
Submitted:
10 January 2024
Posted:
11 January 2024
You are already at the latest version
Abstract
Type IVc Pili (T4cP), also known as Tad or Flp pili, are long thin microbial filaments, which are made-up of small-sized pilins. These appendages serve different functions in bacteria including attachment, biofilm formation, surface sensing, motility, and host colonization. Despite their relevant role in diverse microbial lifestyles, knowledge about T4cP in bacteria that establish symbiosis with legumes, collectively referred to as rhizobia, is still limited. Sinorhizobium meliloti contains two clusters of T4cP-related genes: flp-1 and flp-2, which are located on the chromosome and pSymA megaplasmid, respectively. Bundle-forming pili associated to flp-1 have been involved in competitive nodulation of alfalfa plants but the role of flp-2 remains elusive. In this work, we have performed a comprehensive bioinformatic analyses of T4cP genes in the highly competitive S. meliloti strain GR4 and investigated the role of its flp clusters in pilus biogenesis, motility and in the interaction with alfalfa. Single and double flp cluster mutants were constructed in the wild-type genetic background as well as in a flagellaless derivative strain. Our data demonstrate that both chromosomal and pSymA flp clusters are functional in pili biogenesis, and contribute to surface translocation and nodule formation efficiency of GR4. In this strain, the presence of flp-1 in the absence of flp-2 reduces competitiveness for nodule occupation.
Keywords:
Tad/Flp pili
; Rhizobium
; plant colonization
; nodulation
; competitive ability
1. Introduction
Sinorhizobium meliloti is an alpha-proteobacterium that can live as a saprophyte in the soil but can also establish nitrogen-fixing symbiosis with alfalfa plants [1]. Successful establishment of this symbiosis results in the formation of root nodules, which are infected by bacteria that fix atmospheric dinitrogen into ammonia for direct use by the plant. The progress of this interaction entails different phases and a sophisticated molecular dialogue between the legume host and the rhizobial microsymbiont [1,2,3]. An early and crucial step for the establishment of symbiosis is the bacterial colonization of plant roots, a process that is initiated with the motility of rhizobia towards the plant root and is followed by bacterial attachment to the root surface and the development of a microbial community or biofilm [4,5].
Rhizobial motility is not essential for nodulation or nitrogen fixation but is an important trait for root colonization and infection, which eventually could influence nodulation efficiency and competitiveness [6,7,8,9,10,11]. S. meliloti can move using different mechanisms. Movement of bacteria in aqueous environments or swimming is absolutely reliant on flagellar action whereas translocation of the alfalfa microsymbiont over surfaces can be mediated by both flagella-dependent and –independent mechanisms [10,12,13,14,15]. Swarming is a flagella-driven type of motility characterized by the rapid and coordinated migration of cells across surfaces [16,17,18]. This motility has been described in S. meliloti [10,12,13] and in other rhizobia [19,20,21,22]. S. meliloti flagellaless strains can exhibit a mode of surface spreading referred to as sliding [23]. In this case, bacterial movement is the result of expansive forces of cell growth together with the production of surfactants that reduce surface tension (e.g. exopolysaccharides and the amphiphilic siderophore rhizobactin 1021) [10,13]. Another flagella-independent mode of surface translocation described in many bacteria but not yet in rhizobia is twitching, which is mediated by the extension and retraction of type IV pili (T4P) [24,25]. Like flagella, T4P enable bacteria to not only move but also to attach to surfaces contributing to biofilm formation and host colonization [4,11].
T4P are long, flexible and elastic bacterial extracellular filaments, which are thinner than flagella and are made up of thousands of protein subunits called pilins [24,26,27,28]. These appendages have the peculiarity of being dynamic as rapid polymerization and depolymerization of pilin subunits carried out by a complex machinery permit pili to extend and contract. This characteristic is essential for the different functions associated with T4P [29].
The classification of T4P distinguishes three types: T4aP, T4bP, and the Tight Adherence (Tad) pili, also known as Flp (Fimbrial low-molecular-weight proteins) pili or T4cP [29,30]. One of the features that differentiates the different types of T4P is their major pilin sequences. Pilins of T4aP range between 150-175 amino acids, T4bP pilins are larger (180-200 amino acids) and T4cP are made up of small-sized pilins (50-80 amino acids) [29,31]. In addition, the genes required for T4aP biogenesis are numerous (more than 40) and are often scattered throughout the bacterial genome. In contrast, the synthesis of T4bP and T4cP requires a smaller number of genes (12-14) and they are usually grouped in a region, sometimes with genetic island characteristics [29].
The genes encoding dedicated proteins for polymerization and secretion of T4cP have a very specific and conserved genetic organization as is the case for the Tad system of Aggregatibacter actinomycetemcomitans (formerly Actinobacillus actinomycetemcomitans [32,33,34,35,36]), the cpa (Caulobacter pilus assembly) locus of Caulobacter vibrioides (formerly Caulobacter crescentus [37]), or the ctp (Cluster of type IV pili) locus of Agrobacterium fabrum C58 (formerly Agrobacterium tumefaciens C58 [38]) (Figure 1a). Before the major Flp pilin can be incorporated into the growing T4c pilus, it needs to be processed by a specific prepilin peptidase (TadV/CpaA/CtpB) that removes a leader sequence at a conserved glutamate and tyrosine containing Flp motif (GXXXXEY [34]). TadV also participates in the maturation of the TadE and TadF pseudopilins which share the conserved GXXXXEF sequence at their N-terminal but play an as yet unclear role in pilus biogenesis [35,39,40]. TadA/CpaF/CtpG is the cytosolic ATPase that catalyzes both the extension and retraction of pili with the help of the inner membrane platform proteins TadB/CpaG/CtpH and TadC/CpaH/CtpI [27]. TadZ/CpaE/CtpF is a docking protein for the Tad secretion system, which may help localize pili formation to the poles [40,41,42]. In diderms, T4cP pass through the outer membrane via the secretin RcpA/CpaC/CtpD, which requires the lipoprotein TadD/CpaO that functions as a pilotin enabling the assembly and correct insertion of the secretin into the outer membrane [40,43]. RcpB/CpaD/CtpE and RcpC/CpaB/CtpC probably form a complex with RcpA [36] and possibly are also important for the assembly and stability of the secretin complex [35].
T4cP genes are widely distributed amongst bacteria indicating that they confer significant adaptive advantages [29,35]. They have been involved in diverse functions such as surface sensing, motility, cell adhesion, formation of biofilm, host colonization, virulence, or even in bacterial predation [27,29,44]. Concerning the interaction with plant hosts, T4cP have been involved in the virulence of Ralstonia solanacearum and Pectobacterium atrosepticum [45,46]. In ‘Candidatus Liberibacter asiaticus’, the causal agent of citrus greening disease, Tad pili contribute to the colonization of the insect vector and therefore to disease propagation [47].
There is very little information on T4cP in rhizobia in general, and in S. meliloti in particular. A first indication for pilus like structures in rhizobia and their implication in attachment to roots was described in the soybean symbiont Bradyrhizobium diazoefficiens (formerly Bradyrhizobium japonicum) by Vesper and Bauer (1986) [48]. Information from genome sequencing is revealing that the majority of rhizobial species carry chromosomal gene clusters putatively encoding T4cP, and in many cases additional truncated clusters are also found [11,49,50,51,52,53]. In B. diazoefficiens USDA 110, additional tadGEF genes were found away from the two main T4cP clusters of tad and cpa genes and their loss affected adhesion to soybean roots and caused delayed root infection [50]. In S. meliloti Rm1021, two clusters of genes that encode different components of T4cP have been identified [53]. The flp-1 cluster is located on the chromosome while the second cluster, flp-2, is located on the symbiotic megaplasmid pSymA and appears to be truncated in Rm1021. Deletion of pilA1 that encodes the major pilin of the flp-1 cluster abolishes pili formation and reduces the competitiveness ability of Rm1021 to nodulate alfalfa [53]. However, data available up to now do not implicate these appendages with any type of motility in S. meliloti [53] and the role of each flp cluster in the nodulation process still remains unclear.
S. meliloti GR4 is a highly competitive strain for nodulation of alfalfa [54,55] that exhibits increased biofilm formation on both abiotic and root surfaces compared to Rm1021 [9]. In this work, we have performed a comprehensive bioinformatic analyses of T4cP genes in GR4 and investigated the role of its flp-1 and flp-2 clusters in pilus biogenesis, motility and in the interaction with alfalfa plants by using single and double flp cluster deletion mutants. Our data demonstrate that both chromosomal and pSymA flp clusters are functional in pili biogenesis. These appendages contribute to the surface translocation exhibited by S. meliloti GR4 and influence its symbiotic fitness.
2. Results
2.1. In silico analyses of Type IVc Pili (T4cP) genes of S. meliloti GR4
To identify T4cP-related genes in S. meliloti strain GR4, its genome was scanned for loci similar to tad/cpa/ctp genes (Figure 1, Table 1). We found a total of 29 genes with 16 of them located on the chromosome and 13 on the pSymA megaplasmid. All 29 T4cP-related genes found in GR4 have their corresponding orthologues in the reference strain Rm1021 with sequence identities higher than 98.4%. Therefore, for simplicity, in the text we will refer to the different T4cP genes using the nomenclature of Rm1021 (https://iant.toulouse.inra.fr/bacteria/annotation/cgi/rhime.cgi). In the two S. meliloti strains, GR4 and Rm1021, the majority of the T4cP genes are organized in two clusters: flp-1 located on the chromosome and flp-2 on pSymA (Figure 1b). Additional orphan loci could be identified upstream from flp-1 on the chromosome and a third locus carrying three T4cP genes is located on pSymA (Figure 1b).
The flp-1 cluster shows a synteny very similar to that found in C. vibrioides and carries all of the genes needed to build a complete and functional T4cP apparatus (Figure 1, Figure 2a, Table 1). Indeed, the formation of bundle-forming pili has been associated to this chromosomal cluster in strain Rm1021 [53]. Two copies of the insertion sequence ISRm22 are present in the chromosomal flp cluster of Rm1021 [53] but in GR4 only one copy of this IS element exists (Figure 1b). Nine of the 13 genes in the flp-1 cluster are organized in one operon [56], namely pilA1 coding for the major Flp pilin; cpaA1 that codes for the putative prepilin peptidase; cpaB1, cpaC1 and cpaD1 coding for components of the outer membrane secretin complex; cpaE1 coding for a putative docking protein of the secretion system; cpaF1 encoding the pilus assembly ATPase; and smc02821 and smc02822, orthologues of cpaG and cpaH genes [57], that code for inner membrane platform proteins.
The locus smc02824, located downstream of the ISRm22 element in GR4, codes for a tetratricopeptide repeat (TPR)-containing protein homologous to CpaO. Upstream of pilA1, the locus smc04115 and the operon formed by smc04117 and smc04118 are found. These genes are cpaI, cpaJ and cpaK homologues coding for a CpaC-related secretion pathway protein, and TadE and TadF pseudopilins, respectively [57]. In addition, outside of the flp-1 cluster, two additional chromosomal genes with putative roles in T4cP biogenesis were identified: smc02446 and smc03784 (Figure 1b, Table 1). The 57 amino acid protein encoded by smc02446 is annotated as an Flp pilin component that contains the characteristic Flp motif recognized by the prepilin peptidase, and is a paralogue of PilA1 (60% identity in 71% query coverage) and of the flp-2-encoded pilin PilA2 (64% identity in 43% query coverage) (Figure S1a). According to Schlüter et al. (2013) [56], smc02446 is transcribed in an operon together with smc02447 that codes for a hypothetical transmembrane protein of unknown function. Finally, the orphan smc03784 gene codes for a transmembrane protein that exhibits 39.34% identity with C. vibrioides CpaM, a polar determinant suggested to affect the biogenesis of functional pili portals [57,58]. Both proteins, CpaM and SMc03784, contain a C-terminal domain of divergent polysaccharide deacetylases involved in the removal of N-linked acetyl groups from cell wall polysaccharides.
The flp-2 cluster located on the pSymA megaplasmid lacks IS elements and homologues to four genes found in the chromosomal flp-1 cluster (namely smc04118, cpaC1, cpaD1 and smc02824). In addition, the 10 genes found on the flp-2 cluster exhibit a different organization compared to the flp-1 cluster (Figure 1b). Eight of the ten genes in the flp-2 cluster are organized into two operons [56]. One of them (pilA2-cpaF2-sma1564-sma1562) comprises paralogous genes of the flp-1 cluster encoding a Flp pilin, a pilus assembly ATPase and the two platform proteins of the pilus apparatus, whereas the second operon (cpaA2-cpaB2-cpaE2-sma1572) includes paralogous genes of the flp-1 cluster encoding a prepilin peptidase, a secretin-associated protein, a docking protein for the secretion system, and a pseudopilin. Upstream of cpaA2, two other genes with putative roles in pilus biogenesis could be identified: sma1580 encoding a CpaI homologue, and sma1579 coding for a membrane-associated protein with an N-terminal Tad_N domain found in a family of putative Flp pilus-assembly proteins. No genes homologous to rcpA/cpaC/ctpD coding for the outer membrane secretin were found associated to flp-2, suggesting that this cluster codes for an incomplete secretion system (Figure 2b). Interestingly, outside of the flp-2 cluster on the pSymA, a locus (Locus 3) of three genes (pilQ2-sma0164-sma0166) could be identified, which contains a homolog of cpaC and a gene coding for a protein with characteristic features of T4P pilotins (Figure 1b, Figure 2c, Table 1).
The pilQ2 gene (name denoting its homology to the PilQ secretin of T4aP) of Locus 3 codes for a 466 amino acid (aa) protein with a signal peptide sequence and a secretin-like domain (Figure S2). This protein shows 31% identity with both C. vibrioides CpaC and A. fabrum CtpD, and 30% identity with its chromosomal paralogue CpaC1 (Table 1, Figure S2). Downstream of pilQ2 and forming part of the same operon [56] is sma0164, which codes for a hypothetical 98 aa protein that exhibits significant similarities with proteins encoded by NGR_b10850 and blr3501, which are genes associated to T4cP loci in Sinorhizobium fredii NGR234 and B. diazoefficiens, respectively (Figure 1b, Figure S3). The three proteins contain a prokaryotic membrane lipoprotein lipid attachment profile as identified by InterPro (Figure S3). The profile comprises a precursor signal peptide containing the lipobox motif LXXC that is processed by signal peptidase 2, which cuts upstream of the conserved cysteine residue to which a diacylglycerol is attached [59,60]. The third gene of Locus 3 is sma0166 encoding a conserved hypothetical protein of 176 aas. Although this protein does not exhibit significant homology with CpaO from C. vibrioides or TadD from A. actinomycetemcomitans, all three of these proteins contain a TPR motif (Figure S4). This motif, which is involved in protein-protein interactions and in the assembly of multiprotein complexes [61], is also found in the pilotins TadD and PilF of Pseudomonas aeruginosa [40]. Moreover, and although PROSITE does not identify a prokaryotic membrane lipoprotein profile associated to SMa0166, a lipobox motif is found in the N-terminal region of the protein (Figure S4). The presence in Locus 3 of genes putatively coding for a CpaC homologue and a putative pilotin opens the possibility that this locus complements the flp-2 cluster to function as a complete Flp pilus biogenesis and secretion system.
S. meliloti T4cP-related genes identified in the bioinformatic analyses were compared to T4cP-related genes reported in bacteria belonging to Rhizobiaceae namely the ctp genes of A. fabrum C58 [38], genes of S. fredii NGR234 [51], and the tad/cpa genes of B. diazoefficiens USDA 110 [50] (see Figure 1, Table 1). As expected, the highest similarity both in gene organization as well as in sequence is shared between gene clusters from the same genera, Sinorhizobium. In this manner, the flp-1 gene cluster of S. meliloti shares the highest similarity with the chromosomal Locus 1 of S. fredii NGR324 and the flp-2 cluster with Locus 2 located on the pNGR234b replicon of S. fredii NGR234. The cluster of genes in Locus 3 of S. meliloti GR4 and Sm1021 shares the highest similarity with the cpaC-blr3501-tadD genes of Locus 2 of B. diazoefficiens. The operon encoding the additional Flp pilin found in the chromosome outside of the flp-1 cluster is the most similar to that found upstream of Locus 1 of S. fredii NGR234 and in the linear chromosome of A. fabrum C58. In general, the sequence identity between the gene products of the tad/cpa gene clusters is higher with orthologues in other bacteria than with paralogues of S. meliloti GR4 (Table 1, Table S1) suggesting their origin not to be due to gene duplication events within the same strain.
2.2. Both flp-1 and flp-2 clusters contribute to T4cP production in S. meliloti GR4
In strain Rm1021, formation of Flp pili has been associated to the flp-1 cluster since pilus-like filaments were mostly absent in cells with a pilA1 deletion [53]. However, the rare observation of pilus-like structures in a small percentage of pilA1 mutant cells led the authors to suggest the possibility of pili formation associated either with the expression of the flp-2 cluster or to the incorporation of the Flp pilin encoded by smc02446. To confirm whether both flp-1 and flp-2 clusters are responsible for the formation of Flp pili in S. meliloti GR4, single and double partial cluster mutants were constructed. Although this approach has the drawback of difficulting complementation experiments, it has the advantage of reducing the probability of obtaining unconclusive results due to the possible complementary effect of paralogous genes. In mutant GRflp1, the genes pilA1, cpaA1, cpaB1, cpaC1, cpaD1, cpaE1, and cpaF1 were deleted, whereas mutant GRflp2 harbors a deleted version of the flp-2 cluster in which the pilA2, cpaF2, sma1564 and sma1562 genes were removed (Figure S5). In addition, the GRflp1flp2 double mutant was generated with deleted forms of both the flp-1 and flp-2 clusters. The wild-type and flp single and double mutant strains were examined for the presence of Flp pili by transmission electron microscopy (TEM). For this purpose, cells grown on solid media were used. Two different cell-surface appendages were observed in the wild-type S. meliloti GR4: flagella corresponding to the thicker and more curved filaments, and less abundant, long, thinner and straighter pili-like structures that were often found forming bundles and connecting bacterial cells (Figure 3a,b, Figure S6a,b). The characteristics showed by the the thinner filaments resembled those described for Flp pili of different bacteria, including S. meliloti Rm1021 [38,45,53]. However, neither in the single flp mutants nor in the double mutant could pili-like structures be observed, and only flagella were evident as cell-surface appendages (Figure S6). In order to facilitate the detection of pili structures, flp mutants were created in the flagellaless background offered by the flaAflaB mutant. Due to the lack of the major flagellin FlaA, the flaAflaB mutant cannot produce normal functional flagella and instead short, straight filamentous structures resulting from mutant broken filaments can be observed [10]. Micrographs of the different flagellaless strains revealed that, in addition to the short broken flagellar filaments, pili-like structures were also evident in the flaAflaB mutant (Figure 3c,d) and in its flp simple derivative mutants (Figure 3e,f,g,h) but not in the flagellaless Δflp-1Δflp-2 double mutant (Figure 3i,j). This suggests that both flp clusters function to make pili-like structures.
2.3. Type IVc Pili (T4cP) facilitate surface spreading of S. meliloti GR4
Pili can mediate different types of surface motility [29]. In C. crescentus, Tad pili are able to promote twitching- or walking-like movements [62]. To evaluate the effect of T4cP in S. meliloti GR4 translocation, wild-type as well as single and double flp mutant derivative strains were assessed in different motility assays. Using different media, agar sources and concentrations under the experimental conditions known to promote twitching motility in different bacteria [63], similar limited spreading in the agar/plate interphase was observed for all of the strains (Figure S7). Since no differences were detected between the wild type and the flp mutants in this type of assay, it remains unclear whether the limited spreading observed was due to twitching or to other forms of bacterial motility. Nonetheless, it appears that the flp clusters are not involved in this trait. In contrast, when motility was assessed under conditions that promote swarming in strain GR4 [10], some differences were observed between the wild type and flp derivative mutants. Of the 18 replicates, the frequency of swarming observed for the parental strain was 100%, while 89%, 61% and 67% frequencies were observed for the deletion mutants Δflp-1, Δflp-2, and Δflp-1Δflp-2, respectively. Pairwise comparison using the Chi square statistic indicated significant difference (P<0.01) between the swarming frequency of the wild-type compared to either Δflp-2 or Δflp-1Δflp-2 but not Δflp-1. With respect to the magnitude of surface spreading, surface motility was affected in all mutant strains and significantly so in the double mutant Δflp-1Δflp-2 (Figure 4a,b). These results suggest that the inactivation of the flp-2 cluster has a stronger effect in surface motility than the inactivation of the flp-1 cluster and that this effect was cumulative in the double mutant. When swimming motility was tested in BM (0.3%), all three flp deletion mutants exhibited a slight but significant decrease in their swimming halo compared to that of the wild-type strain, with both the single mutants and the double mutant being equally affected (Figure 4c,d).
2.4. The interaction of S. meliloti GR4 with alfalfa is influenced by T4cP
Deletion of the pilA1 gene in S. meliloti strain Rm1021 reduced competitiveness for nodule occupancy when confronted with the wild type, indicating that Flp pili play a role in Rm1021 symbiotic performance [53]. However, to the best of our knowledge, the role of the pSymA-encoded Flp pili in symbiosis has not yet been reported. In addition, S. meliloti strains Rm1021 and GR4 present remarkable differences including some symbiotic properties with the latter being more efficient in colonizing alfalfa roots and exhibiting higher competitive ability than Rm1021 [9,55]. In this study, the contribution of chromosomal and pSymA-encoded T4cP in the interaction of GR4 with alfalfa plants was investigated. No differences in bacterial attachment to alfalfa roots were found between the wild-type strain or any of the single flp mutants as indicated by the similar number of colony forming units (CFUs) associated to plant roots 2 hours after inoculation (Figure 5). Likewise, the single flp deletion mutants colonized plant roots as efficiently as the wild-type after 24, 48 and 72 hours (Figure 5). In contrast, we found that the double Δflp-1Δflp-2 mutant shows increased adhesion to root surfaces 2 hours post-inoculation compared to the wild-type and the single flp mutants (Figure 5). Remarkably, the increased capacity to attach to root surfaces exhibited by the double mutant does not result in increased bacterial colonization of roots. Indeed, 72 h after inoculation, the number of Δflp-1Δflp-2 CFUs associated to roots was lower than in the wild type, although this difference was not significant. These results indicate that the presence of either of the two Flp pili produced by S. meliloti reduces bacterial attachment to alfalfa roots but this effect does not preclude efficient colonization of the plant host.
Next, to determine whether the flp clusters had an effect on the symbiotic interaction with alfalfa plants, infectivity and competitive abilities of the single and double flp mutants were assayed (Figure 6). All three mutants were able to induce the formation of nitrogen-fixing nodules and no differences in the nodule formation efficiency were detected in the single flp mutants compared with the wild-type (Figure 6a). However, the number of nodules induced by the double mutant was slightly but significantly lower than those of the wild type or single flp mutants at the end of the experiment (13 and 15 days post-inoculation) (Figure 6a). We also determined the competitive ability for nodule occupancy of the flp mutants by inoculating alfalfa plants with bacterial mixtures of the strain to be tested and a marked wild-type strain (S. meliloti GR4 (pGUS3)) at a ratio of 1:1. These competition assays showed that the Δflp-2 mutant showed a significantly lower nodule occupancy than the wild-type strain (Figure 6b). Notably, the reduction in competitiveness was not observed in the double Δflp-1Δflp-2 mutant (Figure 6b), indicating that the lack of T4cP from cluster flp-2 reduces the bacterial competitive ability for nodule occupation only when T4cP from the flp-1 cluster are present. Altogether, these results demonstrate that both flp-1- and flp-2-dependent pili influence S. meliloti symbiotic fitness.
3. Discussion
This work provides insights into T4cP in rhizobia by uncovering the role of the two flp clusters present in the highly competitive S. meliloti strain GR4. We show that both flp-1 and flp-2 are functional in the biogenesis of pili-like structures, and both contribute to the optimal surface translocation exhibited by this rhizobial strain. Our data indicate that, although these flp clusters are dispensable for the formation of nitrogen-fixing nodules, they modulate symbiotic characteristics that might confer adaptive advantages such as bacterial attachment to plant roots, nodule formation efficiency and bacterial competitiveness for nodule occupation.
Despite the participation of T4cP in diverse microbial lifestyles [27,29,44], it is striking that to date, only two studies have addressed the role of T4cP genes in two different rhizobial species [50,53]. Most likely, the abundance of tad/cpa genes in rhizobial genomes [11,35] together with the subtle phenotypes exhibited by T4cP-related mutants under laboratory conditions, discouraged further investigations on these filaments in the legume symbionts. Our bioinformatic analyses identified up to 29 genes putatively related to T4cP in S. meliloti. This number contrasts with the relatively lower number of tad/cpa genes (14-16 genes) found in A. actinomycetemcomitans and C. vibrioides [32,37,57]. Remarkably, the number of loci putatively related to T4cP is even higher in other rhizobia such as S. fredii NGR234 (47 genes) and B. diazoefficiens USDA 110 (39 genes). The maintenance of such a number of pili-related genes throughout evolution could be an indication of the significant adaptive advantages conferred by pili to rhizobial lifestyles. Another remarkable feature of T4cP-related genes in rhizobia compared to tad/cpa genes in A. actinomycetemcomitans and C. vibrioides is their organization in various clusters. In addition to a chromosomal cluster that appears to be conserved in phylogenetically distant bacteria, the three rhizobial species analyzed in our study contain other clusters located in either the chromosome (B. diazoefficiens) or in megaplasmids (pSymA of S. meliloti and pNGR234b of S. fredii NGR234). The existence of more than one cluster of tad/cpa genes is not unique to rhizobia [35,45,64]. Like in S. meliloti, in the bacterial wilt pathogen Ralstonia solanacearum two distinct tad clusters located in the megaplasmid and chromosome were identified [45]. Characterization of a tadA2 mutant of this bacterium revealed that the Flp pili associated to the megaplasmid cluster are required for virulence on potato plants [45] whereas no information exists regarding the function of the chromosomal cluster. The opportunistic animal pathogen Vibrio vulnificus harbors three tad loci and only the triple mutant exhibited decreased virulence in mice, indicating that the tad loci work cooperatively in this species during pathogenicity [64].
In S. meliloti, the chromosomal flp-1 cluster carries all of the genes needed for T4cP biogenesis, and the formation of bundle-forming pili was previously associated to this cluster in strain Rm1021 [53]. In contrast, flp-2 lacks some key genes for the secretion system to be functional. The lack of cpaK and cpaD homologues should not be an impediment for T4cP biogenesis. Indeed, A. fabrum and Pseudomonas aeruginosa are capable of assembling Flp pili in the absence of cpaK and cpaD homologues, respectively [38,65]. However, the lack of genes coding for components of the outer membrane secretin channel (cpaC and cpaO) could prevent pilus biogenesis associated to the flp-2 cluster. Interestingly, an additional set of three genes located on pSymA (Locus 3) was identified that could complement genes in flp-2. Indeed, TEM analysis of flagellaless mutants carrying single large deletions in either the flp-1 or flp-2 clusters revealed the production of filaments while these fimbriae structures were no longer observed in a flagellaless double flp-1flp-2 mutant. These results indicate that, under our experimental conditions, the two flp clusters are functional in the formation of pili. In the study carried out by Zatakia and co-workers (2014)[53], pili-like structures were only rarely observed in the pilA1 mutant of Rm1021, which the authors explained as the result of T4cP assembly from the flp-2 cluster and/or the incorporation of the pilin encoded by the orphan smc02446 gene. The latter is a very likely possibility considering that the two pilin genes in A. fabrum, ctpA and pilA, are functionally interchangeable [38]. The approach used in our study in which deletions of gene clusters were generated in the flp regions instead of deleting individual genes, reduces the possibility of inconclusive results due to possible complementation effects of paralogous genes. In our flaAflaBΔflp-1 mutant, in which not only pilA1 but also cpaA1, cpaB1, cpaC1, cpaD1, cpaE1 and cpaF1 were deleted, pili forming bundles connecting cells were clearly observed that cannot be the result of the incorporation of the smc02446-encoded pilin. Moreover, pili-like structures could not be detected in flaAflaBΔflp-1Δflp-2 indicating that the filaments observed in flaAflaBΔflp-1 are due to a fully functional flp-2 cluster, possibly with the implication of Locus 3 which contains all three genes missing in flp-2. However, experiments are still needed to demonstrate that Locus 3 is essential for pili biogenesis associated to the flp-2 cluster.
The splitting of genes required for the assembly of T4cP in distinct genetic clusters is not usual [28] but at least one example has been reported in the literature. In the bacterial predator Myxococcus xanthus, the assembly of the Tad-like Kil system involved in contact-dependent prey killing is the result of the expression of two clusters that carry complementary sets of genes [44]. In addition, our bioinformatic analyses revealed that in S. fredii, Locus 2 on pNGR234b shows high similarity both in gene organization and sequence with S. meliloti flp-2 and, like the latter, it also lacks genes coding for the secretin complex. Homologs to S. meliloti Locus 3 genes can be identified in a different region in pNGR234b (NGR_b10860, NGR_b10850 and NGR_b10740 in Locus 3), albeit exhibiting a different gene organization and clustered together with additional T4cP-related genes. Clearly, further investigations are needed in order to elucidate if distinct co-existing tad/flp clusters in rhizobia play different or specific roles or if they have complementary functions.
In this work, pili-like structures were more easily detectable in the flagellaless strains containing one or the two flp clusters than in the wild type strain GR4 (Figure 3). A simple explanation could be that the abundant flagellar filaments present in the wild-type strain are hampering the observation of the thinner and scarcer pili-like structures. However, the possibility that the lack of flagellar filaments has an impact on T4cP gene expression in S. meliloti should not be discarded and deserves further investigation. In bacteria in which T4cP have been studied the most, expression of the pilus associated genes and activity of the filament are controlled by complex transcriptional and posttranslational regulatory mechanisms [29,35]. In P. aeruginosa, a feedback regulation between pili and flagellar components occurs through a two-component regulatory system [66]. Moreover, C. crescentus flagellar mutants are significantly deficient in pili biogenesis and it was suggested that different stages of the flagellum assembly act as checkpoints for the regulation of T4cP associated gene expression [67]. Another open question that was not addressed in this study is whether the two flp clusters in S. meliloti are expressed under the same conditions. The presence of an accessory chemotaxis system next to the flp-2 cluster in pSymA is intriguing. This chemotaxis system belongs to the alternative cellular function (ACF) and most likely does not control flagella [11]. It would be worth investigating whether this chemotaxis system controls the activity of T4cP in S. meliloti in an analogous way to how the Pil-Chp pathway controls T4aP in other bacteria [68].
Tad pili are able to promote twitching- or walking-like movements in C. crescentus [62]. Surface motility assays performed with our S. meliloti flp deletion mutants indicate that the flp clusters contribute cooperatively to the optimal surface motility exhibited by strain GR4 with the flp-2 cluster having a stronger effect than flp-1. Under the experimental conditions used in our study, surface motility in GR4 is totally dependent on flagellar action [10]. At present, we can only speculate about how flp-1 and flp-2 affect swarming motility in GR4. Our data indicate that the double flp-1flp-2 mutant exhibits greater attachment to alfalfa roots compared to the wild type (Figure 5). An increased adhesion to surfaces facilitated by the absence of pili could hamper proper bacterial translocation. On the other hand, pili together with flagella participate in surface sensing in bacteria [69] and specifically Tad pili act as surface contact sensors for C. crescentus [30,62,70]. An attractive possibility awaiting investigation is that pilus-based mechanosensation in S. meliloti triggers a signal transduction cascade that promotes swarming and other physiological adaptations to thrive on surfaces.
Information about the role of T4cP in the establishment of symbiosis with legumes is very scarce. Results obtained in this study indicate that T4cP impact the symbiotic fitness of S. meliloti by affecting adhesion to plant roots, nodule formation efficiency and competitiveness. The role of rhizobial T4cP in favoring adhesion to plant roots has been previously proposed in two studies [50,53]. In B. diazoefficiens USDA 110, inactivation of tadG or the cluster tadGEF (Locus 3 of B. diazoefficiens USDA 110 in Figure 1b) impaired adhesion to soybean roots and caused delayed root infection [50]. However, the connection between the symbiotic phenotype of tadGEF mutants and pili formation in B. diazoefficiens USDA 110 was not directly examined. In contrast to B. diazoefficiens and S. fredii NGR234, no tadG homologue could be identified in the genome of S. meliloti. On the other hand, the reduced competitive ability for nodule occupation exhibited by a S. meliloti pilA1 mutant was suggested to be due to a lower bacterial attachment to plant roots, although this possibility was not experimentally tested [53]. Results obtained in our study do not support a role for S. meliloti T4cP in promoting adhesion to root surfaces. As already discussed, the double flp-1flp-2 mutant, which lacks pili-like structures, exhibits greater attachment to plant roots compared to the wild-type and single flp mutant strains (Figure 5). These observations demonstrate that T4cP are not required for attachment of S. meliloti cells to alfalfa roots, and suggest that the presence of T4cP hamper attachment to the root surface mediated by other cell surface structures, perhaps flagella, exopolysaccharides and/or lipopolysaccharide [4].
Our data also demonstrate that the increased binding to root surfaces shown by the double flp cluster mutant does not significantly affect the biofilm formed on plant roots 24, 48 or 72 hours after inoculation, indicating that, under our experimental conditions, S. meliloti T4cP are not essential for plant root colonization during the early stages of the interaction. Likewise, in A. fabrum no differences were detected between the 48 h biofilm formed on Arabidopsis thaliana roots by the wild-type strain and a ctpA derivative mutant [38]. Nevertheless, our S. meliloti double flp mutant showed decreased nodule formation efficiency indicated by the significantly lower number of nodules induced by this strain compared to the wild type or single flp mutants. At present, the reasons for the lower nodulation performance of the double flp mutant are unknown. Perhaps the increased attachment to the root surface shown by the mutant together with the lower ability to move across surfaces reduces the probability of rhizobial cells to find potential infection sites. Another, non-excluding possibility is that T4cP, directly or indirectly, contribute to defend S. meliloti cells against plant defense responses with a role similar to the stealth hiding role proposed for the Tad pili in V. vulnificus [64].
Finally, our data show that deletion of flp-1 in GR4 does not affect its competitive ability for nodule occupation. This result contrasts with the lower competitiveness exhibited by a pilA1 deletion mutant compared with its corresponding wild- type strain Rm1021 [53]. Differences between GR4 and Rm1021 and/or the type of mutation used to analyse the role of T4cP associated to flp-1, could explain the contrasting results obtained in the two studies. Interestingly, although the lack of flp-1 has no influence on GR4 competitiveness, the presence of this cluster in a flp-2 deletion mutant reduces its ability to compete for nodule occupation. This is based on the lower nodule occupancy exhibited by the single flp-2 mutant that is recovered when flp-1 is deleted. A possible explanation for this result is that in the absence of flp-2 pili, interbacterial attachment mediated by Flp-1 pili is increased, which could hamper dissemination to other infection sites. This seems not to be crucial in single inoculation experiments as shown by similar nodulation kinetics of the flp-2 mutant and the wild-type strain, but confers a disadvantage when competing with another bacterium.
In summary, this study increases our knowledge on T4cP in S. meliloti and their role in the interaction with its plant host, and at the same time, highlights the complexity of their study. New questions have arisen and investigations aimed to answer them might help us to better understand how extracellular filament appendages contribute to the symbiotic fitness of rhizobia.
4. Materials and Methods
4.1. Bacterial strains, plasmids and growth conditions
Bacterial strains and plasmids used in this work are listed in Table S2. Escherichia coli cultures were routinely grown in LB medium [71] at 37°C. S. meliloti strains were grown in either complex tryptone yeast (TY) medium [72] or in Robertsen minimal medium (MM) [73] at 28°C. When required, antibiotics were added to final concentrations (µg mL-1) of: 50 kanamycin and 25 hygromycin for E. coli; 200 kanamycin and 75 hygromycin for S. meliloti. All reagents were obtained from Sigma-Aldrich (Steinheim, Germany), unless otherwise specified.
4.2. In silico analyses
In order to identify and compare tad/cpa/ctp genes from S. meliloti GR4, A. actinomycetemcomitans CU1000N, C. vibrioides NA1000, A. fabrum C58, S. fredii NGR234 and B. diazoefficiens USDA110, genome sequences were retrieved from the NCBI database (https://www.ncbi.nlm.nih.gov/datasets/genome/). Comparisons of these genomes were performed in the Rapid Annotations using Subsystems Technology (RAST) server version 2.0 [74]. Further gene identification was performed using resources at KEGG (https://www.genome.jp/kegg/ , [75]) and BV-BRC (https://www.bv-brc.org/ , [76]. Sequence comparisons of individual gene products were performed with blastp (https://blast.ncbi.nlm.nih.gov/Blast.cgi) and Clustal MUSCLE sequence alignment at https://www.ebi.ac.uk/Tools/msa/muscle/. Protein domains within individual gene products were examined using InterPro (https://www.ebi.ac.uk/interpro/ , [77]).
4.3. Construction of S. meliloti mutant strains
flp-1 and flp-2 mutants were obtained by allelic exchange of each flp cluster with markerless deleted versions obtained in vitro by overlap extension PCR using primers delpilA1.1 to delpilA1.4 and delpilA2.1 to delpilA2.4 (Table S3) and Phusion high-fidelity DNA polymerase. The resulting amplicons corresponding to deleted versions of flp-1 and flp-2 were first cloned into pCR-XL-TOPO and confirmed by DNA sequencing and then, subcloned into pK18mobsacB as BamHI or HindIII fragments to give pK18-Δflp1 and pK18-Δflp2, respectively. Each one of these constructions was introduced into S. meliloti GR4 by conjugation using S17-1 as the donor strain [78] and the allelic exchange was selected as previously described [79] to obtain the single GRflp1 and GRflp2 mutants. The double GRflp1flp2 mutant was generated by selecting the allelic exchange of the flp-2 cluster with the corresponding deleted version in the single GRflp1 mutant. To obtain flagellaless flp mutants, plasmid pK18flaAB::Hyg was introduced into single and double flp mutants and replacement of wild type flagellin genes with the mutated version was selected on plates containing hygromycin. All mutants were checked by Southern hybridization with specific probes.
4.4. Transmission electron microscopy (TEM)
Cells for TEM observations were obtained using the same procedures as that described in Calatrava-Morales et al. (2017) [80] from the edge of colonies grown on solid MM (1% agar). In brief, carbon-coated Formvar grids were placed for 5 min on top of a drop of water (sterile MilliQ) previously applied to the colony border. The grids were then washed twice in water for 1 min and stained with 2% (wt/vol) uranyl acetate for 3 min. Grids were allowed to air dry for at least 1 h and visualized using a JEOL JEM-1011 transmission electron microscope with a 100-kV beam at the Microscopy Service of the Estación Experimental del Zaidín (Granada, Spain). Images were captured with an Olympus SIS Megaview III CCD camera for TEM.
4.5. Motility assays
To determine swimming motility, S. meliloti strains were grown in liquid TY medium at 28 ºC until they reached an optical density (OD600nm) of approximately 0.5. Aliquots of 3 μL of each culture were placed onto plates containing Bromfield medium (BM) (0.04% tryptone, 0.01% yeast extract, 0.01% CaCl2×2H2O, 0.3% Bacto Agar, Difco BM). After incubation at 28°C for 72 hours, the dispersion halo of each colony was measured.
Swarming motility assays were performed as described by López-Lara et al. (2018) [81]. Briefly, cultures of the S. meliloti strains in liquid TY medium were grown overnight to an OD600 greater than 1. Subsequently, each culture was washed twice with liquid MM and after the last wash, the pellet was resuspended in 80 μL of liquid MM. 2 μL aliquots of the suspension were inoculated and allowed to dry for 10 minutes onto the surface of MM plates (0.6% noble agar, Difco BM), which were previously dried for 15 min. The plates were incubated at 28°C for 48 hours. To quantify motility, the length of each of the sides of the extended colony was measured in millimeters. Colonies which had extended 7 mm or further were considered positive in swarming.
Twitching motility was assayed according to the protocol described by Turnbull and Whitchurch et al. (2014) [63] with small modifications. Plates containing 10 ml of either TY (1% Bacteriologic Agar), MM (1.3% Agar, Grade A Difco), MM (1% Agar, Grade A Difco) and MM (1% Agar, Noble) were prepared and dried according to the protocol. Strains were stab inoculated with a toothpick to the bottom of the plate and incubated for 2-4 days at 28 °C in a humidified chamber.
4.6. Plant assays
Alfalfa (Medicago sativa L. ‘Victoria’) plants were surface sterilized, germinated, and grown in hydroponic cultures under axenic conditions in glass tubes containing nitrogen-free nutrient solution, as previously described [82].
To test the degree of infectivity of each strain, 9-day-old plants (18-20 replicates, one plant per tube) were inoculated with 1 ml of a rhizobial suspension containing 5 × 106 cells. Prior to inoculation, bacteria were grown to the exponential phase (OD600 = 0.5 to 0.6) in TY broth and diluted 100-fold in sterile water. Four days after inoculation, the number of nodules per plant was recorded over two weeks.
To examine competitive ability, 12 individual plants grown as described for the infectivity test were inoculated with 1 ml of a bacterial suspension prepared as already described, containing mixtures at a ratio of 1:1 of the S. meliloti strain to be tested and GR4 marked with the pGUS3 plasmid [83]. Roots were collected 15 days after inoculation, briefly washed with water, and incubated overnight in the dark at 37°C in 1 mM 5-bromo-4-chloro-3-indolyl-b-D-glucuronide (Apollo Scientific) in 50 mM sodium phosphate buffer (pH 7.5) with 1% SDS. Those nodules occupied by the strain harboring the pGUS3 plasmid stained blue; thus, nodule occupancy could be determined by counting blue and white nodules produced by each bacterial mixture in three independent experiments.
Colonization assays were performed as Calatrava-Morales et al. (2017) [80]. Briefly, 1 week old alfalfa plants (5 plants/tube) were inoculated with 1 ml rhizobial suspension as described above. At different times, 15 roots from each treatment were collected and washed three times with sterile distilled water, and pools of roots from 5 plants introduced onto an Eppendorf tube containing sterile Tris-EDTA buffer (10 mM Tris-HCl [pH 7.5] and 1 mM EDTA). Tubes were sonicated (2x for 1 min each) in a Selecta Ultrasons Sonicator bath. Released rhizobia were quantified by counting colony forming units (CFU) of serial dilutions, and data were plotted as CFU per gram of root fresh weight (RFW).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Multiple sequence alignment by MUSCLE of Flp prepilins; Figure S2: Multiple sequence alignment by MUSCLE of PilQ2-like gene products; Figure S3: Multiple sequence alignment by MUSCLE of SMa0164 with homologous proteins of S. fredii NGR234 (Sf) and B. diazoefficiens (Bd); Figure S4: Multiple sequence alignment by MUSCLE of SMa0166 with TadD-like gene products; Figure S5: Genomic regions corresponding to the flp-1 and flp-2 clusters of S. meliloti GR4 and their corresponding unmarked in-frame deleted versions obtained in this study; Figure S6: Transmission electron microscope (TEM) images of flagella-plus single and double flp deletion mutants; Figure S7: Twitching motility assay of duplicate stab inoculations of the different strains on different media with different agar concentrations and types; Table S1: Percent sequence identities (in parenthesis) among S. meliloti T4cP paralogues; Table S2: Strains and plasmids used in this study; Table S3: List of primers used in this study.
Author Contributions
Conceptualization, P.v.D., and M.J.S.; formal analysis, C.C.-H., V.C., L.M.B-R., and P.v.D.; investigation, C.C.-H., V.C., and L.M.B-R., and M.J.S.; writing—original draft preparation, C.C.-H., and P.v.D.; writing—review and editing, C.C.-H., V.C., L.M.B-R.; P.v.D., and M.J.S.; visualization, C.C.-H., P.v.D., and M.J.S.; supervision, M.J.S.; funding acquisition, M.J.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by PGC2018-096477-B-I00 and PID2021-123540NB-I00 funded by MCIN/AEI/ 10.13039/501100011033 and by “ERDF A way of making Europe”. CCH is supported by CSIC, JAE program.
Data Availability Statement
Not applicable.
Acknowledgments
Microscopy analyses were carried out at the Microscopy service of the Estación Experimental del Zaidín (CSIC). We thank Alicia Rodríguez for help with TEM observations and the assistance of Joaquina Nogales in the design of primers.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Poole, P.; Ramachandran, V.; Terpolilli, J. Rhizobia: from saprophytes to endosymbionts. Nat Rev Microbiol 2018, 16, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Oldroyd, G.E.; Murray, J.D.; Poole, P.S.; Downie, J.A. The rules of engagement in the legume-rhizobial symbiosis. Annu Rev Genet 2011, 45, 119–144. [Google Scholar] [CrossRef] [PubMed]
- Cangioli, L.; Vaccaro, F.; Fini, M.; Mengoni, A.; Fagorzi, C. Scent of a Symbiont: The Personalized Genetic Relationships of Rhizobium-Plant Interaction. Int J Mol Sci 2022, 23, 3358. [Google Scholar] [CrossRef] [PubMed]
- Wheatley, R.M.; Poole, P.S. Mechanisms of bacterial attachment to roots. FEMS Microbiol Rev 2018, 42, 448–461. [Google Scholar] [CrossRef] [PubMed]
- Knights, H.E.; Jorrin, B.; Haskett, T.L.; Poole, P.S. Deciphering bacterial mechanisms of root colonization. Environ Microbiol Rep 2021, 13, 428–444. [Google Scholar] [CrossRef] [PubMed]
- Ames, P.; Bergman, K. Competitive advantage provided by bacterial motility in the formation of nodules by Rhizobium meliloti. J Bacteriol 1981, 148, 728–908. [Google Scholar] [CrossRef]
- Soby, S.; Bergman, K. Motility and Chemotaxis of Rhizobium meliloti in Soil. Appl Environ Microbiol 1983, 46, 995–998. [Google Scholar] [CrossRef] [PubMed]
- Caetano-Anollés, G.; Wrobel-Boerner, E.; Bauer, W.D. Growth and Movement of Spot Inoculated Rhizobium meliloti on the Root Surface of Alfalfa. Plant Physiol 1992, 98, 1181–1189. [Google Scholar] [CrossRef]
- Amaya-Gómez, C.V.; Hirsch, A.M.; Soto, M.J. Biofilm formation assessment in Sinorhizobium meliloti reveals interlinked control with surface motility. BMC Microbiol 2015, 15, 58. [Google Scholar] [CrossRef]
- Bernabéu-Roda, L.; Calatrava-Morales, N.; Cuéllar, V.; Soto, M. Characterization of surface motility in Sinorhizobium meliloti: regulation and role in symbiosis. Symbiosis 2015, 1–12. [Google Scholar] [CrossRef]
- Aroney, S.T.N.; Poole, P.S.; Sanchez-Canizares, C. Rhizobial Chemotaxis and Motility Systems at Work in the Soil. Front Plant Sci 2021, 12, 725338. [Google Scholar] [CrossRef]
- Soto, M.J.; Fernández-Pascual, M.; Sanjuán, J.; Olivares, J. A fadD mutant of Sinorhizobium meliloti shows multicellular swarming migration and is impaired in nodulation efficiency on alfalfa roots. Mol. Microbiol 2002, 43, 371–382. [Google Scholar] [CrossRef]
- Nogales, J.; Bernabéu-Roda, L.; Cuéllar, V.; Soto, M.J. ExpR is not required for swarming but promotes sliding in Sinorhizobium meliloti. J Bacteriol 2012, 194, 2027–2035. [Google Scholar] [CrossRef]
- Dilanji, G.E.; Teplitski, M.; Hagen, S.J. Entropy-driven motility of Sinorhizobium meliloti on a semi-solid surface. Proc Biol Sci 2014, 281, 20132575. [Google Scholar] [CrossRef]
- Ji, Y.Y.; Zhang, B.; Zhang, P.; Chen, L.C.; Si, Y.W.; Wan, X.Y.; Li, C.; Wang, R.H.; Tian, Y.; Zhang, Z.; et al. Rhizobial migration toward roots mediated by FadL-ExoFQP modulation of extracellular long-chain AHLs. ISME J 2023, 17, 417–431. [Google Scholar] [CrossRef]
- Kearns, D.B. A field guide to bacterial swarming motility. Nat Rev Microbiol 2010, 8, 634–644. [Google Scholar] [CrossRef]
- Mattingly, A.E.; Weaver, A.A.; Dimkovikj, A.; Shrout, J.D. Assessing Travel Conditions: Environmental and Host Influences On Bacterial Surface Motility. J Bacteriol 2018, 200, e00014–00018. [Google Scholar] [CrossRef]
- Wadhwa, N.; Berg, H.C. Bacterial motility: machinery and mechanisms. Nat Rev Microbiol 2022, 20, 161–173. [Google Scholar] [CrossRef]
- Daniels, R.; Reynaert, S.; Hoekstra, H.; Verreth, C.; Janssens, J.; Braeken, K.; Fauvart, M.; Beullens, S.; Heusdens, C.; Lambrichts, I.; et al. Quorum signal molecules as biosurfactants affecting swarming in Rhizobium etli. Proc Natl Acad Sci U S A 2006, 103, 14965–14970. [Google Scholar] [CrossRef]
- Tambalo, D.D.; Yost, C.K.; Hynes, M.F. Characterization of swarming motility in Rhizobium leguminosarum bv. viciae. FEMS Microbiol Lett 2010, 307, 165–174. [Google Scholar] [CrossRef]
- Covelli, J.M.; Althabegoiti, M.J.; López, M.F.; Lodeiro, A.R. Swarming motility in Bradyrhizobium japonicum. Res Microbiol 2013, 164, 136–144. [Google Scholar] [CrossRef]
- Alías-Villegas, C.; Fuentes-Romero, F.; Cuéllar, V.; Navarro-Gómez, P.; Soto, M.J.; Vinardell, J.M.; Acosta-Jurado, S. Surface motility regulation of Sinorhizobium fredii HH103 by plant flavonoids and the NodD1, TtsI, NolR, and MucR1 symbiotic bacterial regulators. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Holscher, T.; Kovacs, A.T. Sliding on the surface: bacterial spreading without an active motor. Environ Microbiol 2017, 19, 2537–2545. [Google Scholar] [CrossRef]
- Mattick, J.S. Type IV pili and twitching motility. Annu Rev Microbiol 2002, 56, 289–314. [Google Scholar] [CrossRef]
- Maier, B.; Wong, G.C.L. How bacteria use Type IV pili machinery on surfaces. Trends Microbiol 2015, 23, 775–788. [Google Scholar] [CrossRef]
- Pelicic, V. Type IV pili: e pluribus unum? Mol Microbiol 2008, 68, 827–837. [Google Scholar] [CrossRef]
- Craig, L.; Forest, K.T.; Maier, B. Type IV pili: dynamics, biophysics and functional consequences. Nat Rev Microbiol 2019, 17, 429–440. [Google Scholar] [CrossRef]
- Denise, R.; Abby, S.S.; Rocha, E.P.C. Diversification of the type IV filament superfamily into machines for adhesion, protein secretion, DNA uptake, and motility. PLoS Biol 2019, 17, e3000390. [Google Scholar] [CrossRef]
- Ellison, C.K.; Whitfield, G.B.; Brun, Y.V. Type IV Pili: dynamic bacterial nanomachines. FEMS Microbiol Rev 2022, 46. [Google Scholar] [CrossRef] [PubMed]
- Ellison, C.K.; Kan, J.; Dillard, R.S.; Kysela, D.T.; Ducret, A.; Berne, C.; Hampton, C.M.; Ke, Z.; Wright, E.R.; Biais, N.; et al. Obstruction of pilus retraction stimulates bacterial surface sensing. Science 2017, 358, 535–538. [Google Scholar] [CrossRef] [PubMed]
- Giltner, C.L.; Nguyen, Y.; Burrows, L.L. Type IV pilin proteins: versatile molecular modules. Microbiol Mol Biol Rev 2012, 76, 740–772. [Google Scholar] [CrossRef]
- Kachlany, S.C.; Planet, P.J.; Bhattacharjee, M.K.; Kollia, E.; DeSalle, R.; Fine, D.H.; Figurski, D.H. Nonspecific adherence by Actinobacillus actinomycetemcomitans requires genes widespread in bacteria and archaea. J Bacteriol 2000, 182, 6169–6176. [Google Scholar] [CrossRef]
- Kachlany, S.C.; Planet, P.J.; DeSalle, R.; Fine, D.H.; Figurski, D.H. Genes for tight adherence of Actinobacillus actinomycetemcomitans: from plaque to plague to pond scum. Trends Microbiol 2001, 9, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Kachlany, S.C.; Planet, P.J.; Desalle, R.; Fine, D.H.; Figurski, D.H.; Kaplan, J.B. flp-1, the first representative of a new pilin gene subfamily, is required for non-specific adherence of Actinobacillus actinomycetemcomitans. Mol Microbiol 2001, 40, 542–554. [Google Scholar] [CrossRef] [PubMed]
- Tomich, M.; Planet, P.J.; Figurski, D.H. The tad locus: postcards from the widespread colonization island. Nat Rev Microbiol 2007, 5, 363–375. [Google Scholar] [CrossRef]
- Roux, N.; Spagnolo, J.; de Bentzmann, S. Neglected but amazingly diverse type IVb pili. Res Microbiol 2012, 163, 659–673. [Google Scholar] [CrossRef]
- Skerker, J.M.; Shapiro, L. Identification and cell cycle control of a novel pilus system in Caulobacter crescentus. EMBO J 2000, 19, 3223–3234. [Google Scholar] [CrossRef]
- Wang, Y.; Haitjema, C.H.; Fuqua, C. The Ctp type IVb pilus locus of Agrobacterium tumefaciens directs formation of the common pili and contributes to reversible surface attachment. J Bacteriol 2014, 196, 2979–2988. [Google Scholar] [CrossRef] [PubMed]
- Tomich, M.; Fine, D.H.; Figurski, D.H. The TadV protein of Actinobacillus actinomycetemcomitans is a novel aspartic acid prepilin peptidase required for maturation of the Flp1 pilin and TadE and TadF pseudopilins. J Bacteriol 2006, 188, 6899–6914. [Google Scholar] [CrossRef]
- Tassinari, M.; Rudzite, M.; Filloux, A.; Low, H.H. Assembly mechanism of a Tad secretion system secretin-pilotin complex. Nat Commun 2023, 14, 5643. [Google Scholar] [CrossRef]
- Xu, Q.; Christen, B.; Chiu, H.J.; Jaroszewski, L.; Klock, H.E.; Knuth, M.W.; Miller, M.D.; Elsliger, M.A.; Deacon, A.M.; Godzik, A.; et al. Structure of the pilus assembly protein TadZ from Eubacterium rectale: implications for polar localization. Mol Microbiol 2012, 83, 712–727. [Google Scholar] [CrossRef]
- Perez-Cheeks, B.A.; Planet, P.J.; Sarkar, I.N.; Clock, S.A.; Xu, Q.; Figurski, D.H. The product of tadZ, a new member of the parA/minD superfamily, localizes to a pole in Aggregatibacter actinomycetemcomitans. Mol Microbiol 2012, 83, 694–711. [Google Scholar] [CrossRef]
- Clock, S.A.; Planet, P.J.; Perez, B.A.; Figurski, D.H. Outer membrane components of the Tad (tight adherence) secreton of Aggregatibacter actinomycetemcomitans. J Bacteriol 2008, 190, 980–990. [Google Scholar] [CrossRef] [PubMed]
- Seef, S.; Herrou, J.; de Boissier, P.; My, L.; Brasseur, G.; Robert, D.; Jain, R.; Mercier, R.; Cascales, E.; Habermann, B.H.; et al. A Tad-like apparatus is required for contact-dependent prey killing in predatory social bacteria. eLife 2021, 10. [Google Scholar] [CrossRef]
- Wairuri, C.K.; van der Waals, J.E.; van Schalkwyk, A.; Theron, J. Ralstonia solanacearum needs Flp pili for virulence on potato. Mol Plant Microbe Interact 2012, 25, 546–556. [Google Scholar] [CrossRef]
- Nykyri, J.; Mattinen, L.; Niemi, O.; Adhikari, S.; Kõiv, V.; Somervuo, P.; Fang, X.; Auvinen, P.; Mäe, A.; Palva, E.T.; et al. Role and regulation of the Flp/Tad pilus in the virulence of Pectobacterium atrosepticum SCRI1043 and Pectobacterium wasabiae SCC3193. PLOS ONE 2013, 8, e73718. [Google Scholar] [CrossRef]
- Andrade, M.; Wang, N. The Tad Pilus Apparatus of 'Candidatus Liberibacter asiaticus' and Its Regulation by VisNR. Mol Plant Microbe Interact 2019, 32, 1175–1187. [Google Scholar] [CrossRef]
- Vesper, S.J.; Bauer, W.D. Role of Pili (Fimbriae) in Attachment of Bradyrhizobium japonicum to Soybean Roots. Appl Environ Microbiol 1986, 52, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Krehenbrink, M.; Downie, J.A. Identification of protein secretion systems and novel secreted proteins in Rhizobium leguminosarum bv. viciae. BMC Genomics 2008, 9, 55. [Google Scholar] [CrossRef] [PubMed]
- Mongiardini, E.J.; Parisi, G.D.; Quelas, J.I.; Lodeiro, A.R. The tight-adhesion proteins TadGEF of Bradyrhizobium diazoefficiens USDA 110 are involved in cell adhesion and infectivity on soybean roots. Microbiol Res 2016, 182, 80–88. [Google Scholar] [CrossRef]
- Schmeisser, C.; Liesegang, H.; Krysciak, D.; Bakkou, N.; Le Quere, A.; Wollherr, A.; Heinemeyer, I.; Morgenstern, B.; Pommerening-Roser, A.; Flores, M.; et al. Rhizobium sp. strain NGR234 possesses a remarkable number of secretion systems. Appl Environ Microbiol 2009, 75, 4035–4045. [Google Scholar] [CrossRef] [PubMed]
- Wibberg, D.; Blom, J.; Jaenicke, S.; Kollin, F.; Rupp, O.; Scharf, B.; Schneiker-Bekel, S.; Sczcepanowski, R.; Goesmann, A.; Setubal, J.C.; et al. Complete genome sequencing of Agrobacterium sp. H13-3, the former Rhizobium lupini H13-3, reveals a tripartite genome consisting of a circular and a linear chromosome and an accessory plasmid but lacking a tumor-inducing Ti-plasmid. J Biotechnol 2011, 155, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Zatakia, H.M.; Nelson, C.E.; Syed, U.J.; Scharf, B.E. ExpR coordinates the expression of symbiotically important, bundle-forming Flp pili with quorum sensing in Sinorhizobium meliloti. Appl Environ Microbiol 2014, 80, 2429–2439. [Google Scholar] [CrossRef] [PubMed]
- Casadesús, J.; Olivares, J. Rough and fine linkage mapping of the Rhizobium meliloti chromosome. Mol Gen Genet 1979, 174, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Bellabarba, A.; Bacci, G.; Decorosi, F.; Aun, E.; Azzarello, E.; Remm, M.; Giovannetti, L.; Viti, C.; Mengoni, A.; Pini, F. Competitiveness for Nodule Colonization in Sinorhizobium meliloti: Combined In Vitro-Tagged Strain Competition and Genome-Wide Association Analysis. mSystems 2021, 6, e0055021. [Google Scholar] [CrossRef] [PubMed]
- Schlüter, J.P.; Reinkensmeier, J.; Barnett, M.J.; Lang, C.; Krol, E.; Giegerich, R.; Long, S.R.; Becker, A. Global mapping of transcription start sites and promoter motifs in the symbiotic alpha-proteobacterium Sinorhizobium meliloti 1021. BMC Genomics 2013, 14, 156. [Google Scholar] [CrossRef] [PubMed]
- Christen, M.; Beusch, C.; Bösch, Y.; Cerletti, D.; Flores-Tinoco, C.E.; Del Medico, L.; Tschan, F.; Christen, B. Quantitative Selection Analysis of Bacteriophage φCbK Susceptibility in Caulobacter crescentus. J Mol Biol 2016, 428, 419–430. [Google Scholar] [CrossRef] [PubMed]
- Mignolet, J.; Panis, G.; Viollier, P.H. More than a Tad: spatiotemporal control of Caulobacter pili. Curr Opin Microbiol 2018, 42, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, S.; Wu, H.C. Lipoproteins in bacteria. J Bioenerg Biomembr 1990, 22, 451–471. [Google Scholar] [CrossRef]
- Kaushik, S.; He, H.; Dalbey, R.E. Bacterial Signal Peptides- Navigating the Journey of Proteins. Front Physiol 2022, 13. [Google Scholar] [CrossRef]
- DAndrea, L.D.; Regan, L. TPR proteins: the versatile helix. Trends Biochem Sci 2003, 28, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Sangermani, M.; Hug, I.; Sauter, N.; Pfohl, T.; Jenal, U. Tad Pili Play a Dynamic Role in Caulobacter crescentus Surface Colonization. MBio 2019, 10. [Google Scholar] [CrossRef]
- Turnbull, L.; Whitchurch, C.B. Motility Assay: Twitching Motility. In Pseudomonas Methods and Protocols, Filloux, A.; Ramos, J.-L., Eds. Springer New York: New York, NY, 2014; pp 73-86, ISBN. 978-1-4939-0473-0.
- Duong-Nu, T.M.; Jeong, K.; Hong, S.H.; Puth, S.; Kim, S.Y.; Tan, W. A stealth adhesion factor contributes to Vibrio vulnificus pathogenicity: Flp pili play roles in host invasion, survival in the blood stream and resistance to complement activation. PLoS Pathog 2019, 15, e1007767. [Google Scholar] [CrossRef] [PubMed]
- de Bentzmann, S.; Aurouze, M.; Ball, G.; Filloux, A. FppA, a novel Pseudomonas aeruginosa prepilin peptidase involved in assembly of type IVb pili. J Bacteriol 2006, 188, 4851–4860. [Google Scholar] [CrossRef] [PubMed]
- Kilmury, S.L.N.; Burrows, L.L. The Pseudomonas aeruginosa PilSR Two-Component System Regulates Both Twitching and Swimming Motilities. mBio 2018, 9, e01310–01318. [Google Scholar] [CrossRef]
- Ellison, C.K.; Rusch, D.B.; Brun, Y.V. Flagellar Mutants Have Reduced Pilus Synthesis in Caulobacter crescentus. J Bacteriol 2019, 201. [Google Scholar] [CrossRef] [PubMed]
- Kühn, M.J.; Talà, L.; Inclan, Y.F.; Patino, R.; Pierrat, X.; Vos, I.; Al-Mayyah, Z.; Macmillan, H.; Negrete, J., Jr.; Engel, J.N.; et al. Mechanotaxis directs Pseudomonas aeruginosa twitching motility. Proc Natl Acad Sci U S A 2021, 118, e2101759118. [Google Scholar] [CrossRef]
- Laventie, B.J.; Jenal, U. Surface Sensing and Adaptation in Bacteria. Annu Rev Microbiol 2020, 74, 735–760. [Google Scholar] [CrossRef] [PubMed]
- Del Medico, L.; Cerletti, D.; Schachle, P.; Christen, M.; Christen, B. The type IV pilin PilA couples surface attachment and cell-cycle initiation in Caulobacter crescentus. Proc Natl Acad Sci U S A 2020, 117, 9546–9553. [Google Scholar] [CrossRef]
- Sambrook, J. , Fritsch, E. F. & Maniatis, T. Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, 1989. [Google Scholar]
- Beringer, J.E. R factor transfer in Rhizobium leguminosarum. J Gen Microbiol 1974, 84, 188–198. [Google Scholar] [CrossRef]
- Robertsen, B.K.; Aiman, P.; Darwill, A.G.; Mcneil, M.; Albersheim, P. The structure of acidic extracellular polysaccharides secreted by Rhizobium leguminosarum and Rhizobium trifolii. Plant Physiol 1981, 67, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: rapid annotations using subsystems technology. BMC Genomics 2008, 9, 75. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res 2016, 44, D457–462. [Google Scholar] [CrossRef] [PubMed]
- Olson, R.D.; Assaf, R.; Brettin, T.; Conrad, N.; Cucinell, C.; Davis, J.J.; Dempsey, D.M.; Dickerman, A.; Dietrich, E.M.; Kenyon, R.W.; et al. Introducing the Bacterial and Viral Bioinformatics Resource Center (BV-BRC): a resource combining PATRIC, IRD and ViPR. Nucleic Acids Res 2023, 51, D678–D689. [Google Scholar] [CrossRef] [PubMed]
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; Grego, T.; Pinto, B.L.; Salazar, G.A.; Bileschi, M.L.; Bork, P.; Bridge, A.; Colwell, L.; et al. InterPro in 2022. Nucleic Acids Res 2023, 51, D418–D427. [Google Scholar] [CrossRef] [PubMed]
- Simon, R.; Priefer, U.; Pühler, A. A broad host range mobilization system for in vivo genetic-engineering: transposon mutagenesis in Gram-negative bacteria. Nat Biotechnol 1983, 1, 784–791. [Google Scholar] [CrossRef]
- Schäfer, A.; Tauch, A.; Jager, W.; Kalinowski, J.; Thierbach, G.; Pühler, A. Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli plasmids pK18 and pK19: selection of defined deletions in the chromosome of Corynebacterium glutamicum. Gene 1994, 145, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Calatrava-Morales, N.; Nogales, J.; Ameztoy, K.; van Steenbergen, B.; Soto, M.J. The NtrY/NtrX System of Sinorhizobium meliloti GR4 Regulates Motility, EPS I Production, and Nitrogen Metabolism but Is Dispensable for Symbiotic Nitrogen Fixation. Mol Plant Microbe Interact 2017, 30, 566–577. [Google Scholar] [CrossRef] [PubMed]
- López-Lara, I.M.; Nogales, J.; Pech-Canul, A.; Calatrava-Morales, N.; Bernabéu-Roda, L.M.; Durán, P.; Cuéllar, V.; Olivares, J.; Alvarez, L.; Palenzuela-Bretones, D.; et al. 2-Tridecanone impacts surface-associated bacterial behaviours and hinders plant-bacteria interactions. Environ Microbiol 2018, 20, 2049–2065. [Google Scholar] [CrossRef]
- Olivares, J.; Casadesús, J.; Bedmar, E.J. Method for Testing Degree of Infectivity of Rhizobium meliloti Strains. Appl Environ Microbiol 1980, 39, 967–970. [Google Scholar] [CrossRef]
- García-Rodríguez, F.M.; Toro, N. Sinorhizobium meliloti nfe (nodulation formation efficiency) genes exhibit temporal and spatial expression patterns similar to those of genes involved in symbiotic nitrogen fixation. Mol Plant Microbe Interact 2000, 13, 583–591. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Type IVc pili genes from different Gram negative bacteria. (a) The tad/cpa/ctp loci of Aggregatibacter actinomycetemcomitans CU1000N with gene names according to Kachlany et al. (2001) [34], of Caulobacter vibrioides NA1000 with gene names according to Skerker and Shapiro (2000) [37], Christen et al. (2016) [57], and Mignolet et al. (2018) [58], and of Agrobacterium fabrum C58 with gene names according to Wang et al., 2014 [38]. (b) Rhizobial Type IVc pili gene clusters and loci of S. meliloti GR4, Sinorhizobium fredii NGR234 [51], and Bradyrhizobium diazoefficiens USDA 110 with gene names according to Mongiardini et al. (2016) [50]. In the absence of assigned names, gene identifiers are used. In the case of S. meliloti GR4, assigned names and gene identifiers for S. meliloti Rm1021 were used for short word length. Similar colors indicate similar gene products by identity or assigned function. Dark grey arrows indicate hypothetical proteins putatively associated with pili assembly and light grey arrows hypothetical proteins or proteins annotated with non-pili related functions. The asterisk indicates the position of an additional ISRm22 element in the genome of S. meliloti Rm1021. ‡ BRC ID (https://www.bv-brc.org/).
Figure 1.
Type IVc pili genes from different Gram negative bacteria. (a) The tad/cpa/ctp loci of Aggregatibacter actinomycetemcomitans CU1000N with gene names according to Kachlany et al. (2001) [34], of Caulobacter vibrioides NA1000 with gene names according to Skerker and Shapiro (2000) [37], Christen et al. (2016) [57], and Mignolet et al. (2018) [58], and of Agrobacterium fabrum C58 with gene names according to Wang et al., 2014 [38]. (b) Rhizobial Type IVc pili gene clusters and loci of S. meliloti GR4, Sinorhizobium fredii NGR234 [51], and Bradyrhizobium diazoefficiens USDA 110 with gene names according to Mongiardini et al. (2016) [50]. In the absence of assigned names, gene identifiers are used. In the case of S. meliloti GR4, assigned names and gene identifiers for S. meliloti Rm1021 were used for short word length. Similar colors indicate similar gene products by identity or assigned function. Dark grey arrows indicate hypothetical proteins putatively associated with pili assembly and light grey arrows hypothetical proteins or proteins annotated with non-pili related functions. The asterisk indicates the position of an additional ISRm22 element in the genome of S. meliloti Rm1021. ‡ BRC ID (https://www.bv-brc.org/).
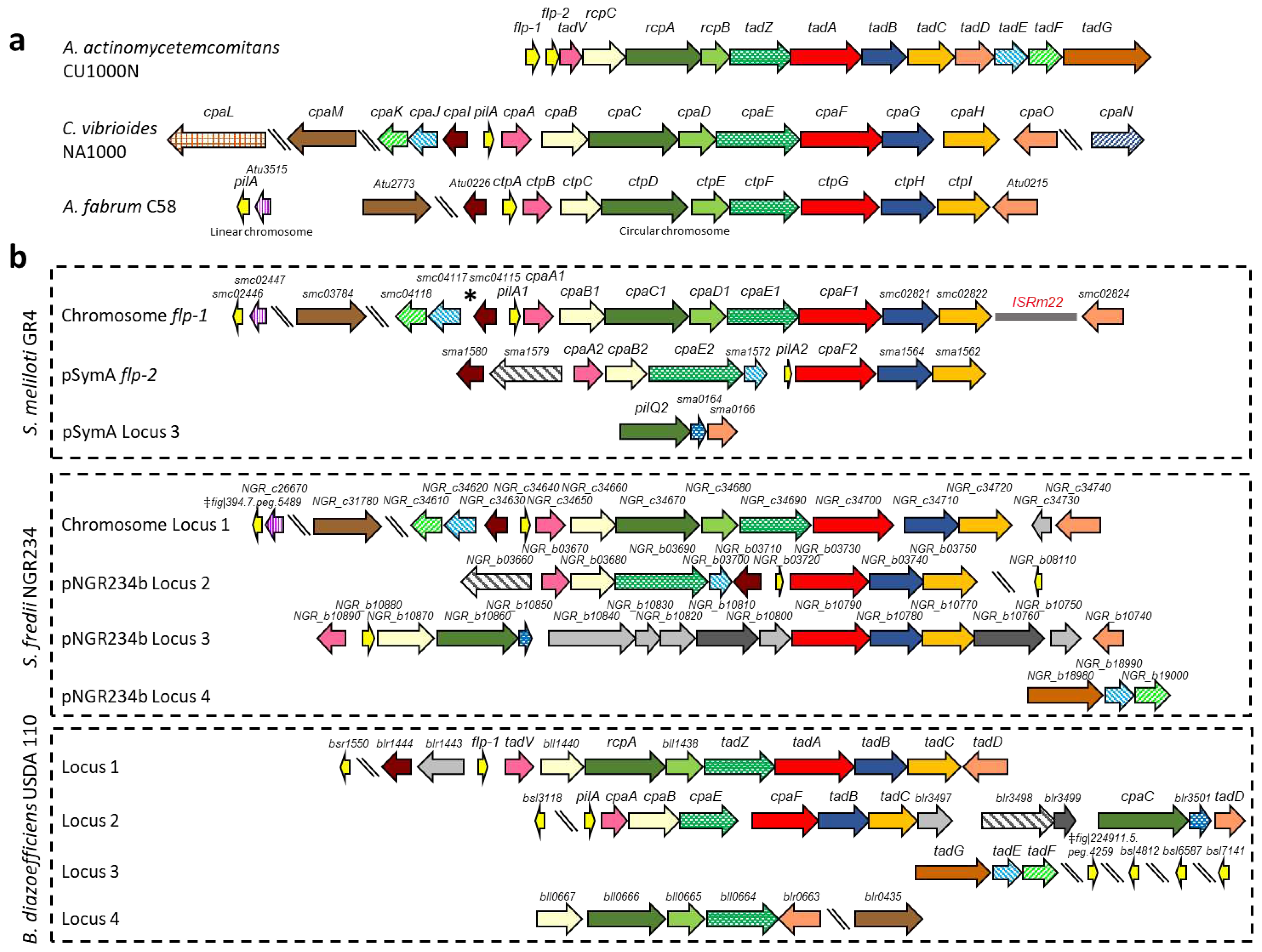
Figure 2.
Predictive protein composition and structure of the secretion systems encoded by S. meliloti tad/cpa genes of the flp-1 cluster, flp-2 cluster and Locus 3. See text for details.
Figure 2.
Predictive protein composition and structure of the secretion systems encoded by S. meliloti tad/cpa genes of the flp-1 cluster, flp-2 cluster and Locus 3. See text for details.
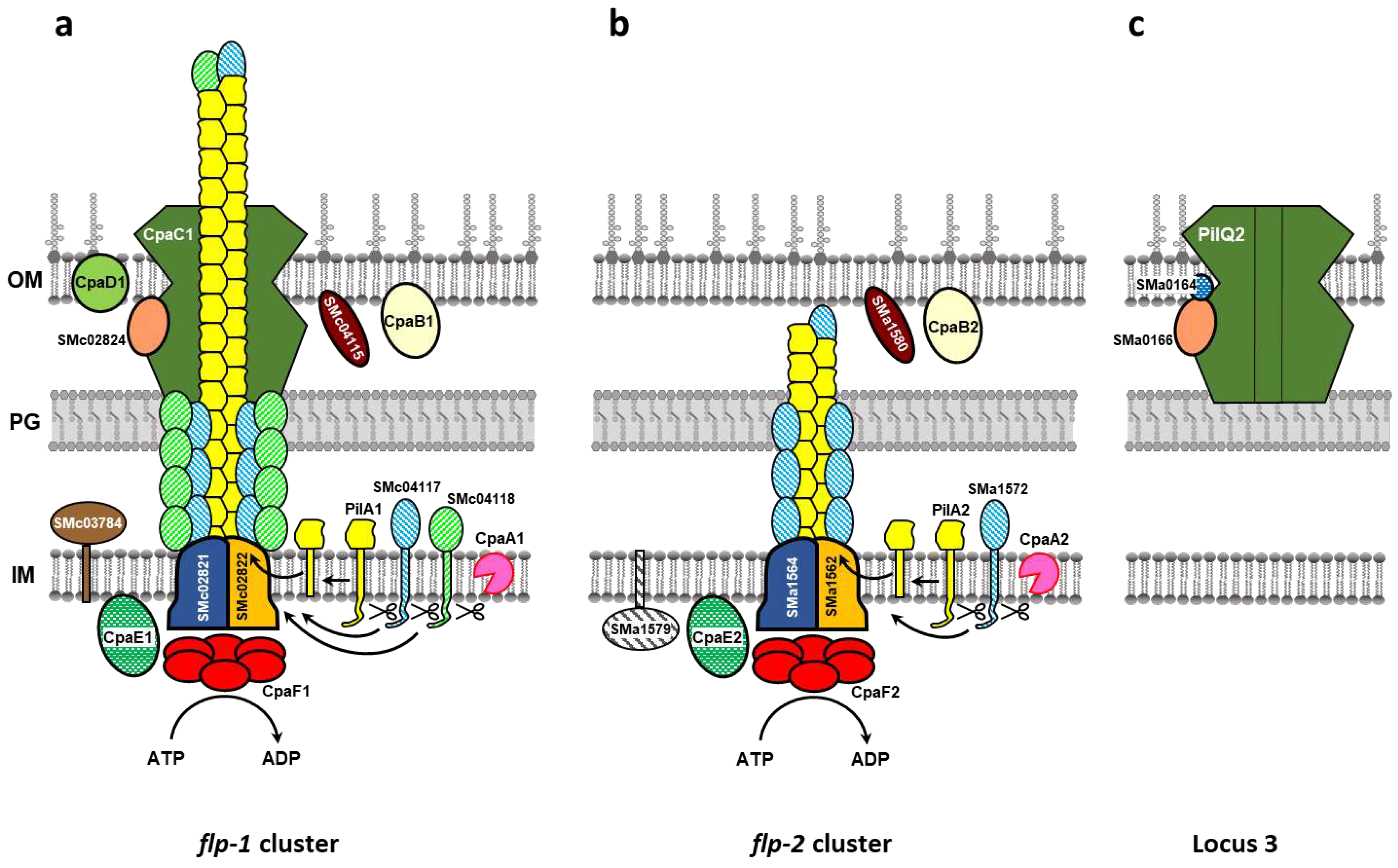
Figure 3.
Transmission electron microscopy of S. meliloti strains. Micrographs were obtained from cells grown overnight on solid MM (1%) and negatively stained with uranyl acetate (2%). Each right panel is a closer view of the red inlet indicated in the corresponding left panel. Wild-type strain GR4 (a-b) and flagellaless strains GR4flaAB (c-d), GRflaflp1 (e-f), GRflaflp2 (g-h), and GRflaflp1flp2 (i-j). Black and white arrowheads indicate flagella and Flp pili, respectively. Yellow arrowheads indicate short broken flagellar filaments characteristic of flaAflaB mutants.
Figure 3.
Transmission electron microscopy of S. meliloti strains. Micrographs were obtained from cells grown overnight on solid MM (1%) and negatively stained with uranyl acetate (2%). Each right panel is a closer view of the red inlet indicated in the corresponding left panel. Wild-type strain GR4 (a-b) and flagellaless strains GR4flaAB (c-d), GRflaflp1 (e-f), GRflaflp2 (g-h), and GRflaflp1flp2 (i-j). Black and white arrowheads indicate flagella and Flp pili, respectively. Yellow arrowheads indicate short broken flagellar filaments characteristic of flaAflaB mutants.
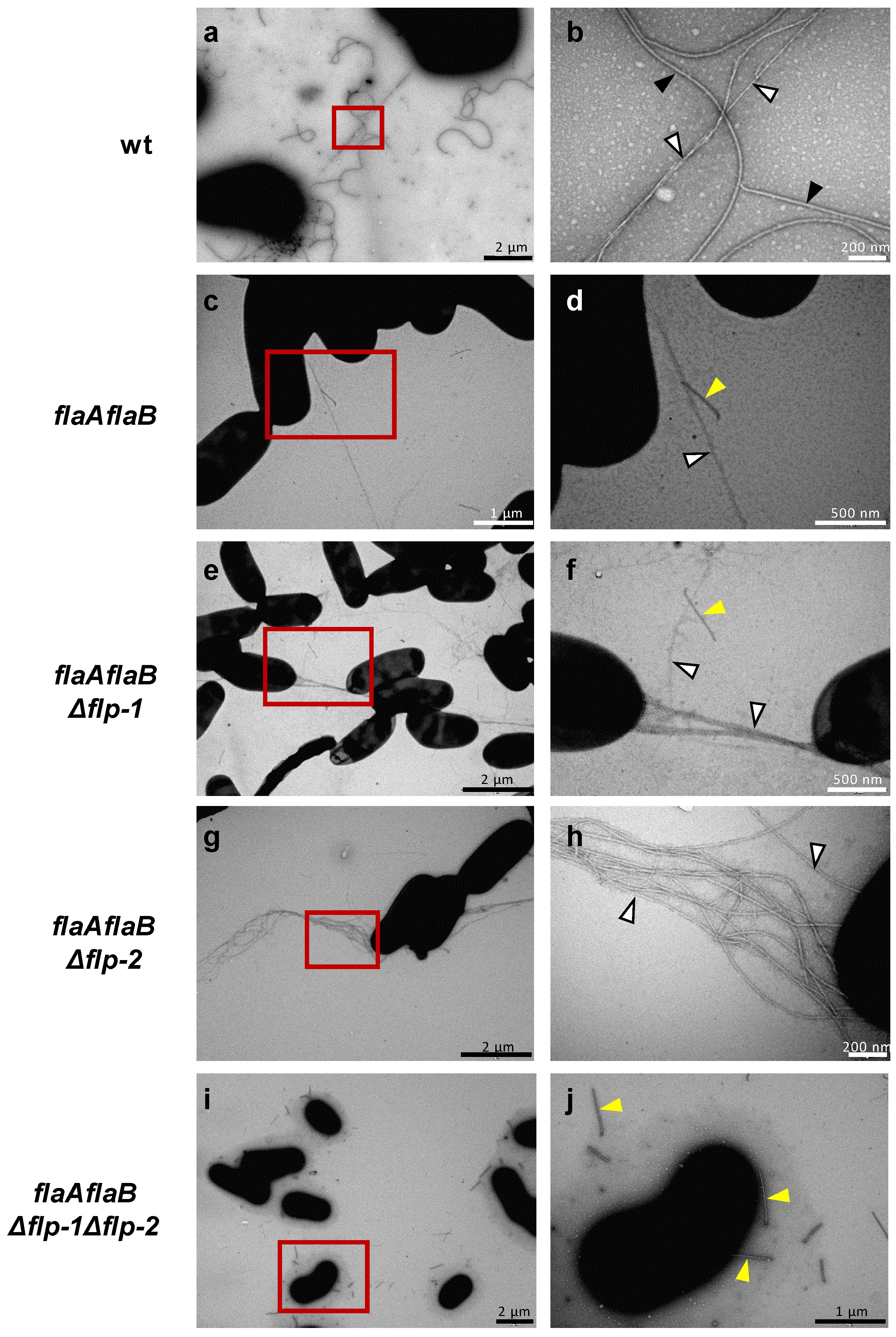
Figure 4.
Surface and swimming motilities exhibited by GR4-derivative flp deletion mutants. (a) Representative pictures and (b) graph of the surface motilities exhibited after 48 h of incubation on MM (0.6 % agar). (c) Representative picture and (d) graph of the swimming motilities in BM (0.3 % agar) after 72 h of incubation. Bars represent standard error of the mean and the different letters indicate significant differences according to an analysis-of-variance (ANOVA) test (P≤0.05).
Figure 4.
Surface and swimming motilities exhibited by GR4-derivative flp deletion mutants. (a) Representative pictures and (b) graph of the surface motilities exhibited after 48 h of incubation on MM (0.6 % agar). (c) Representative picture and (d) graph of the swimming motilities in BM (0.3 % agar) after 72 h of incubation. Bars represent standard error of the mean and the different letters indicate significant differences according to an analysis-of-variance (ANOVA) test (P≤0.05).
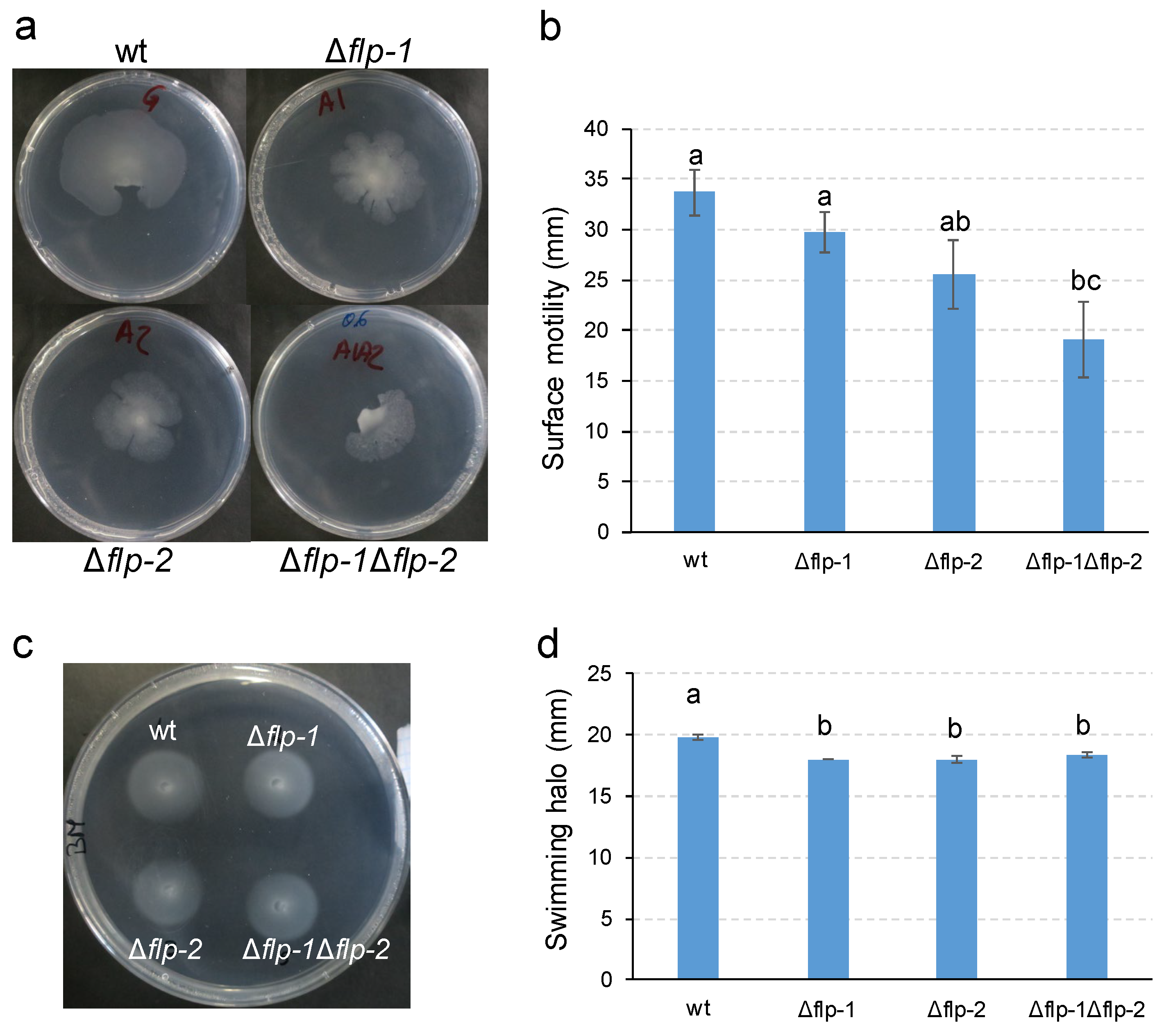
Figure 5.
Bacterial attachment to and colonization of alfalfa roots by flp deletion mutants of S. meliloti. Colony forming units (CFU) recovered from alfalfa roots were determined 2 hours after inoculation to determine bacterial adhesion, and 24, 48 and 72 h post-inoculation to assess the colonization ability of each bacterial strain. Data are expressed per gram of root fresh weight (RFW). Error bars indicate the standard error of the mean. The asterisk indicates significant difference according to an ANOVA test (P≤0.05).
Figure 5.
Bacterial attachment to and colonization of alfalfa roots by flp deletion mutants of S. meliloti. Colony forming units (CFU) recovered from alfalfa roots were determined 2 hours after inoculation to determine bacterial adhesion, and 24, 48 and 72 h post-inoculation to assess the colonization ability of each bacterial strain. Data are expressed per gram of root fresh weight (RFW). Error bars indicate the standard error of the mean. The asterisk indicates significant difference according to an ANOVA test (P≤0.05).

Figure 6.
Symbiotic phenotype of GR4-derivative flp deletion mutants. (a) Nodulation kinetics of alfalfa plants inoculated with S. meliloti wild-type GR4, or their derivative single and double flp deletion mutants. Data represent the average number of nodules per plant. The bars represent standard errors from 38 replicates in two independent experiments. Asterisks indicate significant differences compared to the wild-type according to an ANOVA test (P≤0.05). (b) Competitive nodulation assays. Data represent the percentage of nodules occupied by each rhizobial strain after co-inoculation with the marked strain GR4 (pGUS3) in a mixture ratio of 1:1. Error bars indicate the standard error of the mean obtained from three independent experiments. Different letters indicate significant differences according to an ANOVA test (P≤0.05).
Figure 6.
Symbiotic phenotype of GR4-derivative flp deletion mutants. (a) Nodulation kinetics of alfalfa plants inoculated with S. meliloti wild-type GR4, or their derivative single and double flp deletion mutants. Data represent the average number of nodules per plant. The bars represent standard errors from 38 replicates in two independent experiments. Asterisks indicate significant differences compared to the wild-type according to an ANOVA test (P≤0.05). (b) Competitive nodulation assays. Data represent the percentage of nodules occupied by each rhizobial strain after co-inoculation with the marked strain GR4 (pGUS3) in a mixture ratio of 1:1. Error bars indicate the standard error of the mean obtained from three independent experiments. Different letters indicate significant differences according to an ANOVA test (P≤0.05).
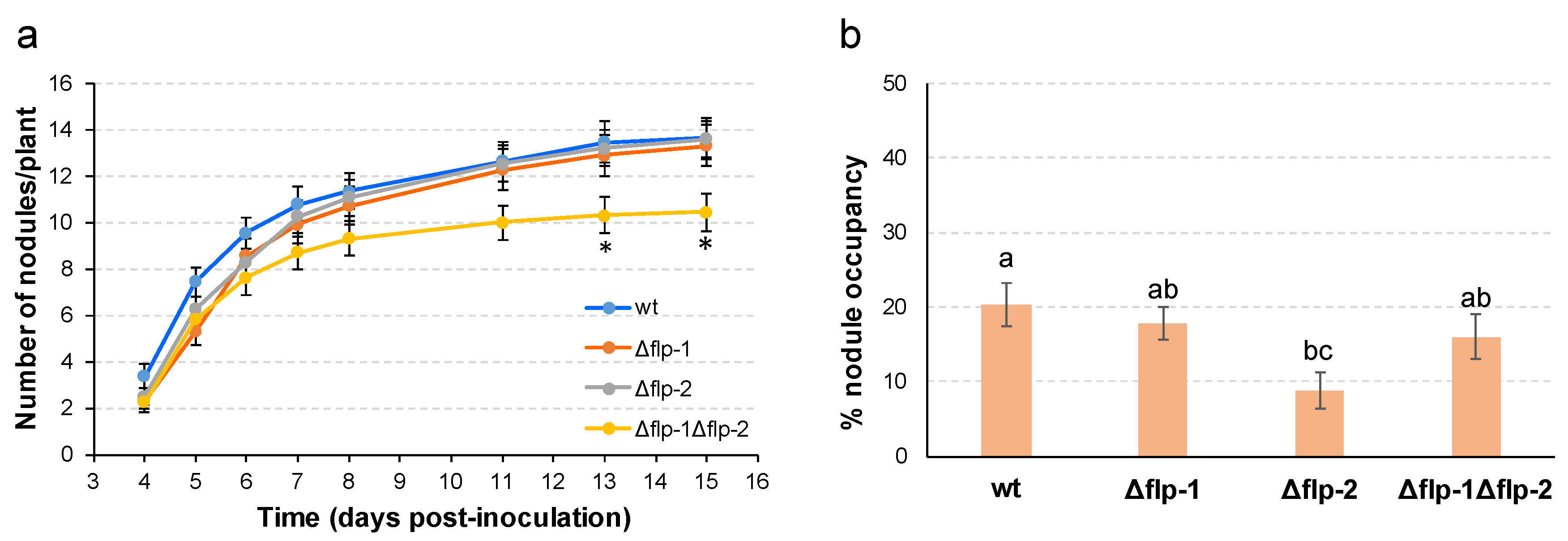

Table 1.
Percent amino acid sequence identities (in parenthesis) between the products of T4cP genes of S. meliloti GR4 and the closest orthologues of the tad genes of A. actinomycetemcomitans CU1000N (Aa), the cpa genes of C. vibrioides NA1000 (Cv), the ctp genes of A. fabrum C58 (Af), the tad/cpa genes of B. diazoefficiens USDA 110 (Bd), and the tad/cpa genes of S. fredii NGR234 (Sf).
Table 1.
Percent amino acid sequence identities (in parenthesis) between the products of T4cP genes of S. meliloti GR4 and the closest orthologues of the tad genes of A. actinomycetemcomitans CU1000N (Aa), the cpa genes of C. vibrioides NA1000 (Cv), the ctp genes of A. fabrum C58 (Af), the tad/cpa genes of B. diazoefficiens USDA 110 (Bd), and the tad/cpa genes of S. fredii NGR234 (Sf).
| Locus GR4/Rm1021 | Description | Aa | Cv | Af | Bd | Sf | |
|---|---|---|---|---|---|---|---|
| Chromosome flp-1 | |||||||
| C770_GR4Chr0141/smc04118 | CpaK-like pseudopilin | CpaK (25) | TadF (26) | NGR_c34610 (60) | |||
| C770_GR4Chr0142/smc04117 | CpaJ-like pseudopilin | CpaJ (32) | TadE (29) | NGR_c34620 (69) | |||
| C770_GR4Chr0143/smc04115 | CpaI-like putative pili related protein | CpaI (26) | Atu0226 (63) | Blr1444 (35) | NGR_c34630 (88) | ||
| C770_GR4Chr0144/smc04114 (pilA1) | Flp pilin subunit | Flp-2 (34) | PilA (51) | CtpA (76) | Bsr1550 (51) | NGR_c34640 (71) | |
| C770_GR4Chr0145/smc04113 (cpaA1) | Putative prepilin peptidase | TadV (31% in 65aa stretch) | CpaA (37) | CtpB (52) | TadV (43) | NGR_c34650 (71) | |
| C770_GR4Chr0146/smc04112 (cpaB1) | Component of the outer membrane secretin complex | CpaB (35) | CtpC (64) | Bll1440 (39) | NGR_c34660 (87) | ||
| C770_GR4Chr0147/smc04111* (cpaC1) | Component of the outer membrane secretin complex | RcpA (26) | CpaC (34) | CtpD (70) | RcpA (40) | NGR_c34670 (87) | |
| C770_GR4Chr0148/smc04110 (cpaD1) | Component of the outer membrane secretin complex | CpaD (39) | CtpE (58) | Bll1438 (35) | NGR_c34680 (80) | ||
| C770_GR4Chr0149/smc04109 (cpaE1) | Putative docking protein of the secretion system | TadZ (23) | CpaE (47) | CtpF (71) | TadZ (49) | NGR_c34690 (87) | |
| C770_GR4Chr0150/smc02820 (cpaF1) | Pilus assembly ATPase | TadA (50) | CpaF (72) | CtpG (81) | TadA (72) | NGR_c34700 (90) | |
| C770_GR4Chr0151/smc02821 | CpaG-like putative inner membrane platform protein | TadB (22) | CpaG (42) | CtpH (64) | TadB Locus 1 (48) | NGR_c34710 (89) | |
| C770_GR4Chr0152/smc02822 | CpaH-like putative inner membrane platform protein | TadC (21) | CpaH (48) | CtpI (69) | TadC Locus 1 (54) | NGR_c34720 (90) | |
| C770_GR4Chr0153/smc04116 (TRm22) | Insertion sequence ISRm22 | ||||||
| C770_GR4Chr0154/smc02824 | CpaO-like TPR containing protein | CpaO (37) | Atu0215 (66) | TadD Locus 1 (45) | NGR_c34740 (84) | ||
| Chromosomal orphan loci | |||||||
| C770_GR4Chr2732/smc02446 | Flp pilin subunit | Flp-2 (35) | PilA (60) | CtpA (51) | Bsl4812 (51) | ‡fig|394.7.peg.5489 (60) | |
| C770_GR4Chr2733/smc02447 | Hypothetical protein | Atu3515 (52) | NGR_c26670 (95) | ||||
| C770_GR4Chr3249/smc03784 | CpaM-like polysaccharide deacetylase | CpaM (39) | Atu2773 (59) | Blr0435 (38) | NGR_c31780 (74) | ||
| pSymA flp-2 | |||||||
| C770_GR4pC0512/sma1580* | CpaI-like putative pili related protein | CpaI (32) | Atu0226 (31) | Blr1444 (37) | NGR_b03710 (59) | ||
| C770_GR4pC0513/sma1579* | Putative pili related protein | Blr3498 (24) | NGR_b03660 (74) | ||||
| C770_GR4pC0514/sma1578* (cpaA2) | Putative prepilin peptidase | TadV (30) | CpaA (38) | CtpB (36) | TadV (43) | NGR_b03670 (71) | |
| C770_GR4pC0515/sma1576* (cpaB2) | Component of the outer membrane secretin complex | CpaB (34) | CtpC (39) | Bll1440 (40) | NGR_b03680 (78) | ||
| C770_GR4pC0516/sma1573 (cpaE2) | Putative docking protein of the secretion system | CpaE (43) | CtpF (40) | TadZ (40) | NGR_b03690 (76) | ||
| C770_GR4pC0517/sma1572 | CpaJ-like pseudopilin protein | CpaJ (41) | NGR_b03700 (75) | ||||
| C770_GR4pC0518/sma1570 (pilA2) | Flp pilin subunit | Flp-1 (34) | PilA (53) | CtpA (55) | Flp-1 (62) | NGR_b03720 (69) | |
| C770_GR4pC0519/sma1568* (cpaF2) | Pilus assembly ATPase | TadA (48) | CpaF (73) | CtpG (69) | TadA (79) | NGR_b03730 (87) | |
| C770_GR4pC0520/sma1564 | CpaG-like putative inner membrane platform protein | TadB (28) | CpaG (43) | CtpH (41) | TadB Locus 1 (46) | NGR_b03740 (71) | |
| C770_GR4pC0521/sma1562 | CpaH-like putative inner membrane platform protein | TadC (20) | CpaH (43) | CtpI (49) | TadC Locus 1 (52) | NGR_b03750 (84) | |
| pSymA Locus 3 | |||||||
| C770_GR4pC1351/sma0163 (pilQ2) | RcpA-like putative component of the outer membrane secretin complex | RcpA (28) | CpaC (31) | CtpD (31) | CpaC (40) | NGR_b10860 (40) | |
| C770_GR4pC1350/sma0164 | Hypothetical lipoprotein | Blr3501 (33) | NGR_b10850 (29) | ||||
| C770_GR4pC1349/sma0166 | CpaO/TadD-like TPR containing protein | Atu0215 (31% in 70 aa middle stretch) | TadD Locus 2 (44) | NGR_b10740 (36) |
* Sequence identity of 98.4-99.8% at the amino acid level between GR4 and Rm1021 gene products otherwise 100% identical. ‡ BRC ID (https://www.bv-brc.org/).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



