Submitted:
20 January 2024
Posted:
22 January 2024
You are already at the latest version
Abstract
Antimicrobial resistance is a major public health concern. Rodents and shrews live in closer proximity to humans and animals and serve as reservoirs for the transmission of several diseases, including AMR pathogens. There is no data available from India on surveillance of AMR bacteria in rodents and shrews. In this study, bacterial pathogens viz, Staphylococcus sp, Escherichia coli and Salmonella sp were isolated from rodents and shrews and genetic elements conferring antimicrobial resistance in the pathogens were screened by PCR. Bacterial pathogens in the oropharyngeal and anal swabs collected from the rodents and shrews trapped (n=100) were cultured on selective media such as Mannitol salt, Mac Conkey and Xylose lysine deoxycholate media to isolate Staphylococcus sp., E. coli and Salmonella sp. respectively. A total of 76 (80%) S. aureus and 19 (20%) Staphylococcus non aureus were isolated. E. coli was isolated in 89 samples and among the Salmonella sp (n=59), 16 (27%) were S. enteritidis and 29 (49.1%) were S. typhimurium. In S. aureus isolates, mec A and mec C (MRSA) were detected in 40 (52.6%) and 6 (7.9%) isolates respectively. Also, 7 (36.84%) and 1 (5.3%) Staphylococcus non aureus isolates were tested to harbour mec A and mec C genes. A total of 77 (86.5%) E. coli isolates had genes encoding ESBL [TEM in 21 (23.59%), SHV in 45 (50.6%) and CTXM in 11 (12.4%)]. Among the 45 pathogenic Salmonella sp, 44 (97.78%) harboured AMR genes [tet in 13 (28.9%), sul3 & sul4 in 20 (44.4%) and qnrA in 11 (24.4%)]. Thirty-eight animals harboured more than one zoonotic pathogen and 21 with multi-drug resistant genes. Our findings signify that these rodents and shrews may serve as sentinels to screen environment spillover of AMR pathogens and also act as reservoir to transmit MRSA, ESBL, Salmonella sp. human/animals. Also these animals aid in the transfer of antimicrobial resistance genetic elements to non-pathogenic organisms.
Keywords:
Rodents and Shrews
; MRSA
; ESBL
; Salmonella sp
; AMR
; Sentinels
; Drug resistance
Short title: Zoonotic bacterial pathogens and AMR in synanthropic rodents
Introduction
Rodents make up around 42% of all known mammalian species with high diversity and they are proven reservoirs for pathogens of public health and animal importance [1]. The faeces, saliva, urine and vectors infesting them act as potential sources of infections to human and animals by contaminating the water, food and feed supplies intended for human and animal use [2]. Rodents, by virtue of colonising the sewerage around the human habitats, feed on human and animal excreta and acquire the multidrug resistant pathogens [2,3]. Increased antibiotic resistance due to antibiotic overuse, may have a great impact on human health especially in an environment where animals and humans share the same ecosystem. As a result, the rodents have become a potent reservoir for hosting, maintaining and dispersing multidrug-resistant bacteria to humans and animals [3].
The mec genes found on the staphylococcal cassette chromosome mec (SCCmec), is a diverse and vast transmissible genetic element that contributes to antimicrobial resistance. There are 14 such SCCmec elements reported. Staphylococcus aureus Penicillin-binding protein (PBP 2a) with a poor affinity for β-lactams, encoded by mec A gene, was the major variant reported to confer methicillin resistance in S. aureus [4]. Later in 2011, mecC, a homologue to mecA, was reported in MRSA in human samples from Ireland. Later several reports of mecC arose globally from humans, animals and the environment [5]. The global spread of mecC was attributed to maritime travel and dispersal by commensal rodents [6].
World Health Organization (WHO report, 2014) has designated E. coli as a critical antimicrobial-resistant bacteria that can cause fatal illnesses such as pneumonia and septicaemia [7]. Extended-spectrumβ-lactamases (ESBL) producing E. coli strains are potentially known pathogens that are resistant to the majority of β-lactam antibiotics [8]. ESBL are plasmid encoded enzymes, which hydrolyses the β – lactam antibiotics including penicillin, cephalosporins and aztreonam. ESBL are further clustered into main three types, TEM, SHV and CTX-M [9]. E. coli with ESBL is acquired from the environment and has been reported in free-roaming rodents, shrews, bats, and wild birds [10].
The genus Salmonella has been associated with enteric diseases in humans, which have been classified into typhoidal illness and non-typhoidal salmonellosis (NTS). NTS, is one of the major aetiologies of enteritis globally, causing 93 million enteric infections with a mortality of 155,000 deaths due to diarrhoea every year [11,12]. Isolates of NTS with genes conferring antimicrobial resistance have been reported against antibiotics such as tetracyclines (tet), sulphonamides (sul3 & sul4), and fluoroquinolones (qnrA), nalidixic acid (nal) and third generation cephalosporins (CEP) [13].
To date, there are no reports on the spectrum of antimicrobial resistance in bacterial pathogens isolated from synanthropic rodents and shrews in India. Hence, the present work was undertaken to understand the dynamics of the spillover of MDR bacterial pathogens into the environment due to overuse of antibiotics in human and animal treatments.
Materials and Methods
Collection and processing of samples
This study was carried out in the Puducherry district, UT of Puducherry, India, which lies between 11’x55’x 48’’ Northern latitude and 79’x49’x 48’’ Eastern longitude with an altitude of 27 feet above the mean sea level. The district is a plain land with almost no mountains and forest. Trapping of rodents was carried out in randomly selected villages of Puducherry district for a period of six months from July to December 2022 (Figure 1). The institutional Animal Ethics Committee approved the study (IAEC/ICMR-VCRC/2018/2). A total of 100 animals were trapped alive in and around Puducherry. The trapped animals were euthanized by exposing them to a chamber saturated with carbon dioxide. The euthanized animals were identified according to their morphological characteristics using standard taxonomical keys [14]. A single oropharyngeal and two anal swabs were collected from each animal using sterile swabs (Himedia, India). The oropharyngeal swabs were transferred individually into the tubes with Luria broth enriched with 7.5% sodium chloride. Each anal swab was individually placed in tubes with Luria broth and selenite F broth respectively (Figure 2 and Figure 3). These samples were transported to the Department of Veterinary Microbiology, Rajiv Gandhi Institute of Veterinary Education and Research, Puducherry under the cold chain, for the isolation and detection of the bacterial pathogens and AMR genes by PCR.
Isolation of the bacterial pathogens:
All the 100 oropharyngeal samples collected in Luria broth enriched with 7.5% sodium chloride were incubated at 37⁰C for 16 to 18 hrs. Individual inoculum from the above was streaked onto Mannitol salt agar to obtain the mannitol fermenting golden yellow colonies. Similarly, 100 anal swabs collected in Luria broth were incubated and individual inoculum was streaked on the MacConkey agar to obtain the lactose fermenting colonies. The anal swabs collected in Selenite F broth were streaked on Brillant Green (BG) agar to isolate pink-coloured colonies. The pink-coloured colonies obtained in BG agar were propagated on XLD agar to obtain growth of the black-centred red-coloured colonies, specific for Salmonella sp.
2.3. Preparation of template DNA
The template DNA from the colonies isolated in Mannitol salt agar, MacConkey agar and XLD agar was extracted by boiling a loop full of colonies for ten minutes followed by snap chilling [15].
2.4. Identification of Staphylococcus spp. and genes conferring methicillin resistance by PCR
The bacterial isolates derived from Mannitol salt agar were primarily screened for Staphylococcus sp. by amplifying 16S rDNA partially [16] and then differentiated into S. aureus and Staphylococcus non aureus by amplifying the partial sequences of nuc gene [17] specific for S. aureus. Uniplex PCR was followed to screen genes conferring methicillin resistance viz mec A [18] and mec C [19] in all the Staphylococcus isolates.
2.5. Detection of Escherichia coli and the presence of Extended Spectrum Beta Lactamase genes (ESBL) by PCR
2.6. PCR Detection of Salmonella sp and non-typhoidal Salmonella sp. and their AMR genes
The bacterial DNA extracted from the black-centred red-coloured colonies on XLD agar were primarily screened for Salmonella sp targeting invA gene [24]. Upon confirmation of Salmonella sp., screening for S. enteritidis and S. typhimurium was done by specific amplification of spvC [25] and fliC [26] genes respectively. AMR genes such as tet [27], sul3 & sul4 [28] and qnrA [29] were screened by PCR in all the non-typhoidal Salmonella isolates.The details of the primers used for the identification of the bacterial pathogens, and their AMR genes, thermal cycling conditions, target amplicon sizes and the relevant references are summarised Table 1.
2.7. Sequencing and analysis of the PCR products
The amplified PCR products from the representative bacterial isolates and the AMR genes were gel extracted and subjected to Sanger sequencing (Genetic Analyzer 3130XL, Applied Biosystems, USA). The nucleotide sequenceswere BLAST analysed to confirm the gene identity(http://www.ncbi.nlm.nih.gov/BLAST). The nucleotide sequences were deposited in GenBank.
3. Results
In our study, a total of 100 animals were trapped, of which 75 were shrews (Suncus murinus) and 25 were rodents (Rattus rattus). From 95/100 (95%) oropharyngeal swabs, we observed the growth of golden yellow colonies in Mannitol salt agar indicating the presence of Staphylococcus sp. Further molecular confirmation was carried out by the successful amplification of 16S rRNA from all the 95 isolates. However, only 76/95 (80%) isolates were found to harbour the thermonuclease gene (nuc), specific for S. aureus. The rest 19 isolates (20%) were identified as Staphylococcus non aureus.
Molecular screening for methicillin resistance indicated that 40 (52.6%) (10 rodents and 30 shrews) and 6 (7.9%) (2 rodents and 4 shrews) among 76 Staphylococcus aureus isolates, were tested to harbour mec A and mec C genetic elements respectively signifying the presence of MRSA in these synanthropic rodents/shrews. Further screening of the 19 Staphylococcus non aureus isolates, indicated the occurrence of mec A and mec C genetic elements in 7 (36.8%) (2 rodents and 5 shrews) and 1 (1 shrew) (5.4%) isolates respectively. A total of five S. aureus (1 rodent and 4 shrews) isolates were found to harbour both mec A and mec C genetic elements conferring resistance to Methicillin.
The growth of characteristic dry, pink lactose fermenting colonies on the Mac Conkey’s agar was witnessed in 89/100 (89%) anal swabs, suggesting the possible presence of E. coli. Further, confirmation of E.coli in all the 89 isolate was done by PCR amplification of the universal stress protein A (uspA) gene. Among the 89 E. coli isolates,77 (86.5%) were identified as ESBL producers by PCR.
Altogether, 21 (23.6%) (6 rodents and 15 shrews), 45 (50.6%) (8 rodents and 37 shrews) and 11 (12.5%) (2 rodents and 9 shrews) of the isolates were found to carry ESBL genes viz. TEM, SHV and CTX-M, respectively. The presence of both TEM and SHV was witnessed in 13 isolates (16.8%) (2 rodents and 11 shrews), SHV and CTX-M in7 isolates (9.1%) (1 rodent and 6 shrews) whereas the occurrence of all 3 ESBL genes (TEM, SHV and CTX-M) was found in 2 (2.6%) isolates (1 shrew and 1 rodent).
The culture of anal swabs (n=100) in XLD agar, resulted in the growth of black-centred red-coloured colonies characteristic of Salmonella sp. from 59 (59%) swabs. PCR amplification of the invA gene in all the 59 isolates confirmed the presence of Salmonella spp. Further the Salmonella enterica serovar enteritidis and Salmonella enterica serovars typhimurium present among the 59 isolates were confirmed by the amplification of spvC and fliC genes in 16 (27.2%) and 29 (49.15%) isolates respectively. Among the 59 isolates, 44 (97.78%) were found positive for the presence of antimicrobial resistance genes. Altogether, 13 (28.8%) (3 rodents and 10 shrews), 20 (44.44%) (5 rodents and 15 shrews) and 11(24.44%) (2 rodents and 9 shrews) isolates were found to carry AMR genes viz tet, sul3& sul4 and qnrA conferring resistance to tetracyclines, sulphonamides and fluoroquinolones respectively. In addition, the occurrence of a combination of AMR genes was also observed in these isolates, tet and sul3 & sul4 in six isolates (13.63%) (2 rodents and 4 shrews), tet and qnrA in two isolates (4.5%) (2 shrews), and the presence of both sul3 & sul4 and qnrA was found in one (2.27%) (1 shrew) isolate (Table 2). The nucleotide sequences of the genes amplified for identification and confirmation of bacterial pathogens and the antimicrobial resistance genes were aligned using ClustalW software and deposited in the GenBank, the details of the accession ID assigned to the respective gene are given in Table 3.
Discussion
AMR is a multifaceted problem and its alarming rise in human and veterinary medicine coupled with its spread to the environment poses a grave challenge containing AMR. Hence, the spread of AMR in the environment is monitored by assessing the presence of AMR pathogens in sentinels such as wild animals [30] and rodents [31]. There is no data on AMR contamination in the environment through rodents and shrews from India. Hence, we assessed the spill over of AMR pathogens in the environment using the synanthropic rodents and shrews as sentinels.
MRSA, being the major nosocomial AMR pathogen, also emerged as a community-associated infection and is the major health care burden in Asia [31]. Hence, we screened the presence of MRSA from the oral swabs collected from the trapped rodents and shrews and found that they harboured S. aureus (80%, n=76) and Staphylococcus non aureus (20%, n=19). The occurrence of S. aureus in the trapped rodents and shrews was 30% and 70% respectively. The prevalence of 80% of S. aureus in the trapped mammals in our study is much higher than the reports of 7.1% [32] to 41.86% [33] in free-living wild animals and rodents trapped in pig farms in Spain and Netherlands respectively.
Among the S. aureus isolates, carriage of AMR genetic elements such as mec A and mec C were evident in 40 and 6 isolates respectively. The presence of both mec A and mec C genetic elements in the trapped rodents confirms the MRSA contamination in the environment from human and livestock sources [34].
Although, S. aureus with mecC genetic element in milk was first reported in India [35], we report the isolation of MRSA with mec C from rodents and shrews for the first time in India. The prevalence of MRSA in shrews and rodents in our study is 73.9% and 26.3% respectively. Ge et al., 2019 [36] reported a higher prevalence of MRSA in Rattus norvegicus (5.6%) than in shrews 0.5%, while we have observed a higher positivity of MRSA in shrews (74%) than in rodents (26%). The difference in the MRSA positivity across species could be attributed to factors such as season, geographical location and sample source [37].
The detection of AMR genetic elements mec A and mec C in Staphylococcus non aureus isolates was very interesting. Of the 19 Staphylococcus non aureus isolates, 7/19 (36.84%) and 1/19 (5.26%) isolates tested positive to harbour mec A and mec C genes. Phumthanakorn et al., 2022 [38] reported that among the 185 coagulase-negative Staphylococci isolated from dogs and cats, 25(13.4%) were tested to harbour the genetic elements conferring resistance to methicillin. Co-habitation of different microbial communities in a common niche leads to the horizontal transfer of AMR genetic elements to the cohabiting microbes [38]. Haaber et al.,2017 [39] reported that the coagulase-negative Staphylococci spp. acquired antimicrobial resistance genes by horizontal transfer from S. aureus.
The prevalence of 89% of E. coli found in the trapped rodents and shrews in our study is slightly higher than the 83.8% reported for rodents at the local zoo in Trinidad and Tobago [39] and 19.88% in rats in the east region of Gabon in Central Africa [40], Vancouver, Canada (41.5%), Berlin, Germany (58.2%), and Nairobi, Kenya (66.7%), [3,36,41,42]. In contrast, Nhung et al.,2015 [43] reported an incidence of 90.91% of E. coli isolates from synanthropic rats and shrews in Vietnam.
Among the 89 E. coli isolates, 77 (86.51%) were confirmed to harbour genetic elements such as TEM (23.59%), SHV (50.56%) and CTX-M (12.35%) respectively. Literak et al., 2009 [43] reported that 2.5% of E. coli isolated from black rats in Africa were ESBL-producers. However, Burriel et al., 2008 [44] reported that 61.5% of the E. coli isolated from R. norvegicus in a port from Greece were tested resistant to twelve antimicrobial agents. Gilliver et al., 1999 [45] reported that 90% of the coliforms isolated from wild rodents in northwest England were beta-lactam resistant.
In addition, some isolates were tested to harbour more than one genetic element encoding ESBL. A total of 13 (16.88%) isolates were tested positive for both TEM and SHV genes, 7 (9.09%) isolates for SHV and CTX-M genes, and 2 (2.59%) isolates were tested to carry TEM, SHV, and CTX-M genes. Out of 77, 22 (28.57%) were found to be positive for more than one AMR gene. A scoping review reported that the order of prevalence of ESBL genotype globally in rats were CTX-M (61%), TEM (26%) and SHV (13%) [46]. Ho et al., 2015 [46] reported a prevalence of 7.7% and 13.9% of CTX-M-producing multidrug-resistant E. coli in black and brown rats trapped in 18 districts of Hong Kong. Onanga et al., 2020 [40] reported human-origin E. coli carrying CTX-M genes in rats in Gabon, Central Africa.
By CTX-M gene characterization, the wild rodents in Hong Kong were identified to have acquired E. coli from humans and livestock [46]. Molecular clonal diversity characterization of the CTX-M gene, would have given an insight into the source of E. coli. However, this remains as the major limitation of our study. However, the presence of the genetic elements conferring resistance to beta-lactams in our study is in line with the spillage of antimicrobial resistance from human and animal pathogens to the environment. Guenther et al., 2012 [42] reported that rodents harbouring multi resistant E. coli are regarded as emerging zoonotic threats to humans. Thus highlighting the risk of transmission of these pathogens to human and animals in our study sites.
In EU/EEA, the salmonellosis was reported to rank second among the zoonoses with a report of 91,857 cases, during 2018 [47]. Interestingly, we observed that 59% of the rodents/shrews in our study to harbour Salmonella spp. However, there are reports of nil [48], and high [41] prevalence of Salmonella spp. in rodents. Further molecular typing indicated the presence of NTS pathogens Salmonella enterica serovar enteritidis (27.7%) and Salmonella enterica serovar typhimurium (49.15%). The major mode of transmission of NTS is through contaminated food [13]. Salmonella enterica serovar enteritidis (28.57%) and Salmonella enterica serovar typhimurium (63.49%) were detected in rats inhabiting poultry farms in South Africa[49]. Salmonella enterica serovars enteritidis and Salmonella enterica serovar typhimurium are the common pathogenic Salmonella spp. reported in 5 developing countries like South Africa, Egypt, Indonesia, India and Romania and the major source of NTS infection in human is the contamination of food with human/ animal wastes[50]. NTS are of public health importance as it is associated with non-invasive enteritis and fatal bloodstream infections in children and adults [51].
We first documented the antimicrobial resistance in Salmonella sp. isolated from rodents and shrews in Puducherry, India. In that, 44/45 (97.78%) were found to carry antimicrobial resistance genes. Altogether, 13/45 (28.88%), 20/45 (44.44%), and 11/45 (24.44%) of the isolates were found to carry AMR genes viz tet, sul3 & sul4 and qnrA respectively. Himsworth et al., 2018 [41] studied the prevalence of AMR in Salmonella sp. isolated from wild urban Norway and black rats from Canada. Out of 633 rodents screened, 13 were positive for Salmonella sp, out of which 2 isolates (15.38%) were tested resistant to sulphonamides and 1 (7.69%) to tetracycline.
Sulfonamides are widely used as antibiotics for the treatment of infectious diseases in human, animals. It is also commonly used for water treatment in poultry houses, thus making sulfonamides as a major environment pollutant in the poultry house effluents [52]. Balakrishnan et al., 2016 [53] isolated fluoroquinolones resistant E. coli isolates of animal origin from Puducherry. In contrast, Ramatla et al., 2022[54] found a higher percentage of qnrA and tet gene resistance in rodents in South Africa. These quinolones are being used to treat human Salmonellosis for over 4 decades. Fluoroquinolones have been considered as one of the last options for the treatment of Salmonella sp [55]. The misuse and overuse of quinolone antibiotics in both veterinary and human medicine have led to the emergence of resistance.
The presence of AMR genes in the pathogens isolated from rodents and shrews clearly indicates the irrational use of antibiotics in veterinary and human treatments, which eventually lead to the spillover of MDR pathogens in the environment. As rodents and shrews mostly populate in the sewage in and around human habitations, they may have picked up the multi-resistant strains from the human and animal excreta [42]. Also, they may act as surrogate indicators of the environmentalspillover of AMR pathogens. To curtail the development of superbugs, necessary intervention measures have to be undertaken for judicious use and disposal of antibiotics.
Conclusions
In conclusion, we report that 89% of the rodents and shrews were tested to harbour more than one zoonotic bacterial pathogens, whereas 55% of them were found to carry multiple AMR genes posing a potential health risk to human, livestockand pet animals. This study underscores the screening of rodents/shrews as sentinels to test the spillover of antimicrobial resistance in the environment.
Author Contributions
HKM, PD & MR contributed to conception and design of the study. PD acquired funds. AC and TNA carried out trapping, morphological identification of rodents and extraction of nucleic acid. ND, KKS, LS, VMS, JV carried out the isolation of the pathogens, PCR detection of pathogens and AMR genetic elements, nucleotide sequencing and analysis. DP, HKM, MR and VP supervised the experiments. PD, ND, HKM, KKS and TNA organized the database and wrote the first draft of the manuscript. All the authors contributed to manuscript revision and approved the final version.
Funding
This extramural study was funded by Department of Health Research, Govt. of India, New Delhi (Grant No: R.11013/57/2021-GIA/HR) for funding the research work.
Acknowledgment
We sincerely acknowledge Department of Health Research, Govt. of India, New Delhi, for funding this study. We thank the Dean, Rajiv Gandhi Institute of Veterinary Education and Research, Puducherry, for supporting this study by providing all the facilities. We thank Mr S. Rajkumar and Mrs. S. Pushpa for their technical support.
Conflicts of Interest
Nil
References
- The Encyclopedia of Mammals, 3rd ed.; Oxford University Press, 2006; ISBN 978-0-19-920608-7.
- Meerburg, B.G.; Singleton, G.R.; Kijlstra, A. Rodent-Borne Diseases and Their Risks for Public Health. Crit. Rev. Microbiol. 2009, 35, 221–270. [Google Scholar] [CrossRef]
- Gakuya, F.M.; Kyule, M.N.; Gathura, P.B.; Kariuki, S. Antimicrobial Resistance of Bacterial Organisms Isolated from Rats. East Afr. Med. J. 2001, 78, 646–649. [Google Scholar] [CrossRef]
- Peacock, S.J.; Paterson, G.K. Mechanisms of Methicillin Resistance in Staphylococcus Aureus. Annu. Rev. Biochem. 2015, 84, 577–601. [Google Scholar] [CrossRef]
- Silva, V.; Gabriel, S.I.; Borrego, S.B.; Tejedor-Junco, M.T.; Manageiro, V.; Ferreira, E.; Reis, L.; Caniça, M.; Capelo, J.L.; Igrejas, G.; et al. Antimicrobial Resistance and Genetic Lineages of Staphylococcus Aureus from Wild Rodents: First Report of mecC-Positive Methicillin-Resistant S. Aureus (MRSA) in Portugal. Animals 2021, 11, 1537. [Google Scholar] [CrossRef]
- Hansen, T.A.; Joshi, T.; Larsen, A.R.; Andersen, P.S.; Harms, K.; Mollerup, S.; Willerslev, E.; Fuursted, K.; Nielsen, L.P.; Hansen, A.J. Vancomycin Gene Selection in the Microbiome of Urban Rattus Norvegicus from Hospital Environment. Evol. Med. Public Health 2016, 2016, 219–226. [Google Scholar] [CrossRef]
- WHO. Antimicrobial Resistance: Global Report on Surveillance; 2014. [Google Scholar]
- Masoud, S.M.; Abd El-Baky, R.M.; Aly, S.A.; Ibrahem, R.A. Co-Existence of Certain ESBLs, MBLs and Plasmid Mediated Quinolone Resistance Genes among MDR E. Coli Isolated from Different Clinical Specimens in Egypt. Antibiotics 2021, 10, 835. [Google Scholar] [CrossRef]
- Sonola, V.S.; Katakweba, A.; Misinzo, G.; Matee, M.I. Molecular Epidemiology of Antibiotic Resistance Genes and Virulence Factors in Multidrug-Resistant Escherichia Coli Isolated from Rodents, Humans, Chicken, and Household Soils in Karatu, Northern Tanzania. Int. J. Environ. Res. Public Health 2022, 19, 5388. [Google Scholar] [CrossRef]
- Ong, K.H.; Khor, W.C.; Quek, J.Y.; Low, Z.X.; Arivalan, S.; Humaidi, M.; Chua, C.; Seow, K.L.G.; Guo, S.; Tay, M.Y.F.; et al. Occurrence and Antimicrobial Resistance Traits of Escherichia Coli from Wild Birds and Rodents in Singapore. Int. J. Environ. Res. Public Health 2020, 17, 5606. [Google Scholar] [CrossRef]
- Ao, T.T.; Feasey, N.A.; Gordon, M.A.; Keddy, K.H.; Angulo, F.J.; Crump, J.A. Global Burden of Invasive Nontyphoidal Salmonella Disease, 2010(1). Emerg Infect. Dis. 2015, 21, 941–949. [Google Scholar] [CrossRef]
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M. International Collaboration on Enteric Disease “Burden of Illness” Studies The Global Burden of Nontyphoidal Salmonella Gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef]
- McDermott, P.F.; Zhao, S.; Tate, H. Antimicrobial Resistance in Nontyphoidal Salmonella. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, V.C. Taxonomic Studies on Indian Muridae and Hystricidae: Mammalia, Rodentia; Records of the Zoological Survey of India; Zoological Survey of India: Calcutta, India, 2000; ISBN 978-81-85874-31-9. [Google Scholar]
- Soumya, M.P.; Pillai, R.M.; Antony, P.X.; Mukhopadhyay, H.K.; Rao, V.N. Comparison of Faecal Culture and IS900 PCR Assay for the Detection of Mycobacterium Avium Subsp. Paratuberculosis in Bovine Faecal Samples. Vet. Res. Commun. 2009, 33, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Sparling, J.; Chow, B.L.; Elsayed, S.; Hussain, Z.; Church, D.L.; Gregson, D.B.; Louie, T.; Conly, J.M. New Quadriplex PCR Assay for Detection of Methicillin and Mupirocin Resistance and Simultaneous Discrimination of Staphylococcus Aureus from Coagulase-Negative Staphylococci. J. Clin. Microbiol. 2004, 42, 4947–4955. [Google Scholar] [CrossRef] [PubMed]
- Brakstad, O.G.; Aasbakk, K.; Maeland, J.A. Detection of Staphylococcus Aureus by Polymerase Chain Reaction Amplification of the Nuc Gene. J. Clin. Microbiol. 1992, 30, 1654–1660. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.C.; de Lencastre, H. Multiplex PCR Strategy for Rapid Identification of Structural Types and Variants of the Mec Element in Methicillin-Resistant Staphylococcus Aureus. Antimicrob. Agents Chemother. 2002, 46, 2155–2161. [Google Scholar] [CrossRef] [PubMed]
- Stegger, M.; Andersen, P.S.; Kearns, A.; Pichon, B.; Holmes, M.A.; Edwards, G.; Laurent, F.; Teale, C.; Skov, R.; Larsen, A.R. Rapid Detection, Differentiation and Typing of Methicillin-Resistant Staphylococcus Aureus Harbouring Either mecA or the New mecA Homologue mecA(LGA251). Clin. Microbiol. Infect 2012, 18, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Griffiths, M.W. PCR Differentiation of Escherichia Coli from Other Gram-Negative Bacteria Using Primers Derived from the Nucleotide Sequences Flanking the Gene Encoding the Universal Stress Protein. Lett. Appl. Microbiol. 1998, 27, 369–371. [Google Scholar] [CrossRef]
- Colom, K.; Pérez, J.; Alonso, R.; Fernández-Aranguiz, A.; Lariño, E.; Cisterna, R. Simple and Reliable Multiplex PCR Assay for Detection of blaTEM, Bla(SHV) and blaOXA-1 Genes in Enterobacteriaceae. FEMS Microbiol. Lett. 2003, 223, 147–151. [Google Scholar] [CrossRef]
- Rasheed, J.K.; Jay, C.; Metchock, B.; Berkowitz, F.; Weigel, L.; Crellin, J.; Steward, C.; Hill, B.; Medeiros, A.A.; Tenover, F.C. Evolution of Extended-Spectrum Beta-Lactam Resistance (SHV-8) in a Strain of Escherichia Coli during Multiple Episodes of Bacteremia. Antimicrob. Agents Chemother. 1997, 41, 647–653. [Google Scholar] [CrossRef]
- Dutta, T.K.; Warjri, I.; Roychoudhury, P.; Lalzampuia, H.; Samanta, I.; Joardar, S.N.; Bandyopadhyay, S.; Chandra, R. Extended-Spectrum-β-Lactamase-Producing Escherichia Coli Isolate Possessing the Shiga Toxin Gene (Stx1) Belonging to the O64 Serogroup Associated with Human Disease in India. J. Clin. Microbiol. 2013, 51, 2008–2009. [Google Scholar] [CrossRef]
- Aabo, S.; Rasmussen, O.F.; Rossen, L.; Sørensen, P.D.; Olsen, J.E. Salmonella Identification by the Polymerase Chain Reaction. Mol. Cell Probes 1993, 7, 171–178. [Google Scholar] [CrossRef]
- Wood, M.W.; Mahon, J.; Lax, A.J. Development of a Probe and PCR Primers Specific to the Virulence Plasmid of Salmonella Enteritidis. Mol. Cell Probes 1994, 8, 473–479. [Google Scholar] [CrossRef]
- Soumet, C.; Ermel, G.; Rose, N.; Rose, V.; Drouin, P.; Salvat, G.; Colin, P. Evaluation of a Multiplex PCR Assay for Simultaneous Identification of Salmonella Sp., Salmonella Enteritidis and Salmonella Typhimurium from Environmental Swabs of Poultry Houses. Lett. Appl. Microbiol. 1999, 28, 113–117. [Google Scholar] [CrossRef]
- Olobatoke, R.Y.; Mulugeta, S.D. Incidence of Non-Typhoidal Salmonella in Poultry Products in the North West Province, South Africa. S. Afr. J. Sci. 2015, 111, 7. [Google Scholar] [CrossRef]
- Chu, C.; Chiu, C.H.; Wu, W.Y.; Chu, C.H.; Liu, T.P.; Ou, J.T. Large Drug Resistance Virulence Plasmids of Clinical Isolates of Salmonella Enterica Serovar Choleraesuis. Antimicrob. Agents Chemother. 2001, 45, 2299–2303. [Google Scholar] [CrossRef]
- Kehrenberg, C.; Friederichs, S.; de Jong, A.; Michael, G.B.; Schwarz, S. Identification of the Plasmid-Borne Quinolone Resistance Gene qnrS in Salmonella Enterica Serovar Infantis. J. Antimicrob. Chemother. 2006, 58, 18–22. [Google Scholar] [CrossRef]
- Vittecoq, M.; Godreuil, S.; Prugnolle, F.; Durand, P.; Brazier, L.; Renaud, N.; Arnal, A.; Aberkane, S.; Jean-Pierre, H.; Gauthier-Clerc, M.; et al. Antimicrobial Resistance in Wildlife. J. Appl. Ecol. 2016, 53, 519–529. [Google Scholar] [CrossRef]
- Chen, C.-J.; Huang, Y.-C. New Epidemiology of Staphylococcus Aureus Infection in Asia. Clin. Microbiol. Infect 2014, 20, 605–623. [Google Scholar] [CrossRef]
- Porrero, M.C.; Mentaberre, G.; Sánchez, S.; Fernández-Llario, P.; Gómez-Barrero, S.; Navarro-Gonzalez, N.; Serrano, E.; Casas-Díaz, E.; Marco, I.; Fernández-Garayzabal, J.-F.; et al. Methicillin Resistant Staphylococcus Aureus (MRSA) Carriage in Different Free-Living Wild Animal Species in Spain. Vet. J. 2013, 198, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Van De Giessen, A.W.; Van Santen-Verheuvel, M.G.; Hengeveld, P.D.; Bosch, T.; Broens, E.M.; Reusken, C.B.E.M. Occurrence of Methicillin-Resistant Staphylococcus Aureus in Rats Living on Pig Farms. Prev. Vet. Med. 2009, 91, 270–273. [Google Scholar] [CrossRef]
- García-Álvarez, L.; Holden, M.T.G.; Lindsay, H.; Webb, C.R.; Brown, D.F.J.; Curran, M.D.; Walpole, E.; Brooks, K.; Pickard, D.J.; Teale, C.; et al. Meticillin-Resistant Staphylococcus Aureus with a Novel mecA Homologue in Human and Bovine Populations in the UK and Denmark: A Descriptive Study. Lancet Infect Dis. 2011, 11, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Dey, T.K.; Shome, B.R.; Bandyopadhyay, S.; Goyal, N.K.; Lundkvist, Å.; Deka, R.P.; Shome, R.; Venugopal, N.; Grace, D.; Sharma, G.; et al. Molecular Characterization of Methicillin-Resistant Staphylococci from the Dairy Value Chain in Two Indian States. Pathogens 2023, 12, 344. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.; Zhong, X.; Xiong, Y.; Qiu, M.; Huo, S.; Chen, X.; Mo, Y.; Cheng, M.; Chen, Q. Methicillin-Resistant Staphylococcus Aureus among Urban Rodents, House Shrews, and Patients in Guangzhou, Southern China. BMC Vet. Res. 2019, 15, 260. [Google Scholar] [CrossRef] [PubMed]
- Erickson, M.C. Overview: Foodborne Pathogens in Wildlife Populations. In Food Safety Risks from Wildlife; Jay-Russell, M., Doyle, M.P., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 1–30. ISBN 978-3-319-24440-2. [Google Scholar]
- Baquero, F.; Coque, T.M.; Martínez, J.-L.; Aracil-Gisbert, S.; Lanza, V.F. Gene Transmission in the One Health Microbiosphere and the Channels of Antimicrobial Resistance. Front. Microbiol. 2019, 10, 2892. [Google Scholar] [CrossRef] [PubMed]
- Nkogwe, C.; Raletobana, J.; Stewart-Johnson, A.; Suepaul, S.; Adesiyun, A. Frequency of Detection of Escherichia Coli, Salmonella Spp., and Campylobacter Spp. in the Faeces of Wild Rats (Rattus Spp.) in Trinidad and Tobago. Vet. Med. Int. 2011, 2011, 686923. [Google Scholar] [CrossRef] [PubMed]
- Onanga, R.; Mbehang Nguema, P.P.; Ndong Atome, G.R.; Mabika Mabika, A.; Ngoubangoye, B.; Komba Tonda, W.L.; Obague Mbeang, J.C.; Lebibi, J. Prevalence of Extended-Spectrum β-Lactamases in E. Coli of Rats in the Region North East of Gabon. Vet. Med. Int. 2020, 2020, 5163493. [Google Scholar] [CrossRef] [PubMed]
- Himsworth, C.G.; Zabek, E.; Desruisseau, A.; Parmley, E.J.; Reid-Smith, R.; Jardine, C.M.; Tang, P.; Patrick, D.M. Prevalence and Characterization of E.Coli and Salmonella Spp in the Feces of Wild Urban Norway and Black Rats (Rattus Norvegius and Rattus Ratttus ) from an Inner City Neighbourhood of Vancouver. J. Wildl. Dis. 2015, 51, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Guenther, S.; Bethe, A.; Fruth, A.; Semmler, T.; Ulrich, R.G.; Wieler, L.H.; Ewers, C. Frequent Combination of Antimicrobial Multiresistance and Extraintestinal Pathogenicity in Escherichia Coli Isolates from Urban Rats (Rattus Norvegicus) in Berlin, Germany. PLoS ONE 2012, 7, e50331. [Google Scholar] [CrossRef]
- Literák, I.; Dolejska, M.; Cizek, A.; Djigo, C.A.T. Reservoirs of Antibiotic-Resistant Enterobacteriaceae among Animals Sympatric to Humans in Senegal: Extended-Spectrum Beta-Lactamases in Bacteria in a Black Rat (Rattus Rattus). Afr. J. Microbiol. Res. 2014, 3, 751–754. [Google Scholar]
- Burriel, A.R.; Kritas, S.K.; Kontos, V. Some Microbiological Aspects of Rats Captured Alive at the Port City of Piraeus, Greece. Int. J. Environ. Health Res. 2008, 18, 159–164. [Google Scholar] [CrossRef]
- Gilliver, M.A.; Bennett, M.; Begon, M.; Hazel, S.M.; Hart, C.A. Antibiotic Resistance Found in Wild Rodents. Nature 1999, 401, 233–234. [Google Scholar] [CrossRef] [PubMed]
- Ho, P.-L.; Lo, W.-U.; Lai, E.L.; Law, P.Y.; Leung, S.M.; Wang, Y.; Chow, K.-H. Clonal Diversity of CTX-M-Producing, Multidrug-Resistant Escherichia Coli from Rodents. J. Med. Microbiol. 2015, 64, 185–190. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority and European Centre for Disease Prevention and Control (EFSA and ECDC) The European Union One Health 2018 Zoonoses Report. EFSA J. 2019, 17, e05926. [CrossRef]
- El-Sharkawy, H.; Tahoun, A.; El-Gohary, A.E.-G.A.; El-Abasy, M.; El-Khayat, F.; Gillespie, T.; Kitade, Y.; Hafez, H.M.; Neubauer, H.; El-Adawy, H. Epidemiological, Molecular Characterization and Antibiotic Resistance of Salmonella Enterica Serovars Isolated from Chicken Farms in Egypt. Gut. Pathog. 2017, 9, 8. [Google Scholar] [CrossRef] [PubMed]
- Taioe, M.O.; Thekisoe, O.M.M.; Syakalima, M.; Ramatla, T. Confirmation of Antimicrobial Resistance by Using Resistance Genes of Isolated Salmonella Spp. in Chicken Houses of North West, South Africa. Jwpr 2019, 9, 158–165. [Google Scholar] [CrossRef]
- Barbour, E.K.; Ayyash, D.B.; Alturkistni, W.; Alyahiby, A.; Yaghmoor, S.; Iyer, A.; Yousef, J.; Kumosani, T.; Harakeh, S. Impact of Sporadic Reporting of Poultry Salmonella Serovars from Selected Developing Countries. J. Infect Dev. Ctries 2015, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Feasey, N.A.; Dougan, G.; Kingsley, R.A.; Heyderman, R.S.; Gordon, M.A. Invasive Non-Typhoidal Salmonella Disease: An Emerging and Neglected Tropical Disease in Africa. Lancet 2012, 379, 2489–2499. [Google Scholar] [CrossRef]
- Bertelloni, F.; Tosi, G.; Massi, P.; Fiorentini, L.; Parigi, M.; Cerri, D.; Ebani, V.V. Some Pathogenic Characters of Paratyphoid Salmonella Enterica Strains Isolated from Poultry. Asian Pac J. Trop. Med. 2017, 10, 1161–1166. [Google Scholar] [CrossRef]
- Balakrishnan, S.; Antony, P.X.; Mukhopadhyay, H.K.; Pillai, R.M.; Thanislass, J.; Padmanaban, V.; Srinivas, M.V. Genetic Characterization of Fluoroquinolone-Resistant Escherichia Coli Associated with Bovine Mastitis in India. Vet. World 2016, 9, 705–709. [Google Scholar] [CrossRef]
- Ramatla, T.; Tawana, M.; Lekota, K.E.; Thekisoe, O. Antimicrobial Resistance Genes of Escherichia Coli, a Bacterium of “One Health” Importance in South Africa: Systematic Review and Meta-Analysis. AIMS Microbiol. 2023, 9, 75–89. [Google Scholar] [CrossRef]
- Abdel-Maksoud, M.; Abdel-Khalek, R.; El-Gendy, A.; Gamal, R.F.; Abdelhady, H.M.; House, B.L. Genetic Characterisation of Multidrug-Resistant Salmonella Enterica Serotypes Isolated from Poultry in Cairo, Egypt. Afr. J. Lab. Med. 2015, 4, 7. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The map representing the sites of rodents trapping in Puducherry. The red dot indicates the villages, where the rodents were trapped.
Figure 1.
The map representing the sites of rodents trapping in Puducherry. The red dot indicates the villages, where the rodents were trapped.
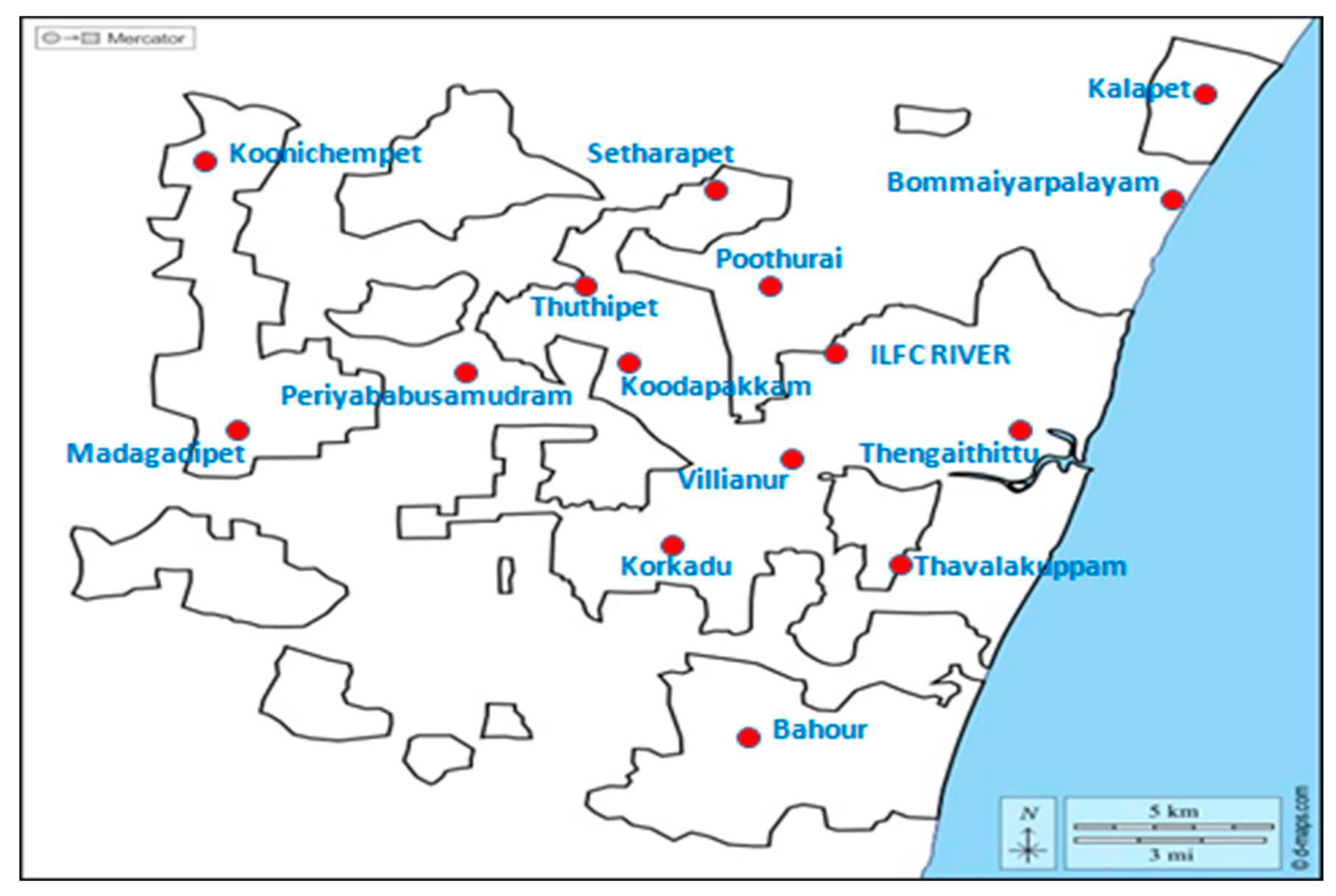
Figure 2.
Collection of the oro-pharyngeal sample for the isolation of Staphylococcus spp. from a shrew.
Figure 2.
Collection of the oro-pharyngeal sample for the isolation of Staphylococcus spp. from a shrew.
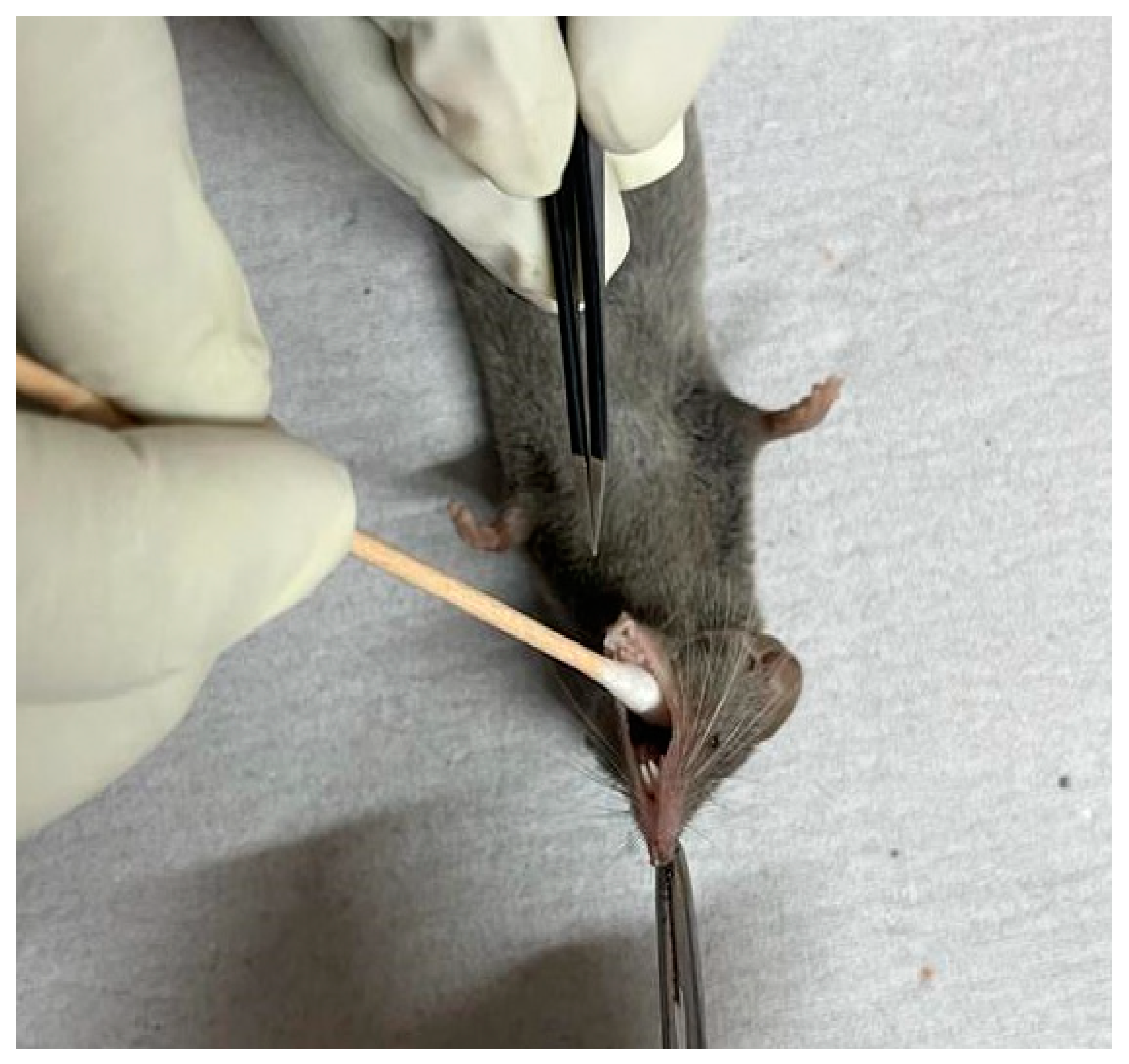
Figure 3.
Collection of the anal swab from a shrew for the isolation of E. coli and Salmonella spp.
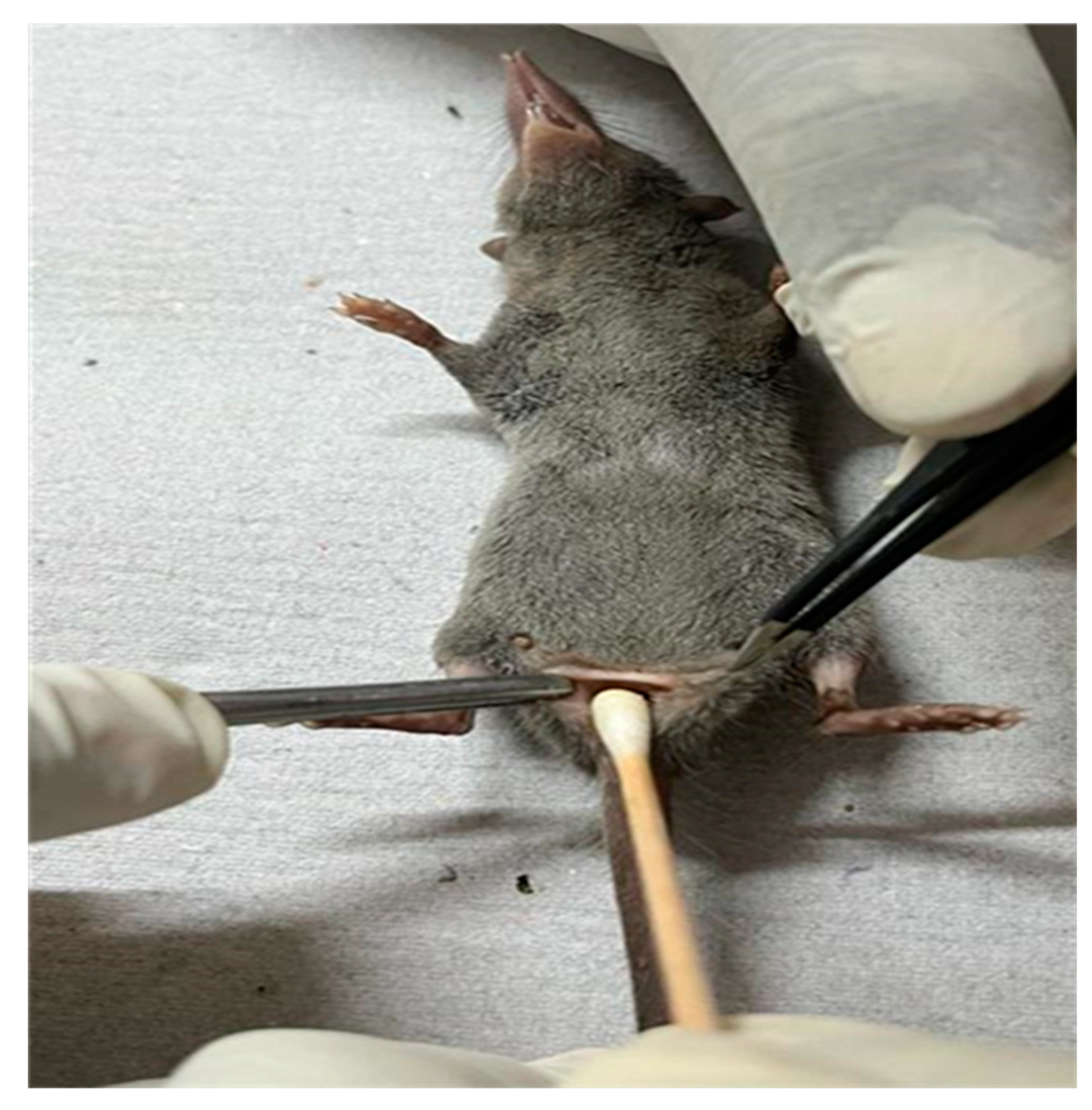

Table 1.
Details of the primers and annealing temperatures for the detection of the pathogen and antibiotic resistance genes harboured by them.
Table 1.
Details of the primers and annealing temperatures for the detection of the pathogen and antibiotic resistance genes harboured by them.
| Bacterial Pathogens | Targeted gene | Sequences | Annealing temperature | References |
|---|---|---|---|---|
| Staphylococcus sp | 16S rRNA | AACTCTGTTATTAGCGAAGAACA CCACCTTCCTCCGGTTTGTCACC |
50°C for 1 min | 16 |
| S. aureus | nuc | GCGATTGATGGTGATACGGT AGCCAAGCCTTGACGAACTAAAGC |
55°C for 30 sec | 17 |
| Methicillin resistance | mec A | TCCAGATTACAACTTCACCAGG CCACTTCATATCTTGTAACG |
50°C for 1 min | 18 |
| mec C | GAAAAAAAGGCTTAGAACGCCC GAAGATCTTTTCCGTTTTCAGC |
19 | ||
| E. coli | uspA | CCGATACGCTGCCAATCAGT ACGCAGACCGTAAGGGCCAGAT |
60°C for 1.5 mins | 20 |
| ESBL-producing E. coli | TEM | ATGAGTATTCAACATTTCCG CTGACAGTTACCAATGCTTA |
56°C for 1 min | 21 |
| SHV | AGGATTGACTGCCTTTTTG ATTTGCTGATTTCGCTCG |
22 | ||
| CTX-M | CAATGTGCAGCACCAAGTAA CGCGATATCGTTGGTGGTG |
59°C for 30 sec | 23 | |
| Salmonella sp | invA | GCCAACCATTGCTAAATTGGCGCA GGTAGAAATTCCCAGCGGGTACTG |
55°C for 45 sec | 24 |
| Salmonella enteritidis | spvC | GCCGTACACGAGCTTATAGA ACCTACAGGGGCACAATAAC |
57°C for 45 sec | 25 |
| Salmonella typhimurium | fliC | CGGTGTTGCCCAGGTTGGTAAT ACTGGTAAAGATGGCT |
55°C for 45 sec | 26 |
| Tetracycline resistance | Tet | GCACTTGTCTCCTGTTTACTCCCC CCTTGTGGTTATGTTTTGGTTCCG |
53°C for 1min | 27 |
| Sulphonamides resistance | sul3 & sul4 | TCAACATAACCTCGGACAGT GATGAAGTCAGCTCCACCT |
60°C for 40 sec | 28 |
| Fluoroquinolone resistance | qnrA | TCAGCAAGAGGATTTCTCA GGCAGCACTATGACTCCCA |
53°C for 30 sec | 29 |
Table 2.
Pathogens isolated from rodents/shrews and results of the molecular screening of genetic elements conferring antimicrobial resistance.
Table 2.
Pathogens isolated from rodents/shrews and results of the molecular screening of genetic elements conferring antimicrobial resistance.
| The animal reservoir | Staphylococcus spp. and the AMR genetic element tested positive in the rodent/shrew screened | |||||
|---|---|---|---|---|---|---|
| Staphylococcus aureus | Staphylococcus non-aureus | Staphylococcus aureus | ||||
| mec A | mec C | mec A | mec C | mec A and mec C | ||
| Rattus rattus | 10 | 2 | 2 | 0 | 1 | |
| Suncus murinus | 30 | 4 | 5 | 1 | 4 | |
| Total | 40 | 6 | 7 | 1 | 5 | |
| The animal reservoir | E.coli and the AMR genetic element tested positive in the rodent/shrew screened | |||||
| TEM | SHV | CTX-M | TEM and SHV | SHV and CTX-M | TEM, SHV, and CTX-M | |
| Rattus rattus | 6 | 8 | 2 | 2 | 6 | 1 |
| Suncus murinus | 15 | 37 | 9 | 11 | 1 | 1 |
| Total | 21 | 45 | 11 | 13 | 7 | 2 |
| The animal reservoir | Salmonella spp. and the AMR genetic element tested positive in the rodent/shrew screened | |||||
| tet | sul 3 and sul 4 | qnrA | tet and qnrA | tet and sul 3 & sul 4 | sul 3 & sul 4 and qnrA | |
| Rattus rattus | 3 | 5 | 2 | - | 2 | - |
| Suncus murinus | 10 | 15 | 9 | 2 | 4 | 1 |
| Total | 13 | 20 | 11 | 2 | 6 | 1 |
Table 3.
Genbank accession ID of the bacterial pathogens and their AMR gene sequences.
| Pathogen | Staphylococcus sp | S. aureus | MRSA | MRSA | ||
|---|---|---|---|---|---|---|
| Gene | (16S rRNA) | (nuc) | (mec A) | (mec C) | ||
| Genbank accession ID | OQ979122, OQ979123 | OQ992661, OQ992662 | OQ992663, OQ992664 | OQ992664 | ||
| Pathogen | E. coli | |||||
| Gene | (uspA) | TEM | SHV | CTX M | ||
| Genbank accession ID | OQ872769 | OR066219 | OR066218 | OR066217 | ||
| Pathogen | Salmonella spp | S. enteritidis | S. typhimurium | |||
| Gene | (invA) | (spvC) | (fliC) | tet | sul3 & sul4 | qnrA |
| Genbank accession ID | OR066220, OR066221 | OR066224, OR066225 | OR066222, OR066223 | OQ992668, OQ992669 | OQ992665, OQ992666 | OQ992667 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



