Submitted:
15 January 2024
Posted:
16 January 2024
You are already at the latest version
Abstract
Demography and behavior of Teinopodagrion oscillans was studied in a protected area from Andean region. Adult damselflies were individually marked, and using their recapture histories we estimated survival, longevity, sex ratio, and population size using Cormack-Jolly-Seber models. Furthermore, aspects of their behaviour were recorded. Survival, recapture, and lifespan were similar for both sexes and all age groups. Mature males were larger and the distance from the water was similar for all individuals. The most supported model was the time-dependent model for survival and recapture (Phi(t) p(t)). This suggests that weather variations affect in a significant way the demography of this population. Individuals exhibited high fidelity to their site perch, perching with open wings near to water on a variety of perch. Mature males make short flights from the perch to intercept conspecific and interspecific males and to hunt prey. Tandem position was formed on macrophytes and then the pair flight away. Oviposition lasted for 11.23 minutes on average, with the females ovipositing by abdomen submersion. Our results offer insights on demographic characteristics and behaviour of this species, providing crucial information for short and long term conservation decisions of the biodiversity in ecosystems of the Andean region.
Keywords:
Damselfly
; dragonfly
; aquatic insects
; mark-recapture
; Neotropics
; Zygoptera
1. Introduction
Research in animal demography has generated a substantial body of literature allowing a good understanding of how populations change over time [1,2,3]. Historically, demographic information has been used in studies focusing on the context of stable population theory [4,5], organisms’ life tables [6], and population biology [7]. However, an emerging sub-discipline of the demography, the bio-demography, suggests going further, using the demographic information to understand and explain the underlying actuarial biological and ecological mechanisms acting on the populations [8]. Current development of approaches based on maximum likelihood models allow estimating demographic parameters using recapture histories of marked individuals, because these methods consider the detection of individuals to estimate sizes and survival rates of the populations [9,10] .
Although mark-recapture and maximum likelihood methods have been widely used in organisms such as damselflies reviewed by [11, 12], research on Neotropical species just is beginning [13,14,15,16,17,18,19,20]. Thus, due to the scarcity of available information about the ecology and biology of Neotropical Odonata species, more studies and field observations are needed to be able to build up a “big picture” of the ecological and biological processes modelling the populations of these insects in tropical habitats [20].
Megapodagrionidae is a pantropical family of Odonata, and includes the genus Teinopodagrion De Marmels, 2001, an endemic genus from the South American Yungas Mountain cloud forest, extending their distribution from Venezuela to northwestern Argentina along the eastern slope of the Andes [21]. To our knowledge, population studies of Teinopodagrion species are inexistent, but are necessary to know and understand population biology and ecology of these tropical species, which are apparently present at very low densities in tropical ecosystems. In this paper, we present the results of a demographic and behavioral study of Teinopodagrion oscillans (Selys) from a population inhabiting in a protected area of the Colombian Andes. The objectives of this research included: i) to estimate population size, sex ratio, survival, and life expectancy, ii) to study the area frequented by the adults, by estimating the distance from the stream in which individuals of different age classes perch, and iii) to describe patterns on coloration and general behavior.
2. Materials and Methods
Study area
The Club Naútico El Muña is a protected area located near Bogotá (Sesquilé, Cundinamarca) at an elevation of 2,600 m at the freshwater reservoir Embalse del Tominé (4°50′9.80” N and 73°55′0.70” W). Although for more than 50 years native flora and fauna species (e.g., Andean forests) have been protected in this area, cosmopolitan species such as Pennisetum clandestinum Hochst. ex Chiov., Eichhornia crassipes (Mart.) Solms, Acacia sp., and Eucalyptus sp. can be found also. This protected area is very important because harbours dragonfly and damselfly species that have disappeared from intervened surrounding areas [22]. The area includes a stone-bed first order stream named Agua Clara, which originates in the Paramo ecosystem. The habitat of T. oscillans is a section of the Agua Clara stream, which is mixed with water of the Tominé dam generating a mesotrophic system of slow current surrounded by the vegetation above mentioned.
Study species
Teinopodagrion oscillans is a medium-sized zygopteran (body length 38.04–45.94 mm; wing length 28-32.99 mm). This species has a bicoloured trapezoidal pterostigma, with sky-blue on the dorsal region and light brown in the ventral region, and is further characterized by long legs, the posterior femur reaching the abdominal segment 3. Postocular spots are also sky-blue. There are age-related colour changes, as indicated in the results section (Table 1, Figure 1a-f). Due to the development of pruinescence, old individuals show blue-greyish on the thorax and the abdominal segments 1-3. The abdomen is brown to black dorsally and light brown on the sides and ventral parts, and shows also temperature-dependent colour changes, being more bluish, greenish and brownish at lower temperatures.
Data Collection
A transect of 500 m parallel to the dam shoreline was demarcated. Every individual observed was caught, marked (at first capture), and its sex, colour (as estimator of age), and recapture data were recorded. Then, it was released at the site of capture within one minute [23,24]. Damselflies were captured with aerial nets and individually marked with a fine-tip Sharpie® black marker on the right forewing using four-digit numbers. A total of 270 h of sampling person effort was used in the study. Fieldwork was done daily between 14 October 2020 and 1 January 2021. However, no individuals were found in the periods 17-19 October and 9-10 November, due to unfavourable weather conditions.
Following well known methods to classify age in Odonata [25] adult individuals in this study were classified into three age categories, i.e., tenerals, those individuals with soft hyaline wings lacking well-defined general body pigmentation; sexually immature, including individuals with entirely flexible wings, or flexible from the node to the tip, with a pale body colour; and sexually mature, those with hard and opaque, inflexible wings, and well defined body colour.
Behavioral study
The observations followed the focal animal temporal sampling technique [26]. Each session took place from 08:00 to 14:00 h (six hours daily) Colombian time (COT, UTC-5), divided into periods of 10 min per individual, in which the sequence and changes in behavior were recorded. Ten minutes later, a different marked animal was observed. A total of 1,800 min were used for collecting observational data into the breeding area. We sorted our observations into the following categories: i) perching; ii) foraging; iii) tandem; iv) mating; v) oviposition. Observations were done with the naked eye or using 10×50 JHOPT™ binoculars (Brighton, CO, USA).
Statistical Analyses
The recapture histories of marked animals were analysed using Cormack-Jolly-Seber models, by means of the software Mark 10.1 [27]. First, we tested the adjustment of the full time-dependent model by sex (model Phi(g*t) p(g*t), where g indicates the group (males or females) and t refers to time (day of sampling)) by means of the program RELEASE. However, this model was not a good starting point, because it did not fit the data (Test 2+ Test 3 by groups, χ2347= 591.8, p<0.001). Therefore, we divided adult males and females into teneral, immature and mature (as described above), and tested the fit of the new model including six groups (three for each sex) and full time-dependence. This model fits the data (Test 2+Test 3 by groups, χ2403= 381.7, p=0.771). In a second step, we estimated the variance inflation parameter (c-hat) by dividing the value of c-hat of the saturated model, by the mean c-hat of 100 bootstrap simulations using Mark. The value obtained (c-hat= 1.0802) was used to correct estimations of standard errors. Models were ranked by their Akaike’s quasi-criterion of Information, corrected for overdispersion (QAICc). Means are presented with the SE and (sample size). Statistical tests were calculated with xlStat 2021.
3. Results
Biometry and population structure
In total, we marked 324 individuals, 167 males and 157 females, and recaptured 276 (85%). Table 2 shows summary information about recaptures, lifespan, and biometry by age class. The proportion of animals that were recaptured was not significantly different by age and sex classes (contingence χ25=9.8, p=0.082). The number of recaptures was on average 5.1±0.20, including animals never resighted, and was not significantly affected by sex and age (ANOVA, F3,320=0.26, p=0.855). Observed lifespan was 14.1±0.59 days, with a maximum of 37 days, and, again, there were no significant differences by sex and age (F3,320=1.20, p=0.311). Body length was significantly different between sexes, males being larger (F1,320= 8.21, p=0.004; Table 2), and also among age classes, mature individuals being larger (F2,320= 4.77, p<0.009; Table 2). However, wing length was not significantly different between sexes and age groups (F3,320=1.27, p=0.285). There was a weak positive correlation between body length and wing length (r=0.13, p=0.017), and no significant correlation of neither variable with date of marking (correlation Body Length-Date, r= -0.07, p= 0.193; Wing-Date, r= 0.07, p= 0.198). Finally, the distance from the water was similar for all groups of individuals, animals being found at 2.7±0.04 m from the stream (F3,320=0.39, p=0.758).
Considering the recapture histories of animals marked as teneral and immature, thanks to the high recapture rate of this population, it is possible to estimate the duration of the juvenile phases, i.e., the time elapsed from teneral to immature, and from immature to mature (Table 2). The teneral phase lasted no more than one day, because all individuals marked at this age that were recaptured the day after, were already immature (N=5 males and 4 females). The time elapsed to achieve mature colouration was on average 8.6±0.40 (70) days for males and 9.3±0.40 (75) days for females. This maturation period was similar for all individuals, irrespective of their sex and age at marking (F2,142=0.994, p=0.372).
Over the period of study, the age composition of the population varied markedly (Figure 2). At the start of the fieldwork, most animals seen for the first time (87-92% of males and 55-88% of females in the period 14-16 October) were sexually mature. Many mature animals were found on 20 and 21 October, but then no new mature individuals were found until 26-30 November, when again mature animals were the majority of the new animals found (Figure 2).
Considering also recaptured animals, the number of individuals found each sampling day was on average 22.2±2.79, but with large variation, between 1 and 48 individuals (Figure 2, line Total). In the first period (14-21 November) the population was large (average of 34.5 individuals), but an abrupt diminution occurred after 22 November. The population recovered after 26 November, when many unmarked mature animals appeared. After that date, there was a continuous diminution of the population, which almost disappeared by the end of December.
Survival and recapture rates
Supplementary Table S1 shows the results of fitting standard models to the recapture histories of marked animals. The most supported model is the time-dependent model for survival and recapture, with no effect of age and sex, model Phi(t) p(t). This model has an Akaike’s weight of 1, indicating that the rest of the models have no support, and implies that there is evidence for daily variability of survival and recapture. Examination of the parameter estimates from this model (Figure 3) shows that after an initial period where estimates of recapture were 0.15-0.35, there was an erratic variation of this parameter starting on 20 November. On 22 November only 2 animals were captured, compared to an average of 38 in the previous 15 days (Figure 3). Daily survival rate was about 1 before 20 November, and then also showed an erratic behaviour but was again around 1 for the last days of fieldwork (Figure 3). This daily variation in survival and recapture is likely due to variable weather conditions. The average daily survival rate was 0.946±0.0032, with a daily recapture rate of 0.269±0.0072, estimated from the reduced model, Phi(.) p(.).
Behavior
We compiled the following information based on observations of the behavior of 324 T. oscillans individuals. Between 08:00 and 14:00 h individuals perched with open wings near to water on macrophytes (36% of the observations), grass (7.6% of the observations; ≤10 cm high); garbage (7.6%), branches, stems and leaves of Acacia sp. (41.2%; range 0.5-12 m high; 2.71±0.75m) and even over us (7.6%). Teneral individuals remained perched with their wings closed. After two hours, they begun to slowly open and close their wings. They perched with their body almost straight, in a vertical or horizontal position. Most of the specimens (80%) were found at the end of the stream, near the still water created by the dam. Our observations indicate high fidelity of the individuals to the perch site, remaining 0.5-122 min on the same perch.
While teneral individuals and mature females remained motionless on the perch, mature males made frequent short flights (<5 sec) returning quickly. From two to five individuals may share the same perch in branches of Acacia sp. Individuals were preferentially found perched on the tip of sunny branches at high temperatures (18-25 °C), but searched for sheltered places at lower temperatures (10-17 °C). Mature males were apparently not aggressive, but inspection flights to other damselflies and dragonflies were commonly observed, including approaches to other conspecifics (53.2% of the observations) and to individuals of coexisting species (29.3% of the observations), including Mesamphiagrion laterale (Selys), Sympetrum gilvum (Selys), Rhionaeschna marchali (Rambur), and Rhionaeschna cornigera (Brauer). Other short flights were predation events (17.5% of the observations). On several occasions, when two or more individuals shared a perch, if any of them changed to a nearby branch, one or more might follow.
At the hottest moments, individuals perched with their body perpendicular to the branch. Mature males approached females, and, if they were receptive, the tandem position was formed, mostly on macrophytes near to the water. Only one pair in copula was observed, because pairs in tandem flight away. We observed only three females ovipositing, all of them not accompanied by the male, but in the two first events, the male remained near the oviposition place, although obvious female guarding was not observed. The oviposition lasted for 11.23±0.61 minutes (N=3), with the females ovipositing by abdomen submersion.
4. Discussion
There is little biological information about most genera included in the Megapodagrionidae sensu latu, and Teinopodagrion is not an exception. Our study is the first to compile demographic information for this genus and yielded some unexpected results. First, we were able to recapture 85% of the animals marked, a value rarely achieved with odonates. Second, males and females were very similar in all demographic parameters, which is unusual. Third, the population studied showed two clear peaks of abundance, separated by six weeks. And finally, both recapture and survival rates showed strong daily variation (Figure 3).
Teinopodagrion oscillans shows little sexual dimorphism in body colouration, as is typical of the species of the genus (for instance, see the pictures of Teinopodagrion meridionale De Marmels in [28]. Males are slightly larger than females, but their wing lengths are not different (Table 2), implying that, to maintain flight performance, wing morphology should be sexually dimorphic, given the different weight of male and female damselflies [29]. Body and wing lengths were not correlated with date of marking. In temperate species, a decrease in body size is frequently observed along the season [29,30], due to time constraints and the effect of rising temperatures, and this is a general pattern in aquatic arthropods [31]. However, in tropical environments, where seasonal limitations are scarce, body size is not expected to correlate with date [32,33,34,35].
In a recent review of mark-recapture studies with odonates, Sanmartín-Villar & Cordero-Rivera [12] found that in general males and females differ in their recapture rates, but not in survival, provided that the study period is long enough to produce unbiased estimates. These authors also found that age at marking affected recapture rates, and also that males are recaptured in a higher proportion than females. In our population, however, both sexes and all age groups showed similar recapture rates and observed lifespan (Table 2). Teinopodagrion oscillans individuals show high site fidelity, and concentrate near the stream, being found on average at a distance lower than 3 m from the water. This behaviour explains the high recapture rates achieved during this study, similar to the case of another damselfly living in tropical streams [36].
The average time between marking and last recapture was two weeks and was not significantly different between sexes. In general, observed lifespan is higher in males, and this has been explained by their behaviour, because males usually remain closer to the water than females, and, as a consequence, male recapture rates are higher e. g., [37]). For instance, in Hypolestes trinitatis (Gundlach), an endemic damselfly from Cuba, which has many ecological similarities with T. oscillans, recapture rates were much higher in males [38]. The estimated survival rate of T. oscillans, 0.946, is one of the highest ever estimated for adult odonates, identical to the male survival rate estimated for H. trinitatis [38], and similar to other species with highly territorial behaviour [12]. Some Calopterygids, Polythorids, Lestids and Thaumatoneurids have daily survival rates above 0.96 [39-41], which translate into expected longevities of around 24 days, using the formula of Cook et al. [42]. In our dataset the expected longevity is 18 days and the maximum observed longevity was 37 days, a value that is nevertheless lower than the maximum longevity of Calopteryx Leach, with is about 40 days e. g., [43].
The first nine days of adult life include the sexual maturation period in T. oscillans. This means that the net reproductive life, for those animals surviving until maturation, is, on average, about one week (Table 2). Sexual maturation is needed in all odonates, because females do not have mature eggs when emerge from the exuvia, and males do not have functional sperm. In general, the pre-reproductive period is longer in females than in males reviewed by [44], but in our population this was not the case. In tropical damselflies, the maturation period can be very long, even more than one month in giant damselflies [45], but species with similar body size as T. oscillans have shorter maturation periods. For instance, male Polythore mutata (McLachlan) need 12.6 days to mature [46], and Palaemnema desiderata Selys and Palaemnema paulitoyaca Calvert 7-10 days [47].
The frequency of reproductive events was low, and this could be due to some reproductive events may have not coincided with the time of our study [48] or they took place in locations difficult to access by the observers; e. g., elevated twigs in the vegetation or quiet sites with high diversity of plants [18].
One of the most unexpected results of our study is the large daily variation in the appearance of mature individuals (Figure 2). Apparently, two cohorts matured with a separation of about six weeks between them, also producing a clear difference in population size between the first and the second half of the study (Figure 2). One possible explanation for this is that these two cohorts represent two generations [49,50]. However, the time elapsed seems too short for larval development. Population size estimates for damselflies from tropical streams are scarce. In a study with a duration of two months of a population of H. trinitatis, male and female abundance were estimated to show little daily variation, with slow increases or decreases, but not sudden changes [38]. Similar patterns emerge from a population study of Argia chelata Calvert [51] in Costa Rica, and two populations of Heteragrion cooki Daigle & Tennessen from Ecuador [16]. However, a mark-recapture study of the damselfly Ischnura pumilio (Charpentier) in Spain, also showed the appearance of two population peaks of mature adults, in this case with a separation of 10 days [52], suggesting immigration from another population. However, we do not know of similar ecosystems close to the area of our study. Thus, the probability of migration from other places seems low.
Growth rates, timing of emergence, and flight seasons can show inter-year variation within species in the same habitat [53]. For several Odonata species it is known that the size of last instar larvae is a complex function of temperature, time of emergence, and species identity. Cothran & Thorp [54] for example found that in the same habitat, individuals of the same species were smaller at the warm micro-habitats, while the largest animals emerged from the cooler micro-habitats. Temperature within lentic freshwater habitats can vary broadly among microhabitats throughout the day and along of the time [55]. In our study area, something similar probably happened with temperature due to the variety of aquatic microhabitats provided by variations in water depth, number and type of macrophytes, and turbidity caused by the mixing of reservoir water with that of the stream. Although these aspects were not measured by us, it is plausible that morphological and behavioral responses can be adapted to the varying conditions of the microhabitat where the animals live.
Our study was also unique because daily recapture and survival rates were clearly variable along the period of fieldwork (Figure 3), despite similar sampling effort. Between 14 October and 19 November, we marked a total of 155 individuals, almost half of the total marked in the whole study, and all of them disappeared by 22 November, producing an abrupt change in population size (Figure 3). Odonate insects strongly depend on ambient temperature to carry out their daily activities [56,57]. The protected area Club Náutico El Muña is in an area where temperatures vary between -2 and 25 ºC throughout the day, over the days and when comparing periods of drought and rain [58]. Therefore, it is plausible that the number of individuals may change rapidly between days due to these temperature variations. We observed that individuals of T. oscillans in this area are indiscriminate when selecting their perch, but on very cold days they stayed on high branches (10-12 m high), where the individuals and their numbers were difficult to track. The influence of microhabitats on T. oscillans seems remarkable, because in addition to the low temperatures, the ecosystem was flooded by the water of the reservoir due to the rains in February 2021, strongly reducing the number of microhabitats available for oviposition and possibly the growth of larvae. Since then, only four adult individuals of T. oscillans have been found in this habitat as of December 2023.
Our study provides crucial information for the understanding of basic population aspects of T. oscillans, a species categorized as Least Concern because it has been reported for two protected areas, one in Colombia [59] and the other in Venezuela [60]. However, our findings support the idea that it is important to know how populations are affected by local conditions, to allow evidence-based categorizations and conservation decisions. Because survival and the probability of recapture were time-dependent, environmental temperature is a factor that alters the presence and size of this population, which we will continue to monitor to try to understand the dynamics that in the medium and long term affect the conservation of this species in habitats of the Andean region.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Table S1: Results of model selection for T. oscillans. Survival rate is denoted by Phi and recapture probability by p. The group is sex (g) and time variation is indicated by t. The best model (in bold) includes variation due to time of recapture and survival.
Author Contributions
Conceptualization, F.P-R.; methodology, F.P-R., A.C-R., D.A.P. and A.C.P.; formal analysis, A.C-R.; investigation, F.P-R., A.C-R., D.A.P. and A.C.P.; data curation, F.P-R., A.C-R.; writing—original draft preparation, F.P-R., A.C-R. and D.A.P.; writing—review and editing, F.P-R., A.C-R., D.A.P. and A.C.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data presented in this study are available on request from the corresponding author (odonata17@hotmail.com).
Acknowledgments
Thanks are due to the Club Naútico Muña board; Alfredo Carlo Amore Pardo, Santiago Rojas, and Camilo Andrés López, for collaborate us during our fieldwork. We thank the Research Group on Odonata and other arthropods in Colombia and the Neotropics for their academic support.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- King, R. C. Handbook of Genetics: Volume 4 Vertebrates of Genetic Interest. (Springer US, 1975). [CrossRef]
- Morin, J. G. & Hastings, J. W. Biochemistry of the bioluminescence of colonial hydroids and other coelenterates. J. Cell. Physiol. 1971, 77, 305–312. [Google Scholar] [PubMed]
- Naranjo, S. E. Influence of Two Mass-Marking Techniques on Survival and Flight Behavior of Diabrotica virgifera virgifera (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1990, 83, 1360–1364. [Google Scholar] [CrossRef]
- Davenport, D. & Nicol, J. A. C. Luminescence in Hydromedusae. Proc. R. Soc. Lond. Ser. B 1955, 144, 399–411. [Google Scholar]
- Bartlett, A. C. Isozyme Polymorphism in Populations of the Pink Bollworm12. Ann. Entomol. Soc. Am. 1981, 74, 9–13. [Google Scholar] [CrossRef]
- Fernandes, O. A. , Wright, R. J., Baumgarten, K. H. & Mayo, Z. B. Use of Rubidium to Label Lysiphlebus testaceipes (Hymenoptera: Braconidae), a Parasitoid of Greenbugs (Homoptera: Aphididae), for Dispersal Studies. Environ. Entomol. 1997, 26, 1167–1172. [Google Scholar]
- Kapp, R. O. How to Know Pollen and Spores; W. C. Brown Company, 1969. [Google Scholar]
- Carey, J. R. Insect biodemography. Annu. Rev. Entomol. 2001, 46, 79–110. [Google Scholar] [CrossRef] [PubMed]
- Kéry, M. & Schaub, M. Bayesian Population Analysis Using WinBUGS: A Hierarchical Perspective; Academic Press, 2012. [Google Scholar]
- Lebreton, J.-D. , Burnham, K. P., Clobert, J. & Anderson, D. R. Modeling Survival and Testing Biological Hypotheses Using Marked Animals: A Unified Approach with Case Studies. Ecol. Monogr. 1992, 62, 67–118. [Google Scholar]
- Cordero-Rivera, A. & Stoks, R. Mark–recapture studies and demography. in Dragonflies and Damselflies: Model Organisms for Ecological and Evolutionary Research (ed. Córdoba-Aguilar, A.) 0 (Oxford University Press, 2008). [CrossRef]
- Sanmartín-Villar, I. & Cordero-Rivera, A. Odonata survival: Insights from mark-recapture experiments. in 129–140 (2022). [CrossRef]
- Altamiranda-Saavedra, M. & Ortega, O. Estructura poblacional de Polythore gigantea (Odonata: Polythoridae) en sistemas lóticos con diferentes estados de conservación en Antioquia, Colombia. Rev. Biol. Trop. 2012, 60, 1205–1216. [Google Scholar]
- Palacino Rodríguez, F. , Sánchez Contreras, N. & Córdoba-Aguilar, A. Population structure in dry and rainy seasons in Erythrodiplax umbrata (Linnaeus, 1758) (Odonata: Libellulidae). Odonatologica 2012, 41, 245–249. [Google Scholar]
- Palacino-Rodríguez, F. , Munguía-Steyer, R., Mesa-Blanco, L. V., Contreras-Sánchez, N. & Córdoba-Aguilar, A. Survival is predicted by territorial status but not wing pigmentation in males of a polythorid damselfly, Euthore fasciata (Odonata: Zygoptera: Polythoridae). Int. J. Odonatol. 2016, 19, 183–190. [Google Scholar]
- Rivas-Torres, A. , Sanmartín-Villar, I., Gabela-Flores, M. V. & Cordero-Rivera, A. Demographics and behaviour of Heteragrion cooki, a forest damselfly endemic to Ecuador (Odonata). Int. J. Odonatol. 2017, 20, 123–135. [Google Scholar]
- Rivas-Torres, A. , Sánchez-Guillén, R. A. & Cordero-Rivera, A. Alternative reproductive strategies in black-winged territorial males of Paraphlebia zoe (Odonata, Thaumatoneuridae). PeerJ 2019, 7, e6489. [Google Scholar] [PubMed]
- Palacino-Rodríguez, F. , Altamiranda-Saavedra, M., Andrés Palacino, D. & Carolina Penagos, A. Ecology of Mesamphiagrion laterale (Odonata: Coenagrionidae): abundance, reproduction and interactions with co-occurring species. Int. J. Odonatol. 2020, 23, 165–182. [Google Scholar]
- Palacino–Rodríguez, F. , Palacino, D. A., Munguia-Steyer, R. & Juen, L. Effects of seasonality and environmental change on an Andean damselfly Mesamphiagrion laterale (Odonata: Coenagrionidae). J. Insect Conserv. 2020, 24, 499–511. [Google Scholar]
- García-Monsalve, M. , Altamiranda-Saavedra, M., Palacino Rodríguez, F. & Cordero-Rivera, A. Demographic Traits and Behavior of Hetaerina cruentata (Odonata: Calopterygidae) in Ecosystems of the Andean Region of Colombia. Int. J. Odonatol. 2021, 24, 261–273. [Google Scholar]
- Ellenrieder, N. The larvae of Teinopodagrion decipiens De Marmels, 2001 and T. meridionale De Marmels, 2001 (Zygoptera: Megapodagrionidae). Odonatologica 2006, 35, 281–287. [Google Scholar]
- Palacino-Rodríguez, F. , Altamiranda-Saavedra, M. A., Palacino-Penagos, D. A., Penagos-Arévalo, A. C. & Ríos-Olaya, K. J. Factors influencing predation on Odonata by Argiope trifasciata (Forsskål, 1775). Int. J. Odonatol. 2023, 26, 36–43. [Google Scholar]
- Alvarez, C. , Álvarez Covelli, M. & Palacino Rodríguez, F. Abdomen or wings? Comparing two body places for marking in Mesamphiagrion laterale (Odonata: Coenagrionidae). Odonatologica 2015, 44, 343–348. [Google Scholar]
- Palacino Rodríguez, F. & Sánchez Contreras, N. Does experimental marking of wings influence resighting success in Mesamphiagrion laterale and Erythrodiplax umbrata? (Odonata: Coenagrionidae, Libellulidae). Odonatologica 2014, 43, 237–246. [Google Scholar]
- Plaistow, S. & Siva-Jothy, M. T. Energetic Constraints and Male Mate-Securing Tactics in the Damselfly Calopteryx splendens xanthostoma (Charpentier). Proc. Biol. Sci. 1996, 263, 1233–1239. [Google Scholar]
- Altmann, J. Observational Study of Behavior: Sampling Methods. Behaviour 1974, 49, 227–267. [Google Scholar] [CrossRef] [PubMed]
- White, G. C. & Burnham, K. P. Program MARK: survival estimation from populations of marked animals. Bird Study 1999, 46, S120–S139. [Google Scholar]
- Ellenrieder, N. & Garrison, R. Libélulas de las Yungas (Odonata). Una Guía de Campo para las especies de Argentina / Dragonflies of the Yungas. A Field Guide to the Species from Argentina. (2007).
- Gallesi, M. M. , Mobili, S., Cigognini, R., Hardersen, S. & Sacchi, R. Season matters: differential variation of wing shape between sexes of Calopteryx splendens (Odonata: Calopterygidae). Zoomorphology 2016, 135, 313–322. [Google Scholar]
- Johansson, F. Latitudinal Shifts in Body Size of Enallagma cyathigerum (Odonata). J. Biogeogr. 2003, 30, 29–34. [Google Scholar] [CrossRef]
- Horne, C. R. , Hirst, Andrew. G. & Atkinson, D. Temperature-size responses match latitudinal-size clines in arthropods, revealing critical differences between aquatic and terrestrial species. Ecol. Lett. 2015, 18, 327–335. [Google Scholar] [PubMed]
- Ashton, K. G. & Feldman, C. R. Bergmann’s rule in nonavian reptiles: turtles follow it, lizards and snakes reverse it. Evolution 2003, 57, 1151–1163. [Google Scholar] [PubMed]
- Phillimore, A. B. , Hadfield, J. D., Jones, O. R. & Smithers, R. J. Differences in spawning date between populations of common frog reveal local adaptation. Proc. Natl. Acad. Sci. U. S. A. 2010, 107, 8292–8297. [Google Scholar] [PubMed]
- Muñoz, M. M. , Wegener, J. E. & Algar, A. C. Untangling intra-and interspecific effects on body size clines reveals divergent processes structuring convergent patterns in Anolis lizards. 2014. [Google Scholar] [CrossRef]
- Boaratti, A. Z. & Da Silva, F. R. Relationships between environmental gradients and geographic variation in the intraspecific body size of three species of frogs (Anura). Austral Ecol. 2015, 40, 869–876. [Google Scholar]
- Cordero Rivera, A. & Zhang, H. Ethological uniqueness of a damselfly with no near relatives: the relevance of behaviour as part of biodiversity. Anim. Biodivers. Conserv. 2018, 41, 161–174. [Google Scholar]
- Stoks, R. Male-biased sex ratios in mature damselfly populations: real or artefact? Ecol. Entomol. 2001, 26, 181–187. [Google Scholar] [CrossRef]
- Torres-Cambas, Y. & Fonseca-Rodríguez, R. Sex ratio, survival, and recapture rate in a Cuban population of the damselfly Hypolestes trinitatis (Odonata: Megapodagrionidae). Acta Ethologica 2011, 14, 69–76. [Google Scholar]
- Cordero-Rivera, A. Estudio ecologico de una poblacion de Lestes viridis Vander Linden, 1825 (Zygoptera, Lestidae). Limnética 1988. [Google Scholar] [CrossRef]
- Munguía-Steyer, R. , Córdoba-Aguilar, A. & Romo-Beltrán, A. Do individuals in better condition survive for longer? Field survival estimates according to male alternative reproductive tactics and sex. J. Evol. Biol. 2010, 23, 175–184. [Google Scholar] [PubMed]
- Cordero–Rivera, A. , Sanmartín–Villar, I., Herrera, M. S., Rivas–Torres, A. & Encalada, A. C. Survival and longevity in neotropical damselflies (Odonata, Polythoridae). (2019).
- Cook, L. M. , Brower, L. P. & Croze, H. J. The Accuracy of a Population Estimation from Multiple Recapture Data. J. Anim. Ecol. 1967, 36, 57–60. [Google Scholar]
- Cordero-Rivera, A. Estructura de tres comunidades de Calopteryx (Odonata: Calopterygidae) con diferente composición específica. Limnética 1989. [Google Scholar] [CrossRef]
- Corbet, P. Biology of Odonata. 2003, 25, 189–217. [Google Scholar] [CrossRef]
- Fincke, O. M. Consequences of Larval Ecology for Territoriality and Reproductive Success of a Neotropical Damselfly. Ecology 1992, 73, 449–462. [Google Scholar] [CrossRef]
- Sanmartín-Villar, I. & Cordero-Rivera, A. Female Colour Polymorphism and Unique Reproductive Behaviour in Polythore Damselflies (Zygoptera: Polythoridae). Neotrop. Entomol. 2016, 45, 658–664. [Google Scholar] [PubMed]
- Garrison, R. & Gonzalez-Soriano, E. Population dynamics of two sibling species of neotropical damselflies, Palaemnema desiderata Selys and P. paulitoyaca Calvert (Odonata: Platystictidae). Folia Entomológica Mex. 1988, 76, 5–24. [Google Scholar]
- Altamiranda-Saavedra, M. , Palacino-Rodríguez, F. & Lobo-Hernández, M. Daily abundance at the breeding site and reproductive behavior of Polythore gigantea (Odonata: Polythoridae). Odonatologica 2015, 43, 69–182. [Google Scholar]
- Corbet, P. , Suhling, F. & Söndgerath, D. Voltinism of Odonata: A review. Int. J. Odonatol. 2005, 9, 1–44. [Google Scholar]
- Mahdjoub, H. , Bouslama, Z., Khelifa, R., Zebsa, R. & Houhamdi, M. Bivoltinism in Coenagrion mercuriale (Zygoptera: Odonata) in the southern margin of its distribution range : emergence pattern and larval growth. Afr. Entomol. 2015, 23, 59–67. [Google Scholar]
- Hamilton, L. D. & Montgomerie, R. D. Population demography and sex ratio in a Neotropical damselfly (Odonata: Coenagrionidae) in Costa Rica. J. Trop. Ecol. 1989, 5, 159–171. [Google Scholar]
- Cordero-Rivera, A. & Abad, J. A. A. Lifetime Mating Success, Survivorship and Synchronized Reproduction in the Damselfly Ischnura pumilio (Odonata: Coenagrionidae). Int. J. Odonatol. 1999, 2, 105–114. [Google Scholar]
- Kormondy, E. J. & Gower, J. L. Life History Variations in An Association of Odonata. Ecology 1965, 46, 882–886. [Google Scholar]
- Cothran, M. L. & Thorp, J. H. Emergence Patterns and Size Variation of Odonata in a Thermal Reservoir. Freshw. Invertebr. Biol. 1982, 1, 30–39. [Google Scholar]
- Ward, J. V. & Stanford, J. A. Thermal Responses in the Evolutionary Ecology of Aquatic Insects. Annu. Rev. Entomol. 1982, 27, 97–117. [Google Scholar]
- Hassall, C. & Thompson, D. J. The effects of environmental warming on Odonata: A review. Int. J. Odonatol. 2008, 11, 131–153. [Google Scholar]
- Hassall, C. Odonata as candidate macroecological barometers for global climate change. Freshw. Sci. 2015, 34, 1040–1049. [Google Scholar] [CrossRef]
- Instituto de Hidrología, Meteorología y Estudios Ambientales - IDEAM. Variabilidad diaria de temperatura. http://www.ideam.gov.co/web/tiempo-y-clima/atlas#_48_INSTANCE_xoDpvO7rhD5O_%3Dhttp%253A%252F%252Fwww.ideam.gov.co%252FAtlasWeb%252Findex.html%253F. (2021).
- Amaya-Vallejo, V. et al. Teinopodagrion oscillans. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7649811/ (2020).
- De Marmels, J. Revision of Megapodagrion Selys, 1886 (Insecta, Odonata: Megapodoagrionidae). ( 2001.
Figure 1.
Teinopodagrion oscillans adult stages; (a) Teneral female; (b) sexually immature male; (c) sexually immature female; (d) sexually immature male; (e) sexually mature male; (d) sexually mature female.
Figure 1.
Teinopodagrion oscillans adult stages; (a) Teneral female; (b) sexually immature male; (c) sexually immature female; (d) sexually immature male; (e) sexually mature male; (d) sexually mature female.

Figure 2.
Daily variation in the number of new individuals found by age class (bars) and the total number found (including recaptures, Total). Note that the number of mature individuals was high at the start of the study and in the period 26-30 November, with a large gap with no mature individuals between these dates and after 30 November. No individuals were found during 17-19 October, and 9-10 November due to bad weather.
Figure 2.
Daily variation in the number of new individuals found by age class (bars) and the total number found (including recaptures, Total). Note that the number of mature individuals was high at the start of the study and in the period 26-30 November, with a large gap with no mature individuals between these dates and after 30 November. No individuals were found during 17-19 October, and 9-10 November due to bad weather.
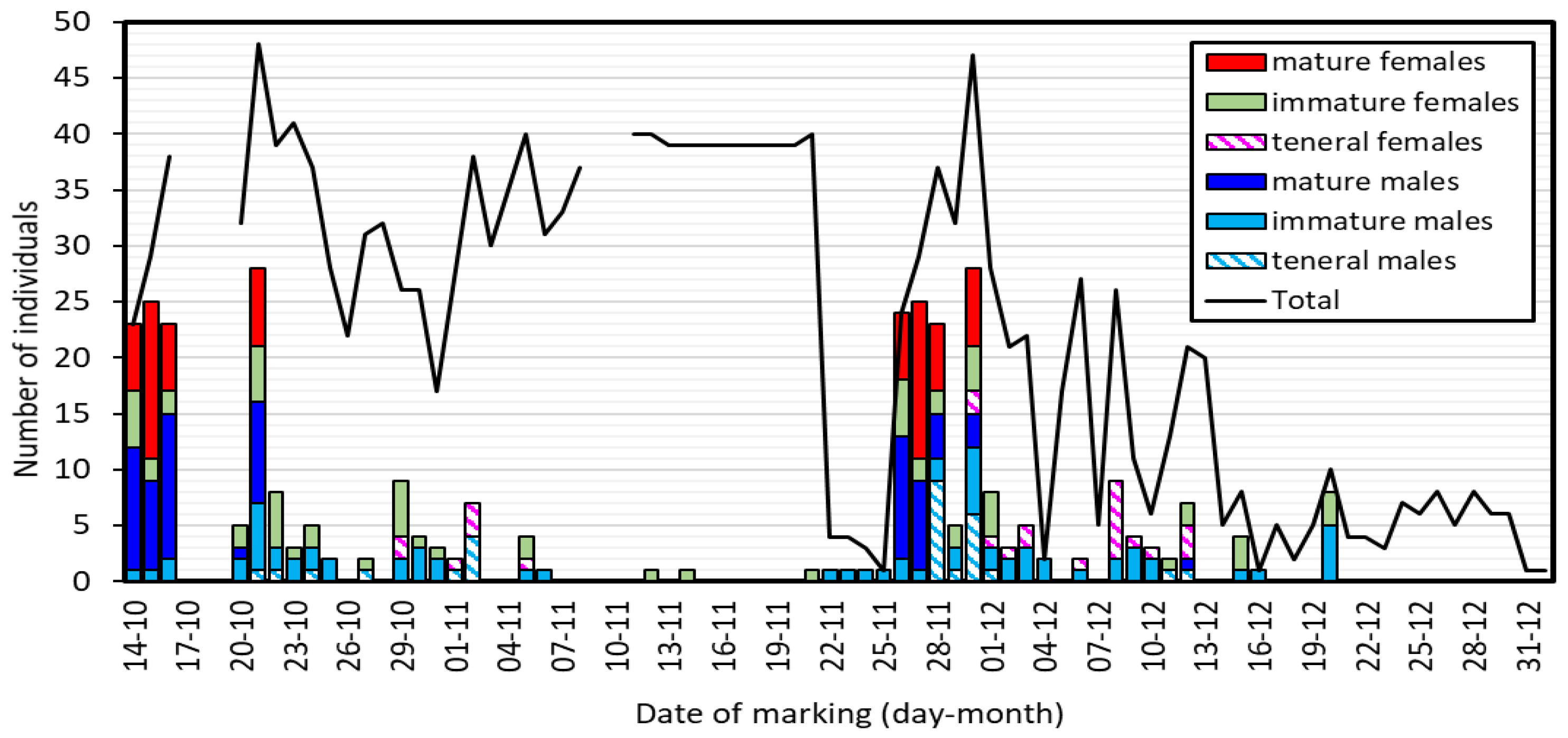
Figure 3.
Estimates of daily survival and recapture rates (±SE) for the period of study, obtained from model Phi(t) p(t). Note the difference between the first period (14 October to 19 November) and the second (20 November to 1 January).
Figure 3.
Estimates of daily survival and recapture rates (±SE) for the period of study, obtained from model Phi(t) p(t). Note the difference between the first period (14 October to 19 November) and the second (20 November to 1 January).
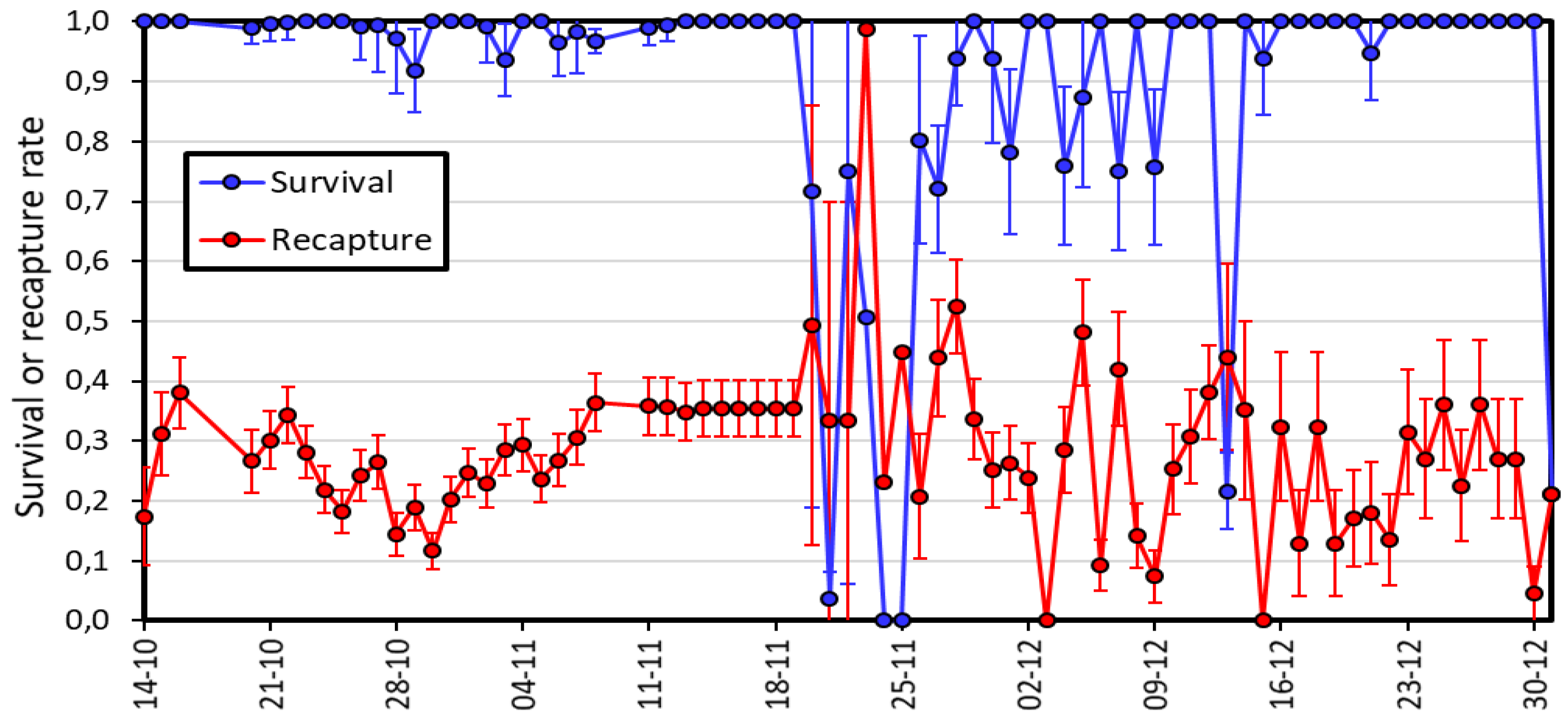

Table 1.
Colour in different body regions of T. oscillans at different ages. PS, Postocular spots; MSP, Mesepisternum; MES, Mesepimeron; MET, Metepisternum; MTP, Metepimeron; S1-3, Abdominal segments 1-3 dorsal and lateral regions; W, Wings; Pt, Pterostigma.
Table 1.
Colour in different body regions of T. oscillans at different ages. PS, Postocular spots; MSP, Mesepisternum; MES, Mesepimeron; MET, Metepisternum; MTP, Metepimeron; S1-3, Abdominal segments 1-3 dorsal and lateral regions; W, Wings; Pt, Pterostigma.
| Age | Teneral | Sexually immature | Sexually mature |
|---|---|---|---|
| PS | Yellow- Light Brown | Light blue | Light blue |
| MSP | 1 longitudinal dark brown stripe and 1 longitudinal yellow stripe | Light blue (80%) and Light brown (20%) | 1 longitudinal black stripe and 1 longitudinal light blue stripe |
| MES | 1 longitudinal brown stripe | Ligth blue (60%) and Light brown (40%) | Light blue (80%) and Light brown (20%) |
| MET | 1 longitudinal yellow stripe | Light blue (90%) and Light brown (10%) | Light blue |
| MTP | Yellow- Light brown | Light blue (80%) and Light brown (20%) | Light blue |
| S1-3 | Yellow- Light brown | Light blue (50%) and Light brown (50%) | Light blue (80%) and Light brown (20%) |
| W | Infumated | Hyaline | Hyaline |
| Pt | Yellow- Ligth brown | Light blue | Light blue |
Table 2.
Summary statistics of the population of Teinopodagrion oscillans marked in this study. Recaptures indicates the number of days in which the individual was found, and Lifespan refers to the number of days between marking and last recapture. Maturation indicates the time in days to achieve mature colouration, only for animals marked as teneral or immature. Distance indicates the distance to the water in m, on the moment of first capture. Body size measurements in mm. Sample size is the number of marked specimens, except indicated otherwise. A, Age; T, Teneral; I, Immature; M, Mature; Mk, Marked individuals; R, Recaptured (%) individuals; SE, standard error of mean; N, sample size. .
Table 2.
Summary statistics of the population of Teinopodagrion oscillans marked in this study. Recaptures indicates the number of days in which the individual was found, and Lifespan refers to the number of days between marking and last recapture. Maturation indicates the time in days to achieve mature colouration, only for animals marked as teneral or immature. Distance indicates the distance to the water in m, on the moment of first capture. Body size measurements in mm. Sample size is the number of marked specimens, except indicated otherwise. A, Age; T, Teneral; I, Immature; M, Mature; Mk, Marked individuals; R, Recaptured (%) individuals; SE, standard error of mean; N, sample size. .
| Recaptures | Lifespan | Maturation | Body length | Wing length | Distance | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sex | A | Mk | R | Mean | SE | Mean | SE | Mean | SE (N) | Mean | SE | Mean | SE | Mean | SE |
| Male | T | 28 | 25 (89.3) | 5.18 | 0.55 | 12.75 | 1.56 | 9.43 | 0.57 (21) | 41.86 | 0.31 | 30.69 | 0.30 | 2.82 | 0.15 |
| I | 70 | 53 (75.7) | 4.64 | 0.41 | 12.87 | 1.28 | 8.16 | 0.47 (49) | 41.50 | 0.23 | 30.51 | 0.16 | 2.64 | 0.08 | |
| M | 69 | 48 (70.0) | 5.70 | 0.48 | 16.12 | 1.38 | - | - | 42.01 | 0.25 | 30.40 | 0.17 | 2.71 | 0.08 | |
| Female | T | 26 | 24 (92.3) | 4.31 | 0.51 | 10.46 | 1.26 | 8.95 | 0.52 (29) | 40.89 | 0.29 | 30.34 | 0.28 | 2.77 | 0.13 |
| I | 65 | 51 (78.5) | 5.80 | 0.46 | 16.08 | 1.29 | 9.22 | 0.48 (55) | 40.79 | 0.23 | 30.14 | 0.16 | 2.69 | 0.10 | |
| M | 66 | 46 (70.0) | 4.77 | 0.43 | 13.39 | 1.43 | - | - | 41.68 | 0.22 | 30.92 | 0.16 | 2.71 | 0.10 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



