Submitted:
03 January 2024
Posted:
04 January 2024
You are already at the latest version
Abstract
Expansins (EXPs) which are the essential components of plant cell wall function as relaxation factors to directly promote turgor-driven expansion of cell wall and control plant growth and development and diverse environmental stress responses. A lot of EXPs genes have been widely identified and characterized in numerous plant species, but not in sweetpotato. In the present study, a total of 59 EXPs genes that unevenly distributed onto 14 of 15 chromosomes were identified from sweetpotato genome, and segmental and tandem duplications were exhibited to act as dominant functions in IbEXPs diversity. Phylogenetic analysis exhibited that IbEXPs members were clustered into four subfamilies according to EXPs in Arabidopsis and rice and the regularity of motif, domain and gene structures was in accordance with the subfamily classification. The collinearity analysis between IbEXPs and related homologous sequences in 9 plants provided the phylogenetic insights of EXPs gene family. Cis-element analysis futhrer revealed the potential roles of IbEXPs in sweetpotato development and stress responses. Evidence from RNA-seq and qRT-PCR analysis of eight selected IbEXPs genes displayed their various tissue specificity in different tissues, and their transcripts were varyingly induced or suppressed under multiple hormone treatments (ABA, SA, JA and ACC) and abiotic stresses (low and high temperature). These results supply worthy foundations to further comprehensively investigate the exact functions of IbEXPs genes, and multiple members may function as promising regulators to control plant development and enhance the stress resistance of transgenic plants.
Keywords:
Expansin gene family
; Systematic characterization
; Expression analysis
; Tuberous root development
; Abiotic stress
; Hormones
; Sweetpotato
1. Introduction
Among the variousplant cell components, the cell wall which is a kind of distinct, crucial and dynamic structure has the good extensibility. It determines and maintains the cell size and shape and serves as the protective barrier [1,2,3]. The plant cell wall is highly complex structures constituted by various polysaccharides that vary in abundance, function, and structure [4]. The cell wall plays crucial roles in supplies of stiffness and mechanical support to plant body, resistance to abiotic and biotic stresses, conduction of nutrients and water, determination of plant architecture and morphogenesis [5]. Nowdays, the study on cell wall extension mechanism has become the research priority due to the significance of cell wall enlargement during plant morphogenesis [6]. The increase of cell volume and quantity which hinges on the cell wall enlargement and loosening is crucial for plant growth [7]. Cell wall loosening is the significant precondition of cell wall remodeling, in which the physical structure is altered or new components are added into the cell wall, inducing alteration of shape and anisotropic growth in the cell [7]. The modified proteins which attach to the cell wall serve as the vital roles in cell wall enlargement and loosening, the most widely recognized of which are the expansins (EXPs) [8].
EXPs are the crucial proteins related to cell wall that participate in cell wall enlargement and loosening and are commonly found in plants [9]. As the primary factor of enlargement and loosening, EXPs can control the cell relaxation without any chemical energy through the non-enzymatic activity [7]. Expansin can act directly on plant cell wall to loosen the cell wall via binding to the cellulose in cell wall to disrupt hydrogen bonds which exist in wall matrix polysaccharides and cellulose microfibrils in the pH-dependent manner, and also take part in the decomposition, remodeling, extension and assembly of cell wall [7,9,10,11]. Plant expansins usually are composed of a signal peptide (SP) at the N-terminal (about 20-30 amino acid residues) and two domains. Domain I is a six-stranded double-psi beta-barrel (DPBB) and locates at the N-terminal. It shares the homologous with the catalytic domain of GH45 proteins (glycoside hydrolase family 45) and harbors a conserved His-Phe-Asp (HFD), but do not has the β-1, 4-glucanase activity [12]. This region is rich in cysteine (Cys) residues with one characteristic catalytic domain which may have something to do with the disulfide bond formation [13]. The Domain II which harbors a β-sandwich fold and shares about 50% similarity with the group-II pollen allergen protein (pollen_allerg_1, G2A family) is considered as the polysaccharide binding domain due to it cantains the conserved aromatic amino acids and polar tryptophan residues on its surface. It cantains 90-120 amino acid residues and was classified as the family-63 carbohydrate binding module (CBM63) [7,14].
According to the standardized nomenclature and phylogenetic analysis, plant EXPs proteins are divided into 4 subfamilies: EXPA (α-expansin), EXPB (β-expansin), EXLA (expansin-like A) and EXLB (expansin-like B) [15,16]. Nowadays, numerous EXPs members of this four subfamilies have been identified in numerous plants. Among them, the roles of EXPA and EXPB have been widely studied, which show the wall-loosening activities to participate in cell expansion and plant developmental processes [15,16]. While members of EXLA and EXLB mainly play functions in stress response, hypocotyl length and root architecture [17,18,19]. Based on the previous investigations, EXPs are regarded as the main determinant of cell shape in many cell developmental processes, especially in the regulation of cell-wall extensibility [20,21,22], including elongation and expansion [23]. Since the first identification in cucumber hypocotyl [24], EXPs proteins have been commonly found in numerous plant species.
It has been widely shown that EXPs play crucial functions in multiple biological processes by cell-wall modification and elongation, such as biotic and abiotic stress, the root and fiber development, root nodule formation, fruit development and ripening, and other developmental processes [7,10,20,22]. For instance, overexpression of Osmanthus fragrans OfEXLA1 gene increased the resistance to salt and drought stress in Arabidopsis [25]. Ectopic ovexpression of wild Arachis AdEXLB8 in tobacco increased tolerance to biotic (Meloidogyne incognita and Sclerotinia sclerotiorum) and abiotic (drought) stresses [26]. The wheat TaEXPA2 can significantly elevate the resistance of transgenic plants to Cd toxicity and multiple abiotic stresses (drought, oxidative and salt) [27,28,29,30]. Ectopic expression of poplar PttEXPA8 in tobacco enhanced the heat resistance in transgenic plants [31]. Three expansin genes, Tomato SlExp1, apple MdEXLB1, mango MiExpA1 have been identified as the crucial determinants during fruit softening and ripening [32,33,34,35]. Two β-Expansin Gene GmINS1 and GmEXPB2 play significant roles in nodule formation and development [8,36]. The rice OsEXPB2 and OsEXPA8, Soybean GmEXLB1 and GmEXPB2 function as the important regulators in root system architecture [18,36,37,38,39]. Stylosanthes SgEXPB1, rice OsEXPA10, and Arabidopsis AtEXPA7 are required for root development [6,40,41]. Upregulation of GhEXPA8 or GbEXPATR can increase the fibre length in cotton, while reduced EXPA results in shorter fibre [42]. In addition, plant expansin genes also play vital roles in height and leaf growth [43,44], pollen tube and stem elongation [45,46,47], seed development, germination and yield [48,49,50], flower development [51], and so on.
As the seventh most important food crop, sweetpotato (Ipomoea batatas) is the only crop that generates starch storage roots among plant species in the Convolvulaceae [52,53]. It has been one of the most widely cultivated food crops worldside due to its numerous advantages, such as low input requirements, strong stress resistance, wide adaptability, high yield and starch content, and make it a significant food crop in the world [54]. At present, sweetpotato has been broadly used in alcohol and starch production, animal feed, starch processing, and human food. And it also ensures the food security in a lot of developing countries on account of its adaptation in various environmental conditions [53]. In previous study, 37 EXP genes were identified from Ipomoea trifida, which is the most possible diploid wild relative species of sweetpotato [55]. However, the Ipomoea trifida genome does not adequately represent the whole sweetpotato genome information. Recently, the completion of hexaploid sweetpotato genome sequencing provides sufficient and valuable information for the identification and characterization of gene families [56], whereas the genome-wide identification of sweetpotato EXP gene family is still lagging and so it is necessary for us to comprehensively identify and characterize the EXP gene in sweetpotato.
In the current stage of cropmolecular breeding, one of the main focuses is to improve environmental stress resistance and promote plant growth. Related investigations have been performed systematically in a variety of plant species due to the compatiblity between the crucial functions of EXPs genes and the demand of breeding. Nowadays, a large number of EXPs genes have been broadly identified from diverse plants, for instance, a total number of EXPs genes have been identified in monocots, such as 92, 58, 241, 88, 46, 38 genes in sugarcane [4], rice [15], common wheat [57], maize [58], barley [59], Brachypodium distachyon [60], respectively, and in dicotyledons, such as 36, 75, 46, 93, 52 genes were found in Arabidopsis [15], soybean [61], gingkgo [62], cotton [63], tobacco [64], respectively. The genome sequencing of hexaploid sweetpotato (Taizhong6) has been completed [56], however, no systematical identification and characterization on EXPs genes in sweetpotato (Ipomoea batatas L.) is available. The identification of molecular feature of significant EXPs gene family will contribute to further understand the regulatory mechanism of plant development and adaptation to environmental stresses. In this study, 59 EXPs genes which were divided into four subfamilies (36 IbEXPAs, 10 IbEXPBs, 2 IbEXLAs, 11 IbEXLBs,) were indentified from sweetpotato genome. To standardize the nomenclature of EXPs proteins in sweetpotato and evaluate their possible functions and relationships in development and stress responses, the comprehensively systematical characterization of 59 identified sweetpotato EXPs genes was conducted. The phylogenetic relationship, chromosomal location, conserved motif and domain, gene structure, molecular characterization, cis-element, gene duplication and the expression pattern analysis of IbEXPs in different tissues and various treatments of hormones and abiotic stresses were investigated in this study. All data obtained in this study will lay the worthy foundation for the further screening and functional investigation of valuable EXPs genes that play crucial roles in tuberous root development and stress tolerance in sweetpotato.
2. Results
2.1. IbEXP gene identification and characterization in sweetpotato
In present study, a total number of 59 IbEXP genes were identified from sweetpotato genome and were named following the AtEXPs and OsEXPs classification in Arabidopsis and and rice their position on chromosome. These 59 IbEXP proteins were divided into four subfamilies, namely IbEXPA, IbEXPB, IbEXLA and IbEXLB, with 36, 10, 2 and 11 members, respectively. After that, the protein size (aa), theoretical isoelectric point (pI), molecular weight (Mw) and phosphorylation site of 59 IbEXP proteins were explored. The detailed data were shown in Table 1. The protein length and Mw of IbEXPs varied widely, with the length of proteins varied from 183 aa (IbEXPA31) to 670 aa (IbEXPA19) and the Mw ranging from 20.1739 KD (IbEXPB31) to 74.052 KD (IbEXPA19). The PI ranged from 4.63 (IbEXLB11) to 9.82 (IbEXPA34). The subcellular location prediction exhibited that most of IbEXPs were located on cell wall, and very few IbEXPs were located in nucleus (IbEXPA19), and simultaneously located in chloroplast and on cell wall (IbEXPA22 and IbEXPA33). The results of phosphorylation site prediction of IbEXPs displayed that significant changes from 24 (IbEXPA17 and IbEXPA31) to 109 (IbEXPA12), and the vast majority of IbEXPs harbored more Ser sites than that of Tyr and Thr sites. Moreover, over 83.05% of IbEXPs have at least 30 phosphorylation sites.
2.2. Phylogenetic relationship of IbEXPs in sweetpotato
To explore the evolutionary relationships of IbEXPs in sweetpotato, phylogenetic analysis was carried out using the protein sequences of 59 IbEXPs, 36 Arabidopsis AtEXPs and 58 rice OsEXPs (Supplementary file S1, Table S1). Then the unrooted phylogenetic tree was constructed using the neighbor-joining bootstrap method by the MEGA software (version 11.0). Phylogenetic tree analysis exhibited that 59 IbEXPs were divided into four subfamilies according to the topology of the tree, clades support values, and reported studies about the EXPs classification in Arabidopsis and rice [15] (Figure 1), and the sizes of every subfamily vary greatly. The number of IbEXP members contained in these four subfamilies ranged from 2-36, with the EXPA subfamily containing the most IbEXPs (36) and the EXLA subfamily with only two members of IbEXPs, which are consistent with that in Arabidopsis and rice. The member differences among IbEXPs, AtEXPs and OsEXPs divided into in the same subfamily indicated that apparent interspecific divergences of EXP gene family exist among sweetpatato, Arabidopsis and rice.
2.3. Chromosome localization of sweetpatato IbEXP genes
Chromosome distribution analysis based on sweetpatato GFF3 genome annotations exhibited that 59 IbEXPs genes were located on 14 chromosomes of sweetpatato and no IbEXP genes were found on LG 9. In general, IbEXP genes are unevenly distributed on the 14 chromosomes, which may be the results of uneven gene replication of chromosome fragments. Among them, LG 14 contained the most IbEXP genes (11), and the LG 7 contained the second number of IbEXP genes (9), while LG 11 had only one IbEXP gene. Furthermore, there were eight on LG 4, five on LG 5, and two to four on other Chrs (Figure 2). These results suggested that IbEXPs distribution had high variable densities and was disproportionate to the length of chromosome. For instance, the largest chromosome (LG 11) contained only one IbEXP gene, while the smallest chromosome (LG 10) contains three IbEXP genes.
2.4. Collinearity analysis of sweetpatato IbEXP proteins
In plants, genome duplication facilitate the expansions and evolutions of gene families [65]. In order to explore the potential gene duplications amone all 59 IbEXP genes, the collinearity analysis of IbEXPs was performed using the MCScanX and BlastP programs. The results exhibited that 7 gene pairs with tandem duplication among IbEXP genes were indentified, including IbEXPA2-IbEXPA3, IbEXPB2-IbEXPB3, IbEXLB8-IbEXLB9, IbEXLB9-IbEXLB10, IbEXPA29-IbEXPA30, IbEXPA30-IbEXPA31, IbEXPA32-IbEXPA33 (Figure 2, Table S2-1). And these IbEXP genes exhibiting tandem duplications belonged to the same subfamily. In addition, MCScanX and BlastP programs were also carried out to identify the fragment duplications and 3 gene pairs of only EXPA subfamily were found on only 5 (LG1-3, LG5, LG7) of 15 chromosomes as follows: IbEXPA2-IbEXPA16, IbEXPA4-IbEXPA6, IbEXPA14-IbEXPA17 (Figure 3, Table S2-2). However, there were no more fragment duplications of IbEXPA genes were found on other then chromosomes (LG4, LG6, LG8-15) and no fragment duplications were detected in other three subfamilies. These segmental duplications occured only between genes in the EXPA subfamily may be one of the reason that the number of EXPA members was larger than that of other three subfamilies and also indicate the functions of EXPA genes in regulating plant development and response to stress were more significant than that of genes in other three subfamilies. In brief, these results suggest that gene duplication are conducive to the expansion of sweetpatato IbEXP gene family.
2.5. Collinearity analysis of EXP genes between sweetpatato and other plants
To further investigate the origin and evolutionary relationships of sweetpatato IbEXP genes, collinearity relationships among IbEXPs and orthologous genes in nine representative plants was explored, including Ipomoea triloba (the likely diploid wild relative of sweetpotato), two cereal plants, two representative model plants, two Solanaceae plants, and two Brassica plants. The results exhibited that a total of 30 (50.8%) IbEXPs shared the collinear relationships with that in Ipomoea triloba, followed with Solanum lycopersicum (13), Arabidopsis (5), Capsicum annuum (4), Brassica oleracea (3), Brassica rapa (2), while there were no collinear relationships of IbEXPs with that in Triticum aestivum, Oryza sativa and Zea mays (Figure 4, Table S3-1/-2/-3). Therefore, the largest number of collinearity relationships exist between sweetpatato IbEXPs and Ipomoea triloba genes suggesting a closer relation between these two plants. Moreover, these data also showed that multiple IbEXPs had the collinear relationships with two genes in other plant species, particularly with Ipomoea triloba. Analogously, two IbEXPs genes shared the collinearity with the same one gene of detected four plants (Ipomoea triloba, Arabidopsis thaliana, Brassica oleracea, and Solanum lycopersicum) were also found (Table S3-1/-2/-3). These data indicated that a number of orthologous genes might originate from the common ancestor in these plants.
2.6. Motif, conserved domain and gene structure analysis of IbEXPs
To further evaluate the sequence characteristics of sweetpatato IbEXPs, the investigation of conserved motif composition was performed by the MEME tool. The results exhibited that a total number of 20 distinct motifs were detected from IbEXPs proteins that are based upon the settings of Arabidopsis and rice [15]. The results showed that the IbEXPs belonging to the same subfamilies generally harbored the similar motif compositions, and these results further supported our subfamily classification (Figure 5 A-B). Motifs 2, 5 and 11 widely existed in the most IbEXPs, and other motifs distributed in certain IbEXP proteins. Universally, multiple motifs almost existed in all IbEXP members in the same subfamily and there were some composition differences of motifs among different subfamilies. For example, almost all members of EXPA, EXPB, EXLA, EXLB subfamilies harbored motifs 1, 2, 3, 4, 5, 6, 7, 8, 11, 20, motifs 2, 3, 5, 7, 10, 11, 16, motifs 2, 6, 9, 10, 11, 13, motifs 2, 5, 6, 11, 13, 15, respectively. And the results also displayed that some IbEXPs in the same subfamily contained specific motifs except the common motifs they had, such as motif 4, 8, 17 and 18 in EXPA, motif 14, 16 and 19 in EXPB. These results suggested that the compositions and number vary observably in these four subfamilies, and the existence of these specific motifs may suggest that sweetpatato IbEXPs had distinct and diverse functions.
To expore the sequence diversity of IbEXPs genes, the exon-intron compositions and conserved domain in each IbEXPs genes were examined. Conserved domain examination using Batch CD-Search showed that five domains with one pollen_allerg_1 domain and four typical EXP domains were examined from all IbEXPs (Figure 5 C). In the same subfamilies, most IbEXPs contain the same EXP domain and the pollen_allerg_1 domain, and the EXP domain located on the similar position with few exceptions. These data indicated that the EXP domain is the most valuable information construct distinctly the phylogenetic relationships among IbEXPs members. Gene structure detection displayed that the exon numbers of each IbEXPs gene varied from 1-14, with one IbEXPs genes containing no introns and four IbEXPs genes only having 1 intron (Figure 5 D). IbEXPA19 harbored the most exon (14), following by IbEXPA22 and IbEXPB3 with 11 exons. Moreover, this result displayed that most of IbEXPs genes generally harbored similar gene structures and exhibited similar exon length. A certain difference of intron numbers among IbEXPs genes in the same subfamily were also found, which may associated with the function diversity of IbEXPs genes. All these results obtained here demonstrated that the phylogenetic relationships of IbEXPs were mainly related to their conserved EXP domains and gene structures.
2.7. Cis-element prediction in IbEXPs promoter regions
The cis-elements which locate in gene promoter region are the non-coding sequences. They are also vital for gene expression and widely regulate numerous biological processes [66]. To expore the possible regulatory mechanism of IbEXPs genes in controlling plant growth and response to stresses and hormones, the 2000bp sequences upstream of the start codon ATG of each IbEXPs gene was used to detect the cis-elements via the PlantCARE database. The results showed that a total of 742 cis-elements were found in the pormoter regions of IbEXPs genes (Table S4-1). All these cis-elements were associated with 19 types of biological processes (Figure 6 A, Table S4-2). Among them, IbEXLB5 (g15300.t1) contained the largest number (31) of cis-elements. The cis-element numbers in each subfamily varies greatly (Figure 6 B) and all detected cis-elements in this study can be classified into three categories (Figure 6C):
The first category relates to the hormone responses (404), including the salicylic acid responsive element (21, 2.83%), MeJA responsive element (90, 12.13%), gibberellin responsive element (34, 4.58%), ethylene responsive element (59, 7.95%), auxin responsive element (40, 5.39%), and abscisic acid responsive element (160, 21.56%). All subfamilies of IbEXPs genes abundantly contain the abscisic acid responsive elements (G-Box and ABRE), gibberellin responsive elements (GARE, P-box, TATC-box and CARE) and ethylene responsive element (ERE) in their promoter regions. Members in EXPA subfamily contained the maximum number of abscisic acid responsive elements, auxin responsive elements, MeJA responsive element, ethylene responsive elements and salicylic acid responsive elements compared with other three subfamilies, while members in EXLA harbored the smallest number of these five hormone related elements. It is worth noting that only one gibberellin responsive element were found in promoter region of EXLA members. Among these five hormone responsive element, the number (43 IbEXPs genes) of IbEXPs genes that contained the abscisic acid responsive element was the largest, followed by ethylene responsive element (34 IbEXPs genes). Besides, some IbEXPs genes, such as IbEXPA2/8/9/17/23/24/26/34/36, IbEXPB3/4, IbEXLA1 and IbEXLB6/10/11 had multiple common hormone responsive elements, manifesting the possibility that these members had more rapid intense and response to certain hormones. Simultaneously, IbEXPA1/2/8/911/17/24/25/27/31/37/39, IbEXPB3/7/8/12 and IbEXLB1/2/3/9/10 harbored diverse hormone responsive elements manifesting their potential roles in many networks of hormone regulation.
The second category relates to growth and development (123), including cell cycle regulatory element (3, 0.4%), circadian control element (11, 1.48%), endosperm expression element (11, 1.48%), flavonoid biosynthetic gene element (2, 0.26%), growth and development related element (48, 6.47%), meristem expression element (34, 4.58%), seed specific regulatory element (7, 0.94%), palisade mesophyll cell differentiation element (7, 0.94%). As shown in Figure 6B,D, some cis-elements were absent from certain subfamily, such as endosperm expression element, cell cycle regulatory element, flavonoid biosynthetic gene element, growth and development related element, meristem expression element and seed specific regulatory element. Cis-elements related to meristem expression had the largest number of IbEXPs (26), followed by growth and development related element (12 IbEXPs genes). The EXPA subfamily contained all these eight elements and had the maximum number of cis-elements that related to endosperm expression, growth and developmen and meristem expression compared with other three subfamilies. These results indicated that members of the EXPA subfamily may be the major regulator during plant growth and development.
The third category relates to stress responsiveness (214), including defense and stress responsive element (26, 3.5%), drought inducibility, anthocyanin pathway element (70, 9.43%), drought responsive element (99, 13.34%), low temperature responsive element (18, 2.43%) and wound responsive element (1, 0.13%). Similar to the two categories above, the EXPA subfamily also contained the largest number of cis-elements that related to drought inducibility, defense and stress responsive, anthocyanin pathway, drought responsive and low temperature responsive. The wound responsive element was absent from EXPA, EXLA and EXLB subfamily, and only one of this element existed in EXPB. Members in the EXPA subfamily contained the maximum number of drought responsive element, indicating that the EXPA members may be the major regulator in drought responsive. Moreover, the drought responsive element also had the largest number of IbEXPs genes, suggesting the major roles of of IbEXPs genes may mainly in drought responsive.
In short, the number and composition of cis-elements in promoter sequences of different IbEXPs genes exhibited great diversity in and among subfamilies. These results suggeseted that the gene expression levels of IbEXPs in sweetpotato are controled by diverse cis-elements in connection with growth and development, hormones and stress response.
2.8. Transcriptome-wide identification of IbEXPs genes associated with tuberous root development and their expression profiles in different tissues
Increasing evidence suggested that EXP genes played critical and various roles in different developmental processes, such as root growth and architecture, fruit softening and ripening, seed production, nodule formation and development. To investigate the potential biological roles of IbEXPs genes in tuberous root formation- and development, their transcript levels were explored in different developmental stages (FR, DR and MR) based on our previous transcriptome data [67]. We found that more than 20 of the 59 identified IbEXPs genes were significantly expressed in these three periods. To verify our transcriptome data, the expression patterns of eight selected IbEXPs genes which showed distinct expression changes in our RNA-seq data were analyzed in different periods of tuberous roots and other tissues by qRT-PCR assay. The results displayed that the transcript abundances of these IbEXPs genes showed varying degrees in different tissues (Figure 7). The expression levels of IbEXPA4, IbEXPA17, IbEXPA25, IbEXPB10 and IbEXLA1 were increased during tuberous root development (DR1, DR2, DR3 and MR), and were significantly higher in MR than other three stages (DR1, DR2 and DR3). Moreover, IbEXPA17, IbEXPA25, IbEXPB5, IbEXLA1 and IbEXLA2 were also highly expression in leaves. In the ealy stage DR1, the expression of IbEXLA2 was lower, and then was dramatically increased at DR2. After that, its transcripts were induced with the development of tuberous root. The expression of IbEXLB11 was down-regulated with the development of tuberous root development and was significantly accumulated in stems. The expression of IbEXPB5 were increased following tuberous root development and showed the highest expression levels in DR3 and finally were reduced in MR. While the expression levels of IbEXLA1 in stems and leaves and IbEXLB11 in stems were significantly higher than that in different stages of tuberous roots. These results suggested that these selected EXP genes might play signifcant roles in different tissues or developmental stages of sweetpotato.
2.9. Expression pattern analysisis of IbEXPs genes under multiple hormone treatment and abiotic stresses
In addition to the crucial functions in plant development, EXPs were also verified to participate in response to multiple abiotic stresses and exogenous plant hormones [27,37,68]. Thus, the transcript accumulation of selected eight IbEXPs genes under the treatments of stress and plant hormones were investigated. The results showed the expression levels of these IbEXPs were enhanced or decreased to varying degrees under different hormone treatments (Figure 8). Wherein, the transcriptional levels of IbEXPA4/17/25, IbEXPB5/10, IbEXLA1, IbEXLB11 can be increased to diverse degrees by these four hormones (ACC, ABA, JA and SA). On the contrary, the expression levels of IbEXLA2 were reduced under these four hormones. The transcripts of IbEXPB5/10, and IbEXLA1 displayed higher fold changes with 6.6-7.9 fold compared to 0h data by ABA treatment, while other four IbEXPs genes (IbEXPA4/17/25, IbEXLB11) exhibited lower fold changes with less than 2 fold. The expression levels of IbEXPA4/17/25, IbEXPB5 and IbEXLB11 were up-regulated about 2.9-26.3 fold compared with that in 0h example under the ACC treatment. After being treated by JA, the transcript levels of IbEXPA17/25, IbEXPB5, IbEXLA1 and IbEXLB11 were induced by 2.9-13.8 fold at certain time points. While being treated by SA, only IbEXLB11 displayed the increased 2 fold changes of transcript abundances, the transcripts of other seven EXP genes were decreased to varying degrees at all or some time points.
In consideration of the crucial functions of IbEXPs genes in various abiotic stresses reported in previous studies, we also performed the expression pattern analysis of selected IbEXPs genes under low and high temperature according to our previous study [69]. The results displayed that transcripts of selected eight IbEXPs increased or reduced to different degrees (Figure 9). The transcripts of IbEXPA4/25, IbEXPB10, IbEXLA1 and IbEXLA2 were significantly reduced at all time points. The transcripts of IbEXPA17 were markedly reduced at all or some time points under low temperature stress, while its expression were increased about 2.3 fold at 48h. On the contrary, the expression levels of IbEXPB5 were dramatically reduced at all or some time points under high temperature stress, but were observably increased about 4.1-9 fold at 24h and 48h. These results suggested that many IbEXPs genes may function as crucial regulators in response to some hormone (especially ABA, ACC and JA), multiple abiotic stresses and/or signal transduction.
3. Discussion
Sweetpotato is a significant food crop and is broadly used in industrial materials, animal feed and human food. It has various natural advantages, such as strong stress resistance, high yield, wide adaptability [53]. Plant growth is determined by cell enlargement and proliferation and is restricted by cell wall, which limits the protoplasm increases of plant cells. The mechanism of cell wall extension has always been the focus of investigation due to the crucial functions of cell wall enlargement during plant morphogenesis [6]. Expansins are necessary for plant development and play critical roles in the relaxation of plant cell wall. As one of the most investigated structural proteins, expansins promote the growth of primary cell wall in plants. Expansins can loosen cell wall via cleaving hydrogen bonds between hemicellulose and cellulose microfibrils and can fill microfibrils and combine cellulose networks to form reticular systems [21,70], resulting in the increases of cell-wall strength and toughness and the continuous extension of cell wall [64]. However, members of expansin gene family in Ipomoea batatas are still not systematacially and comprehensively characterized. The completed genome sequencing of sweetpotato and advanced bioinformatics provide excellent foundations to perform the identification and characterisation of specific gene families [56]. In this study, the IbEXPs genes in Ipomoea batatas were systematically characterized and analyzed and their multifarious molecular characterizations were further implemented. Our study will provide lay the fundation for the further investigatins of regulatory modes and molecular functions of IbEXPs genes in sweetpotato growth and development and stress tolerance.
3.1. Characterization of IbEXPs in sweetpotato
In this study, 59 IbEXPs genes were identified from Ipomoea batatas genome, while only 37 ItfEXP genes were identified in Ipomoea trifida, suggesting that the number expansion of EXP gene family has occurred in Ipomoea batatas compared with to its diploid wild relative [55]. These 59 IbEXPs genes were divided into four subfamilies, IbEXPA, IbEXPB, IbEXLA and IbEXLB, which were similar to other plant species. The number of 59 IbEXPs genes is more than 46, 38, 36, 46, 36 and 38 in barley [59], Brachypodium distachyon [60], Arabidopsis [15], gingkgo [62], potato [71] and tomato [72], and is similar to 52 and 58 in tobacco [64] and rice [15], while is obviously less than 75, 88, 92, 93 and 241 in soybean [61], maize [58], sugarcane [4], cotton [63] and common wheat [57], suggesting that member number of EXP genes has changed significantly in different plants during evolution. Likewise, the sizes of expansin subfamilies are unevenly distributed in different plant species and the EXPA subfamily occupies the highest proportion compared with other three subfamilies in alll plants (Table 2). In addition, 59 IbEXPs genes were disproportionately distributed on 14 chromosomes. Similar uneven distributions were also observed in wheat [57], cotton [63], barley [59] and potato [71] et al., indicating that the number of IbEXPs genes on each chromosome is irrelevant to the size of chromosomes. Therefore, IbEXPs genes on chromosomes were relatively clustered instead of averagely distributed, which may be due to the uneven gene replications of chromosome fragments.
Although the IbEXPs protein properties diplayed significant differences, IbEXPs in the same clade harbored the relatively conserved gene structures, motifs and domains, which could provide an important reference for the phylogenetic analysis and functional investigation. These results verify that genes originated from ancestor can gradually expand and evolve [63]. IbEXPs members in the same subfamily tend to display similar compositions of motifs, domains and exon/intron structure, which were similar to that in other plant species such as tobacco, soybean, and cotton [61,63,64]. Gene structure exploration exhibited that about 83.1% of IbEXPs genes harbored 1-5 exons, which displayed the similarity with EXP genes in maize [58], soybean [61], tea [73], Populus [74] and banana [75]. However, several IbEXPs genes exhibited a distinct exception with over 8 exons, indicating the high divergences among them. The N-terminal conserved motifs (DPPB domains) of IbEXPs proteins which is rich in cysteine (Cys) residues with a characteristic catalytic domain may be related to the disulfide bond formation [13]. The C-terminal motif (pollen_allerg_1 domain) which cantains the conserved aromatic amino acids and polar tryptophan residues on its surface was considered as the polysaccharide binding domain. These motifs exist in IbEXPs proteins suggest that these motifs are crucial for the functions of IbEXPs in cell wall enlargement and loosening. Though the conserved DPBB and pollen_allerg_1 motifs are similar among most IbEXPs in the same subfamily, the IbEXPs showed significant differences of the molecular characteristics, which may be generated by the sequence differences in their non-conservative regions. Some specific motifs are only found in certain subfamilies, suggesting that some IbEXPs may play specific roles in plant development, because previous studies have widely reported that EXPs perform functions in various biological processes [17,26,34,37,41].
3.2. Evolutionary relationship and collinearity analysis of IbEXPs in sweetpotato
Phylogenetic relationship analysis showed that 59 IbEXPs were grouped into 4 subfamilies based on sequence homologies and subfamily classifications in Arabidopsis AtEXPs and rice OsEXPs [15,20,58]. At least one IbEXPs protein was found in each subfamily of rice and Arabidopsis [15,20], indicating that the discrepancies of EXP proteins may be earlier than dicots and monocots. The memebers of IbEXPs protiens in sweetpotato were unevenly distributed in the four subfamilies and most of them were the members of EXPA and EXPB subfamilies, while only a few of IbEXPs proteins blong to the EXLA and EXLB subfamilies. Similar uneven distribution events of members in these four subfamilies are also found in other plants, such as sugarcane [4], rice [15], Arabidopsis [15], cotton [63], potato [71] and tomato [72]. These results indicated that members of EXPA and EXPB subfamily may play more crucial roles in plant growth and development. For instance, the rice OsEXPA10, an Al-inducible expansin gene in rice, plays significant roles in cell elongation of roots [41]. Overexpression of GhEXPA8 in cotton improves the length of fibers and micronaire value [76]. TaEXPA2 plays crucial functions in seed production, and multiple abiotic stresses (salt, drought, oxidative and Cd) [27,28,29,30,77]. The β-expansin gene OsEXPB2 participate in the architecture of root system in rice [37]. Moreover, the proportion of members in the same subfamily of different plants varies greatly. The classification of IbEXPs proteins displayed similarities and discrepancies compared to that in other plants, suggesting that more diversity of functions and structures exist in EXP proteins in different plants.
Gene duplications are vital driving forces during the processes of expansion and evolution of many gene families in plants, which can promote the generation of novel functional genes and species, and then plant species can better resist adverse environmental conditions in the process of evolution [65,78,79]. Previous investigations in moso bamboo [13], maize [58], barley [59], potato [71], banana [75], and Brassica species [79] suggest that the duplication events of tandem, segments and genome may explain IbEXPs expansion and evolution in sweetpotato. Analogously, collinearity analysis displayed that multiple IbEXPs were identified to have the duplications of tandems and segmentals, suggesting that some IbEXPs genes may be generated via gene duplications, further supporting the mechanism that brings about the expansion of IbEXPs genes. And the segmental and tandem duplications have similar contributions to the increase of IbEXPs genes. Additionally, the IbEXPs genes exhibiting segmental duplications and tandem repeats usually come from the same subfamily, and the IbEXPs gene pairs were mainly from EXPA, EXPB and EXLB subfamily, indicating that the expansions of IbEXPs genes in specific subfamilies may be beneficial for sweetpotato growth and development and adaptation to changing environmental conditions. These results are similar to those of IbEXPs genes in moso bamboo [13], maize [58], and potato [71], and so on, indicating the critical evolutionary functions of egmental and tandem duplications in gene expansions.
Furthermore, synteny analysis estimating the relations between IbEXPs genes and the counterparts of nine plants were studied, including Ipomoea triloba, Oryza sativa, Arabidopsis, Zea mays, Triticum aestivum, Capsicum annuum, Solanum lycopersicum, Brassica oleracea and Brassica rapa. Among them, the largest orthologous gene numbers between sweetpotato and Ipomoea triloba were identified, which further proved their close evolutionary relations, following by Solanum lycopersicum, Arabidopsis, Capsicum annuum, Brassica oleracea and Brassica rapa. These gene pairs of orthologous may come from the common ancestor of sweetpotato and its corresponding plant species. In addition, more complex relations such as multiple Ipomoea trilobato-single sweetpotato genes or multiple sweetpotato-single Ipomoea trilobato genes was also found, indicating that the orthologous genes might have vital functions in IbEXPs evolutions in sweetpotato. Howerover, there were no orthologous genes between sweetpotato and three gramineae plants (Oryza sativa, Zea mays, and Triticum aestivum) and some EXP subfamlies such as EXLB were lost in many plants [4,57,58,59], which probably due to the abundant chromosomal fusions and rearrangements took place in the genomes, and the selective gene loss seriously obscured the identification of synteny relations [80]. These findings may might have something to do with phylogenetic relations between sweetpotato and these nine plants. And large-scale duplications predate plant divergence and play vital functions in IbEXPs in the expansions of IbEXPs gene family.
3.3. Expression patterns and functional prediction of IbANCs in sweetpotato
EXP gene family have received increasing attentions on account of their wide participation in various biological processes of plant development and responses to diverse stresses. Numerous documents have manifested that EXP genes are excellent candidates for regulating growth and development and improving stress tolerance of crops via molecular breeding [20,36,68]. For example, OsEXPA8, OsEXPA10 and OsEXPB2 in rice [37,39,41], GmEXPB2, GmEXLB1 and GmINS1 in soybean [8,18,36,38], TaEXPA2, TaEXPA8, TaEXPB23 in wheat [27,28,30,68,81] etc. have been proved their crucial functions in plant development and/or responses to various adverse environmental conditions. Up to now, the functions of sweetpotato IbEXPs genes in regulating stress resistance and plant development are still poorly understood. The expression profiles of genes are closely related to their biological functions, and the identification of gene expression can be conducive to accelerate the characterisation of gene functions. In this study, our results of RNA-seq and qRT-PCR showed that the examined IbEXPs genes exhibited obvious differential expression in different tissues and after various treatments of hormones and abiotic stresses, suggesting their diverse and critical functions of IbEXPs genes in plant development and stress response. For instance, the expression levels of multiple IbEXPs genes, particularly IbEXPA25, IbEXPB5, IbEXLA2 and IbEXLB11 showed obvious differential expression in different tissues, and IbEXPA4, IbEXPB5 and IbEXLB11 were dramatically induced or suppressed under different hormone and abiotic stresses, manifesting that they may play significant functions in sweetpotato growth and development and stress/hormone response, and/or help plants to reduce the damage caused by various stress.
In addition, the potential functions of IbEXPs genes in growth and development and stress/hormone response were further supported by cis-element and evolutionary relationship analysis. EXP genes in the close subfamilies may derive from the same gene/fragment duplication events, and functional investigations of EXPs have verified their conserved and/or similar roles in the same subfamily of plants [8,20,22,36,39,41]. Previous studies have manifested that TaEXPA2 [27,28,29,30], OsEXPA8 [39], OsEXPA10 [41], TaEXPA8 [81] and AtEXPA1 [82,83] in EXPA subfamily, and AtEXPB1 [84], GmINS1 [8], OsEXPB2 [37], GmEXPB2 [36,38] and TaEXPB23 [68] in EXPB subfamily, AtEXLA2 in EXLB subfamily [19,85], GmEXLB1 [18], MdEXLB1 [34] and BrEXLB1 [86] and in EXLB subfamily function as crucial participants in tissue growth and development and/or various stress responses. The phylogenetic analysis showed showed that the IbEXPA1/-25/-26 genes was closed linked to AtEXPA1/-10/-15, IbEXPB10 gene was closed linked to AtEXPB1/-3 and OsEXPB16/-17, IbEXLA1/2 genes were closed linked to AtEXLA1/-2/-3. And the expression levers of detected eight genes (IbEXPA4/-17/25, IbEXPB5/-10, IbEXLA1/-2, IbEXLB11) displayed tissue specificity in different tissues, and their transcripts were observably induced or suppressed under various hormone and stress treatments, implying that they might play vital roles in regulating the pathways of plant development and stress responses. Furthermore, IbEXP members in the same phylogenetic branch exhibited similar and discrepant expression profiles, suggesting the diversity of their potential functions. Therefore, development and/or stress response related IbEXPs which were grouped in the same subfamily were speculated to take part in controlling plant development and/or stress/hormone responses.
Previous investigations have showed that the phytohormones play significant roles in controling plant development and adaptation of various adverse environmental conditions [87,88]. Moreover, the cis-elements which lacated in gene promoter regions also function as crucial regulators in gene expression. In this study, numerous cis-elements in connection with hormones, growth and development, and different stresses including ABRE, ARE, ERE, TGACG-motif, CCGTCC-box, TGA-element, P-box, as-1, CAT-box, LTR, MSA-like and TATC-box and so on were detected in IbEXPs promoters, suggesting that they might function as the necessary participants in growth and development regulation, hormone and/or stress signaling. And the expression levels of examined IbEXPs genes (IbEXPA4/-17/-25, IbEXPB5/-10, IbEXLA1/-2 and IbEXLB11) were markedly increased or reduced by one or some hormones and abiotic stress. These results are similar to previous investigations in moso bamboo [13], Ipomoea trifida [55], tobacco [64], Brachypodium distachyon [60] and banana [75]. However, the exact biological functions of sweetpotato IbEXPs genes remain to be studied. Therefore, investigating the involvements of IbEXPs genes in develpoment and stress regulation can provide valuable information to illustrate their potential functions in growth and development and stress tolerance.
4. Conclusion
In present study, 59 IbEXPs genes were systematically identified and characterized from sweetpotato genome and unevenly distributed on 14 of 15 chromosomes. Their phylogenetic classification, conserved motif and domain, collinearity relationship, gene structure, cis-elements detection, molecular characterization and chromosome localization were systematically investigated. Duplication events such as the tandem and segmental duplication are conducive to the expansion of sweetpotato IbEXP gene family, and synteny analysis of orthologous genes between sweetpotato and typical plant species provided significant insights about the phylogenetic characteristics of IbEXPs genes in sweetpotato. Moreover, the results of transcriptome and qRT-PCR displayed differential and various expression patterns of IbEXPs genes in different developmental tissues and divers stress and hormone treatments. Multiple tissue-specific expression and hormone/stress-induced IbEXPs genes may have very close relationships with the transcriptional regulation of sweetpotato development and stress responses. In conclusion, the present results can not only contribute to further understanding the complexity and importance of EXP gene family, but also lay the foundation for the comprehensive analysis of potential functions in regulating plant growth and development and stress tolerance.
5. Materials and Methods
5.1. Identification of IbEXPs genes in sweetpotato
The genome data and GFF annotations files of sweetpotato were obtained from the online database Ipomoea Genome Hub (http://sweetpotao.com) [56]. The protein sequences of EXP members of rice and Arabidopsis were downloaded from rice genome annotation database (http://rice.plantbiology.msu.edu/) and TAIR (https://www.arabidopsis.org/) according to previous study [15], respectively. Then the EXP protein sequences of Arabidopsis and rice were employed as the query sequences to carry out the BLASTP program againsting all of the Ipomoea batatas protein sequences to identify all the possible EXP members in sweetpotato via the TBtools software with E-value ≤ 1e-5, NumofHits of 500 and NumofAligns of 250 [89]. Subsequently, the Prosite database (https://prosite.expasy.org/), Pfam database (http://pfam.xfam.org/) and NCBI batch CD-search (https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi) were used to confirm all obtained candidate EXP members and to exclude the protein sequences that lacked the EXP domain. Finally, a total of 59 non-redundant proteins sequences were identified as the putative EXP members and were further studied. Each IbEXPs sequence of nucleotide and protein can be found in Supplementary file S1.
5.2. Sequence alignment and phylogenetic analysis of EXP proteins
For phylogenetic analysis, the reported 36 Arabidopsis AtEXP proteins, 58 rice OsEXP proteins [15] and 59 sweetpotato IbEXP proteins were used to perform the phylogenetic tree analysis. Finally, 153 EXP protein sequences were used to carry out the multiple sequence alignmen employing the ClustalW with default parameters. After that, the MEGA software (version 11.0) was used to construct the unrooted phylogenetic tree by the neighbor-joining bootstrap method. The detailed parameters were as follows: poisson model, pairwise deletion and 1,000 replicates. The Arabidopsis and rice EXP protein sequences were provided in Supplementary file S1.
5.3. Analysis of motif pattern, conserved domain and protein property of IbEXPs
The online MEME Suite (version 5.5.4, https://meme-suite.org/meme/tools/meme) and the Batch CD-Search database (https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi) were employed to explore the motif and conserved domains (standard results) of each IbEXP protein, respectively. The ExPASy database (http://expasy.org/) were used to evaluate each IbEXP protein property, including the Mw (molecular weight) and pI (theoretical isoelectric point). The prediction of subcellular locations and phosphorylation sites of each IbEXP protein were also performed using the online datebase Plant-mPLoc (http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/) and NetPhos 3.1 (https://services.healthtech.dtu.dk/service.php?NetPhos-3.1), respectively.
5.4. Chromosomal location and collinearity analysis of IbEXPs
The IbEXPs location on the chromosome of sweetpotato was investigated using the GFF annotation information obtained from the Ipomoea genome website (https://sweetpotao.com/). To preform the synteny analysis bewteen IbEXPs and genes in other plants, the genome and GFF annotation files of Ipomoea batatas and other nine representative plant species (Ipomoea triloba, rice, Arabidopsis, maize, wheat, Brassica rapa, pepper, Brassica oleracea and tomato) were downloaded from TAIR, Ipomoea Genome Hub, Sol Genomics Network, EnsemblPlants database (http://plants.ensembl.org/index.html) and phytozome database (https://phytozome-next.jgi.doe.gov/). The collinearity relationships and gene duplications were investigated using Multiple Collinearity Scan toolkit (MCScanX) by a default parameter [90]. Then the circos and TBtools softwares with the 30 value as the minimum block size were employed to visualize these results [89,91].
5.5. Plant materials and treatments of abiotic stresses and hormones
To be consistent with the plant materials used in the RNA-seq in our previous study [67], the Taizhong 6 sweetpotato (Ipomoea batatas) plants were cultivated in a greenhouse in Jiangsu Normal University. Then the developing tuberous roots at 30 dap (DR1), 60 dap (DR2) and 100 dap (DR3)), mature tuberous roots at 120 dap (MR), mature leaves at 60 dap (L) and stems at 60 dap (S) were collected to analysis the expression patterns of selected IbEXPs genes. For the treatments of different hormones and abiotic stresses, plant seedlings of Ipomoea batatas (XuShu 22) harboring 3-4 mature leaves were cultured in a greenhouse. After a week of cultivation in water, the consistent seedlings were selected for the hormone treatments and abiotic stress. For hormone treatment, the seedlings were cultivated in water that contained 100 μM ACC, 100 μM ABA, 100 μM JA and 2mM SA, respectively. For abiotic stresses, seedlings were cultivated at 4 ˚C and 42 ˚C to simulate the cold and heat stresses, respectively. All needed leaf samples of controls and treated plants were harvested at 0, 1, 12, 24 and 48 h after various treatments. At least three independent biological replicates were collected for each treatment.
5.6. RNA extraction and qRT-PCR analysis
Total RNA in each collected sample was extracted using the RNA Extraction Kits (OMEGA, USA) following the method provided by manufacturers. The first-strand cDNA was synthesized via reverse transcription using 800 ng total RNA following the same methods in our study [92]. Then the synthesized cDNAs was diluted by the RNase/DNase-free water. The qRT-PCR experiments according to the same the same method were implemented on the CFX96™ Real-Time System (Bio-Rad, USA) as described before [92]. The sweetpotato IbARF (accession number: JX177359) gene was applied as the reference gene [93]. All used primers in qRT-PCR can be found in Table S5. Three independent technical and biological repetitions were performed for each sample in the qRT-PCR.
5.7. Statistical analyses
Data were presented as mean ± SE standard deviation. A cut-off two-fold value for differential gene expression which was regarded as the biological significance was adopted [94]. The originPro software (v8.0, SAS Institute) were graphs were employed to visualize the results of qRT-PCR experiments.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, J.Z. and Z.Z.; Data curation, J.Z. and M.Z.; Funding acquisition, J.Z., Z.Z., Q.Y., Y.S. and Y.S.; Project administration, Z.Z. and Y.S.; Software, J.Z and M.Z.; Supervision, Z.Z., T.D. and Y.S.; Validation, T.D. and M.Z.; Visualization, T.D. and Q.Y.; Writing – original draft, J.Z.; Writing – review & editing, M.Z., Y.S. and R.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Natural Science Foundation of Shandong Province (no. ZR2021MC079, no. ZR2021QC200, no. ZR2020QB154, no. ZR2020QC037), Science and technology Huimin demonstration special of Qingdao (solanaceous vegetables new varieties breeding and demonstration promotion, 23-2-8-xdny-15-nsh), National Natural Science Foundation of China (no. 32300170).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data and materials supporting the conclusions of this study are included within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cosgrove, D. J. Growth of the plant cell wall. Nature Reviews Molecular Cell Biology 2005, 6, 850–861. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, D. J. Plant cell wall extensibility: connecting plant cell growth with cell wall structure, mechanics, and the action of wall-modifying enzymes. Journal of Experimental Botany 2015, 67, 463–476. [Google Scholar] [CrossRef] [PubMed]
- Höfte, H.; Voxeur, A. Plant cell walls. Current Biology 2017, 27, R865–R870. [Google Scholar] [CrossRef] [PubMed]
- Santiago, T. R.; Pereira, V. M.; de Souza, W. R.; Steindorff, A. S.; Cunha, B. A. D. B.; Gaspar, M.; Fávaro, L. C. L.; Formighieri, E. F.; Kobayashi, A. K.; Molinari, H. B. C. Genome-wide identification, characterization and expression profile analysis of expansins gene family in sugarcane (Saccharum spp.). PLOS ONE 2018, 13, e0191081. [Google Scholar]
- Zhang, B.; Gao, Y.; Zhang, L.; Zhou, Y. The plant cell wall: Biosynthesis, construction, and functions. Journal of Integrative Plant Biology 2021, 63, 251–272. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Choi, H.-S.; Cho, H.-T. Root Hair-Specific EXPANSIN A7 Is Required For Root Hair Elongation in Arabidopsis. Molecules and cells 2011, 31, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, D. J. Plant expansins: diversity and interactions with plant cell walls. Current Opinion in Plant Biology 2015, 25, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zheng, J.; Zhou, H.; Chen, S.; Gao, Z.; Yang, Y.; Li, X.; Liao, H. The soybean β-expansin gene GmINS1 contributes to nodule development in response to phosphate starvation. Physiologia Plantarum 2021, 172, 2034–2047. [Google Scholar] [CrossRef]
- Cosgrove, D. J. Loosening of plant cell walls by expansins. Nature 2000, 407, 321. [Google Scholar] [CrossRef]
- Marowa, P.; Ding, A.; Kong, Y. Expansins: roles in plant growth and potential applications in crop improvement. Plant Cell Reports 2016, 35, 949–965. [Google Scholar] [CrossRef] [PubMed]
- McQueen-Mason, S. J.; Cosgrove, D. J. Expansin Mode of Action on Cell Walls (Analysis of Wall Hydrolysis, Stress Relaxation, and Binding). Plant Physiology 1995, 107, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Yennawar, N. H.; Li, L.-C.; Dudzinski, D. M.; Tabuchi, A.; Cosgrove, D. J. Crystal structure and activities of EXPB1 (Zea m 1), a β-expansin and group-1 pollen allergen from maize. Proceedings of the National Academy of Sciences 2006, 103, 14664–14671. [Google Scholar] [CrossRef] [PubMed]
- Jin, K.-M.; Zhuo, R.-Y.; Xu, D.; Wang, Y.-J.; Fan, H.-J.; Huang, B.-Y.; Qiao, G.-R. Genome-Wide Identification of the Expansin Gene Family and Its Potential Association with Drought Stress in Moso Bamboo. International Journal of Molecular Sciences 2020, 21, 9491. [Google Scholar] [CrossRef] [PubMed]
- Georgelis, N.; Yennawar, N. H.; Cosgrove, D. J. Structural basis for entropy-driven cellulose binding by a type-A cellulose-binding module (CBM) and bacterial expansin. Proceedings of the National Academy of Sciences 2012, 109, 14830–14835. [Google Scholar] [CrossRef] [PubMed]
- Sampedro, J.; Cosgrove, D. J. The expansin superfamily. Genome biology 2005, 6, 242.1–242.11. [Google Scholar] [CrossRef] [PubMed]
- Kende, H.; Bradford, K.; Brummell, D.; Cho, H.-T.; Cosgrove, D.; Fleming, A.; Gehring, C.; Lee, Y.; McQueen-Mason, S.; Rose, J. Nomenclature for members of the expansin superfamily of genes and proteins. Plant Molecular Biology 2004, 55, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Chang, L.; Sun, W.; Ullah, A.; Yang, X. Overexpression of an expansin-like gene, GhEXLB2 enhanced drought tolerance in cotton. Plant Physiology and Biochemistry 2021, 162, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Wang, B.; Du, H.; Li, W.; Li, X.; Zhang, C. GmEXLB1, a Soybean Expansin-Like B Gene, Alters Root Architecture to Improve Phosphorus Acquisition in Arabidopsis. Frontiers in Plant Science 2019, 10, 808. [Google Scholar] [CrossRef]
- Boron, A. K.; Van Loock, B.; Suslov, D.; Markakis, M. N.; Verbelen, J.-P.; Vissenberg, K. Over-expression of AtEXLA2 alters etiolated arabidopsis hypocotyl growth. Annals of Botany 2014, 115, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.; Cho, H.-T.; Lee, Y. Expansins: expanding importance in plant growth and development. Physiologia Plantarum 2006, 126, 511–518. [Google Scholar] [CrossRef]
- Li, Y.; Jones, L.; McQueen-Mason, S. Expansins and cell growth. Current Opinion in Plant Biology 2003, 6, 603–610. [Google Scholar] [CrossRef]
- Choi, D.; Kim, J. H.; Lee, Y. Expansins in Plant Development. In Advances in Botanical Research; Academic Press, 2008; Vol. 47, pp. 47–97. [Google Scholar]
- Ashwin Narayan, J.; Chakravarthi, M.; Nerkar, G.; Manoj, V. M.; Dharshini, S.; Subramonian, N.; Premachandran, M. N.; Arun Kumar, R.; Krishna Surendar, K.; Hemaprabha, G.; Ram, B.; Appunu, C. Overexpression of expansin EaEXPA1, a cell wall loosening protein enhances drought tolerance in sugarcane. Industrial Crops and Products 2021, 159, 113035. [Google Scholar] [CrossRef]
- McQueen-Mason, S.; Durachko, D. M.; Cosgrove, D. J. Two endogenous proteins that induce cell wall extension in plants. The Plant Cell 1992, 4, 1425–1433. [Google Scholar] [PubMed]
- Dong, B.; Wang, Q.; Zhou, D.; Wang, Y.; Miao, Y.; Zhong, S.; Fang, Q.; Yang, L.; Xiao, Z.; Zhao, H. Abiotic stress treatment reveals expansin like A gene OfEXLA1 improving salt and drought tolerance of Osmanthus fragrans by responding to abscisic acid. Horticultural Plant Journal 2023. [Google Scholar] [CrossRef]
- Brasileiro, A. C. M.; Lacorte, C.; Pereira, B. M.; Oliveira, T. N.; Ferreira, D. S.; Mota, A. P. Z.; Saraiva, M. A. P.; Araujo, A. C. G.; Silva, L. P.; Guimaraes, P. M. Ectopic expression of an expansin-like B gene from wild Arachis enhances tolerance to both abiotic and biotic stresses. The Plant Journal 2021, 107, 1681–1696. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, G.; An, J.; Li, Q.; Chen, Y.; Zhao, X.; Wu, J.; Wang, Y.; Hao, Q.; Wang, W.; Wang, W. Expansin gene TaEXPA2 positively regulates drought tolerance in transgenic wheat (Triticum aestivum L.). Plant Science 2020, 298, 110596. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Ren, Y.; Zhang, G.; An, J.; Yang, J.; Wang, Y.; Wang, W. Overexpression of the wheat expansin gene TaEXPA2 improves oxidative stress tolerance in transgenic Arabidopsis plants. Plant Physiology and Biochemistry 2018, 124, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Chen, Y.; An, J.; Zhao, Z.; Zhang, G.; Wang, Y.; Wang, W. Wheat expansin gene TaEXPA2 is involved in conferring plant tolerance to Cd toxicity. Plant Science 2018, 270, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Han, Y.; Kong, X.; Kang, H.; Ren, Y.; Wang, W. Ectopic expression of wheat expansin gene TaEXPA2 improved the salt tolerance of transgenic tobacco by regulating Na+/K+ and antioxidant competence. Physiologia Plantarum 2017, 159, 161–177. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, H.; Zhang, H.; Li, J.; Xie, B.; Xu, J. The expansin gene PttEXPA8 from poplar (Populus tomentosa) confers heat resistance in transgenic tobacco. Plant Cell, Tissue and Organ Culture (PCTOC) 2016, 126, 353–359. [Google Scholar] [CrossRef]
- Minoia, S.; Boualem, A.; Marcel, F.; Troadec, C.; Quemener, B.; Cellini, F.; Petrozza, A.; Vigouroux, J.; Lahaye, M.; Carriero, F.; Bendahmane, A. Induced mutations in tomato SlExp1 alter cell wall metabolism and delay fruit softening. Plant Science 2016, 242, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Kaur, P.; Samuel, D. V. K.; Bansal, K. C. Fruit-specific Over-expression of LeEXP1 Gene in Tomato Alters Fruit Texture. Journal of Plant Biochemistry and Biotechnology 2010, 19, 177–183. [Google Scholar] [CrossRef]
- Chen, Y.-h.; Xie, B.; An, X.-h.; Ma, R.-p.; Zhao, D.-y.; Cheng, C.-g.; Li, E.-m.; Zhou, J.-t.; Kang, G.-d.; Zhang, Y.-z. Overexpression of the apple expansin-like gene MdEXLB1 accelerates the softening of fruit texture in tomato. Journal of Integrative Agriculture 2022, 21, 3578–3588. [Google Scholar] [CrossRef]
- Sane, V. A.; Chourasia, A.; Nath, P. Softening in mango (Mangifera indica cv. Dashehari) is correlated with the expression of an early ethylene responsive, ripening related expansin gene, MiExpA1. Postharvest Biology and Technology 2005, 38, 223–230. [Google Scholar] [CrossRef]
- Li, X.; Zhao, J.; Tan, Z.; Zeng, R.; Liao, H. GmEXPB2, a Cell Wall β-Expansin, Affects Soybean Nodulation through Modifying Root Architecture and Promoting Nodule Formation and Development. Plant Physiology 2015, 169, 2640–2653. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Wenwen, Y.; Zang, G.; Kang, Z.; Zhang, Z.; Huang, J.; Wang, G. OsEXPB2, a β-expansin gene, is involved in rice root system architecture. Molecular Breeding 2015, 35, 41. [Google Scholar] [CrossRef]
- Guo, W.; Zhao, J.; Li, X.; Qin, L.; Yan, X.; Liao, H. A soybean β-expansin gene GmEXPB2 intrinsically involved in root system architecture responses to abiotic stresses. The Plant Journal 2011, 66, 541–552. [Google Scholar] [CrossRef]
- Wang, Y.; Ma, N.; Qiu, S.; Zou, H.; Zang, G.; Kang, Z.; Wang, G.; Huang, J. Regulation of the α-expansin gene OsEXPA8 expression affects root system architecture in transgenic rice plants. Molecular Breeding 2014, 34, 47–57. [Google Scholar] [CrossRef]
- Wang, L.; Wang, W.; Miao, Y.; Peters, M.; Schultze-Kraft, R.; Liu, G.; Chen, Z. Development of transgenic composite Stylosanthes plants to study root growth regulated by a β-expansin gene, SgEXPB1, under phosphorus deficiency. Plant Cell Reports 2023, 42, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Che, J.; Yamaji, N.; Shen, R. F.; Ma, J. F. An Al-inducible expansin gene, OsEXPA10 is involved in root cell elongation of rice. The Plant Journal 2016, 88, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tu, L.; Pettolino, F. A.; Ji, S.; Hao, J.; Yuan, D.; Deng, F.; Tan, J.; Hu, H.; Wang, Q.; Llewellyn, D. J.; Zhang, X. GbEXPATR, a species-specific expansin, enhances cotton fibre elongation through cell wall restructuring. Plant Biotechnology Journal 2016, 14, 951–963. [Google Scholar] [CrossRef] [PubMed]
- Ma, N.; Wang, Y.; Qiu, S.; Kang, Z.; Che, S.; Wang, G.; Huang, J. Overexpression of OsEXPA8, a Root-Specific Gene, Improves Rice Growth and Root System Architecture by Facilitating Cell Extension. PLOS ONE 2013, 8, e75997. [Google Scholar] [CrossRef] [PubMed]
- Pien, S.; Wyrzykowska, J.; McQueen-Mason, S.; Smart, C.; Fleming, A. Local expression of expansin induces the entire process of leaf development and modifies leaf shape. Proceedings of the National Academy of Sciences 2001, 98, 11812–11817. [Google Scholar] [CrossRef] [PubMed]
- Gray-Mitsumune, M.; Blomquist, K.; McQueen-Mason, S.; Teeri, T. T.; Sundberg, B.; Mellerowicz, E. J. Ectopic expression of a wood-abundant expansin PttEXPA1 promotes cell expansion in primary and secondary tissues in aspen. Plant Biotechnology Journal 2008, 6, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Xu, L.; Lin, H.; Cao, J. Two Expansin Genes, AtEXPA4 and AtEXPB5, Are Redundantly Required for Pollen Tube Growth and AtEXPA4 Is Involved in Primary Root Elongation in Arabidopsis thaliana. Genes 2021, 12, 249. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.; Lee, Y.; Cho, H.-T.; Kende, H. Regulation of Expansin Gene Expression Affects Growth and Development in Transgenic Rice Plants. The Plant Cell 2003, 15, 1386–1398. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Bradford, K. J. Expression of an Expansin Is Associated with Endosperm Weakening during Tomato Seed Germination1. Plant Physiology 2000, 124, 1265–1274. [Google Scholar] [CrossRef] [PubMed]
- Calderini, D. F.; Castillo, F. M.; Arenas-M, A.; Molero, G.; Reynolds, M. P.; Craze, M.; Bowden, S.; Milner, M. J.; Wallington, E. J.; Dowle, A.; Gomez, L. D.; McQueen-Mason, S. J. Overcoming the trade-off between grain weight and number in wheat by the ectopic expression of expansin in developing seeds leads to increased yield potential. New Phytologist 2021, 230, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Dahal, P.; Bradford, K. J. Two Tomato Expansin Genes Show Divergent Expression and Localization in Embryos during Seed Development and Germination. Plant Physiology 2001, 127, 928–936. [Google Scholar] [CrossRef]
- Zenoni, S.; Reale, L.; Tornielli, G. B.; Lanfaloni, L.; Porceddu, A.; Ferrarini, A.; Moretti, C.; Zamboni, A.; Speghini, A.; Ferranti, F.; Pezzotti, M. Downregulation of the Petunia hybrida α-Expansin Gene PhEXP1 Reduces the Amount of Crystalline Cellulose in Cell Walls and Leads to Phenotypic Changes in Petal Limbs. The Plant Cell 2004, 16, 295–308. [Google Scholar] [CrossRef] [PubMed]
- Arisha, M. H.; Aboelnasr, H.; Ahmad, M. Q.; Liu, Y.; Tang, W.; Gao, R.; Yan, H.; Kou, M.; Wang, X.; Zhang, Y.; Li, Q. Transcriptome sequencing and whole genome expression profiling of hexaploid sweetpotato under salt stress. BMC genomics 2020, 21, 197. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q. Improvement for agronomically important traits by gene engineering in sweetpotato. Breeding Science 2017, 67, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Ahn, Y. O.; Kim, S. H.; Kim, C. Y.; Lee, J.-S.; Kwak, S.-S.; Lee, H.-S. Exogenous sucrose utilization and starch biosynthesis among sweetpotato cultivars. Carbohydrate Research 2010, 345, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Chen, L.; Lang, T.; Qu, H.; Zhang, C.; Feng, J.; Pu, Z.; Peng, M.; Lin, H. Genome-Wide Identification and Expression Analysis of Expansin Gene Family in the Storage Root Development of Diploid Wild Sweetpotato Ipomoea trifida. Genes 2022, 13, 1043. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Moeinzadeh, M. H.; Kuhl, H.; Helmuth, J.; Xiao, P.; Haas, S.; Liu, G.; Zheng, J.; Sun, Z.; Fan, W.; Deng, G.; Wang, H.; Hu, F.; Zhao, S.; Fernie, A. R.; Boerno, S.; Timmermann, B.; Zhang, P.; Vingron, M. Haplotype-resolved sweet potato genome traces back its hexaploidization history. Nature Plants 2017, 3, 696–703. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Liu, Y.; Deng, X.; Liu, D.; Liu, Y.; Hu, Y.; Yan, Y. Genome-wide identification and expression analysis of expansin gene family in common wheat (Triticum aestivum L.). BMC genomics 2019, 20, 101. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yan, H.; Chen, W.; Liu, J.; Jiang, C.; Jiang, H.; Zhu, S.; Cheng, B. Genome-wide identification and characterization of maize expansin genes expressed in endosperm. Molecular Genetics and Genomics 2014, 289, 1061–1074. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Fu, M.; Guo, F.; Wu, C. Genome-wide identification of expansin gene family in barley and drought-related expansins identification based on RNA-seq. Genetica 2021, 149, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Luo, Y.; Wang, G.; Feng, C.; Li, H. Genome-wide identification of expansin genes in Brachypodium distachyon and functional characterization of BdEXPA27. Plant Science 2020, 296, 110490. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wu, N.; Song, W.; Yin, G.; Qin, Y.; Yan, Y.; Hu, Y. Soybean (Glycine max) expansin gene superfamily origins: segmental and tandem duplication events followed by divergent selection among subfamilies. BMC Plant Biology 2014, 14, 93. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Guo, J.; El-Kassaby, Y. A.; Wang, G. Genome-Wide Identification of Expansin Gene Family and Their Response under Hormone Exposure in Ginkgo biloba L. International Journal of Molecular Sciences 2023, 24, 5901. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.-M.; Zuo, D.-Y.; Wang, X.-F.; Cheng, H.-L.; Zhang, Y.-P.; Wang, Q.-L.; Song, G.-L.; Ma, Z.-Y. Genome-wide identification of the expansin gene family reveals that expansin genes are involved in fibre cell growth in cotton. BMC Plant Biology 2020, 20, 223. [Google Scholar] [CrossRef] [PubMed]
- Ding, A.; Marowa, P.; Kong, Y. Genome-wide identification of the expansin gene family in tobacco (Nicotiana tabacum). Molecular Genetics and Genomics 2016, 291, 1891–1907. [Google Scholar] [CrossRef] [PubMed]
- Cannon, S. B.; Mitra, A.; Baumgarten, A.; Young, N. D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC plant biology 2004, 4, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhai, Z.; Li, Y.; Geng, S.; Song, G.; Guan, J.; Jia, M.; Wang, F.; Sun, G.; Feng, N.; Kong, X.; Chen, L.; Mao, L.; Li, A. Genome-Wide Identification and Expression Profiling of the TCP Family Genes in Spike and Grain Development of Wheat (Triticum aestivum L.). Frontiers in Plant Science 2018, 9. [Google Scholar]
- Li, D.; Zhu, J.; Zhang, J.; Yang, Y.; Jia, Q.; Li, Z.; Zhu, M.; Dong, T. Combination analysis of genome-wide association and transcriptome sequencing identifies tuberous root development-related homeobox transcription factors and functional analysis of IbHB040 in sweetpotato. Scientia Horticulturae 2024, 326, 112753. [Google Scholar] [CrossRef]
- yang Han, Y.; xiu Li, A.; Li, F.; rong Zhao, M.; Wang, W. Characterization of a wheat (Triticum aestivum L.) expansin gene, TaEXPB23, involved in the abiotic stress response and phytohormone regulation. Plant Physiology and Biochemistry 2012, 54, 49–58. [Google Scholar]
- Meng, X.; Liu, S.; Dong, T.; Xu, T.; Ma, D.; Pan, S.; Li, Z.; Zhu, M. Comparative transcriptome and proteome analysis of salt-tolerant and salt-sensitive sweet potato and overexpression of IbNAC7 confers salt tolerance in Arabidopsis. Frontiers in Plant Science 2020, 11, 1342. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.-T.; Cosgrove, D. J. Altered expression of expansin modulates leaf growth and pedicel abscission in Arabidopsis thaliana. Proceedings of the National Academy of Sciences 2000, 97, 9783–9788. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, B.; Li, C.; Lei, C.; Kong, C.; Yang, Y.; Gong, M. A comprehensive expression analysis of the expansin gene family in potato (Solanum tuberosum) discloses stress-responsive expansin-like B genes for drought and heat tolerances. PLOS ONE 2019, 14, e0219837. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Liu, L.; Wang, X.; Han, Z.; Ouyang, B.; Zhang, J.; Li, H. Genome-wide identification and expression analysis of the expansin gene family in tomato. Molecular Genetics and Genomics 2016, 291, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Bordoloi, K.; Dihingia, P.; Krishnatreya, D.; Agarwala, N. Genome-wide identification, characterization and expression analysis of the expansin gene family under drought stress in tea (Camellia sinensis L.). Plant Science Today 2021, 8, 32–44. [Google Scholar] [CrossRef]
- Yin, Z.; Zhou, F.; Chen, Y.; Wu, H.; Yin, T. Genome-Wide Analysis of the Expansin Gene Family in Populus and Characterization of Expression Changes in Response to Phytohormone (Abscisic Acid) and Abiotic (Low-Temperature) Stresses. International Journal of Molecular Sciences 2023, 24, 7759. [Google Scholar] [CrossRef] [PubMed]
- Backiyarani, S.; Anuradha, C.; Thangavelu, R.; Chandrasekar, A.; Renganathan, B.; Subeshkumar, P.; Giribabu, P.; Muthusamy, M.; Uma, S. Genome-wide identification, characterization of expansin gene family of banana and their expression pattern under various stresses. 3 Biotech 2022, 12, 101. [Google Scholar] [CrossRef] [PubMed]
- Bajwa, K. S.; Shahid, A. A.; Rao, A. Q.; Bashir, A.; Aftab, A.; Husnain, T. Stable transformation and expression of GhEXPA8 fiber expansin gene to improve fiber length and micronaire value in cotton. Frontiers in Plant Science 2015, 6, 838. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Han, Y.; Zhang, M.; Zhou, S.; Kong, X.; Wang, W. Overexpression of the Wheat Expansin Gene TaEXPA2 Improved Seed Production and Drought Tolerance in Transgenic Tobacco Plants. PLOS ONE 2016, 11, e0153494. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Conery, J. S. The Evolutionary Fate and Consequences of Duplicate Genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Ma, B.; Shen, J.; Zhao, S.; Ma, X.; Wang, Z.; Fan, Y.; Tang, Q.; Wei, D. The evolution of the expansin gene family in Brassica species. Plant Physiology and Biochemistry 2021, 167, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Paterson, A. H.; Wang, X.; Tang, H.; Lee, T. H. Synteny and Genomic Rearrangements. In Plant Genome Diversity Volume 1: Plant Genomes, their Residents, and their Evolutionary Dynamics; Wendel, J. F., Greilhuber, J., Dolezel, J., Leitch, I. J., Eds.; Springer Vienna: Vienna, 2012; pp. 195–207. [Google Scholar]
- Peng, L.; Xu, Y.; Wang, X.; Feng, X.; Zhao, Q.; Feng, S.; Zhao, Z.; Hu, B.; Li, F. Overexpression of paralogues of the wheat expansin gene TaEXPA8 improves low-temperature tolerance in Arabidopsis. Plant Biology 2019, 21, 1119–1131. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-Q.; Wei, P.-C.; Xiong, Y.-M.; Yang, Y.; Chen, J.; Wang, X.-C. Overexpression of the Arabidopsis α-expansin gene AtEXPA1 accelerates stomatal opening by decreasing the volumetric elastic modulus. Plant Cell Reports 2011, 30, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Liu, K.; Lu, Y. T. Specific roles of AtEXPA1 in plant growth and stress adaptation. Russian Journal of Plant Physiology 2010, 57, 241–246. [Google Scholar] [CrossRef]
- Kwon, Y. R.; Lee, H. J.; Kim, K. H.; Hong, S.-W.; Lee, S. J.; Lee, H. Ectopic expression of Expansin3 or Expansinβ1 causes enhanced hormone and salt stress sensitivity in Arabidopsis. Biotechnology Letters 2008, 30, 1281–1288. [Google Scholar] [CrossRef] [PubMed]
- Abuqamar, S.; Ajeb, S.; Sham, A.; Enan, M. R.; Iratni, R. A mutation in the expansin-like A2 gene enhances resistance to necrotrophic fungi and hypersensitivity to abiotic stress in Arabidopsis thaliana. Molecular Plant Pathology 2013, 14, 813–827. [Google Scholar] [CrossRef] [PubMed]
- Muthusamy, M.; Kim, J. Y.; Yoon, E. K.; Kim, J. A.; Lee, S. I. BrEXLB1, a Brassica rapa Expansin-Like B1 Gene Is Associated with Root Development, Drought Stress Response, and Seed Germination. Genes 2020, 11, 404. [Google Scholar] [CrossRef] [PubMed]
- Verma, V.; Ravindran, P.; Kumar, P. P. Plant hormone-mediated regulation of stress responses. BMC plant biology 2016, 16, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Introduction to Phytohormones. The Plant Cell 2010, 22, 1.
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H. R.; Frank, M. H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Molecular Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; DeBarry, J. D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-h.; Jin, H.; Marler, B.; Guo, H.; Kissinger, J. C.; Paterson, A. H. MCScanX: a toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Research 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S. J.; Marra, M. A. Circos: An information aesthetic for comparative genomics. Genome research 2009, 19, 1639–1645. [Google Scholar] [CrossRef]
- Zhang, J.; Hu, Z.; Yao, Q.; Guo, X.; Nguyen, V.; Li, F.; Chen, G. A tomato MADS-box protein, SlCMB1, regulates ethylene biosynthesis and carotenoid accumulation during fruit ripening. Scientific reports 2018, 8, 3413–3427. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-j.; Zou, W.-s.; Fei, C.-y.; Wu, G.; Li, X.-y.; Lin, H.-h.; Xi, D.-h. α-Expansin EXPA4 Positively Regulates Abiotic Stress Tolerance but Negatively Regulates Pathogen Resistance in Nicotiana tabacum. Plant and Cell Physiology 2018, 59, 2317–2330. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, C.; Zhu, Q.; Guo, F.; Chai, R.; Wang, M.; Deng, X.; Dong, T.; Meng, X.; Zhu, M. Genome- and transcriptome-wide systematic characterization of bZIP transcription factor family identifies promising members involved in abiotic stress response in sweetpotato. Scientia Horticulturae 2022, 303, 111185. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic tree of 59 IbEXPs and EXPs in Arabidopsis and rice. The phylogenetic relationships were constructed by the MEGA 11.0 software using the neighbor-joining bootstrap method according to the following parameters: poisson model, pairwise deletion and 1,000 replicates. Different subfamilies are named following the studies in Arabidopsis and rice, and different colors were used to distinguish each subfamily. Red circles, green triangles and blue triangles represent sweetpotato IbEXPs, Arabidopsis AtEXPs and rice OsEXPs, respectively.
Figure 1.
Phylogenetic tree of 59 IbEXPs and EXPs in Arabidopsis and rice. The phylogenetic relationships were constructed by the MEGA 11.0 software using the neighbor-joining bootstrap method according to the following parameters: poisson model, pairwise deletion and 1,000 replicates. Different subfamilies are named following the studies in Arabidopsis and rice, and different colors were used to distinguish each subfamily. Red circles, green triangles and blue triangles represent sweetpotato IbEXPs, Arabidopsis AtEXPs and rice OsEXPs, respectively.
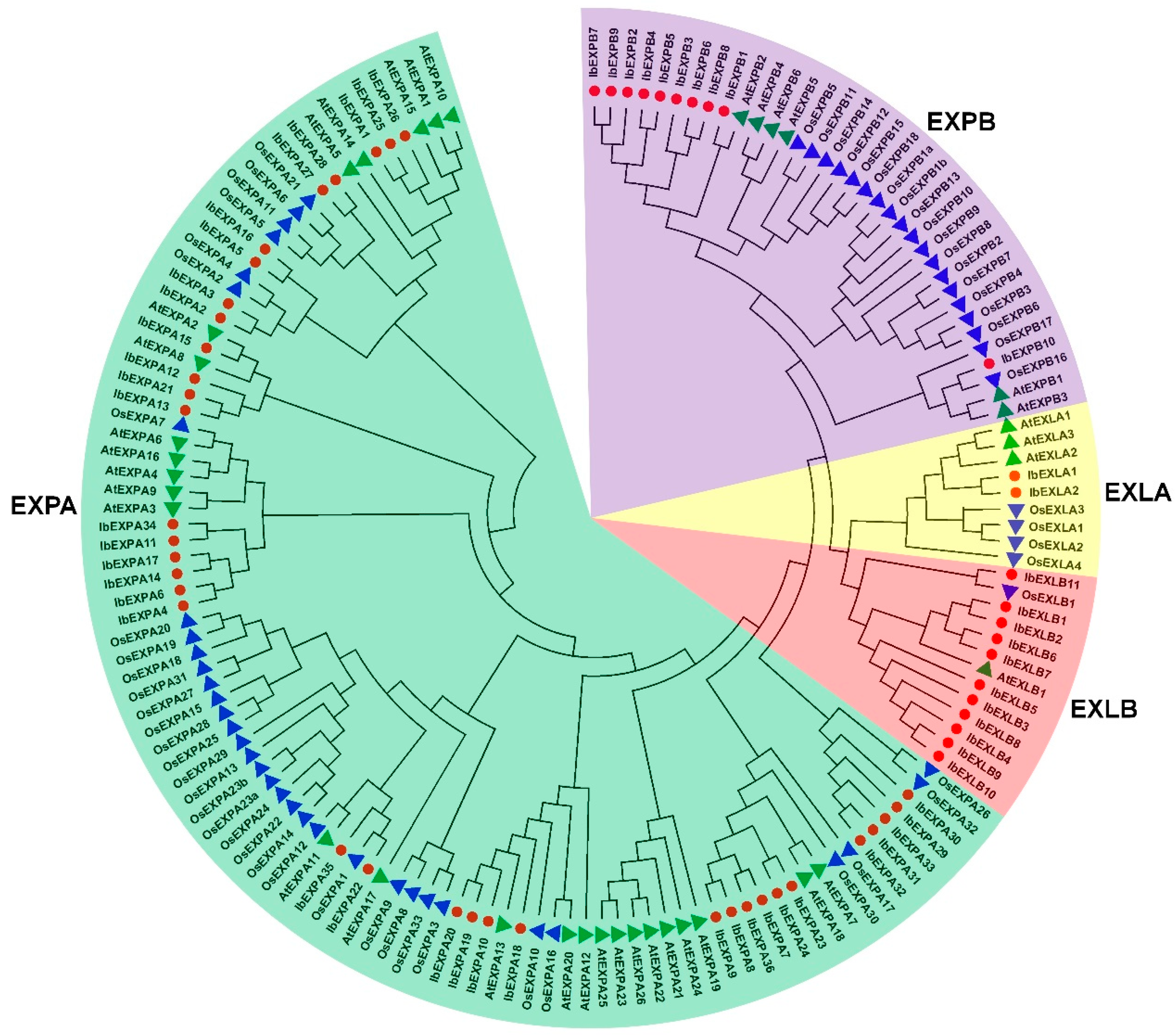
Figure 2.
Localizations of 59 IbEXPs on sweetpotato chromosomes (LG1-LG15). The red arc behind some IbEXPs represent the gene duplication of part of the EXP genes in sweetpotato.
Figure 2.
Localizations of 59 IbEXPs on sweetpotato chromosomes (LG1-LG15). The red arc behind some IbEXPs represent the gene duplication of part of the EXP genes in sweetpotato.

Figure 3.
Segmental duplications and collinearity analysis of IbEXPs in sweetpotato. LG1- LG15 are represented by different colored rectangles. The heatmap and polyline along each rectangle depict the gene density of each chromosome. Duplicated IbEXP gene pairs on sweetpotato chromosomes are indicated by colored lines, and these corresponding genes are also marked with colors. Other IbEXPs genes which exhibit no collinear relationships were marked with black color.
Figure 3.
Segmental duplications and collinearity analysis of IbEXPs in sweetpotato. LG1- LG15 are represented by different colored rectangles. The heatmap and polyline along each rectangle depict the gene density of each chromosome. Duplicated IbEXP gene pairs on sweetpotato chromosomes are indicated by colored lines, and these corresponding genes are also marked with colors. Other IbEXPs genes which exhibit no collinear relationships were marked with black color.

Figure 4.
Synteny analyses of sweetpotato IbEXPs with other nine representative plants. These plants include Ipomoea triloba (A), Oryza sativa and Arabidopsis thaliana (B), Brassica rapa and Brassica oleracea (C), Triticum aestivum and Zea mays (D), Solanum lycopersicum and Capsicum annuum (E). The chromosomes of various plants are distinguished with differential colors.
Figure 4.
Synteny analyses of sweetpotato IbEXPs with other nine representative plants. These plants include Ipomoea triloba (A), Oryza sativa and Arabidopsis thaliana (B), Brassica rapa and Brassica oleracea (C), Triticum aestivum and Zea mays (D), Solanum lycopersicum and Capsicum annuum (E). The chromosomes of various plants are distinguished with differential colors.
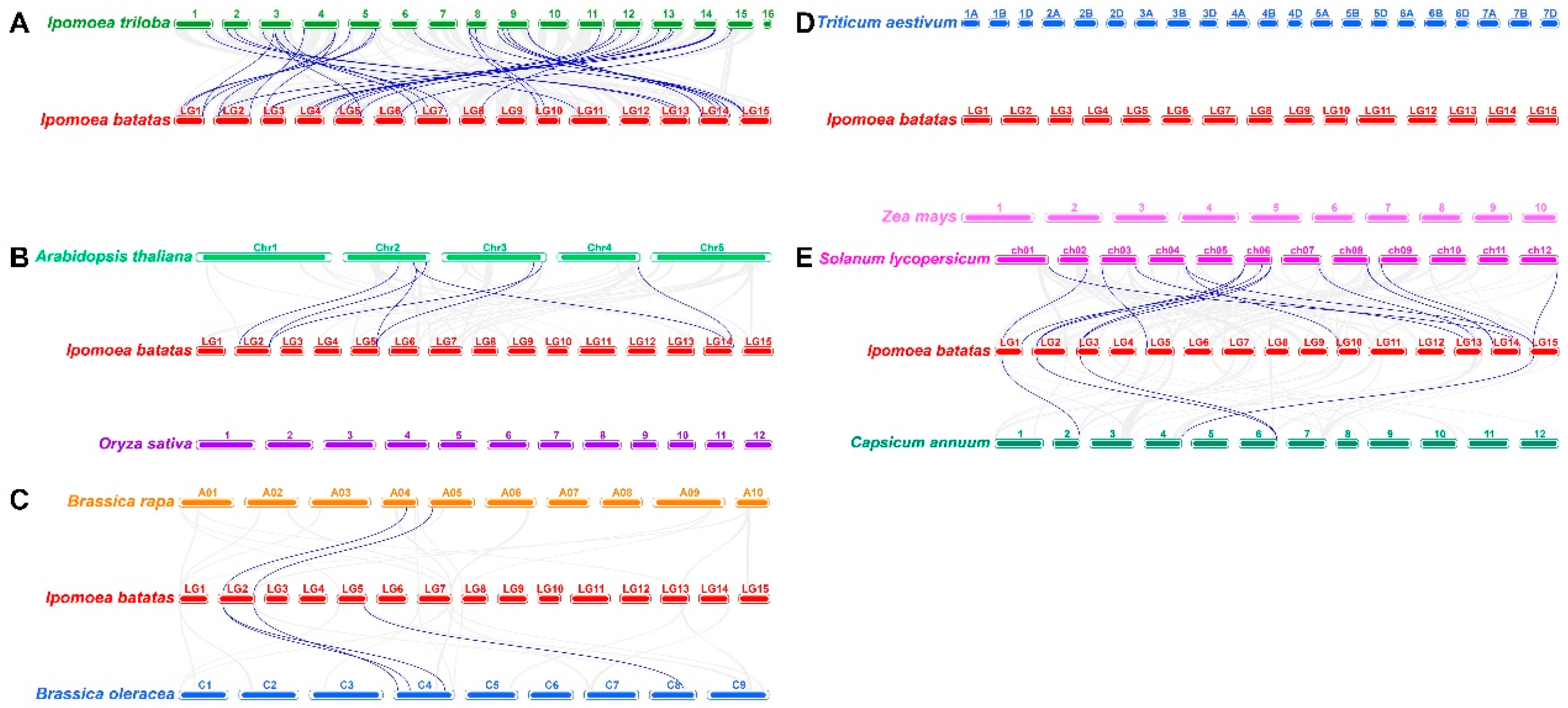
Figure 5.
Phylogenetic tree, motif pattern, protein domain and gene structures of 59 sweetpotato IbEXPs. (A). The phylogenetic tree of 59 IbEXPs was constructed by MEGA 11.0 based on the consistent parameters used in Figure 1. (B). Distributions of motifs in each IbEXP protein. 20 motifs were identified by MEME data. (C). Conserved domain distributions of IbEXP proteins. The CD-search of NCBI database was used to detect the distributions of conserved domains of IbEXP proteins. The different colorful boxes present diverse conserved domains of each subfamily. (D). Gene structures of 59 sweetpotato IbEXPs. Green and yellow bars were used to represent the UTR and exons, respectively. The black lines were employed to indicate the introns. The length of IbEXP proteins or genes were estimated using the scale at the bottom.
Figure 5.
Phylogenetic tree, motif pattern, protein domain and gene structures of 59 sweetpotato IbEXPs. (A). The phylogenetic tree of 59 IbEXPs was constructed by MEGA 11.0 based on the consistent parameters used in Figure 1. (B). Distributions of motifs in each IbEXP protein. 20 motifs were identified by MEME data. (C). Conserved domain distributions of IbEXP proteins. The CD-search of NCBI database was used to detect the distributions of conserved domains of IbEXP proteins. The different colorful boxes present diverse conserved domains of each subfamily. (D). Gene structures of 59 sweetpotato IbEXPs. Green and yellow bars were used to represent the UTR and exons, respectively. The black lines were employed to indicate the introns. The length of IbEXP proteins or genes were estimated using the scale at the bottom.
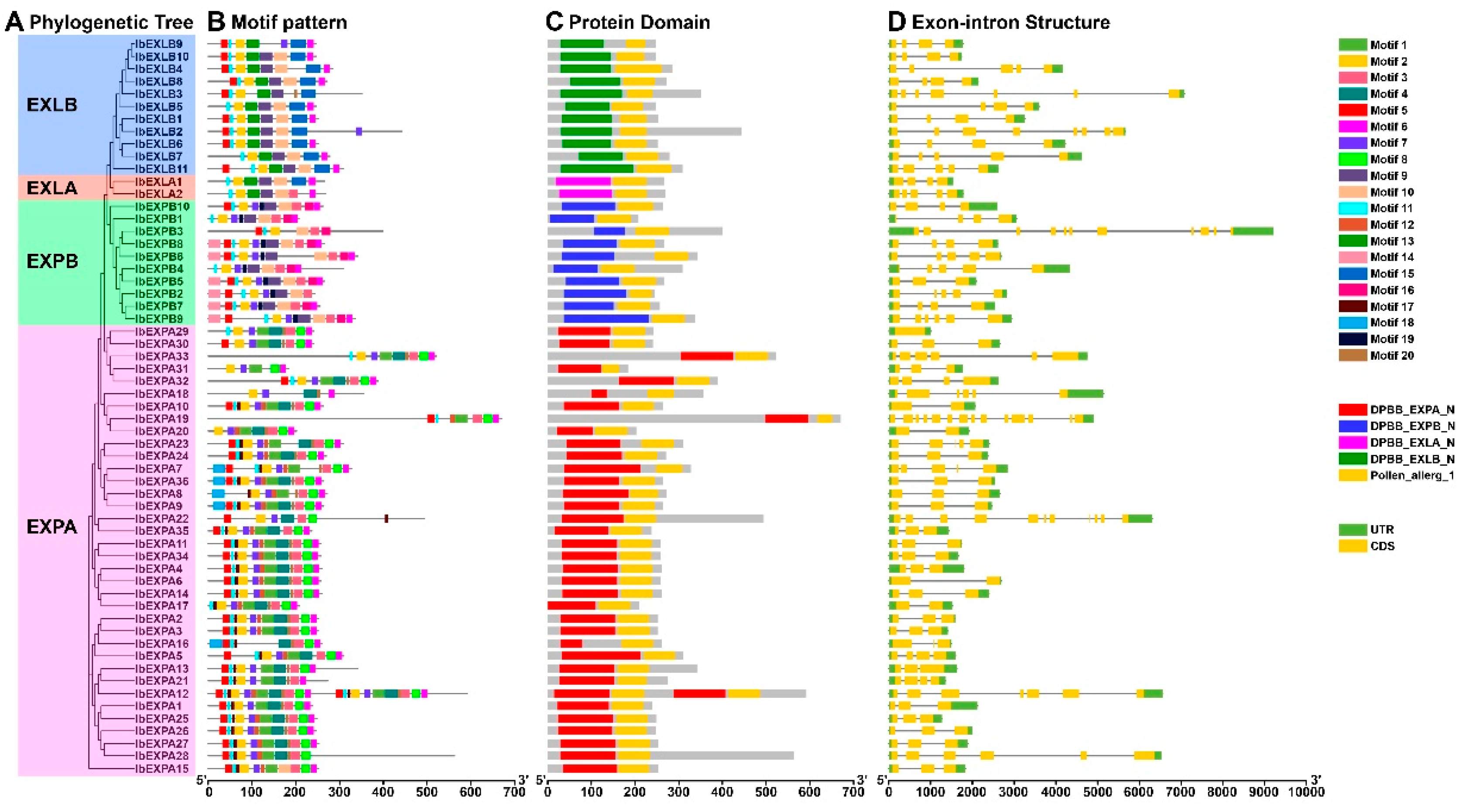
Figure 6.
Predicted cis-elements in the promoters of 59 sweetpotato IbEXPs. (A). The phylogenetic tree and predicted cis-elements detected from 2000 bp promoter regions of each IbEXPs gene by PlantCARE database. The same phylogenetic tree as Figure 5 was used. All cis-elements are classified into three categories: hormone responsiveness, growth and development, and stress responsiveness. (B). The number of cis-elements in promoter of IbEXPs. The left table represents the number of each kind of cis-element found in each subfamily. The red rectangles indicate the gene number in each subfamily. (C). Venn diagram of three categories of cis-elements.
Figure 6.
Predicted cis-elements in the promoters of 59 sweetpotato IbEXPs. (A). The phylogenetic tree and predicted cis-elements detected from 2000 bp promoter regions of each IbEXPs gene by PlantCARE database. The same phylogenetic tree as Figure 5 was used. All cis-elements are classified into three categories: hormone responsiveness, growth and development, and stress responsiveness. (B). The number of cis-elements in promoter of IbEXPs. The left table represents the number of each kind of cis-element found in each subfamily. The red rectangles indicate the gene number in each subfamily. (C). Venn diagram of three categories of cis-elements.
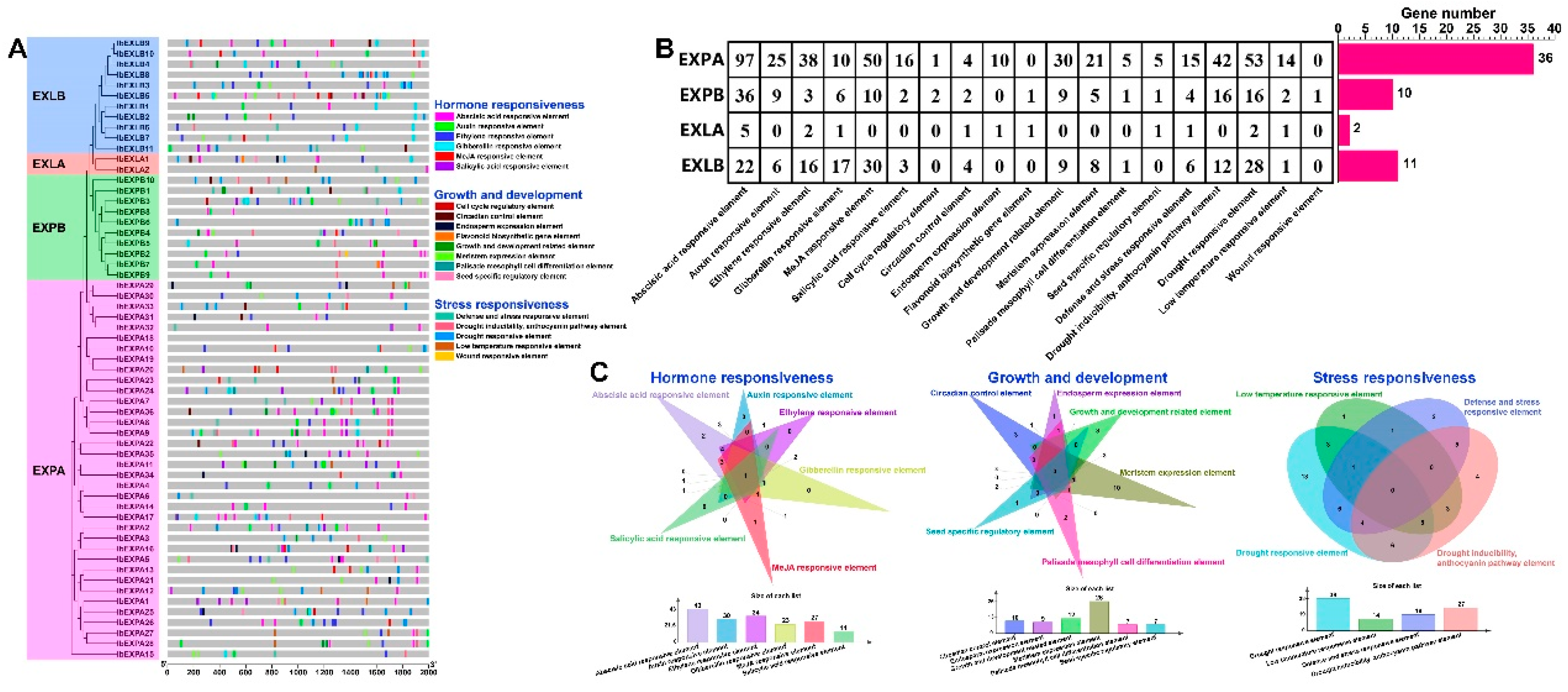
Figure 7.
Expression profiles analysis of 8 IbEXPs in different tissues by qRT-PCR. L, mature leaves at 60 dap; S, stems at 60 dap; DR1, tuberous roots at 30 dap; DR2, tuberous roots at 60 dap; DR3, tuberous roots at 100 dap; MR, mature tuberous roots at 120 dap. Bars indicate the mean of three biological replicates ± SE.
Figure 7.
Expression profiles analysis of 8 IbEXPs in different tissues by qRT-PCR. L, mature leaves at 60 dap; S, stems at 60 dap; DR1, tuberous roots at 30 dap; DR2, tuberous roots at 60 dap; DR3, tuberous roots at 100 dap; MR, mature tuberous roots at 120 dap. Bars indicate the mean of three biological replicates ± SE.
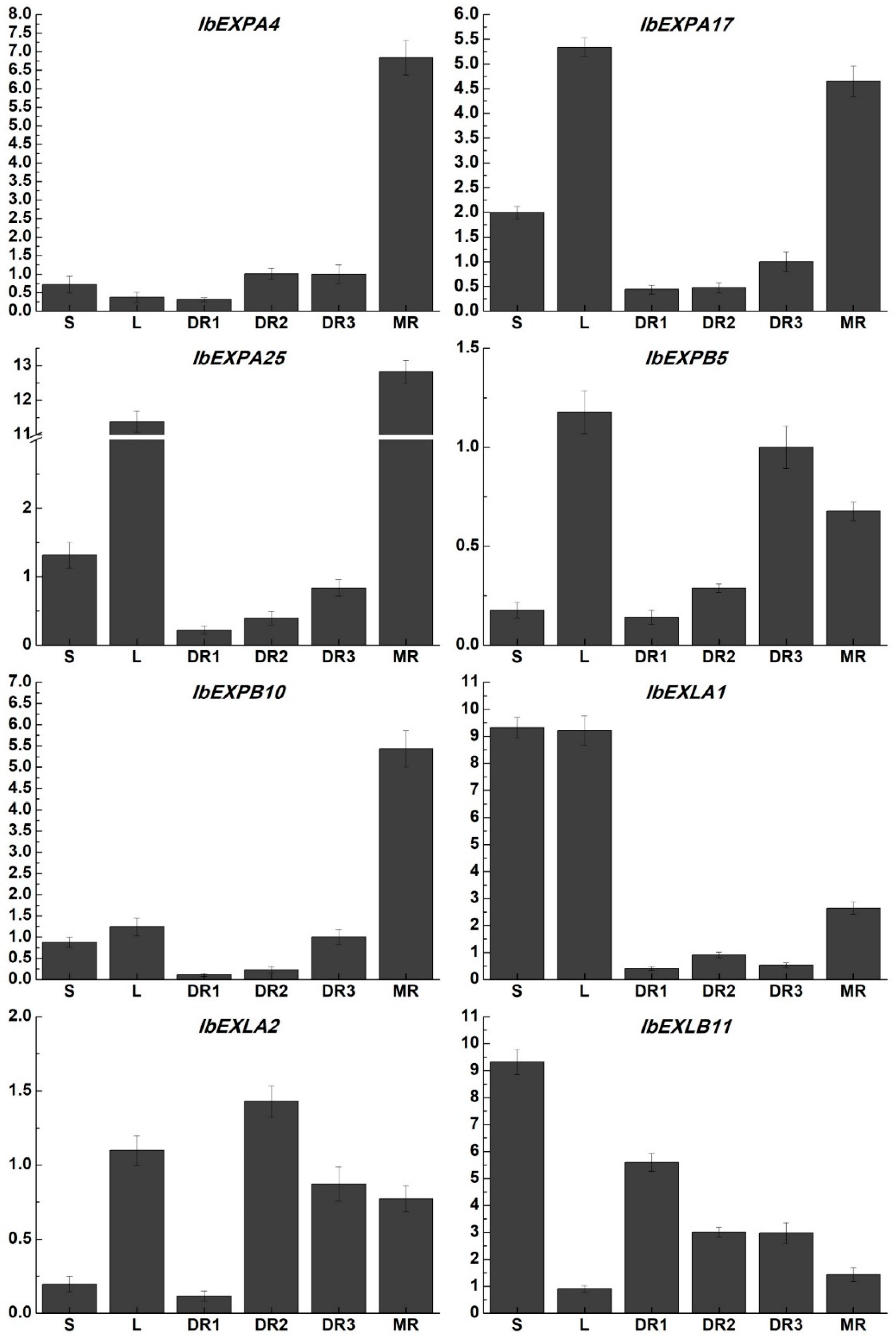
Figure 8.
Relative expression levels of 8 IbEXPs detected by qRT-PCR under diverse hormone treatments. These hormone treatments include ABA (100 μM), ACC (100 μM), JA (100 μM) and SA (2mM). 0h represent the WT seedlings that were not treated by each treatment. Bars indicate the mean of three biological replicates ± SE. The two-fold expression changes of IbEXPs in each treated sample compared to 0h sample is considered to be the significant expression changes.
Figure 8.
Relative expression levels of 8 IbEXPs detected by qRT-PCR under diverse hormone treatments. These hormone treatments include ABA (100 μM), ACC (100 μM), JA (100 μM) and SA (2mM). 0h represent the WT seedlings that were not treated by each treatment. Bars indicate the mean of three biological replicates ± SE. The two-fold expression changes of IbEXPs in each treated sample compared to 0h sample is considered to be the significant expression changes.
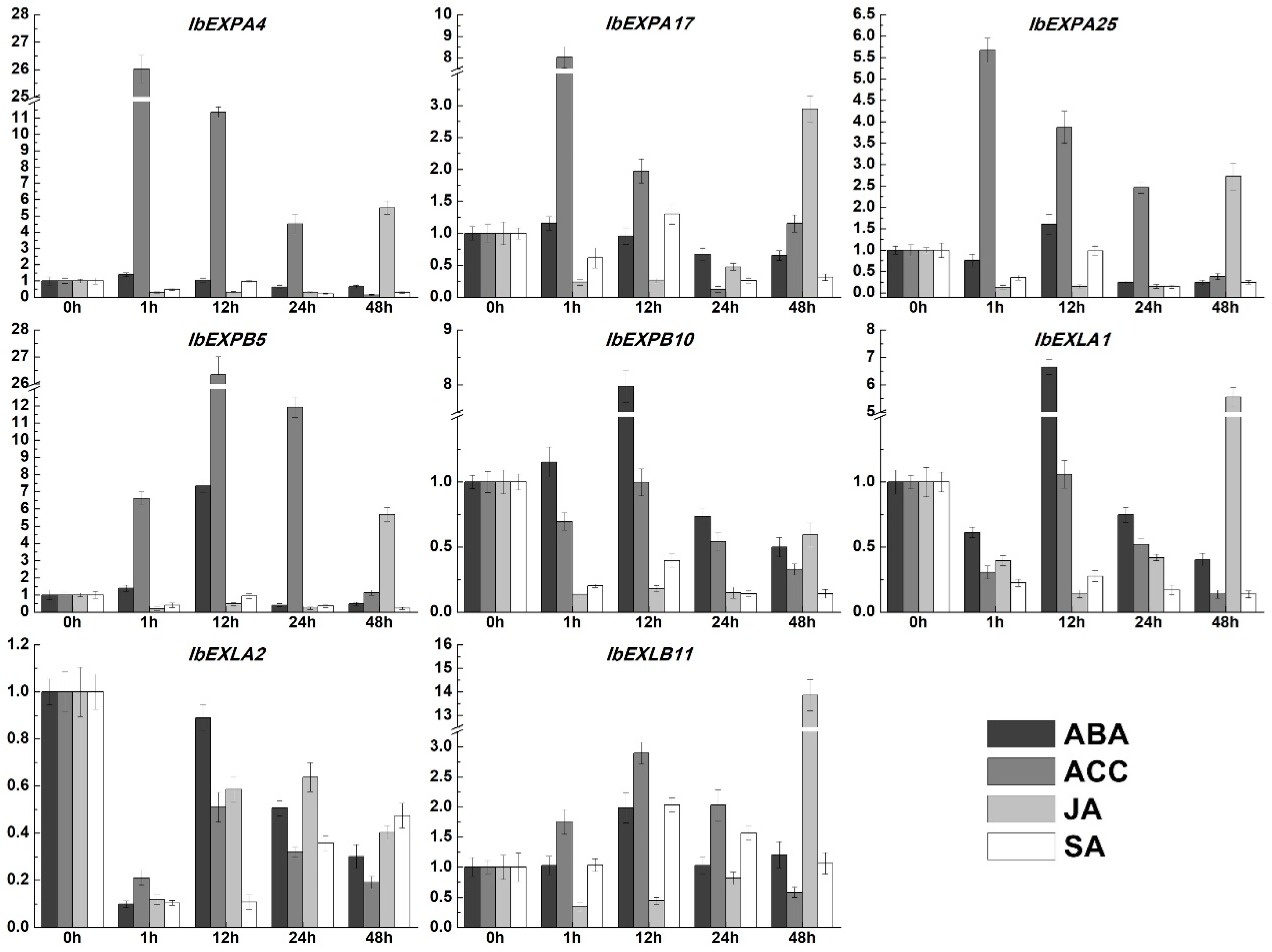
Figure 9.
Expression profiles analysis of 8 IbEXPs under abiotic stresses by qRT-PCR. The abiotic stress treatments include low temperature (LT) and high temperature (HT). 0h represent the WT seedlings that were not treated by each treatment. Bars indicate the mean of three biological replicates ± SE. The two-fold expression changes of IbEXPs in each treated sample compared to 0h sample is considered to be the significant expression changes.
Figure 9.
Expression profiles analysis of 8 IbEXPs under abiotic stresses by qRT-PCR. The abiotic stress treatments include low temperature (LT) and high temperature (HT). 0h represent the WT seedlings that were not treated by each treatment. Bars indicate the mean of three biological replicates ± SE. The two-fold expression changes of IbEXPs in each treated sample compared to 0h sample is considered to be the significant expression changes.
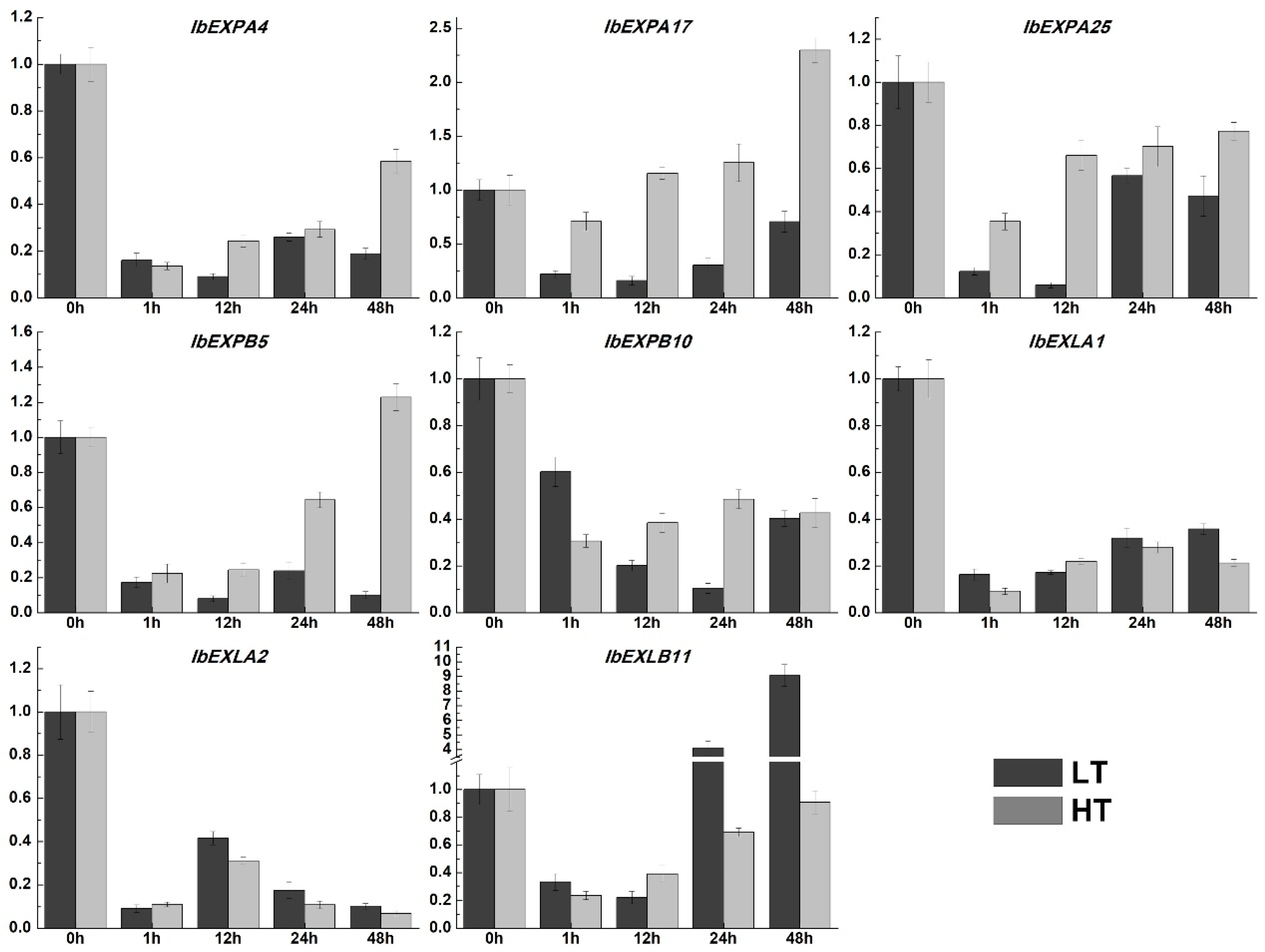

Table 1.
Characteristics of IbEXP proteins in Ipomoea batatas.
| Gene name | Gene ID | Amino acids | MW (Da) | PI | Subcellularlocation | phosphorylation cite | |||
|---|---|---|---|---|---|---|---|---|---|
| Ser site (S) | Tyr cite (Y) | Thr cite (T) | Total | ||||||
| IbEXPA1 | g1306.t1 | 238 | 25431.71 | 9.53 | Cell wall. | 20 | 3 | 8 | 31 |
| IbEXPA2 | g3925.t1 | 251 | 26960.41 | 8.07 | Cell wall. | 14 | 4 | 8 | 26 |
| IbEXPA3 | g3926.t1 | 251 | 26740.92 | 8.36 | Cell wall. | 21 | 4 | 6 | 31 |
| IbEXPA4 | g4923.t1 | 260 | 28213.08 | 9.27 | Cell wall. | 26 | 3 | 7 | 36 |
| IbEXPA5 | g9889.t1 | 310 | 33660.23 | 9.25 | Cell wall. | 26 | 5 | 20 | 51 |
| IbEXPA6 | g10004.t1 | 258 | 27595.27 | 9.3 | Cell wall. | 19 | 3 | 4 | 26 |
| IbEXPA7 | g15786.t1 | 327 | 35627.31 | 9.29 | Cell wall. | 45 | 5 | 12 | 62 |
| IbEXPA8 | g15816.t1 | 272 | 29860.26 | 9.36 | Cell wall. | 25 | 4 | 12 | 41 |
| IbEXPA9 | g15820.t1 | 264 | 28775.77 | 9.37 | Cell wall. | 29 | 4 | 9 | 42 |
| IbEXPA10 | g16869.t1 | 263 | 28645.8 | 8.56 | Cell wall. | 17 | 4 | 13 | 34 |
| IbEXPA11 | g17886.t1 | 258 | 27852.84 | 9.5 | Cell wall. | 13 | 4 | 10 | 27 |
| IbEXPA12 | g18537.t1 | 591 | 63172.9 | 7.19 | Cell wall. | 77 | 12 | 20 | 109 |
| IbEXPA13 | g18539.t1 | 341 | 35684.91 | 7.05 | Cell wall. | 57 | 6 | 10 | 73 |
| IbEXPA14 | g20283.t1 | 260 | 28086.01 | 9.4 | Cell wall. | 20 | 2 | 12 | 34 |
| IbEXPA15 | g24580.t1 | 252 | 27413.57 | 6.42 | Cell wall. | 12 | 4 | 11 | 27 |
| IbEXPA16 | g25447.t1 | 260 | 27406.89 | 9.16 | Cell wall. | 45 | 3 | 8 | 56 |
| IbEXPA17 | g25606.t1 | 209 | 22967.29 | 9.73 | Cell wall. | 13 | 3 | 8 | 24 |
| IbEXPA18 | g29706.t1 | 355 | 39001.06 | 9.64 | Cell wall. | 47 | 2 | 9 | 58 |
| IbEXPA19 | g29707.t1 | 670 | 74052.16 | 8.27 | Nucleus. | 72 | 5 | 23 | 100 |
| IbEXPA20 | g30029.t1 | 202 | 22488.71 | 9.44 | Cell wall. | 12 | 2 | 12 | 26 |
| IbEXPA21 | g33328.t1 | 273 | 28718.04 | 6.2 | Cell wall. | 30 | 5 | 7 | 42 |
| IbEXPA22 | g38554.t1 | 493 | 53544.79 | 8.24 | Cell wall, Chloroplast | 53 | 9 | 7 | 69 |
| IbEXPA23 | g39420.t1 | 309 | 33789.29 | 9.29 | Cell wall. | 29 | 4 | 20 | 53 |
| IbEXPA24 | g39467.t1 | 270 | 29209.17 | 9.1 | Cell wall. | 26 | 5 | 11 | 42 |
| IbEXPA25 | g42794.t1 | 248 | 26037.26 | 9.34 | Cell wall. | 16 | 2 | 7 | 25 |
| IbEXPA26 | g47278.t1 | 246 | 25903.09 | 8.64 | Cell wall. | 18 | 4 | 3 | 25 |
| IbEXPA27 | g53361.t1 | 253 | 27102.64 | 8.87 | Cell wall. | 22 | 3 | 6 | 31 |
| IbEXPA28 | g53365.t1 | 563 | 61262.75 | 9.32 | Cell wall. | 56 | 5 | 25 | 86 |
| IbEXPA29 | g58047.t1 | 241 | 26303.82 | 6 | Cell wall. | 23 | 3 | 13 | 39 |
| IbEXPA30 | g58048.t1 | 240 | 26373.45 | 9.21 | Cell wall. | 22 | 5 | 14 | 41 |
| IbEXPA31 | g58049.t1 | 183 | 20173.91 | 6.81 | Cell wall. | 10 | 2 | 12 | 24 |
| IbEXPA32 | g58052.t1 | 388 | 43264.45 | 9.2 | Cell wall. | 38 | 2 | 15 | 55 |
| IbEXPA33 | g58053.t1 | 521 | 58831.3 | 6.61 | Cell wall, Chloroplast | 26 | 9 | 27 | 62 |
| IbEXPA34 | g58951.t1 | 257 | 27894.88 | 9.82 | Cell wall. | 17 | 2 | 7 | 26 |
| IbEXPA35 | g60585.t1 | 237 | 25641.21 | 9.35 | Cell wall. | 20 | 2 | 9 | 31 |
| IbEXPA36 | g61698.t1 | 264 | 28887.9 | 9.39 | Cell wall. | 28 | 4 | 10 | 42 |
| IbEXPB1 | g9145.t1 | 207 | 21810.79 | 9.49 | Cell wall. | 22 | 1 | 10 | 33 |
| IbEXPB2 | g24143.t1 | 243 | 25960.18 | 5.75 | Cell wall. | 31 | 4 | 4 | 39 |
| IbEXPB3 | g24144.t1 | 400 | 44133.02 | 7.09 | Cell wall. | 56 | 7 | 21 | 84 |
| IbEXPB4 | g27129.t1 | 308 | 33145.35 | 7.52 | Cell wall. | 47 | 5 | 20 | 72 |
| IbEXPB5 | g27144.t1 | 265 | 28079.59 | 6.19 | Cell wall. | 37 | 2 | 5 | 44 |
| IbEXPB6 | g27334.t1 | 342 | 36757.24 | 5.16 | Cell wall. | 50 | 5 | 7 | 62 |
| IbEXPB7 | g27336.t1 | 256 | 27320.01 | 8.37 | Cell wall. | 31 | 5 | 3 | 39 |
| IbEXPB8 | g48839.t1 | 266 | 27877.16 | 4.89 | Cell wall. | 35 | 3 | 8 | 46 |
| IbEXPB9 | g48841.t1 | 336 | 36036.73 | 6.69 | Cell wall. | 32 | 5 | 6 | 43 |
| IbEXPB10 | g54432.t1 | 262 | 28422.34 | 8.74 | Cell wall. | 19 | 1 | 14 | 34 |
| IbEXLA1 | g904.t1 | 266 | 28622.45 | 5.29 | Cell wall. | 22 | 4 | 9 | 35 |
| IbEXLA2 | g59839.t1 | 268 | 29409.37 | 6.96 | Cell wall. | 17 | 4 | 12 | 33 |
| IbEXLB1 | g14386.t1 | 252 | 28275.14 | 5.45 | Cell wall. | 12 | 14 | 10 | 36 |
| IbEXLB2 | g14427.t1 | 443 | 49325.01 | 8.93 | Cell wall. | 37 | 15 | 21 | 73 |
| IbEXLB3 | g14812.t1 | 351 | 38060.5 | 7.91 | Cell wall. | 40 | 10 | 21 | 71 |
| IbEXLB4 | g15079.t1 | 284 | 30949.46 | 8.57 | Cell wall. | 29 | 4 | 8 | 41 |
| IbEXLB5 | g15300.t1 | 247 | 26876.61 | 8.32 | Cell wall. | 20 | 7 | 6 | 33 |
| IbEXLB6 | g33144.t1 | 251 | 28135.78 | 5.41 | Cell wall. | 17 | 15 | 10 | 42 |
| IbEXLB7 | g33212.t1 | 278 | 31034.24 | 5.52 | Cell wall. | 17 | 13 | 10 | 40 |
| IbEXLB8 | g55672.t1 | 272 | 28559.26 | 6.58 | Cell wall. | 24 | 5 | 17 | 46 |
| IbEXLB9 | g55673.t1 | 247 | 27029.36 | 9.33 | Cell wall. | 30 | 8 | 18 | 56 |
| IbEXLB10 | g55674.t1 | 246 | 26984.94 | 8.46 | Cell wall. | 16 | 14 | 13 | 43 |
| IbEXLB11 | g55709.t1 | 308 | 32778.84 | 4.63 | Cell wall. | 24 | 7 | 12 | 43 |
Table 2.
Summary of each expansin subfamily in 15 plant species.
| Species | EXPA | EXPB | EXLA | EXLB | Total | Reference |
|---|---|---|---|---|---|---|
| Ipomoea batatas | 36 (61%) | 10 (17%) | 2 (3.4%) | 11 (18.6%) | 59 | In this study |
| Ipomoea trifida | 23 (62.2%) | 4 (10.8%) | 2 (5.4%) | 8 (21.6%) | 37 | [55] |
| sugarcane | 51 (55.4%) | 38 (41.3%) | 3 (3.3%) | 0 (0%) | 92 | [4] |
| Oryza sativa | 34 (58.6%) | 19(32.8%) | 4 (6.9%) | 1 (1.7%) | 58 | [15] |
| Triticum aestivum | 121 (50.2%) | 104 (43.2%) | 16 (6.6%) | 0 (0%) | 241 | [57] |
| maize | 36 (40.9%) | 48 (54.5%) | 4 (4.5%) | 0 (0%) | 88 | [58] |
| barley | 24 (52.2%) | 16 (34.8%) | 6 (13%) | 0 (0%) | 46 | [59] |
| Brachypodium distachyon | 30 (79%) | 4 (10.5%) | 3 (7.9%) | 1 (2.6%) | 38 | [60] |
| Arabidopsis | 26 (72.2%) | 6 (16.7%) | 3 (8.3%) | 1 (2.8%) | 36 | [15] |
| soybean | 49 (65.3%) | 9 (12%) | 2 (2.7%) | 15 (20%) | 75 | [61] |
| Ginkgo biloba | 32 (69.5%) | 4 (8.7%) | 5 (10.9%) | 5 (10.9) | 46 | [62] |
| cotton | 67 (72%) | 8 (8.6%) | 6 (6.5%) | 12 (12.9%) | 93 | [63] |
| tobacco | 36 (69.2%) | 6 (11.5%) | 3 (5.8%) | 7 (13.5%) | 52 | [64] |
| potato | 24 (66.6%) | 5 (13.9%) | 1 (2.8%) | 6 (16.7%) | 36 | [71] |
| tomato | 25 (65.8%) | 8 (21.1%) | 1 (2.6) | 4 (10.5%) | 38 | [72] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



