2011 as a key property of obesity associated non-healthy microbiomes [16,24] suggesting that the development of the mean F-/B-ratio after surgery observed with the patients presented here can be judged as a positive trend. In consequence, also due to associated fermentation activities, occurrence of lower levels of Firmicutes could result in a reduction of energy harvest besides caloric restriction and thus might benefit sustained weight loss and maintenance [56]. It has been reported that Bacteroidetes produce less butyrate than Firmicutes, but more acetate and propionate [78]. Butyrate is generally considered as health-promoting due to its influence [79] on critical obesity related parameters and comorbidities [80,81,82]. Propionate, however, stimulates the secretion of the obesity-related gut hormones glucagon-like peptide-1 (GLP-1) and peptide YY (PYY) leading to inhibition of appetite [36,83]. The coexistence of Bacteroidetes and Firmicutes in the gut has been suspected to imply minimized competition for resources through cooperation or specialization, with a yet unknown mechanism shifting this delicate balance towards Firmicutes [22,34]. In principle, the expected negative effects of a lack of reduction of the Firmicutes abundance may be compensated by the metabolic activities of the Bacteroidetes leading to a behavioral optimization of the patients regarding food intake as a consequence of the physiologic interplay of the fermentation products with the humoral system responsible for appetite control.
The median relative abundance of the phylum Actinobacteria was raised after surgery as the Bacteroidetes by 15 %. Actinobacteria have been designated a relevant minority for the maintenance of gut homeostasis [39]. They also produce the carbohydrate fermentation products propionate and butyrate and, in addition, acetate, which are essential as a source of energy for epithelial cells [37,38]. Moreover, among them, Bifidobacteria have beneficial effects in the maintenance of gut barrier based on their enormous capability to produce such fermentation end-products [84]. Acetate for example can protect the host from enteropathogenic infections such as entero-hemorrhagic Escherichia coli and Shigella [39,85]. Interestingly, Bifidobacteria in our study represented the genus with the strongest increase within the group of health-relevant bacteria, whereas the genera Escherichia and Pseudomonas which both can contain important human pathogens were drastically reduced. Without overinterpreting this coincidence of the changes in the relevant abundances of these prominent probiotics and pathogens, these developments suggest that bariatric surgery and the resulting weight loss may really tip the balance towards microbiomes of increased “healthiness”. Another piece of evidence comes from the phylum Proteobacteria, which in our cohort of patients showed the most pronounced decrease of relative abundance. Apart from the fact that Escherichia, Shigella, Pseudomonas and other Gram-negative pathogens belong to this phylum. In general, Proteobacteria have been described to be present in the gut of healthy humans at low abundances and elevated levels have been designated as a signature of microbial dysbiosis of the gut and even as a probably reliable biomarker [40]. As in our study, the major changes of the gut microbiome of animals and humans have been described to affect Proteobacteria independently from the host species, type of diet and metabolic phenotypes before surgeries [86,87,88,89].
Apart from pathogens as certainly health-relevant bacteria and in addition to Bifidobacteria and Lactobacilli as prominent and for long time commercially available probiotics several gut bacteria arising from studies on diseases different as the Alzheimer disease, other neurodegenerative diseases, chronic intestinal inflammation (e.g. Ulcus ulcerosa, inflammatory bowel disease and Crohn´s disease) are frequently suggested as promising next generation probiotics [90,91]. Roseburia, especially R. intestinalis, has been associated with positive effects on inflammatory processes in the gut presumably mostly due to its high butyrate production capability [92,93]. The fact that Roseburia showed a considerable reduction may be interpreted as indicative for a probable shortfall in necessity of its anti-inflammatory metabolic benefits in the background of abundance adjustments by other bacteria which may compensate for (e.g., Bifidobacteria). In contrast, the genus Akkermansia gained drastic relative abundance. The most eminent example, A. muciniphila, is characterized by its name giving ability to degrade mucin residing in the gut of mammals, including humans [93,94]. The impact of A. muciniphila on health has been discussed in the context of different diseases including metabolic disorders such as obesity [53], diabetes mellitus [95], and neurodegenerative diseases such as multiple sclerosis [96,97], Alzheimer´s [98], and Parkinson´s disease [99]. In mouse models of Alzheimer´s disease, the development of symptoms is accompanied with a reduction of A. muciniphila in the gut microbiome [91]. A marker-like characteristic of microbiomes in obesity is a low abundance of A. muciniphila [55] and upon weight loss a restoration occurs, which is also the case of environmentally induced metabolic switches in animals [100]. A. muciniphila supplementation can be of therapeutic relevance to reduce bodyweight in obese humans [54].
One composition of the human intestinal microbiome that can be influenced by diet are the enterotypes first introduced by Arumugam et al. in 2011 [47]. These states can be identified by their most prominent relative abundance of the genera Bacteroides (Enterotype 1), the in Prevotella enriched enterotype 2, and in this study the most viable Ruminococcus enterotype 3 [47]. The enterotype 1 enriched in Bacteroides was linked to a high protein and animal fat diet, whereas the Prevotella enterotype 2 was related to a diet with a great amount of carbohydrates [101]. Previous studies have demonstrated that these enterotypes tend to be relatively stable over extended periods of time. In an analysis based on two single collection points during the Human Microbiome Project [102], it was found that 84 % of individuals did not switch their enterotype [49]. Over our cohort of twelve severely obese females, a broad variation in enterotypes can be observed with seven patients belonging to the first enterotype, four clustering into enterotype 2 and only one into enterotype 3 before bariatric surgery. Furthermore, we revealed a significant amount of four changes of enterotypes in total post-surgery, divided in three alterations from the in Ruminococcus enriched enterotype 3 prior-surgery to the enterotype 1 containing mostly Bacteroides as is true for the control volunteers.
Our study represents the next important piece of evidence for understanding the development of the microbiome composition upon bariatric surgery and/or the corresponding weight loss. The term “and/or” in this context is not only correct but crucial since after years of small-scale studies with intrinsically extremely limited numbers of patients it is still not possible to unequivocally distinguish between the causes and effects or in other words whether the changes in bacterial abundance distinctly result from surgery or weight loss. However, several properties of the post-surgery microbiome composition and the resulting changes behind it point into the direction of a normalization towards conditions in healthy people. Interestingly, prominent and well-established as well as emerging future probiotics played major roles. The question whether early supplementation with such present or next generation probiotics can support or even prepone the desired “healing” of obesity impaired microbiomes needs to be answered on the way to additional obesity therapies. However, this will require far more detailed and systematic studies with more patients and consequent monitoring not only of comorbidities, but also strict control of the development in dietary behavior and lifestyle after surgery.
4. Materials and Methods
4.1 Study Cohort
The study included twelve adult (≥ 18 years) female patients who underwent bariatric surgery at Ulm University Hospital in the timespan from 2020 to 2022. Eleven out of twelve received a sleeve gastrectomy and one a bypass surgery. These patients also met the criteria outlined in the “German national S3-guideline for the surgical treatment of morbid obesity and metabolic diseases” (BMI ≥ 40 kg/m2 with one or more obesity-associated comorbidities) [15]. Furthermore, patients with inflammatory bowel disease, systemic inflammatory disease, acute infections, cancer, autoimmune disease, or receiving immunosuppressive therapy were excluded from the study. The study participants included in the patient group were of non-Hispanic white ethnicity.
4.2 Anthropometric Measurements and Clinical Data
The body mass index (BMI) in kg/m2, represents the ratio of an individual´s weight in kilograms to the square of their height in meters. Prior to surgery and during follow-up examinations at respective timepoints at Ulm University Hospital, body weight and height were measured.
4.3 16S rDNA Next-Generation Sequencing
Fecal samples of twelve patients before and after bariatric surgery as well as fecal samples of two lean and young control volunteers (one female, one male) were sent to BIOMES laboratory (Wildau, Germany) for bacterial abundance analysis using 16S rRNA next-generation Sequencing. The sequencing was performed using INTEST.pro (Biomes Laboratory, Wildau, Germany), following the method by Lilja et al. [64]. In summary, microbial genomic DNA was extracted using a bead-beating technique, and the V3–V4 region of the 16S rRNA gene [63] was amplified and sequenced on the Illumina MiSeq platform using a 2 × 300 bp paired-end protocol (Illumina, San Diego, CA, USA). Normalized counts (abundance) were calculated by applying biological normalization of the copy number to the raw counts. The relative abundance was then normalized to 100 %, and the resulting data space was transferred to a 0 to 1 value range [64]. The absolute and relative sequence counts for each taxonomical unit were provided.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Classification of normal body weight and different stages of obesity by BMI-scale. The BMI is generally calculated according to the formula given in the “BMI”-field of A. Figure S2: Relative abundance [%] of the identified major phyla of the gut microbiome, their respective medians and the respective percentage delta. Data from patients (P-ID 003-060) before and after bariatric surgery, as well as controls (C1, C2), are displayed. A) Actinobacteria, B) Bacteroidetes, C) Firmicutes, D) Proteobacteria, E) “Unspecific”, F) “Phylum < 2 %”. (G) Mean relative abundance [%] of the individual phyla summarized as “Phylum < 2 %”.
Author Contributions
Conceptualization, F.R.; methodology, A.-K.K.; and F.P.; validation, A.-K.K., F.P., H.-M.R. and I.D.; formal analysis, A.-K.K.; and F.P.; investigation, A.-K.K.; F.P.; A.G. and L.E.; resources, U.K. and F.R.; data curation, A.-K.K.; F.P. and I.D.; writing—original draft preparation, A.-K.K., F.P. and F.R.; writing—review and editing, A.-K.K., F.P., H.-M.R., K.R.S., T.W., U.K. and F.R.; visualization, A.-K.K.; and F.P.; supervision, F.R.; project administration, F.R. All authors have read and agreed to the published version of the manuscript.
Funding
This work did not receive external funding.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Ethics Committee of University Ulm (30/20-TR, 5 April 2022).
Informed Consent Statement
Written informed consent has been obtained from the patients to publish this paper.
Data Availability Statement
Data is contained within the article and Supplementary Material.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Harnack, L.J.; Jeffery, R.W.; Boutelle, K.N. Temporal Trends in Energy Intake in the United States: An Ecologic Perspective. Am J Clin Nutr 2000, 71, 1478–1484. [CrossRef]
- Zhang, H.; DiBaise, J.K.; Zuccolo, A.; Kudrna, D.; Braidotti, M.; Yu, Y.; Parameswaran, P.; Crowell, M.D.; Wing, R.; Rittmann, B.E.; et al. Human Gut Microbiota in Obesity and after Gastric Bypass. Proceedings of the National Academy of Sciences 2009, 106, 2365–2370. [CrossRef]
- Stein, C.J.; Colditz, G.A. The Epidemic of Obesity. J Clin Endocrinol Metab 2004, 89, 2522–2525. [CrossRef]
- Fernandez, M.L. The Metabolic Syndrome. Nutr Rev 2008, 65, S30–S34. [CrossRef]
- Obesity: Preventing and Managing the Global Epidemic. Report of a WHO Consultation. World Health Organ Tech Rep Ser 2000, 894, i–xii, 1–253.
- Green, M.; Arora, K.; Prakash, S. Microbial Medicine: Prebiotic and Probiotic Functional Foods to Target Obesity and Metabolic Syndrome. Int J Mol Sci 2020, 21, 2890. [CrossRef]
- Payne, J.H.; DeWind, L.T.; Commons, R.R. Metabolic Observations in Patients with Jejunocolic Shunts. The American Journal of Surgery 1963, 106, 273–289. [CrossRef]
- Siezen, R.J.; Kleerebezem, M. The Human Gut Microbiome: Are We Our Enterotypes? Microb Biotechnol 2011, 4, 550–553. [CrossRef]
- Wijngaarden, L.H.; Taselaar, A.E.; Nuijten, F.; Harst, E. van der; Klaassen, R.A.; Kuijper, T.M.; Jongbloed, F.; Ambagtsheer, G.; Klepper, M.; IJzermans, J.N.M.; et al. T and B Cell Composition and Cytokine Producing Capacity Before and After Bariatric Surgery. Front Immunol 2022, 13. [CrossRef]
- Hotamisligil, G.S. Inflammation and Metabolic Disorders. Nature 2006, 444, 860–867. [CrossRef]
- Van Gaal, L.F.; Mertens, I.L.; De Block, C.E. Mechanisms Linking Obesity with Cardiovascular Disease. Nature 2006, 444, 875–880. [CrossRef]
- Calle, E.E.; Thun, M.J. Obesity and Cancer. Oncogene 2004, 23, 6365–6378. [CrossRef]
- Buchwald, H.; Avidor, Y.; Braunwald, E.; Jensen, M.D.; Pories, W.; Fahrbach, K.; Schoelles, K. Bariatric Surgery. JAMA 2004, 292, 1724. [CrossRef]
- NIH Conference. Gastrointestinal Surgery for Severe Obesity. Consensus Development Conference Panel. Ann Intern Med 1991, 115, 956–961.
- Runkel, N.; Colombo-Benkmann, M.; Hüttl, T.P.; Tigges, H.; Mann, O.; Sauerland, S. Bariatric Surgery. Dtsch Arztebl Int 2011. [CrossRef]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity Alters Gut Microbial Ecology. Proceedings of the National Academy of Sciences 2005, 102, 11070–11075. [CrossRef]
- Zhou, H.; Urso, C.J.; Jadeja, V. <p>Saturated Fatty Acids in Obesity-Associated Inflammation</P>. J Inflamm Res 2020, Volume 13, 1–14. [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An Obesity-Associated Gut Microbiome with Increased Capacity for Energy Harvest. Nature 2006, 444, 1027–1031. [CrossRef]
- Quigley, E.M.M. Gut Bacteria in Health and Disease. Gastroenterol Hepatol (N Y) 2013, 9, 560–569.
- Gomaa, E.Z. Human Gut Microbiota/Microbiome in Health and Diseases: A Review. Antonie Van Leeuwenhoek 2020, 113, 2019–2040. [CrossRef]
- Whitman, W.B.; Coleman, D.C.; Wiebe, W.J. Prokaryotes: The Unseen Majority. Proceedings of the National Academy of Sciences 1998, 95, 6578–6583. [CrossRef]
- Ley, R.E.; Peterson, D.A.; Gordon, J.I. Ecological and Evolutionary Forces Shaping Microbial Diversity in the Human Intestine. Cell 2006, 124, 837–848. [CrossRef]
- Hildebrandt, M.A.; Hoffmann, C.; Sherrill–Mix, S.A.; Keilbaugh, S.A.; Hamady, M.; Chen, Y.; Knight, R.; Ahima, R.S.; Bushman, F.; Wu, G.D. High-Fat Diet Determines the Composition of the Murine Gut Microbiome Independently of Obesity. Gastroenterology 2009, 137, 1716-1724.e2. [CrossRef]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A Core Gut Microbiome in Obese and Lean Twins. Nature 2009, 457, 480–484. [CrossRef]
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.-M.; Kennedy, S.; et al. Richness of Human Gut Microbiome Correlates with Metabolic Markers. Nature 2013, 500, 541–546. [CrossRef]
- Liu, B.-N.; Liu, X.-T.; Liang, Z.-H.; Wang, J.-H. Gut Microbiota in Obesity. World J Gastroenterol 2021, 27, 3837–3850. [CrossRef]
- Waters, D.L.; Ward, A.L.; Villareal, D.T. Weight Loss in Obese Adults 65years and Older: A Review of the Controversy. Exp Gerontol 2013, 48, 1054–1061. [CrossRef]
- Kong, F.; Deng, F.; Li, Y.; Zhao, J. Identification of Gut Microbiome Signatures Associated with Longevity Provides a Promising Modulation Target for Healthy Aging. Gut Microbes 2019, 10, 210–215. [CrossRef]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A Human Gut Microbial Gene Catalogue Established by Metagenomic Sequencing. Nature 2010, 464, 59–65. [CrossRef]
- Wu, G.D.; Bushmanc, F.D.; Lewis, J.D. Diet, the Human Gut Microbiota, and IBD. Anaerobe 2013, 24, 117–120. [CrossRef]
- Ghosh, S.; Pramanik, S. Structural Diversity, Functional Aspects and Future Therapeutic Applications of Human Gut Microbiome. Arch Microbiol 2021, 203, 5281–5308. [CrossRef]
- Ciobârcă, D.; Cătoi, A.F.; Copăescu, C.; Miere, D.; Crișan, G. Bariatric Surgery in Obesity: Effects on Gut Microbiota and Micronutrient Status. Nutrients 2020, 12, 235. [CrossRef]
- Coimbra, V.O.R.; Crovesy, L.; Ribeiro-Alves, M.; Faller, A.L.K.; Mattos, F.; Rosado, E.L. Gut Microbiota Profile in Adults Undergoing Bariatric Surgery: A Systematic Review. Nutrients 2022, 14, 4979. [CrossRef]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Human Gut Microbes Associated with Obesity. Nature 2006, 444, 1022–1023. [CrossRef]
- de Wit, N.; Derrien, M.; Bosch-Vermeulen, H.; Oosterink, E.; Keshtkar, S.; Duval, C.; de Vogel-van den Bosch, J.; Kleerebezem, M.; Müller, M.; van der Meer, R. Saturated Fat Stimulates Obesity and Hepatic Steatosis and Affects Gut Microbiota Composition by an Enhanced Overflow of Dietary Fat to the Distal Intestine. American Journal of Physiology-Gastrointestinal and Liver Physiology 2012, 303, G589–G599. [CrossRef]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes Ratio: A Relevant Marker of Gut Dysbiosis in Obese Patients? Nutrients 2020, 12, 1474. [CrossRef]
- Scaldaferri, F.; Pizzoferrato, M.; Gerardi, V.; Lopetuso, L.; Gasbarrini, A. The Gut Barrier. J Clin Gastroenterol 2012, 46, S12–S17. [CrossRef]
- Purchiaroni, F.; Tortora, A.; Gabrielli, M.; Bertucci, F.; Gigante, G.; Ianiro, G.; Ojetti, V.; Scarpellini, E.; Gasbarrini, A. The Role of Intestinal Microbiota and the Immune System. Eur Rev Med Pharmacol Sci 2013, 17, 323–333.
- Binda, C.; Lopetuso, L.R.; Rizzatti, G.; Gibiino, G.; Cennamo, V.; Gasbarrini, A. Actinobacteria: A Relevant Minority for the Maintenance of Gut Homeostasis. Digestive and Liver Disease 2018, 50, 421–428. [CrossRef]
- Shin, N.-R.; Whon, T.W.; Bae, J.-W. Proteobacteria: Microbial Signature of Dysbiosis in Gut Microbiota. Trends Biotechnol 2015, 33, 496–503. [CrossRef]
- Shannon, C.E. A Mathematical Theory of Communication. The Bell system technical journal 1948, 27, 379–423.
- Goodrich, J.K.; Waters, J.L.; Poole, A.C.; Sutter, J.L.; Koren, O.; Blekhman, R.; Beaumont, M.; Van Treuren, W.; Knight, R.; Bell, J.T.; et al. Human Genetics Shape the Gut Microbiome. Cell 2014, 159, 789–799. [CrossRef]
- Beaumont, M.; Goodrich, J.K.; Jackson, M.A.; Yet, I.; Davenport, E.R.; Vieira-Silva, S.; Debelius, J.; Pallister, T.; Mangino, M.; Raes, J.; et al. Heritable Components of the Human Fecal Microbiome Are Associated with Visceral Fat. Genome Biol 2016, 17, 189. [CrossRef]
- Guo, Y.; Liu, C.; Shan, C.; Chen, Y.; Li, H.; Huang, Z.; Zou, D. Gut Microbiota after Roux-en-Y Gastric Bypass and Sleeve Gastrectomy in a Diabetic Rat Model: Increased Diversity and Associations of Discriminant Genera with Metabolic Changes. Diabetes Metab Res Rev 2017, 33. [CrossRef]
- Guo, Y.; Huang, Z.-P.; Liu, C.-Q.; Qi, L.; Sheng, Y.; Zou, D.-J. Modulation of the Gut Microbiome: A Systematic Review of the Effect of Bariatric Surgery. Eur J Endocrinol 2018, 178, 43–56. [CrossRef]
- Shao, Y.; Ding, R.; Xu, B.; Hua, R.; Shen, Q.; He, K.; Yao, Q. Alterations of Gut Microbiota After Roux-En-Y Gastric Bypass and Sleeve Gastrectomy in Sprague-Dawley Rats. Obes Surg 2017, 27, 295–302. [CrossRef]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.-M.; et al. Enterotypes of the Human Gut Microbiome. Nature 2011, 473, 174–180. [CrossRef]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.-Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking Long-Term Dietary Patterns with Gut Microbial Enterotypes. Science (1979) 2011, 334, 105–108. [CrossRef]
- Vandeputte, D.; De Commer, L.; Tito, R.Y.; Kathagen, G.; Sabino, J.; Vermeire, S.; Faust, K.; Raes, J. Temporal Variability in Quantitative Human Gut Microbiome Profiles and Implications for Clinical Research. Nat Commun 2021, 12, 6740. [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics Consensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat Rev Gastroenterol Hepatol 2014, 11, 506–514. [CrossRef]
- Douillard, F.P.; de Vos, W.M. Functional Genomics of Lactic Acid Bacteria: From Food to Health. Microb Cell Fact 2014, 13, S8. [CrossRef]
- Dao, M.C.; Everard, A.; Aron-Wisnewsky, J.; Sokolovska, N.; Prifti, E.; Verger, E.O.; Kayser, B.D.; Levenez, F.; Chilloux, J.; Hoyles, L.; et al. Akkermansia Muciniphila and Improved Metabolic Health during a Dietary Intervention in Obesity: Relationship with Gut Microbiome Richness and Ecology. Gut 2016, 65, 426–436. [CrossRef]
- Plovier, H.; Everard, A.; Druart, C.; Depommier, C.; Van Hul, M.; Geurts, L.; Chilloux, J.; Ottman, N.; Duparc, T.; Lichtenstein, L.; et al. A Purified Membrane Protein from Akkermansia Muciniphila or the Pasteurized Bacterium Improves Metabolism in Obese and Diabetic Mice. Nat Med 2017, 23, 107–113. [CrossRef]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia Muciniphila in Overweight and Obese Human Volunteers: A Proof-of-Concept Exploratory Study. Nat Med 2019, 25, 1096–1103. [CrossRef]
- Dao, M.C.; Belda, E.; Prifti, E.; Everard, A.; Kayser, B.D.; Bouillot, J.-L.; Chevallier, J.-M.; Pons, N.; Le Chatelier, E.; Ehrlich, S.D.; et al. Akkermansia Muciniphila Abundance Is Lower in Severe Obesity, but Its Increased Level after Bariatric Surgery Is Not Associated with Metabolic Health Improvement. American Journal of Physiology-Endocrinology and Metabolism 2019, 317, E446–E459. [CrossRef]
- Damms-Machado, A.; Mitra, S.; Schollenberger, A.E.; Kramer, K.M.; Meile, T.; Königsrainer, A.; Huson, D.H.; Bischoff, S.C. Effects of Surgical and Dietary Weight Loss Therapy for Obesity on Gut Microbiota Composition and Nutrient Absorption. Biomed Res Int 2015, 2015, 1–12. [CrossRef]
- Gihring, A.; Gärtner, F.; Mayer, L.; Roth, A.; Abdelrasoul, H.; Kornmann, M.; Elad, L.; Knippschild, U. Influence of Bariatric Surgery on the Peripheral Blood Immune System of Female Patients with Morbid Obesity Revealed by High-Dimensional Mass Cytometry. Front Immunol 2023, 14. [CrossRef]
- Badal, V.D.; Vaccariello, E.D.; Murray, E.R.; Yu, K.E.; Knight, R.; Jeste, D. V.; Nguyen, T.T. The Gut Microbiome, Aging, and Longevity: A Systematic Review. Nutrients 2020, 12, 3759. [CrossRef]
- Tseng, C.-H.; Wu, C.-Y. The Gut Microbiome in Obesity. Journal of the Formosan Medical Association 2019, 118, S3–S9. [CrossRef]
- Min, Y.; Ma, X.; Sankaran, K.; Ru, Y.; Chen, L.; Baiocchi, M.; Zhu, S. Sex-Specific Association between Gut Microbiome and Fat Distribution. Nat Commun 2019, 10, 2408. [CrossRef]
- Robles-Alonso, V.; Guarner, F. [Progress in the Knowledge of the Intestinal Human Microbiota]. Nutr Hosp 2013, 28, 553–557. [CrossRef]
- Jethwani, P.; Grover, K. Gut Microbiota in Health and Diseases – A Review. Int J Curr Microbiol Appl Sci 2019, 8, 1586–1599. [CrossRef]
- Huse, S.M.; Dethlefsen, L.; Huber, J.A.; Welch, D.M.; Relman, D.A.; Sogin, M.L. Exploring Microbial Diversity and Taxonomy Using SSU RRNA Hypervariable Tag Sequencing. PLoS Genet 2008, 4, e1000255. [CrossRef]
- Lilja, S.; Stoll, C.; Krammer, U.; Hippe, B.; Duszka, K.; Debebe, T.; Höfinger, I.; König, J.; Pointner, A.; Haslberger, A. Five Days Periodic Fasting Elevates Levels of Longevity Related Christensenella and Sirtuin Expression in Humans. Int J Mol Sci 2021, 22, 2331. [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The Human Microbiome Project. Nature 2007, 449, 804–810. [CrossRef]
- Lloyd-Price, J.; Abu-Ali, G.; Huttenhower, C. The Healthy Human Microbiome. Genome Med 2016, 8, 51. [CrossRef]
- Konopiński, M.K. Shannon Diversity Index: A Call to Replace the Original Shannon’s Formula with Unbiased Estimator in the Population Genetics Studies. PeerJ 2020, 8, e9391. [CrossRef]
- Ghaffari, S.; Abbasi, A.; Somi, M.H.; Moaddab, S.Y.; Nikniaz, L.; Kafil, H.S.; Ebrahimzadeh Leylabadlo, H. Akkermansia Muciniphila: From Its Critical Role in Human Health to Strategies for Promoting Its Abundance in Human Gut Microbiome. Crit Rev Food Sci Nutr 2023, 63, 7357–7377. [CrossRef]
- Nie, K.; Ma, K.; Luo, W.; Shen, Z.; Yang, Z.; Xiao, M.; Tong, T.; Yang, Y.; Wang, X. Roseburia Intestinalis: A Beneficial Gut Organism From the Discoveries in Genus and Species. Front Cell Infect Microbiol 2021, 11. [CrossRef]
- Elder, K.A.; Wolfe, B.M. Bariatric Surgery: A Review of Procedures and Outcomes. Gastroenterology 2007, 132, 2253–2271. [CrossRef]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of Diet in Shaping Gut Microbiota Revealed by a Comparative Study in Children from Europe and Rural Africa. Proceedings of the National Academy of Sciences 2010, 107, 14691–14696. [CrossRef]
- Ismail, N.A.; Ragab, S.H.; ElBaky, A.A.; Shoeib, A.R.S.; Alhosary, Y.; Fekry, D. Frequency of Firmicutes and Bacteroidetes in Gut Microbiota in Obese and Normal Weight Egyptian Children and Adults. Archives of Medical Science 2011, 3, 501–507. [CrossRef]
- Armougom, F.; Henry, M.; Vialettes, B.; Raccah, D.; Raoult, D. Monitoring Bacterial Community of Human Gut Microbiota Reveals an Increase in Lactobacillus in Obese Patients and Methanogens in Anorexic Patients. PLoS One 2009, 4, e7125. [CrossRef]
- Bäckhed, F.; Ding, H.; Wang, T.; Hooper, L. V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The Gut Microbiota as an Environmental Factor That Regulates Fat Storage. Proceedings of the National Academy of Sciences 2004, 101, 15718–15723. [CrossRef]
- Bervoets, L.; Van Hoorenbeeck, K.; Kortleven, I.; Van Noten, C.; Hens, N.; Vael, C.; Goossens, H.; Desager, K.N.; Vankerckhoven, V. Differences in Gut Microbiota Composition between Obese and Lean Children: A Cross-Sectional Study. Gut Pathog 2013, 5, 10. [CrossRef]
- Krajmalnik-Brown, R.; Ilhan, Z.; Kang, D.; DiBaise, J.K. Effects of Gut Microbes on Nutrient Absorption and Energy Regulation. Nutrition in Clinical Practice 2012, 27, 201–214. [CrossRef]
- Xu, P.; Li, M.; Zhang, J.; Zhang, T. Correlation of Intestinal Microbiota with Overweight and Obesity in Kazakh School Children. BMC Microbiol 2012, 12, 283. [CrossRef]
- Fei, N.; Zhao, L. An Opportunistic Pathogen Isolated from the Gut of an Obese Human Causes Obesity in Germfree Mice. ISME J 2013, 7, 880–884. [CrossRef]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The Role of Short-Chain Fatty Acids in the Interplay between Diet, Gut Microbiota, and Host Energy Metabolism. J Lipid Res 2013, 54, 2325–2340. [CrossRef]
- Gao, Z.; Yin, J.; Zhang, J.; Ward, R.E.; Martin, R.J.; Lefevre, M.; Cefalu, W.T.; Ye, J. Butyrate Improves Insulin Sensitivity and Increases Energy Expenditure in Mice. Diabetes 2009, 58, 1509–1517. [CrossRef]
- Säemann, M.D.; Böhmig, G.A.; Österreicher, C.H.; Burtscher, H.; Parolini, O.; Diakos, C.; Stöckl, J.; Hörl, W.H.; Zlabinger, G.J. Anti-inflammatory Effects of Sodium Butyrate on Human Monocytes: Potent Inhibition of IL-12 and Up-regulation of IL-10 Production. The FASEB Journal 2000, 14, 2380–2382. [CrossRef]
- Soliman, M.M.; Ahmed, M.M.; Salah-eldin, A.; Abdel-Aal, A.A.-A. Butyrate Regulates Leptin Expression through Different Signaling Pathways in Adipocytes. J Vet Sci 2011, 12, 319. [CrossRef]
- Chambers, E.S.; Viardot, A.; Psichas, A.; Morrison, D.J.; Murphy, K.G.; Zac-Varghese, S.E.K.; MacDougall, K.; Preston, T.; Tedford, C.; Finlayson, G.S.; et al. Effects of Targeted Delivery of Propionate to the Human Colon on Appetite Regulation, Body Weight Maintenance and Adiposity in Overweight Adults. Gut 2015, 64, 1744–1754. [CrossRef]
- Hardy, H.; Harris, J.; Lyon, E.; Beal, J.; Foey, A. Probiotics, Prebiotics and Immunomodulation of Gut Mucosal Defences: Homeostasis and Immunopathology. Nutrients 2013, 5, 1869–1912. [CrossRef]
- Fukuda, S.; Toh, H.; Taylor, T.D.; Ohno, H.; Hattori, M. Acetate-Producing Bifidobacteria Protect the Host from Enteropathogenic Infection via Carbohydrate Transporters. Gut Microbes 2012, 3, 449–454. [CrossRef]
- Furet, J.-P.; Kong, L.-C.; Tap, J.; Poitou, C.; Basdevant, A.; Bouillot, J.-L.; Mariat, D.; Corthier, G.; Doré, J.; Henegar, C.; et al. Differential Adaptation of Human Gut Microbiota to Bariatric Surgery–Induced Weight Loss. Diabetes 2010, 59, 3049–3057. [CrossRef]
- Kong, L.-C.; Tap, J.; Aron-Wisnewsky, J.; Pelloux, V.; Basdevant, A.; Bouillot, J.-L.; Zucker, J.-D.; Doré, J.; Clément, K. Gut Microbiota after Gastric Bypass in Human Obesity: Increased Richness and Associations of Bacterial Genera with Adipose Tissue Genes. Am J Clin Nutr 2013, 98, 16–24. [CrossRef]
- Li, J. V.; Ashrafian, H.; Bueter, M.; Kinross, J.; Sands, C.; le Roux, C.W.; Bloom, S.R.; Darzi, A.; Athanasiou, T.; Marchesi, J.R.; et al. Metabolic Surgery Profoundly Influences Gut Microbial-Host Metabolic Cross-Talk. Gut 2011, 60, 1214–1223. [CrossRef]
- Zhang, H.; DiBaise, J.K.; Zuccolo, A.; Kudrna, D.; Braidotti, M.; Yu, Y.; Parameswaran, P.; Crowell, M.D.; Wing, R.; Rittmann, B.E.; et al. Human Gut Microbiota in Obesity and after Gastric Bypass. Proceedings of the National Academy of Sciences 2009, 106, 2365–2370. [CrossRef]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia Muciniphila in Overweight and Obese Human Volunteers: A Proof-of-Concept Exploratory Study. Nat Med 2019, 25, 1096–1103. [CrossRef]
- Raber, H.F.; Kubiczek, D.H.; Bodenberger, N.; Kissmann, A.K.; D’souza, D.; Hu, X.; Mayer, D.; Xu, P.; Knippschild, U.; Spellerberg, B.; et al. Flucell-selex Aptamers as Specific Binding Molecules for Diagnostics of the Health Relevant Gut Bacterium Akkermansia Muciniphila. Int J Mol Sci 2021, 22. [CrossRef]
- Louis, P.; Duncan, S.H.; McCrae, S.I.; Millar, J.; Jackson, M.S.; Flint, H.J. Restricted Distribution of the Butyrate Kinase Pathway among Butyrate-Producing Bacteria from the Human Colon. J Bacteriol 2004, 186, 2099–2106. [CrossRef]
- Tamanai-Shacoori, Z.; Smida, I.; Bousarghin, L.; Loreal, O.; Meuric, V.; Fong, S.B.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A. Roseburia Spp.: A Marker of Health? Future Microbiol 2017, 12, 157–170. [CrossRef]
- Derrien, M.; Vaughan, E.E.; Plugge, C.M.; de Vos, W.M. Akkermansia Muciniphila Gen. Nov., Sp. Nov., a Human Intestinal Mucin-Degrading Bacterium. Int J Syst Evol Microbiol 2004, 54, 1469–1476. [CrossRef]
- Larsen, N.; Vogensen, F.K.; van den Berg, F.W.J.; Nielsen, D.S.; Andreasen, A.S.; Pedersen, B.K.; Al-Soud, W.A.; Sørensen, S.J.; Hansen, L.H.; Jakobsen, M. Gut Microbiota in Human Adults with Type 2 Diabetes Differs from Non-Diabetic Adults. PLoS One 2010, 5, e9085. [CrossRef]
- Chen, J.; Chia, N.; Kalari, K.R.; Yao, J.Z.; Novotna, M.; Paz Soldan, M.M.; Luckey, D.H.; Marietta, E. V.; Jeraldo, P.R.; Chen, X.; et al. Multiple Sclerosis Patients Have a Distinct Gut Microbiota Compared to Healthy Controls. Sci Rep 2016, 6, 28484. [CrossRef]
- Cantarel, B.L.; Waubant, E.; Chehoud, C.; Kuczynski, J.; DeSantis, T.Z.; Warrington, J.; Venkatesan, A.; Fraser, C.M.; Mowry, E.M. Gut Microbiota in Multiple Sclerosis: Possible Influence of Immunomodulators. Journal of Investigative Medicine 2015, 63, 729–734. [CrossRef]
- Ou, Z.; Deng, L.; Lu, Z.; Wu, F.; Liu, W.; Huang, D.; Peng, Y. Protective Effects of Akkermansia Muciniphila on Cognitive Deficits and Amyloid Pathology in a Mouse Model of Alzheimer’s Disease. Nutr Diabetes 2020, 10, 12. [CrossRef]
- Heintz-Buschart, A.; Pandey, U.; Wicke, T.; Sixel-Döring, F.; Janzen, A.; Sittig-Wiegand, E.; Trenkwalder, C.; Oertel, W.H.; Mollenhauer, B.; Wilmes, P. The Nasal and Gut Microbiome in Parkinson’s Disease and Idiopathic Rapid Eye Movement Sleep Behavior Disorder. Movement Disorders 2018, 33, 88–98. [CrossRef]
- Kissmann, A.K.; Rosenau, F.; Herwig, A.; Diedrich, V. Short Photoperiod-Dependent Enrichment of Akkermansia Spec. as the Major Change in the Intestinal Microbiome of Djungarian Hamsters (Phodopus Sungorus). Int J Mol Sci 2023, 24. [CrossRef]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.-Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking Long-Term Dietary Patterns with Gut Microbial Enterotypes. Science (1979) 2011, 334, 105–108. [CrossRef]
- Peterson, J.; Garges, S.; Giovanni, M.; McInnes, P.; Wang, L.; Schloss, J.A.; Bonazzi, V.; McEwen, J.E.; Wetterstrand, K.A.; Deal, C.; et al. The NIH Human Microbiome Project. Genome Res 2009, 19, 2317–2323. [CrossRef]
Figure 1.
A) Comparison of stomachs and indicated direction in which the food is digested by the stomach (black) and the flow of the digestive juice (blue). Left: Stomach after sleeve gastrectomy where ≥ 80 % of the stomach is resected. Middle: Stomach before bariatric surgery. Right: Stomach after gastric bypass where the stomach is taken down right after the gastric inlet to create a gastric pouch that the food bypasses a major part of the stomach. B) Expectations (Exp.) of the influence of weight reduction from morbid obesity (BMI > 40) to normal weight (BMI 18.5-24.9) through bariatric surgery on the human gut microbiome and other factors are listed. In addition to the expectations, the correlations found in our study (real) are shown as far as information can be provided, in case of the enterotype, there was a change to mostly type 1.
Figure 1.
A) Comparison of stomachs and indicated direction in which the food is digested by the stomach (black) and the flow of the digestive juice (blue). Left: Stomach after sleeve gastrectomy where ≥ 80 % of the stomach is resected. Middle: Stomach before bariatric surgery. Right: Stomach after gastric bypass where the stomach is taken down right after the gastric inlet to create a gastric pouch that the food bypasses a major part of the stomach. B) Expectations (Exp.) of the influence of weight reduction from morbid obesity (BMI > 40) to normal weight (BMI 18.5-24.9) through bariatric surgery on the human gut microbiome and other factors are listed. In addition to the expectations, the correlations found in our study (real) are shown as far as information can be provided, in case of the enterotype, there was a change to mostly type 1.
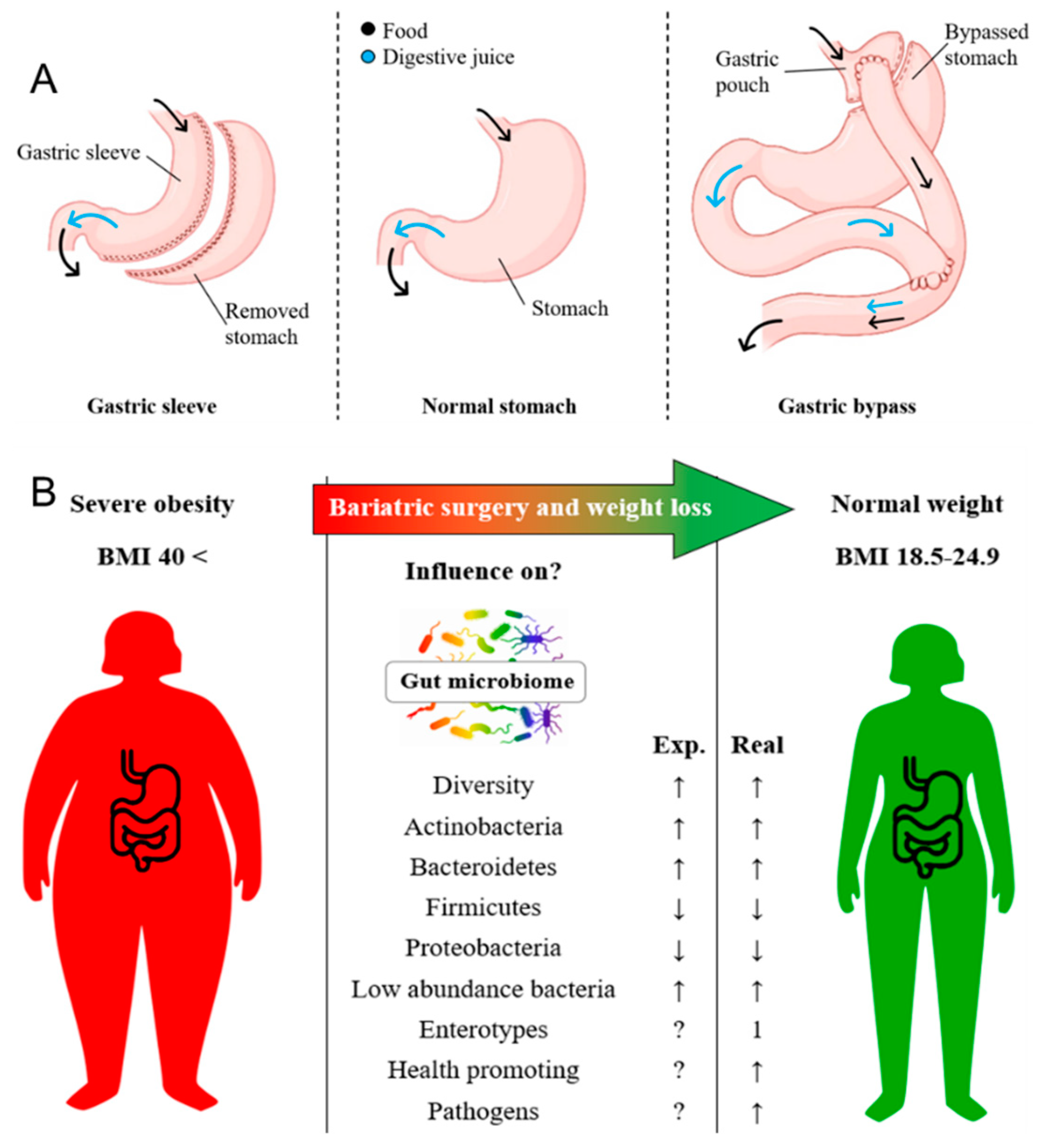
Figure 2.
Timeline overview divided in quarters of all in this study included twelve patient-IDs (P-ID 003-060) and corresponding fecal sampling points. Day -1 describes the day prior to bariatric surgery for all patient samples. The shortest extraction point post-surgery was Day 122, the longest day 559. In three patients, two stool samples were taken after surgery, whereby the respective last time point was included as post-operative in the following considerations.
Figure 2.
Timeline overview divided in quarters of all in this study included twelve patient-IDs (P-ID 003-060) and corresponding fecal sampling points. Day -1 describes the day prior to bariatric surgery for all patient samples. The shortest extraction point post-surgery was Day 122, the longest day 559. In three patients, two stool samples were taken after surgery, whereby the respective last time point was included as post-operative in the following considerations.
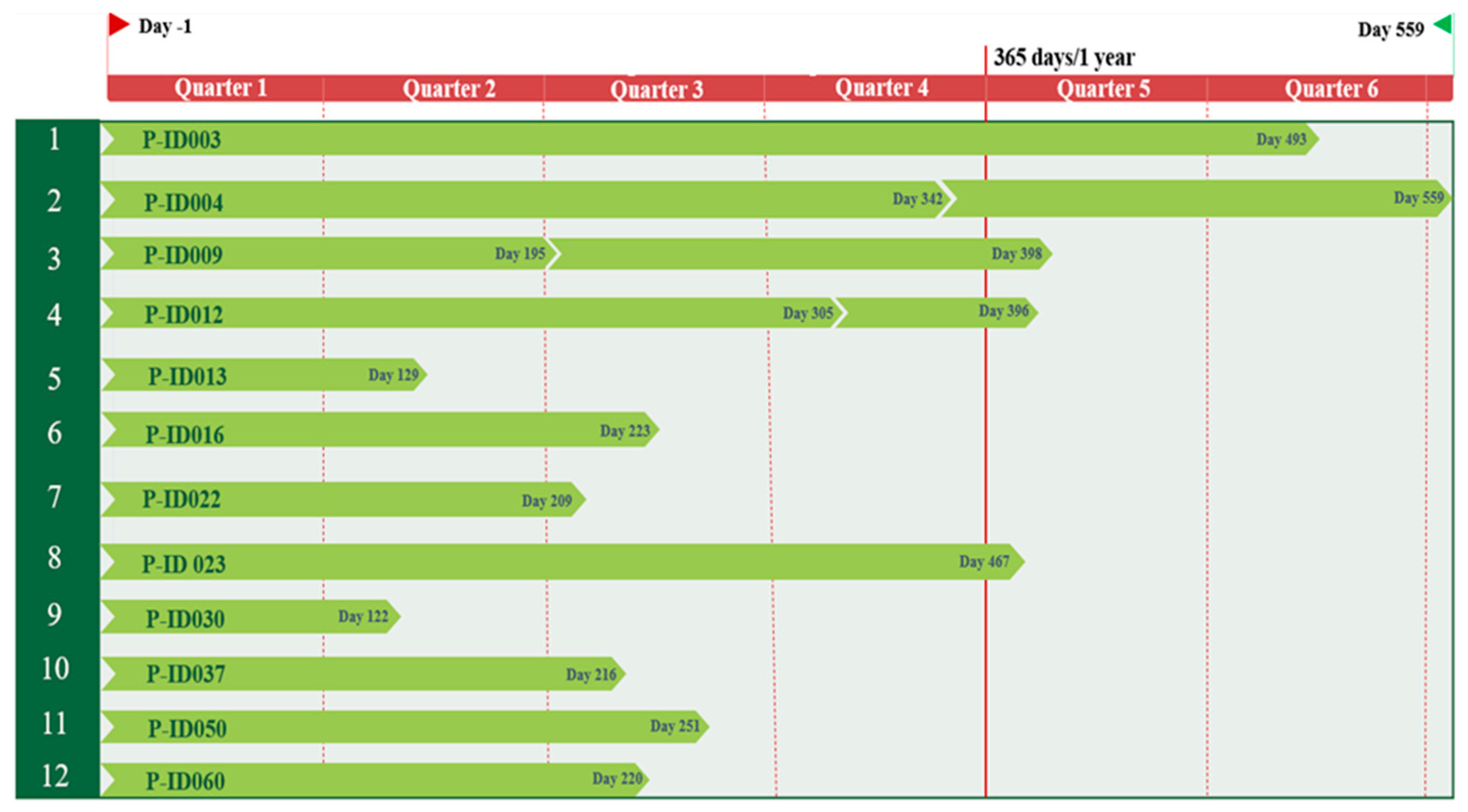
Figure 3.
Changes in patient BMI (P-ID 003-060) per week ascending by age. Distinction between rapid decrease (∆BMI > 0.3/week; green) and slow decrease (∆BMI < 0.3/week; red). Shown are the clusters under 50 years and above.
Figure 3.
Changes in patient BMI (P-ID 003-060) per week ascending by age. Distinction between rapid decrease (∆BMI > 0.3/week; green) and slow decrease (∆BMI < 0.3/week; red). Shown are the clusters under 50 years and above.
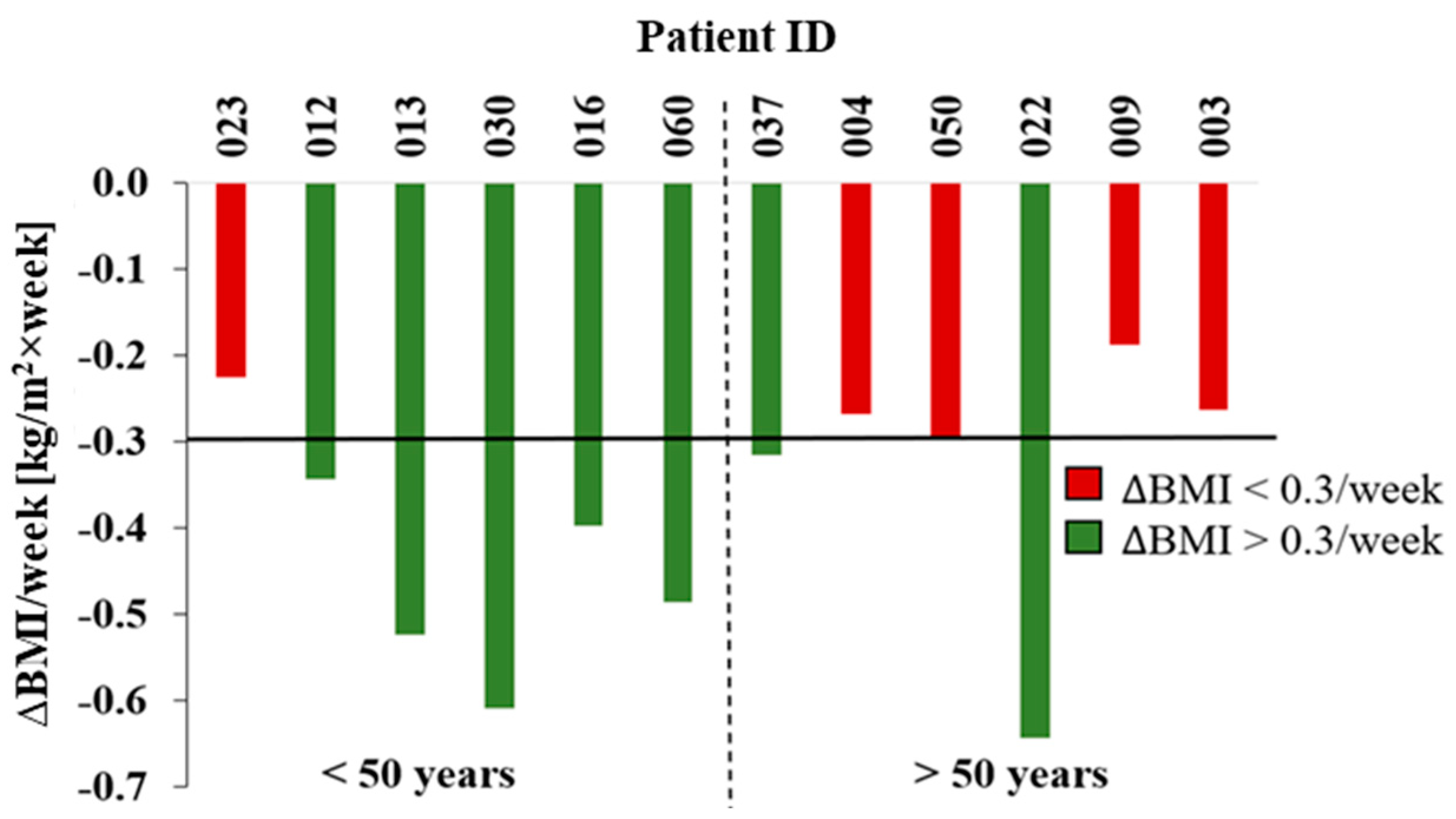
Figure 4.
Alpha-diversity analysis of microbiomes of patients (P-ID 003-060) before (red columns) and after (green columns) bariatric surgery through the Shannon index. Above the paired columns the percentage delta of the entropies is shown.
Figure 4.
Alpha-diversity analysis of microbiomes of patients (P-ID 003-060) before (red columns) and after (green columns) bariatric surgery through the Shannon index. Above the paired columns the percentage delta of the entropies is shown.
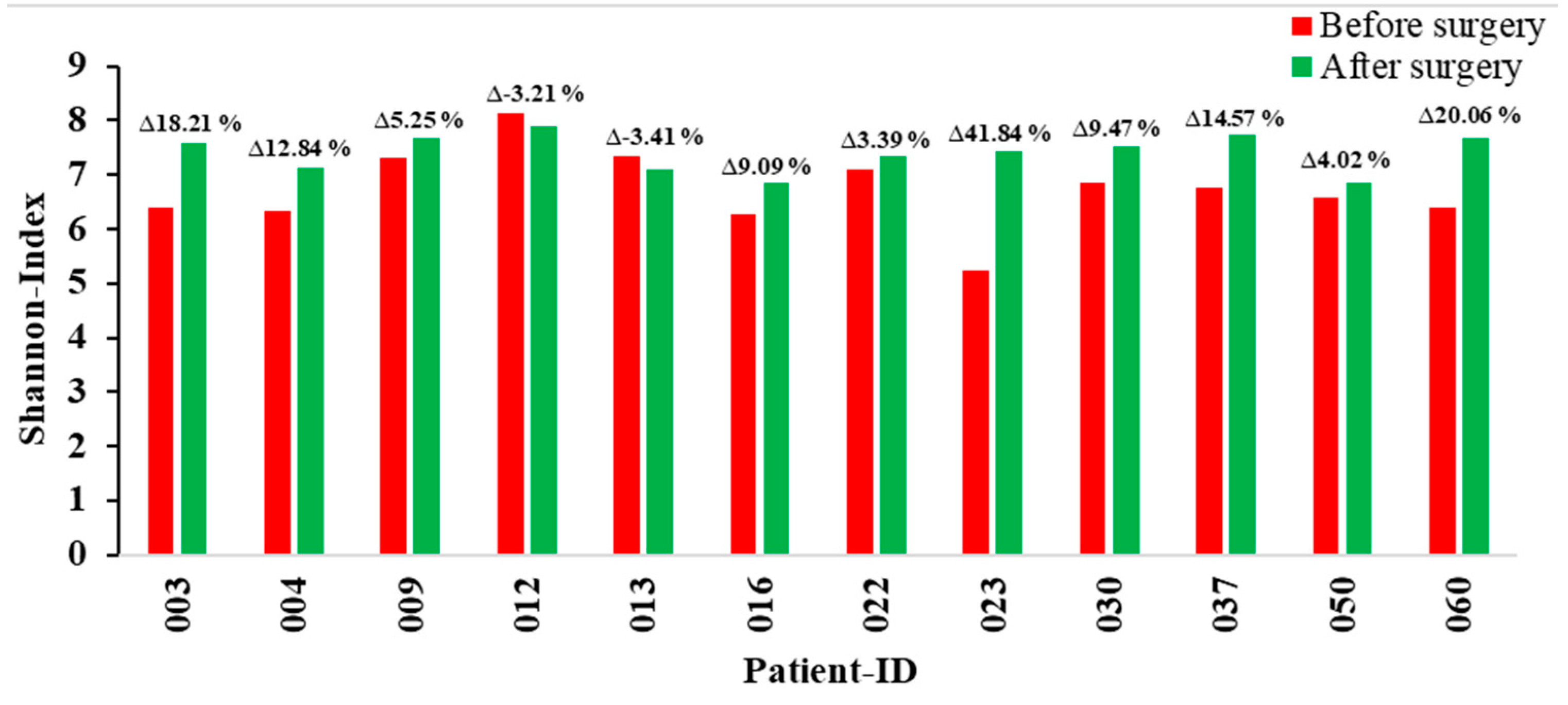
Figure 5.
A) Box-Whisker-Plot of the major phylum groups Actinobacteria, Bacteroidetes, Firmicutes, Proteobacteria, Phylum < 2 % and Unspecific of the patients (P-ID 003-060) as well as the per-centage delta of each median is shown above the boxplot groups. Left boxes (bright colors) represent the relative abundance of the phylum before, right boxes (pale colors) the values after bariatric surgery. B) Ratio of the relative abundance of the gut microbiome phyla Firmicutes to Bacteroidetes. Depict-ed are data of patients before (green columns) and after (red columns) bariatric surgery (P-ID 003-060). The percentage difference of the F/B-ratio from before to after bariatric surgery of the patients is shown above the columns.
Figure 5.
A) Box-Whisker-Plot of the major phylum groups Actinobacteria, Bacteroidetes, Firmicutes, Proteobacteria, Phylum < 2 % and Unspecific of the patients (P-ID 003-060) as well as the per-centage delta of each median is shown above the boxplot groups. Left boxes (bright colors) represent the relative abundance of the phylum before, right boxes (pale colors) the values after bariatric surgery. B) Ratio of the relative abundance of the gut microbiome phyla Firmicutes to Bacteroidetes. Depict-ed are data of patients before (green columns) and after (red columns) bariatric surgery (P-ID 003-060). The percentage difference of the F/B-ratio from before to after bariatric surgery of the patients is shown above the columns.
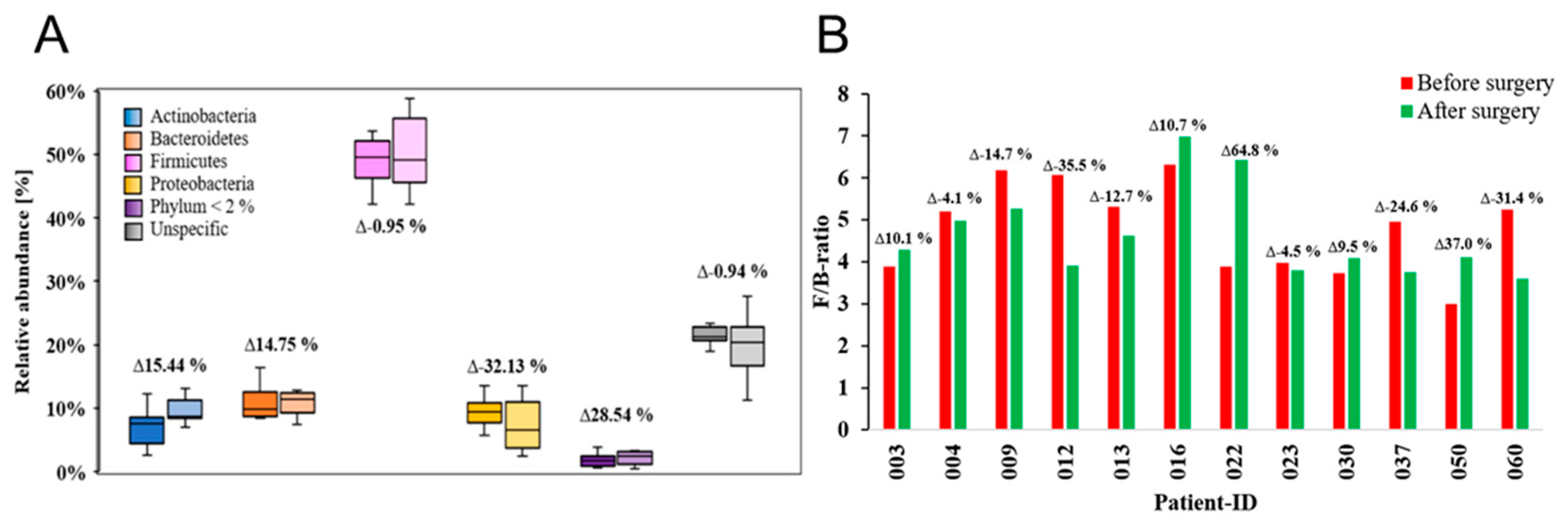
Figure 6.
Associated relative abundance of enterotypes of patients (P-ID 003 to 060), as well as their respective value and lean controls (C1 and C2). The genera Bacteroides (enterotype 1), Prevotella (enterotype 2) and Ruminococcus (enterotype 3) are shown.
Figure 6.
Associated relative abundance of enterotypes of patients (P-ID 003 to 060), as well as their respective value and lean controls (C1 and C2). The genera Bacteroides (enterotype 1), Prevotella (enterotype 2) and Ruminococcus (enterotype 3) are shown.
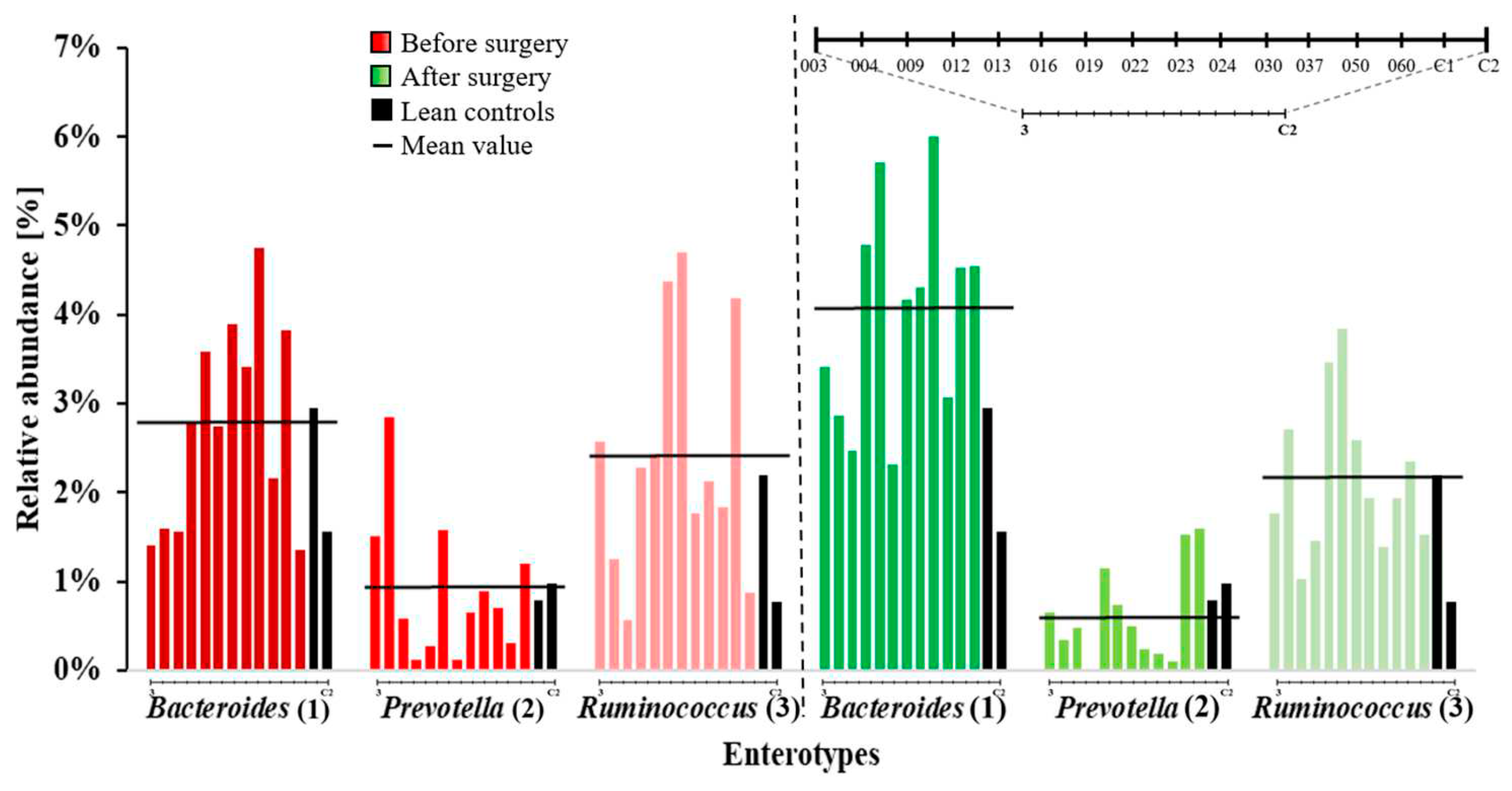
Figure 7.
A) Mean values of percent relative abundances in complete microbiomes of known health-related bacteria in patients (P-ID 003-060) before and after surgery, as well as in lean controls are displayed. The group of known bacteria relevant to health consist of: Akkermansia, Bifidobacterium, Blautia, Escherichia, Faecalibacterium, Lactobacillus, Pseudomonas, Rikenella and Roseburia. B) Changes (∆) in the mean relative abundance of known health-related bacteria in patients sorted by greatest increase to greatest decrease from before to after surgery.
Figure 7.
A) Mean values of percent relative abundances in complete microbiomes of known health-related bacteria in patients (P-ID 003-060) before and after surgery, as well as in lean controls are displayed. The group of known bacteria relevant to health consist of: Akkermansia, Bifidobacterium, Blautia, Escherichia, Faecalibacterium, Lactobacillus, Pseudomonas, Rikenella and Roseburia. B) Changes (∆) in the mean relative abundance of known health-related bacteria in patients sorted by greatest increase to greatest decrease from before to after surgery.

Table 1.
Overview of patients and lean controls. Assigned to their respective sample-IDs (003-060, C1, C2) are gender, surgery type (S ≙ Gastric sleeve, B ≙ Gastric bypass), age, height, starting and end BMI, weight before and after bariatric surgery for patients or weight for lean controls, as well as pre-existing conditions and whether they smoke (Ex ≙ Ex-smoker) through checkmarks (✓). Pre-existing conditions (Arterial hypertonia, Hyperthyroidism, Arthrosis, Prediabetes, Depressive disorder) are marked with a checkmark if they apply to the patients or lean controls.
Table 1.
Overview of patients and lean controls. Assigned to their respective sample-IDs (003-060, C1, C2) are gender, surgery type (S ≙ Gastric sleeve, B ≙ Gastric bypass), age, height, starting and end BMI, weight before and after bariatric surgery for patients or weight for lean controls, as well as pre-existing conditions and whether they smoke (Ex ≙ Ex-smoker) through checkmarks (✓). Pre-existing conditions (Arterial hypertonia, Hyperthyroidism, Arthrosis, Prediabetes, Depressive disorder) are marked with a checkmark if they apply to the patients or lean controls.
| S-ID | Sex | Surgery | Age [years] | Height [cm] | Start BMI [kg/m2] | Weight before surgery [kg] |
Weight after surgery [kg] |
End BMI [kg/m2] | Arterial hypertonia | Hyperthyroidism | Arthrosis | Prediabetes | Depressive disorder | Smoking |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 003 | F | S | 65 | 163 | 44 | 117 | 72 | 27 | ✓ | ✓ | ✓ | |||
| 004 | F | S | 54 | 162 | 61 | 159 | 103 | 39 | ✓ | ✓ | ✓ | |||
| 009 | F | S | 57 | 177 | 42 | 131 | 98 | 31 | ✓ | ✓ | ✓ | ✓ | ||
| 012 | F | S | 30 | 158 | 47 | 118 | 70 | 28 | ✓ | ✓ | ||||
| 013 | F | S | 30 | 173 | 50 | 150 | 121 | 40 | ✓ | ✓ | ✓ | ✓ | ✓ | |
| 016 | F | S | 39 | 163 | 41 | 109 | 75 | 28 | ✓ | |||||
| 022 | F | S | 55 | 173 | 49 | 147 | 89 | 30 | ✓ | |||||
| 023 | F | S | 27 | 167 | 41 | 114 | 72 | 26 | ✓ | ✓ | ✓ | |||
| 030 | F | S | 33 | 170 | 42 | 122 | 91 | 32 | ✓ | ✓ | Ex | |||
| 037 | F | S | 45 | 165 | 43 | 117 | 91 | 33 | ✓ | |||||
| 050 | F | S | 54 | 152 | 50 | 115 | 91 | 39 | ✓ | |||||
| 060 | F | B | 40 | 170 | 50 | 144 | 100 | 35 | ✓ | |||||
| C1 | F | - | 27 | 158 | 22 | 56 | 22 | |||||||
| C2 | M | - | 32 | 179 | 22 | 70 | 22 | |||||||
Table 2.
Overview of sample IDs and respective enterotypes before and after surgery of patients (P-ID003-060) and enterotype of lean controls (C1 and C2). Changes in the enterotype of the patients after surgery are indicated by checkmarks (✓).
Table 2.
Overview of sample IDs and respective enterotypes before and after surgery of patients (P-ID003-060) and enterotype of lean controls (C1 and C2). Changes in the enterotype of the patients after surgery are indicated by checkmarks (✓).
| S-ID | Before Surgery | After Surgery | Change of Enterotype |
|---|---|---|---|
| 003 | III | I | ✓ |
| 004 | II | I | ✓ |
| 009 | I | I | |
| 012 | I | I | |
| 013 | I | I | |
| 016 | III | III | |
| 022 | III | I | ✓ |
| 023 | I | I | |
| 030 | I | I | |
| 037 | I | I | |
| 050 | III | I | ✓ |
| 060 | I | I | |
| C1 | I I |
||
| C2 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



