
Submitted:
25 December 2023
Posted:
26 December 2023
You are already at the latest version
Abstract
Toxic runoff from heavy rains on 26 September 2019 caused a complete fish kill on the 2-km-long headwater reach of Garvin Brook, Winona County, Minnesota, USA. This project examined recovery of the slimy sculpin (Uranidea cognata) population within the lower 900 m of the kill zone, comparing relative abundance estimates and size structures between the kill zone and a downstream, unimpacted reference section. Electrofishing surveys were conducted at 24 sites (12 within both kill and reference zones) at six, 11, 18, 28, and 42 months post-kill to assess relative abundance (catch-per-effort [CPE], fish/min) and population age structures (based on total length). At six months post-kill, sculpin were present throughout the kill zone. However, adult CPE declined significantly with upstream distance, suggestive of on-going immigration of adult sculpin from downstream. Age structures were similar in kill and reference zones with all size/age groups present in both zones after six months, while CPE was twice as high in the reference (6.3 fish/min) versus the kill zone (3.0 fish/min). After 11 months, CPE did not differ between zones (14 fish/min) and remained that way for the remainder of the study. However, age structures differed dramatically between zones at both 11 and 18 months post-kill; adult sculpin were significantly more abundant in the reference zone, whereas juvenile fish dominated the kill zone. By 28 months post-kill, both sculpin abundance and age structure within the kill zone had fully recovered from the kill. Sculpin recovery was accomplished mostly through enhanced reproduction within the kill zone following initial post-kill movements of both adult and juvenile fish into the kill zone from downstream. Low predation on juvenile sculpin due to reduced abundance of trout and adult sculpin post-kill likely allowed more rapid recovery of the sculpin population within the kill zone.
Keywords:
fish kill
; recovery
; sculpin
; instream movements
; reproduction
; Uranidea cognata
1. Introduction
Fish kills caused by anthropogenic activities are commonplace in a range of aquatic habitats worldwide [1,2,3]. Kills can range from minor with little to no observable effect on aquatic ecosystems, to extreme, decimating stream and river communities over vast distances [3,4]. Concern over fish kills is amplified when they affect valuable commercial and recreational fisheries [1–3, 5].
In many fish kill events, most of the fish killed are non-game species, including cyprinids (e.g., minnows, shiners, dace, chubs), catostomids (suckers), clupeids (shad), percids (darters), cottids (sculpin), and others [3,4,6]. Although not as highly valued by the general public, these species serve important roles in community stability and energy transfer to higher trophic levels [7,8,9]. Their recovery after a fish kill is just as important for overall ecosystem function as the recovery of the top-level piscivorous game fishes [4].
Many fish species are highly mobile, and can quickly recolonize defaunated reaches after a fish kill if nearby source populations are present [4,5,10,11]. However, some species are much less mobile, greatly extending the time required for recolonization after a fish kill [12]. Many small-bodied, non-game fish in streams and rivers are considered largely non-mobile or sedentary [13,14,15], especially bottom-dwelling species such as darters and sculpins [16,17,18,19]. Individuals of these species may spend their entire lives within a short stream reach, with home ranges often spanning <30 m of stream length [19].
The slimy sculpin (Uranidea cognata) has the largest geographic distribution of any sculpin species in North America, spanning the breadth of Canada and extending into the northern United States and Alaska [20,21]. The species is restricted to coldwater streams, rivers, and lakes, often co-occurring with brook trout (Salvelinus fontinalis) and brown trout (Salmo trutta) [22,23]. In streams, it prefers clean, coarse substrates and shallow (<40 cm) water with slow (<20 cm/sec) current velocities [24]. Due to its wide geographic range, sensitivity to a variety of anthropogenic disturbances, relatively high abundance, short life span, high reproductive output, and easy to measure biological parameters, the species has been recommended for use as a sentinel species for monitoring studies throughout the northern regions of North America [19,25]. Although generally considered as non-migratory with low mobility and strong site fidelity [18], slimy sculpin can display long-range movements under certain conditions [26,27].
During September 2019, a fish kill resulting from organic-rich runoff after a rain event [28,29] occurred in a 2-km-long, spring-fed headwater reach of Garvin Brook, a coldwater trout stream in southeastern Minnesota, USA [5]. All brook trout, brown trout, and slimy sculpin were killed, an estimated 1440 fish total [29]. After the fish kill, brown trout within the kill reach were allowed to recover naturally via migration from downstream, unimpacted sections and reproduction by those in-migrants. Brown trout quickly repopulated the kill area, spawning within the entire kill zone less than two months after the kill [5]. However, it was not known if the slimy sculpin population would respond similarly post-kill, given its absence of any defined spawning migration and its presumed general lack of mobility overall [19].
The present study was initiated to examine the recovery of slimy sculpin in Garvin Brook after the September 2019 fish kill. Specifically, I examined the abilities of slimy sculpin (1) to recolonize the kill zone via upstream movements only from unimpacted downstream reaches (no unimpacted upstream source of potential colonizers), and (2) to reproduce successfully within the kill zone. These were assessed by a series of five, relative-abundance electrofishing surveys spanning three spawning seasons over a 42-month period post-kill. Surveys were conducted both within the kill zone and a similar, unimpacted reference zone downstream from the kill zone.
2. Methods
2.1. Study Site
Garvin Brook is a 1st- to 3rd-order coldwater stream in Winona County, Minnesota, USA (43.999792 N, -91.813290 W). It originates from a series of groundwater springs, flowing as a designated coldwater trout stream for 26 km before joining with the Mississippi River at Minnesota City, Minnesota. Native fish species present within the stream include brook trout, slimy sculpin, and brook stickleback (Culaea inconstans), as well as introduced brown trout. The stream is managed primarily as a coldwater fishery for brown trout by the Minnesota Department of Natural Resources [30].
A fish kill was reported on 26 September 2019 in the headwaters section of Garvin Brook. Dead fish were observed throughout the reach from the main spring source to the stream’s confluence with Peterson Creek, a stream length of 2043 m. State agency personnel investigating the kill estimated that the majority (>90%) of fish impacted had died in the lower half of the impacted reach, within a county park that borders the stream for 900 m above the Garvin Brook–Peterson Creek confluence [29,31]. No fish were believed to have survived the kill within the entire 2-km+ reach upstream from the Peterson Creek confluence [28]. The fish community below the confluence was unimpacted, likely due to the dilution effect of the flow from Petersen Creek [29]. No specific cause of the fish kill was determined, but post-kill investigations suggested that organic runoff after a rain event had produced short-term toxic conditions [29,31].
2.2. Fieldwork
Slimy sculpin were surveyed in Garvin Brook during five time periods post-kill: March 2020, August 2020, March 2021, January 2022, and March 2023. These times represented six, 11, 18, 28, and 42 months after the fish kill. March and January were selected to permit identification of mature male and female sculpin prior to spring spawning, whereas August was chosen to determine relative abundance of young-of-year (YOY) sculpin after the first spawning period post-kill.
Each survey consisted of brief electrofishing passes at each of 12 stream sites (except only 10 sites were surveyed in the reference reach during March 2020) spaced evenly throughout two, 900-m sections of Garvin Brook: one 900-m section within the kill zone (upstream from Peterson Creek) and another section downstream (below Peterson Creek) in a reference zone not impacted by the fish kill (Figure 1). At each of the 24 sites, a short section (approximately 10–20 m) of stream that included the preferred habitats of both juvenile and adult sculpin [24] was electrofished (Smith-Root model LR-24 or model LR-20B backpack electrofishers, along with a single dip netter) for a time period (typically 1 to 3 minutes) sufficient to capture approximately 10 or more sculpin. Electrofisher “on” time was recorded for each site, and all sculpin were measured (nearest mm total length [TL]), recorded as male, female, or immature (based on external sex characteristics just prior to spawning; presence of urogenital papilla in adult males, abdomen swollen with eggs and no urogenital papilla in adult females) [32], and returned alive to the stream. Sex was not determined for adult sculpin during the August 2020 survey.
2.3. Data Analyses
Sculpin populations were compared between kill and reference zones using a variety of approaches. Size structures of sculpin were compared graphically between kill and reference zones using simple length-frequency plots, and average sculpin sizes were compared with Student t-tests. In addition, sizes of male and female sculpin were compared within each zone–date combination to examine possible differences between the sexes. Chi-square goodness-of-fit tests were used to assess male-to-female sex ratios in each zone on each of the five survey dates, and sex ratios were compared between zones on each survey data with contingency table Chi-square tests.
At each site and date surveyed, sculpin catch-per-effort (CPE) was calculated as fish/minute for juvenile, adult, and total sculpin. CPE values were compared between kill and reference zones for each date with Student t-tests (with adjusted df values if variances were significantly different between zones, based on results of an F test; www.VassarStats.net). Possible patterns in CPE (total, adult, juvenile) among sample sites (downstream to upstream) within each zone during the first survey (March 2020) were examined with simple linear regression to assess possible migration of sculpin into the kill zone from downstream locations. Decreasing CPE moving upstream within the kill zone would suggest ongoing immigration of sculpin into the zone from downstream.
3. Results
3.1. Survey Effort
In total, 1834 slimy sculpin were collected and examined across the five survey dates: 963 sculpin in the kill zone and 871 in the reference zone. Across all dates, there was no significant difference between zones in either the electrofishing time per site (t118 = 1.74, P = 0.084) or the number of sculpin collected per site (t118 = 1.00, P = 0.320), averaging 103 sec (SD = 45 sec; range = 35–310 sec) and 16 sculpin (SD = 6 fish; range = 3–32 fish) per site overall.
3.2. Sculpin Age Structure
Length-frequency distributions of slimy sculpin displayed a variety of patterns across the five survey periods (Figure 2). Sculpin ranged in size from 24 mm up to 119 mm TL. During the March 2020 survey (6 months after the fish kill), all age groups of sculpin were present throughout the entirety of the kill zone, with yearling fish dominating the population. The same age groups were present in the reference zone, although yearling fish were relatively less dominant than in the kill zone. By August 2020, YOY representing the 2020 year class were very dominant at kill zone sites, and dominant at reference zone sites. Older, larger sculpin were relatively more abundant within the reference zone than in the kill zone. By March 2021, the 2020 year class (now yearling fish) dominated both zones, with older fish again relatively more common in the reference zone. In January 2022, the 2021 sculpin year class (now yearlings) were dominant in the reference zone, but were less common in the kill zone, clearly outnumbered by 2-year-old fish representing the 2020 year class. Finally, in March 2023, yearling fish (representing the 2022 year class) were dominant within the kill zone, whereas that age class was much reduced in the reference zone, which continued to be dominated by the 2021 year class (now 2-year-old fish). Across all time periods, the full size range of sculpin were present within both the kill and reference zones (Figure 2).
Average sizes of sculpins within the two zones varied across time periods based on changing dominance of various year classes (Table 1). During the initial survey, average sculpin size was nearly identical between kill and reference zones. Sculpin were significantly larger in the reference versus the kill zone in August 2020 and in March 2021, but significantly larger in the kill zone than in the reference zone in both January 2022 and March 2023 (Table 1).
3.3. Adult Sculpin Sex Ratios and Size Differences
During winter surveys when sex of adult sculpin could be determined, male fish generally averaged 10 mm longer than female fish (Figure 3). Male fish were significantly larger than females during all surveys in the reference zone, but only during two of the surveys (March 2020, January 2022) in the kill zone.
Female sculpin were more abundant than male fish in five of the eight zone X date combinations examined (Figure 4). However, only three of those five had sex ratios that differed significantly from 1:1. When sex ratios were compared between kill and reference zones, they differed only during March 2023 (Table 2). Overall, there is no evidence of any pattern in sculpin sex ratios throughout the study period.
3.4. Sculpin CPE Relative Abundances
Sculpin CPE displayed some distinctive patterns across the five survey time periods (Figure 5). During the first survey 6 months post-kill, total sculpin relative abundance was significantly higher (Table 3) in the reference zone than in the kill zone (Figure 5A), largely due to differing abundances of adult fish in the two zones (Figure 5B). Higher relative abundances of adult fish in the reference zone were maintained across the next two survey periods (Figure 5A, Table 3). However, beginning in August 2020 and continuing through March 2021, relative abundance of juvenile sculpin was significantly higher (1.5 to 2.7 times) in the kill zone than in the reference zone (Figure 5C, Table 3). Consequently, total sculpin CPE in the kill zone was equal to or slightly higher than that in the reference zone by August 2020 (Figure 5A), 11 months after the fish kill. Total sculpin CPE did not differ between the two zones for the remainder of the study period, even though adult and juvenile CPE continued to display some significant differences (Figure 5B, C, Table 3).
Slimy sculpin CPE displayed various patterns among sample sites during the initial survey in March 2020. Within the kill zone, both total and adult sculpin CPE declined significantly from downstream to upstream, whereas juvenile CPE did not (Table 4). Within the reference zone, total CPE increased significantly from downstream to upstream, but neither adult nor juvenile CPE changed significantly (Table 4).
4. Discussion
After slimy sculpin were eliminated from a 2-km headwater reach of Garvin Brook by a fish kill in September 2019, recovery was driven by upstream movement of fish of all ages into the defaunated reach within six months after the kill, reproduction by the adult segment of the in-migrant group, and good survival of the 2020 and 2021 year classes of sculpin. Abundance recovery relative to a reference reach was achieved within six months of the kill, but age structure recovery was not accomplished until 28 months post-kill.
Instream movements by slimy sculpin usually have been characterized as minimal, with individual fish residing within a short section (<30 m) of stream for much of their lifetimes [18,19,33]. Adults (especially males) move very little due to being territorial [18,34,35], so instream movements usually are made by subadults seeking suitable, unoccupied habitat [26]. However, even those movements may be very limited if habitat is plentiful and food resources are abundant [36,37].
In the present study, all age groups of slimy sculpin were present throughout the lower 900 m of the kill zone by the first survey date, six months post-kill. However, decreasing adult sculpin CPE with increasing distance upstream (60% decline in CPE between lowermost and uppermost sites) within the kill zone suggests that adult sculpin may still have been actively moving into that section from downstream six months after the kill. Fewer than half as many adult fish had reached the upstream site compared to the downstream site. In fact, adult sculpin continued moving into the kill zone after March 2020, with adult CPE in the kill zone nearly doubling between March and August 2020, then remaining constant through March 2021. There was no obvious dominance of one sex over the other in the adult sculpin that moved into the kill zone. In contrast, juvenile sculpin CPE did not differ throughout the kill zone, indicating that young sculpin may have been more migratory than older fish, achieving relatively similar abundances throughout the 900 m of the kill zone. These observations appear to contradict previous studies that reported very limited movements of sculpin within streams [18,19,33]. However, those studies were conducted in streams with intact sculpin populations, not a system that had experienced a fish kill. Many small-bodied fishes that normally display limited mobility can quickly repopulate kill zones [38,39], so it should not be surprising that slimy sculpin can quickly repopulate stream habitats lacking any sculpin. In fact, even in studies where sculpin have been found to move very little, some individuals exhibited movements exceeding several hundred meters during the study periods [18,19,33], suggesting that sculpin are capable of extensive movements even within more crowded habitats. Reduced movements of adult versus juvenile sculpin lend support to previous studies [26,27] indicating greater movement by sculpin during their first year or two followed by greatly reduced movements as adults.
There is additional support for the observation that slimy sculpin can quickly populate a stream reach where they previously were extirpated. Beginning in 2003, the Minnesota Department of Natural Resources initiated a slimy sculpin reintroduction program to stock sculpin into nine coldwater trout streams where they were lacking (likely eliminated many decades prior due to historic poor water quality), to restore an ecologically important native species and provide additional forage for existing wild trout populations [40]. Sculpin dispersed rapidly both upstream and downstream from their reintroduction sites, and several studies subsequently examined various aspects of the reintroductions [41,42,43]. Sculpin recently were reintroduced to an additional eight streams in 2022/2023 [44]. Coincidentally, Garvin Brook (specifically the reference reach) served as one of three donor streams from which sculpin were collected to stock elsewhere [40].
Alternatively, it is possible that slimy sculpin may not have needed to move into the kill zone to repopulate it. Instead, the September 2019 fish kill may not have been a complete kill, leaving some survivors to gradually repopulate the kill zone via subsequent reproduction. The fish kill investigation estimated that only 172 sculpin died throughout the 2-km-long kill zone [28], an estimated density of <0.02 sculpin/m2. Based on previous work in Garvin Brook that found sculpin present at densities >2 fish/m2 (N. Mundahl, unpublished data), a complete fish kill may have killed over 18,000 sculpin in the identified kill zone, 100 times higher than the official count. The sculpin’s small size, cryptic coloration, and preferred rocky habitats [19,24] likely caused most of the dead fish to be missed during the kill count. But those rocky interstices also may have provided reduced- or no-flow microhabitats that protected some sculpin from short-term toxic stream flows moving past overhead during the September 2019 kill event [45], allowing fish to survive an otherwise catastrophic event. However, lacking any electrofishing survey data immediately after the kill event to support these speculations, it cannot be stated with any degree of certainty that some sculpin may actually have survived the kill.
Although adult slimy sculpin CPE in the kill zone was less than 40% of that observed in the reference zone in March 2020 shortly before the onset of sculpin spring spawning, production of juvenile sculpin by those adult fish was high. During August surveys, juvenile sculpin CPE in the kill zone was two to three times greater than in the reference zone. Some of this difference may have resulted from a greater proportion of large (>100 mm TL) adult sculpin, with greater potential fecundity [46], in the kill zone compared to the reference zone. But greater fecundity in the kill zone could have been counteracted by the significantly skewed sex ratio of adult sculpin (more females than males) within the reference zone. However, much of the higher juvenile CPE in the kill zone simply may have resulted from better survival of young sculpin due to reduced mortality from fewer potential predators (i.e., adult sculpin, brown trout) within that zone [5,46]. In fact, reduced numbers of potential predators of young sculpin (i.e., adult sculpin, adult brown trout) continued through the March 2021 survey period [5]. Much of the recovery of the slimy sculpin population within the kill zone of Garvin Brook was directly the result of reproduction by the adult sculpin that migrated into the kill zone, and the subsequent growth and maturation of the 2020 and 2021 year-classes of sculpin.
5. Conclusions
After an autumn fish kill event, it required approximately two years for the slimy sculpin population to fully recover within the headwaters of Garvin Brook. Although recovery of total sculpin CPE was evident within the kill zone in surveys conducted 11 months post-kill, sculpin age structures remained different between the kill and reference zones through 18 months post-kill. Age structures finally looked similar between the two zones in surveys conducted 28 months after the kill, as the large, 2020 sculpin year class within the kill zone reached maturity. Coincidentally, this was the same pattern and same duration of population recovery exhibited by brown trout that recolonized the kill zone after the fish kill [5]. Considering the differences in fish size, expected mobilities, and season of spawning (spring for sculpin, fall for trout) of these two species, the similarity in both abundance and age structure recovery was unexpected. But with similar life spans in this headwaters system, population recovery based on strong reproductive output, good survival of young, and a short time (approximately two years) to sexual maturity, is likely highly adaptive.
Funding
This project received in-kind support from the Department of Biology and Large River Studies Center, Winona State University.
Data Availability Statement
Data are available directly from the author per reasonable request.
Acknowledgments
I thank V. Snook and T. Burri (Minnesota Department of Natural Resources–Fisheries) for sharing data and other details from the 2019 fish kill investigation. V. Snook and B. Nerbonne (Minnesota Department of Natural Resources–Fisheries) provided information about the slimy sculpin reintroduction program.
Conflicts of Interest
The author declares no competing interests.
Ethics approval – Compliance with ethical standards
Fish collecting permits were provided by the Minnesota Department of Natural Resources–Fisheries, and collections were made with the approval of the Winona State University Institutional Animal Care and Use Committee (1317064-1, 1310072-2).
References
- Thronson, A.; Quigg, A. Fifty-five years of fish kills in coastal Texas. Estuaries Coasts 2008, 31, 802–813. [CrossRef]
- La, V.T.; Cooke, S.J. Advancing the science and practice of fish kill investigations. Rev. Fish. Sci. 2011, 19, 21–33. [CrossRef]
- Phelps, N.B.D.; Bueno, I.; Poo-Muñoz, D.A., Knowles, S.J.; Massarani, S.; Rettkowski, R.; Shen, L.; Rantala, H.; Phelps, P.L.F.; Escobar, L.E. Retrospective and predictive investigation of fish kill events. J. Aquat. Anim. Health 2019, 31, 61–70. [CrossRef]
- Meade, R. Fish and invertebrate recolonization in a Missouri prairie stream after an acute pollution event. N. Am. J. Fish. Manage. 2004, 24, 7–19. [CrossRef]
- Schnaser, A.C.; Mundahl, N.D. Recovery of a headwater stream population of brown trout after a fish kill in southeastern Minnesota, USA. Environ. Biol. Fish. 2022, 105, 1179–1192. [CrossRef]
- Hunt, L. South Branch Whitewater River unified fish kill response. Minnesota Department of Agriculture, Saint Paul, Minnesota, USA. 2015. https://files.dnr.state.mn.us/areas/fisheries/lanesboro/unified-fish-kill-response12-22-15.pdf Accessed 21 December 2021.
- Roell, M.J.; Orth, D.J. The roles of predation, competition, and exploitation in the trophic dynamics of a warmwater stream: a model synthesis, analysis, and application. Hydrobiologia 1994, 291, 157–178. [CrossRef]
- Fitzgerald, D.G.; Kott, E.; Lanno, R.P.; Dixon, D.G. A quarter century of change in the fish communities of three small streams modified by anthropogenic activities. J. Aquat. Ecosyst. Stress Recovery 1997, 6, 111–127. [CrossRef]
- Cooke, S.J.; Bunt, C.M.; Hamilton, S.J.; Jennings, C.A.; Pearson, M.P.; Cooperman, M.S.; Markle, D.F. Threats, conservation strategies, and prognosis for suckers (Catostomidae) in North America: insights from regional case studies of a diverse family of non-game fishes. Biol. Conserv. 2005, 121, 317–331. [CrossRef]
- Peterson, J.T.; Bayley, A.B. Colonization rates of fishes in experimentally defaunated warmwater streams. Trans. Am. Fish. Soc. 1993, 122, 199–207.
- Winona Daily News. DNR: Whitewater trout stream that saw fish kill is recovering. Winona Daily News, Winona, Minnesota, USA. 2016. https://www.winonadailynews.com/news/local/dnr-whitewater-trout-stream-that-saw-fish-kill-is-recovering/article_3bf19ec9-ec08-58d2-80f0-2bbd6be1a15c.html Accessed: 21 December 2021.
- Freeman, M.; Elkins, D.; Maholland, P.; Butler, Z.; Kleinhans, M.; Skaggs, J.; Stowe, E.; Straight, C.; Wegner, S. Slow recovery of headwater-stream fishes following a catastrophic poisoning event. J. Fish. Wildl. Manage. 2021, 12, 362–372. https://doi.org/10.3996/JFWM-20-080.
- Mundahl, N.D.; Ingersoll, C.G. 1989 Home range, movements, and density of the central stoneroller, Campostoma anomalum, in a small Ohio stream. Environ. Biol. Fish. 1989, 24, 307–311. [CrossRef]
- Rasmussen, J.E.; Belk, M.C. Individual movement of stream fishes: linking ecological drivers with evolutionary processes. Rev. Fish. Sci. Aquac. 2017, 25, 70–83. [CrossRef]
- De Fries, L.; Camana, M.; Hartz, S.M.; Becker, F.G. Heterogenous movement by a small non-migratory stream fish. Environ. Biol. Fish. 2022, 105, 1873–1885. [CrossRef]
- Mundahl, N.D.; Ingersoll, C.G. Early autumn movements and densities of johnny (Etheostoma nigrum) and fantail (E. flabellare) daters in a southwestern Ohio stream. Ohio J. Sci. 1983, 83, 103–108.
- Ingersoll, C.G.; Hlohowskyj, I.; Mundahl, N.D. Movements and densities of the darters Etheostoma flabellare, E. spectabile, and E. nigrum during spring spawning. J. Freshw. Ecol. 1984, 2, 345–351.
- Keeler, R.A.; Breton, A.R.; Peterson, D.P.; Cunjak, R.A. Apparent survival and detection estimates for PIT-tagged slimy sculpin in five small New Brunswick streams. Trans. Am. Fish. Soc. 2007, 131, 281–292. [CrossRef]
- Gray, M.A.; Curry, R.A.; Arciszewski, T.J.; Munkittrick, K.R.; Brasfield, S.M. The biology and ecology of slimy sculpin: a recipe for effective environmental monitoring. FACETS 2018, 3, 103–127. [CrossRef]
- Wallace, R.L.; McAllister, D.E.; and Rankin, M. Cottus cognatus Richardson, slimy sculpin, p. 808. In Lee, D.S.; Gilbert, C.R.; Hocutt, C.H.; Jenkins, R.E.; McAllister, D.E.; Stauffer Jr., J.R. Atlas of North American freshwater fishes. North Carolina Biological Survey Publication #1980-12. 1980.
- Page, L.M.; Burr, B.M. A field guide to freshwater fishes: North America north of Mexico. Houghton Mifflin Company, Boston, Massachusetts, USA. 1991. [CrossRef]
- Eddy, S.; Underhill, J.C. Northern fishes, with special reference to the Upper Mississippi Valley. University of Minnesota Press, Minneapolis, 1974.
- Kelly, B.B.; Siepker, M.J.; Weber, M.J. 2023. Effects of non-native Salmo trutta and multiscale habitat factors on native fishes in the Driftless Area. Can. J. Fish. Aquat. Sci. 2023, 80, 942–957. [CrossRef]
- Mundahl, N.D.; Thomas, K.N.; Mundahl, E.D. Selected habitats of slimy sculpin in coldwater tributaries of the Upper Mississippi River in Minnesota. Am. Midl. Nat. 2012, 168, 144–161. [CrossRef]
- Gray, M.A.; Munkittrick, K.R. An effects-based assessment of slimy sculpin (Cottus cognatus) populations in agricultural regions of northwestern New Brunswick. Water Qual. Res. J. Canada 2005, 40, 16–27. [CrossRef]
- Clarke, A.D.; Telmer, K.H.; Shrimpton, J.M. Movement patterns of fish revealed by otolith microchemistry: a comparison of putative migratory and resident species. Environ. Biol. Fish. 2015, 98, 1583–1597. [CrossRef]
- Weinstein, S.Y.; Coombs, J.A.; Nislow, K.H.; Riley, C.; Roy, A.H.; Whiteley, A.R. Evaluating the effects of barriers on slimy sculpin movement and population connectivity using novel sibship-based and traditional genetic metrics. Trans. Am. Fish. Soc. 2019, 148, 1117–1131. [CrossRef]
- Snook, V. Fish kill incident report: Garvin Brook at Farmer’s Community Park. Minnesota Department of Natural Resources-Fisheries, Lanesboro, Minnesota, USA. 2019.
- Burri. T.; Fier-Tucker, D.; Ganske, L.; Hunt, L.; Wieland, R. Response to 2019 fish kill on Garvin Brook, Winona County. Minnesota Pollution Control Agency, Saint Paul, Minnesota, USA. 2020 https://www.pca.state.mn.us/sites/default/files/tdr-gl-22.pdf.
- Minnesota Department of Natural Resources. Garvin Brook aquatic management area. 2023. https://www.dnr.state.mn.us/amas/detail_report.html?id=AMA00210 (accessed 10 December 2023).
- Minnesota Pollution Control Agency. Agencies conclude investigation: contaminated runoff suspected in Garvin Brook fish kill. Minnesota Pollution Control Agency, Saint Paul, Minnesota, USA. 2020. https://pca.state.mn.us/news/agencies-conclude-investigation-contaminated-runoff-suspected-garvin-brook-fish-kill (Accessed 13 January 2022).
- Arciszewski, T.J.; Gray, M.; Munkittrick, K.; Baron, C. Guidance for the collection and sampling of slimy sculpin (Cottus cognatus) in northern Canadian lakes for environmental effects monitoring (EEM). Canadian Technical Report of Fisheries and Aquatic Sciences 2909. Fisheries and Oceans Canada, Winnipeg, Manitoba, Canada. 2010.
- Breen, M.J.; Ruetz III, C.R.; Thompson, K.J.; Kohler, S.L. Movements of mottled sculpins (Cottus bairdii) in a Michigan stream: how restricted are they? Can. J. Fish. Aquat. Sci. 2009, 66, 31–41.
- Mousseau, T.A.; Collins, N.C.; Cabana, G. A comparative study of sexual selection and reproductive investment in the slimy sculpin, Cottus cognatus. Oikos 1987, 51, 156–162. [CrossRef]
- Majeski, M.J.; Cochran, P.A. Spawning season and habitat use of slimy sculpin (Cottus cognatus) in southeastern Minnesota. J. Freshw. Ecol. 2009, 24, 301–307. [CrossRef]
- Petty, T.D.; Grossman, G.D. Restricted movement by mottled sculpin (Pisces: Cottidae) in a southern Appalachian stream. Freshw. Biol. 2004, 49, 631–645.
- Schmetterling, D.A.; Adams, S.B. Summer movements within the fish community of a small montane stream. N. Am. J. Fish. Manag. 2004, 24, 1163–1172.
- Larimore, R.W.; Childers, W.F.; Heckrotte, C. Destruction and re-establishment of stream fish and invertebrates affected by drought. Trans. Am. Fish. Soc. 1959, 88, 261–285. [CrossRef]
- Harrel, R.C.; Davis, B.J.; Dorris, T.C. Stream order and species diversity of fishes in an intermittent Oklahoma stream. Am. Midl. Nat. 1967, 78, 428–436.
- Minnesota Department of Natural Resources. Southeast Minnesota Sculpin Reintroduction Plan. Minnesota Department of Natural Resources–Fisheries, Lanesboro, Minnesota, USA. 2003. https://files.dnr.state.mn.us/areas/fisheries/lakecity/semn_sculpin_reintroplan.pdf.
- Huff, D.D.; Miller, L.M.; Vondracek, B. Patterns of ancestry and genetic diversity in reintroduced populations of the slimy sculpin: implications for conservation. Conserv. Genet. 2010, 11, 2379–2391.
- Huff, D.D.; Miller, L.M.; Chizinski, C.J.; Vondracek, B. 2011. Mixed-source reintroductions lead to outbreeding depression in second-generation descendants of a native North American fish. Mol. Ecol. 2011, 20, 4246–4258. [CrossRef]
- Mundahl, N.D.; Mundahl, D.E.; Merten, E.C. Success of slimy sculpin introductions: influence of feeding and diets. Am. Midl. Nat. 2012, 168, 162-183.
- Nerbonne, B. Personal communication. Minnesota Department of Natural Resources–Fisheries, Saint Paul, Minnesota, USA. 2023.
- Detenbeck, N.; DeVore, P.W.; Niemi, G.J.; Lima, A. Recovery of temperate-stream fish communities from disturbance: a review of case studies and synthesis of theory. Environ. Manag. 1992, 16, 33–53. [CrossRef]
- Anderson, C.S. The structure of sculpin populations along a stream size gradient. Environ. Biol. Fish. 1985, 13, 93-102. [CrossRef]
Figure 1.
Map of the Garvin Brook study area in southeastern Minnesota, USA, indicating 900-m stream sections assessed within the kill zone (upstream from the confluence with Peterson Creek) and an unimpacted reference zone (downstream from the confluence). Yellow stars indicate survey sites where slimy sculpin were collected, and the red star on the inset map indicates the location of the study area in North America.
Figure 1.
Map of the Garvin Brook study area in southeastern Minnesota, USA, indicating 900-m stream sections assessed within the kill zone (upstream from the confluence with Peterson Creek) and an unimpacted reference zone (downstream from the confluence). Yellow stars indicate survey sites where slimy sculpin were collected, and the red star on the inset map indicates the location of the study area in North America.
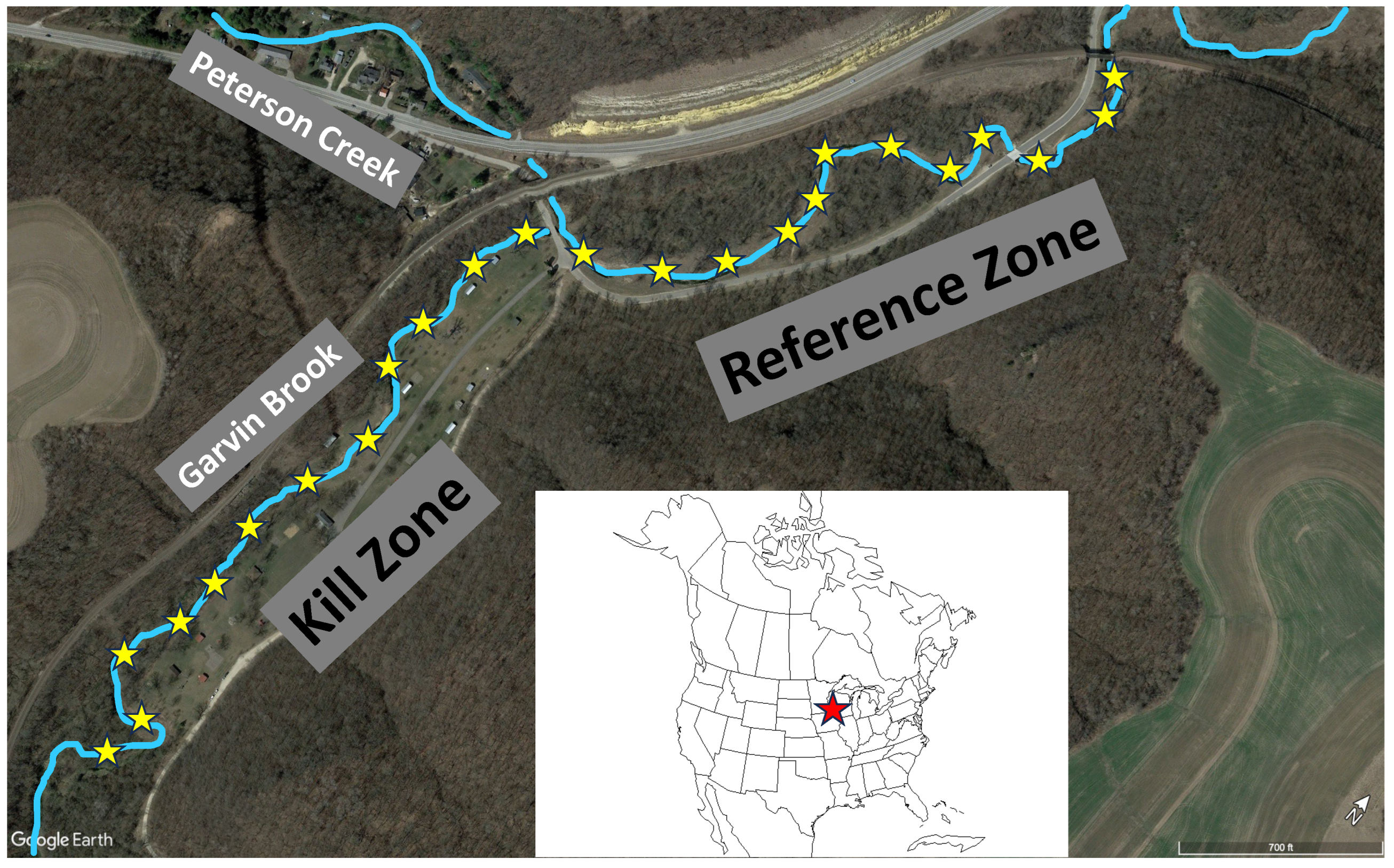
Figure 2.
Length-frequency distributions for slimy sculpin collected from kill and reference zones of Garvin Brook on five dates post-kill.
Figure 2.
Length-frequency distributions for slimy sculpin collected from kill and reference zones of Garvin Brook on five dates post-kill.
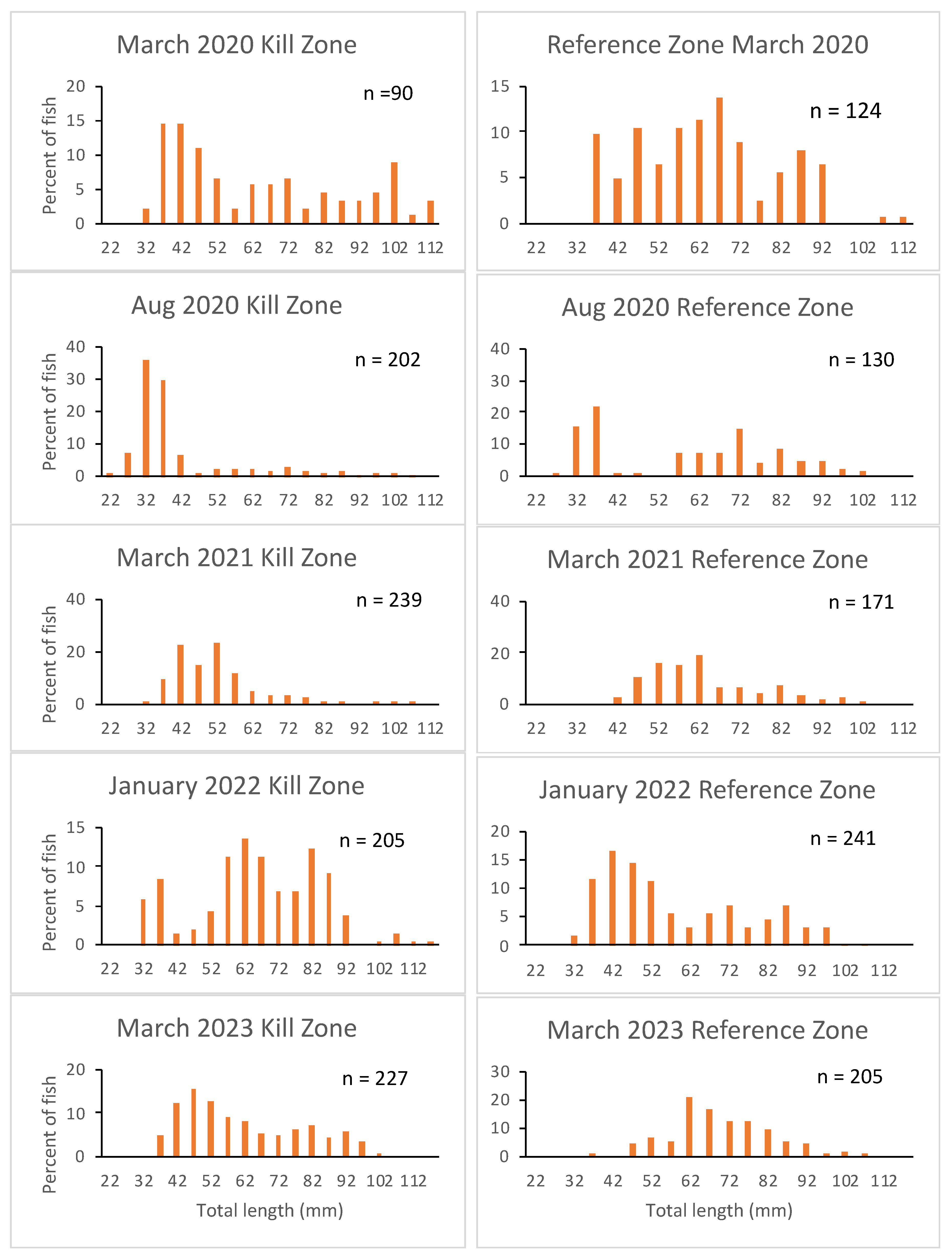
Figure 3.
Total lengths of adult female and male slimy sculpin collected from kill and reference zones of Garvin Brook prior to spring spawning during four years post-kill. Bars are means and lines indicate standard deviations. Numbers above bars are P values from t tests comparing male and female lengths within each zone on each date.
Figure 3.
Total lengths of adult female and male slimy sculpin collected from kill and reference zones of Garvin Brook prior to spring spawning during four years post-kill. Bars are means and lines indicate standard deviations. Numbers above bars are P values from t tests comparing male and female lengths within each zone on each date.
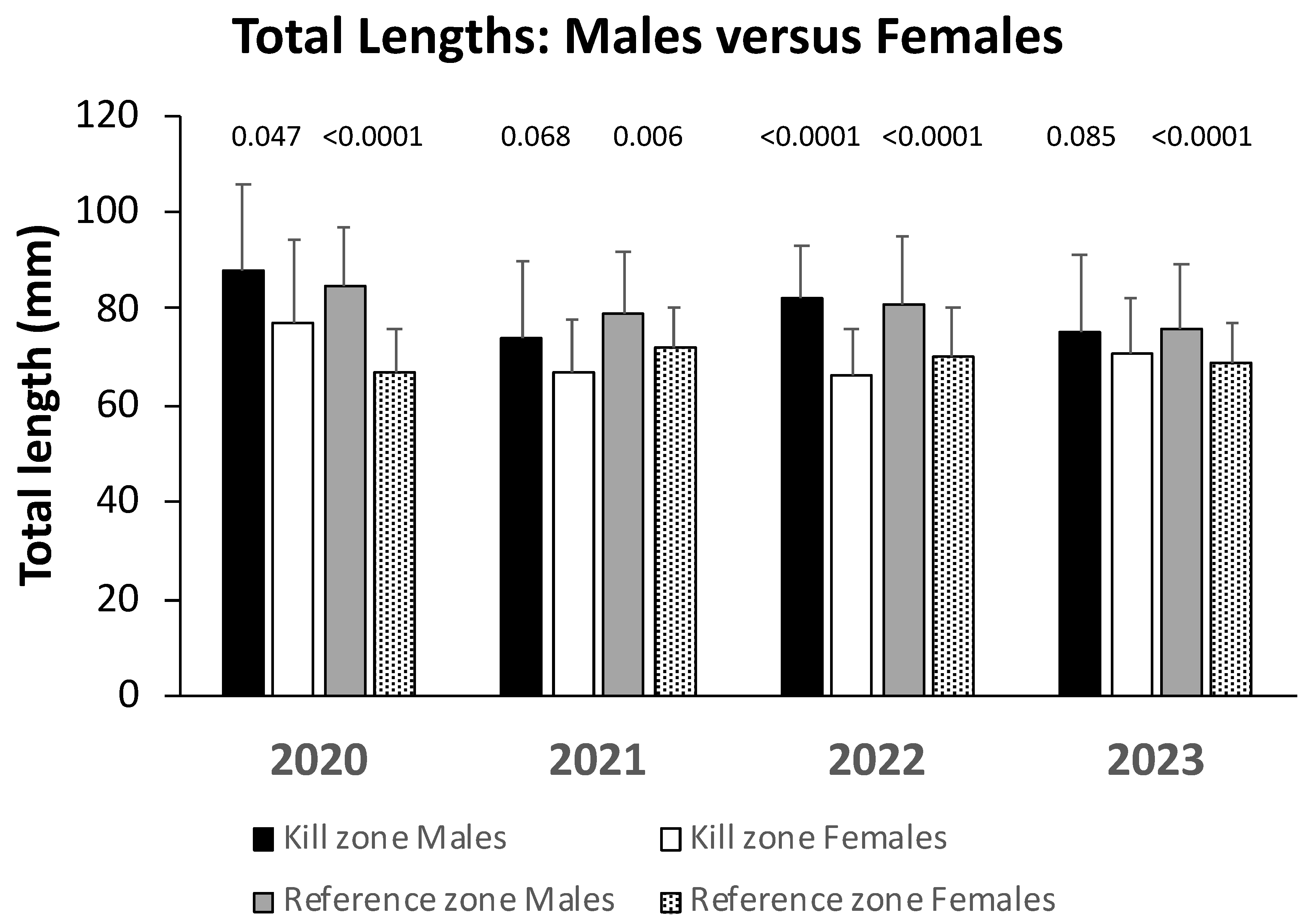
Figure 4.
Female-to-male ratios of adult slimy sculpin collected from kill and reference zones of Garvin Brook prior to spring spawning during four years post-kill. Stars above bars indicate zone-date combinations where ratios differed significantly from 1:1 (indicated by the horizontal dashed line).
Figure 4.
Female-to-male ratios of adult slimy sculpin collected from kill and reference zones of Garvin Brook prior to spring spawning during four years post-kill. Stars above bars indicate zone-date combinations where ratios differed significantly from 1:1 (indicated by the horizontal dashed line).
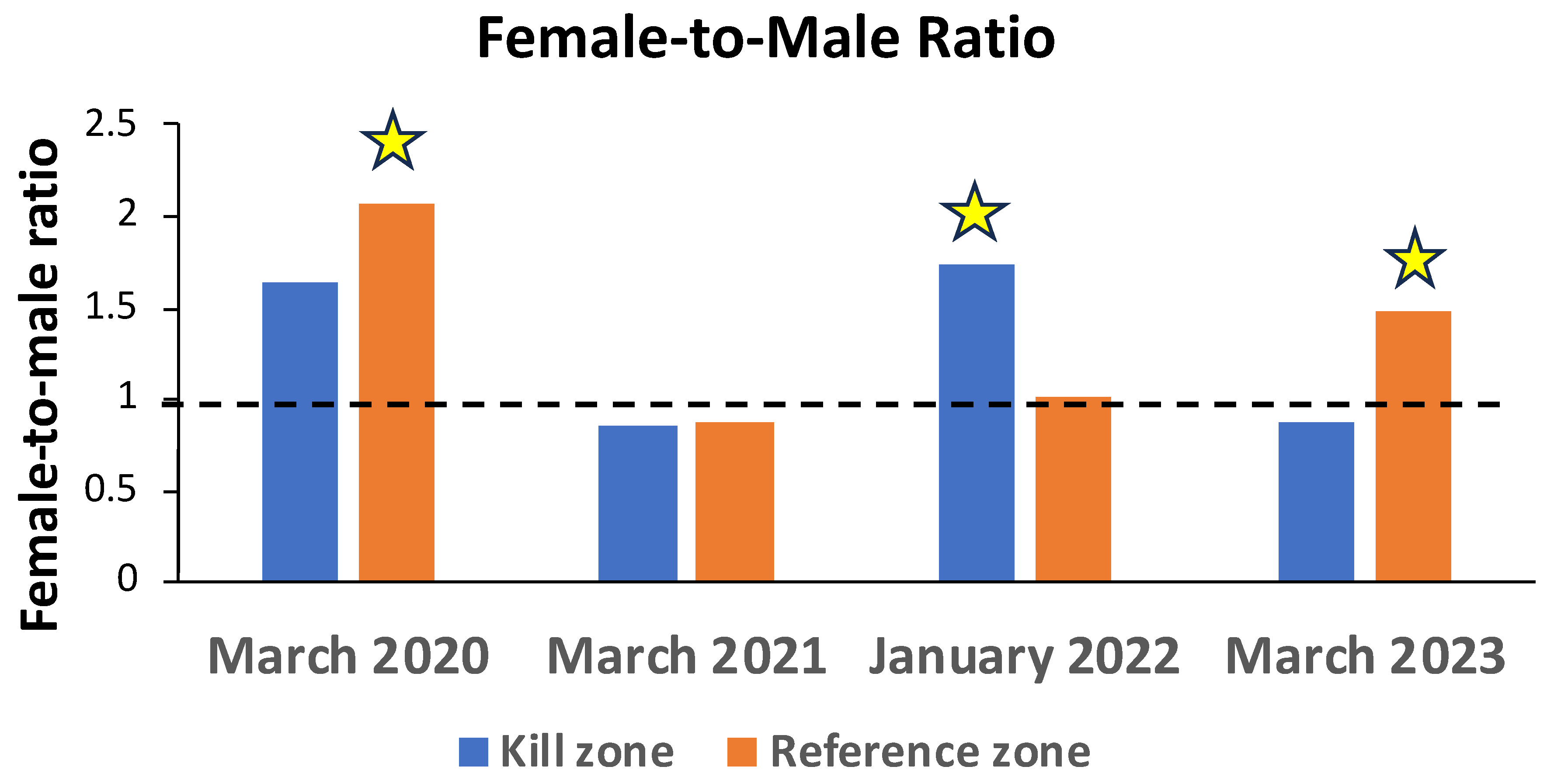
Figure 5.
Electrofishing catch per effort (fish/minute; CPE) for total (A), adult (B), and juvenile (C) slimy sculpins collected from kill and reference zones of Garvin Brook on five dates post-kill. Dots represent means and vertical bars indicate ± SD. Stars indicate dates during which mean CPE differed significantly (P < 0.05; see Table 2) between kill and reference zones, based on t test comparisons.
Figure 5.
Electrofishing catch per effort (fish/minute; CPE) for total (A), adult (B), and juvenile (C) slimy sculpins collected from kill and reference zones of Garvin Brook on five dates post-kill. Dots represent means and vertical bars indicate ± SD. Stars indicate dates during which mean CPE differed significantly (P < 0.05; see Table 2) between kill and reference zones, based on t test comparisons.
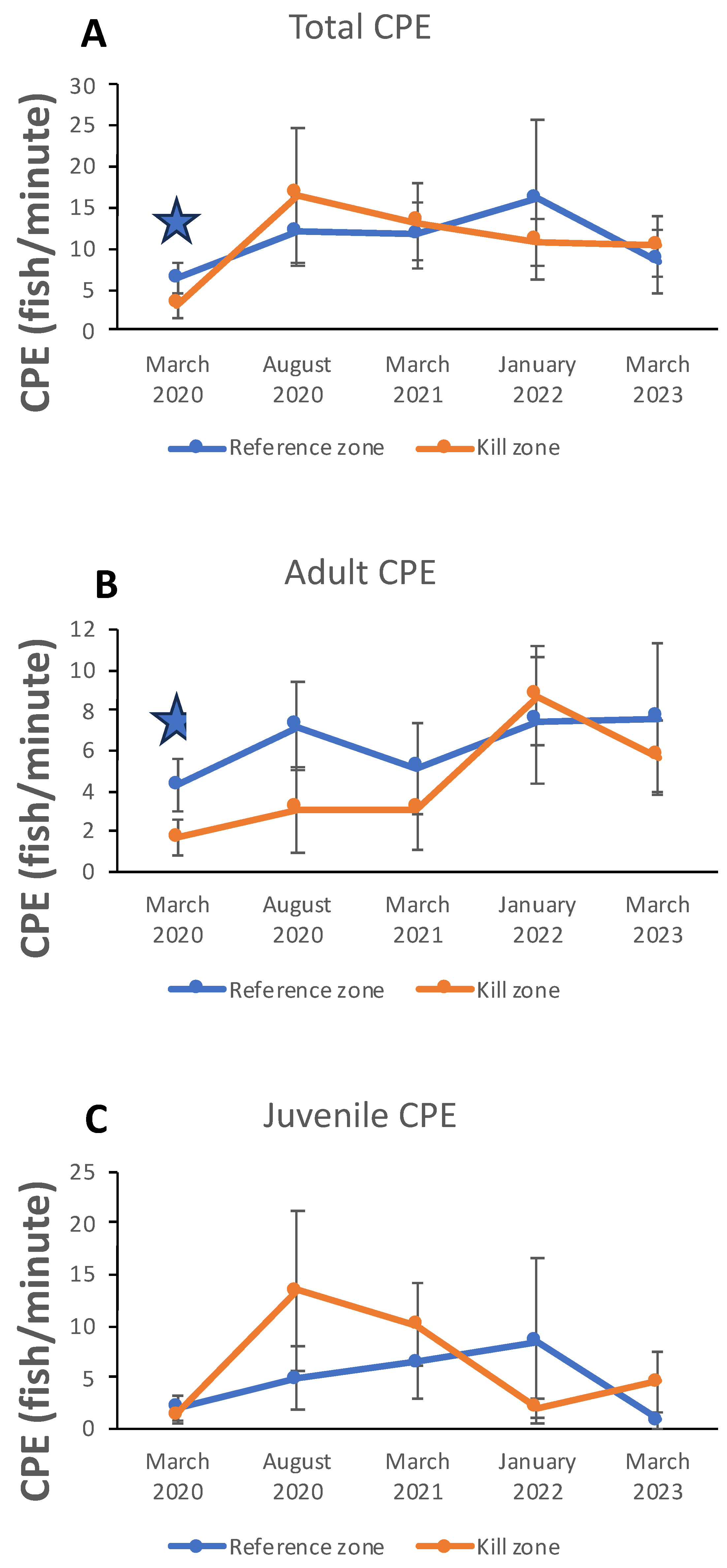

Table 1.
Average total lengths (mm TL) of slimy sculpin collected from kill and reference zones in Garvin Brook on five dates post-kill. Values are means with SD in parentheses. N = sample size. Results of t tests comparing sculpin size between zones are included.
Table 1.
Average total lengths (mm TL) of slimy sculpin collected from kill and reference zones in Garvin Brook on five dates post-kill. Values are means with SD in parentheses. N = sample size. Results of t tests comparing sculpin size between zones are included.
| Date | Kill zone | N | Reference | N | t value | P | |||
|---|---|---|---|---|---|---|---|---|---|
| Mar-20 | 64 (24) | 90 | 64 (17) | 124 | 0.05 | 0.959 | |||
| Aug-20 | 41 (17) | 202 | 59 (22) | 130 | 7.97 | <0.001 | |||
| Mar-21 | 52 (13) | 239 | 63 (14) | 171 | 8.55 | <0.001 | |||
| Jan-22 | 66 (18) | 205 | 58 (19) | 241 | 4.39 | <0.001 | |||
| Mar-23 | 70 (13) | 227 | 61 (18) | 205 | 5.66 | <0.001 |
Table 2.
Abundances of male and female slimy sculpin within kill and reference zones in Garvin Brook on five dates post-kill. Contingency table Chi-square test results comparing.
Table 2.
Abundances of male and female slimy sculpin within kill and reference zones in Garvin Brook on five dates post-kill. Contingency table Chi-square test results comparing.
| Kill zone | Reference zone | |||||||
|---|---|---|---|---|---|---|---|---|
| Date | Males | Females | Males | Females | X2 | P | ||
| Mar-20 | 19 | 31 | 27 | 56 | 0.43 | 0.521 | ||
| Mar-21 | 29 | 25 | 40 | 35 | 0.002 | 0.967 | ||
| Jan-22 | 68 | 99 | 54 | 55 | 2.08 | 0.149 | ||
| Mar-23 | 69 | 60 | 74 | 110 | 5.38 | 0.02 |
Table 3.
Summary t-test statistics (t values and P values) comparing total, adult, and juvenile sculpin electrofishing catch per effort (CPE) between kill and reference zones in Garvin Brook on five dates post-kill.
Table 3.
Summary t-test statistics (t values and P values) comparing total, adult, and juvenile sculpin electrofishing catch per effort (CPE) between kill and reference zones in Garvin Brook on five dates post-kill.
| Total CPE | Adult CPE | Juvenile CPE | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Date | t | P | t | P | t | P | |||
| Mar-20 | 4.49 | 0.0002 | 5.48 | <0.0001 | 1.54 | 0.14 | |||
| Aug-20 | 1.66 | 0.111 | 4.65 | 0.0001 | 3.56 | 0.002 | |||
| Mar-21 | 0.9 | 0.375 | 2.32 | 0.03 | 2.42 | 0.024 | |||
| Jan-22 | 1.79 | 0.097 | 1.07 | 0.298 | 2.77 | 0.018 | |||
| Mar-23 | 1.18 | 0.252 | 1.57 | 0.136 | 4.27 | <0.0001 |
Table 4.
Change in slimy sculpin catch per effort (CPE, fish/min) versus sampling site (downstream to upstream) in kill and reference zones of Garvin Brook, March 2020, based on linear regression statistics.
Table 4.
Change in slimy sculpin catch per effort (CPE, fish/min) versus sampling site (downstream to upstream) in kill and reference zones of Garvin Brook, March 2020, based on linear regression statistics.
| Zone/statistic | Total CPE | Adult CPE | Juvenile CPE | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Kill zone | |||||||||
| Slope | –0.226 | –0.136 | –0.089 | ||||||
| Y intercept | 4.5 | 2.524 | 1.976 | ||||||
| r2 | 0.276 | 0.295 | 0.141 | ||||||
| t value (one-tailed) | 1.95 | 2.05 | 1.28 | ||||||
| Degrees of freedom | 10 | 10 | 10 | ||||||
| P value | 0.04 | 0.034 | 0.115 | ||||||
| Reference zone | |||||||||
| Slope | 0.356 | 0.162 | 0.194 | ||||||
| Y intercept | 4.329 | 3.334 | 0.995 | ||||||
| r2 | 0.339 | 0.141 | 0.249 | ||||||
| t value (one-tailed) | 2.03 | 1.25 | 1.63 | ||||||
| Degrees of freedom | 8 | 8 | 8 | ||||||
| P value | 0.038 | 0.142 | 0.071 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



