Submitted:
28 December 2023
Posted:
29 December 2023
You are already at the latest version
Abstract
This study investigated whether metabolomic fingerprints of bovine embryo growth media improve the prediction of successful embryo implantation. In this prospective cohort study, the metabolome from in vitro produced Day 7 blastocysts with successful implantation (n=11), blas-tocysts with failed implantation (n=10), and plain culture media without embryos (n=5) was in-cluded. Samples were analyzed using AbsoluteIDQ® p180 Targeted Metabolomics Kit with LC-MS/MS, and a total of 189 metabolites were analyzed from each sample. Blastocysts that re-sulted in successful embryo implantation had significantly higher levels of methionine sulfoxide (p<0.001), DOPA (p<0.05), spermidine (p<0.001), acetylcarnitine to free carnitine ratio (p<0.05), C2+C3 to free carnitine ratio (p<0.05), and lower levels of threonine (nep<0.001) and phosphati-dylcholine PC ae C30:0 (p<0.001) compared to control media. However, when compared to embryos that failed to implant, only DOPA, spermidine, C2/C0, (C2+C3)/C0 and PC ae C30:0 levels differentiated significantly. In summary, our study identifies a panel of differential metabolites in the culture media of bovine blastocysts that could act as potential biomarkers for the selection of viable blastocysts before embryo transfer.
Keywords:
bovine
; embryo
; metabolomics
; in vitro
; pregnancy
; biomarkers
1. Introduction
Fertility in cattle has decreased dramatically in recent decades and is currently one of the major challenges in the dairy industry [1]. Assisted reproductive technologies (ART) play a crucial role in addressing low fertility and have been widely used in cattle worldwide to enhance the reproduction of animals with high genetic value [2,3]. Moreover, bovine in vitro embryo production (IVEP) enables the use of sub-fertile donors and repeat breeders. IVEP has evolved into a significant international business with over a million transfers of in vitro produced bovine embryos recorded worldwide [4].
Successful conception, embryo survival, and the birth of a healthy calf are major factors influencing the production and economic efficiency of a beef or dairy farm. Several conditions are known to lead to implantation failure after embryo transfer (ET), including the health status of the recipient animal, reduced endometrial receptivity, genetic abnormalities of an embryo, errors in the embryo transfer technique, or a combination of these influences [5]. One of the first essential steps in achieving a successful pregnancy is the selection and transfer of highly competent in vitro produced embryos, which is critical for the application of IVF technology in large animal breeding toolbox.
There are currently three main trending research areas in the embryo industry: time-lapse microscopy empowered by AI-based computational modelling [6]; the analysis of extracellular vesicles (EVs) secreted by blastocysts [7]; and lastly, the correlation of metabolomic/proteomic markers in spent culture medium to embryo viability [8,9,10,11].
In practice, however, the most prevalent method to assess bovine embryos' developmental potential in vitro is to determine cleavage rate paired with morphological assessment, which sadly is a highly subjective method based on the experience of the embryologist [12]. Even though the first calf after the transfer of an IVEP embryo was born 40 years ago, the selection of viable IVEP embryos in the livestock industry is still insufficient, and the industry is missing a reliable non-invasive, and cost-effective technique to assess embryos before ET [13]. Low IVEP embryo transfer success rates inevitably bring higher overall cost per calf born.
Despite a considerable amount of promising research results, a practical and accurate non-invasive assessment system has not yet been developed for the cattle industry [14]. The need for a fast, precise, and practical method of determining embryo viability and predicting its ability to implant has driven years of research directed at finding the most suitable method for clinical application.
The outstanding advances in non-invasive metabolomic analysis in recent years, enabling the assessment of single embryo physiology, are now swiftly becoming a part of everyday work in human IVF labs and hopefully will soon be applied in bovine IVF labs as well. Changes in embryo metabolism during early development mirror cellular metabolic activities and the overall viability of the embryo [15]. As the metabolome reflects the end-products of embryos' physiological activities, our primary objective in this study was to evaluate the metabolome through metabolomics and explore its relationship with successful pregnancy outcomes. This approach enhances our comprehension of the health status in pre-implantation bovine embryos.
Given this robust background, the present study aimed to test the effectiveness of targeted metabolomic analysis using the AbsoluteIDQ® p180 Targeted Metabolomics Kit coupled with liquid chromatography-tandem mass spectrometry (LC-MS/MS). This approach allows for the comprehensive capture of a broad range of molecular content in individually cultured bovine embryo culture media. We hypothesized that metabolomic profiles of embryos with successful implantation differ from those of embryos with failed implantation. To the best of our knowledge, this study is the first of its kind to utilize the AbsoluteIDQ® p180 Targeted Metabolomics Kit to determine metabolomic profiles of individually cultured bovine IVEP embryos, aiming to identify metabolomic signatures associated with successful implantation and those associated with failed implantation. A non-invasive approach for predicting embryo viability could offer multiple benefits, reducing the risk of impairing the embryo and its environment, thereby maintaining development potential. Additionally, it has the potential to lower the overall cost of IVEP and embryo transfer by contributing to higher pregnancy rates and reducing the number of open recipients [16].
2. Materials and Methods
2.1. Media
Serum-free media for every step of in vitro embryo production, including in vitro maturation, fertilization, and individual cultivation of embryos, were obtained from IVF Limited T/A IVF Bioscience (Bickland Industrial Park, Falmouth, Cornwall, TR11 4TA, United Kingdom), unless stated otherwise. Care was taken to select media for all experiments from the same production batch.
2.2. Experimental design
We embraced embryos produced from OPU-derived oocytes in our study to enable the transfer of such embryos and to compare the metabolic fingerprints of blastocysts resulting in pregnancy with those that did not.
2.3. Oocyte retrieval and in vitro maturation
Transvaginal ultrasound-guided follicular puncture (Ovum Pick-Up; OPU) and in vitro embryo production were employed to generate blastocysts for embryo transfers. Holstein donor cows, pregnant between 45 and 100 days of gestation, and non-pregnant Holstein donor cows, 40 to 90 days after parturition, were utilized in the OPU procedure. No superovulation of donor cows was used before OPU. Epidural anesthesia was administered using Xylazine (0.05 mg/kg body weight). Ovaries were scanned with a 7.5 MHz micro-convex transducer, and follicles ≥ 3 mm were punctured using an 18G needle coupled to the aspiration device and vacuum system. Follicular fluid, mixed with BO-OPU media, was transported to the laboratory within one hour.
Cumulus-oocyte complexes (COCs) with three or more layers of unexpanded cumulus cells and morphologically bright, evenly granulated cytoplasm were selected for in vitro maturation. COCs were washed thrice in HEPES buffered medium (BO-Wash) and once with maturation medium (BO-IVM). COCs from each donor cow were matured separately in four-well plates (Nunclon, Roskilde, Denmark), each containing 500 µl BO-IVM medium. Oocytes were incubated at 38.5 ˚C with 5% CO2 in humidified air for 24 h.
2.4. In vitro fertilization
Frozen-thawed semen (commercially produced at Animal Breeders' Association of Estonia, Keava) from a single bull was prepared by thawing straws in water at 35 ˚C for 30 seconds. In vitro fertilization (IVF) was conducted using semen from the bull EHF ZIARD 27481 (ID EE13993023). Semen from one straw was gently expelled into a 15 ml centrifuge tube containing 3 ml of BO-SemenPrep medium and centrifuged for 5 minutes at 328 x g at 23 ˚C. The supernatant was removed, 3 ml of BO-SemenPrep was added, and the same centrifugation process was repeated. Matured oocytes were washed once in BO-IVF medium and placed in four-well plates with 500 µl BO-IVF medium per well. The cumulus-oocyte complexes (COCs) and sperm were co-incubated at a final concentration of approximately 1 × 106 motile sperm/ml at 38.5 ˚C with 5% CO2 in air with maximum humidity for 18 h.
2.5. In vitro cultivation and embryo transfer
Presumptive zygotes were denuded 18 hours after IVF by vortexing for 120 seconds in BO-Wash medium, rinsed thrice in BO-Wash medium, and once in BO-IVC medium. Zygotes were cultured in 60 µl BO-IVC culture medium droplets (one zygote per droplet) overlaid with mineral oil in 100 mm Petri dishes (Sigma-Aldrich) at 38.5˚C under an atmosphere of 5% CO2, 5% O2, and 90% N2 for up to 7 days. Controls were created by placing 60 µl of plain BO-IVC culture media droplets that were never in contact with an embryo, under oil, and were kept under the same conditions as zygotes (38.5 ˚C under an atmosphere of 5% CO2, 5% O2, 90% N2). Fresh Day 7 IETS Grade 1 blastocysts were transferred to the synchronized EHF recipient heifers using BO-Transfer media.
2.6. Collection of media for LC-MS/MS and categorization of samples
20 µl of culture media samples were collected from droplets of individually cultured OPU-derived Day 7 blastocysts. Morphological assessments were conducted on each embryo at the time of media collection, adapted from previously published work [17,18]. Any embryos that were arrested in development by Day 7 were excluded from the experiment. Subsequently, samples were categorized according to their pregnancy outcomes: “ET-P” for blastocysts resulting in pregnancy and “ET-NP” for blastocysts that failed to result in pregnancy at Day 30 following embryo transfer. Plain media samples were also collected on Day 7 and processed as controls (hereafter referred to as “Control”), following the approach adopted from previous research [19]. All collected media samples were labeled and stored immediately after collection at -20°C.
2.7. Preparation of culture media samples for LC-MS/MS and spectrometry
Each 20 µl culture media sample was prepared following The AbsoluteIDQ® p180 Targeted Metabolomics Kit sample preparation protocol for Agilent Infinity high-performance liquid chromatography (Agilent, CA, USA) coupled to the 4500 QTRAP® ion trap mass spectrometer (Sciex, MA, USA).
2.8. Statistical analysis
Based on the raw chromatography-mass spectrometry data, all missing values were treated as such due to being below the detection limit and were subsequently substituted with zero. Univariate comparisons of each metabolite or derived indexes (metabolite sums or ratios) between different media samples were conducted. Metabolites with more than 30% missing values were compared using the Chi-square test to determine whether the metabolite is significantly more frequently detectable in any of the study groups. If less than 30% of values were missing, the Shapiro-Wilk test was used to assess whether the data resembled a normal distribution. Depending on this test, either ANOVA with Tukey honest significance post-hoc or Kruskal-Wallis with Dunn post-hoc test was applied. Results were considered statistically significant at p < 0.001, and p-values between 0.05 and 0.001 were considered borderline significant. All statistical analyses were performed, and all figures were constructed using R version 4.2.0 (R Foundation for Statistical Computing, Vienna, Austria).
3. Results
A total of 26 spent media samples were analyzed using the AbsoluteIDQ® p180 Targeted Metabolomics Kit with LC-MS/MS, covering a total of 189 metabolites: 40 acylcarnitines, 42 amino acids/biogenic amines, 91 phospholipids, 15 sphingolipids, and the sum of hexoses (Supplemental Table S1). The culture media samples were derived from 11 Day 7 ET blastocysts with confirmed pregnancy (ET-P BL), 10 Day 7 ET blastocysts with failed implantation (ET-NP BL), and 5 Day 7 plain growth media (CM) for control.
3.1. Metabolites Differing in Culture Media of Blastocysts and Controls
In total, 8 metabolites showed significant differences (p < 0.001), and 27 exhibited borderline significance (p between 0.05 and 0.001) between the blastocyst culture media and empty controls (Table 1). Spermidine was not detectable in the empty media (control) but was found in most of the media from pregnancy-yielding blastocysts and in a few media samples from blastocysts without pregnancy.
Among amino acids, only threonine (Thr) was significantly consumed by blastocysts. Glutamine (Gln) was secreted into the culture media (p = 0.007), and dihydroxyphenylalanine (DOPA), a metabolite of phenylalanine (Phe) and tyrosine (Tyr), was detectable in 64% of ET-P samples and 20% of ET-NP culture media samples, while being completely absent in empty media. Additionally, oxidized methionine (Met-SO) levels rose in media containing blastocysts.
Among acylcarnitines, methylglutaryl- (C5M-DC), and decanoylcarnitines (C10) were increased in media containing blastocysts. Interestingly, pimeloylcarnitine (C7-DC) levels were reduced in media containing blastocysts (Table 1). Blastocysts secreted carnitine esters with C4:1, C4, C5, C18:2, C5:1-dicarboxylic acid, and hydroxylated C14:1-OH, C16:1-OH, C18:1-OH fatty acid residues into the culture media with borderline significances.
Thirteen phosphatidylcholines (PCs) out of the measured 76 and 3 lysophosphatidylcholines (lysoPCs) out of the 14 measured ones had at least borderline significance in groupwise comparison. Both increases and decreases in their concentration in culture media were observed due to blastocyst presence (Figure 1). PC ae C30:0 and PC aa C28:1 in culture media were significantly depleted by blastocysts, whereas the concentration of PC ae C30:3 was distinct in all three experimental groups.
3.2. Derived Indicators Differing Between Culture Media of Blastocysts and Controls
Metabolites of the same class share metabolic pathways. Summary concentrations of these, or ratios of a pathway's start and end product or enzyme substrate and product, may be more informative than individual metabolites. Forty-four sums or ratios of metabolites were calculated (Supplemental Table S1). The ratio of oxidized methionine to methionine was the only calculated indicator of metabolic activity that significantly differentiated control media from media containing blastocysts (Table 2). The summary concentration of all PCs, mainly due to PC aa type lipids and saturated fatty acid residues, had borderline significance. The relative amount of dicarboxylic acid esters to all carnitine esters also had borderline significance between the groups.
3.3. Metabolites Differing Between Media of Pregnancy-Yielding and Unyielding Blastocysts
Post-hoc analysis revealed significant differences in PC ae C30:0 concentrations among all three groups (Table 1). Among the metabolites with borderline significance, PC aa C42:1, PC ae C34:3, and linoleic acid carnitine ester differentiated in the ET-P BL and ET-NP BL groups. All PC-s had a lower concentration in the ET-P BL group, and the carnitine ester had a higher concentration in the culture media of ET-P BL group. The total concentration of PC aa was significantly lower in the culture media of pregnancy-yielding blastocysts than in the ET-NP BL group and controls (Table 2). The acetylcarnitine to free carnitine ratio was significantly higher in the ET-P BL group than in either of the other groups.
4. Discussion
Maximizing pregnancy outcomes relies on the non-invasive assessment of blastocysts, and metabolomic characterization of pre-implantation growth media has shown some success. Although various metabolites have been proposed as embryo viability markers, the results are scarce, and reproducibility seems to be a challenge. Certainly, the development and putative markers may be influenced by the experimental setup and many other factors [20].
4.1. Glucose Metabolism
The dominant hexose in growth media is glucose, and, as expected, both ET-P BL and ET-NP BL blastocysts consumed a fraction of it in our experiment. Glucose serves as a substrate for ATP and NADPH production once the biosynthesis gears up after the initial embryo cleavages [21]. In some studies, the level of glucose consumption has been linked to the quality and success of pregnancy [22]. However, glucose usage intensity of embryos was not correlated with pregnancy in our results.
4.2. Amino Acid Metabolism
Amino acids are utilized in the de novo synthesis of nitrogen-containing compounds (e.g., nucleobases and heme) and lipids (serving as precursors for sphingolipids and choline), and, of course, proteins. Additionally, they can serve as a source of ATP. Essential amino acids cannot be synthesized by mammals; thus, their levels are expected to decrease in growth media. Glutamate (Glu), Gln, and Ala are central in transamination, thereby linking the metabolisms of nitrogen-containing and nitrogen-free metabolism. Depending on blastocyst activity, their levels in growth media may also increase, even if nitrogen waste is in other forms [11].
It has been previously shown that the consumption of essential amino acids can vary among embryos. Two decades ago, it was discovered that embryos require different amino acids at various developmental stages [23,24]. Gln, arginine (Arg), Met, Ala, asparagine (Asn), serine (Ser), and leucine (Leu) were highlighted then. However, there were disagreements between the early reports, possibly due to suboptimal culture conditions such as oxygen levels in incubators and the absence of serum in culture media, which may have led to erroneous conclusions [25].
In a recent bovine pregnancy prediction study [26] it was found that Glu, proline (Pro), Met, Arg, lysine (Lys), and Thr concentrations in embryo culture media have the best pregnancy-predicting power, but only when a single metabolite was used in the assessment. Lechniak et al. [10] compared different culture setups and found distinct differences in metabolic pathways of Phe, Tyr, Met, aspartate (Asp), Arg, Pro, and histamine (His) with regard to the culture system used. In the same study, cysteine (Cys), Ala, Glu, glycine (Gly), Ser, Phe, Tyr, Lys, and Asp-dependent pathways appeared to be associated with zygotic cleavage pattern, serving as potential viability indicators. While these lists may seem extensive, many amino acids are directly related to each other; for example, Phe and Tyr probably mark a single process. Ser and Gly can be converted to each other, potentially affecting Met levels. Ser and Gly also relate to Ala and Thr in one or two-step conversions [27].
In our study, most of the amino acids did not differ significantly between media containing blastocysts and empty control media, indicating that the consumption and secretion of amino acids by embryos were relatively modest compared to their levels in growth media. This aligns with the theory that viable bovine embryos exhibit a 'quiet' metabolism rather than an 'active' one, although the limits of this quietness are yet to be defined, and there is still much to be learned [28]. Similarities between the successful and failed implantation groups could also be attributed to the selection of embryos with similar morphological qualities for embryo transfer. Interestingly, Thr, an essential amino acid, was a striking exception. Thr levels were significantly lower in media from successfully implanted embryos relative to empty control media; however, we did not observe significant differences between the successful and failed implantation groups. This finding is consistent with other recent works [8].
Whether Thr is indeed more special than other amino acids remains to be confirmed, but one theory suggests that threonine dehydrogenase (TDH) might be key for bovine blastocyst development [29]. This enzyme uses Thr to produce a one-carbon fragment and reduced NADH. Both products can be further used in anabolic processes. Met, a usual donor of one-carbon fragments for methylation reactions, can thus be reserved for other uses, such as sulfur-based components of the antioxidant system.
Thr plays an important role in the growth and proliferation of embryonic stem cells (ES) in pre-implantation embryos, where it is converted to Gly and acetyl−CoA by TDH. Gly and acetyl-CoA aid nucleotide biosynthesis, epigenetic modifications, and mitochondrial free energy conversions in ES cells [30].
In relation to Met, an essential amino acid whose metabolism has been prominently featured in recent blastocyst quality assessments and embryo implantation studies [31], it is noteworthy that numerous studies have confirmed an increase in the concentration of Met in viable embryos [32] as well as in the uterine lumen of pregnant animals [33,34]. Met is oxidized to Met-SO, and its levels, which were significantly different in our study, are considered to protect cells from oxidative damage, although a more complex bioregulatory role cannot be excluded [35]. Specifically, we observed that Met-SO levels were significantly higher in media from successfully implanted embryos and in the failed implantation group, relative to the empty control media.
Aromatic amino acids did not exhibit any significant distinctions, except for DOPA, which was unexpectedly measurable in 64% of samples from embryos with successful pregnancies and in 20% of samples from embryos with implantation failure, while being completely absent in empty culture media. This suggests that DOPA is synthesized and secreted into the culture media by embryos. To date, no study has demonstrated the metabolism of DOPA or its role in early-developing bovine embryos. However, it is well-known that catecholamine synthesis in the fetus begins at mid-gestation with the conversion of Tyr into 3,4-dihydroxyphenylalanine, mediated by the enzyme tyrosine hydroxylase (TH). DOPA can subsequently be converted to dopamine by aromatic L-amino acid decarboxylase [36]. While previous studies have provided little evidence for any role of DOPA in early embryo development, further research is needed to investigate this peculiar finding. Catecholamine precursors such as Phe and Tyr or related compounds are commonly reported in metabolomic studies [9,10]. The role of catecholamines in blastocyst development has also been suggested by Wang et al. [36]. There are hints that catecholamines might regulate lipid usage [37] and the synthesis of cell differentiation-regulating polyamines [38], but this knowledge needs to be explored further before definitive conclusions can be drawn.
4.3. Polyamine Metabolism
The most outstanding difference in our study between culture media of pregnancy-yielding and non-yielding blastocysts was spermidine, a polyamine. Spermidine is a polycationic substance synthesized from Arg and Met and is known to regulate embryonic gene expression and protein synthesis, playing a crucial role in the growth and proliferation of embryos [39]. The essential role of spermidine in developing blastocysts has been reported previously [40,41]. In our experiment, spermidine was not detectable in empty media (control), but it was found in 90.91% of media from successfully implanted blastocysts and in 20% of media from embryos with failed implantation. Therefore, spermidine could be a potential biomarker for selecting viable bovine embryos for embryo transfer, as it appears to be closely related to successful implantation and pregnancy. Several gaps in our knowledge around spermidine metabolism in early bovine blastocysts could benefit from further research, including larger trial groups of transferrable embryos and a higher number of embryo transfers and recipients.
4.4. Lipid Metabolism
Most of the changes observed in any comparison among the three growth media groups were related to lipids. The multifaceted roles of lipids in early embryo development were recently reviewed by Melo-Sterza and Poehland [20]. Beta-oxidation and carnitine esterification of fatty acids emerge as particularly crucial aspects of embryo lipid metabolism [42]. In an experimental study on bovine pregnancy prediction, Gomez et al. [9] identified free fatty acids C10, C16, C18, and C18 glycerol ester as potential viability markers in later stages of embryo development. Although Lechniak et al. [10] did not report individual metabolites for embryo selection, they found that beta-oxidation of fatty acids seems highly dependent on embryo culture conditions, providing insight into the embryo's response to environmental factors.
To provide a broader perspective beyond individual lipid species, we observed that media containing blastocysts in our study were richer in oxidized (DC- dicarboxylic and OH- hydroxylated) fatty acid residues, as well as short-chain fatty acids (SCFA). This indicates active oxidation (beta or, in the case of DC variants, omega oxidation) of fatty acids during embryo development. When comparing the ET-P and ET-NP groups, differences in the concentration of long-chain acylcarnitines in culture media were noted, along with acetylcarnitine and the C2+C3 to free carnitine ratio, which was significantly higher in the culture media of blastocysts with successful implantation. The acetyl group and long-chain fatty acids mark the beginning and end of fatty acid synthesis in one direction and beta-oxidation in the other. Higher levels of their carnitine esters suggest more efficient ester synthesis. Carnitine supplementation during in vitro culture is known to improve embryo potential for implantation and successful pregnancy [43]. The stimulatory effect of carnitine on lipid metabolism has been confirmed in cattle, mice, and pig oocytes [44]. L-carnitine, a powerful antioxidant, reduces the accumulation of reactive oxygen species (ROS) and decreases apoptosis in animal cells [45]. Our results suggest that it's not just the level of carnitine itself, but the efficiency of its use—such as the acetylcarnitine to free carnitine ratio and C2+C3 to free carnitine ratio—that might be a decisive factor for blastocyst quality and subsequent implantation.
4.5. Phosphatidylcholine Metabolism
PCs play a crucial role in membrane synthesis during cell division and can also function as substrates for beta-oxidation, as well as storage for polyunsaturated fatty acids, which serve as precursors for signaling molecules [46]. PCs, being dominant membrane phospholipids, play a vital role in maintaining the structural integrity of the lipid bilayer during embryonic development [47].
In our study, there was a significant difference in PC ae C30:0 levels in culture media between blastocysts with successful and failed implantation. PC ae C30:0 is a saturated plasminogen. Additionally, the concentration of PC ae C30:3 was significantly lower in culture media of blastocysts with successful implantation compared to the ET-NP and empty culture media groups. Notably, the sum of all saturated fatty acid-containing PCs exhibited a similar trend, with lower concentrations in the ET-P group and higher concentrations in the ET-NP group.
While the biological function of PC ae C30:0 in early embryo development remains undescribed, our study is the first to investigate PC ae C30:0 levels in culture media and their association with viable embryos. Although studies in human plasma have explored PC ae C30:0 concentration and its correlation with conditions such as ovarian endometriosis and prostate cancer [48], further replication of our findings is essential before PC ae C30:0 can be considered a biomarker for selecting blastocysts with the highest implantation potential.
5. Conclusions
In conclusion, our study affirms the significance of lipid beta-oxidation, Met, aromatic amino acids, and polyamine metabolisms as pivotal processes in blastocyst preimplantation development. When considering our findings alongside existing literature, these pathways emerge as the most promising candidates for providing biomarkers to predict blastocyst viability and assess the embryo's potential for successful implantation. While individual metabolites may vary across studies, the core processes likely remain consistent, with the specific metabolite identified as a biomarker influenced by experimental conditions and culture media. Undoubtedly, our unexpected revelations regarding DOPA, spermidine, Met-SO, C2/C0, (C2+C3)/C0, and PC ae C30:0 metabolism and their association with successfully implanted bovine embryos warrant careful consideration and further investigation.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, E.Ts., K.K. and Y.J.; Methodology, E.T., K.K., E.Ta.; Software, K.K.; Validation, E.Ts., K.K.; Formal analysis, K.K.; Investigation, Y.J.; Resources E.Ts., K.K., P.P., A.V.-S., A.K.; Data curation, E.Ts., K.K.; Writing—original draft preparation, E.Ts., K.K.; writing—review and editing, E.Ts., K.K., Y.J.; Visualization, K.K.; Supervision, K.K., Y.J.; Project administration, E.Ts.; Funding acquisition, E.Ts., K.K., Y.J.; All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the European Union through the European Regional Development Fund, grant number 616215780014.
Data Availability Statement
The data presented in this study are available on reasonable request from the corresponding author. The data are not publicly available due to privacy and/or ethical concerns.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study, in the collection, analyses, or interpretation of data, in the writing of the manuscript, or in the decision to publish the results.
References
- Pohler, K.G.; Reese, S.T.; Franco, G.A.; Oliveira Filho, R. Vander; Paiva, R.; Fernandez, L.; Melo, G. de; Moraes Vasconcelos, J.L.; Cooke, R.; Poole, R.K. New Approaches to Diagnose and Target Reproductive Failure in Cattle. Anim Reprod 2020, 17, 1–19. [Google Scholar] [CrossRef]
- Mebratu, B. Embryo Transfer in Cattle Production and Its Principle and Applications. International Journal of Pharmacy & Biomedical Research 2020, 7, 40–54. [Google Scholar] [CrossRef]
- Nowicki, A. Embryo Transfer as an Option to Improve Fertility in Repeat Breeder Dairy Cows. Journal of Veterinary Research (Poland) 2021, 65, 231–237. [Google Scholar] [CrossRef]
- 2021 Statistics of Embryo Production and Transfer in Domestic Farm Animals; 2022.
- Hansen, P.J. The Incompletely Fulfilled Promise of Embryo Transfer in Cattle-Why Aren’t Pregnancy Rates Greater and What Can We Do about It? J Anim Sci 2020, 98. [Google Scholar] [CrossRef]
- Magata, F. Time-Lapse Monitoring Technologies for the Selection of Bovine in Vitro Fertilized Embryos with High Implantation Potential; Vol. 69. [CrossRef]
- Imakawa, K.; Matsuno, Y.; Fujiwara, H. New Roles for EVs, MiRNA and LncRNA in Bovine Embryo Implantation. Front Vet Sci 2022, 9. [Google Scholar] [CrossRef]
- Cheredath, A.; Uppangala, S.; Asha, C.S.; Jijo, A.; Vani Lakshmi, R.; Kumar, P.; Joseph, D.; Nagana, N.G.; Kalthur, G.; Adiga, S.K. Combining Machine Learning with Metabolomic and Embryologic Data Improves Embryo Implantation Prediction. Reproductive Sciences 2023, 30, 984–994. [Google Scholar] [CrossRef] [PubMed]
- Gomez, E.; Canela, N.; Herrero, P.; Cereto, A.; Gimeno, I.; Carrocera, S.; Martin-gonzalez, D.; Murillo, A.; Muñoz, M. Metabolites Secreted by Bovine Embryos in Vitro Predict Pregnancies That the Recipient Plasma Metabolome Cannot, and Vice Versa. Metabolites 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Lechniak, D.; Sell-Kubiak, E.; Warzych, E. The Metabolic Profile of Bovine Blastocysts Is Affected by in Vitro Culture System and the Pattern of First Zygotic Cleavage. Theriogenology 2022, 188, 43–51. [Google Scholar] [CrossRef]
- Pallisco, R.; Lazzarino, G.; Bilotta, G.; Marroni, F.; Mangione, R.; Saab, M.W.; Brundo, M.V.; Pittalà, A.; Caruso, G.; Capoccia, E.; et al. Metabolic Signature of Energy Metabolism Alterations and Excess Nitric Oxide Production in Culture Media Correlate with Low Human Embryo Quality and Unsuccessful Pregnancy. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Rubessa, M.; Wheeler, M.B. Label-Free Microscopy: A Non-Invasive New Tool to Assess Gametes and Embryo Quality. Theriogenology 2020, 150, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Ferré, L.B.; Kjelland, M.E.; Strøbech, L.B.; Hyttel, P.; Mermillod, P.; Ross, P.J. Review: Recent Advances in Bovine in Vitro Embryo Production: Reproductive Biotechnology History and Methods. Animal 2020, 14, 991–1004. [Google Scholar] [CrossRef]
- McLennan, H.J.; Saini, A.; Dunning, K.R.; Thompson, J.G. Oocyte and Embryo Evaluation by AI and Multi-Spectral Auto-Fluorescence Imaging: Livestock Embryology Needs to Catch-up to Clinical Practice. Theriogenology 2020, 150, 255–262. [Google Scholar] [CrossRef]
- Lipinska, P.; Pawlak, P.; Warzych, E. Species and Embryo Genome Origin Affect Lipid Droplets in Preimplantation Embryos. Front Cell Dev Biol 2023, 11. [Google Scholar] [CrossRef]
- Zmuidinaite, R.; Sharara, F.I.; Iles, R.K. Current Advancements in Noninvasive Profiling of the Embryo Culture Media Secretome. Int J Mol Sci 2021, 22, 1–13. [Google Scholar] [CrossRef]
- Merton, J.S.; Vermeulen, Z.L.; Otter, T.; Mullaart, E.; de Ruigh, L.; Hasler, J.F. Carbon-Activated Gas Filtration during in Vitro Culture Increased Pregnancy Rate Following Transfer of in Vitro-Produced Bovine Embryos. Theriogenology 2007, 67, 1233–1238. [Google Scholar] [CrossRef] [PubMed]
- Farin, P.W.; Farin, C.E. Transfer of Bovine Embryos Produced In Vivo or In Vitro: Survival and Fetal Development; 1995; Vol. 52. [CrossRef]
- Yagi, A.; Miyanaga, S.; Shrestha, R.; Takeda, S.; Kobayashi, S.; Chiba, H.; Kamiya, H.; Hui, S.P. A Fatty Acid Profiling Method Using Liquid Chromatography-High Resolution Mass Spectrometry for Improvement of Assisted Reproductive Technology. Clinica Chimica Acta 2016, 456, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Melo-Sterza, F. de A.; Poehland, R. Lipid Metabolism in Bovine Oocytes and Early Embryos under in Vivo, in Vitro, and Stress Conditions. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Banliat, C.; Mahé, C.; Lavigne, R.; Com, E.; Pineau, C.; Labas, V.; Guyonnet, B.; Mermillod, P.; Saint-Dizier, M. The Proteomic Analysis of Bovine Embryos Developed in Vivo or in Vitro Reveals the Contribution of the Maternal Environment to Early Embryo. BMC Genomics 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Ferrick, L.; Lee, Y.S.L.; Gardner, D.K. Metabolic Activity of Human Blastocysts Correlates with Their Morphokinetics, Morphological Grade, KIDScore and Artificial Intelligence Ranking. Human Reproduction 2020, 35, 2004–2016. [Google Scholar] [CrossRef] [PubMed]
- Brison, D.R.; Houghton, F.D.; Falconer, D.; Roberts, S.A.; Hawkhead, J.; Humpherson, P.G.; Lieberman, B.A.; Leese, H.J. Identification of Viable Embryos in IVF by Non-Invasive Measurement of Amino Acid Turnover. Human Reproduction 2004, 19, 2319–2324. [Google Scholar] [CrossRef] [PubMed]
- Houghton, F.D.; Hawkhead, J.A.; Humpherson, P.G.; Hogg, J.E.; Balen, A.H.; Rutherford, A.J.; Leese, H.J. Non-Invasive Amino Acid Turnover Predicts Human Embryo Developmental Capacity; 2002; Vol. 17. [CrossRef]
- de Lima, C.B.; dos Santos, É.C.; Ispada, J.; Fontes, P.K.; Nogueira, M.F.G.; dos Santos, C.M.D.; Milazzotto, M.P. The Dynamics between in Vitro Culture and Metabolism: Embryonic Adaptation to Environmental Changes. Sci Rep 2020, 10. [Google Scholar] [CrossRef]
- Gimeno, I.; García-Manrique, P.; Carrocera, S.; López-Hidalgo, C.; Valledor, L.; Martín-González, D.; Gómez, E. The Metabolic Signature of in Vitro Produced Bovine Embryos Helps Predict Pregnancy and Birth after Embryo Transfer. Metabolites 2021, 11. [Google Scholar] [CrossRef]
- Motiei, M.; Vaculikova, K.; Cela, A.; Tvrdonova, K.; Khalili, R.; Rumpik, D.; Rumpikova, T.; Glatz, Z.; Saha, T. Non-Invasive Human Embryo Metabolic Assessment as a Developmental Criterion. J Clin Med 2020, 9, 1–13. [Google Scholar] [CrossRef]
- Leese, H.J.; Brison, D.R.; Sturmey, R.G. The Quiet Embryo Hypothesis: 20 Years On. Front Physiol 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Van Winkle, L.J. Amino Acid Transport and Metabolism Regulate Early Embryo Development: Species Differences, Clinical Significance, and Evolutionary Implications. Cells 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, D.P.; Van Winkle, L.J.; Díaz de la Garza, R.I.; Dubrovsky, J.G. Interkingdom Comparison of Threonine Metabolism for Stem Cell Maintenance in Plants and Animals. Front Cell Dev Biol 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Ye, Q.; Zeng, X.; Yang, G.; Ye, C.; Chen, M.; Yu, H.; Wang, Y.; Wang, G.; Huang, S.; et al. CBS and MAT2A Improve Methionine-Mediated DNA Synthesis through SAMTOR/MTORC1/S6K1/CAD Pathway during Embryo Implantation. Cell Prolif 2021, 54. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Kang, J.; Su, J.; Zhang, J.; Zhang, L.; Liu, X.; Zhang, J.; Wang, F.; Lu, Z.; Xing, X.; et al. Methionine Adenosyltransferase 2A Regulates Mouse Zygotic Genome Activation and Morula to Blastocyst Transition. Biol Reprod 2019, 100, 601–617. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, P.D.; Baez, G.M.; Lobos, N.E. Potential Benefits of Feeding Methionine on Reproduction Efficiency of Lactating Dairy Cows.
- Hugentobler, S.A.; Diskin, M.G.; Leese, H.J.; Humpherson, P.G.; Watson, T.; Sreenan, J.M.; Morris, D.G. Amino Acids in Oviduct and Uterine Fluid and Blood Plasma during the Estrous Cycle in the Bovine. Mol Reprod Dev 2007, 74, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Drazic, A.; Winter, J. The Physiological Role of Reversible Methionine Oxidation. Biochim Biophys Acta Proteins Proteom 2014, 1844, 1367–1382. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, Y.; Zhou, M.; Zhang, L.; Pang, Y.; Wang, C.; Xiao, Q.; Liu, L. Metabolism and Secretion Mechanism of Catecholamine Syndrome and Related Treatment Strategies. J Xiangya Med 2020, 5. [Google Scholar] [CrossRef]
- Zhuan, Q.; Ma, H.; Chen, J.; Luo, Y.; Luo, Y.; Gao, L.; Hou, Y.; Zhu, S.; Fu, X. Cytoplasm Lipids Can Be Modulated through Hormone-Sensitive Lipase and Are Related to Mitochondrial Function in Porcine IVM Oocytes. Reprod Fertil Dev 2020, 32, 667–675. [Google Scholar] [CrossRef]
- Elmetwally, M.A.; Lenis, Y.; Tang, W.; Wu, G.; Bazer, F.W. Effects of Catecholamines on Secretion of Interferon Tau and Expression of Genes for Synthesis of Polyamines and Apoptosis by Ovine Trophectoderm †. Biol Reprod 2018, 99, 611–628. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Ren, W.; Rahu, N.; Kalhoro, D.H.; Yin, Y. Exploring Polyamines: Functions in Embryo/Fetal Development. Animal Nutrition 2017, 3, 7–10. [Google Scholar] [CrossRef]
- Minois, N. Molecular Basis of the “anti-Aging” Effect of Spermidine and Other Natural Polyamines - A Mini-Review. Gerontology 2014, 60, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Muzikova, E.; Clark, D.A. Polyamines May Increase the Percentage of In-Vitro Fertilized Murine Oocytes That Develop into Blastocysts; 1995; Vol. 10. [CrossRef]
- Dunning, K.R.; Cashman, K.; Russell, D.L.; Thompson, J.G.; Norman, R.J.; Robker, R.L. Beta-Oxidation Is Essential for Mouse Oocyte Developmental Competence and Early Embryo Development. Biol Reprod 2010, 83, 909–918. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-González, D.F.; Rodríguez-Osorio, N.; Long, C.R.; Vásquez-Araque, N.A.; Maldonado-Estrada, J.G. L-Carnitine Supplementation during in Vitro Maturation and in Vitro Culture Does Not Affect the Survival Rates after Vitrification and Warming but Alters Inf-T and Ptgs2 Gene Expression. Int J Mol Sci 2020, 21, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Inaba, Y.; Somfai, T.; Kaneda, M.; Geshi, M.; Nagai, T.; Manabe, N. Supplementation of Culture Medium with Class L-Carnitine Improves Development and Cryotolerance of Bovine Embryos Produced in Vitro. Reprod Fertil Dev 2013, 25, 589–599. [Google Scholar] [CrossRef]
- Ye, J.; Li, J.; Yu, Y.; Wei, Q.; Deng, W.; Yu, L. L-Carnitine Attenuates Oxidant Injury in HK-2 Cells via ROS-Mitochondria Pathway. Regul Pept 2010, 161, 58–66. [Google Scholar] [CrossRef]
- Sudano, M.J.; Santos, V.G.; Tata, A.; Ferreira, C.R.; Paschoal, D.M.; Machado, R.; Buratini, J.; Eberlin, M.N.; Landim-Alvarenga, F.D.C. Phosphatidylcholine and Sphingomyelin Profiles Vary in Bos Taurus Indicus and Bos Taurus Taurus in Vitro- and in Vivo-Produced Blastocysts. Biol Reprod 2012, 87. [Google Scholar] [CrossRef]
- Wu, G.; Aoyama, C.; Young, S.G.; Vance, D.E. Early Embryonic Lethality Caused by Disruption of the Gene for Choline Kinase α, the First Enzyme in Phosphatidylcholine Biosynthesis. Journal of Biological Chemistry 2008, 283, 1456–1462. [Google Scholar] [CrossRef] [PubMed]
- Kühn, T.; Floegel, A.; Sookthai, D.; Johnson, T.; Rolle-Kampczyk, U.; Otto, W.; von Bergen, M.; Boeing, H.; Kaaks, R. Higher Plasma Levels of Lysophosphatidylcholine 18:0 Are Related to a Lower Risk of Common Cancers in a Prospective Metabolomics Study. BMC Med 2016, 14. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
A Venn diagram summarizing the metabolites shared or differentiating among growth media of pregnancy-yielding blastocysts (“ET-P BL”), blastocysts that did not result in a successful pregnancy (“ET-NP BL”), and empty culture media.
Figure 1.
A Venn diagram summarizing the metabolites shared or differentiating among growth media of pregnancy-yielding blastocysts (“ET-P BL”), blastocysts that did not result in a successful pregnancy (“ET-NP BL”), and empty culture media.
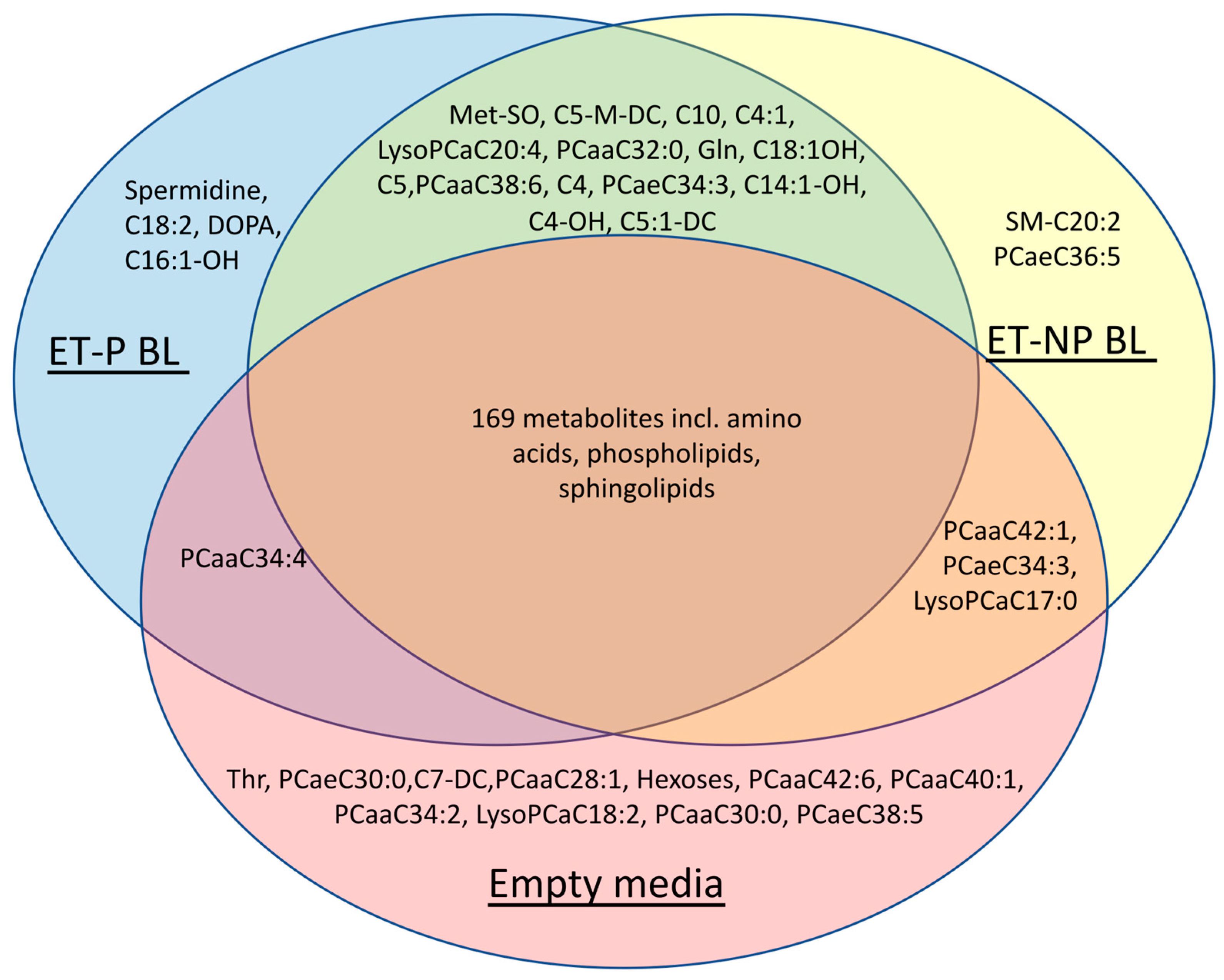

Table 1.
Metabolites exhibiting a significant difference in concentrations (indicated by a bolded p-value) or those displaying borderline significance. In instances where metabolites were detected in only a limited number of samples, the Chi-square test was employed, and the count of samples with detectable concentrations was provided. For metabolites demonstrating an approximately normal distribution, the analysis involves ANOVA with a Tukey HSD post-hoc test. Conversely, for metabolites featuring a non-normal distribution of values, the Kruskal-Wallis test, coupled with a Dunn post-hoc test, was utilized.
Table 1.
Metabolites exhibiting a significant difference in concentrations (indicated by a bolded p-value) or those displaying borderline significance. In instances where metabolites were detected in only a limited number of samples, the Chi-square test was employed, and the count of samples with detectable concentrations was provided. For metabolites demonstrating an approximately normal distribution, the analysis involves ANOVA with a Tukey HSD post-hoc test. Conversely, for metabolites featuring a non-normal distribution of values, the Kruskal-Wallis test, coupled with a Dunn post-hoc test, was utilized.
| Metabolites | Concentrations in culture media, μM | Significance | |||
|---|---|---|---|---|---|
| Metabolite | Pregnancy | No pregnancy | Control media |
p-value | Test |
| Thr | 113.5 (±12.7)b | 121.7 (±12.9)b |
205.7 (±43.5)a | 1.20E-07 | ANOVA |
| PC ae C30:0 | 0.029 (±0.003)b |
0.033 (±0.003)c |
0.041 (±0.005)a | 6.70E-06 | ANOVA |
| C7-DC | 0.013 (±0.002)b |
0.015 (±0.002)b |
0.02 (±0.003)a |
3.20E-05 | ANOVA |
| Met-SO | 1.1 (±0.26)b |
0.98 (±0.28)b |
0.33 (±0.28)a | 7.10E-05 | ANOVA |
| PC aa C28:1 | 0.008 (±0.003)b |
0.009 (±0.003)b |
0.015 (±0.001)a | 0.00019 | ANOVA |
| Spermidine | 10 (90.91 %) |
2 (20 %) |
0 (0 %) |
0.00035 | Chi-square |
| C5M-DC | 0.042 (±0.004)b |
0.04 (±0.003)b |
0.032 (±0.005)a | 0.00038 | ANOVA |
| C10 | 0.114 (±0.013)b |
0.106 (±0.006)b |
0.091 (±0.008)a | 0.00095 | ANOVA |
| DOPA | 7 (64%) |
2 (20%) |
0 (0%) |
0.021 | Chi-square |
a b c Culture media groups that do not exhibit statistical differences from each other were assigned the same letter, while groups with differing letters indicate significant differences between them.
Table 2.
Metabolite sums and ratios exhibiting significant differences (indicated by bolded p-values) or borderline significance in concentrations were examined. In cases where metabolites were detected in only a few samples, the Chi-square test was employed, providing the count of samples with detectable concentrations. For metabolites displaying a roughly normal distribution, ANOVA with a Tukey HSD post-hoc test was applied. In instances of metabolites featuring a non-normal distribution of values, the Kruskal-Wallis test with a Dunn post-hoc test was utilized.
Table 2.
Metabolite sums and ratios exhibiting significant differences (indicated by bolded p-values) or borderline significance in concentrations were examined. In cases where metabolites were detected in only a few samples, the Chi-square test was employed, providing the count of samples with detectable concentrations. For metabolites displaying a roughly normal distribution, ANOVA with a Tukey HSD post-hoc test was applied. In instances of metabolites featuring a non-normal distribution of values, the Kruskal-Wallis test with a Dunn post-hoc test was utilized.
| Metabolites | Concentrations in culture media | Significance | |||
|---|---|---|---|---|---|
| Metabolite | Pregnancy | No pregnancy | Control media |
p-value | Test |
| Met-SO/Met | 0.022 (±0.005)b | 0.02 (±0.007)b | 0.006 (±0.005)a | 1.00E-04 | ANOVA |
| SFA/PC | 0.76 (±0.017)b | 0.77 (±0.024)b | 0.80 (±0.015)a | 0.0011 | ANOVA |
| Total PC/SM | 1.77 (±0.057)b | 1.77 (±0.043)b | 1.853 (±0.035)a | 0.0083 | ANOVA |
| Total PC aa | 0.88 (0.86-0.9a)b | 0.9 (0.89-0.92)b | 0.97 (0.94-0.98)a | 0.00904 | Kruskal |
| Total PC | 1.69 (±0.063)b | 1.70 (±0.043)b | 1.78 (±0.03)a | 0.013 | ANOVA |
| C2/C0 | 0.17 (0.17-0.19)a | 0.16 (0.15-0.17)b | 0.17 (0.16-0.17)b | 0.0186 | Kruskal |
| (C2+C3)/C0 | 0.21 (0.20-0.22)b | 0.2 (0.19-0.21)a | 0.2 (0.199-0.211)a | 0.0285 | Kruskal |
| Total AC-DC / Total AC | 0.19 (±0.008)b | 0.19 (±0.005)b | 0.198 (±0.006)a | 0.043 | ANOVA |
a b c Culture media groups that do not exhibit statistical differences from each other were assigned the same letter, while groups with differing letters indicate significant differences between them.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



