
Submitted:
25 January 2024
Posted:
26 January 2024
You are already at the latest version
Abstract
Cryptococcus neoformans is a fungus that mainly affects the respiratory system and the central nervous system. One of the main virulence factors is the capsule, constituted by the polysaccharides glucuronoxylomannan (GXM) and glucuronoxylomanogalactan (GXMGal). Polysaccharides are immunomodulators. One of the target cell populations for modulation are macrophages, part of the first line of defense, important for innate and adaptive immunity. It has been reported that macrophages can be modulated to act as a "Trojan horse" taking phagocytosed yeasts to strategic sites or having their machinery activation compromised. The scarcity of information on canine cryptococcosis led us to assess whether the purified capsular polysaccharides from C. neoformans would be able to modulate the microbicidal action of macrophages. In the present study, we observed that the capsular polysaccharides, GXM, GXMGal or capsule total did not present toxic effects for the DH82 macrophage cell line. However, it was possible to demonstrate that phagocytic activity was decreased after treatment with polysaccharides. In addition, yeasts recovered from macrophages treated with the polysaccharides, after phagocytosis, could be cultured, showing that their viability was not altered. The polysaccharides led to a reduction in ROS production and the expression of IL-12 and IL-6. We observed that GXMGal inhibits MHC class II expression and GXM reduces ERK phosphorylation. In contrast, GXMGal and GXM were able to increase the PPAR- expression. Furthermore, our data suggests that capsular polysaccharides can reduce the microbicidal activity of canine macrophages DH82.
Keywords:
Canine Macrophages
; Capsular Polysaccharides
; Inhibition
; Cryptococcosis
1. Introduction
Cryptococcosis is a systemic disease that affects animals and people worldwide caused by an encapsulated yeast species of the genus Cryptococcus with predilection for the nervous and respiratory systems [1]. The infection in dogs is frequently caused by Cryptococcus neoformans and Cryptococcus gattii. Clinical signs depend on the sites of infection, but the involvement of critical organs, such as the CNS, eyes, gastrointestinal tract, myocardium, adrenal glands, and pancreas in dogs with cryptococcosis is frequently observed [2]. Even though the reported incidence of cryptococcosis is lower in dogs than in cats, CNS involvement may be more common in dogs, with widespread dissemination to other organs [3].
Macrophages are crucial for cryptococcosis control. It has been demonstrated that macrophage and dendritic cell depletion significantly reduced mice survival after infection with C. neoformans. Besides, the Trojan horse hypothesis suggest that fungus entry into the blood brain barrier through dissemination within the macrophage [4,5]. In this context for C. neoformans, a Trojan horse mechanism for dissemination is supported by the results obtained after depletion of alveolar macrophages, preventing brain dissemination [6]. In addition, increased brain fungal burden was observed in experiments with mice injected with ex vivo infected macrophages [7].
Although numerous proteins are important in interaction with macrophages [8], capsular polysaccharides have shown an important ability to deactivate the immune system [9]. The capsular polysaccharides of C. neoformans are the main virulence factor. The capsule is composed of glucuronoxylomannan (GXM), glucuronoxylomanogalactan (GXMGal) and small amounts of mannoproteins. GXM represents 90% of the capsule, GXMGal around 8% and mannoproteins have been identified (2%) was not studied in detail yet [10].
Previous studies have demonstrated that capsular polysaccharides as GXM and GXMGal were able to induce impairment of macrophage activity, demonstrated by modulation of proinflammatory cytokines, apoptosis induction and modulated production of neutrophil extracellular traps (NETs) by human neutrophils [11,12]. GXM can be captured by different receptors particularly TLRs, CD14, CD18 and FcγRIIB [13]. Inside the macrophage, GXM produces multiple effects, for example, reduction of the APC function [14], increase FasL and deregulation of pro-inflammatory and anti-inflammatory secretion of cytokines. Indeed, low doses of GXMGal can be able to induce FasL expression and inhibit proliferation in macrophage cell line, indicating that GXMGal is more potent to produce immunomodulatory effects [11].
Most data in the literature refer to analyzes carried out in mouse macrophages, clearly showing a lack of information on the effect of capsular polysaccharides from C. neoformans. In general, the polysaccharides seem to exert an immunomodulatory effect on several effective functions of murine macrophages [8,11,15,16]. However, there are no reports on the possible effects of these polysaccharides on canine macrophages. Understanding these mechanisms may bring light on the understanding of the initial moments of the interaction with C. neoformans. Therefore, in this study the objective was to evaluate the ability of the purified capsular polysaccharides from C. neoformans to modulate the activity of canine macrophage cell line DH82.
2. Materials and Methods
2.1. DH82 Cell Line Culture
Canine macrophages DH82 (ATCC) were grown in 75 cm2 tissue culture flasks (T75) (Nunc, Roskilde, Denmark) with Dulbecco’s Modified Eagle Medium (DMEM; Sigma-Aldrich) supplemented with 10% fetal calf serum (FCS; Gibco), 1% MEM non-essential amino acids (Sigma-Aldrich), 100 μg/mL of streptomycin and 100 Units/mL of penicillin and 2 mM of l-glutamine were cultured at 37 0C with 5% CO2. Subcultures were carried out once a week when they reached a confluence of 95–100%. Cells were released with 0.25% of trypsin (Difco) and 1 mM of EDTA (Sigma-Aldrich), harvested, and washed twice in HBSS by centrifugation at room temperature for 10 min at 250 g. The supernatant was discarded, cell pellet was resuspended in DMEM containing 10% FCS, cultured into new T75 flasks, and culture medium was changed after 3 days in culture. Before to use in all assays, cells’ viability was verified by trypan blue exclusion.
2.2. Isolation and Purification of C. neoformans Capsular Polysaccharides (GXMGal and GXM)
Polysaccharides were isolated and purified as previously described [15]. Briefly, cells grown in defined liquid medium at 37 0C with continuous shaking (100 rpm) for 5 days were removed through centrifugation (12,000 g for 1 h at 4 0C), and the capsular polysaccharides in the supernatant were precipitated after adding three volumes of cold ethanol. To separate GXMGal from mannoproteins, the capsular polysaccharides were fractionated using lectin-affinity chromatography on a XK-26 column (Pharmacia) packed with 70 mL Concanavalin A-Sepharose 4B (Pharmacia) at 4 0C. The unbound polysaccharide fractions, containing GXMGal, were localized through a phenol-sulfuric reaction, pooled, dialyzed against distilled water and lyophilized. Pure GXMGal was obtained through anion-exchange chromatography on a MonoQ (HR16/10) column using a 50 mL super loop.
GXM was purified by differential precipitation with CTAB. Briefly, capsular polysaccharides isolated from the culture supernatant by precipitation with ethanol were dissolved in 0.2 M NaCl (10 mg ml-1) and CTAB (3 mg mg-1 of polysaccharides) was added slowly. Then, a solution of 0.05% CTAB was added and the GXM was selectively precipitated. The precipitate was collected by centrifugation and washed with 2% acetic acid in ethanol then in 90% ethanol. The precipitate was dissolved in 1 M NaCl, and three volumes of ethanol were added to precipitate the GXM. The precipitate was centrifuged, washed with 2% acetic acid in ethanol and then with ethanol, dissolved in water and lyophilized.
To eliminate potential LPS contamination, 10 mg of GXMGal or GXM preparations were dissolved in LPS-free water and further purified through chromatography on a column of Polimixin B-Agarose (Sigma) equilibrated with LPS free-water. Purified GXMGal or GXM were eluted with 12 mL of LPS free water, recovered and lyophilized.
2.3. Cytotoxicity Assay
The verification of the possible toxic effects of capsular polysaccharides (GXMGal, GXM and total capsule) on canine macrophages DH82 was performed using the XTT colorimetric assay (Sigma), which quantifies mitochondrial metabolism and respiratory chain activity in the presence of the electron acceptor PMS. DH82 cells (1x105/well) were cultured in 96-well plates and incubated with GXMGal, GXM and total capsule at concentrations of 10, 50 and 100 µg/mL for 24 h and 48 h. After incubation, an XTT solution (1.8 mg XTT, 75 µL PMS and 1425 µL PBS) was prepared and 50 µL was added per well. Cultures were incubated for 3 h and quantification was performed using a 490 nm filter in a Spectramax M3 reader.
2.4. Assessment of Binding and Phagocytosis
DH82 cells were plated on glass coverslips (1x104 cells/well) and incubated overnight with different concentrations (10, 50 and 100 µg/mL) of GXM, GXMGal and total capsule. The used concentrations of polysaccharides were based on our previous work [11,12]. After that, Saccharomyces cerevisiae yeast was added to the cells in a 10:1 ratio for 40 minutes (binding assay) or 4 h (phagocytosis assay) and incubated at 37 0C and 5% CO2. After incubation, the cells were washed twice with PBS to remove non- binding and non-phagocytosed yeasts. The coverslips were washed with HBSS, fixed with methanol, and stained with Diff-Quick (Thermo Fisher, Waltham, MA, USA). Yeasts were counted at 100x oil immersion on a (Olympus) microscope. The number of yeasts was estimated in 100 cells per coverslip, and the frequency was compared among six coverslips per time point.
2.5. Viability of Yeasts after Phagocytosis
Canine DH82 macrophages were grown in 24-well culture plates (1x104 cells/well) and incubated with the capsular polysaccharides and total capsule for 24 h at the concentrations described above. After this period, S. cerevisiae yeasts were added at a ratio of 10:1 (yeasts/macrophage) and incubated for 4 h. Subsequently, the cultures were washed twice with PBS and lysed with ice water (500 µL). The lysate was diluted 1:10 in 1x PBS. The 30 µL volume of the diluted lysate was seeded in Petri dishes containing Sabouraud agar. The plates were incubated at 37 0C for 48 h and then the colony forming units were counted [16].
2.6. Reactive Oxygen Species (ROS) Detection
The quantification of ROS intracellular levels was made by oxidation of non-fluorescent 2′,7′-dichlorofluorescein probe, delivered as diacetate form (DCFH-DA), to the fluorescent product 2′,7′-dichlorofluorescein [17]. Macrophages DH82 were seeded in 96-well plate (5x104 cells/well), incubated with polysaccharides (GXMGal, GXM and capsule) 100 µg/mL, stimulated or not with LPS (400 ng/mL, Sigma-Aldrich) and recombinant IFN-γ (1.5 ng/mL, Serotech) and after 24 h later cells were washed and loaded with 10 μM DCFH-DA (Invitrogen) for 20 minutes at 37 0C. Untreated and unstimulated cells were used as a negative control. After incubation, the cells were washed, and fluorescence was measured (excitation = 485 nm excitation; λ emission = 535 nm) in an FLx800 Fluorescence Microplate Reader (BioTek).
2.7. Real-Time RT-PCR Quantification
DH82 macrophage (1x106 cells) was plated and incubated with GXMGal, GXM and capsule (100 µg/mL), stimulated or not with LPS (400 ng/mL) and IFN-γ (1.5 ng/mL) for 24 h. After this time, total RNA was extracted with RNeasy Plus Mini Kit (Qiagen 74134, Germany), 1 μg aliquot was reverse transcribed to first-strand cDNA with ImProm-II (Promega) and oligo(dT) primer according to the manufacturer's instructions. The DNA sequences of the primers used were IL-6-F: 5′-GCGTCTTCCCTCATGACC-3′, IL-12-R: 5′-GGGTGCCAGTCCAACTCTAC-3′, IL-6-F:5′-GGGAAAGCAGTAGCCATCAC-3′,IL-6-R:5′CAGGACCCCAGCTATGAACT-3′,TGF-β-F:5′-CGAAGCCCTCGACTCC-3′,TGF-β-R:5′TGGCTGYCCTTTGATGTCAC-3′, GAPDH-F: 5′-TGCACCACCAACTGCTTAGC-3′ and GAPDH-R: 5′-GGCATGGACTGTGGTCATGAG-3′. qRT-PCR data from the experiments were normalized using Gapdh primers as an endogenous control. Amplicon specificity was carefully verified by the presence of a single melting temperature peak in dissociation curves run after real-time RT-PCR. Real-time quantitative RT-PCR (qRT-PCR) was performed via the Applied Biosystems StepOne™ detection system (Applied Biosystems) using GoTaq® qPCR Master Mix (Promega Corp., Madison, WI, USA). All expression ratios were computed via the analysis of relative gene expression ΔΔCt method through the StepOne software version 2.0 (Applied Biosystems).
All real-time PCR reactions were previously standardized. Including both the concentration of primers in relation to the melting curve and a standard curve between the endogen used and the targets, to achieve efficiency across all targets. It was only after these tests that we carried out the analysis of the relative expression in relation to the endogenous one, which in this case is GAPDH. Among all samples tested, the Ct (“Cycle Threshold”) was similar and close to less than 1 log, thus being considered a good endogenous target to normalize during reactions.
2.8. MHC Class II Expression
DH82 macrophages were grown in 6-well culture plates (1x106 cells/well) and incubated for 24 h with the capsular polysaccharides (100 µg/mL), LPS (400 ng/mL) and IFN-γ (1.5 ng /mL). After 24 h, the cells were detached, washed, adjusted to a concentration of 5x105 cells/tube, and incubated with blocking buffer (CD16/CD32 Fc Block-eBioscience) for 15 min on ice to prevent non-specific antibody binding to Fc receptors. Cells were stained with anti-dog MHC class II-FITC (Serotech). All washing steps were performed with PBS containing 3% FCS and 0.02% sodium azide. Data was acquired (10,000 events), evaluated on FACSCalibur™ cytometer and analyzed using CellQuest® software (BD Biosciences, Heidelberg, Germany).
2.9. Assay of Protein Phosphorylation
DH82 macrophages were grown in 6-well culture plates (1x106cells/well) and incubated for 24 h with the polysaccharides GXMGal, GXM and total capsule (100 µg/mL), stimulated with canine IFN-γ (1.5 ng/mL) and LPS (400 ng/ml). The western blotting protocol has been described [18]. The following antibodies were used: anti-ERK (4696S - Cell Signaling), anti-pERK (4370S-CellSignaling), anti-PPAR-γ (2435 - Cell Signaling) and anti-β actin (Sigma-Aldrich) as primary antibody and anti-IgG (whole molecule)-peroxidase (Sigma-Aldrich) as secondary antibody. Reactions were developed using a chemiluminescence kit (ECL Western Blotting Substrate-Promega-w1015) according to the manufacturer's instructions. The band densitometry of western blotting was analyzed by the Scion Image software 4.0.3.2.
3. Results
3.1. Purified Polysaccharides and the Total Capsule Have No Toxic Effects on Canine Macrophages DH82
Capsular polysaccharides from C. neoformans were previously described as inducers of apoptosis in murine macrophages [11]. Thus, it was initially analyzed whether the purified polysaccharides would have any toxic effect on canine macrophages DH82 cell line.
For this, cell cultures of the DH82 macrophage were grown and treated with GXMGal, GXM or C. neoformans total capsule at different concentrations and incubated for 24 h and 48 h. After the times established by the assay, they were analyzed by the XTT test, which evaluates the mitochondrial activity of the cells. We observed in our experiments that the polysaccharides and the total capsule showed no impairment of the viability of DH82 macrophages after 24 h (Figure 1A) or 48 h of incubation (Figure 1B).
3.2. Purified Polysaccharides from C. neoformans Inhibit Binding and Phagocytic Activity of Canine Macrophages DH82
The ability to binding and be phagocytosed is critical for the elimination of infectious agents [19]. As binding and phagocytosis is one of the main activities of this cell type, we started to evaluate the effect of the contact of DH82 cells with the polysaccharides GXMGal, GXM or the total capsule on the mechanisms of binding and the ability of macrophages to phagocytose. For this, DH82 cells were maintained in culture in the presence of purified polysaccharides or the total capsule and, later, incubated with Saccharomyces cerevisiae yeasts for 40 min. After incubation, we observed that the cells treated with the polysaccharides showed a reduction in the binding capacity, demonstrated by the lower number of yeasts attached to the surface of DH82 cells (Figure 2A).
A similar phenomenon was observed in relation to the phagocytic activity of the treated DH82 cells. We established those cells in the presence of polysaccharides showed a significant reduction in phagocytosis. This was demonstrated by the decrease in the number of cells that phagocytize yeast (Figure 2B) and in the phagocytosed yeast by macrophage (Figure 2C). Our results indicate that contact with C. neoformans polysaccharides may decrease the ability of DH82 cells to binding to particles and subsequent phagocytosis.
3.3. Yeasts Recovered from DH82 Macrophages Treated with Polysaccharides Show Increased Growth
Since we observed an inhibition of phagocytosis in the presence of polysaccharides, we decided to evaluate if the yeasts that were phagocytized would be viable or if the possible modulation of macrophages would have supported their viability.
DH82 cells were plated and incubated with GXMGal, GXM or the whole capsule. Then, S. cerevisiae was added to the culture and incubated for a period of 4 h. The phagocytosed yeasts were recovered through macrophage lysis and seeded on Sabouraud agar, using the protocol previously described [16]. A greater number of colonies forming units (CFU) of yeasts recovered from DH82 cells that were treated with the polysaccharides or the total capsule was observed. Yeasts recovered from cells treated with C. neoformans polysaccharides at a concentration of 100 μg/mL showed significant growth when compared with yeasts recovered from cells not treated with polysaccharides (Figure 3), indicating a possible change in the microbicidal mechanisms of the canine macrophage.
3.4. Polysaccharides from C. neoformans Induce the Production of Reactive Oxygen Species in Canine Macrophage Lineage DH82
An important mechanism for macrophage microbicidal function is the production of reactive oxygen species (ROS). Therefore, we evaluated the effect of treatment with purified polysaccharides and the total capsule on ROS production in DH82 macrophages. For this, DH82 macrophages were plated in the presence of purified polysaccharides or the total capsule of C. neoformans and incubated with a fluorescent probe that measures the intracellular concentration of ROS. The result was obtained after 24 h of incubation (Figure 4). Surprisingly, we observed that GXMGal, GXM and the total capsule were able to induce a significant decrease in ROS production of cells activated with LPS and IFN-γ. Indicating that this pathway was probably strongly modulated in the presence of fungal polysaccharides, which may have compromised the ability to kill yeast.
3.5. GXMGal Purified from C. neoformans Modulates the Expression of MHC Class II Molecules
Another way of evaluating the role of the stimulus that the polysaccharides exert on the DH82 cell is through the evaluation of the expression of surface molecules, especially cellular markers associated with antigen presentation, such as MHC Class II (MHC-II) [20]. DH82 macrophages were plated and incubated with GXMGal, GXM or the total capsule, in the presence or absence of stimulation with LPS and IFN-γ, so that the expression of the MHC-II molecule was evaluated by flow cytometry. Our results showed that polysaccharides were not able to alter MHC-II expression in non-activated cells. In the presence of additional LPS and IFN-γ stimuli, GXMGal induced a statistically significant reduction in the expression of these molecules (Figure 5A and 5B). Our results suggest that cells in contact with the capsular polysaccharides of C. neoformans do not change the expression of MHC-II, however if they receive an activating stimulus, they seem to lose the ability to respond at full capacity.
3.6. Purified Polysaccharides from C. neoformans Alter Cytokine Production by DH82 Macrophages
It has already been shown that C. neoformans polysaccharides can modulate the production of pro- or anti-inflammatory cytokines in murine macrophages and dendritic cells [11,15]. Likewise, we evaluated the role of polysaccharides and the total capsule in the expression of pro-inflammatory cytokines, such as IL-12 and IL-6, as well as the expression of the anti-inflammatory cytokine TGF-β. DH82 macrophages were cultured for 24 h in the presence of GXMGal, GXM and the total capsule, stimulated or not with LPS and IFN-γ and the cytokine expression were quantified from the lysate of the cultures by the RT-PCR technique. The analysis of our results showed that in DH82 macrophages treated with the polysaccharides there was alteration in the expression of IL-12, IL-6 and TGF-β (Figure 6A, 6B and 6C). The expression of IL-12 and IL-6 was not altered in non-stimulated cells, however, when they were stimulated with LPS and IFN-γ in the presence of polysaccharides, we observed a decrease in IL-12 expression by 1.9 times in the presence of GXMGal, 1.5 times in the presence of GXM and 1.8 times in the presence of total capsule (Figure 6A). Similarly, we observed a 1.6-fold decrease in IL-6 expression in stimulated macrophages in the presence of GXMGal, 1.4-fold in the presence of GXM, and 1.7-fold in the presence of total capsule (Figure 6B). Interestingly, TGF-β expression was increased 1.4 times in the presence of GXMGal, 1.8 times in the presence of GXM and 1.9 times in the presence of total capsule in unstimulated macrophages. However, there was a reduction of this expression in stimulated cells where we observed a decrease in TGF-β expression by 1.2 times in the presence of GXMGal, 1.1 times in the presence of GXM and 1.2 in the presence of total capsule (Figure 6C).
The results obtained analyzing the m-RNA expression of cytokines in DH82 cells suggest that the contact with the capsular polysaccharides of C. neoformans could negatively modulate the production of pro-inflammatory cytokines, directing the cells to an anti-inflammatory profile that would contribute to the permanence of yeasts inside macrophages.
3.7. Purified Polysaccharides from C. neoformans Modulate ERK Phosphorylation and PPAR-γ Expression
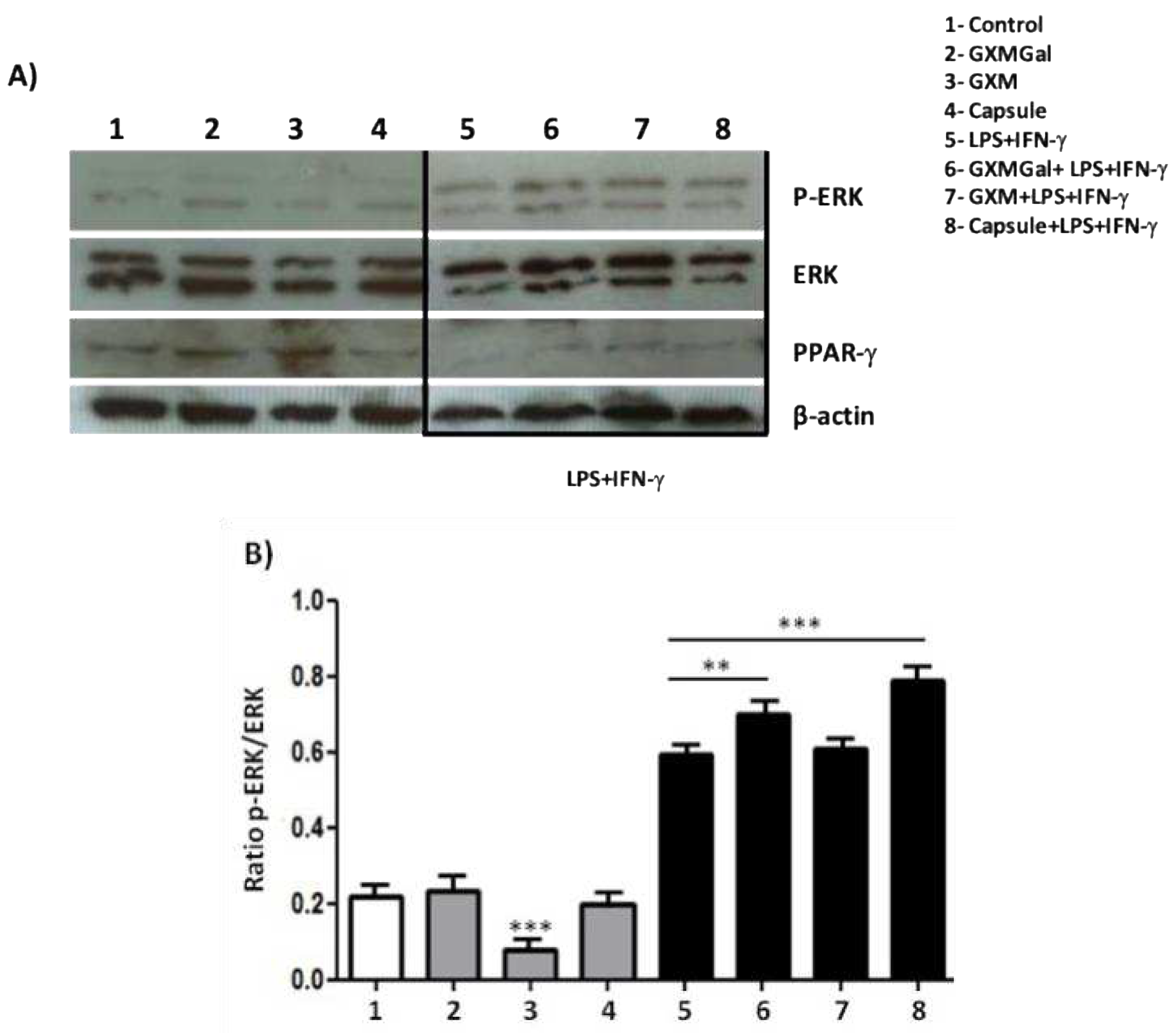
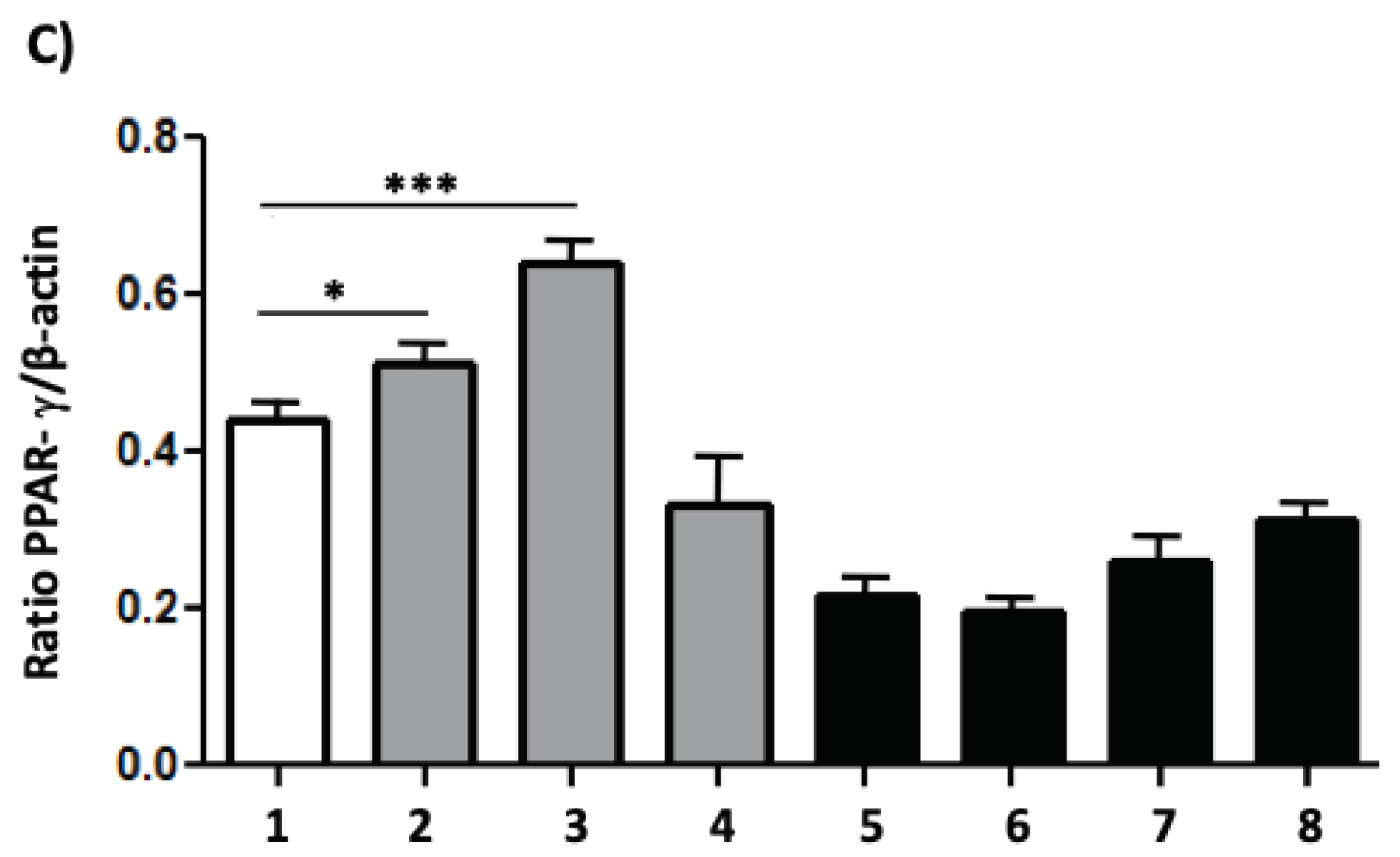
To evaluate the effects of treatment with polysaccharides or total capsule of C. neoformans on intracellular signaling of DH82 cells, we decided to analyze the phosphorylation of the ERK protein kinase, which is fundamental in the processes of macrophage activation. In addition to protein kinases, some nuclear factors may be related to suppressive phenomena in macrophages [21,22]. Thus, we decided to evaluate whether the expression of the nuclear factor PPAR-γ could be a target of capsular polysaccharides. We performed assays with the aim of verifying whether the modulation of ERK phosphorylation or PPAR-γ expression could be responsible for the tendency to inhibit important microbicidal activities for macrophages such as phagocytosis and expression of cytokines with a pro-inflammatory profile that we have observed so far. For this, DH82 cells were incubated with GXMGal, GXM or total capsule, in the presence or absence of LPS and IFN-γ, for 24 h. After incubation, the protein extract was analyzed by the immunoblot technique. Analyzing ERK phosphorylation, our results showed that unstimulated macrophages in the presence of GXM had a statistically significant decrease in ERK phosphorylation. However, when DH82 cells were activated with LPS and IFN-γ, we observed increased ERK phosphorylation in the presence of GXMGal, and total capsule (Figure 7A and 7B). This suggests that non-activated DH82 cells can be negatively modulated by GXM, while activated cells in contact with polysaccharides increase ERK-dependent signaling. An antagonistic panorama was observed in the expression of PPAR-γ. DH82 macrophages have the expression of this nuclear factor increased in the presence of GXM and GXMGal (Figure 7A and 7C), suggesting that DH82 cells in the presence of these polysaccharides may be directed to acquire an inhibitory profile, which would support the permanence of the fungus in the organism.
4. Discussion
Different models been extensively studied to reveal the mechanisms of interaction between host cells and parasitic agents [23,24,25]. For many infectious agents, the type of interaction with host cells can determine the resolution or success of the infection. Among the host cells, macrophages stand out for their importance in innate and adaptive immunity [8,25]. Cryptococcus neoformans is an infectious agent capable of replicating within macrophages and this has been reported by several authors [26,27,28]. Furthermore, it was demonstrated that macrophages could be used by C. neoformans as a Trojan horse to transport yeast to strategic points in the body, such as the CNS [7,29].
Although several studies point to the relevance of the interaction of macrophages with C. neoformans, there is no study carried out with the aim of understanding the effect of the interaction of the capsular components of C. neoformans with canine macrophages. In this work we used the canine macrophage DH82 cell line and our group has demonstrated the importance of this cell line in the interaction with parasites as a replacement method [30,31]. Using DH82 canine macrophages treated with purified C. neoformans polysaccharides, we demonstrated for the first time their immunomodulatory effects. Previous work has shown that purified polysaccharides from C. neoformans can lead to death by apoptosis of murine macrophages [11]. Thus, we first treated canine macrophages DH82 with different concentrations of GXMGal, GXM and total capsule with incubation of 24 and 48h to see if the polysaccharides would have any toxic effect on canine macrophages. Surprisingly, it was not possible to observe a change in cell viability at any of the concentrations used. This data suggests that macrophages from different species may show variations in resistance to toxic effects resulting from contact with capsular polysaccharides.
The ability to binding and phagocytose infectious agents is a fundamental microbiological function for macrophages. However, it is known that components of the C. neoformans capsule inhibit phagocytic activity and may compromise efficient elimination by this mechanism [16,32]. To assess whether purified polysaccharides would reproduce this effect, we incubated canine macrophages DH82 with GXMGal, GXM and whole capsule and offered Saccharomyces cerevisiae yeasts. We observed that the polysaccharides were able to inhibit the ability of macrophages to binding to yeasts, as well as phagocytose. The polysaccharides indistinctly led to a decrease in phagocytic activity. We observed a decrease in the number of yeasts attached to macrophages, number of cells that phagocytosed and number of phagocytosed yeasts. This data corroborates findings from other authors who previously showed that the C. neoformans capsule can inhibit phagocytosis by macrophages through various mechanisms [33,34,35].
Based on the observation that GXMGal, GXM and the total capsule of C. neoformans influenced phagocytosis, we investigated whether phagocytosed S. cerevisiae yeasts would undergo fungicidal action by DH82 macrophages by counting CFU of recovered yeasts after phagocytosis. Experiments performed on murine macrophages [16], suggest vesicles secreted by C. neoformans influence the amount of CFU produced by recovered yeasts after being phagocytosed. In our experiment, the high number of yeast colonies recovered from cells treated (100 µg/mL) with polysaccharides suggests a relationship with a modulating action of the microbicidal activities of the macrophage by the polysaccharides, consistent with previous work that showed inhibition of the production of pro-inflammatory cytokines [11] and can be extrapolated to the physiological model, where the impairment of macrophages is related to inhibition of the immune system leading to a greater fungal burden [4,5,11,14,36]. The presence of numerous CFU in macrophages with polysaccharides suggests that they decrease the macrophage's fungicidal activity.
Despite the reduced rate of phagocytosis in the presence of the capsular polysaccharides of C. neoformans, it was shown that the yeasts phagocytosed by these macrophages remained viable and able to grow. For this reason, we analyzed a microbicidal mechanism of oxidant production by phagocytes. ROS play a crucial role in protection against fungal infections, as observed in patients with chronic granulomatous disease (CGD) which are very susceptible to fungal infections [37,38]. Furthermore, the decrease in ROS production may compromise the differentiation of T lymphocytes to the Th1 profile, which is the protective profile [39]. ROS is a factor that participates in the inflammatory process observed in canine cryptococcosis associated with excessive granuloma formation [3,40,41]. Our results show that polysaccharides were able to inhibit ROS production in DH82 macrophages, suggesting that the inhibitory effect on ROS production may have favored yeast survival.
After phagocytosis, within the phagosome, yeasts are under the action of low pH, ROS, reactive nitrogen species, and nutrients deprivation [42]. These challenges are neutralized by yeast through powerful mechanisms. Phagocytosed yeasts can upregulate gene expression of oxidative stress enzymes, starvation responses, and the autophagic machinery [43,44]. In a model of NADPH oxidase-null mice, cryptococcal infection is contained and the fungal load in both brain and lung is decreased [39], suggesting that inflammatory ROS are prejudicial also to the host.
Although ROS act in the death of fungi, this usually happens in a pro-inflammatory environment [45], in our model, where canine macrophages treated with polysaccharides showed ROS inhibition, this suppressor environment, points to the important role of ROS in the microbicidal activity of DH82 canine macrophages.
Macrophages act in the processing and presentation of antigens, providing the adaptive immune response. Thus, the expression of MHC molecules is crucial, because through the expression of MHC-II, macrophages are upregulated, acting as APC and functioning as fundamental elements in the activation of lymphocytes, which, when stimulated, will lead to the adaptative immune response [46]. DH82 macrophages treated with capsular polysaccharides did not increase the basal expression of MHC-II, contributing to the non-responsiveness of the immune system imposed by C. neoformans on the host [47]. The basal expression of MHC-II presented in our results is similar to that found previously [48], but slightly lower [49,50]. DH82 canine macrophages stimulated with LPS and IFN-γ increase the expression of MHC-II, due to the already known capacity that these factors exert on the increase of the expression of this protein [49,50]. However, we observed statistically significant decrease in MHC class II expression in the presence of GXMGal, indicating a possible suppression of the DH82 canine macrophage response mediated by this purified C. neoformans polysaccharide, data shown for the first time in this cell type, since most of the works refer to GXM [14]. However, it is known that the C. neoformans cell induces the expression of MHC-II in glial cells when in contact with the fungus [51].
Macrophages produce cytokines that mediate and direct the immune response to an infection. In cryptococcosis, the crucial role of this function has been demonstrated, the polarization of macrophages, through the cytokine profile, influencing the course of infection by C. neoformans [52]. We evaluated the cytokines IL-12 and IL-6 that help protect the host and TGF-β that can contribute to the permanence of the pathogen, since cryptococcosis is a balance infection between Th1 and Th2 profiles [53].
Our results show that GXMGal, GXM and the total capsule inhibit the expression of IL-12, IL-6 and TGF-β. Although non-stimulated DH82 macrophages do not alter the expression of these cytokines, we clearly saw that when stimulated these cells have a significant reduction in the expression of IL-12 and IL-6. On the other hand, TGF-β had its expression increased in non-stimulated cells and reduced in response to stimuli. This modulation in cytokine production suggests that the increase in TGF-β in initial moments of contact with capsular polysaccharides may compromise the production of pro-inflammatory cytokines in canine DH82 macrophages. The inhibition of pro-inflammatory cytokines such as IL-12, which is fundamental in autocrine signaling in the activation of macrophages [54], as well as the inhibition of IL-6 can be determinant not only for macrophage activation, but also in the induction of the adaptive response [55]. Thus, the action of polysaccharides may compromise the activation of macrophages. Previous works have demonstrated the anti-inflammatory role being predominantly exerted by GXM [11,47], however, our results suggest that, for canine macrophages DH82, GXMGal, GXM and total capsule exert the same inhibitory effect on the expression of the evaluated cytokines.
The results presented so far lead to the profile of canine DH82 macrophage suppression, with this, we investigated the intracellular signaling in the face of stimulation with the capsular polysaccharides of C. neoformans. The results obtained show that GXMGal, GXM or the total capsule do not induce ERK phosphorylation, which occurs in the presence of LPS and IFN-γ. We also observed a decrease in cell phosphorylation in the presence of GXM, showing once again the suppressive characteristic exerted by polysaccharides, since ERK is part of the large family of MAPK that are involved in several cellular mechanisms, including proliferation and cell activation upon stimulation [56,57]. Another intracellular marker investigated was PPAR-γ, a nuclear factor that is activated in the presence of suppressive stimuli [21,58]. Based on the already known activation that LPS and IFN-γ cause in macrophages, we observed through the increase in PPAR-γ expression that capsular polysaccharides, especially GXM of C. neoformans induce the suppression of canine macrophages DH82.
All aspects evaluated in the present study, such as decreased phagocytosis, inhibition of ROS, IL-12 and IL-6 production, increased TGF-β production, combined with decreased MHC class II expression, suggest that polysaccharides lead to the impairment of fundamental microbicidal activities of macrophages. In addition, the evaluation of intracellular signaling revealed that in the presence of polysaccharides there is a reduction in ERK phosphorylation and an increase in PPAR-γ expression, reinforcing the suppressor aspect exerted by the polysaccharides on canine macrophages DH82. These data may contribute to the understanding that the impairment of the microbicidal activities of macrophages could contribute to the permanence and spread of the fungus in the canine host.
5. Conclusions
The present work suggests a suppressive role of capsular polysaccharides from C. neoformans in canine macrophages DH82. Through this canine macrophage suppression profile, we can infer that the fungus could remain viable within the macrophage and capable of triggering a systemic infection. The macrophage showing signs of suppression such as decreased phagocytosis, inhibition of ROS, decrease of the expression of pro-inflammatory cytokines and increased expression of PPAR-γ would be able to carry C. neoformans to reach different sites of the host, including the CNS, because it is a cell able to cross the blood-brain barrier.
We demonstrate for the first time the modulatory role of capsular polysaccharides from C. neoformans acting on cells of canine origin. We understand the importance of more detailed evaluations on DH82 cells. In addition, a need of further confirmation of these data in canine macrophages obtained from peripheral blood.
Author Contributions
D.D.-R., C.G.F.-d.-L., I.F.L-F. and E.B.S.J. wrote the main manuscript text; I.F.L-F., E.B.S.J., L.P.G., B.S.D.L., A.C.V., U.G.L., L.F.-d-L. and D.D.-R. prepared all figures; I.F.L-F., E.B.S.J., L.P.G., B.S.D.L., A.C.V., A.M., J.O.P., L.M.-P.,C.G.F.-d.-L., and D.D.-R. conducted the experiments; I.F.L-F., E.B.S.J., A.C.V., A.M, J.O.P., L.M.-P.,C.G.F.-d.-L., and D.D.-R. analyzed the data and revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Brazilian National Research Council (CNPq), Rio de Janeiro State Science Foundation (FAPERJ) and Fundação Oswaldo Cruz (FIOCRUZ).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data is contained within this article.
Acknowledgments
This work was supported by the Brazilian National Research Council (CNPq), Rio de Janeiro State Science Foundation (FAPERJ) and Fundação Oswaldo Cruz (FIOCRUZ). We thank Renata Sant’Anna Rezende Ribeiro and Lindomar Miranda for helpful technical assistance.
References
- Trivedi, S.R.; et al. Clinical features and epidemiology of cryptococcosis in cats and dogs in California: 93 cases (1988-2010). J Am Vet Med Assoc 2011, 239, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Malik, R.; et al. Cryptococcosis in dogs: a retrospective study of 20 consecutive cases. J Med Vet Mycol 1995, 33, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Sykes, J.E.; et al. Clinical signs, imaging features, neuropathology, and outcome in cats and dogs with central nervous system cryptococcosis from California. J Vet Intern Med 2010, 24, 1427–1438. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.K.; et al. Monocyte/macrophage traffic in HIV and SIV encephalitis. J Leukoc Biol 2003, 74, 650–656. [Google Scholar] [CrossRef] [PubMed]
- Coelho, C.; Bocca, A.L.; Casadevall, A. The intracellular life of Cryptococcus neoformans. Annu Rev Pathol 2014, 9, 219–238. [Google Scholar] [CrossRef]
- Kechichian, T.B.; Shea, J.; Del Poeta, M. Depletion of alveolar macrophages decreases the dissemination of a glucosylceramide-deficient mutant of Cryptococcus neoformans in immunodeficient mice. Infect Immun 2007, 75, 4792–4798. [Google Scholar] [CrossRef] [PubMed]
- Charlier, C.; et al. Evidence of a role for monocytes in dissemination and brain invasion by Cryptococcus neoformans. Infect Immun 2009, 77, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Jung, E.H.; et al. Cryptococcus neoformans releases proteins during intracellular residence that affect the outcome of the fungal-macrophage interaction. Microlife 2022, 3, uqac015. [Google Scholar] [CrossRef] [PubMed]
- Decote-Ricardo, D.; et al. Immunomodulatory Role of Capsular Polysaccharides Constituents of Cryptococcus neoformans. Front Med (Lausanne) 2019, 6, 129. [Google Scholar] [CrossRef]
- McClelland, E.E.; Bernhardt, P.; Casadevall, A. Estimating the relative contributions of virulence factors for pathogenic microbes. Infect Immun 2006, 74, 1500–1504. [Google Scholar] [CrossRef]
- Villena, S.N.; et al. Capsular polysaccharides galactoxylomannan and glucuronoxylomannan from Cryptococcus neoformans induce macrophage apoptosis mediated by Fas ligand. Cell Microbiol 2008, 10, 1274–1285. [Google Scholar] [CrossRef] [PubMed]
- Rocha, J.D.; et al. Capsular polysaccharides from Cryptococcus neoformans modulate production of neutrophil extracellular traps (NETs) by human neutrophils. Sci Rep 2015, 5, 8008. [Google Scholar] [CrossRef]
- Monari, C.; et al. Microbial immune suppression mediated by direct engagement of inhibitory Fc receptor. J Immunol 2006, 177, 6842–6851. [Google Scholar] [CrossRef] [PubMed]
- Retini, C.; et al. Encapsulation of Cryptococcus neoformans with glucuronoxylomannan inhibits the antigen-presenting capacity of monocytes. Infect Immun 1998, 66, 664–669. [Google Scholar] [CrossRef] [PubMed]
- LaRocque-de-Freitas, I.F.; et al. Involvement of the capsular GalXM-induced IL-17 cytokine in the control of Cryptococcus neoformans infection. Sci Re 2018, 8, 16378. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.L.; et al. Extracellular vesicles from Cryptococcus neoformans modulate macrophage functions. Infect Immun 2010, 78, 1601–1609. [Google Scholar] [CrossRef] [PubMed]
- Sundaresan, M.; et al. Requirement for generation of H2O2 for platelet-derived growth factor signal transduction. Science 1995, 270, 296–299. [Google Scholar] [CrossRef] [PubMed]
- Benschop, R.J.; et al. Activation and anergy in bone marrow B cells of a novel immunoglobulin transgenic mouse that is both hapten specific and autoreactive. Immunity 2001, 14, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; et al. Reduced phagocytosis and killing of Cryptococcus neoformans biofilm-derived cells by J774.16 macrophages is associated with fungal capsular production and surface modification. Fungal Genet Biol 2019, 132, 103258. [Google Scholar] [CrossRef]
- Kotsias, F.; Cebrian, I.; Alloatti, A. Antigen processing and presentation. Int Rev Cell Mol Biol 2019, 348, 69–121. [Google Scholar]
- Johann, A.M.; et al. Recognition of apoptotic cells by macrophages activates the peroxisome proliferator-activated receptor-gamma and attenuates the oxidative burst. Cell Death Differ 2006, 13, 1533–1540. [Google Scholar] [CrossRef]
- Freire-de-Lima, C.G.; et al. Apoptotic cells, through transforming growth factor-beta, coordinately induce anti-inflammatory and suppress pro-inflammatory eicosanoid and NO synthesis in murine macrophages. J Biol Chem 2006, 281, 38376–38384. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Díaz, R.A.; et al. Biological characterization of Trypanosoma cruzi strains. Mem Inst Oswaldo Cruz 2001, 96, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Bogdan, C.; Röllinghoff, M. How do protozoan parasites survive inside macrophages? Parasitol Today 1999, 15, 22–28. [Google Scholar] [CrossRef]
- Bauman, S.K.; Nichols, K.L.; Murphy, J.W. Dendritic cells in the induction of protective and nonprotective anticryptococcal cell-mediated immune responses. J Immunol 2000, 165, 158–167. [Google Scholar] [CrossRef] [PubMed]
- De Leon-Rodriguez, C.M.; et al. The Capsule of Cryptococcus neoformans Modulates Phagosomal pH through Its Acid-Base Properties. mSphere 2018, 3. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, A.S.; Wheeler, R.T.; May, R.C. Fungal Pathogens: Survival and Replication within Macrophages. Cold Spring Harb Perspect Med 2014, 5, a019661. [Google Scholar] [CrossRef] [PubMed]
- Mansour, M.K.; et al. Macrophage Cryptococcus interactions: an update. Curr Fungal Infect Rep 2014, 8, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Santiago-Tirado, F.H.; et al. Trojan Horse Transit Contributes to Blood-Brain Barrier Crossing of a Eukaryotic Pathogen. mBio 2017, 8. [Google Scholar] [CrossRef]
- Mendonça, P.H.B.; et al. Canine Macrophage DH82 Cell Line As a Model to Study Susceptibility to Trypanosoma cruzi Infection. Front Immunol 2017, 8, 604. [Google Scholar] [CrossRef]
- Nadaes, N.R.; et al. DH82 Canine and RAW264.7 Murine Macrophage Cell Lines Display Distinct Activation Profiles Upon Interaction With Leishmania infantum and Leishmania amazonensis. Front Cell Infect Microbiol 2020, 10, 247. [Google Scholar] [CrossRef]
- Feldmesser, M.; Tucker, S.; Casadevall, A. Intracellular parasitism of macrophages by Cryptococcus neoformans. Trends Microbiol 2001, 9, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Liu, O.W.; et al. Systematic genetic analysis of virulence in the human fungal pathogen Cryptococcus neoformans. Cell 2008, 135, 174–188. [Google Scholar] [CrossRef] [PubMed]
- Stano, P.; et al. App1: an antiphagocytic protein that binds to complement receptors 3 and 2. J Immunol 2009, 182, 84–91. [Google Scholar] [CrossRef] [PubMed]
- García-Rodas, R.; Zaragoza, O. Catch me if you can: phagocytosis and killing avoidance by Cryptococcus neoformans. FEMS Immunol Med Microbiol 2012, 64, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Murphy, J.W.; Cozad, G.C. Immunological unresponsiveness induced by cryptococcal capsular polysaccharide assayed by the hemolytic plaque technique. Infect Immun 1972, 5, 896–901. [Google Scholar] [CrossRef] [PubMed]
- Pollock, J.D.; et al. Mouse model of X-linked chronic granulomatous disease, an inherited defect in phagocyte superoxide production. Nat Genet 1995, 9, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Mesa-Arango, A.C.; et al. The production of reactive oxygen species is a universal action mechanism of Amphotericin B against pathogenic yeasts and contributes to the fungicidal effect of this drug. Antimicrob Agents Chemother 2014, 58, 6627–6638. [Google Scholar] [CrossRef]
- Snelgrove, R.J.; et al. In the absence of reactive oxygen species, T cells default to a Th1 phenotype and mediate protection against pulmonary Cryptococcus neoformans infection. J Immunol 2006, 177, 5509–5516. [Google Scholar] [CrossRef]
- Lavely, J.; Lipsitz, D. Fungal infections of the central nervous system in the dog and cat. Clin Tech Small Anim Pract 2005, 20, 212–219. [Google Scholar] [CrossRef]
- Vorathavorn, V.I.; Sykes, J.E.; Feldman, D.G. Cryptococcosis as an emerging systemic mycosis in dogs. J Vet Emerg Crit Care (San Antonio) 2013, 23, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Shoham, S.; Levitz, S.M. The immune response to fungal infections. Br J Haematol 2005, 129, 569–582. [Google Scholar] [CrossRef] [PubMed]
- Derengowski Lda, S.; et al. The transcriptional response of Cryptococcus neoformans to ingestion by Acanthamoeba castellanii and macrophages provides insights into the evolutionary adaptation to the mammalian host. Eukaryot Cell 2013, 12, 761–774. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; et al. Cryptococcus neoformans gene expression during murine macrophage infection. Eukaryot Cell 2005, 4, 1420–1433. [Google Scholar] [CrossRef] [PubMed]
- Husain, N.; Mahmood, R. Copper(II) generates ROS and RNS, impairs antioxidant system and damages membrane and DNA in human blood cells. Environ Sci Pollut Res Int 2019, 26, 20654–20668. [Google Scholar] [CrossRef] [PubMed]
- Ramachandra, L.; Simmons, D.; Harding, C.V. MHC molecules and microbial antigen processing in phagosomes. Curr Opin Immunol 2009, 21, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Casadevall, A.; et al. The capsule of Cryptococcus neoformans. Virulence 2019, 10, 822–831. [Google Scholar] [CrossRef] [PubMed]
- Barnes, A.; et al. Immunological and inflammatory characterisation of three canine cell lines: K1, K6 and DH82. Vet Immunol Immunopathol 2000, 75, 9–25. [Google Scholar] [CrossRef] [PubMed]
- Harrus, S.; et al. Down-regulation of MHC class II receptors of DH82 cells, following infection with Ehrlichia canis. Vet Immunol Immunopathol 2003, 96, 239–243. [Google Scholar] [CrossRef]
- Wasserman, J.; et al. Suppression of canine myeloid cells by soluble factors from cultured canine tumor cells. Vet Immunol Immunopathol 2012, 145, 420–430. [Google Scholar] [CrossRef]
- Aguirre, K.; Miller, S. MHC class II-positive perivascular microglial cells mediate resistance to Cryptococcus neoformans brain infection. Glia 2002, 39, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.J.; et al. Macrophage M1/M2 polarization dynamically adapts to changes in cytokine microenvironments in Cryptococcus neoformans infection. mBio 2013, 4, e00264–13. [Google Scholar] [CrossRef] [PubMed]
- Li, S.S.; Mody, C.H. Cryptococcus. Proc Am Thorac Soc 2010, 7, 186–196. [Google Scholar] [CrossRef] [PubMed]
- Pillarisetty, V.G.; et al. Natural killer dendritic cells have both antigen presenting and lytic function and in response to CpG produce IFN-gamma via autocrine IL-12. J Immunol 2005, 174, 2612–2618. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; et al. Therapeutic effects of an anti-IL-6 antibody in fungal keratitis: Macrophage inhibition and T cell subset regulation. Int Immunopharmacol 2020, 85, 106649. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, A.B.; et al. Lysophosphatidylcholine triggers TLR2- and TLR4-mediated signaling pathways but counteracts LPS-induced NO synthesis in peritoneal macrophages by inhibiting NF-κB translocation and MAPK/ERK phosphorylation. PLoS One 2013, 8, e76233. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Bernabé, A.; et al. Leishmania mexicana lipophosphoglycan activates ERK and p38 MAP kinase and induces production of proinflammatory cytokines in human macrophages through TLR2 and TLR4. Parasitology 2014, 141, 788–800. [Google Scholar] [CrossRef]
- von Knethen, A.; et al. 5-Lipoxygenase contributes to PPARγ activation in macrophages in response to apoptotic cells. Cell Signal 2013, 25, 2762–2768. [Google Scholar] [CrossRef]
Figure 1.
Effect of purified polysaccharides and the total capsule on the viability of DH82 cells. Canine macrophage DH82 cultures were grown at a concentration of 105 cells/well and incubated for 24 h (A) and 48 h (B). Cells were treated with GXMGal, GXM and total capsule at the indicated concentrations (μg/mL). The dead control received the addition of 0,1% of Triton before the addition of XTT. All cultures were performed in triplicate and bars show the mean + SD. Statistical analysis was performed by t-test from representative results of three similar experiment (*p<0.05, **p<0.01, ***p<0.001).
Figure 1.
Effect of purified polysaccharides and the total capsule on the viability of DH82 cells. Canine macrophage DH82 cultures were grown at a concentration of 105 cells/well and incubated for 24 h (A) and 48 h (B). Cells were treated with GXMGal, GXM and total capsule at the indicated concentrations (μg/mL). The dead control received the addition of 0,1% of Triton before the addition of XTT. All cultures were performed in triplicate and bars show the mean + SD. Statistical analysis was performed by t-test from representative results of three similar experiment (*p<0.05, **p<0.01, ***p<0.001).
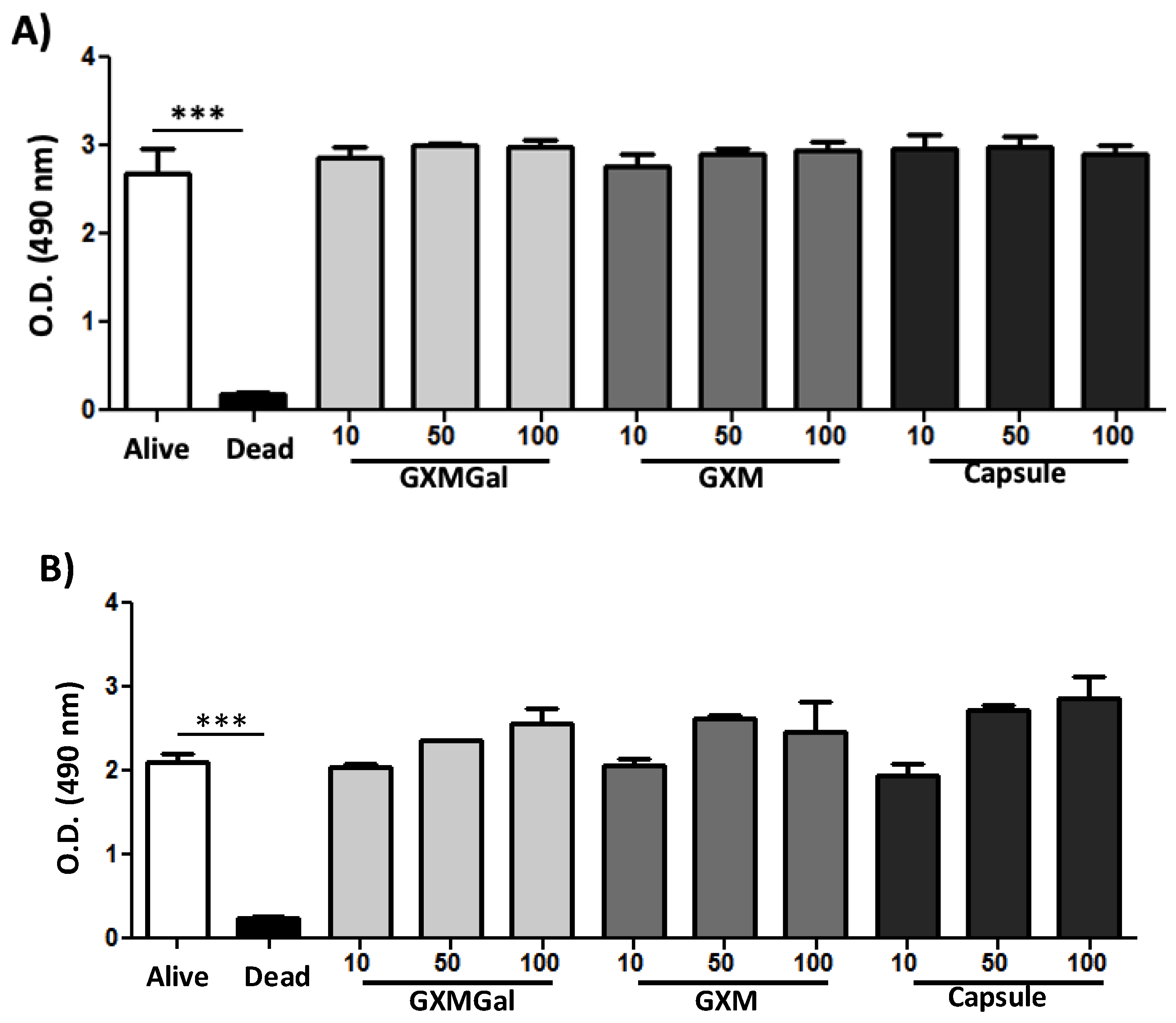
Figure 2.
Adherence and phagocytosis by DH82 in the presence of capsular polysaccharides of C. neoformans. DH82 cells were plated on a glass coverslip (1x104 cells/well) and incubated with GXMGal, GXM and capsule at the indicated concentrations (μg/mL). After 24 h, S. cerevisiae (10x104 cells/well) was added to the culture. After 40 minutes for the evaluation of binding (A), 4h to evaluate the percentage of cells that phagocytized (B) and number of phagocytosed yeasts (C). The cells were then stained, and the count was performed at 100x magnification. All cultures were performed in triplicate and bars show the mean + SD. Statistical analysis was performed by t-test from representative results of three similar experiment (*p<0.05, **p<0.01, ***p<0.001).
Figure 2.
Adherence and phagocytosis by DH82 in the presence of capsular polysaccharides of C. neoformans. DH82 cells were plated on a glass coverslip (1x104 cells/well) and incubated with GXMGal, GXM and capsule at the indicated concentrations (μg/mL). After 24 h, S. cerevisiae (10x104 cells/well) was added to the culture. After 40 minutes for the evaluation of binding (A), 4h to evaluate the percentage of cells that phagocytized (B) and number of phagocytosed yeasts (C). The cells were then stained, and the count was performed at 100x magnification. All cultures were performed in triplicate and bars show the mean + SD. Statistical analysis was performed by t-test from representative results of three similar experiment (*p<0.05, **p<0.01, ***p<0.001).
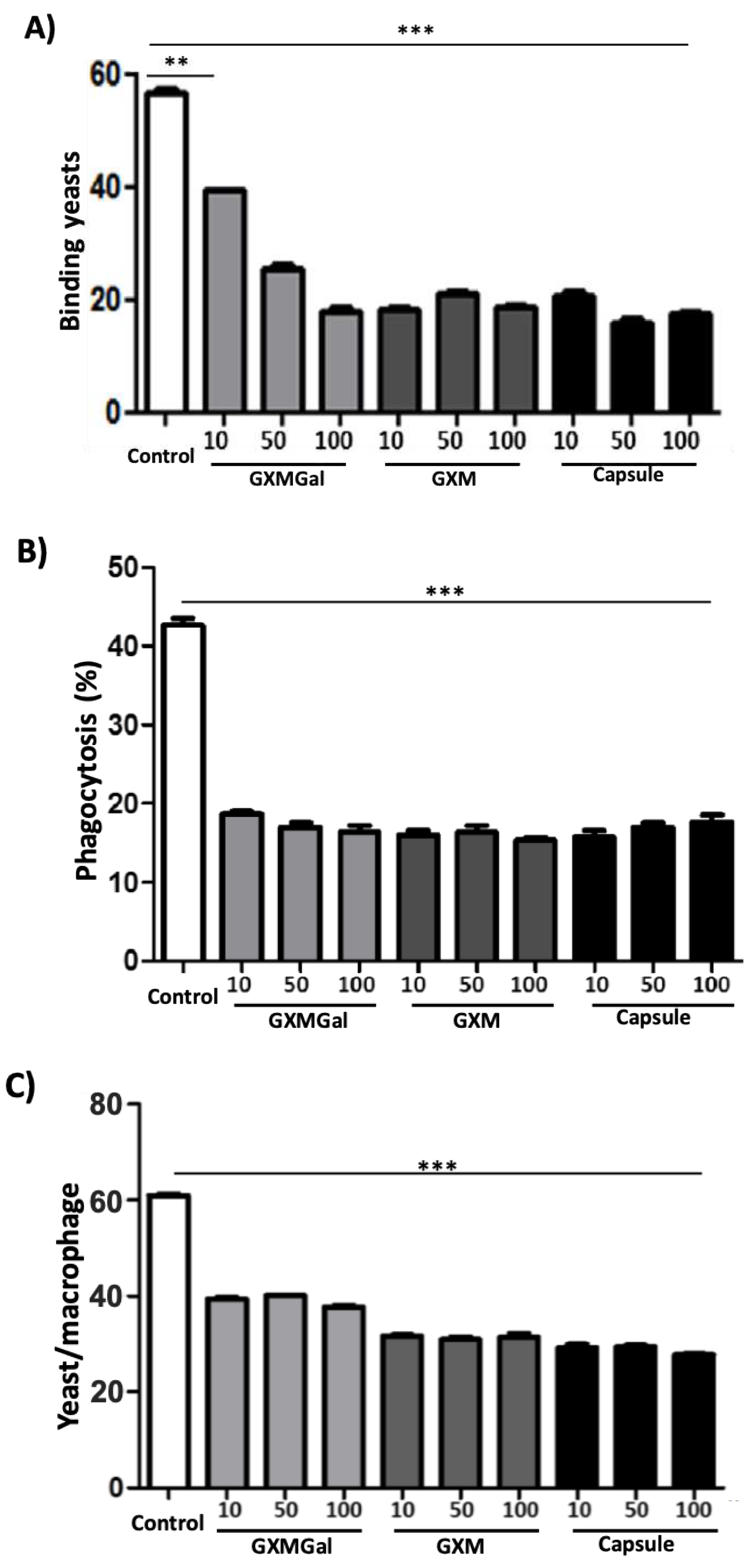
Figure 3.
Fungicidal activity of canine macrophages DH82 treated with purified polysaccharides from C. neoformans. DH82 cell cultures were plated (1x104 cells/well) and treated with GXMGal, GXM or capsule at the indicated concentrations (μg/mL). Yeasts of S. cerevisiae (10x104cells/well) were added to the culture and incubated for 4 h. After incubation, the macrophages were lysed, and the recovered yeasts were seeded on Sabouraud agar and incubated for 3 days. After incubation, colony forming units were counted. All cultures were performed in triplicate and bars show the mean + SD. Statistical analysis was performed by t-test from representative results of three similar experiment (*p<0.05, **p<0.01, ***p<0.001).
Figure 3.
Fungicidal activity of canine macrophages DH82 treated with purified polysaccharides from C. neoformans. DH82 cell cultures were plated (1x104 cells/well) and treated with GXMGal, GXM or capsule at the indicated concentrations (μg/mL). Yeasts of S. cerevisiae (10x104cells/well) were added to the culture and incubated for 4 h. After incubation, the macrophages were lysed, and the recovered yeasts were seeded on Sabouraud agar and incubated for 3 days. After incubation, colony forming units were counted. All cultures were performed in triplicate and bars show the mean + SD. Statistical analysis was performed by t-test from representative results of three similar experiment (*p<0.05, **p<0.01, ***p<0.001).
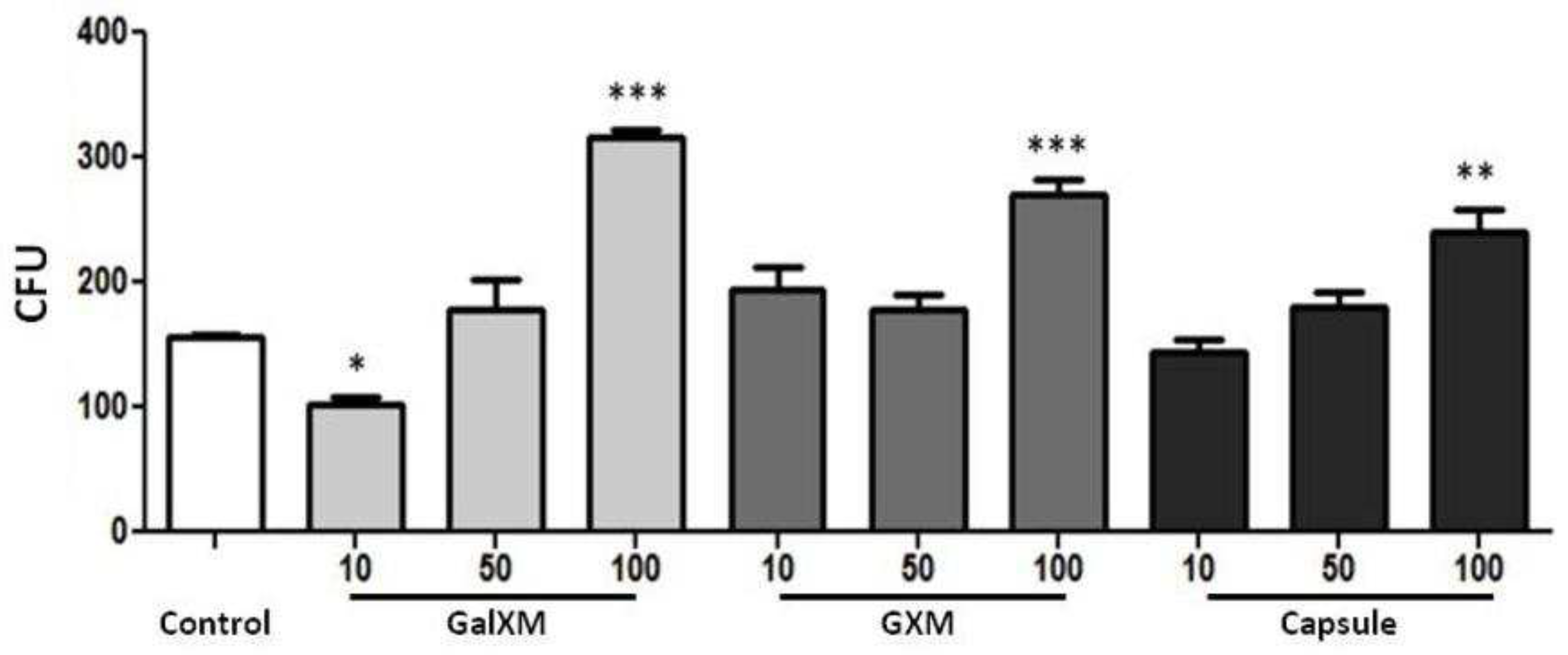
Figure 4.
Reactive species of oxygen (ROS) production by DH82 cells treated with purified capsular polysaccharides from C. neoformans. DH82 canine macrophages were cultured (5x104/mL) and incubated with H2DCFDA, followed by washing the cells were treated with GXMGal, GXM or capsule (100 μg/mL) and some cultures were stimulated with LPS (400 ng/mL) and IFN-γ (1.5 ng/mL) for 24 h. All cultures were performed in triplicate are shown as the mean arbitrary fluorescence units (AU) + SD. Statistical analysis was performed by t-test from representative results of three similar experiment (*p<0.05, **p<0.01, ***p<0.001).
Figure 4.
Reactive species of oxygen (ROS) production by DH82 cells treated with purified capsular polysaccharides from C. neoformans. DH82 canine macrophages were cultured (5x104/mL) and incubated with H2DCFDA, followed by washing the cells were treated with GXMGal, GXM or capsule (100 μg/mL) and some cultures were stimulated with LPS (400 ng/mL) and IFN-γ (1.5 ng/mL) for 24 h. All cultures were performed in triplicate are shown as the mean arbitrary fluorescence units (AU) + SD. Statistical analysis was performed by t-test from representative results of three similar experiment (*p<0.05, **p<0.01, ***p<0.001).
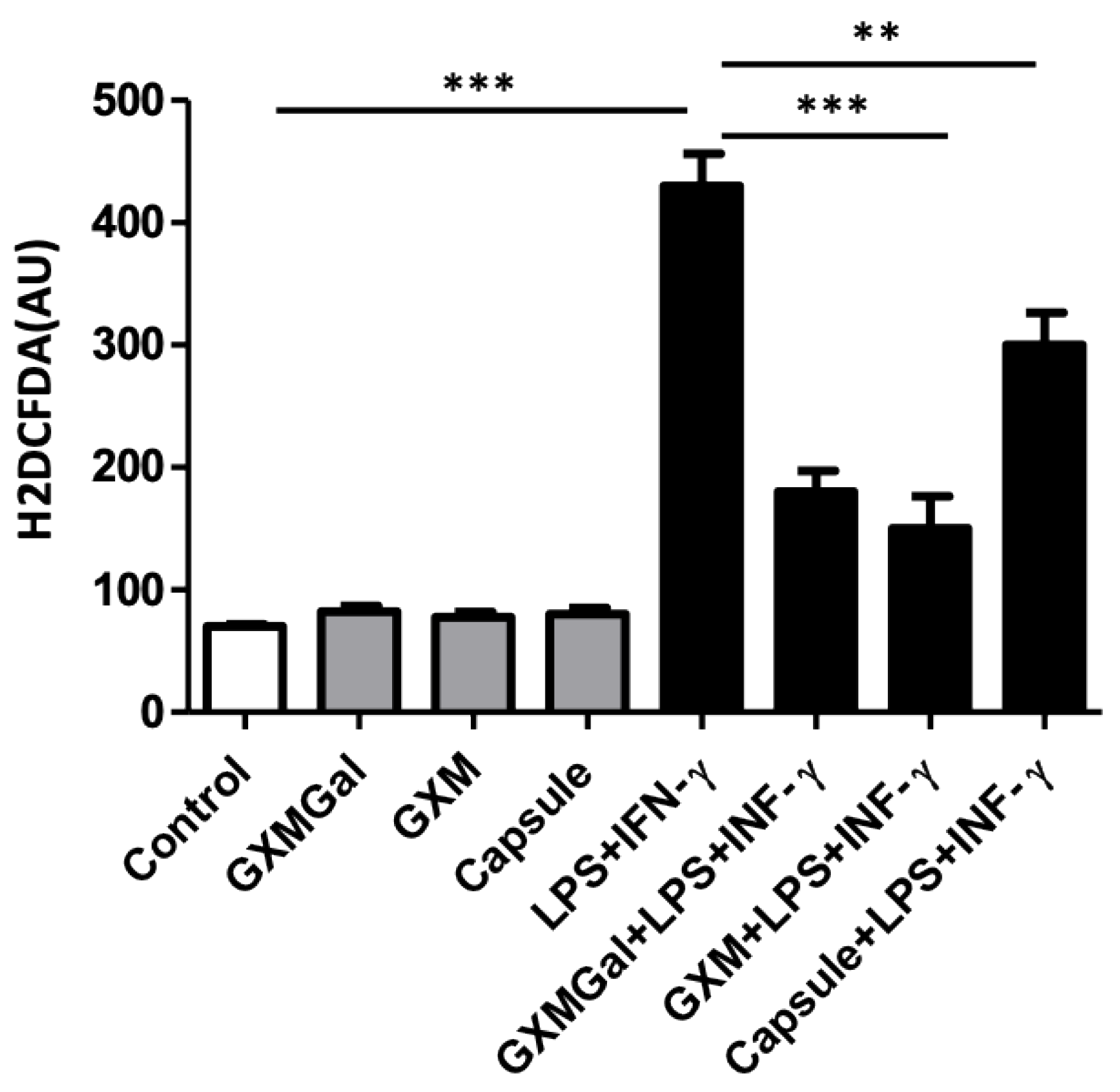
Figure 5.
Measurement of MHC class II expression in canine macrophage DH82 treated with powdered polysaccharides from C. neoformans. DH82 macrophages (5x105/mL) were treated with GXMGal, GXM (100 μg/mL). Some cultures were stimulated with LPS (400 ng/mL) and IFN-γ (1.5 ng/mL). After 24 h, the cells were prepared for analysis by flow cytometry. The M1 marker shows the percentage of cells identified by MHC class II (A, and B). All cultures were performed in triplicate are shown as the mean arbitrary fluorescence units (AU) + SD. Statistical analysis was performed by t-test from representative results of three similar experiment (*p<0.05, **p<0.01, ***p<0.001).
Figure 5.
Measurement of MHC class II expression in canine macrophage DH82 treated with powdered polysaccharides from C. neoformans. DH82 macrophages (5x105/mL) were treated with GXMGal, GXM (100 μg/mL). Some cultures were stimulated with LPS (400 ng/mL) and IFN-γ (1.5 ng/mL). After 24 h, the cells were prepared for analysis by flow cytometry. The M1 marker shows the percentage of cells identified by MHC class II (A, and B). All cultures were performed in triplicate are shown as the mean arbitrary fluorescence units (AU) + SD. Statistical analysis was performed by t-test from representative results of three similar experiment (*p<0.05, **p<0.01, ***p<0.001).
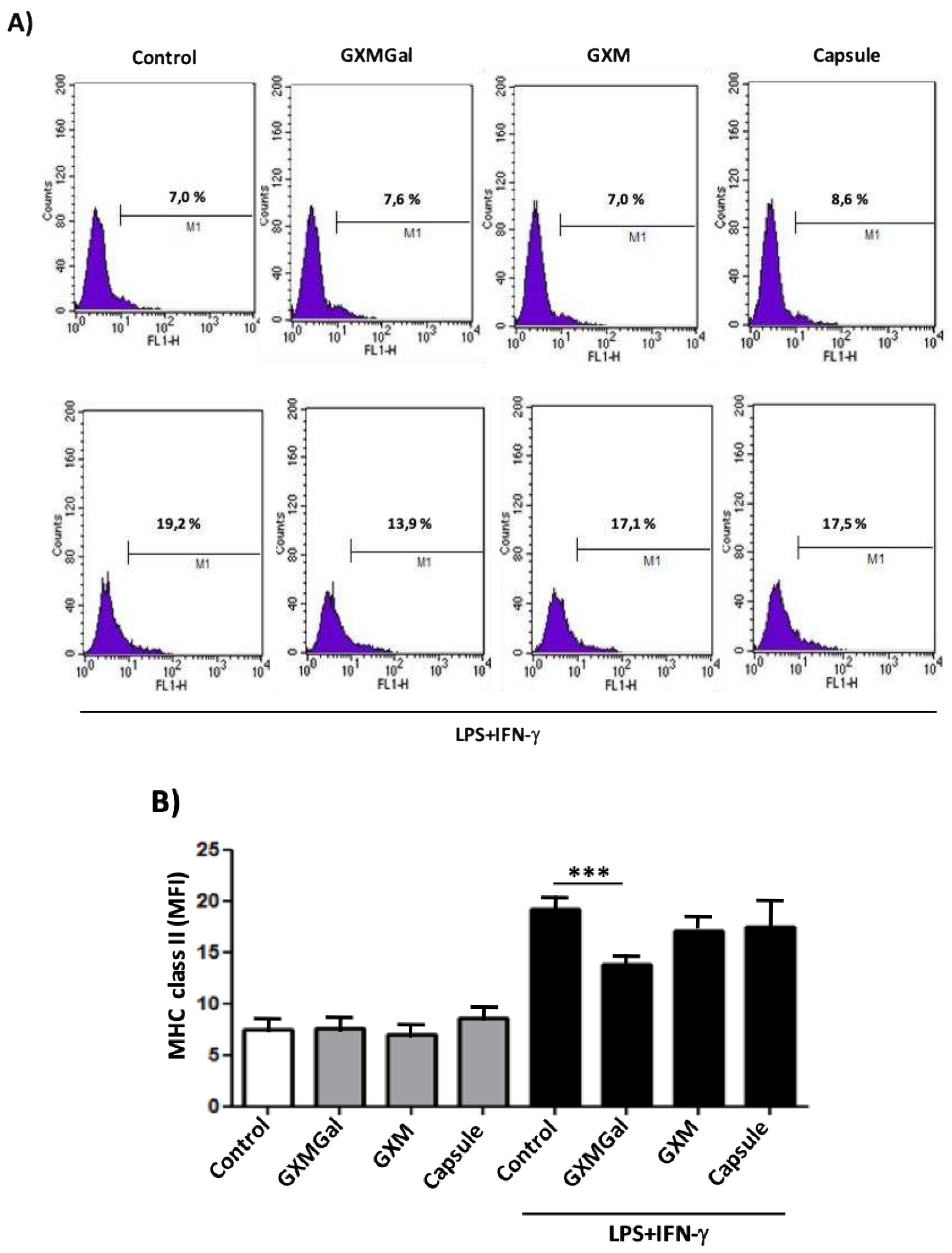
Figure 6.
Cytokine expression of the canine macrophage DH82. Cytokine profile in canine macrophage DH82 (1x106/mL) were plated and treated with GXMGal, GXM and capsule (100 μg/mL). Some cultures were stimulated with LPS (400 ng/mL) and IFN-γ (1.5 ng/mL). After 24 h of incubation, the cells were processed and mRNA for the cytokines quantified by qRT-PCR. Data from the experiments were normalized using GADPH primers as an endogenous control. All cultures were performed in triplicate and bars show the mean + SD. Statistical analysis was performed by t-test from representative results of three similar experiment (*p<0.05, **p<0.01, ***p<0.001).
Figure 6.
Cytokine expression of the canine macrophage DH82. Cytokine profile in canine macrophage DH82 (1x106/mL) were plated and treated with GXMGal, GXM and capsule (100 μg/mL). Some cultures were stimulated with LPS (400 ng/mL) and IFN-γ (1.5 ng/mL). After 24 h of incubation, the cells were processed and mRNA for the cytokines quantified by qRT-PCR. Data from the experiments were normalized using GADPH primers as an endogenous control. All cultures were performed in triplicate and bars show the mean + SD. Statistical analysis was performed by t-test from representative results of three similar experiment (*p<0.05, **p<0.01, ***p<0.001).
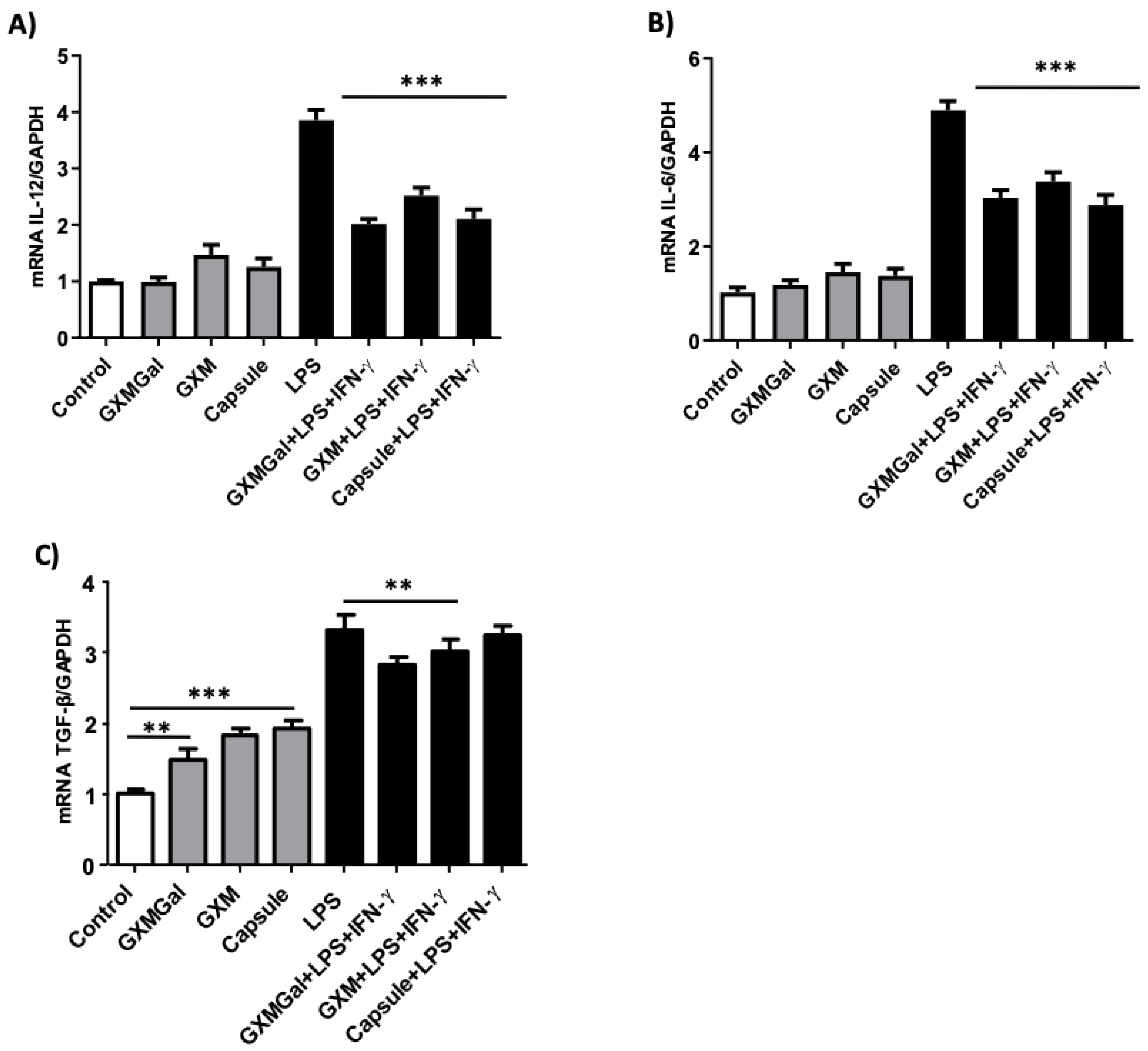
Figure 7.
MAPK phosphorylation and PPAR- γ activation. Canine macrophage DH82 (5x105) were plated and treated GXMGal, GXM and capsule (100 μg/mL). Some cultures were stimulated with LPS and IFN-γ. After 24 h of incubation whole-cell lysates were loaded onto SDS-PAGE gels. The blot was run and probed with the following antibodies: (A) anti-phospho ERK antibody and (B) anti-PPAR-γ antibody. The same blots were then stripped and reprobed with antibodies to non-phosphorylated proteins to determine absolute protein levels. Bar graphics shows the ratio of phosphorylated and total proteins. The band densitometry of western blotting was analyzed by the Scion Image software 4.0.3.2. All cultures were performed in triplicate and bars show the mean + SD. Statistical analysis was performed by t-test from representative results of three similar experiment (*p<0.05, **p<0.01, ***p<0.001).
Figure 7.
MAPK phosphorylation and PPAR- γ activation. Canine macrophage DH82 (5x105) were plated and treated GXMGal, GXM and capsule (100 μg/mL). Some cultures were stimulated with LPS and IFN-γ. After 24 h of incubation whole-cell lysates were loaded onto SDS-PAGE gels. The blot was run and probed with the following antibodies: (A) anti-phospho ERK antibody and (B) anti-PPAR-γ antibody. The same blots were then stripped and reprobed with antibodies to non-phosphorylated proteins to determine absolute protein levels. Bar graphics shows the ratio of phosphorylated and total proteins. The band densitometry of western blotting was analyzed by the Scion Image software 4.0.3.2. All cultures were performed in triplicate and bars show the mean + SD. Statistical analysis was performed by t-test from representative results of three similar experiment (*p<0.05, **p<0.01, ***p<0.001).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



