Submitted:
29 January 2024
Posted:
30 January 2024
You are already at the latest version
Abstract
The urgent global health challenge posed by methicillin-resistant Staphylococcus aureus (MRSA) infections demands effective solutions. Antimicrobial peptides (AMPs) represent promising tools of research of new antibacterial agents and LyeTx I mn∆K, a short synthetic peptide based on the Lycosa erythrognatha spider venom, is a good representative. This study focused on analyzing some antimicrobial activities of LyeTx I mn∆K, including minimum inhibitory and bactericidal concentrations, synergy and resensitization assays, lysis activity, the effect on biofilm, and the bacterial death curve in MRSA. Additionally, its characterization was conducted through isothermal titration calorimetry, dynamic light scattering, calcein release, and finally, efficacy in a mice wound model. The peptide demonstrating remarkable efficacy against planktonic cells and biofilms of MRSA, outperforms vancomycin in terms of rapid bactericidal action and anti-biofilm effects. The mechanism involves significant membrane damage. Interactions with bacterial model membranes, including those with lysylphosphatidylglycerol (LysylPOPG) modifications, highlight versatility and selectivity of this compound. Also, the peptide has the ability to sensitize resistant bacteria to conventional antibiotics, showing potential for combinatory therapy. Furthermore, using an in vivo model, was showed that a formulated gel containing the peptide proved superior to vancomycin in treating MRSA-induced wound in mice. Together the results highlight LyeTx I mnΔK as a promising prototype for the development of effective therapeutic strategies against superficial MRSA infections.
Keywords:
Antimicrobial peptides
; Methicillin-resistant Staphylococcus aureus (MRSA)
; Lycosa erythrognatha
; Skin infections
; Action on membrane.
1. Introduction
Staphylococcus aureus is a gram-positive coccus, catalase positive, coagulase positive and PYR-positive bacteria that is recognized as one of the major infectious disease-causing pathogens in humans. Antimicrobials such as β-lactams are generally used for chemotherapy of these diseases; however, the emergence of methicillin-resistant S. aureus (MRSA), which is virtually resistant to all antimicrobials from this class, has posed serious problems [1]. Currently, it is estimated that 11,000 people die each year from MRSA-related infections in the USA, representing nearly half of all fatalities caused by multidrug-resistant (MDR) pathogens in the country [2]. Furthermore, estimates indicate that MRSA kills more Americans each year than HIV/AIDS, Parkinson’s disease, tuberculosis, emphysema, and homicide combined [3]. In cases of MRSA infections, few pharmacological options are clinically available, such as vancomycin, mupirocin and daptomycin, which are drugs that have a slow bactericidal effect, significant adverse reactions and are expensive [4]. Thus, the development of new effective and safe therapeutic agents against MRSA-induced infections is imperative to control the public health emergency related to this superbug [5].
In this scenario, antimicrobial peptides (AMPs) highlight as a promising source of new antimicrobials against MDR pathogens, especially against MRSA [6,7]. The AMPs are components of innate immunity found in various species, from bacteria and fungi to animals and plants [8,9]. There are eight AMPs approved for clinical use by the Food and Drug Administration (FDA) (i.e., gramicidin, colistin, polymyxin B, daptomycin, vancomycin, oritavancin, dalbavancin and telavancin), all of them are considered “last line” antimicrobials and active even against MDR specimens [10]. These agents are known to act on the cell membrane of bacterial pathogens stimulating the lysis and release of cytoplasmic contents. Furthermore, they can cross the membrane inhibiting different metabolic pathways in bacterial cell and can modulate the antibacterial immune response [11,12,13]. AMPs have a series of advantages over conventional therapies such as a rapid bactericidal effect, low potential for inducing resistance, absence of the formation of active residues that can contaminate the environment and action on MDR isolates [14,15].
One of the main sources of AMPs in nature are animal toxins. In fact, numerous compounds with antimicrobial activity have been isolated from the venom of spiders, scorpions, snakes, frogs, bees, wasps, ants and other animals, highlighting this biological matrix as a potential source for the search for new antimicrobials against MRSA. Herein, our group has isolated a peptide containing 25 amino acid residues (H-IWLTALKFLGKNLGKHLAKQQLAKL-NH2) from the venom of the Brazilian spider Lycosa erythrognatha, popularly known as the “aranha-lobo”, “aranha-de-jardim”, or “tarantula”, which has demonstrated good antimicrobial activity. In vitro studies shown that the LyeTx I peptide proved to be active against Escherichia coli (MIC: 1.41-7.81 μM), Pseudomonas aeruginosa (MIC: 2.82 μM), Acinetobacter baumannii (MIC: 0.70 μM), Staphylococcus aureus (MIC: 0.70-3.79 μM), Staphylococcus epidermidis (MIC: 0.70 μM), Pichia kudriavzevii (formerly Candida krusei) (MIC: 14.23-26.30 μM), Cryptococcus gattii (MIC: 2.23 μM) and Cryptococcus neoformans (MIC: 7.11-13.20 μM) [16]. However, peptides that have many amino acid residues end up having a restriction on their availability since the synthesis process of this molecules is laborious and expensive, with the costs being directly proportional to the size of the primary sequence. Thus, in order to obtain a smaller molecule while retaining antimicrobial activity, LyeTx I was minimized from the C-terminal region by sequentially removing amino acid residues up to the alanine at position 15 [17]. Additionally, this shortened derivative was changed by insertion of a lysine residue at position 5, resulting in a new peptide named LyeTx I mnΔK that contains only 16 amino acid residues (H-IWLTKALKFLGKNLGK-NH2). This compound, in addition to presenting a spectrum of activity very similar to that of LyeTx I [17], was able to act on both, planktonic cells and biofilms of Candida spp. [18] and carbapenem-resistant Acinetobacter baumannii (CRAB) [18]. Moreover, intranasal instillation of LyeTx I mnΔK (1-10 mg/Kg) was able to reduce the pulmonary bacterial load of animals with CRAB-induced lower airway infection, indicating its potential therapeutic use against superbug infections [19].
Considering the clinical relevance of MRSA infections and the limited therapeutic arsenal available against this pathogen, and taking into account the good antibacterial activity obtained in vitro and in vivo with LyeTx I mnΔK against MDR bacteria, the use of this compound to treat MRSA infections is promising. In this study, we investigated the therapeutic potential of the LyeTx I mnΔK for non-surgical wounds caused by MRSA in mice, and in addition, we employed biophysical studies to evaluate the membrane binding properties of this peptide, in the presence of S. aureus mimetic membranes.
2. Results
2.1. LyeTx I mn∆K Shows Good In Vitro Activity against MRSA
The antibacterial activity of LyeTx I mnΔK was evaluated by a broth microdilution assay against MRSA isolated from infected wounds of patients hospitalized in a tertiary hospital in southeast Brazil. As shown in Table 1, LyeTx I mn∆K inhibited the growth of all isolates tested, with MICs ranging from 8 to 32 μM, where the MIC required to inhibit 50% of MRSA isolates (MIC50) was 8 µM. The antimicrobial effect observed was mostly bactericidal, and it was shown that LyeTx I mn∆K could kill MRSA cells at concentrations ranging from 16 to 32 μM. Vancomycin, in turn, shows a MIC50 of 1 μM and MBCs between 2 and 8 μM (Table 1).
2.2. LyeTx I mn∆K Exhibits Rapid Bactericidal Effects on Cells in Logarithmic Growth of MRSA
The bactericidal action in function of time of LyeTx I mn∆K was assessed, comparing it with vancomycin, through death curves assay against MRSA USA300. The LyeTx I mn∆K demonstrated high bactericidal efficacy against MRSA cells, with complete reduction of the bacterial load within a short time period. This peptide was able to eliminate a high bacterial load (106 UFC/mL) of MRSA after only 1 hour of exposure at concentrations of 5x and 10x the MIC (Figure 1). At a concentration of 2x the MIC, LyeTx I mn∆K was able to completely reduce the bacterial load in just 3 hours. In contrast, the bactericidal effect of vancomycin was less potent. After 6 h of incubation, vancomycin produced approximately four-log reduction and required 24 h to completely eliminate MRSA cells. These results highlight the efficient and rapid ability of the LyeTx I mn∆K to kill MRSA.
2.3. LyeTx I mn∆K Reduces Preformed Biofilms of MRSA
MRSA has the ability to form biofilms on various surfaces and tissues, including in wounds, and these bacterial structures are particularly problematic because they make the infection chronic and difficult to treat [20]. As LyeTx I mn∆K showed good activity against planktonic MRSA cells, we evaluated whether this compound is also active against mature biofilms formed by this pathogen, which have low susceptibility to currently available antimicrobials.
According to the results presented in Figure 2A, the peptide demonstrated a greater ability to reduce biofilm when compared to vancomycin. LyeTx I mnΔK at 2x MIC (16 µM) was able to reduce mature biofilms to 32.28±1.62% of the biomass observed in nontreated cells, while vancomycin did not show any effect not even at the highest concentration tested. Furthermore, we studied the effect of LyeTx I mn∆K and vancomycin on inhibiting MRSA biofilm formation. The peptide reduces the biomass of biofilms formed by MRSA at concentrations of 1/4x MIC (2 µM; 39.28±13.41%) and 1/2x MIC (4 µM; 32.90±5.14%) at a level similar to that observed with the same concentrations of vancomycin (1/4x MIC: 33.93±2.07% and 1/2x MIC: 40.01±5.13%) (Figure 2B). Only at the lowest concentration tested did LyeTx I mn∆K show no significant effect, while vancomycin at 1/8x MIC (0.125 µM) reduces the biofilm formed to 70.43±26.09% when compared to untreated cells (99.91±14.02%).
2.4. Behavior of LyeTx I mnΔK after Combination with Conventional Antimicrobial Agents against MRSA
The combination of the LyeTx I mnΔK peptide with antimicrobials frequently used in the treatment of S. aureus infections (i.e., oxacillin and vancomycin) was studied by the synergism and re-sensitization assay. Initially, the synergism between the peptide and the conventional antimicrobials was evaluated against MRSA using a checkerboard test (Table 2). The combination of vancomycin and LyeTx I mnΔK resulted in a fractional inhibitory concentration index (FICI) of 1.50, indicating an indifferent effect. The combination of peptide with oxacillin showed a FICI of 1.02, which also indicates an indifferent effect. These findings were corroborating with the analysis by isobolograms, which show that the relationship between the FIC of the peptide and the FIC of vancomycin (Figure 3A) or oxacillin (Figure 3B) results in a straight line indicating an additive or indifferent effect.
Next, the MIC values of the antimicrobials (vancomycin and oxacillin) were determined after exposure of MRSA USA300 for 1h to a subinhibitory concentration of LyeTx I mnΔK. According to Table 3, the results showed that the previous exposition to peptide resulted in a significant increase in the sensitivity of bacteria to the antimicrobials vancomycin and oxacillin. The MIC of vancomycin and oxacillin decreased from 1.0 µg/mL to 0.5 µg/mg and 128 µg/mL to 64 µg/mL, respectively, resulting in a resensitization by two-fold to each antimicrobial agent.
2.5. LyeTx I mn∆K Induces Membranolytic Effect on MRSA Cells
To assess the ability of LyeTx I mn∆K to damage bacterial membranes the material with absorbance at 260 nm was quantified, suggesting the release of microbial genetic material into the medium after exposure to the compound. In accordance with Figure 4, the treatment of bacterial suspensions of MRSA USA300 with LyeTx I mn∆K at 10x MIC induced a marked time-dependent increase in the 260 nm-absorbance, suggesting that this peptide can disrupt the integrity of the bacterial membrane. In contrast, the activity of vancomycin was similar to the untreated control, indicating that this compound has a low membranolytic effect.
2.6. LyeTx I mn∆K Interacts with POPG:CL Membranes
Due to the membranolytic effect observed by the release of intracellular material absorbing at 260-nm, we studied the ability of the LyeTx I mnΔK peptide to interact with the S. aureus membrane using techniques that simulate the lipid bilayer of this microorganism. Thus, the membrane-mimetic interactions of S. aureus with LyeTx I mn∆K were investigated in aqueous solution and in the presence of membrane-mimetic environments using circular dichroism (CD) spectroscopy. Whereas the CD spectrum of the peptides in aqueous solution is indicative of a predominantly random coil conformation (Figure 5B), the stepwise addition of anionic POPG:CL (2:1) unilamellar phospholipid vesicles (LUVs) to 50 µM of LyeTx I mn∆K suspended in 10 mM Tris, 50 mM NaCl, pH 7.4 results in the appearance of pronounced minima around 208 and 222 nm.
The interaction between LyeTx I mn∆K and POPG:CL (2:1, mol/mol) LUVs was further investigated by isothermal titration calorimetry (ITC) (Figure 5A). This technique has been extensively used for the characterization of the apparent thermodynamic parameters of the interaction (∆G, ∆H, ∆S, the binding constant K, and the stoichiometric coefficient, n) of some compounds [22,23]. Titration was performed with the peptide solution in the calorimeter cell and the LUVs suspension in the syringe. Figure 5A presents the raw data for the titration of a 25 µM solution of LyeTx I mn∆K with 20 µM POPG:CL LUVs. The first injections of LUVs into the peptide solution shows an exothermic heat (~ -55 µcal/s) associated with the peptide-membrane interaction, which reaches a plateau when a LUVs/peptide molar ratio of 20 is reached. The enthalpy calculated for each injection of LUVs subtracted from the enthalpy of LUVs dilution is plotted as a function of the phospholipid/peptide ratio in Figure 5A.
The inflection point of the isotherm corresponds to the stoichiometric molar ratio (n). The n of approximately 12 indicates a supramolecular peptide-LUVs structure containing 12 lipids for each peptide molecule of the system under analysis. The binding isotherm was calculated by non-linear curve fitting using the model of equivalent binding sites (eq. 1) and the thermodynamic parameters for the peptide-membrane interaction are shown in Figure 5A. The peptide-membrane binding is driven by both enthalpic and entropic contributions, where the entropic term (588 cal/mol) is around 5 times greater than the enthalpic contribution (-122 cal/mol).
2.7. LyeTx I mn∆K Has Low Interaction with Artificial Vesicles That Mimicking Eukaryotic Membranes
The conformational behavior and lytic activity of LyeTx I mn∆K in the presence of mimetics of eukaryotic membranes were investigated by CD and calcein release, respectively. The LyeTx I mn∆K spectra reveals any defined minimal can be recorded in the presence of zwitterionic POPC LUVs containing 33% cholesterol (Chol) mol/mol (Figure 6).
The ability of the peptide to induce leakage from calcein-loaded LUVs made of POPC:Chol (2:1) was evaluated at 100 µM lipid concentration. The calcein-loaded LUVs with the addition of the peptides are presented in Figure 7A,B.
2.8. Effect of LysylPOPG on the Peptide-Membrane Interaction
In order to show the effect of the membrane-mimetic interactions of S. aureus with peptide upon the hydrodynamic diameter (Dh) and zeta potential (ζP) of POPG:CL (2:1) and POPG:CL:LysylPOPG (2:2:1) LUVs, time lapse dynamic light scattering (DLS) experiment was performed (Figure 8) [24,25,26]. The DLS experiments were carried out by incubating the respective LUVs suspension in the absence and presence of 5-75 μM LyeTx I mnΔK for 30 minutes. Figure 8A shows the changes in the Dh of vesicles upon addition of the peptide. The control vesicles were monodisperse with a Dh of 101±5 nm, while for POPG:CL:LysylPOPG a Dh of 107±4 nm was observed. Both LUVs incubated with peptide showed an increased hydrodynamic diameter in a concentration-dependent manner. The Dh slightly increase until 10 µM of peptide concentration, with a ∆Dh of 13 nm for the LUVs in absence of LysylPOPG and 7 nm for the LUVs containing LysylPOPG. Above this concentration, the Dh values ramp up with addition of the peptide in both lipid systems reaching around four and five times higher Dh than those observed for the control vesicles (410 ± 60 nm for POPG:CL and 515 ± 15 nm for POPG:CL:LysylPOPG).
The electrophoretic mobility of LUVs in the presence of LyeTx I mn∆K was measured to evaluate the effect of the peptide on the zeta potential (ζP) of both lipid systems. Figure 8B presents the ζ-potential experiments in which LUV suspensions were titrated with solutions of LyeTx I mn∆K. The initial values of ζP are near −41 mV and −26 mV for POPG:CL and POPG:CL:LysylPOPG LUVs, respectively. The addition of peptide led to a continuous increase of the ζP of the anionic vesicles until a peptide concentration of 20 µM in the presence of POPG:CL and 40 µM in the presence of POPG:CL:LysylPOPG LUVs. From these values a plateau is reached with a change of ζP around − 30 mV and -25 mV in the presence of the POPG:CL and POPG:CL:LysylPOPG LUVs, respectively.
The ability of LyeTx I mn∆K to induce leakage on calcein-loaded LUVs was studied on POPG:CL (2:1) or POPG:CL:LysylPOPG (2:2:1), both at 100 µM of lipid concentration. The results of the leakage measurements of the calcein-loaded LUV with the addition of LyeTx I mn∆K at 35◦C are presented in Figure 9. The calcein leakage from both LUVs is virtually negligible in the absence of the peptide (first 5 minutes). Calcein release is immediately observed after the LyeTx I mn∆K is added (Figure 9A-B). Kinetics of the calcein release were recorded for 15 min and the maximum effect of the peptide on both LUVs is observed around after 6 min in POPG:CL and 9 min in POPG:CL:LysylPOPG, after which the disrupting effect was stable and did not increase with time. The addition of the peptide in increasing concentrations to the dye-containing LUV induced calcein release in both systems in a concentration-dependent manner. The rate of calcein leakage from the vesicles was calculated by the increase of fluorescence emission as a function of time. The data were analyzed as a first-order process and a rate constant, Kobs, was calculated for each run.
The kinetics of calcein release from POPG:CL and POPG:CL:LysylPOPG LUVs are presented in Figure 9C and the rate constants are shown in Table 4. For both LUVs, increments in the LyeTx I mn∆K concentration lead to a linear increase in the rate of calcein leakage (kobs). The slope for the plot of the kobs vs. peptide concentration for POPG:CL:LysylPOPG LUVs is slightly lower than the value obtained for POPG:CL LUVs, indicating a lower kinetics of calcein release. The percentage of calcein release (% calcein release), after 15 min, was determined by using Eq. (2) (see Section 4.10) and the values were plotted as a function of peptide concentration (Figure 9D) in the presence of either POPG:CL or POPG:CL:LysylPOPG lipid systems. In the presence of 4 µM LyeTx I mn∆K the disrupting effect was higher for POPG:CL LUVs (28.5%±2.5) when compared to 16.0±1.0% for POPG:CL:LysylPOPG. Nevertheless, the disrupting effect of the peptide was similar in both vesicles at the highest (32 µM) LyeTx I mn∆K molar concentration assayed; the leakage was 82.2±3.0% for POPG:CL and 77.2±2.0% for POPG:CL:LysylPOPG vesicles.
In order to evaluate the lytic activity of LyeTx I mn∆K in the eukariotic membranes, similar leakage experiments were also carried out in the presence of calcein-loaded LUVs composed by POPC:Chol (2:1) (Figure S1). Notably, the calcein leakage from zwitterionic LUVs is at least 8 times lower than the values observed in the presence of both anionic LUVs. Whereas the calcein leakage from POPG LUVs is around 80%, the maximum value reached in the presence of POPC:Chol LUVs is less than 10%.
2.9. Gel Containing LyeTx I mnΔK is Effective in MRSA-Induced Wounds in Mice
Due to the relevant antibacterial activity of the LyeTx I mnΔK peptide in vitro, we evaluated the use of a gel formulated with this active compound in the treatment of wounds induced by MRSA in vivo. Thus, to assess the potential of LyeTx I mnΔK as a topical pharmacological agent, mice were infected with S. aureus MRSA USA 300 by intradermal injection allowing the formation of an open wound/abscess. After treatment, a significant reduction in the bacterial load from lesions was observed in mice exposed to LyeTx I mnΔK when compared to the untreated groups (6.66±0.71 Log10 CFU/g of wound) or those treated with base gel (6.25±0.67 Log10 CFU/g of wound) (p-value <0.05; Figure 10). Mice treated with gel containing 0.50% LyeTx I mnΔK showed the greatest reduction in bacterial load (3.18±2.64 Log10 CFU/g of wound), followed by those that received 0.25% LyeTx I mnΔK (3.81±1.90 Log10 CFU/g of wound) and 1.00% LyeTx I mnΔK (3.93±0.85 Log10 CFU/g of wound) (Figure 10). Therefore, the effect of the peptide in vivo did not show dose-dependent behavior. In turn, the gel containing 1% vancomycin reduced only a little the bacterial load of the wound (4.78±0.52 Log10 CFU/g of wound) when compared to the untreated group, revealing that the treatment with LyeTx I mnΔK, even at the lowest concentration tested (0.25%), was more effective than the vancomycin. These results corroborate the finding that LyeTx I mnΔK is a promising molecule for topical administration and is, thus, superior to reference drugs.
Corroborating these findings, the animals that received the gel containing the peptide at 0.50% showed a significantly lower weight loss compared to untreated animals (Figure 11). This result suggests that the discomfort of the wound, which leads to less water and food intake with consequent weight loss in animals, was reduced after using of formulation contained the peptide.
3. Discussion
Several studies indicate that the spread of infections caused by MRSA is a major public health challenge worldwide [27]. In this sense, the World Health Organization (WHO) categorized this superbug as a high priority microorganism for the search and development of new antimicrobials [27], and AMPs stand out as relevant agents in this scenario. Here, we showed that a synthetic peptide inspired by a compound isolated from the venom of the L. erythrognatha spider, called LyeTx I mnΔK, has good activity against planktonic cells and biofilms of MRSA, in addition to being useful in the treatment of wounds caused by this superbug.
The peptide LyeTx I mnΔK showed remarkable antibacterial activity against the MRSA isolates tested, having a pronounced bactericidal action at low concentrations and with rapid maximum effect. In the treatment of infections, it is important that the antimicrobial agent acts quickly and can completely eradicate the microorganism to prevent the development of microbial resistance and complications of infection [19,21]. In this regard, the predominantly bactericidal activity of peptide has shown promise, especially in highly resistant microorganisms such as MRSA. Despite the rapid bactericidal effect of LyeTx I mnΔK, vancomycin exhibits a very slow antibacterial activity, which may compromise its efficacy. Clinical use of vancomycin often leads to therapeutic failure due to its limited bactericidal capacity [28], which makes LyeTx I mnΔK advantageous because of its efficient microbicidal property.
In addition to its potential to acquire antimicrobial resistance genes, S. aureus exhibits another virulence factors as its ability to form biofilms. Bacterial biofilms are clusters of bacteria that are attached to a surface and/or to each other and embedded in a self-produced exopolymer matrix that protect them from adverse conditions such as sanitizers and antimicrobials agents [29]. When MRSA-formed biofilms are established in wounds, they exhibit resistance to destruction by conventional antimicrobials, which involves a high risk of inflammation and/or purulence [30]. Therefore, the activity of the LyeTx I mnΔK peptide was evaluated against MRSA biofilms and the results revealed that this compound has a promising anti-biofilm effect. These results were similar to those observed by Lima et al. (2021) who showed that LyeTx I mnΔK has the ability to disrupt mature biofilms and inhibit the formation of this bacterial structure in carbapenem-resistant Acinetobacter baumannii cells [19]. It is worth noting that the effect of the peptide on mature biofilms formed by MRSA was superior to that observed by vancomycin. Conventional antimicrobials have a low ability to act on biofilms because, in addition to the exopolymeric matrix that physically protects bacterial cells from interaction with these compounds, the microorganisms within the biofilm are in a stationary phase of growth [31]. As all available antibacterial agents have activity only against cells in the logarithmic growth phase, bacteria within the biofilm are naturally insensitive to the action of these drugs [32,33]. Therefore, the present result shows that the LyeTx I mnΔK peptide, different from vancomycin, can act in both phases of microbial growth, i.e., logarithmic and stationary.
A rapid bactericidal effect and the activity on bacterial biofilms is many times associated with the ability of compound to damage the bacterial cell membrane, which can result in lysis. In fact, the LyeTx I mnΔK peptide significantly increases the loss of genetic material from MRSA cells as indicated by the release of 260-nm-absorbing intracellular compounds, showing that this AMP damages the bacterial plasma membrane. The characteristic of AMPs to cause membrane damage, associated with a rapid and instantaneous bactericidal action, has been highlighted by several authors such as Brogden (2005) [11] and Avelar (2015) [34]. In addition to being associated with a rapid antimicrobial effect, the action of LyeTx I mnΔK on the MRSA membrane may come with other advantages such as the reduced potential for inducing resistance and low doses required to clinical use. In this context, it is widely known that compounds that act on plasma membrane have a low capacity to induce antimicrobial resistance because membrane redesign by bacteria would be a “costly” solution for most microbial species [35,36]. Moreover, due to the low MIC values related to these antimicrobial agents, the concentrations normally required for clinical use are low, which reduces the possibility of adverse events related to this medication [10]. Vancomycin, in turn, did not alter the release of materials with absorption at 260 nm, which suggests a discrete effect on the MRSA membrane. This glycopeptide acts by binding to the end of the cross-links that form the bacterial wall (specifically, to the D-Ala-D-Ala residues) [37], therefore, it does not interfere with the bacterial lipid membrane, which is confirmed by our results.
In order to comprehend better the mechanism of action of LyeTx I mn∆K on MRSA membranes, we studied the peptide-membrane interaction in the presence of POPG:CL (2:1, mol/mol). This phospholipid composition was selected as it represnets the predominant constituents of S. aureus membranes [38,39,40]. Initially, the thermodynamic studies regarding the membrane interactions of LyeTx ImnΔK with POPG:CL LUVs indicated that the peptide presents from moderate to high affinity to negatively charged vesicles at 35 ◦C. Although, the negative enthalpy values indicate exothermic binding of the peptides to the anionic vesicles and the predominance of attractive Coulomb forces, the binding process for the peptide is mainly driven by entropic factors, since |TΔS| >> |ΔH|. This interaction should occur by partial peptide incorporation in the bilayers interface, as it has been shown the increase of the vesicle size observed by the DLS experiments [41]. Interestingly, above this critical concentration the PDI values ramp up (>0,3) and two size populations were noted. This suggests that the peptide adsorption to the membranes is followed by vesicle aggregation at high peptide concentration (>20 μM) [42]. Notably, the peptide concentration dependence is also observed in the ζ-potencial measurements. Initially, the interaction of the positively charged peptide with the negatively charged bilayers results in a linear increase of ΔP, reaching a plateau between 15 – 20 μM peptide.
Regarding the secondary structure of the peptide, our results show α-helical structures when in contact with membrane surface of POPG:CL [43]. LyeTx I mn∆K presents a significant affinity to both phospholipid vesicles, with an association constant (5,9 x 103 M-1) that falls within the range of 103 – 104 M-1 [41,44]. However, in the presence of POPC:Chol (2:1) lipid bilayers, mimicking eukaryotic membranes, the LyeTx I mn∆K spectra reveal no defined conformation. These results suggest that shortened form presents lower affinity to the mimetics of eukaryotic membranes. The same can be observed with the calcein release, which the LyeTx I mn∆K reached a level of less than 20% in vesicles that simulate mammalian cells compared to membranes related to the envelope of S. aureus. Corroborating these findings, previous studies have shown that the LyeTx I mnΔK peptide has low toxicity against eukariotic cells. The high concentration required to kill 50% (CC50) of Lund human mesencephalic (LUHMES; CC50 15.02 µM) [17] and african green monkey kidney epithelium (Vero; CC50 55.31±5.0 µM) [19] cells in culture indicate that this peptide preferentially binds to bacterial cells. Furthermore, LyeTx I mnΔK has low hemolytic activity, with a cytotoxic concentration for 50% of erythrocytes ranging from 44.67 µM to 77.07 µM [17,19].
Lysyl phosphatidylglycerol (LysylPOPG) is commonly found in the multidrug-resistant Staphylococcus aureus such as MRSA by modification of POPG. This process is driven by multiple peptide resistance factors (MprF), which modifies the phopholipids with positively charged lysine residues in the cytoplasmic membrane leaflet and translocate the LysylPOPG to the outer cytoplasmic membrane [45]. This modification has shown resistance to cationic antimicrobial peptides such as colistin, nisin, human-β-defensin 3, and polymyxin B.
Therefore, to explore the effect of the addition of LysylPOPG lipid in membrane permeabilization ability of the LyeTx I mn∆K, we evaluate the Dh, ζP and calcein release from LUVs containing LysylPOPG. Although the addition of LysylPOPG decreases the kinetics (kobs) of lytic activity for LyeTx I mn∆K, this lipid does not affect the maximum release of calcein caused by the peptide. The effect the peptide was also virtually equivalent to the changes of ζP of the LUVs in the absence and in the presence of LysylPOPG (ζP ~ − 30 mV for POPG:CL and -25 mV for POPG:CL:LysylPOPG). The polydispersity index (PDI) of the suspensions increased, suggesting that peptide-membrane interaction affects the originally homogeneous vesicles resulting in a deformed lipid-peptide complex. Interestingly, two populations can be observed with increasing of peptide concentration, one centered around 100 nm and the other around 1000 nm (Figure S1). These findings suggest a fusion process as compared to aggregation which results in the polydispersity of the size population [46,47]. Similarly, the effect of the peptide in the Dh of both LUVs was comparable, resulting in vesicle agglutination. These findings are in accordance with the antimicrobial assays, which have demonstrated high activity of LyeTx I mn∆K even against the MRSA USA300 strains (MIC50 and CBM50 of 8 µM and 32 µM, respectively). Thus the presence of LysylPOPG which constitutes part of the adaptation of MRSA to antibiotic treatments is surpassed by LyeTx I mn∆K, indicating its effectiveness even in challenging conditions.
One of the most commonly used strategies for treating infections caused by MDR bacteria, such as MRSA, is the combination of different antimicrobial agents [48]. This therapeutic protocol has several advantages such as rapid antibacterial effect, lower possibility of therapeutic failure due to selection of resistant bacteria, reduced concentration of each antimicrobial used in the combination and lower risk of serious adverse reactions associated with high doses of the drug [49]. LyeTx I mn∆K is indifferent to the activity of the antimicrobial’s oxacillin and vancomycin. This indicates that both compounds can be combined to broaden the spectrum of antimicrobial action, thus helping to treat polymicrobial infections [36]. Furthermore, prior exposure to LyeTx I mn∆K reduces the active concentrations of these two antimicrobials, which may generate a benefit in reducing the dose of these agents with a consequent lower frequency of adverse reactions [49].
Clinical manifestations of MRSA range of auto-limited superficial infection to life-challenging bacteremia, however the wounds are still most commonly problem caused by this pathogen [50]. MRSA-infected wounds are difficult to heal, purulent, painful and treatment is restricted to a few options such as vancomycin, mupirocin and clindamycin [51]. Here, we show that a formulated gel containing LyeTx I mnΔK was effective in treating MRSA-induced non-surgical wounds in mice, reducing bacterial load in a non-dose-dependent manner. Interestingly, this effect was superior to that of the reference drug (vancomycin). Due to their amphoteric nature, AMPs are able to interact better with the corneal extract of the epidermis. This characteristic facilitates absorption through the skin, allowing them to reach deeper layers of the dermis, including infection sites. Thus, the LyeTx I mnΔK highlighted as a promising alternative for the treatment of cutaneous infections by MRSA. On the other hand, vancomycin, being a hydrophilic molecule [52], faces limitations in penetrating through the skin. This characteristic hinders its access to deeper layers of the dermis, compromising its effectiveness in treating deep skin infections.
4. Materials and Methods
4.1. Microorganisms
The antibacterial activity was evaluated against six clinical isolates of methicillin-resistant Staphylococcus aureus (MRSA). All isolates were recovered from wounds of patients with acute infections admitted to Santa Casa de Belo Horizonte, been the procedure approved by the ethics committee for human subjects of Group Santa Casa BH (CAAE: 09187719.4.0000.5138). Bacterial identification was performed by automated biochemical-physiological methods (BD Phoenix®; Frankfurt, Germany) and MRSA phenotype was confirmed by disk-diffusion assay using cefoxitin [53]. The reference strain S. aureus MRSA USA300 was kindly provided by the Reference Laboratory in Microbiology at the Oswaldo Cruz Foundation (FIOCRUZ-RJ, Brazil).
4.2. Reagents
Vancomycin (Inlab, São Paulo, SP, Brazil), dimethyl sulphoxide (DMSO), sodium chloride (NaCl), ethylic alcohol, glacial acetic acid, violet crystal (Synth, São Paulo, SP, Brazil), and oxacillin (Sigma-Aldrich, San Louis, USA) were all purchased from commercial suppliers. The peptide LyeTx mnΔK was obtained by solid phases synthesis and acquired from GL Biochem (Shangai, China) with a purity of 99%, which was confirmed by spectrometric analysis. Mueller–Hinton broth, Mueller-Hinton agar, Mannitol agar and trypticase soy broth (TSB) were purchased from Kasvi (São José do Pinhais, PR, Brazil).
4.3. Antimicrobial Activity
4.3.1. Preparation of the Inoculum
The bacterial inoculum used in the susceptibility test was standardized according to the Clinical Laboratory and Standard Institute (CLSI) M07 document (CLSI, 2018). Initially, bacterial streaks were performed on Mueller-Hinton agar. Subsequently, three to five isolated colonies from the S. aureus cultures were selected and transferred to tubes containing sterile saline solution (0.9% NaCl). The resulting suspension was adjusted to the 0.5 McFarland scale (correspondent to 108 CFU/mL) using a spectrophotometer (Shimadzu UVmini-1240 Spectrophotometer UV/Vis Reader). Finally, 50 μL of this suspension was transferred to 10 mL of Mueller-Hinton broth, resulting in a final inoculum at 106 CFU/mL, which was employed in antimicrobial tests.
4.3.2. Determination of Minimum Inhibitory Concentration (MIC)
Mueller-Hinton broth [19] was used to determination of minimum inhibitory concentration (MIC) by broth microdilution method, according the CLSI document M07 for aerobically growing bacteria. The compounds (LyeTx I mnΔK and vancomycin) were dissolved in sterile water and serially diluted two-fold ranging from 0.5–128 µM. Finally, the results were visualized after 24 h of incubation at 37 °C and the MIC value was considered as the lowest concentration of tested compound able to prevent visible bacterial growth. Vancomycin and wells with inoculum and culture medium were included as positive and negative controls, respectively. Furthermore, wells containing only culture medium were included as a control for medium sterility, all experiments were performed in triplicate.
4.3.3. Determination of Minimum Bactericidal Concentration (MBC)
The minimum bactericidal concentration (MBC) was determined by transferring 100 µL from the wells optically free of growth of MIC assay to plates of Mueller-Hinton agar plates. After 24 hours of incubation at 37 °C the MBC was considered the lowest concentration of the tested agent capable of reduce 99% of colonies in relation to negative control. Vancomycin was used as positive control, and the experiment was realized in three replicates.
4.4. Time-Kill Curve
The peptide activity as a function of time was studied by death curve assay. Initially, an inoculum of MRSA USA300 at 106 UFC/mL was prepared conformed to item 4.3.1. Bacteria were then challenged with LyeTx I mn∆K at the 2x, 5x, and 10x MIC. Untreated cells and vancomycin at 10x MIC were used as negative and positive control, respectively. The tubes were then incubated at 37°C with aeration at 225 rpm. At intervals of 0, 30, 60, 120, 150 and 180 min, 100 μL of this bacterial suspension was serially diluted (10-1 to 10-5) in sterile saline (0.9% NaCl) and plated onto mannitol agar. The plates were finally incubated for 24 h and the CFU/mL of morphotypes colonies of S. aureus on mannitol agar (i.e., lysis, medium and yellow colonies) was calculated [29].
4.5. Anti-Biofilm Activity
To assess the effect of the LyeTx I mn∆K on pre-formed biofilm of MRSA USA300, as well as on the formation of biofilm by this bacterium, the methodology described by Herrera et al. (2020) was utilized [54]. Mueller-Hinton broth supplemented with glucose 1% (MHBg) and a bacterial suspension prepared according to item 4.3.1. were used. In the biofilm formation inhibition assay, bacterial suspension in MHBg was incubated at 37° C for 24 h with the compounds (LyeTx I mnΔK and vancomycin) at concentrations below the MIC (1/8x, ¼x, 1/2x MIC), and inoculum without treatment was considered a control. Next, the medium was removed, and the microtiter plates washed three times with saline (NaCl 0.9% w/v). In the established biofilm disruption assay, bacterial suspension in MHBg was incubated at 37°C for 24 h to enable biofilm formation. After removal of the medium, the microtiter plates were washed tree times with saline and the biofilm attached to the wells treated with the compounds (LyeTx I mnΔK and vancomycin) at 1x, 2x and 4x MIC. Bacterial suspension without treatment was considered a control (untreated group). The microtiter plates were incubated at 37°C for 24 h, the medium was removed and next washed three times with saline.
In both assays, the biofilm was fixed in wells with 100 µL methanol for 30 min at 37oC. Next, the wells were washed with saline (three times) and biofilms were stained with 1% w/v crystal violet for 30 min at room temperature. Finally, the wells were washed with saline (three times), air-dried by 2 h, and the biofilm masses solubilized in 30% glacial acetic acid. The readings were taken on a microplate reader (Bio-Tek Instruments, Winooski, VT) at 600 nm. The biofilm biomass percentages in treated groups were calculated by comparing their absorbance values with those of untreated wells (100% of biomass). Results of biofilm assays were presented graphically using the arithmetic mean±standard deviation of six independent experiments.
4.6. Membranolytic Effect
In this study, the effect of compounds (LyeTx I mn∆K and vancomycin) on membrane of MRSA USA300 was evaluated through of the release of intracellular material with absorbance at 260 nm [36]. Tubes containing 5 mL of a suspension of MRSA USA 300 at 108 CFU/mL in saline, prepared according to item 3.4.1, were exposed to the LyeTx I mn∆K or vancomycin at 10x the MIC of each one. After incubation at 37oC, a 500 µL sample of tubes was removed at different time intervals (0, 0.5, 1, 3, 6 and 24 h), centrifuged (1500 g for 25 min at 4 oC) and the supernatant removed to measure the optical density at 260 nm in a spectrophotometer (Bio-Tek Instruments, Winooski, VT). Untreated cells were included as controls and the results were expressed graphically as OD260nm vs. time (hours).
4.7. Combination Assays
4.7.1. Synergism
The interaction of LyeTx I mnΔK with vancomycin and oxacillin against MRSA USA300 was evaluated by the checkerboard assay as described previously [55]. S. aureus suspensions (106 CFU/mL) were incubated with the combination of LyeTx I mnΔK and antimicrobials (0.05–40 μg/mL) at ratio of 1:1 for 24 h at 37 °C. Next, the fractional inhibitory concentration index (FICI) was calculated as the sum of the FICs of the peptide and the FIC of the antimicrobials, which are defined by the following equation:
The results were interpreted according to Oroojalian et al. (2002) [56], which considered synergic effect if FICI ≤ 0.5, additive effects to 0.5 < FICI ≤ 1.0, indifferent activity if 1.0 < FICI ≤ 4., and antagonist effect to FICI > 4.0. In addition, the results were interpreted by plotting the FIC values of each compound involved in the combinations. Thus, a graph called isobologram is generated, where agents are considered synergistic when the orthogonal projection is concave, additive or indifferent if the projection behaves like a straight line, and antagonistic in cases where the projection takes a convex form [36].
4.7.2. Resensitization
The effect of previous exposure of MRSA USA300 to LyeTx I mnΔK on MIC values of antimicrobial agents (oxacillin and vancomycin) were performed as described previously [57], with modifications. Briefly, 1/4× MIC of LyeTx I mnΔK was incubated with the bacterial suspension (106 CFU/mL) in MHB at room temperature for 60 min. After incubation, the tube was centrifugated at 1500g for 25 min at 4 oC, the supernatant was removed e 10 mL of sterilized MHB was newly added in tubes. Next, peptide-treated bacteria were added to 96-well plates and the MIC of oxacillin and vancomycin was determined. Bacteria peptide-untreated served as negative control. The result was expressed as the fold-change of resensitization, calculated by the ratio of the MIC of antimicrobial alone and the MIC of the antimicrobial after resensitization with 1/4× MIC of LyeTx I mnΔK.
4.8. Preparation of Large Unilamellar Vesicles (LUVS)
LUVs were prepared using the phospholipids, 1-palmitoyl-2-oleoyl-sn-glycero-3-phospho-(1′-rac-glycerol) (POPG), diphosphatidylglycerol (Cardiolipin - CL) and 1,2-dioleoyl-sn-glycero-3-[phospho-rac-(3-lysyl(1-glycerol)) (LysylPOPG). The appropriate amount of phospholipid in a molar ratio of POPG:CL 2:1 or POPG:CL:LysylPOPG 2:2:1 was transferred to a glass tube and suspended with 1 mg per mL of chloroform at room temperature. The organic solvent was removed in a rotary evaporator to form a lipid film. Multilamellar vesicles (MLVs) were prepared according to the freeze thawing method in aqueous buffer (10 mM Tris-HCl, 50 mM NaCl, pH 7.4), eventually containing calcein (50 mM) at 25 °C. The calcein buffer was prepared by diluting 545 mg of calcein in 2,5 ml of 1 M NaOH, followed by addition of 1.25 ml of buffer (100 mM Tris-HCl, 50 mM NaCl, pH 7.4) was added, together with 2,5 ml of Milli-Q water (Millipore, Bedford, MA). The solution was then pH-adjusted to 7.4 by dropwise addition of 1 M HCl while being stirred constantly to avoid precipitation. Milli-Q water was then added to bring the calcein buffer to a final volume of 12,5 ml and the calcein concentration to 50 mM. The resulting MLVs were subjected to eight freeze/thaw cycles using liquid nitrogen and a water bath at 35 °C (five cycles). The MLVs were extruded in a 10 mL stainless steel extruder (Lipex Biomembranes Inc., Vancouver, Canada) at 25 °C to obtain LUVs of 100 nm. For calcein release experiments the dye-containing vesicles were separated from non-entrapped calcein using a size exclusion chromatographic column (Sephadex G50 Fine, 10 ×150 mm), eluted with 10 mM Tris-HCl buffer that did not contain calcein. The total lipid concentration of the extruded LUVs was estimated by a colorimetric method as described by Stewart [58].
4.9. Isothermal Titration Calorimetry (ITC)
ITC analyses were performed in a Malvern® VP-ITC microcalorimeter (Worcestershire, United Kingdom), at 25 °C. The calibration of the equipment was performed with deionized water Milli-Q® type 1. The solutions were previously degassed in a Microcal Thermovac® (Worcestershire, Kingdom) accessory from Malvern®. Each experiment consisted of 25 successive injections of 5 μL of 20 mM POPG:CL (2:1, mol/mol) LUVs into the calorimetric cell filled with 1.400 mL of peptide solution (25 μM), at 300 s intervals. To eliminate the diffusion effects of material from the syringe to the calorimetric cell, the first injection of 1 μL was discarded. Dilution experiments of LUVs were performed by injections of the samples in 10 mM Tris-HCl, 50 mM NaCl, pH 7.4. The acquisition and isothermal treatments for each analysis were performed using the Microcal Origin 7.0 software for ITC. Non-linear fitting was performed by using the model of one-site binding based on the Wiseman Isotherm [22] (equation 1) to obtain the partial molar enthalpy of complexation (= dQ/d[X]) at constant pressure.
where X is the molar ratio of titrant and M (XR = [X]t/[M]t) molar ratio of titrated at any point during the titration. The r parameter is the composition variable (r = 1/[M]t.Kapp) and the ∆intH°, V0 and Keq parameters are the standard complexation enthalpy, effective volume of the solution in the titration cell and apparent equilibrium constant, respectively.
4.10. Dynamic Light Scattering (DLS)
The changes of hydrodynamic diameter (Dh) and zeta potential (ζP) of POPG:CL (2:1, mol/mol) LUVs and or POPG:CL:LysylPOPG (2:2:1) with addition of the peptide solutions were measured at 35 ºC on a Z98 Zeta sizer nano ZS Malvern® model BI-900 particle analyzer (Worcestershire, UK). The light intensity of the monochromatic laser (4 mW Ne laser, λ of 633 nm) scattered was detected at a 90° angle. The experiments were collected in triplicate on independent experiments using 10 samples of 100 μM LUVs solution containing different peptide concentration with a final volume of 800 μL. For all samples were used 1 mM peptide and 400 μM LUVs stock solution, both suspended in 10 mM Tris-HCl, 50 mM NaCl, pH 7.4. Samples compositions are detailed in Table 5. The samples were incubated at 35 °C for 30 minutes before reading the hydrodynamic diameter and zeta potential.
4.11. Calcein Release
Calcein release experiments were carried in polystyrene microplates (128 × 86 × 14.5 mm) for fluorescence emission measurement in media containing LUVs suspension (100 μM) of POPG:CL (2:1), POPG:CL:LysylPOPG (2:2:1) or POPC:Chol (2:1), different volumes of peptide (250 μM) and buffer (10 mM Tris-HCl, 50 mM NaCl, pH 7.4) solutions to a final volume of 300 μL as detailed in Table 6.
The increase of CF fluorescence as a function of time at 25 °C was recorded every 1 minute in the Spectra Max®Paradigm detection platform (Molecular Devices, LLC, Sunnyvale, US) at excitation and emission wavelengths of 480 and 520 nm, respectively. The stability of the LUVs was monitored for 5 min before the peptide was added. The calcein release after the addition of the peptide was recorded for 15 min. The total calcein released from the LUVs (100%) was determined in a similar experiment using 0.1% Triton-X 100 in the absence of the peptide. Calcein leakage was calculated using equation 2:
where, F0, F, FT denotes the basal fluorescence intensity, florescence intensity after addition of peptides and maximum florescence intensity obtained after addition of 0.1 % Triton X-100, respectively. Tris-HCl buffer was used as negative control. The experiments were performed in triplicate on independent samples of calcein-loaded LUVs. The results are presented as the average with their standard deviations. The observed rate constant of calcein release, kobs, was calculated by using a single-exponential fitting of the recorded data of fluorescence intensity as a function of time [41].
4.12. Peptide Formulation
The formulation used for the experiment was prepared according to the Brazilian Pharmacopoeia 6th edition [59]. It consisted of a gel prepared by dissolving methylparaben (Nipagin®) in water heated to 70 °C. Then, under constant stirring on a hotplate with a magnetic stirrer hydroxyethylcellulose (Natrosol®) was added to form the gel. The process involved alternating stirring with resting periods, and the resulting gel had colorless characteristics and a pH of 6. Additionally, sodium metabisulfite was incorporated as an antioxidant to protect peptides from degradation by oxidation. The components and quantities used are described in Table 7.
4.13. In Vivo assays
4.13.1. Animals
Six-week-old male BALB/c mice (Biotério Central da UFMG, Belo Horizonte, MG, Brazil) were used in this study. The animals were kept in polypropylene boxes measuring 30x19x13 cm in an environment with a controlled temperature of 25±2°C, 40% humidity, and a 12/12 h light/dark cycle in Bioterio of Faculdade de Saúde Santa Casa de Belo Horizonte. All experimental procedures strictly followed the international protocols for laboratory animal management, and the methods were approved by the Laboratory Animal Research Ethics Committee of the Faculdade de Saúde Santa Casa de Belo Horizonte (CEUA-Santa Casa: 001-2023).
4.13.2. Murine Model of Non-Surgical MRSA-Infected Wounds
An in vivo model of MRSA USA300 skin infection was conducted according to Lima et al. [36]. Following the induction of general anesthesia (60 mg/kg ketamine + 8 mg/kg xylazine, i.p.), the dorsal hair was removed, and the skin was cleaned with 70% (v/v) ethanol. Inside the biological security cabin (Veco® Bioseg 18, Campinas, SP, Brazil), 50 μL of a bacterial suspension at 108 CFU/mL was injected subcutaneously into the dorsal area of the animals. After infection, the animals were placed in previously sterilized polypropylene boxes and kept in this environment with access to autoclaved water and feed ad libitum. After 48 h, an open wound/abscess was observed at the injection site.
4.13.3. Treatment of Animals
After MRSA-induced open wound/abscess, the animals were divided into six experimental groups (n = 5). These groups were treated topically with the previously formulated gel containing different concentrations of LyeTx I mnΔK (0.25%, 0.50%, or 1.00%). The positive control received 1.00% vancomycin-containing gel. Groups of animals treated topically with saline (untreated control) and base gel (base control) were also included. All animals were treated for three consecutive days with two daily administrations. Twente hours after the last treatment, the animals were euthanized, the wounds were aseptically removed, weighted, homogenized in saline, and plated on mannitol agar to determine the bacterial load.
4.14. Statistical Analysis
For the interpretation of the results statistical analyses were conducted. Initially, the normality of the data was assessed using the Shapiro-Wilk test in SPSS Statistics v. 19 software. If the data does not follow a normal distribution, the histogram, median, standard deviation, and kurtosis were used to check the normality of the data. Unidirectional analysis of (One-way ANOVA) was used. Tukey’s post-test was selected for comparisons between all groups, and Dunnett’s test was used for comparisons with the control group (untreated). Results were expressed as mean±standard deviation and p-value less that 0.05 was considered to statistic significancy.
5. Conclusions
The synthetic peptide LyeTx I mnΔK emerges as a potent antimicrobial agent with notable efficacy against methicillin-resistant Staphylococcus aureus (MRSA). There is a rapid bactericidal action, significant anti-biofilm effects, and membrane damaging mechanism making it a promising candidate for the urgent challenge of MRSA infections, besides showing low apparent toxicity to cells of mammals. The peptide demonstrates superiority activity over conventional antibiotics like vancomycin and oxacillin, particularly in eradicating MRSA biofilms and action on bacterial membranes. The LyeTx I mnΔK maintains effectiveness even mimetic membrane with lysylphosphatidylglycerol (LysylPOPG) modifications, showing its potential in challenging conditions. Furthermore, its indifference in combination with conventional antibiotics and higher resensibilization suggests a potential role in combination therapy, offering a multifaceted approach to combatting MRSA. The peptide’s application in a formulated gel proves effective in treating MRSA-induced non-surgical mice, unparalleled by the performance of vancomycin. These findings underscore LyeTx I mnΔK as a promising antimicrobial agent for MRSA infections, emphasizing its multifaceted efficacy, low resistance induction, specificity and potential for combination therapy.
6. Patents
LIMA, W.G.; BRITO, J.C.M.; COELHO, A.P.G.; RESENDE, J.M.; VERLY, R.M.; LIMA, M. E. Formulação para aplicação tópica de peptídeo antimicrobiano e seu uso para produzir medicamentos para tratamento de infecções dermatológicas. 2023, Brasil. Patente: Privilégio de Inovação. Número do registro: BR13202302645, Instituição de registro: INPI - Instituto Nacional da Propriedade Industrial. Depósito: 15/12/2023
Author Contributions
Conceptualization; methodology, validation, formal analysis: M.E d L., R.M.V, W.G.L, M.C.M.B, J.M.R, B.B; software: W.G.L.; investigation, A.P.G.C, A.N.S, W.G.L.; resources, supervision; project administration; fundraising: M.E.d.L., R.M.V, J.M.R, W.G.L., B.B. Writing—original draft preparation, W.G.L P.G.C., ANS; writing—review and editing, W.G.L., R.M.V., J.C.M.B., A.N.S., M.E d Lima; Methodology: S.O.A.G, L.P.B.C, D.C.S, L.V.R.G. All authors have read and agreed to the published version of the manuscript.
Funding
Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) [Chamada CNPq 25/2021 - Pós-Doutorado Júnior - PDJ 2021]; [Chamada CNPq nº 26/2021 - Doutorado-Sanduíche no Exterior (SWE) 2022/2023]; CNPq [304337/2019-6] - Chamada CNPq 06/2019 - Bolsas de Produtividade em Pesquisa]; FAPEMIG (Fundação de auxílio`a Pesquisa do Estado de Minas Gerais), Centre national de la recherche scientifique (CNRS); Université de Strasbourg the Agence Nationale de la Recherche (projects Naturalarsenal 19-AMRB-0004-02, AmphiPep 20-CE18-0021, SAFEST 21-CE18-0043, and the LabEx Chemistry of Complex Systems 10-LABX-0026_CSC), the Foundation Jean-Marie Lehn / Interdisciplinary Thematic Institute SysChem / IdEx Unistra (ANR-10-IDEX-0002).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Ethics committee for human subjects of Group Santa Casa BH (CAAE: 09187719.4.0000.5138). Laboratory Animal Research Ethics Committee of the Faculdade de Saúde Santa Casa de Belo Horizonte (CEUA-Santa Casa: 001-2023)
Acknowledgments
W.G.L. acknowledge grants from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) by a post-doctoral scholarship and doctorate-sandwich abroad. And, financial support from CNRS and Université de Strasbourg.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sharma, N.K.; Garg, R.; Baliga, S.; Bhat, G.J.J.o.c.; JCDR, d.r. Nosocomial infections and drug susceptibility patterns in methicillin sensitive and methicillin resistant Staphylococcus aureus. 2013, 7, 2178.
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M.; Holland, T.L.; Fowler Jr, V.G.J.N.R.M. Methicillin-resistant Staphylococcus aureus: an overview of basic and clinical research. 2019, 17, 203-218.
- Ventola, C.L.J.P.; therapeutics. The antibiotic resistance crisis: part 1: causes and threats. 2015, 40, 277.
- Mahjabeen, F.; Saha, U.; Mostafa, M.N.; Siddique, F.; Ahsan, E.; Fathma, S.; Tasnim, A.; Rahman, T.; Faruq, R.; Sakibuzzaman, M.J.C. An Update on Treatment Options for Methicillin-Resistant Staphylococcus aureus (MRSA) Bacteremia: A Systematic Review. 2022, 14.
- Oliveira, C.F.d.; Morey, A.T.; Biasi-Garbin, R.P.; Perugini, M.R.E.; Yamauchi, L.M.; Yamada-Ogatta, S.F. Emergência de Staphylococcus aureus resistentes aos antimicrobianos: um desafio contínuo. 2014.
- Mba, I.E.; Nweze, E.I.J.T.Y.j.o.b.; medicine. Focus: Antimicrobial Resistance: Antimicrobial Peptides Therapy: An Emerging Alternative for Treating Drug-Resistant Bacteria. 2022, 95, 445.
- Mwangi, J.; Hao, X.; Lai, R.; Zhang, Z.-Y.J.Z.r. Antimicrobial peptides: new hope in the war against multidrug resistance. 2019, 40, 488.
- Vale, N.; Aguiar, L.; Gomes, P.J.F.i.P. Antimicrobial peptides: a new class of antimalarial drugs? 2014, 5, 275.
- Bechinger, B.; Gorr, S.-U.J.J.o.d.r. Antimicrobial peptides: mechanisms of action and resistance. 2017, 96, 254-260.
- Chen, C.H.; Lu, T.K.J.A. Development and challenges of antimicrobial peptides for therapeutic applications. 2020, 9, 24.
- Brogden, K.A.J.N.r.m. Antimicrobial peptides: pore formers or metabolic inhibitors in bacteria? 2005, 3, 238-250.
- Huang, H.W.; Charron, N.E.J.Q.r.o.b. Understanding membrane-active antimicrobial peptides. 2017, 50, e10.
- Espeche, J.C.; Varas, R.; Maturana, P.; Cutro, A.C.; Maffía, P.C.; Hollmann, A.J.P.S. Membrane permeability and antimicrobial peptides: Much more than just making a hole. 2023, e24305.
- Marr, A.K.; Gooderham, W.J.; Hancock, R.E.J.C.o.i.p. Antibacterial peptides for therapeutic use: obstacles and realistic outlook. 2006, 6, 468-472.
- Liu, Y.; Shi, J.; Tong, Z.; Jia, Y.; Yang, K.; Wang, Z.J.M. Potent broad-spectrum antibacterial activity of amphiphilic peptides against multidrug-resistant bacteria. 2020, 8, 139.
- Santos, D.; Verly, R.; Piló-Veloso, D.; De Maria, M.; De Carvalho, M.; Cisalpino, P.; Soares, B.; Diniz, C.; Farias, L.; Moreira, D.J.A.A. LyeTx I, a potent antimicrobial peptide from the venom of the spider Lycosa erythrognatha. 2010, 39, 135-144.
- Fuscaldi, L.L.; de Avelar Júnior, J.T.; Dos Santos, D.M.; Boff, D.; de Oliveira, V.L.S.; Gomes, K.A.G.G.; Cruz, R.d.C.; de Oliveira, P.L.; Magalhães, P.P.; Cisalpino, P.S.J.E.B., et al. Shortened derivatives from native antimicrobial peptide LyeTx I: In vitro and in vivo biological activity assessment. 2021, 246, 414-425.
- Cardoso, B.G.; de Lima, W.G.; Fernandes, S.O.A.; de Lima, M.E.; Cardoso, V.N.J.N.P.R. Antifungal activity of a shortened analogue of the natural peptide LyeTx I isolated from the venom of the spider Lycosa erythrognatha. 2023, 37, 759-763.
- Lima, W.G.; Brito, J.C.M.; de Lima, M.E.; Pizarro, A.C.S.T.; Vianna, M.A.M.d.M.; de Paiva, M.C.; de Assis, D.C.S.; Cardoso, V.N.; Fernandes, S.O.A.J.T.J.o.A. A short synthetic peptide, based on LyeTx I from Lycosa erythrognatha venom, shows potential to treat pneumonia caused by carbapenem-resistant Acinetobacter baumannii without detectable resistance. 2021, 74, 425-434.
- Santos Neto, N.A.d. United we stand, divided we fall: antibiofilm activity and mechanisms of action of synthetic peptides in combination with ciprofloxacin against Staphylococcus aureus biofilm. 2023.
- de Brito, J.C.M. Peptídeo antimicrobiano PEGuilado como potencial biofármaco para o tratamento de pneumonia causada por Acinetobacter baumannii. 2020.
- Wiseman, T.; Williston, S.; Brandts, J.F.; Lin, L.-N.J.A.b. Rapid measurement of binding constants and heats of binding using a new titration calorimeter. 1989, 179, 131-137.
- Seelig, J.J.B.e.B.A.-B. Thermodynamics of lipid–peptide interactions. 2004, 1666, 40-50.
- Abrunhosa, F.; Faria, S.; Gomes, P.; Tomaz, I.; Pessoa, J.C.; Andreu, D.; Bastos, M.J.T.J.o.P.C.B. Interaction and lipid-induced conformation of two cecropin− melittin hybrid peptides depend on peptide and membrane composition. 2005, 109, 17311-17319.
- Niu, Z.; Prade, E.; Malideli, E.; Hille, K.; Jussupow, A.; Mideksa, Y.G.; Yan, L.M.; Qian, C.; Fleisch, M.; Messias, A.C.J.A.C. Structural Insight into IAPP-Derived Amyloid Inhibitors and Their Mechanism of Action. 2020, 132, 5820-5830.
- Junior, E.F.; Guimarães, C.F.; Franco, L.L.; Alves, R.J.; Kato, K.C.; Martins, H.R.; de Souza Filho, J.D.; Bemquerer, M.P.; Munhoz, V.H.; Resende, J.M.J.A.a. Glycotriazole-peptides derived from the peptide HSP1: synergistic effect of triazole and saccharide rings on the antifungal activity. 2017, 49, 1389-1400.
- Hassoun, A.; Linden, P.K.; Friedman, B.J.C.c. Incidence, prevalence, and management of MRSA bacteremia across patient populations—a review of recent developments in MRSA management and treatment. 2017, 21, 1-10.
- Wijesekara, P.N.K.; Kumbukgolla, W.W.; Jayaweera, J.A.A.S.; Rawat, D.J.V.s. Review on usage of vancomycin in livestock and humans: maintaining its efficacy, prevention of resistance and alternative therapy. 2017, 4, 6.
- Lima, W.G.; Batista Filho, F.L.; Lima, I.P.; Simião, D.C.; Brito, J.C.M.; da Cruz Nizer, W.S.; Cardoso, V.N.; Fernandes, S.O.A.J.N.P.R. Antibacterial, anti-biofilm, and anti-adhesive activities of melittin, a honeybee venom-derived peptide, against quinolone-resistant uropathogenic Escherichia coli (UPEC). 2022, 36, 6381-6388.
- Percival, S.L.; McCarty, S.M.; Lipsky, B.J.A.i.w.c. Biofilms and wounds: an overview of the evidence. 2015, 4, 373-381.
- Tolker-Nielsen, T.J.M.B. Biofilm development. 2015, 51-66.
- Venkatesan, N.; Perumal, G.; Doble, M.J.F.m. Bacterial resistance in biofilm-associated bacteria. 2015, 10, 1743-1750.
- Hall, C.W.; Mah, T.-F.J.F.m.r. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. 2017, 41, 276-301.
- de Avelar Júnior, J.T. Estudo de três peptídeos sintéticos com atividade antimicrobiana, derivados da toxina LyeTx I da aranha Lycosa erythrognatha (Lucas, 1836). 2015.
- Wang, S.; Zeng, X.; Yang, Q.; Qiao, S.J.I.j.o.m.s. Antimicrobial peptides as potential alternatives to antibiotics in food animal industry. 2016, 17, 603.
- Lima, W.G.; de Brito, J.C.M.; Cardoso, V.N.; Fernandes, S.O.A.J.E.J.o.P.S. In-depth characterization of antibacterial activity of melittin against Staphylococcus aureus and use in a model of non-surgical MRSA-infected skin wounds. 2021, 156, 105592.
- Stogios, P.J.; Savchenko, A.J.P.S. Molecular mechanisms of vancomycin resistance. 2020, 29, 654-669.
- Hayami, M.; Okabe, A.; Kariyama, R.; Abe, M.; Kanemasa, Y.J.M.; immunology. Lipid composition of Staphylococcus aureus and its derived L-forms. 1979, 23, 435-442.
- Hernández-Villa, L.; Manrique-Moreno, M.; Leidy, C.; Jemioła-Rzemińska, M.; Ortíz, C.; Strzałka, K.J.B.c. Biophysical evaluation of cardiolipin content as a regulator of the membrane lytic effect of antimicrobial peptides. 2018, 238, 8-15.
- Perez, M.I.; Trier, S.M.; Bernal, A.; Vargas, J.C.; Herrfurth, C.; Feussner, I.; Gonzalez, J.M.; Leidy, C.J.B.J. S. Aureus adapt to growth conditions by changing membrane order. 2014, 106, 580a.
- Verly, R.M.; Rodrigues, M.A.; Daghastanli, K.R.P.; Denadai, A.M.L.; Cuccovia, I.M.; Bloch Jr, C.; Frézard, F.; Santoro, M.M.; Piló-Veloso, D.; Bemquerer, M.P.J.P. Effect of cholesterol on the interaction of the amphibian antimicrobial peptide DD K with liposomes. 2008, 29, 15-24.
- Moreira Brito, J.C.; Carvalho, L.R.; Neves de Souza, A.; Carneiro, G.; Magalhães, P.P.; Farias, L.M.; Guimarães, N.R.; Verly, R.M.; Resende, J.M.; Elena de Lima, M.J.F.i.M.B. PEGylation of the antimicrobial peptide LyeTx Ib maintains structure-related biological properties and improves selectivity. 2022, 9, 1001508.
- Greenfield, N.J.J.N.p. Using circular dichroism spectra to estimate protein secondary structure. 2006, 1, 2876-2890.
- Verly, R.M.; Resende, J.M.; Junior, E.F.; de Magalhães, M.T.; Guimarães, C.F.; Munhoz, V.H.; Bemquerer, M.P.; Almeida, F.C.; Santoro, M.M.; Piló-Veloso, D.J.S.r. Structure and membrane interactions of the homodimeric antibiotic peptide homotarsinin. 2017, 7, 40854.
- Rashid, R.; Cazenave-Gassiot, A.; Gao, I.H.; Nair, Z.J.; Kumar, J.K.; Gao, L.; Kline, K.A.; Wenk, M.R.J.P.o. Comprehensive analysis of phospholipids and glycolipids in the opportunistic pathogen Enterococcus faecalis. 2017, 12, e0175886.
- Van Bambeke, F.; Tulkens, P.M.; Brasseur, R.; Mingeot-Leclercq, M.-P.J.E.J.o.P.M.P. Aminoglycoside antibiotics induce aggregation but not fusion of negatively-charged liposomes. 1995, 289, 321-333.
- Van Bambeke, F.; Mingeot-Leclercq, M.-P.; Brasseur, R.; Tulkens, P.M.; Schanck, A.J.C.; lipids, p.o. Aminoglycoside antibiotics prevent the formation of non-bilayer structures in negatively-charged membranes. Comparative studies using fusogenic (bis (β-diethylaminoethylether) hexestrol) and aggregating (spermine) agents. 1996, 79, 123-135.
- Parmanik, A.; Das, S.; Kar, B.; Bose, A.; Dwivedi, G.R.; Pandey, M.M.J.C.m. Current treatment strategies against multidrug-resistant bacteria: a review. 2022, 79, 388.
- Tängdén, T.J.U.j.o.m.s. Combination antibiotic therapy for multidrug-resistant Gram-negative bacteria. 2014, 119, 149-153.
- Hatlen, T.J.; Miller, L.G.J.I.D.C. Staphylococcal skin and soft tissue infections. 2021, 35, 81-105.
- Simonetti, O.; Marasca, S.; Candelora, M.; Rizzetto, G.; Radi, G.; Molinelli, E.; Brescini, L.; Cirioni, O.; Offidani, A.J.A.m. Methicillin-resistant Staphylococcus aureus as a cause of chronic wound infections: Alternative strategies for management. 2022, 8, 125.
- Santana, H.J.A. Interação do glicopeptídeo Vancomicina em monocamadas de Langmuir como modelos de membrana celular. 2022.
- (BrCAST), C.B.D.T.D.S.A.A. Orientações do EUCAST para a detecção de mecanismos de resistência e resistências específicas de importância clínica e/ou epidemiológica. 2017.
- Herrera, K.M.; da Silva, F.K.; de Lima, W.G.; Barbosa, C.d.S.; Gonçalves, A.M.; Viana, G.H.; Soares, A.C.; Ferreira, J.M.J.M.C.R. Antibacterial and antibiofilm activities of synthetic analogs of 3-alkylpyridine marine alkaloids. 2020, 29, 1084-1089.
- Orhan, G.; Bayram, A.; Zer, Y.; Balci, I.J.J.o.c.m. Synergy tests by E test and checkerboard methods of antimicrobial combinations against Brucella melitensis. 2005, 43, 140-143.
- Oroojalian, F.; Kasra-Kermanshahi, R.; Azizi, M.J.P.M. Synergistic antibacterial activity of Bunium persicum and Cuminum cyminum essential oils. 2008, 74, PI36.
- Mohamed, M.F.; Abdelkhalek, A.; Seleem, M.N.J.S.r. Evaluation of short synthetic antimicrobial peptides for treatment of drug-resistant and intracellular Staphylococcus aureus. 2016, 6, 29707.
- Stewart, J.C.M.J.A.b. Colorimetric determination of phospholipids with ammonium ferrothiocyanate. 1980, 104, 10-14.
- Brazil. AGENCIA NACIONAL DE VIGILÂNCIA SANITÁRIA (ANVISA). 5ª ed.; Farmacopeia Brasileira, Brasilia: 2010b; Vol. 1.
Figure 1.
Time-kill curves of LyeTx I mn∆K and vancomycin against a suspension of methicillin-resistant Staphylococcus aureus (MRSA) on logarithmic growth phase (106 CFU/mL). MIC: Minimum inhibitory concentration.
Figure 1.
Time-kill curves of LyeTx I mn∆K and vancomycin against a suspension of methicillin-resistant Staphylococcus aureus (MRSA) on logarithmic growth phase (106 CFU/mL). MIC: Minimum inhibitory concentration.
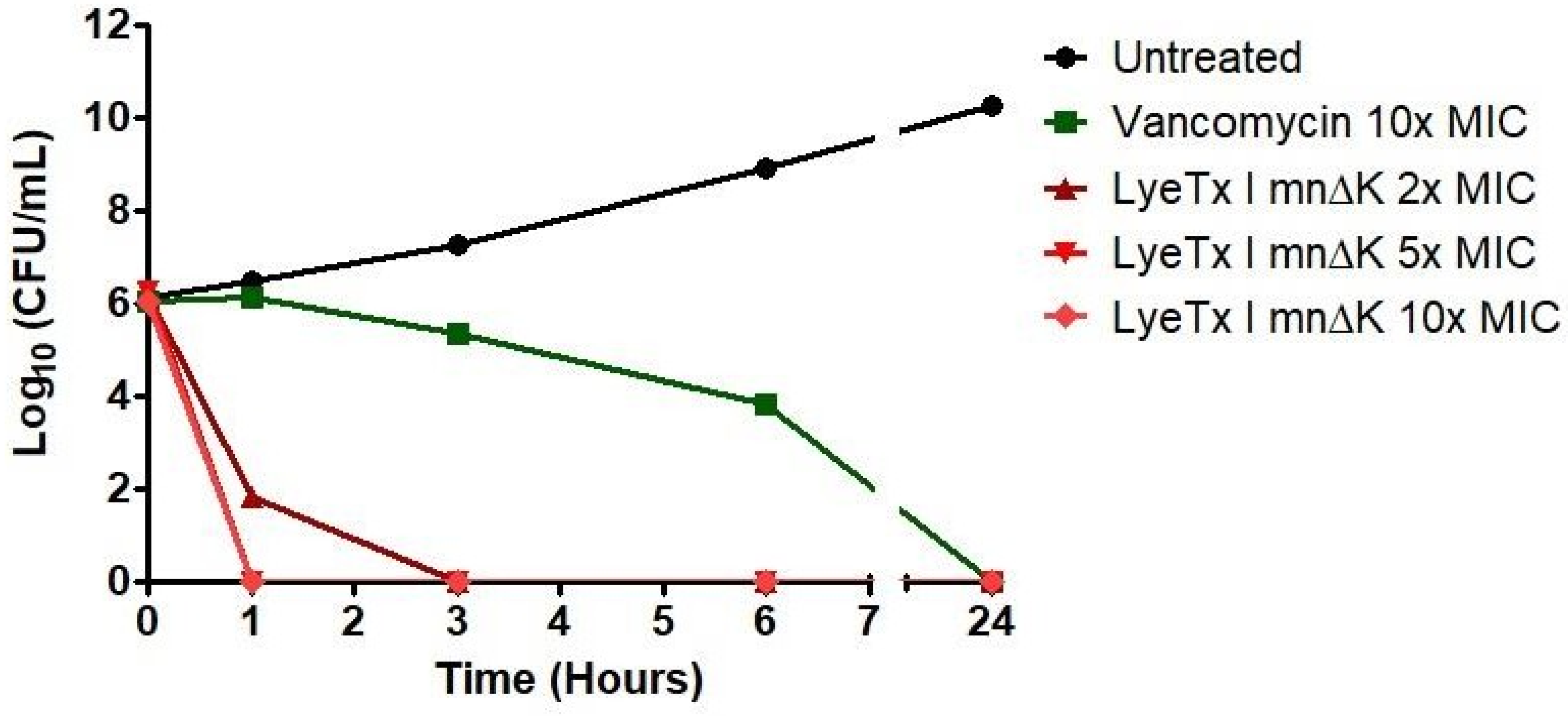
Figure 2.
Evaluation of the effect of different concentrations of LyeTx I mnΔK and the control (vancomycin) on the formation of biofilm and pre-formed biofilm of methicillin-resistant Staphylococcus aureus (MRSA). (A) Reduction of mature biofilm; (B) prevention of the formation of biofilm. Two asterisks (**) indicate statistically significant difference compared to the control, with p-value <0.01. Three asterisks (***) indicate statistically significant difference compared to the control, with p-value < 0.0001. The results were analyzed by one-way ANOVA with Dunnett’s post-test.
Figure 2.
Evaluation of the effect of different concentrations of LyeTx I mnΔK and the control (vancomycin) on the formation of biofilm and pre-formed biofilm of methicillin-resistant Staphylococcus aureus (MRSA). (A) Reduction of mature biofilm; (B) prevention of the formation of biofilm. Two asterisks (**) indicate statistically significant difference compared to the control, with p-value <0.01. Three asterisks (***) indicate statistically significant difference compared to the control, with p-value < 0.0001. The results were analyzed by one-way ANOVA with Dunnett’s post-test.
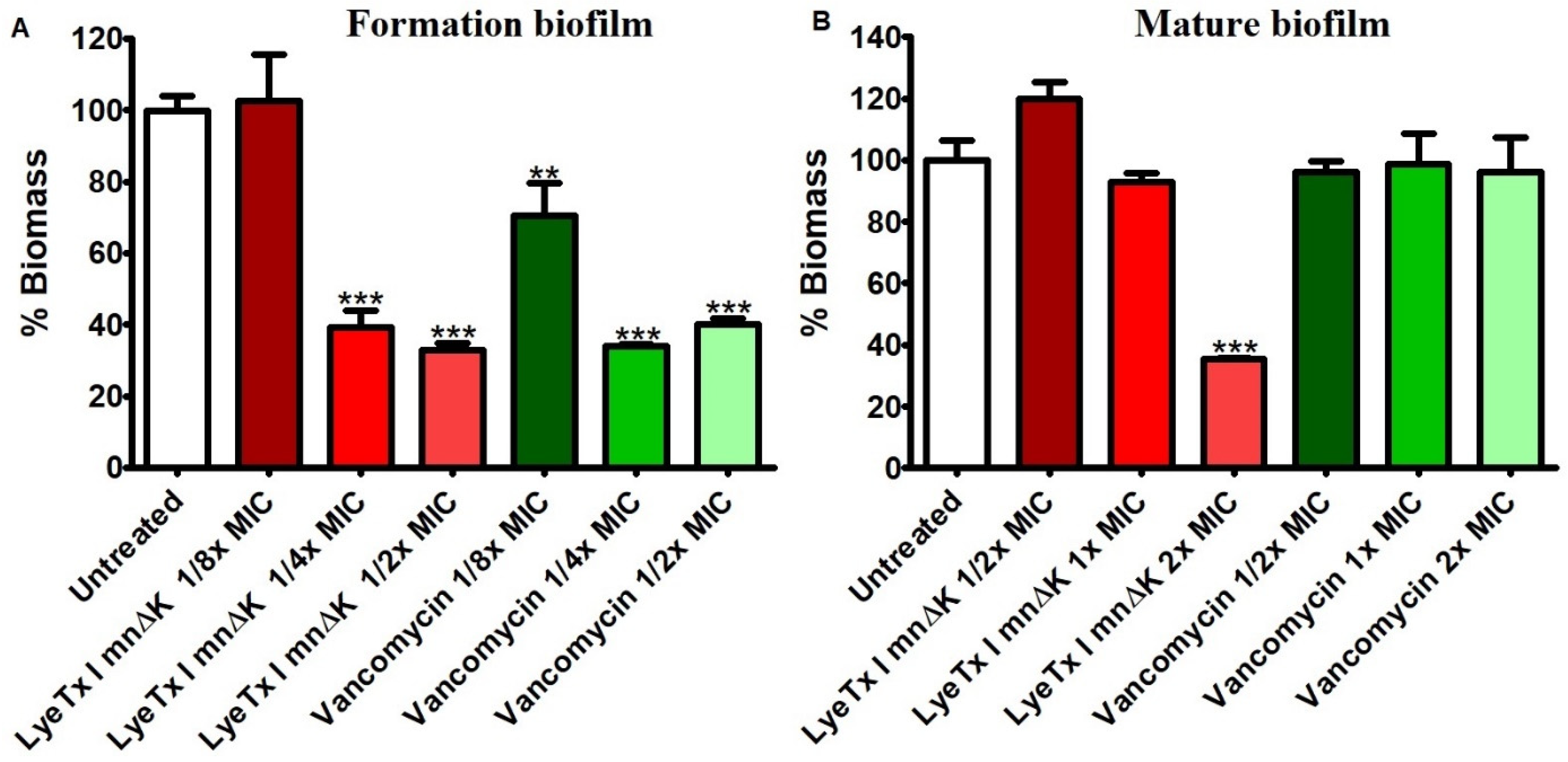
Figure 3.
Isobolograms of combination of peptide LyeTx I mn∆K with vancomycin (A) and oxacillin (B) against methicillin-resistant Staphylococcus aureus (MRSA).
Figure 3.
Isobolograms of combination of peptide LyeTx I mn∆K with vancomycin (A) and oxacillin (B) against methicillin-resistant Staphylococcus aureus (MRSA).
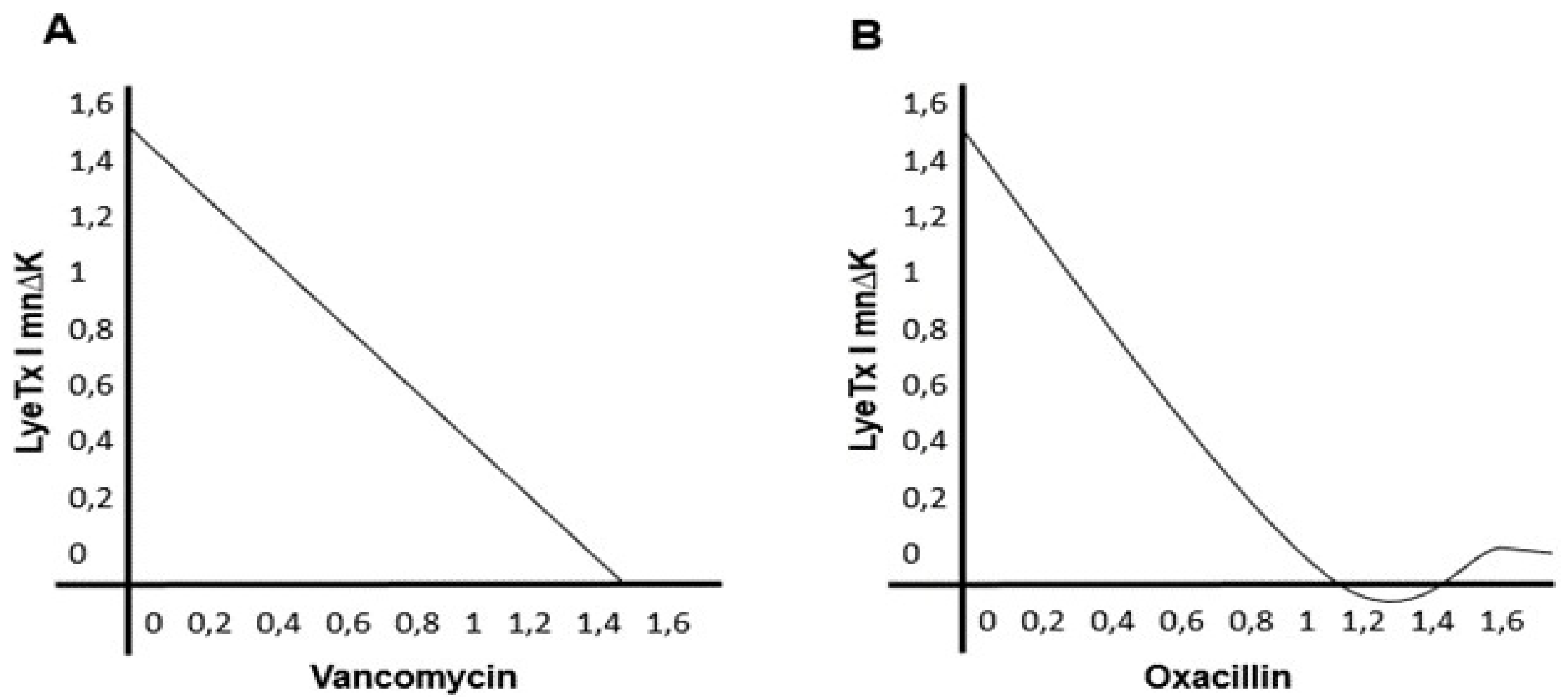
Figure 4.
Release of 260-nm-absorbing intracellular material over time for methicillin-resistant Staphylococcus aureus (MRSA) suspension treated with LyeTx I mn∆K (80 µM) or vancomycin (10 µM) at 10x MIC. Two asterisks (**) indicate statistically significant difference compared to the control, with p-value <0.01. Three asterisks (***) indicate statistically significant difference compared to the control, with p-value <0.0001. Results were analyzed by One-way ANOVA with Dunnett’s post-test.
Figure 4.
Release of 260-nm-absorbing intracellular material over time for methicillin-resistant Staphylococcus aureus (MRSA) suspension treated with LyeTx I mn∆K (80 µM) or vancomycin (10 µM) at 10x MIC. Two asterisks (**) indicate statistically significant difference compared to the control, with p-value <0.01. Three asterisks (***) indicate statistically significant difference compared to the control, with p-value <0.0001. Results were analyzed by One-way ANOVA with Dunnett’s post-test.
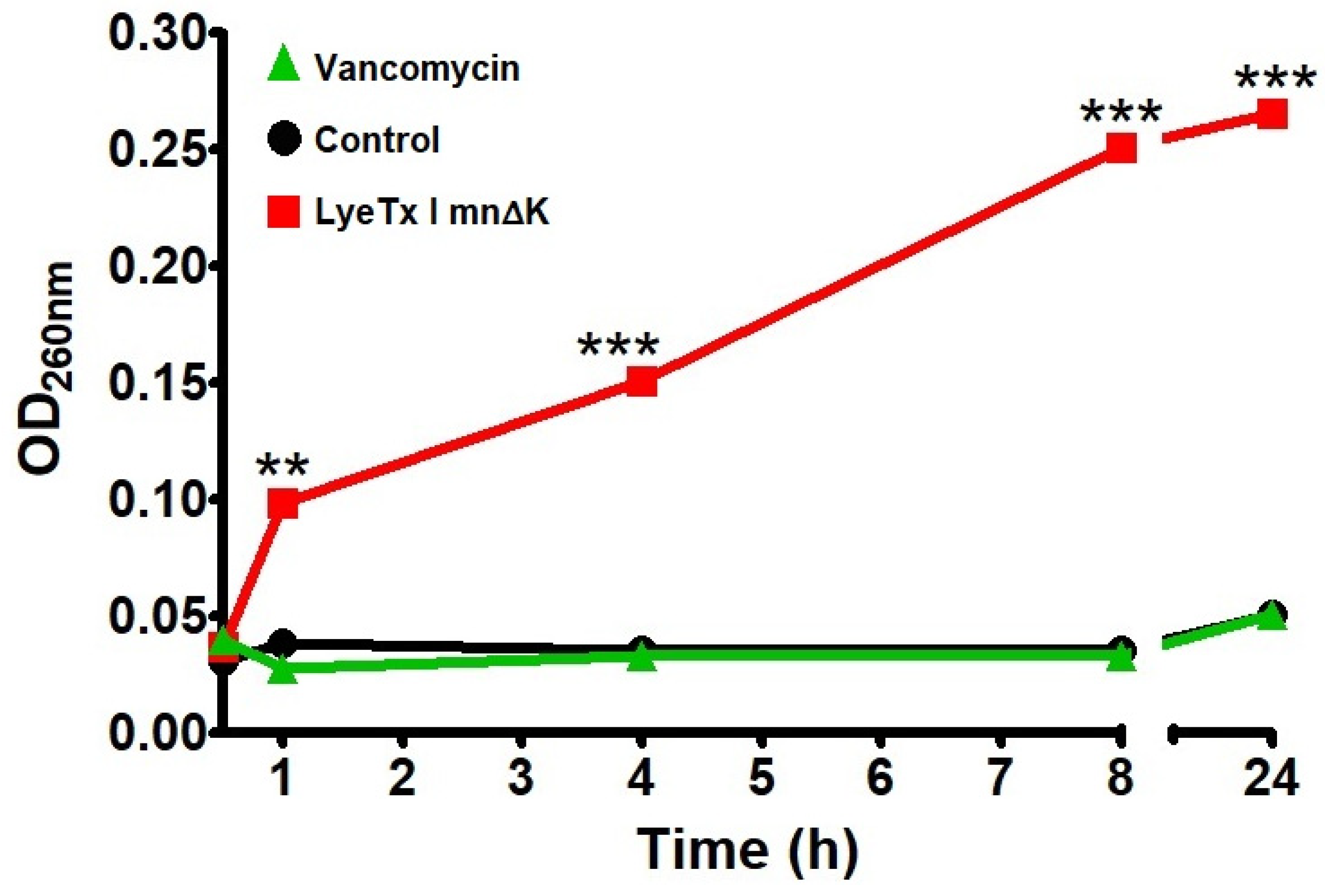
Figure 5.
Calorimetric titration isotherms obtained from the titration of a solution of 25 μM LyeTx I mn∆K with 20 mM POPG:CL (2:1, mol/mol) unilamellar phospholipid vesicles (LUVs) (A). In the upper panel the heat flow graphs for each LUVs injection as a function of time. The lower panel shows the enthalpy as a function of the phospholipid/peptide molar ratio (the red curve represents the non-linear fitting by eq. (1)). Circular dichroism (CD) spectra of 50 µM LyeTx I mn∆K in the presence of POPG:CL (2:1) (B). CD spectra of the peptides in 10 mM Tris, 50 mM NaCl, pH 7.4 are presented by black opened circles, and after addition of different phospholipid concentration: 100 µM by red filled squares, 250 µM by blue filled upper triangles, 500 µM by magenta filled down triangles and 750 µM by filled green circles.
Figure 5.
Calorimetric titration isotherms obtained from the titration of a solution of 25 μM LyeTx I mn∆K with 20 mM POPG:CL (2:1, mol/mol) unilamellar phospholipid vesicles (LUVs) (A). In the upper panel the heat flow graphs for each LUVs injection as a function of time. The lower panel shows the enthalpy as a function of the phospholipid/peptide molar ratio (the red curve represents the non-linear fitting by eq. (1)). Circular dichroism (CD) spectra of 50 µM LyeTx I mn∆K in the presence of POPG:CL (2:1) (B). CD spectra of the peptides in 10 mM Tris, 50 mM NaCl, pH 7.4 are presented by black opened circles, and after addition of different phospholipid concentration: 100 µM by red filled squares, 250 µM by blue filled upper triangles, 500 µM by magenta filled down triangles and 750 µM by filled green circles.
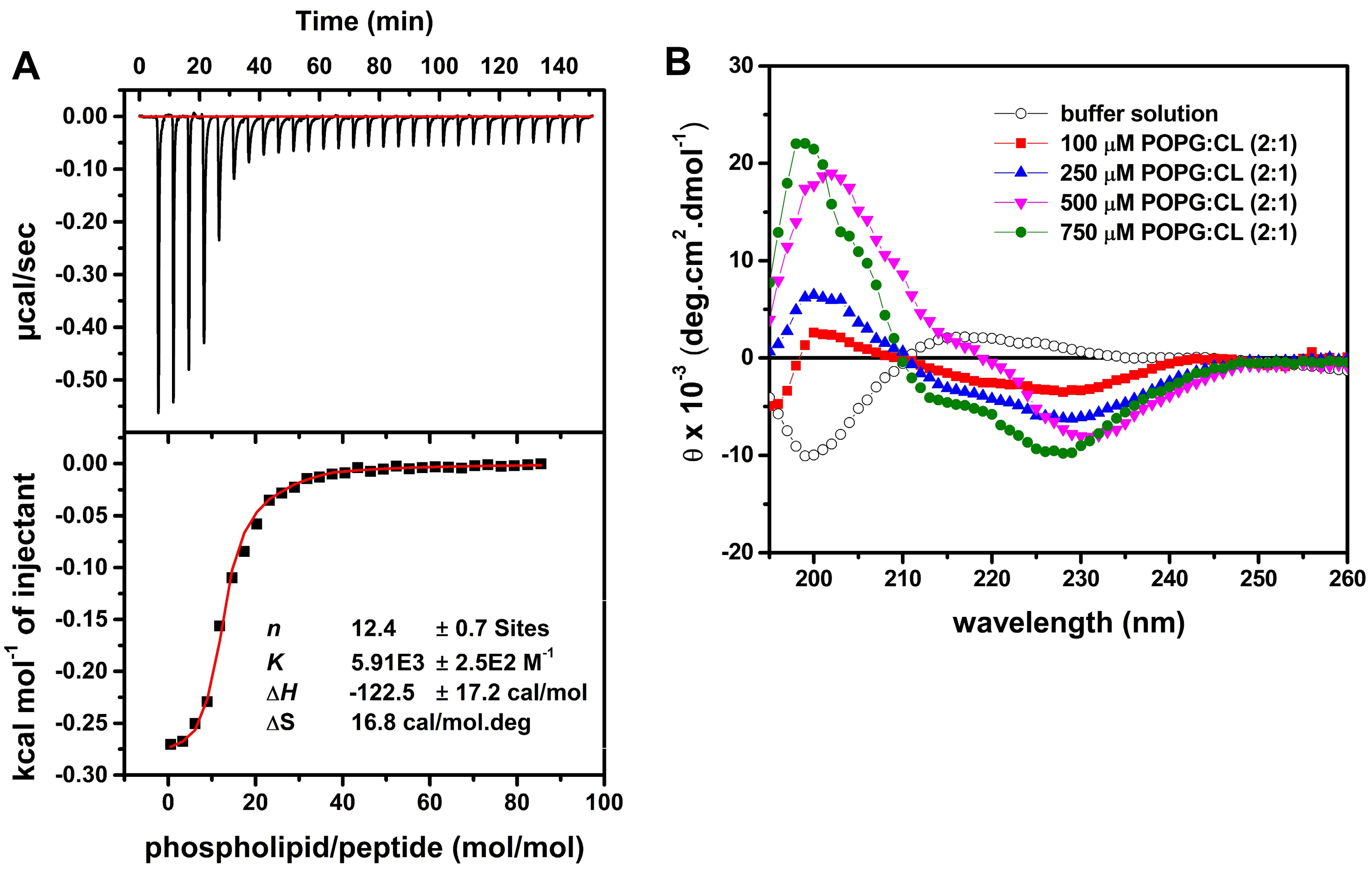
Figure 6.
Circular dichroism (CD) spectra of 50 µM LyeTx I mn∆K in the presence of POPC:Chol (2:1). The CD spectra of the peptide in 10 mM Tris, 50 mM NaCl, pH 7.4 are presented by black open circles, and after addition of different phospholipid concentrations: 100 µM by red filled squares, 250 µM by blue filled upper triangles, 500 µM by magenta filled down triangles and 750 µM by filled green circles.
Figure 6.
Circular dichroism (CD) spectra of 50 µM LyeTx I mn∆K in the presence of POPC:Chol (2:1). The CD spectra of the peptide in 10 mM Tris, 50 mM NaCl, pH 7.4 are presented by black open circles, and after addition of different phospholipid concentrations: 100 µM by red filled squares, 250 µM by blue filled upper triangles, 500 µM by magenta filled down triangles and 750 µM by filled green circles.
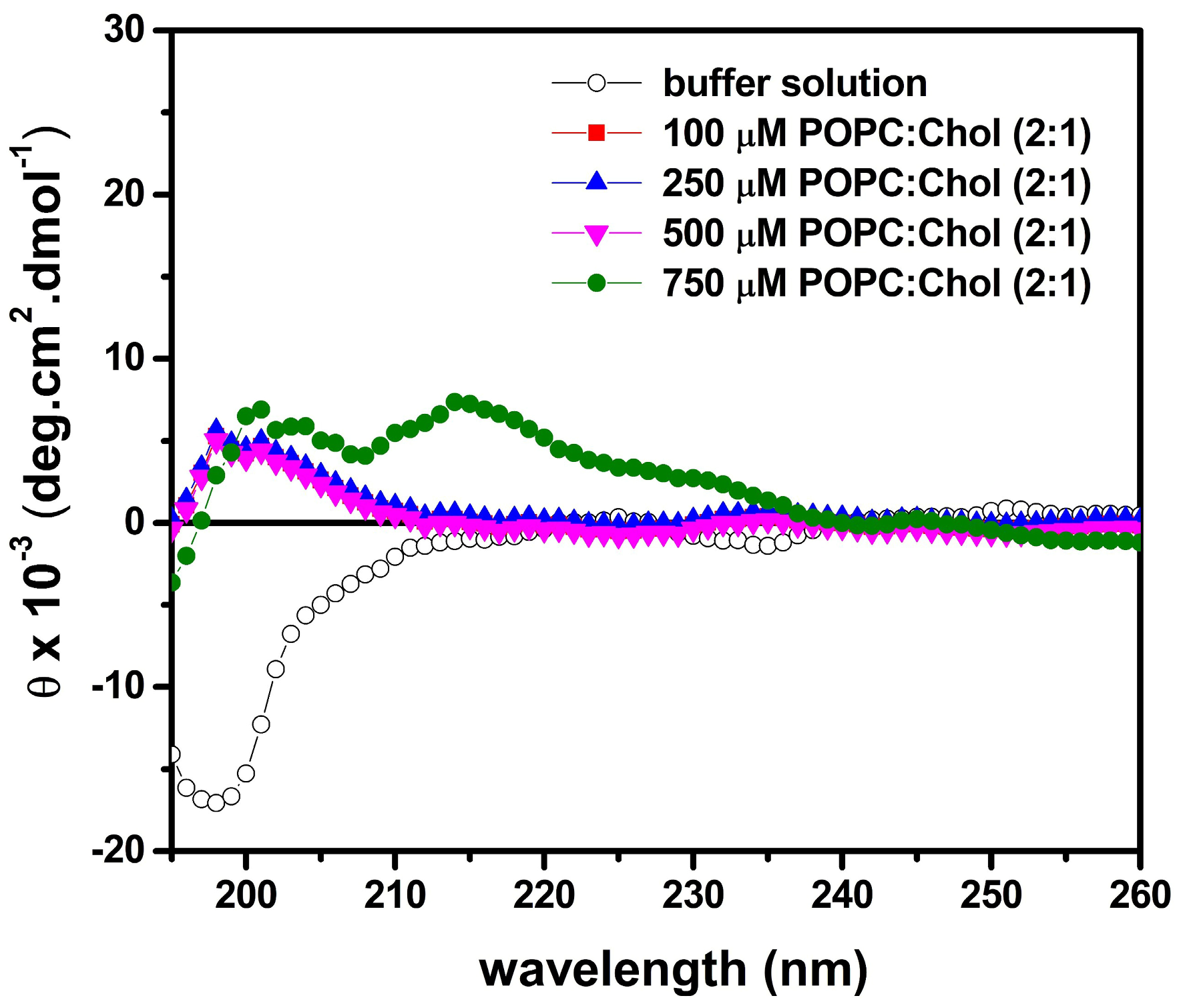
Figure 7.
Effect of LyeTx I mn∆K on phospholipid membranes as assessed by calcein release from POPC:Chol (2:1) at 100 µM lipid concentration. (A) Time dependence of membrane permeability with LyeTx I mn∆K. (B) Percentage of calcein release as a function of peptide concentration (positive control - 2% Triton X-100, considered as 100%). Data are representative of three independent experiments.
Figure 7.
Effect of LyeTx I mn∆K on phospholipid membranes as assessed by calcein release from POPC:Chol (2:1) at 100 µM lipid concentration. (A) Time dependence of membrane permeability with LyeTx I mn∆K. (B) Percentage of calcein release as a function of peptide concentration (positive control - 2% Triton X-100, considered as 100%). Data are representative of three independent experiments.
Figure 8.
Effect of LyeTx I mn∆K on the size and charge of 200 µM POPG:CL (2:1 mol/mol) and 200 µM POPG:CL:LysylPOPG LUVs (2:2:1 mol/mol/mol). (A) Changes in the hydrodynamic diameter (Dh) and (B) in the zeta potential as a function of the peptide concentration. Data are representative of three independent experiments.
Figure 8.
Effect of LyeTx I mn∆K on the size and charge of 200 µM POPG:CL (2:1 mol/mol) and 200 µM POPG:CL:LysylPOPG LUVs (2:2:1 mol/mol/mol). (A) Changes in the hydrodynamic diameter (Dh) and (B) in the zeta potential as a function of the peptide concentration. Data are representative of three independent experiments.
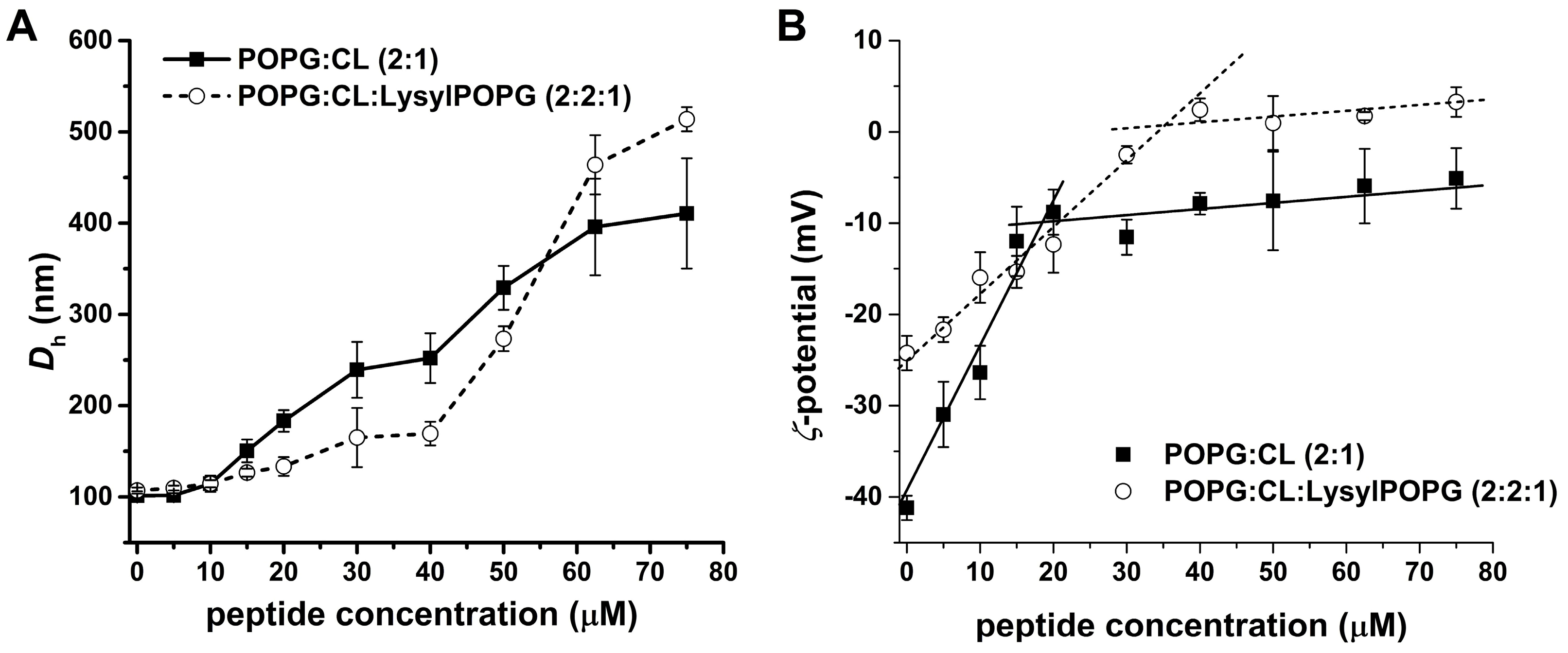
Figure 9.
Effect of [LyeTx I mn∆K] on phospholipid membranes as assessed by calcein release from POPG:CL (2:1) and POPG:CL:LysylPOPG LUVs (2:2:1), both at 100 µM of lipid concentration. Time dependence of membrane permeability with LyeTx I mn∆K (a) POPG:CL (2:1); (b) POPG:CL:LysylPOPG LUVs (2:2:1); (c) observed rate constants, kobs, of CF release vs. peptide concentration and (d) percentage of calcein release compared to what was observed after addition of 2% Triton X-100 (positive control, considered as 100%). Data are representative of three independent experiments.
Figure 9.
Effect of [LyeTx I mn∆K] on phospholipid membranes as assessed by calcein release from POPG:CL (2:1) and POPG:CL:LysylPOPG LUVs (2:2:1), both at 100 µM of lipid concentration. Time dependence of membrane permeability with LyeTx I mn∆K (a) POPG:CL (2:1); (b) POPG:CL:LysylPOPG LUVs (2:2:1); (c) observed rate constants, kobs, of CF release vs. peptide concentration and (d) percentage of calcein release compared to what was observed after addition of 2% Triton X-100 (positive control, considered as 100%). Data are representative of three independent experiments.
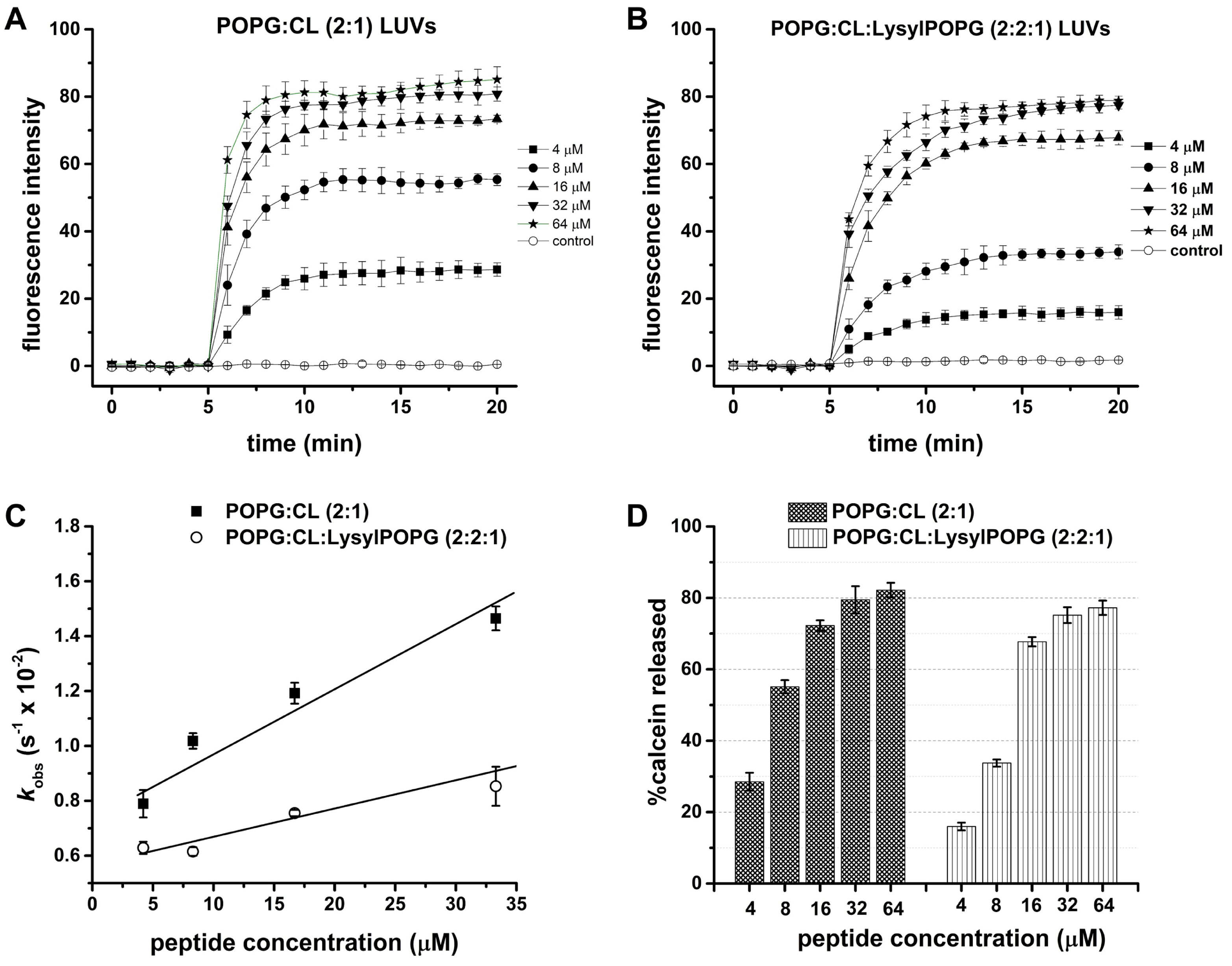
Figure 10.
Bacterial load (Log10CFU/g of wound) after the topical treatment of non-surgical methicillin-resistant Staphylococcus aureus (MRSA)-infected wounds with gel contained LyeTx I mnΔK (0.25%, 0.50%, or 1.00%), vancomycin 1%, gel ointment or saline (control) (n = 5). Mice were intradermally injected with 107 CFU of highly virulent MRSA USA300. Followed 48 h after injection, the mice developed an open wound/abscess at the local site of injection and were treated twice daily for 3 days. An asterisk (*) indicates a statistically significant difference compared to the control with p-value <0.05. All results were analyzed by one-way ANOVA with Dunnett’s post-test.
Figure 10.
Bacterial load (Log10CFU/g of wound) after the topical treatment of non-surgical methicillin-resistant Staphylococcus aureus (MRSA)-infected wounds with gel contained LyeTx I mnΔK (0.25%, 0.50%, or 1.00%), vancomycin 1%, gel ointment or saline (control) (n = 5). Mice were intradermally injected with 107 CFU of highly virulent MRSA USA300. Followed 48 h after injection, the mice developed an open wound/abscess at the local site of injection and were treated twice daily for 3 days. An asterisk (*) indicates a statistically significant difference compared to the control with p-value <0.05. All results were analyzed by one-way ANOVA with Dunnett’s post-test.
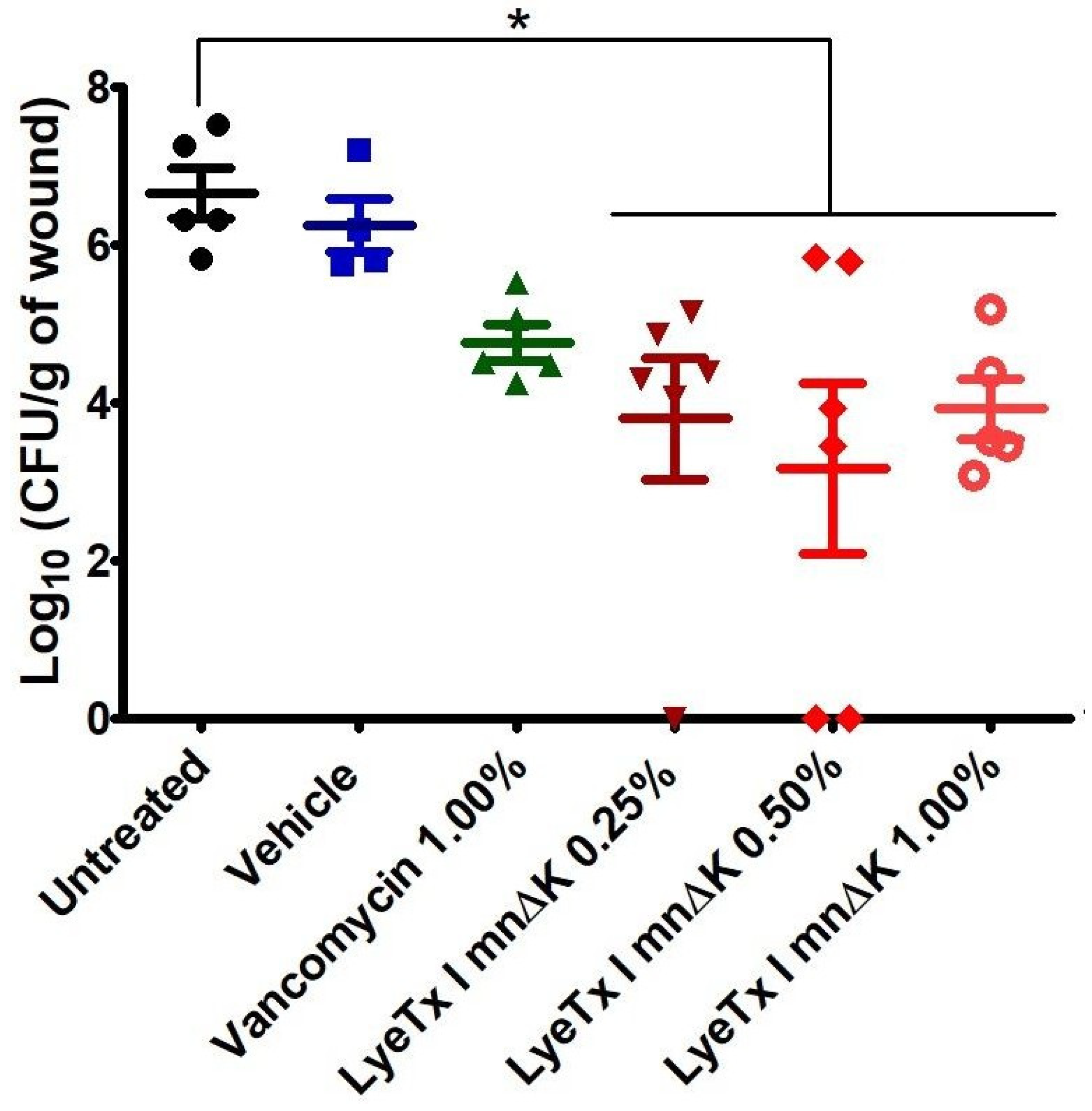
Figure 11.
Effect of the peptide on changes in body weight in mice infected with methicillin-resistant Staphylococcus aureus (MRSA). An asterisk (*) indicates a statistically significant difference between LyeTx I mnΔK 0.50% and untreated group with p-value <0.05. All results were analyzed by one-way ANOVA with Dunnett’s post-test.
Figure 11.
Effect of the peptide on changes in body weight in mice infected with methicillin-resistant Staphylococcus aureus (MRSA). An asterisk (*) indicates a statistically significant difference between LyeTx I mnΔK 0.50% and untreated group with p-value <0.05. All results were analyzed by one-way ANOVA with Dunnett’s post-test.

Table 1.
Minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) of LyeTx I mnΔK and vancomycin against clinical isolates of methicillin-resistant Staphylococcus aureus (MRSA).
Table 1.
Minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) of LyeTx I mnΔK and vancomycin against clinical isolates of methicillin-resistant Staphylococcus aureus (MRSA).
|
Microorganism |
Clinical origin |
Antibacterial activity (µM) | |||
|---|---|---|---|---|---|
| LyeTx I mnΔK | Vancomycin | ||||
| MIC | MBC | MIC | MBC | ||
| S. aureus 11 | Infected wound | 8 | 32 | 1 | 8 |
| S. aureus 29 | Infected wound | 8 | 16 | 1 | 2 |
| S. aureus 130 | Infected wound | 8 | 32 | 1 | 2 |
| S. aureus 366 | Infected wound | 16 | 32 | 1 | 8 |
| S. aureus 524 | Infected wound | 16 | 32 | 1 | 2 |
| S. aureus 526 | Infected wound | 16 | 32 | 1 | 4 |
| MRSA USA300 | Infected wound | 8 | 16 | 1 | 2 |
| MIC50 | 8 | 1 | |||
| MBC50 | 32 | 2 | |||
MIC50: Minimum inhibitory concentration required to inhibit 50% of MRSA isolates tested; MBC50: Minimum bactericidal concentration required to inhibit 50% of MRSA isolates tested.
Table 2.
Range of fractional inhibitory concentration (FIC) and fractional inhibitory concentration index (FICI) of LyeTx I mn∆K in combination with vancomycin or oxacillin against methicillin-resistant Staphylococcus aureus (MRSA).
Table 2.
Range of fractional inhibitory concentration (FIC) and fractional inhibitory concentration index (FICI) of LyeTx I mn∆K in combination with vancomycin or oxacillin against methicillin-resistant Staphylococcus aureus (MRSA).
|
Class |
Antimicrobials |
FIC | FICI (ΣFIC) |
Effect | |
|---|---|---|---|---|---|
| LyeTx I mnΔK | Antimicrobial | ||||
| Glycopeptide | Vancomycin | 1.00 | 0.50 | 1.50 | Indifferent |
| β-lactam | Oxacillin | 1.00 | 0.02 | 1.02 | Indifferent |
ΣFICI: Fractional inhibitory concentration index. The FICI was interpreted as follows: A FICI ≤0.5 is considered evidence of synergy. Additive was defined as 0.5< FICI <1. A 1≤ FICI ≤4 was considered an indifferent effect. Antagonism was defined as a FICI >4 [21].
Table 3.
Resensitization of methicillin-resistant Staphylococcus aureus (MRSA) to vancomycin and oxacillin using a sub-inhibitory concentration (1/4x MIC) of LyeTx I mnΔK for one hour.
Table 3.
Resensitization of methicillin-resistant Staphylococcus aureus (MRSA) to vancomycin and oxacillin using a sub-inhibitory concentration (1/4x MIC) of LyeTx I mnΔK for one hour.
| Antibacterial | MICs (µg/mL) | Fold resensitization | |
|---|---|---|---|
| Not exposed to LyeTx I mnΔK | Exposed to LyeTx I mnΔK | ||
| Vancomycin | 1.0 | 0.5 | 2 |
| Oxacillin | 128 | 64 | 2 |
Fold of resensitization: It is the ratio of the MIC of antibacterial alone divided by the MIC of antibacterial after re-sensitization with (1/4x MIC) of LyeTx I mnΔK.
Table 4.
Effect of LyeTx I mn∆K on kobs for the leakage of calcein from POPG:CL and POPG:CL:LysylPOPG LUVs.
Table 4.
Effect of LyeTx I mn∆K on kobs for the leakage of calcein from POPG:CL and POPG:CL:LysylPOPG LUVs.
| LyeTx I mn∆K (mM) | POPG:CL | POPG:CL:Lysyl:POPG | ||||||
| %calcein released |
SD* | kobs (s-1) x10-2 | SD* x10-4 |
%calcein released |
SD* | kobs (s-1) x10-2 | SD* x10-4 |
|
| 4 | 28.51 | 2.50 | 0.78 | 5.01 | 15.98 | 1.06 | 0.629 | 2.23 |
| 8 | 55.11 | 1.83 | 1.01 | 2.82 | 33.74 | 1.01 | 0.615 | 1.56 |
| 18 | 72.23 | 1.49 | 1.19 | 3.81 | 67.73 | 1.30 | 0.755 | 1.13 |
| 32 | 79.45 | 3.82 | 1.46 | 4.36 | 75.17 | 2.20 | 0.853 | 7.10 |
| 64 | 82.17 | 2.04 | 2.16 | 12.5 | 77.23 | 1.99 | 1.225 | 4.89 |
*SD - Standard deviation.
Table 5.
Sample composition used for hydrodynamic diameter (Dh) and zeta potential ( P) measurements.
Table 5.
Sample composition used for hydrodynamic diameter (Dh) and zeta potential ( P) measurements.
| Sample | Vpep ( L) | VLUVs ( L) | Vbuffer ( L) | [LyeTx I mn K] ( M) |
| 1 | 0 | 400 | 400 | 0 |
| 2 | 4 | 400 | 396 | 5 |
| 3 | 8 | 400 | 392 | 10 |
| 4 | 12 | 400 | 388 | 15 |
| 5 | 16 | 400 | 384 | 20 |
| 6 | 24 | 400 | 376 | 30 |
| 7 | 32 | 400 | 368 | 40 |
| 8 | 40 | 400 | 360 | 50 |
| 9 | 50 | 400 | 350 | 62.5 |
| 10 | 60 | 400 | 340 | 75 |
Vpep = volume of 1 mM peptide stock solution; VLUVs = volume of 400 μM POPC:POPG LUVs stock solution and Vbuffer = volume of 10 mM Tri-HCl buffer solution (pH 7.4).
Table 6.
Samples composition used for calcein release measurements.
| Sample | Vpep (μL) | VLUVs (μL) | Vbuffer (μL) | [LyeTx I mnΔK] (μM) |
| 1 | 0 | 150 | 150 | 0 |
| 2 | 5 | 150 | 145 | 4,2 |
| 3 | 10 | 150 | 140 | 8,3 |
| 4 | 20 | 150 | 130 | 16,7 |
| 5 | 40 | 150 | 110 | 33,3 |
| 6 | 80 | 150 | 70 | 66,7 |
Vpep = volume of 2 mM peptide stock solution; VLUVs = volume of 200 μM POPC: POPG LUVs stock solution and Vbuffer = volume of 10 mM Tri-HCl buffer solution (pH 7.4).
Table 7.
Components used for the formulation of LyeTx I mnΔK.
| Component | Concentration |
|---|---|
| Hydroxyethylcellulose (Natrosol®) | 2.2% |
| Sodium metabisulfite | 0.6% |
| Methylparaben (Nipagin®) | 0.2% |
| Distilled water | q.s. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



