
Submitted:
19 January 2024
Posted:
22 January 2024
You are already at the latest version
Abstract
This study focused on evaluating the potential of natural fermentation of pea flour (F) to improve the release of antioxidant compounds. Two fermentation conditions of F were performed: 1: 36.4% w/w, 24 h, 30°C (FF1); 2: 14.3% w/w, 24 h, 37°C (FF2); pH values decreased to 4.4-4.7, with a predominance of lactic acid bacteria. An increment of the proteolysis degree (TNBS method) (greater for FF2), polypeptides aggregation and decrease of solubility, increase of < 2 kDa peptides, and increase in the ORAC potency of PBS-soluble fractions after fermentation was demonstrated. After simulated gastrointestinal digestion (SGID, COST-INFOGEST), FF1D and FF2D showed greater proteolysis degrees (63 and 53%) than F1D and F2D (45%), with differences in the molecular composition of the different digests. ORAC and HORAC potencies increased in all cases. FF2D presented the greater ORAC value, with higher activities for > 4 kDa, some in the range 2 to 0.3 kDa. and < 0.10 kDa fractions. Fermentation also increased the 60 % ethanol extracted phenolic compounds, mainly flavonoids, and the ORAC activity. After SGID, the flavan-3-ols disappeared but some phenolic acids increased respect to F. Fermentation in condition 2 was considered the most appropriate to obtain a functional antioxidant ingredient.
Keywords:
yellow peas
; natural fermentation
; gastrointestinal digestion
; antioxidant properties
1. Introduction
Legumes have a high content of protein, essential amino acids, fiber, vitamins B including folic acid, thiamine, and niacin, and minerals. In comparison, legumes contain twice the amount of protein as whole grain cereals (wheat, oats, and barley) and triple that of rice. Particularly, peas (Pisum sativum) have a protein content of 20–23.5 % w/w [1], which positions them as an important source of protein. Also, pulses are good sources of various phenolic compounds (PCs) including phenolic acids, flavonoids, isoflavones and tannins. Contents and types of PCs, which can be free, esterified or linked to other components, vary with the pulse type and genotype; for example, the PCs composition change with the seed coat color of the legume [2].
Some suitable options to increase the consumption of legumes in the daily diet is to use their flours in the formulas for making baked goods, and to improve their sensory and functional features through fermentation [3] that is one of the oldest biotechnological processes in the production of foods based on cereals and legumes. As indicated by Adebo et al. [4], fermented food products are sometimes classified as "functional foods" due to their potential health benefits. Natural or spontaneous fermentation is the most common way sourdough preparation in developing countries. The sourdough consists of a mixture of flour and water fermented with the native microbiota of the seeds. This natural fermentation occurs through the sequential and competitive action of an abundance of microorganisms, with the best-adapted strain(s) having a better growth rate, eventually dominating the microbiota [3, 4]. The original microbiota of grains, and therefore flour, is affected by various factors such as climate (temperature and humidity), storage conditions, insect attacks, and fungicide application. Another variable that affects the fermentation process is the type of grain due to differences in the quantity and quality of carbohydrates as fermentation starter substrates, in nitrogen sources, and in growth factors. This microbiota is composed of a great diversity of bacteria and especially lactic acid bacteria (LAB), and yeasts [4]. Throughout the fermentation, the microorganisms most adapted to the environmental conditions will prevail, generally the LABs.
Technological parameters used in the preparation of fermented products such as time, temperature, flour to water ratio making fermentation occur in a solid or liquid medium, and agitation, also have a key influence on microbial communities. In this sense, fermented doughs or flours can be classified into three types: type I prepared by a process of daily refreshments, type II obtained in a single fermentation step generally in a liquid medium in bioreactors, and type III corresponded to those of type II that are subsequently dried and stabilized. Each type will have different characteristics and uses, and the prevalence of different types of microorganisms has been shown [4,5,6].
Fermentation of seed-based products produces several shelf life, texture, flavor, and nutritional improvements. Several studies have demonstrated that fermentation (with starters or through the native flora) of legumes enhances their nutritive value, reduces some anti-nutritional endogenous compounds such as phytic acid, exerts beneficial effects on protein digestibility and their biological value, and produces the release of bioactive compounds [4]. It has beneficial effects on the irritable bowel syndrome since various compounds, including gluten, digestive enzyme inhibitors and certain carbohydrates or fermentable polyols could be hydrolyzed preventing the symptoms [4,7]. Besides, it influences the bioavailability of phytochemicals, particularly on phenolic compounds (PCs) by releasing bound or non-extractable PCs and their aglycones. Also, proteolysis occurs through endogenous seed proteases that can be activated by lowering the pH, or by microbial proteases and peptidases. In addition to the functional, physical and chemical aspects of protein modification, fermentation can produce the liberation of bioactive peptides by protein hydrolysis which would confer positive effects on human health [8]. Regarding antioxidant activity, fermentation could produce an increase through different mechanisms, such as the release of antioxidant peptides and the release and/or transformation of PCs. It has also been reported that microorganisms could increase the antioxidant capacity of fermented products through the secretion of antioxidant enzymes, glutathione, and other biomolecules such as exopolysaccharides with this activity [4,8].
The literature on the antioxidant effect of pea-derived peptides is scarce. Therefore, a first evaluation of the antioxidant capacity of yellow pea flour and protein isolate was carried out before and after being subjected to a simulated gastrointestinal digestion process (SGID) [1]. Also, we have studied the profile and antioxidant activity of PCs and the effect of SGID (Cipollone et al., under revision). An interesting antioxidant activity of peptides and PCs after gastrointestinal digestion of protein isolates and flour was demonstrated. Furthermore, while there is quite a bit of work done on legume fermentation and some on pea fermentation with starters, so far we have not found bibliography specifically studying indigenous pea fermentation, let alone the effect on gastrointestinal digestion and antioxidant activity. In the present investigation, natural fermentation of yellow pea flour was performed under different conditions, evaluating its effect on the proteolysis, PCs profile, and antioxidant activity of peptides and PCs fractions before and after a SGID, in order to obtain an ingredient with improved antioxidant activity potential.
2. Materials and Methods
2.1. Chemicals and Samples
Alfa-amylase from Bacillus subtilis (10070, 57.4 U/mg), pepsin from porcine gastric mucosa P6887 (3200-4500 U/mg), porcine pancreatin 4XUSP P1750, bovine bile salts B3883, Trolox (6-hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid), blue dextran, aprotinin, hippuric acid, and 2,4,6-trinitrobencenesulfonic acid (TNBS) were purchased from Sigma Chemical Co. (St. Louis, MO, USA). Fluorescein sodium 2,2′-Azinobis-(3-ethylbenzthiazolin-6-sulfonic acid) (ABTS) were from Fluka (Steinheim, Germany), AAPH (2,2´-Azo-bis-(2-methylpropionamidine) dihydrochloride) was from Aldrich (Wisconsin, USA). Nutritive agar (NA) was from Britania (Argentina); Man, Rogosa & Sharpe (MRS) media and Chloramphenicol Yeast Glucose Agar (YGC) were from Biokar Diagnostics (Argentine). Other reagents were of analytical grade.
Yellow pea seeds (Pisum sativum var. Yams) cultivated in Buenos Aires (Argentina) were ground in an Udy mill (0.5 mm mesh) to prepare the flour (F).
2.2. Autochthonous Fermentation
2.2.1. Preliminary Tests
A preliminary screening to analyze different fermentation conditions was carried out. In a first stage, the tests were carried out in tubes with agitation in a rotary shaker. Flour was dispersed in distilled water in a ratio of 1/1.75 (flour concentration: 36.4 % w/w); two times (24 and 48 h) and two temperatures (30 and 37°C) of incubation were evaluated. After fermentation, the samples were frozen. The final pH, proteolysis degree, total and soluble protein contents, and antioxidant activity (ORAC) of the four fermented flours were analyzed according to the methodologies described later.
2.2.2. Preparation of Fermented Flour
After the preliminary tests, fermentation process was performed in a bioreactor with a glass jacket connected to a water bath (LAUDA RMT6) that allows the recirculation of thermostatic water. Agitation was achieved by using a vertical stirrer Dlab OS20-PRO20L (250 rpm). Two tests were carried out under different conditions. In the first one, flour was dispersed in distilled water in a ratio of 1/1.75 (similar to tube assay) and the mixture was incubated at 30°C for 24 h (FF1). In the second, the flour/distilled water ratio was 1/6 and the fermentation was done at 37°C for 24 h (FF2). Dispersions of flour in distilled water without fermentation were used as a control (F1 and F2). pH measurements were taken at different times during the incubations. After fermentation, the samples were lyophilized.
2.2.3. Microbiological Analysis
The fermented (FF1/FF2) and non-fermented (F1/F2) samples were serially diluted in selective agar media for isolation and counting of different microorganisms: MRS incubated for 48 h at 30°C in anaerobiosis; NA incubated for 24 h at 30°C; and YGC incubated for 48 h at 30°C. All the colonies obtained were tested by the Gram staining and the catalase test.
2.3. Centesimal Composition
The protein content was determined by the micro-Kjeldahl method (f = 5.6 g protein/g N [9]) followed by colorimetric determination [10]; moisture and ash were determined according to AOAC 1984 (24.002 and 24.009) [11], lipids according to AOAC 1990 (920.39) [12]. Megazyme kit (Megazyme International Ltd., Wicklow, Ireland) was used for determining total dietary fibre (TDF) (AOAC 1995, 991.43) [13]. Total carbohydrates were obtained by difference [14].
2.4. Simulated Gastrointestinal Digestion (SGID)
The protocol of Minekus et al. [15] was applied on fermented and non-fermented samples obtaining the corresponding digest (FF1D, FF2D and F1D, F2D).The process was performed in a bioreactor with a glass jacket connected to a water bath (LAUDA RMT6) at 37°C with agitation (90 rpm oral and gastric phase, 100 rpm intestinal phase, vertical stirrer Dlab OS20-PRO20L). The pH was monitored during the digestion process using a pH-meter (Van London Co.). Oral phase: samples (about 50 g FF1, FF2 and F1, F2) were homogenized with 35 mL of electrolite solution for the simulated salival fluid (SSF, pH = 7), 5 mL of α-amilase solution in SSF (26 mg/mL), 250 µL 0.3 mol/L CaCl2 and 9.75 mL H2O were added (all reactives preincubated at 37°C). The mixture was agitated and incubated 2 min at 37°C. Gastric phase: the oral solution was mixed with 75 mL of the electrolyte solution for the simulated gastric fluid (SGF, pH = 3), 16 mL of pepsin solution (25000 Anson U/ml in SGF) and 50 µL 0.3 mol/L CaCl2, adjusting the pH to 3 with 2 eq/L HCl and adding water to complete 100 mL of SGF. The mixture was incubated along 2 h at 37°C. Intestinal phase: gastric solution (200 mL) was mixed with 110 mL of the electrolyte solution for the simulated intestinal fluid (SIF), 50 mL of pancreatin solution (800 TAME U/ml in SIF), 25 mL of bovine bile salts (150 mg/mL), and 400 μL 0.3 mol/L CaCl2, the pH was adjusted to 7 with 1 mol/L NaOH and water was added to complete 200 mL of SIF. The mix was incubated along 2 h at 37°C. After that, enzyme activities were inactivated by incubation at 85°C for 10 min. Electrolyte solutions for SSF, SGF and SIF were prepared according to [15].
2.5. Characterization of Polypeptide/Peptide Fraction
2.5.1. Protein Hydrolysis Degree (HD)
HD was measured by the 2,4,6-trinitrobencenesulfonic acid (TNBS) method [16,17]. HD was calculated as follows: HD % = 100×[-NH2]h/[-NH2]∞; where [-NH2] indicates the concentration of free amino groups in the hydrolyzed samples (h). The parameter [-NH2]∞ was estimated according to [-NH2]∞ = 1/Maa×(1+fLys)×Cprot, where Maa is the average molecular weight of amino acids (169.42 g/mol), fLys is the proportion of lysine (1/17.8) (values calculated from amino acid composition of peas [18]), and Cprot is the protein concentration
2.5.2. Glycine-SDS-PAGE [19]
Freeze-dried samples were dispersed in 0.0625 mol/L Tris-HCl, 2 % SDS, 10 % v/v glycerol buffer (pH = 8.8) and centrifuged before loading in the gel. Separating and stacking gels (120 and 40 g/L acrylamide, respectively) were used. Runs were carried out in a Mini Protean II Dual Slab Cell (BIO-RAD) equipment at room temperature, applying a constant current (30 mA/gel) in the case of SDS-PAGE and varying between 30 mA/gel and 100 mA/gel after passing the stacking gel for tricine-SDS-PAGE. Gels were stained with Coomasie Brilliant Blue R-250 (1 g/L). Silver staining was applied to increase analytical sensitivity [20].
2.5.3. Aqueous Extraction
Suspensions (20 mg/mL) of freeze-dried samples in PBS (KH2PO4 1.5 mmol/L, NaCl 138 mmol/L, KCl 3 mmol/L, Na2HPO4 8.1 mmol/L, pH = 7.4) were prepared by agitation at 500 rpm (1 h, 37°C) (Termomixer Eppendorf), and then centrifugation (10000×g, 10 min, room temperature, Hermle, Labortechnik GmbH, Germany). Soluble protein concentration was determined by the Lowry method [21].
2.5.4. Gel Filtration FPLC Chromatography
Soluble fractions (see 2.5.3) were analyzed in ÄKTA purifier (GE Healthcare) equipment using a molecular exclusion column. Superdex Peptide 10/300 GL (GE Healthcare) (exclusion limit = 10 kDa; separation range = 0.1-7 kDa) was calibrated with blue dextran (exclusion volume Vo = 7.60 mL), aprotinin (6.5 kDa), vitamin B12 (1.35 kDa) and hypuric acid (0.18 kDa) obtaining the following calibration curve: log MM = 4.84 – 3.30 Kav, where Kav = (Ve - Vo)/(Vt - Vo), Ve is the elution volume of the resolved species, Vo is the void volume, and Vt is the total volume of the column (Vt = 24 ml). Samples were filtered by a 0.45 µm nylon filter, 200 µL of sample were loaded and eluted with PBS buffer at 0.5 mL/min for Superdex 30. Detection at 210 nm was performed. Fractions (500 µL) were collected.
2.6. Characterization of PCs
2.6.1. Ethanol Extraction
Extractions were performed according to previous optimization in our lab [unpublished]. Ultrasound-assisted extraction (UAE) in a VCX 750 ultrasonic processor (Sonics & Materials Inc., Newtown, USA) was applied using a mixture 60:40 ethanol/water and the following conditions: 15 min, 40 % amplitude. The extractions were carried out in an ice bath to avoid excessive increases in temperature, which was maintained in all cases below 42°C. The extracts were centrifuged (Hermle, Labortechnik GmbH, Germany, 5 min, 10000×g, room temperature). The supernatants were evaporated (30°C, 2 h cycles, Concentrator plus/Vacufuge® plus) and resuspended in PBS buffer.
2.6.2. Total PCs Content (TPC)
TPC was determined using Folin-Ciocalteu method [22]. To 325 µl of sample, 50 µl of 1 eq/L Folin reagent was added, mixed by shaking and 3 min later, 375 µl of 20% w/v Na2CO3 were added. The mixture was allowed to stand for one hour in the dark. After that, the absorbance at 760 nm was measured in a microplate reader (SYNERGY HT SIAFRT, Biotek Instruments, USA). In parallel, a standard curve was made with gallic acid (0-0.06 mg/mL). Results were expressed as mg gallic acid equivalent (GAE)/g sample in dry matter (dm). All determinations were performed at least in duplicate.
2.6.3. Gel Filtration FPLC Chromatography
Ethanolic extracts were analyzed according to 2.5.4.
2.6.4. Profiling and Quantification of PCs
For the determination of the qualitative and quantitative profiles of the PCs, dry extracts were dissolved in the initial mobile phase of the chromatographic method at the time of analysis. Separation and determination of PCs was performed in a high performance liquid chromatography coupled with diode-array and fluorescence (HPLC-DAD-FLD) detectors (Dionex Ultimate 3000 system, Dionex Softron GmbH, Thermo Fisher Scientific Inc., Germering, Germany) and a reversed-phase Kinetex C18 column (3.0×100 mm, 2.6 mm; Phenomenex, Torrance, CA, USA). The software Chromeleon 7.1 was used to control all the parameters of the system and to process the obtained data. The list of PCs determined, chromatographic and detection conditions were those reported by Ferreyra et al. [23], with little modifications. The mobile phases were an aqueous solution of 0.1% formic acid (eluent A) and acetonitrile (eluent B). The gradient applied was as follows: 0-1.7 min, 5% B; 1.7-10 min, 30% B; 10-13.5 min, 95% B; 13.5-15 min, 95% B; 15-16 min, 5% B; 16-19, 5% B. The flow rate was set at 0.8 mL/min, the column temperature was 35°C and the injection volume was 10 μL. The analytical flow cell for DAD was set to scan from 200 nm to 400 nm; a data collection rate of 5 Hz, a band width of 4 nm and a response time of 1 s were used. Different wavelengths (254, 280, 320 and 370 nm) were used according on the maximum absorbance of analytes for DAD. For FLD, an excitation wavelength of 290 nm and a monitored emission response of 315, 360 and 400 nm were used depending on the targeted analytes. A data collection rate of 10 Hz was used for FLD. The retention times of compounds in samples were compared with those of authentic standards for the identification of PCs. Calibration plots for studied analytes showed linear ranges between 0.05 and 40 mg/L (r2 > 0.993) for most of the analytes.
2.7. Antioxidant Activity
2.7.1. Oxygen Radical Absorbance Capacity (ORAC)
The ORAC assay was carried out using previously optimized protocols in our laboratory [24]. A 53.3 nmol/L fluorescein solution in phosphate buffer (150 μL) was mixed with 25 μL of sample or the same volume of either the phosphate buffer (negative control) or Trolox (positive control), and then pre-incubated at 37°C for 10 min; 160 mmol/L AAPH (25 μL) in phosphate buffer was added and the reaction mixture was incubated at 37°C for 45 min. The fluorescence intensity (λexc: 485, λem: 535 nm) was read every min in a SYNERGY HT– SIAFRT™ multidetection microplate reader (Biotek Instruments, USA) to obtain the fluorescein-decay curve. The area under curve was obtained as AUC = 0.5 + f1/f0 + f2/f0 + …+ fi-1/f0 + 0.5 fi/f0, where f is the fluorescence value at a particular time during the decay. A blank without AAPH was included and the % scavenging calculated as: % ROO· scavenging = [(AUCS–AUCNC)/ (AUCB-AUCNC)]×100; where S = sample, B = blank, NC: negative control. Trolox (6.25–75.0 μmol/L) was used as a reference compound. The concentration that inhibits the 50 % of radicals (IC50) was obtained from dose-response curves. PBS and separated FPLC fractions as well as ethanolic extracts were analyzed.
2.7.2. Hydroxyl Radical Averting Capacity (HORAC) [24]
The hydroxyl radical was generated by a cobalt-mediated Fenton-like reaction with fluorescein used as a probe. Either samples or buffer (20 μL) were mixed with 190 μL of 60.3 nmol/L fluorescein solution in phosphate buffer, 15 μL of 0.75 mol/L H2O2 solution, and 75 μL of the cobalt solution (10 mg of picolinic acid and 11 mg of CoCl2.6H2O in 50 mL of water) were added. Mixture was incubated at 37°C for 3 h in the SYNERGY HT microplate reader; and the fluorescence (λexc: 485, λem: 535 nm) was read at 1-min intervals to obtain the AUC. The percent inhibition was calculated as: % OH· inhibition = [(AUCS– AUCNC)/(AUCB-AUCNC)]×100; where: S = sample, B = blank (without addition of the cobalt and hydrogen-peroxide solutions), and NC = negative control. Chlorogenic acid (0.05–0.5 mg/mL) was used as a reference compound. The concentration that inhibits the 50 % of radicals (IC50) was obtained. PBS fractions were analyzed.
2.7.3. ABTS Free Radical Scavenging Capacity
The ABTS•+ radical cation decolorization method was carried out according to Tironi and Añón [25], with some modifications. The ABTS•+ radical was obtained by reaction of a 7 mmol/L solution of ABTS with potassium persulfate (final concentration: 2.45 mmol/L) incubated at room temperature in the dark for at least 16 h. Prior to the assay, the ABTS•+ solution was diluted in PBS to obtain an absorbance at 734 nm of 0.70 ± 0.02 in a 1 cm cuvette with a Beckman DU 650 spectrophotometer. Mixtures of 1000 µL of the diluted ABTS•+ solution with 100 µL of the resuspended extracts were prepared in 48-well plates and the absorbance at 734 nm was measured at various times (0 – 15 min) in a SYNERGY HT microplate reader. SIAFRT (Biotek Instruments, USA). Trolox (0.05-0.20 mmol/L) was used as the reference compound. With the measurements made for each sample and the blank (PBS buffer), the % inhibition of the radical was calculated using the following equation: % de ABTS•+ inhibition = [(AbsB0 – AbsS15) - (AbsB0 – AbsB15) x 100] / AbsB0; where AbsB0 and AbsB15 refer to the absorbance of the blank at 0 and 15 min, respectively, and AbsS15 refers to the absorbance of the sample at 15 min. Ethanolic fractions were analyzed.
2.8. Statistical Analysis
Determinations were performed on at least two independent batches. Differences between samples were analyzed by one way-ANOVA multiple comparisons. Significant differences (p < 0.05) among mean values were evaluated by the Tukey HSD test (Statgraphics Centurion XVI).
3. Results and Discussion
3.1. Preliminary Assays on Natural Fermentation of F
Our first studies on the natural fermentation of yellow pea flour were related to the evaluation of fermentation conditions that produced a certain degree of proteolysis with the potential release of antioxidant peptides. In this way, a dispersion containing 36.4 % w/w of F and four time/temperature pairs were evaluated (24 and 48 h, 30 and 37°C). The final pH, proteolysis degree (HD), protein solubility, and antioxidant activity (ORAC, PBS-soluble fractions) of the four fermented flours were analyzed (Table 1). The final pH value achieved was different according to the incubation time as was expected, reaching 3.6 and 3.9 ± 0.1 after 48 h at 30 and 37°C, respectively, and 5.1 ± 0.1 and 4.7 ± 0.2 after 24 h at 30 and 37°C, respectively, showing no significant differences (p > 0.05) between both incubation temperatures at any time. In addition, no significant (p > 0.05) differences in the proteolysis degree (HD) and significant (p < 0.05) but small differences in the ORAC IC50 value were obtained among the different fermented products (Table 1). According to these results, it was decided to continue working with fermentations of 24 h (30 and 37°C) since a prolongation of the time up to 48 h did not generate significant changes neither in HD % nor in the solubility of polypeptides/peptides, and only very minor differences in ORAC activity.

Table 1.
Preliminary tests of natural fermentation of yellow pea flour dispersions (36.4 % F w/w) performed in tube (10 mL) under different time/temperature conditions.
Table 1.
Preliminary tests of natural fermentation of yellow pea flour dispersions (36.4 % F w/w) performed in tube (10 mL) under different time/temperature conditions.
| Fermentation | Proteolysis | Protein solubility | ORAC | |
|---|---|---|---|---|
| condition | Final pH | HD %1 | (g SP/100gTP)2 | IC50 (mg SP/mL) |
| 24 h/30°C | 5.1 ± 0.1b | 16.8 ± 0.7a | 52 ± 5a | 0.071 ± 0.004a |
| 48 h/30°C | 3.6 ± 0.1a | 17 ± 2a | 54 ± 4a | 0.093 ± 0.004b |
| 24 h/37°C | 4.7 ± 0.2b | 13 ± 2a | 54 ± 3a | 0.087 ± 0.005b |
| 48 h/37°C | 3.9 ± 0.1a | 13 ± 2a | 57 ± 8a | 0.066 ± 0.002a |
1 determined in the corresponding dispersions. 2 SP: PBS-soluble protein. Different letters in the same column indicates significant differences (p < 0.05).
3.2. Preparation of Fermented Flours in Bioreactor
Taking into account the information obtained in the preliminary tests, natural fermentation was carried out in a bioreactor. Two different systems were studied:
Test 1. The fermented flour FF1 was obtained by using 36.4 % w/w F dispersion and incubating for 24 h at 30°C. The evolution of the pH was recorded, values drop from 6.2-6.3 (t = 0) up to a final value (24 h) of 4.75 (Table 2). Thus, pH values decreased to a greater extent than in tube fermentation in the same conditions (Table 1). It should be noted that this dispersion presented a high viscosity being in consequence difficult to stir.
Test 2. The fermented flour FF2 was obtained using a lower F concentration (14.3 % w/w) and incubating for 24 h at 37°C to achieve a greater fluidity of the dispersion and an easy agitation. The pH value achieved after 20 h of incubation was 4.75 (similar to those reached at 24 h by FF1) and after 24 h was about 4.4 (Table 2), showing a slightly higher decline (p < 0.05) than in FF1. These results suggest an increase in the fermentation rate under conditions of lower flour/water ratio. Sáez et al. [26] informed a pH decrease from 6.30-6.43 to 4.80-4.83 after the first back-slopping for the natural fermentation (24 h at 37°C) of different varieties of beans in 1 g/mL flour/water dispersions.
A microbiological screening of the non-fermented and fermented samples using nutrient agar (NA) (total mesophilic aerobic bacteria), YGC (fungi and yeasts), and MRS with selection factor to observe growth of LAB was performed. Microbiological counts of samples before fermentation (t = 0) were in the range of 4.2 and 4.5 log cfu/g both in NA and MRS, with no evident growth on YGC medium (Table 2). These results are comparable to other previously reported which presented values of 4.6 log cfu/g for unfermented chickpea flour [27] and are slightly higher than those obtained by Rizzello et al. [28] for Faba bean flour (3.6 log cfu/g). After fermentation, counts around 9 log cfu/g (NA and MRS) were registered for FF1 and FF2 (Table 2). The bacterial colonies that grew in the MRS medium presented smooth edges, white color, and a creamy appearance. When observing under the microscope, these colonies presented chained coccus-type and bacilli morphologies. According to the Bergey's Manual, Gram staining (positive) and catalase test (negative) carried out on the different colony-forming units, indicated that these colonies could be presumptively identified as LAB. Generally, a sourdough contains a variable number of LAB ranging from 7 to 9 log cfu/g [29]. Values around 8 log cfu/g have been reported for different beans sourdoughs after 6 days of fermentation [26]. No growth was observed in the YGC medium indicating undetectable count of yeasts after the fermentation process in the lower dilution performed.
At this point it is worth mentioning that tests 1 and 2 were carried out with different batches of peas (different harvest years and different storage times) and microbiological counts were similar both for the initial microbiota and for the fermented samples.
Table 2.
Natural fermentation of yellow pea flour dispersions performed in bioreactor: final pH values, and microbial count in YGC, MRS and NA.
Table 2.
Natural fermentation of yellow pea flour dispersions performed in bioreactor: final pH values, and microbial count in YGC, MRS and NA.
| Sample | pH | Microbial count (log cfu/g) | ||
| YGC | MRS | NA | ||
| F1 | 6.2 ± 0.1c | nd | 4.2 ± 0.1a | 4.2 ± 0.1a |
| FF1 | 4.75 ± 0.03b | nd | 9.1 ± 0.6b | 8.9 ± 0.6b |
| F2 | 6.29 ± 0.01c | nd | 4.5 ± 0.3a | 4.4 ± 0.2a |
| FF2 | 4.43 ± 0.01a | nd | 9.5 ± 0.3b | 8.8 ± 0.8b |
F1 and F2: pea flour dispersions in condition 1 and 2, respectively. FF1 and FF2: Fermented pea flour dispersions in conditions 1 and 2, respectively. Condition 1: 36.4 % w/w F, 24 h, 30°C; Condition 2: 14.3 % w/w F, 24 h, 37°C. nd: not detected growth in dilution -2. Different letters in the same column indicates significant differences (p < 0.05).
3.3. Composition of the Fermented Flours
The macro-components composition of the freeze-dried fermented flours was determined in comparison with the non-fermented flour (Table 3). There were no significant differences (p > 0.05) in the ash and lipid contents for any of the samples, with values comparable with those previously informed for Canadian peas [30]. Fermented flours did not present significant changes in the protein content respect to the corresponding non-fermented samples. Regarding the dietary fiber, the content obtained for F1 and F2 was comparable to others previously reported for pea seeds (15.3 %, [31]). In relation to the effect of fermentation on this component, there was a significant increase (p < 0.05) in the case of FF2. It has been informed that spontaneous fermentation of mung bean increased the crude fiber content [32]. However, other study [33] reported that pea, chickpea, and grass pea flours containing high levels of dietary fiber did not show significant variations after the fermentation process (L. plantarum or L. brevis, 24 h, 30°C). Also, natural fermentation of lupin and soy did not affect the contents of soluble, insoluble, and total fiber [34]. Further studies -which are not the subject of this work- will be necessary to analyze the effect of fermentation on the fiber composition and try to explain the small increase recorded for FF2 and its potential health benefit.
Table 3.
Composition % of freeze-dried yellow pea flour before and after fermentation in bioreactor.
Table 3.
Composition % of freeze-dried yellow pea flour before and after fermentation in bioreactor.
| Sample | Proteins1 | Lipids1 | Glucides1* | Fiber1 | Ash1 | Moisture |
|---|---|---|---|---|---|---|
| F1 | 17.9 ± 0.3a | 2.3 ± 0.3a | 61.6 | 15.3 ± 0.8a | 2.8 ± 0.5a | 4.61 ± 0.06ab |
| F2 | 24.2 ± 0.9c | 2.3 ± 0.2a | 55.4 | 15.2 ± 0.8a | 2.8 ± 0.5a | 4 ± 1a |
| FF1 | 22.1 ± 0.3bc | 2.1 ± 0.1a | 55.4 | 17.2 ± 0.2ab | 3.1 ± 0.1a | 4.12 ± 0.04a |
| FF2 | 21.9 ± 0.3b | 2.3 ± 0.1a | 53.1 | 18.8 ± 0.6b | 3.8 ± 0.1a | 5.73 ± 0.08b |
1Contents are expressed as g/100 dry matter. *Carbohydrates were obtained by difference. F1 and F2: pea flour dispersions in condition 1 and 2, respectively. FF1 and FF2: Fermented pea flour dispersions in conditions 1 and 2, respectively. Condition 1: 36.4 % w/w F, 24 h, 30ºC; Condition 2: 14.3 % w/w F, 24 h, 37ºC. Different letters in the same column indicates significant differences (p < 0.05).
3.4. Changes in the Protein Fraction of Fermented Flours
As in the case of 10 mL (tube) fermentations, a partial proteolysis was evidenced according to the increment of the HD value. Although FF1 and FF2 presented similar HD values, we can remark that the HD value doubled in the case of FF1 and increased 5 times in the case of FF2 with respect to the corresponding initial values (Table 4). In this way, there was a greater level of protein hydrolysis in the case of FF2. A complementary test was carried out to determine if endogenous proteases from pea seeds could be activated by the drop in pH, producing proteolysis. The mobilization of storage proteins in germinating seeds is initiated by endo-proteases which convert the water insoluble storage proteins into soluble peptides. Most of the plant proteases are neutral or alkaline and there are few acid proteases (pH optimum: 2-3) widely distributed in the plant seeds [35]. In the case of cereals, the comparison of wheat and rye sourdoughs and chemically acidified doughs indicated that primary proteolysis is mainly attributable to endogenous proteases [36]. To evaluate that, the pH of an F dispersion (14.3 % w/w) was lowered with 2 N HCl to the final value obtained in the fermentations (4.4) and the degree of protein hydrolysis was measured (TNBS method). A very low value (close to 0), even lower than those registered for flour dispersions in water before fermentation, was obtained. According to this, no activation of endogenous proteases was evidenced. In agreement, Akhtaruzzaman et al. [37] extracted the proteases from seven overnight imbibed leguminous seeds and found that the alkaline proteases involved in all seeds were more potent than the acidic proteases. In consequence, the proteolysis in the fermented samples would be the product of the action of proteases from microorganisms. LAB strains displayed a wide range of proteolytic activities [27].
Comparing the results obtained for 10 mL (tube) fermentations and in reactor under the same conditions (36.4 % w/w, 24 h, and 30 °C), there was no significant difference (p > 0.05) in the protein solubility values (52 and 56 %, respectively, Table 1 and Table 4). In all cases, the solubility decreased in the fermented samples with respect to the non-fermented ones, which could be due to the formation of aggregates during the fermentation process, as will be discussed next.
Table 4.
Protein-related analysis (proteolysis degree1, protein solubility and antioxidant activity of PBS-soluble fractions2) of yellow pea flour before and after fermentation and after SGID.
Table 4.
Protein-related analysis (proteolysis degree1, protein solubility and antioxidant activity of PBS-soluble fractions2) of yellow pea flour before and after fermentation and after SGID.
| Sample | Proteolysis | Soluble Protein | Protein solubility | ORAC IC50 | HORAC IC50 |
|---|---|---|---|---|---|
| HD % | (SP) (mg/mL) | (g SP/100gTP) | (mg SP/mL) | (mg SP/mL) | |
| F1 | 8.5 ± 0.6a | 2.6 ± 0.1 | 75 ± 5cd | 0.178 ± 0.019d | 7.4 ± 0.5b |
| FF1 | 17 ± 2b | 2.3 ± 0.1 | 56 ± 4ab | 0.071 ± 0.007c | 7.7 ± 0.5b |
| F1D | 45 ± 2c | 4.1 ± 0.6 | 86 ± 10cd | 0.049 ± 0.003bC | 3.7 ± 0.2a |
| FF1D | 64 ± 4e | 3.8 ± 0.7 | 76 ± 10cd | 0.024 ± 0.001aB | 3.8 ± 0.3a |
| F2 | 4 ± 1a | 3.3 ± 0.1 | 71 ± 2bc | 0.089 ± 0.001c | 7.9 ± 0.9b |
| FF2 | 20 ± 2b | 2.1 ± 0.2 | 49 ± 3a | 0.033 ± 0.007ab | 7 ± 1b |
| F2D | 44 ± 3c | 3.5 ± 0.4 | 88 ± 8d | 0.017 ± 0.001aA | 3.6 ± 0.4a |
| FF2D | 53 ± 4d | 2.9 ± 0.2 | 79 ± 5cd | 0.017 ± 0.001aA | 3.6 ± 0.3a |
F1 and F2: pea flour dispersions in condition 1 and 2, respectively. FF1 and FF2: Fermented pea flour dispersions in conditions 1 and 2, respectively. F1D, F2D, FF1D, FF2D: simulated gastrointestinal digests. Condition 1: 36.4 % w/w F, 24 h, 30ºC; Condition 2: 14.3 % w/w F, 24 h, 37ºC. 1 determined in the corresponding dispersions. 2 determined in 20 mg. mL-1 dispersions of freeze dried samples. Different lowercase letters in the same column indicates significant differences (p < 0.05) when a global Tuckey analysis was performed. Capital letters in case of ORAC assay indicates significant differences (p < 0.05) when a Tuckey analysis among the four digests was performed.
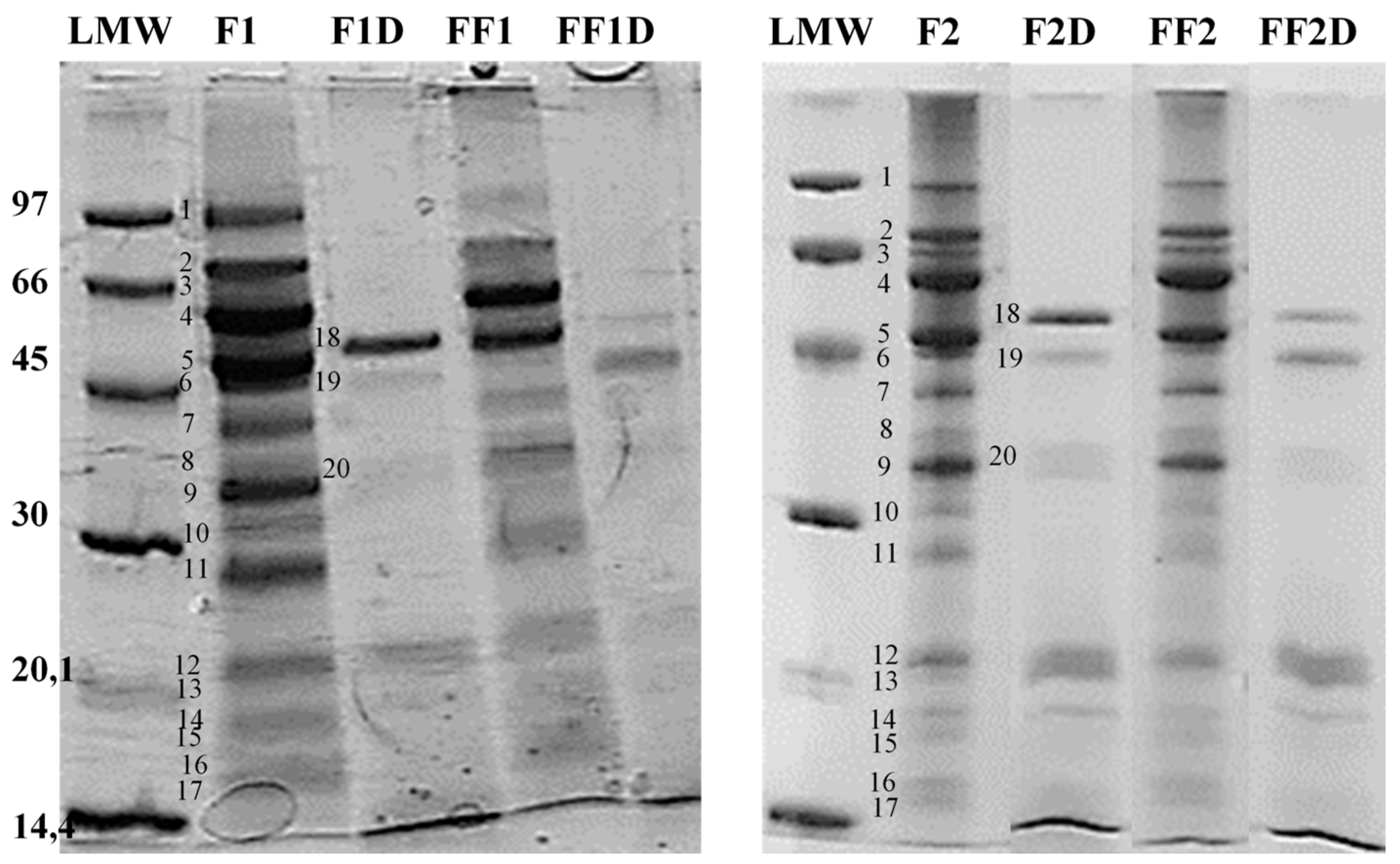
The changes in the peptide/polypeptide profile produced by the fermentation were analyzed by glycine-SDS-PAGE. The glycine-SDS-PAGE profile of F1 and F2 (Figure 1) showed a great variety of polypeptides between 14 and 97 kDa. It was possible to detect bands tentatively belonging to linoleate 9S-lipoxygenase (band 1, 93 kDa); alpha-dioxygenase (band 2, 77 kDa); convicilin (an important storage protein of peas, 70 kDa, band 3); legumin subunits (59 kDa, band 4); free acidic (40 kDa, band 7) and basic (band 13, 20 kDa) legumin subunits; vicilin subunits (53 kDa, band 5; 34 kDa, band 9; pea vicilin is heterogeneous, variable polypeptides could be produced by different gene coding), band 6 (probably alpha-galactosidase, 45 kDa); band 11 and 12 (28 and 25, probably subunits/polypeptides of albumin-2); and bands 14 to 17 (20-14 kDa) would correspond to albumins. The recognition of the pea polypeptides was carried out according to Ma et al. [38].
After fermentation, a decrease in the intensity of all bands was observed, being more evident for 93 kDa (band 1) and for bands corresponding to MW < 40 kDa. Also, an increase of high MW molecules that did not enter the gel could be appreciated in fermented samples suggesting the presence of aggregates that remain even in the presence of SDS and urea (Figure 1). The formation of aggregates could explain the decrease in solubility observed in fermented samples (Table 4); this fact can be at least partially explained since pea proteins have their minimum solubility at the isoelectric point (between 4 and 5), coinciding with the final pH value in fermented flours. Band 10 (31 kDa, which could correspond to the anti-nutritional factor lectin [38]) appeared much more intense in samples F2 and FF2 than in F1 and FF1, while band 12 (25 kDa, which could include Kunitz-type trypsin inhibitor-like 2 protein [38]) has a higher intensity for F1 and FF1 respect to F2 and FF2 (Figure 1). Beyond these differences between both dispersions, the intensity of these bands decreased after fermentations suggesting a diminution in the mentioned anti-nutritional factors. The reduction of the color intensity of several bands after fermentation could be associated with polypeptides diminution due to proteolytic activity. Byanju et al. [39] also observed this pattern of band discoloration in pea, lentil, and soybean flours after fermentation with Lactiplantibacillus plantarum and Pediococcus acidilactici. Also, the fermentation of pea flour with three LAB (Pediococcus pentosaceus, Lactococcus raffinolactis, and Lactiplantibacillus plantarum) resulted in similar patterns of the Coomassie brilliant blue stained gels, not very different from the extract of the unfermented flour, except for the disappearance of some high molecular weight bands [40].
Figure 1.
Electrophoresis SDS-PAGE of freeze-dried samples solubilized in electrophoresis buffer. F1: 36.4 % w/v F dispersion; F2: 14.3 % w/v F dispersion; FF1: fermented F in condition 1 (36.4 % w/v F dispersion, 24 h, 30°C); FF2: fermented F in condition 2 (36.4 % w/v flour dispersion, 24 h, 37 °C); F1D: F1 after SGDI; F2D: F2 after SGDI; FF1D: FF1 after SGDI; FF2D: FF2 after SGDI. LMW: low molecular weight standard.
Figure 1.
Electrophoresis SDS-PAGE of freeze-dried samples solubilized in electrophoresis buffer. F1: 36.4 % w/v F dispersion; F2: 14.3 % w/v F dispersion; FF1: fermented F in condition 1 (36.4 % w/v F dispersion, 24 h, 30°C); FF2: fermented F in condition 2 (36.4 % w/v flour dispersion, 24 h, 37 °C); F1D: F1 after SGDI; F2D: F2 after SGDI; FF1D: FF1 after SGDI; FF2D: FF2 after SGDI. LMW: low molecular weight standard.
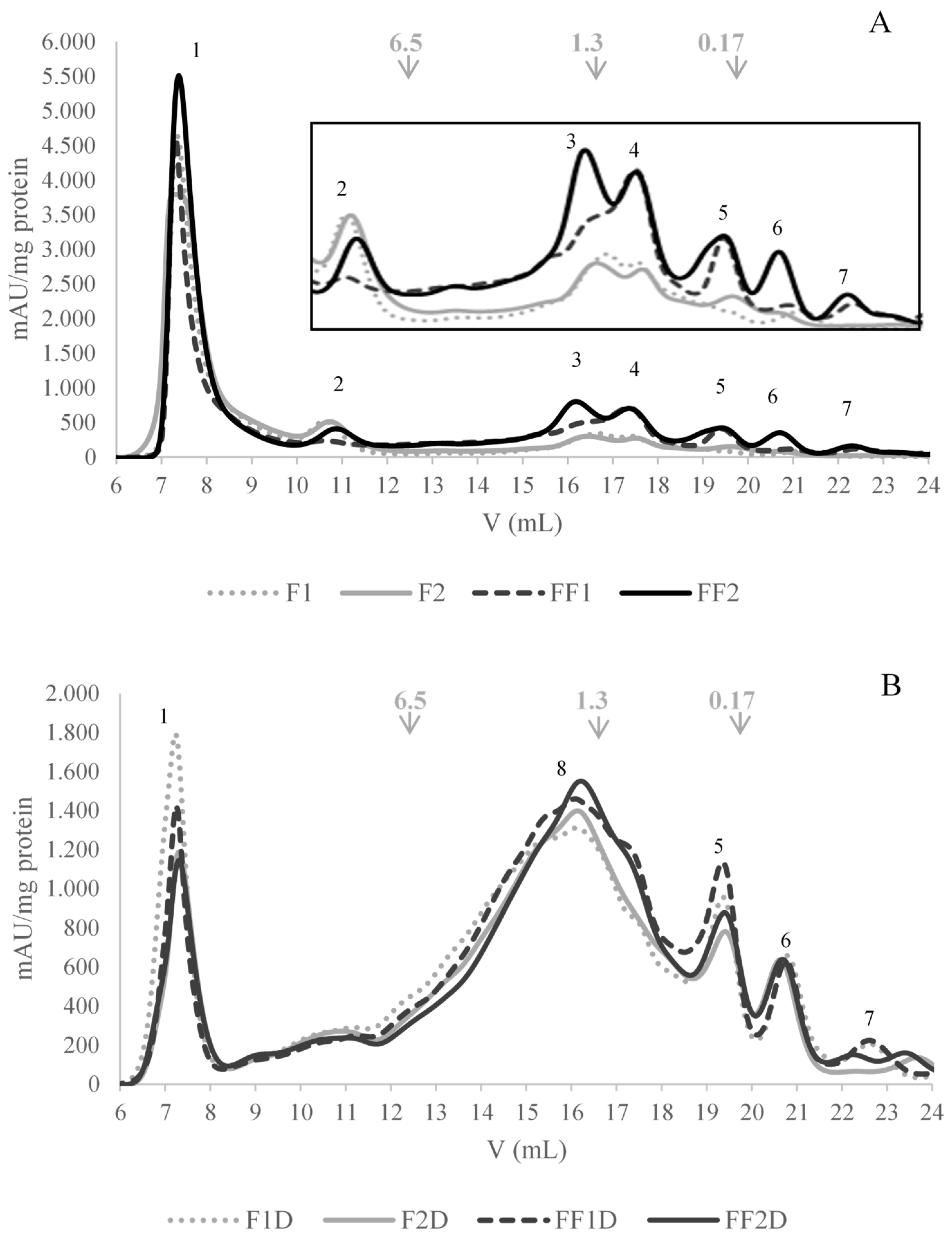
The peptide/polypeptide composition of the soluble fractions of non-fermented and fermented flours was analyzed by gel filtration chromatography using a Superdex 30 column (optimal separation in the range for MW < 10 kDa) in order to evaluate low MW peptides. As expected, the chromatograms of F1 and F2 (Figure 2A) were similar. Fermentation caused an increase in molecules smaller than 6.5 kDa in both conditions (FF1 and FF2). However, some differences between FF1 and FF2 could be described: peak 2 (MW > 6.5 kDa) decreased more in the case of FF1, while peaks 1 (MW > 10 kDa), 3 (1.5-0.8 kDa), 5 (0.47-0.18 kDa) and 6 (0.18-0.08 kDa) increased more in the case of FF2 (respect to the non-fermented flour), showing a greater occurrence of small molecules in FF2.
Figure 2.
Gel filtration (FPLC) chromatograms (Superdex 30 column, optimal separation range < 10 kDa) of PBS-soluble fractions. (A) F1: 36.4 % w/v F dispersion; F2: 14.3 % w/v F dispersion; FF1: fermented F in condition 1 (36.4 % w/v F dispersion, 24 h, 30°C); FF2: fermented F in condition 2 (36.4 % w/v F dispersion, 24 h, 37°C). (B) F1D: F1 after SGDI; F2D: F2 after SGDI; FF1D: FF1 after SGDI; FF2D: FF2 after SGDI. Molecular weight markers are shown in the top of chromatograms.
Figure 2.
Gel filtration (FPLC) chromatograms (Superdex 30 column, optimal separation range < 10 kDa) of PBS-soluble fractions. (A) F1: 36.4 % w/v F dispersion; F2: 14.3 % w/v F dispersion; FF1: fermented F in condition 1 (36.4 % w/v F dispersion, 24 h, 30°C); FF2: fermented F in condition 2 (36.4 % w/v F dispersion, 24 h, 37°C). (B) F1D: F1 after SGDI; F2D: F2 after SGDI; FF1D: FF1 after SGDI; FF2D: FF2 after SGDI. Molecular weight markers are shown in the top of chromatograms.
According to the electrophoresis and FPLC analysis, the fermentation produced some minor changes in the protein profile of the pea flour related with the appearance of aggregates and soluble proteolytic fragments with MW < 2 kDa, with some differences between the two fermentation conditions assayed. These results together with the registered proteolysis degree showed that the LAB strains present in the fermented flours produced a moderate proteolysis of the pea proteins.
3.5. Effect of Fermentation on Protein Fraction Bioaccesibility (SGID) and Antioxidant Activity
After SGID, as expected, HD significantly increased (p < 0.05) for all samples (Table 4). However, the values were significantly greater (p < 0.05) when F was previously fermented (FF1D and FF2D respect to F1D and F2D, respectively), indicating that the fermentation process improved the protein gastrointestinal digestion. FF1D presented a significantly greater (p < 0.05) HD value than FF2D. However, the HD value of FF1D was 7.5 times greater than those of F1, while the HD value of FF2D was 13 times greater than those of F2 showing a greater proportion of proteolysis in the second case (Table 4). SDS-PAGE (Figure 1) showed that in the samples subjected to SGID most of the polypeptides disappeared, appearing some bands such as 18 (51 kDa), 19 (43 kDa) and 20 (a broad band about 35 kDa), and partially remaining bands of MW < 25 kDa, legumins subunits and albumins) for all the digests (F1D, F2D, FF1D and FF2D). In this way, some pea polypeptides resisted the gastrointestinal digestion. This fact has been previously observed when the gastrointestinal digests of flours and protein isolates from two pea varieties were analyzed [1]. Ma et al. [38] reported that a pea protein hydrolysate obtained by the action of a mixture of trypsin, chymotrypsin and peptidase presented a reduction of most of the bands present in the raw pea profile but with persistence of bands with MW between 10 and 30 kDa. Some differences could be detected among digests, mainly in the molecules generated by SGID. The intensity of the bands 18 and 20 was greater for digests from non-fermented flour (F1D and F2D) while the intensity of band 19 was greater for digests from fermented flour (FF1D and FF2D) (Figure 1). Analyzing the effect of fermentation on the subsequent SGID, the electrophoretic profiles showed a lower intensity in some of the remaining bands in the fermented meals, in agreement with the highest HD values obtained for digests of fermented flours. The partial proteolysis due to the fermentation process made the sequences more susceptible to further degradation by the digestive enzymes, as has been previously reported [40].
Analyzing the composition of the PBS soluble fractions of gastrointestinal digests, gel filtration chromatograms (Figure 2B) showed that the peaks corresponding to the exclusion volume (> 10 kDa) decreased with respect to the undigested samples, and significantly increased the amount of molecules smaller than 6.5 kDa in all digested samples. Similar behavior has been previously reported for flours and protein isolates of two pea varieties and their corresponding digested samples [1]. Considering each particular peak, only minor differences in the area were observed among the four digests. The peak 8 (0.4 to 8 kDa) constituted the greatest modification after gastrointestinal digestion and presented the highest area in the four digests, representing about the 60-63% of the total area. Peak 1 (the remaining MW > 10 kDa molecules) accounted for around 30-33% of the area, presenting F1D the highest value and FF1D the lowest one. Peak 5 (0.47-0.18 kDa) represented about 2.5 and 4 %, and peak 6 (< 0.18 kDa) between 2.5 and 3 %.
The antioxidant activity of PBS-soluble fractions of non-fermented and fermented pea flour, before and after SGID, was evaluated. ORAC assay method measures the scavenging capacity against peroxyl radicals (generated from AAPH at 37 °C) by the oxidative degradation of the fluorescein [41]. Dose-response curves for ORAC (ROO· scavenging % versus peptide concentration) were obtained and IC50 values were calculated (Table 4). The ORAC activity was significantly (p < 0.05) increased by the fermentation process, with a diminution of the IC50 values of 2.5 times for FF1 respect to F1, and 2.7 times for FF2 respect to F2, with a significantly (p < 0.05) lower IC50 value for FF2 (Table 4). Also, HORAC assay was performed, in which the oxidative degradation of fluorescein occurs by hydroxyl radicals generated by the Fenton reaction [42]. Dose-response curves presented a linear fitting in this case. There was no significant difference (p > 0.05) between the IC50 values of non-fermented and fermented flours in any fermentation condition (Table 4), indicating that fermentation had not effect on this activity.
The SGID produced a significant increase (p < 0.05) of the ORAC activity, both in case of F1D and FF1D being that increase of about 4 and 9 times with respect to F1, respectively. F2D and FF2D also presented a significant increase (p < 0.05) in ORAC activity with respect to the initial sample (F2) with a potency increment of 5 times. FF2D presented an IC50 value that was slightly (but significantly) lower than FF1D (Table 4). SGID process produced an increase in antioxidant HORAC potency since the IC50 values were reduced by half, without significant difference between the different digests (p > 0.05). Based on these results, we can conclude that the natural fermentation of pea flour produced an increase in the ORAC activity associated in principle with the release of peptides, but had no noticeable effect on the HORAC activity. The difference in the sensitivity and in the mechanisms of action related to these both methods could explain the differences in the behavior of fermented flours.
Taking into account the previous results and some practical considerations related to the ease of agitation and dispersion, it was decided to continue studying the flour fermented in condition 2 (14.3 % w/w, 24 h, and 37 °C). In order to learn more about the distribution of molecules that contribute to the antioxidant activity of these samples, fractions of different MW were separated by FPLC gel filtration from F2, FF2, F2D, and FF2D, to which their peptide concentration and ROO· scavenging activity were determined using the ORAC test (Figure 3). In F2, as expected, the fractions with the greatest polypeptide concentration were those with MW > 10 kDa (fractions 1 to 9). These fractions presented ROO· scavenging activity (40-60 %); however, the fractions 23-26 (MW between 0.29 and 0.59 kDa) presented the highest activities (66 to 81 %, Figure 3A), but low or non-detectable concentration of peptides (Figure 3B). According to the MW of these fractions, they could involve peptides between 3 and 5 amino acids, although the presence of other components such as phenolic compounds cannot be ruled out, all of which would present significant ORAC activity.
Figure 3.
FPLC fractions separated from F1 (36.4 % w/v F dispersion), F2 (14.3 % w/v F dispersion), FF1 fermented F in condition 1 (36.4 % w/v F dispersion, 24 h, 30 °C); FF2 (fermentation in condition 2 (36.4 % w/v F dispersion, 24 h, 37 °C); F1D: F1 after SGDI; F2D: F2 after SGDI; FF1D: FF1 after SGDI; FF2D: FF2 after SGDI. A: Protein concentration (Lowry method). B: % ROO˙ scavenging (ORAC method).
Figure 3.
FPLC fractions separated from F1 (36.4 % w/v F dispersion), F2 (14.3 % w/v F dispersion), FF1 fermented F in condition 1 (36.4 % w/v F dispersion, 24 h, 30 °C); FF2 (fermentation in condition 2 (36.4 % w/v F dispersion, 24 h, 37 °C); F1D: F1 after SGDI; F2D: F2 after SGDI; FF1D: FF1 after SGDI; FF2D: FF2 after SGDI. A: Protein concentration (Lowry method). B: % ROO˙ scavenging (ORAC method).
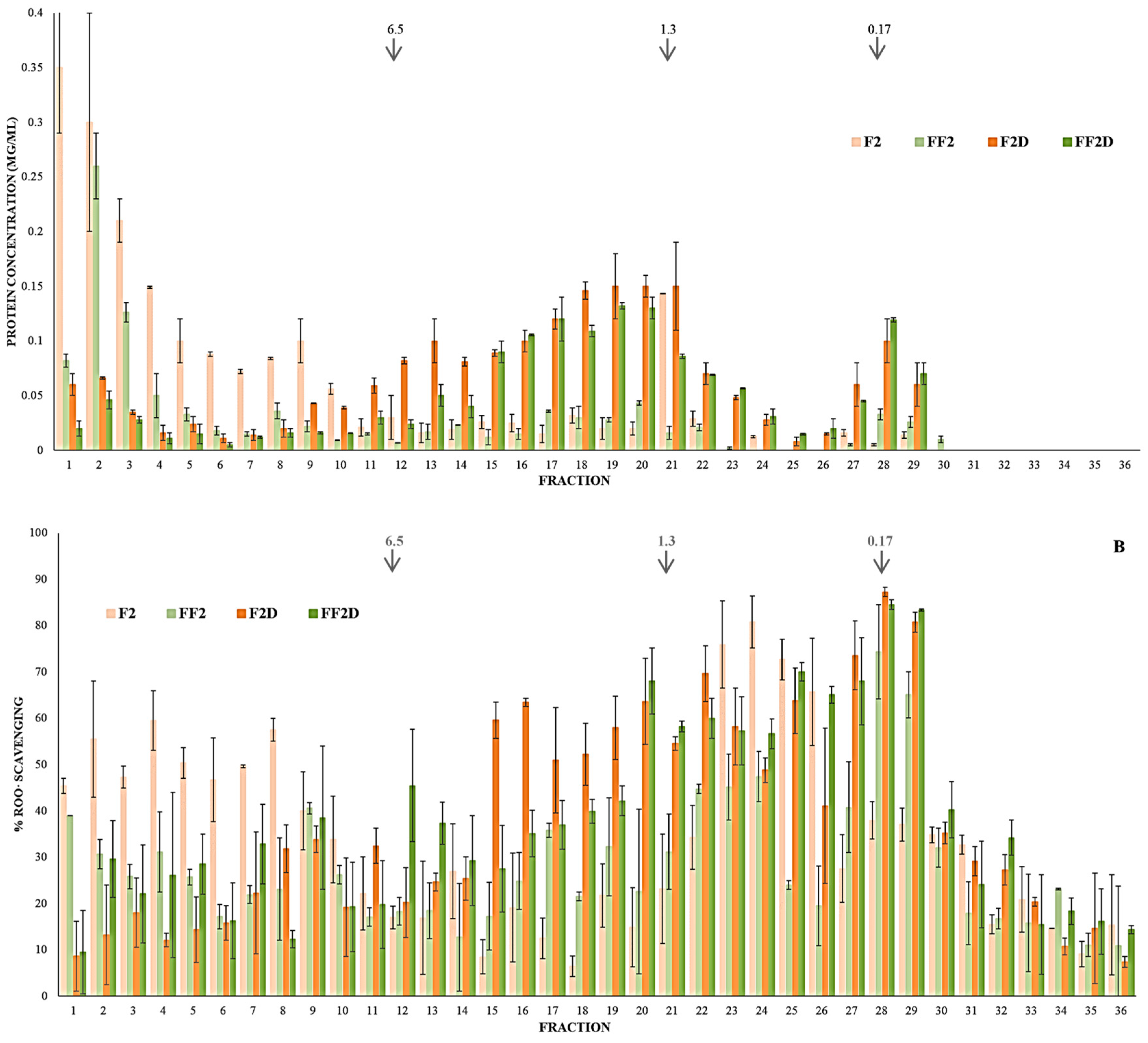
After fermentation (FF2), fractions 1 to 8 (>10 kDa) decreased their polypeptide concentration (Figure 3A) and also their ORAC activity (Figure 3B). Also, fractions 23 to 26 (0.29-0.59 kDa) diminished their ROO· scavenging activity, while several fractions in the range of 0.75 to 4 kDa (fractions 15-22) and 0.18-0.23 kDa (fractions 28-29, 65-74 %) increased it (Figure 3B). In this way, the increment of the ORAC activity registered after fermentation of pea flour could be mainly related to the appearance of molecules in the range of 0.75-4 kDa and 0.18-0.3 kDa with improved ROO· scavenging. Most of the studies involving the formation of bioactive peptides by fermentation were carried out by LAB which possesses a complex system of proteases and peptidases [43]. As reported by Venegas-Ortega et al. [44], the differences found within LAB proteinases explain the variety of bioactive peptides produced, even when the same protein matrix is used. In a previous work [45], nine Lactobacillus strains were evaluated for their ability to grow in pea seed protein-based medium, and to hydrolyze purified pea proteins to produce peptides with antioxidant activity. Two strains, Lacticaseibacillus rhamnosus BGT10 and Lacticaseibacillus zeae LMG17315, exhibited strong proteolytic activity against pea proteins. These authors showed that the antioxidant activity (DPPH assay) of the fraction with MW < 10 kDa increases after 12 h of fermentation with Lacticaseibacillus rhamnosus BGT10. This fraction presented antioxidant activity by different assays and when performing a separation by ion exchange chromatography, they showed that a low abundance sub-fraction of basic peptides presented the highest activity.
The SGID process (F2D and FF2D) produced an increase in the peptide concentration of all fractions with MW < 3 kDa (Figure 3A), and an increment of the ROO· scavenging % for almost all fractions with MW < 6.5 kDa (Figure 3B). F2D showed the higher scavenging % values (41–87 %) for fractions between 0.14 and 4 kDa (fractions 15-29). FF2D presented higher scavenging values with respect to F2D in almost all fractions greater than 4 kDa (< 45 % scavenging), some of the fractions in the range 2 to 0.3 kDa (18-26) and less than 0.10 kDa (< 40 % scavenging), and both digests presented their maximum ROO· inhibition in the fractions around 0.14-0.18 kDa (28 and 29, probably free aminoacids), being 84 and 87 % for FF2D and F2D, respectively (Figure 3B). These results also showed some differences in the molecular composition of the gastrointestinal digest of non-fermented and fermented pea flour.
3.6. Effect of the Fermentation on PCs Bioaccesibility (SGID) and Antioxidant Activity
Given the importance that PCs have in antioxidant activity, whether fermentation modified the content of PCs was evaluated on 60 % ethanol extracts of F2 and FF2. Fermentation process significantly (p < 0.05) increased (about 3 times) the TPC measured by the Folin-Ciocalteu method (Table 5).
Table 5.
Total phenolic content (TPC) and antioxidant activity of 60 % ethanol UAE extracts from yellow pea flour (F2), fermented flour (FF2) and the gastrointestinal digests (F2D, FF2D).
Table 5.
Total phenolic content (TPC) and antioxidant activity of 60 % ethanol UAE extracts from yellow pea flour (F2), fermented flour (FF2) and the gastrointestinal digests (F2D, FF2D).
| Sample | TPC | ORAC | ABTS |
|---|---|---|---|
| (µg GAE/mL) | IC50 (µg GAE/mL) | IC50 (µg GAE/mL) | |
| F2 | 33 ± 1a | 1.2 ± 0.1b | 23 ± 2a |
| FF2 | 96 ± 2b | 1.4 ± 0.3b | 48 ± 10b |
| F2D | 181 ± 4c | 0.8 ± 0.1a | 29 ± 4a |
| FF2D | 193 ± 4d | 0.8 ± 0.1a | 22 ± 3a |
Condition 2: 14.3 % w/w F, 24 h, 37ºC. UAE: ultrasound assisted extraction (15 min, 40 % amplitude). GAE: gallic acid equivalent. Different letters in the same column indicates significant differences (p < 0.05).
Gan et al. [46] reported that natural fermentation increased TPC in most legumes, especially in the mottled cowpea, where it increased by about 80 %. Xiao et al. [47] performed extractions with different solvents (80 % methanol, 80 % ethanol, 80 % acetone, and water) of fermented mung bean and in all of them, the TPC increased with respect to the non-fermented samples. These authors suggested that the chemical structures, polarities, and solubilities of the mung bean PCs were significantly influenced by the fermentation process.
The PCs profile of FF2 was analysed by HPLC-DAD-FLD and compared to those of F2 (Table 6).
Table 6.
Phenolic compound profile of ethanolic UAE extracts from yellow pea flour (F), fermented flour (FF2) and the gastrointestinal digests (F2D, FF2D).
Table 6.
Phenolic compound profile of ethanolic UAE extracts from yellow pea flour (F), fermented flour (FF2) and the gastrointestinal digests (F2D, FF2D).
| Compound | F | FF2 | FD | FF2D |
|---|---|---|---|---|
| OH-tyrosol | 1.7 ± 0.1a | nd | 13.6 ± 0.1c | 7.2 ± 0.1b |
| Phenolic acids | ||||
| Ellagic acid | 0.28 ± 0.02b | 0.44 ± 0.01c | 0.22 ± 0.01a | 0.24 ± 0.01ab |
| Gallic acid | 0.78 ± 0a | nd | nd | 0.82 ± 0b |
| Syringic acid | nd | nd | nd | 4.13 ± 0.03 |
| Caffeic acid | 2.1 ± 0.5ab | 8.9 ± 0.5c | 0.7 ± 0.2a | 2.9 ± 0.6b |
| p-coumaric acid | 1.53 ± 0.01d | 0.22 ± 0a | 1.11 ± 0.04c | 0.46 ± 0.01b |
| Ferulic acid | 0.45 ± 0.09b | 0.19 ± 0.01a | 0.80 ± 0.03c | 0.46 ± 0.02b |
| Rosmarinic acid | 5.2 ± 0.4b | 6.4 ± 0.1c | 3.30 ± 0.02a | 4.69 ± 0.03b |
| Total phenolic acids | 10±1 | 16.1 ± 0.7 | 6.1±0.2 | 13.7 ± 0.7 |
| Stilbenes | ||||
| Polydatin | 26.05 ± 0.04c | 25.61 ± 0.01c | 23.2 ± 0.3b | 22.44 ± 0a |
| trans-resveratrol | 2.6 ± 0.1a | 4.8 ± 0.1b | 7.5 ± 0.1d | 6.67 ± 0c |
| Total stilbenes | 28.6±0.1 | 30.39 ± 0.09 | 30.7±0.3 | 29.11±0 |
| Flavonoids | ||||
| Rutin | 5.2 ± 0.4a | 13.2 ± 0.7b | nd | nd |
| Quercetin-3-glucoside | 0.88 ± 0.01a | 1.59 ± 0.01b | nd | 0.96 ± 0.04a |
| Kaempferol-3-glucoside | 2.3 ± 0.3b | 6.5 ± 0.5c | 0.8 ± 0.1a | 1.1 ± 0.1ab |
| Quercetin | nd | 3.01 ± 0.01 | nd | nd |
| Procyanidin B1 | 13 ± 6a | 21 ± 8a | nd | nd |
| (+)-catechin | 1.05 ± 0.04a | 1.25 ± 0.04b | nd | nd |
| (-)-epigallocatechin | 59.7 ± 0,2 | 82 ± 6 | 27 ± 4 | 0.06 ± 0.08 |
| (-)-epicatechin | 0.55 ± 0.02 | nd | 27 ± 5c | nd |
| (-)-gallocatechin gallate | nd | nd | 6.3 ± 0.2 | nd |
| Naringenin | 0.32 ± 0.02 | nd | nd | nd |
| Hesperetin | 0.71 ± 0.08a | 1.65 ± 0.07b | nd | 1.71 ± 0.02b |
| Total flavonoids | 84±6 | 140±12 | 40±5 | 3.86±0.05 |
| Total | 125 ± 6 | 187 ± 12 | 90 ± 5 | 47 ± 1 |
Contents are expresed as µg/g d.m. In the case of FD and FF2D, content are referred to the original F. dm: dry matter. nd: not detected. Condition 2: 14.3 % w/w F, 24 h, 37ºC. Different letters in the same column indicates significant differences (p < 0.05).
The PCs composition of yellow pea flour has been previously studied in our lab (Cipollone et al., under revision), with (-)-epigallocatechin (a flavan-3-ol) and polydatin (stilbene) as major PCs. Several changes were observed after fermentation. Increments in ellagic, rosmarinic and specially caffeic acids, but diminution in gallic (an hydroxibenzoid acid), p-coumaric, and ferulic acids (hydroxicinamic acids) were observed. OH-tyrosol was not detectable after fermentation. Among the flavonoids (majority in F), only (-)-epicatechin and naringenin (flavanone) decreased, while the flavanone hesperitin, the flavan-3-ols (-)-epigallocatechin and (+)-catechin, and the flavonols rutin (quercetin-3-O-rutinoside), quercetin-3-glucoside, and kaempferol-3-glucoside and quercetin-3-glucoside increased; inclusive quercetin that was not found in F, appeared in FF2 (Table 6). The total amount of HPLC-DAD-FLD detected PCs increased after fermentation, mainly due to a flavonoids family increment. Dueñas et al. [48] carried out spontaneous and with Lactobacillus plantarum ATCC 14917 fermentations of cowpea flour (48 h, 37°C); both fermentations modified the content of PCs in different manner. They found -as in our case- that the fermentation gave rise to the appearance of some PCs compounds not detected in raw flour such as quercetin. That was explained due to the pH lowering could activate some enzymes that hydrolyse the quercetin glycosides, thus yielding quercetin. Lactobacillaceae possess a broad spectrum of enzymatic activities for biotransformation of dietary PCs that could have participated in the change of the PC profile previously described. Esterases, reductases, and decarboxylases would participate in the conversion of hydroxycinnamic- and hydoxybenzoic acids. In addition, LAB contain glycosyl hydrolases that seems to be dedicated to the hydrolysis of glycosides of plant secondary metabolites such as glycosylated flavonoids, although little is known about the substrate specificity of these enzymes [49].
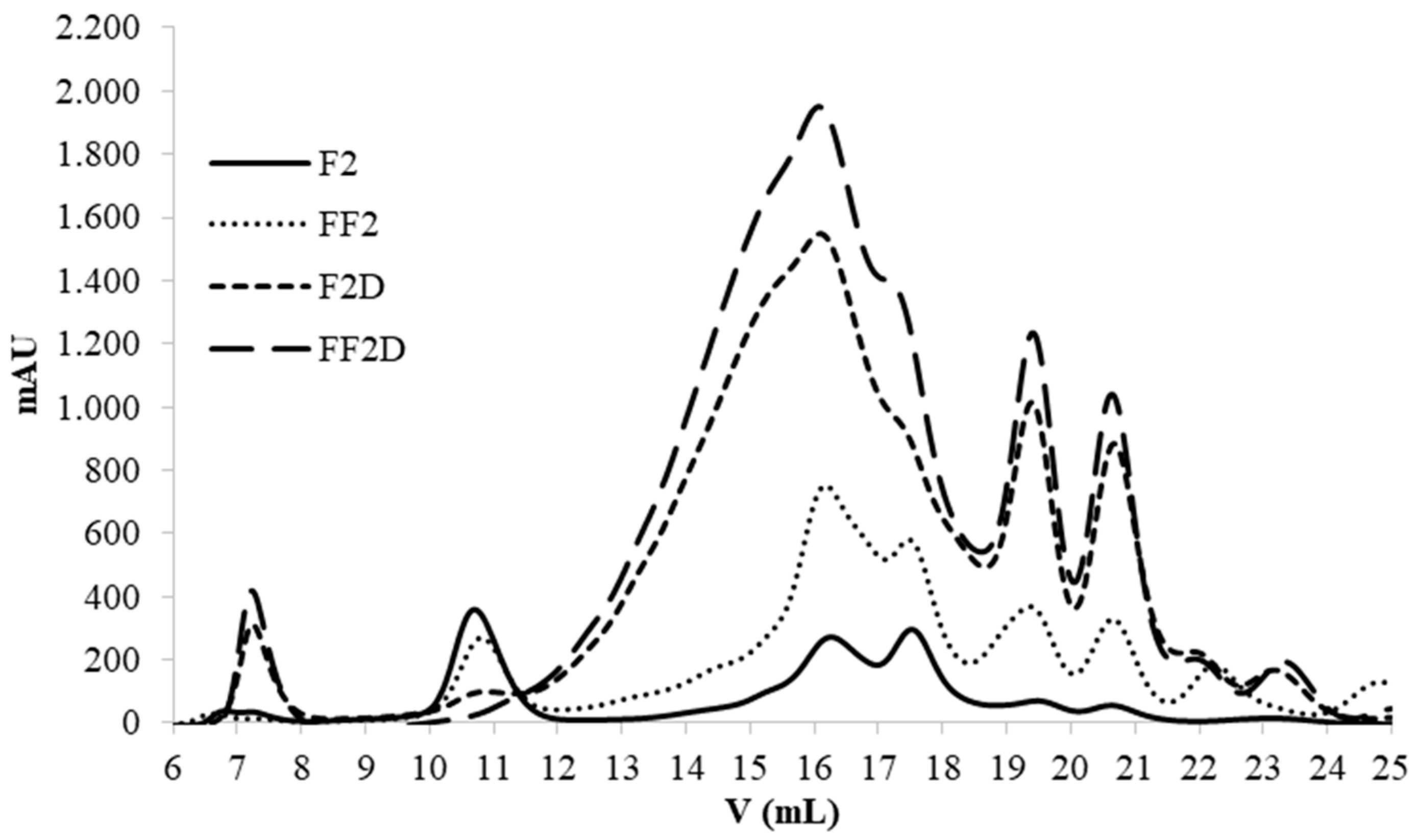
Analyzing the antioxidant activity of the ethanolic extracts, it was observed that the fermentation ABTS activity significantly decreased (p < 0.05) after fermentation (Table 5). In addition, fermentation did not have effect on ROO· scavenging since FF2 presented a similar (p > 0.05) IC50 value for ORAC than F2 (Table 5). This behavior was different from that recorded for the fractions soluble in PBS in which ORAC activity increased after fermentation (Table 4). Gel filtration FPLC chromatograms (Figure 4) of the ethanol extracts showed that both F2 and FF2 presented molecules with MW in a broad range (< 10 kDa), but they did not contain the larger polypeptides (> 10 kDa) that appear in the peak corresponding to the exclusion volume (unlike the fractions soluble in PBS, Figure 2). As in the PBS-soluble fractions, the increment of molecules lower than 2 kDa was evident after fermentation (Figure 4).
Figure 4.
Gel filtration (FPLC) chromatograms (Superdex 30 column, optimal separation range < 10 kDa) of 60 % ethanol extracts from F2: 14.3 % w/v F dispersion; FF2: fermented F in condition 2 (36.4 % w/v F dispersion, 24 h, 37°C); F2D: F2 after SGDI; FF2D: FF2 after SGDI. Molecular weight markers are shown in the top of chromatograms.
Figure 4.
Gel filtration (FPLC) chromatograms (Superdex 30 column, optimal separation range < 10 kDa) of 60 % ethanol extracts from F2: 14.3 % w/v F dispersion; FF2: fermented F in condition 2 (36.4 % w/v F dispersion, 24 h, 37°C); F2D: F2 after SGDI; FF2D: FF2 after SGDI. Molecular weight markers are shown in the top of chromatograms.
After SGID, an increase in the TPC was observed in F2D and FF2D with respect to their non-digested samples, being greater when the flour had been previously fermented (Table 5). According to this, Ketnawa and Ogawa [50] reported an increase in TPC values after subjecting fermented soybeans to a SGID process. SGID produced several changes in the PCs profile of F (Cipollone et al., under revision). The gastrointestinal digest of fermented flour (FF2D) presented a higher content of some phenolic acids than F2D, such as gallic, syringic, caffeic and rosmarinic acids (Table 6). However, the greatest difference was found in flavonoids, whose content was much lower in FF2D since compounds from the flavan-3-ol family (catechins and procyanidin) were not found. These results suggest that, after fermentation, these compounds were more available for the modifications that can occur during the gastrointestinal digestion process, such as instability of catechins at neutral pH [51] and of procyanidin at gastric acidic pH [52]. The SGID produced a significant decrease (p < 0.05) in the IC50 values of both digests, without significant differences between them (Table 5). It also led to an increase in ABTS activity in the case of FF2, with both digests showing similar IC50 values. Thus, although fermentation produced modifications in the PCs profile of F, these did not translate into important changes in ORAC and ABTS activities after SGID. Sancho et al. [53] measured the antioxidant activity in methanol extracts of raw red and black beans before and after digestion and reported that there was no significant difference in the ABTS values and there was only a difference in the extract of black beans when measured by the ORAC method.
It is important to note that although the total content of PCs detected by HPLC-DAD-FLD was much lower in the case of FF2D, its TPC determined by Folin was somewhat higher than for F2D. In addition, both digests presented higher TPC but lower HPLC-detected PCs than non-digested samples (Table 5 and Table 6). These facts suggested that other substances reactive to Folin were present in the extracts. To analyze this, gel filtration FPLC of the ethanol extracts was performed. After SGID, the presence of molecules with MW < 6.5 kDa increased strongly, and to a much lesser extent molecules with MW > 10 kDa (Figure 4); the latter presented much lower abundance than in the case of the fractions soluble in PBS (Figure 2). These analysis demonstrated the presence of other kinds of compounds in the ethanol extracts, such as peptides and amino acids, with higher abundance in FF2D. Therefore, the antioxidant activity of these ethanol extracts showed the contribution of both PCs and peptides and free amino acids that could be solubilized in the extraction conditions.
4. Conclusions
A single fermentation step of yellow pea flour in a liquid medium (14.3 % F dispersion) in bioreactor (24 h, 37 °C) allowed a product with a pH of 4.4 and a count of about 9 log CFU/g of LABs, and significant changes in protein and PCs which could modify the nutritional and bio-functional value of this legume. Partial proteolysis and increased protein digestion after SGID were evident after fermentation, which could be potentially associated to a better nutritional quality of the fermented flour. Also, the fermentation produced an increase in extractable PCs, mainly flavonoids, some of which, such as flavan-3-ols, disappeared after SGID, with these digests presenting a higher content of some phenolic acids. The ORAC potency augment observed after fermentation could be mainly related to the appearance of small molecules, mostly peptides with a size lower than 4 kDa; After SGID, fermented flour showed increased ROO· scavenging activity associated with molecules in a broad MW range (> 4 kDa to < 0.10 kDa).
Subsequent studies will be carried out to study in greater depth the microbial populations responsible for the observed changes, as well as the reproducibility and the effect of seed storage conditions. Also, natural fermentation can improve other biological properties in addition to antioxidant, which should be analyzed in order to fully exploit all its benefits. These first results show that natural fermentation could be used as an economical and easy-to-implement tool to achieve a yellow pea ingredient with improved antioxidant properties whose application in food formulation must be evaluated from a technological point of view.
Author Contributions
Conceptualization, V.T.; methodology, M.A.C, A.A., A.F., V.T.; formal analysis, M.A.C, A.A., A.F., V.T; investigation, M.A.C.; resources, A.F, V.T.; writing—original draft preparation, M.A.C, V.T.; writing—review and editing, M.A.C, A.A., A.F., V.T.; visualization, M.A.C; supervision, V.T.; project administration, A.F., V.T.; funding acquisition, A.F., V.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Universidad Nacional de La Plata (UNLP), grant X870, and by Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT), PICT-2020-1367. Authors are members of CONICET (Argentina).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cipollone, M.A.; Tironi, V. Yellow pea flour and protein isolate as potentially antioxidant ingredients. Legume Sci. 2020, 2, 4–e59. [Google Scholar] [CrossRef]
- Vaz Patto, M.; Amarowicz, R.; Aryee, A.; Boye, J.; Chung, H.J.; Martín-Cabrejas, M.; Domoney, C. Achievements and challenges in improving the nutritional quality of food legumes. Crit. Rev. Plant Sci. 2015, 34, 105–143. [Google Scholar] [CrossRef]
- Gobbetti, M.; De Angelis, M.; Di Cagno, R.; Calasso, M.; Archetti, G.; Rizzello, C.G. Novel insights on the functional/nutritional features of the sourdough fermentation. Int. J. Food Microb. 2019, 302, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Adebo, J.A.; Njobeh, P.B.; Gbashi, S.; Oyedeji, A.B.; Ogundele, O.M.; Oyeyinka, S.A.; Adebo, O.A. Fermentation of cereals and legumes: Impact on nutritional constituents and nutrient bioavailability. Fermentation 2022, 8, 63. [Google Scholar] [CrossRef]
- Carnevali, P.; Ciati, R.; Leporati, A.; Paese, M. Liquid sourdough fermentation: Industrial application perspectives. Food Microb. 2007, 24, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Rizzello, C.G.; Lorusso, A.; Montemurro, M.; Gobbetti, M. Use of sourdough made with quinoa (Chenopodium quinoa) flour and autochthonous selected lactic acid bacteria for enhancing the nutritional, textural and sensory features of white bread. Int. J. Food Microb. 2016, 56, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gan, R.Y.; Li, H.B.; Gunaratne, A.; Sui, Z.Q.; Corke, H. Effects of fermented edible seeds and their products on human health: Bioactive components and bioactivities. Compreh. Rev. Food Sci. Food Saf. 2017, 16, 489–531. [Google Scholar] [CrossRef]
- Adebo, O.A.; Njobeh, P.B.; Adebiyi, J.A.; Gbashi, S.; Phoku, J.Z.; Kayitesi, E. Fermented pulse-based food products in developing nations as functional foods and ingredients. In Functional Food - Improve Health through Adequate Food; Hueda, M.C. Ed.; IntechOpen, Rijeka, 2017, chapter 5. [CrossRef]
- Gueguen, J.; Barbot, J. Quantitative and qualitative variability of pea (Pisum sativum L.) protein composition. J. Sci. Food Agric. 1988, 42, 209–224. [Google Scholar] [CrossRef]
- Nkonge, C.; Balance, M. A sensitive colorimetric procedure for nitrogen determination in micro-Kjeldhal digests. J. Agric. Food Chem. 1982, 30, 416–420. [Google Scholar] [CrossRef]
- AOAC (1984). Official Methods of Analysis. Association of Official Analytical Chemists, 24.002 and 24.009.
- AOAC (1990). Official Methods of Analysis of the Association of Official Analytical Chemists. Association of Official Analytical Chemists, 920.39.
- AOAC (1995) Official Methods of Analysis of the Association of Official Analytical Chemists, 991.43.
- García Fillería, S.; Tironi, V. Intracellular antioxidant activity and intestinal absorption of amaranth peptides released using simulated gastrointestinal digestion with Caco-2 TC7 cells. Food Biosci. 2021, 2021 41, 101086. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Balance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; Dufour, C.; Egger, L.; Golding, M.; Karakaya, S.; Kirkhus, B.; Le Feunteun, S.; Lesmes, U.; Macierzanka, A.; Mackie, A.; Marze, S.; McClements, D.J.; Ménard, O.; Recio, I.; Santos, C.N.; Singh, R.P.; Vegarud, G.E.; Wickham, M.S.; Weitschies, W.; Brodkorb, A. A standardised static in vitro digestion method suitable for food - an international consensus. Food Function 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Adler – Nissen, J. Determination of degree of hydrolysis of food proteins hydrolysates by trinitrobencensulfonic acid. J. Agric. Food Chem. 1979, 27, 1256–1262. [Google Scholar] [CrossRef] [PubMed]
- Orsini Delgado, M.C.; Tironi, V.; Añón, M.C. Antioxidant activity of amaranth proteins or their hydrolysates under simulated gastrointestinal digestion. LWT Food Sci Technol 2011, 44, 1752–1760. [Google Scholar] [CrossRef]
- Boye, J.I.; Zare, F.; Pletch, A. Pulse proteins: Processing, characterization, functional properties and applications in food and feed. Food Res. Int. 2010, 43, 414–431. [Google Scholar] [CrossRef]
- Laemmli, U. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Blum, H.; Beier, H.; Gross, H. Improved silver staining of plant proteins, RNA and DNA in polyacrylamide gels. Electrophoresis 1987, 8, 93–99. [Google Scholar] [CrossRef]
- Lowry, O.; Rosebrough, N.; Farr, A.; Randall, R. Protein measurement with the Folin phenol reagent. J. Biolog. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics whit phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Viticult. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Ferreyra, S.; Bottini, R.; Fontana, A. Tandem absorbance and fluorescence detection following liquid chromatography for the profiling of multiclass phenolic compounds in different winemaking products. Food Chem. 2021, 338, 128030. [Google Scholar] [CrossRef]
- Orsini Delgado, M.C.; Galleano, M.; Añón, M.C.; Tironi, V. Amaranth peptides from gastrointestinal digestion: Antioxidant activity against physiological reactive species. Plant Foods Human Nut. 2015, 70, 27–34. [Google Scholar] [CrossRef]
- Tironi, V.A.; Añón, M.C. Amaranth proteins as a source of antioxidant peptides: Effect of proteolysis. Food Res. Int. 2010, 43, 315–322. [Google Scholar] [CrossRef]
- Sáez, G.D.; Hébert, E.M.; Saavedra, L.; Zárate, G. Molecular identification and technological characterization of lactic acid bacteria isolated from fermented kidney beans flours (Phaseolus vulgaris L. and P. coccineus) in northwestern Argentina. Food Res. Int. 2017, 102, 605–615. [Google Scholar] [CrossRef]
- Sáez, G.D.; Saavedra, L.; Hebert, E.M.; Zárate, G. Identification and biotechnological characterization of lactic acid bacteria isolated from chickpea sourdough in northwestern Argentina. LWT Food Sci Technol 2018, 93, 249–256. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Coda, R.; Wang, Y.; Verni, M.; Kajala, I.; Katina, K.; Laitila, A. Characterization of indigenous Pediococcus pentosaceus, Leuconostoc kimchii, Weissella cibaria and Weissella confusa for Faba bean bioprocessing. Int. J. Food Microb. 2019, 302, 24–34. [Google Scholar] [CrossRef]
- Corsetti, A. Technology of sourdough fermentation and sourdough applications. In Handbook on sourdough biotechnology; Gobbetti, M., Gänzle, M., Eds.; Springer: New York, NY, USA, 2013; pp. 85–103. [Google Scholar]
- Boye, J.I.; Zare, F.; Pletch, A. Pulse proteins: Processing, characterization, functional properties and applications in food and feed. Food Res. Int. 2010, 43, 414–431. [Google Scholar] [CrossRef]
- Martín-Cabrejas, M.A.; Ariza, N.; Esteban, R.; Mollá, E.; Waldron, K.; López-Andréu, F.J. Effect of germination on the carbohydrate composition of the dietary fiber of peas (Pisum sativum L.). J. Agric. Food Chem. 2003, 51, 1254–1259. [Google Scholar] [CrossRef] [PubMed]
- Onwurafor, E.U.; Onweluzo, J.C.; Ezeoke, A.M. Effect of fermentation methods on chemical and microbial properties of mung bean (Vigna radiata) flour. Nigerian Food J. 2014, 32, 89–96. [Google Scholar] [CrossRef]
- Curiel, J.A.; Coda, R.; Centomani, I.; Summo, C.; Gobbetti, M.; Rizzello, C.G. Exploitation of the nutritional and functional characteristics of traditional Italian legumes: The potential of sourdough fermentation. Int. J. Food Microb. 2015, 196, 51–61. [Google Scholar] [CrossRef]
- Kaczmarska, K.; Chandra-Hioe, M.; Zabaras, D.; Frank, D.; Arcot, J. Effect of germination and fermentation on carbohydrate composition of Australian sweet lupin and soybean seeds and flours. J. Agric. Food Chem. 2017, 65, 10064–10073. [Google Scholar] [CrossRef]
- Ramakrishna, V.; Rao, P.R. Purification of acidic protease from the cotyledons of germinating Indian bean (Dolichos lablab L. var lignosus) seeds. African J. Biotech. 2005, 4, 703–707. [Google Scholar]
- Gänzle, M.G.; Loponen, J.; Gobbetti, M. Proteolysis in sourdough fermentations: mechanisms and potential for improved bread quality. Trends in Food Sci. Tech. 2008, 19, 513–521. [Google Scholar] [CrossRef]
- Akhtaruzzaman, N.H.M.; Rubel, M.; Ripa, J.; Atikur, R.; Tanjina, R. Isolation and characterization protease enzyme from leguminous seeds. Agric. Sci. Res. J. 2012, 2, 434–440. [Google Scholar]
- Ma, Z.; Boye, J.I.; Hu, X. In vitro digestibility, protein composition and techno-functional properties of Saskatchewan grown yellow field peas (Pisum sativum L.) as affected by processing. Food Res. Int. 2017, 92, 64–78. [Google Scholar] [CrossRef]
- Byanju, B.; Hojilla-Evangelista, M.P.; Lamsal, B.P. Fermentation performance and nutritional assessment of physically processed lentil and green pea flour. J. Sci. Food Agric. 2021, 101, 5792–5806. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Tagliazucchi, D.; Babini, E.; Rutella, G.S.; Taneyo Saa, D.L.; Gianotti, A. Bioactive peptides from vegetable food matrices: research trends and novel biotechnologies for synthesis and recovery. J. Funct. Foods 2016, 27, 549–569. [Google Scholar] [CrossRef]
- Ou, B.; Hampsch-Woodil, M.; Prior, R. Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J. Agric. Food Chem. 2016, 49, 4619–4626. [Google Scholar] [CrossRef] [PubMed]
- Ou, B.; Hampsch-Woodill, M.; Flanagan, J.; Deemer, E.K.; Prior, R.L.; Huang, D. Novel fluorometric assay for hydroxyl radical prevention capacity using fluorescein as the probe. J. Agric. Food Chem. 2002, 50, 2772–2777. [Google Scholar] [CrossRef]
- Verni, M.; Verardo, V.; Rizzello, C.G. How fermentation affects the antioxidant properties of cereals and legumes. Foods 2019, 8, 362. [Google Scholar] [CrossRef]
- Venegas-Ortega, M.G.; Flores-Gallegos, A.C.; Martinez-Hernandez, J.L.; Aguilar, C.N.; Nevarez-Moorillon, G.V. Production of bioactive peptides from lactic acid bacteria: A sustainable approach for healthier foods. Compreh. Rev. Food Sci. Food Saf. 2019, 18, 1039–1051. [Google Scholar] [CrossRef] [PubMed]
- Stanisavljević, N.S.; Vukotić, G.N.; Pastor, F.T.; Sužnjević, D.; Jovanović, Z.S.; Strahinić, I.D.; Fira, D.A.; Radović, S.S. Antioxidant activity of pea protein hydrolysates produced by batch fermentation with lactic acid bacteria. Arch. Biol. Sci. Belgrade 2015, 67, 1033–1042. [Google Scholar] [CrossRef]
- Gan, R.Y.; Shah, N.P.; Wang, M.F.; Lui, W.Y.; Corke, H. Fermentation alters antioxidant capacity and polyphenol distribution in selected edible legumes. International Journal of Food Science & Technology 2016, 51, 875–884. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhang, Q.; Miao, J.; Rui, X.; Li, T.; Dong, M. Antioxidant activity and DNA damage protection of mung beans processed by solid state fermentation with Cordyceps militaris SN-18. Innov. Food Sci. Emerging Tech. 2015, 31, 216–225. [Google Scholar] [CrossRef]
- Dueñas, M.; Fernández, D.; Hernandez, T.; Estrella, I.; Muñoz, R. Bioactive phenolic compounds of cowpeas (Vigna sinensis L). Modifications by fermentation with natural microflora and with Lactobacillus plantarum ATCC 14917. J. Sci. Food and Agric. 2005, 85, 297–304. [Google Scholar] [CrossRef]
- Gaur, G.; Gänzle, M. Conversion of (poly)phenolic compounds in food fermentations by lactic acid bacteria: Novel insights into metabolic pathways and functional metabolites. Current Res. Food Sci. 2023, 6, 100448. [Google Scholar] [CrossRef]
- Ketnawa, S.; Ogawa, Y. Evaluation of protein digestibility of fermented soybeans and changes in biochemical characteristics of digested fractions. J. Funct. Foods 2019, 52, 640–647. [Google Scholar] [CrossRef]
- Oh, J.H.; Lee, C.Y.; Lee, Y.E.; Yoo, S.H.; Chung, J.O.; Rha, C.S.; Park, M.Y.; Hong, Y.D.; Shim, S.M. Profiling of in vitro bioaccessibility and intestinal uptake of flavonoids after consumption of commonly available green tea types. Molecules 2021, 26, 1518. [Google Scholar] [CrossRef] [PubMed]
- Bouayed, J.; Deuber, H.; Hoffmann, L.; Bohn, T. Bioaccessible and dialysable polyphenols in selected apple varieties following in vitro digestion vs. their native patterns. Food Chem. 2012, 131, 4–1466. [Google Scholar] [CrossRef]
- Sancho, R.A.S.; Pavan, V.; Pastore, G.M. Effect of in vitro digestion on bioactive compounds and antioxidant activity of common bean seed coats. Food Res. Int. 2015, 76, 74–78. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



