Submitted:
09 December 2023
Posted:
11 December 2023
You are already at the latest version
Abstract
Training has a significative effect on physiology of blood coagulation both in humans and in horses. Several hemostatic changes have been reported after exercise in the horse but data available are inconclusive. The aim of this study was to investigate the platelet activation and primary platelet-related hemostasis modifications in young untrained Thoroughbred racehorses in the first 4-month timeframe incremental training period in order to improve knowledge on this topic.
Twenty-nine clinically healthy, untrained, 2-year-old Thoroughbred racehorses.
Never trained Thoroughbreds were followed during their incremental 4-month sprint exercise schedule. Blood collection was performed once a month (T-30, T0, T30, T60, and T90). Platelet aggregation was measured by light transmission aggregometer in response to various agonists: adenosine diphosphate (ADP), collagen and calcium ionophore A23187. Platelet function was evaluated using the Platelet Function Analyzer (PFA-100®) using collagen/ADP and collagen/adrenaline cartridges. Nitrite-nitrate (NOx) plasma concentrations were measured by a colorimetric assay to assess in vivo nitric oxide bioavailability. Platelet activation was investigated also through gene expression analyses (selectin P-SELP, ectonucleotidase CD39-ENTPD1, prostaglandin I2 synthase-PTGIS, endothelial nitric oxide synthase 3-NOS3). Differences among the time points were analyzed and mean±SEM were calculated. Significant modifications were identified compared with T-30 with increase of platelet aggregation (collagen:32.6±4.8 vs 21.6±4.9%; ADP: 35.5±2.0 vs 24.5±3.1%; A23187: 30±4.7 vs 23.8±4%) and a shorter closure time of C-ADP cartridges (75.6±4.4 vs 87.7±3.4 sec) that tended to return to baseline value at T90. NOx concentrations in plasma significantly increased after 30 days of training program compared with baseline. The first long-term training period induce platelet hyperactivity already after 30 days in untrained Thoroughbreds. Regular physical training blunts the negative effects of acute efforts on platelets activation.
Keywords:
coagulation
; platelet aggregation
; equine
; first long-term training
; Thoroughbred racehorse
1. Introduction
Intense physical training has been proven to have a strong effect on blood coagulation in humans and in horses [1,2,3,4,5,6,7]. In horses, contrasting data have been reported regarding hemostatic changes after exercise. Moreover, most studies have been performed on the occasion of acute exercise [4,6,7,8,9,10].
Blood coagulation and fibrinolysis represent two vital physiological systems, regulated by a complex balance between activators and inhibitors. Several studies have shown that exercise affects the hemostatic equilibrium with a transient increase in blood coagulability counterbalanced by a simultaneous increase in the fibrinolytic status and that it depends on the intensity and duration of exercise [5,11]. S studies explored the long-term effects of exercise on the coagulative balance in racehorses and they only focused on the modifications of clotting parameters [1,5] and platelet aggregation [3,12]. Similarly to humans, these studies seem to indicate a condition of hypercoagulability in athletic horses, instrumental to a prompt reaction to possible microvascular trauma induced by exercise [1,13]. This has been considered as a physiological reaction of the hemostatic system to long-term exercise. Moreover, a variable degree of enhanced platelet aggregation has been reported during different training schedules and interpreted as a protective endothelial mechanism [3]. Nevertheless, data about platelet aggregation in athletic horses are few and conflicting [1,3,12].
The physiological regulation of platelet reactivity is tightly controlled by balancing prothrombotic and antithrombotic signals [14,15]. In humans, nitric oxide (NO), a gaseous signaling molecule involved in many physiological functions, is one of the potent inhibitors of platelet activation. Its antithrombotic effect is the consequence of the blunting of human platelet adhesion to the endothelium and aggregation [14,15].
The potential effect of NO on platelet activation pathways has not been explored in detail in horses and no studies have been carried out on NO regulation of platelet function in race horses during the training period.
Aim of our study was to investigate changes in platelet activation and NO activity as well as of the expression of some genes related to hemostasis and inflammation (selectin P-SELP, ectonucleotidase triphosphate diphosphohydrolase-1 CD39-ENTPD1, prostaglandin I2 synthase-PTGIS, and nitric oxide synthase 3-NOS3) [16,17,18,19,20,21], in young, never trained, Thoroughbreds during the first incremental exercise period, in order to deeply understand the dynamics of the coagulative system in this athletic horse. Monitoring of blood hemostasis could provide useful information about the fitness level of the animals during exercise and competition and might contribute to the development of more adequate training programs.
2. Material and Methods
2.1. Animals
Twenty-nine Thoroughbreds (17 males and 12 females; two years old, weight 450–550 kg, height 165–168 cm) were enrolled (same animals included in our previous studies [13,22]. Horses were clinically healthy (no previous coagulative abnormalities) based on heart examination, thorax auscultation and rectal temperature. Hemato-biochemical and hemostatic analyses were within reference ranges. No pharmacological treatment was assumed prior and during the study.
Horses were housed in an approved facility and in accordance with protocols prescribed by the Institutional Animal Care and Use Committee. All horses were managed with same feeding and housing schedule. These animals were never trained for flat racing (canter and gallop) before the beginning of the study. Each horse followed a feeding program to maintain health, performance and well-being during training. Water was available ad libitum.
Horses were trained with the same training schedule (Table 1) for a 4-time months’ period (from April to July 2018, from Monday to Saturday with Sunday rest). Clinical evaluation and blood samples were taken at rest: 30 days before the beginning of the official training program when horses had never trained before when they were houses together (T-30), on the day of the beginning of the training program which included canter and gallop (T0) and 30 days (T30), 60 days (T60) and 90 days (T90) after the incremental increase of the training program, in order to assess whether or not animals have adapted to the exercise (Table 1). Each animal had one or more races after the end of the experimental period. No horses showed poor-performance syndrome during the study period or the races.
2.2. Sample Collection
Blood sampling activity was performed once a month, from March 2018 to July 2018, at 6:30 am, at rest, before training and feeding. March (T-30, untrained horses), horses were houses together and begin to have the same environmental conditions of nutrition and milde exercise; April (T0, horses starting gallop) was the first month of beginning of training simulating competitions (gallop; samples were taken at rest on the first day of starting training) From April to July, training was incremental (T30, T60, T60, T90, horses under incremental training program). Blood samples were collected directly into vacutainer tubes by venipuncture of the jugular vein, avoiding unnecessary manipulation that could induce activation of coagulation. Blood samples were collected into Vacutainer tubes (Terumo Corporation, BD brand; Tokyo, Japan) with K3-ethylenediamine tetra acetic acid-EDTA 3.8% sodium citrate). K3-EDTA tubes were used to evaluate the expression of target genes in RT-qRCR system. Sodium citrated tubes was used to evaluate platelet function and for nitrite-nitrate (NOx) assay. Sodium citrated blood tubes were centrifuged at 1500g for 15 min to obtain citrated plasma samples within 15 minutes following collection. All samples were immediately transported to the coagulation laboratory and were analyzed within 3h hours after blood collection.
2.3. Light Transmission Aggregometry
To study platelet aggregation, platelet-rich plasma (PRP) was obtained by centrifugation of citrated blood at 120g for 15 min at room temperature. The upper 2/3 of PRP were carefully removed using a plastic pipette and transferred into plastic tubes. Platelet-poor plasma (PPP) was prepared by further centrifugation of the remaining blood at 4000g for 10min [23].
Platelet aggregation was carried out using an optical aggregometer (APACT-4, Helena Biosciences, UK), as previously described [24]. Agonists used were adenosine diphosphate (ADP) 1, 5, and 10µM, collagen 4 µg/mL and calcium ionophore A23187 5µM [23,25].
Results are expressed as maximal amplitude of aggregation measured within 5 minutes of stimulation.
2.4. Platelet Function Analyzer
Platelet function analyzer (PFA-100®, Siemens Healthcare, Malvern, USA) is a point-of-care platelet function analyzer that evaluates platelet function in a small volume of whole blood in high shear-stress conditions and in a semiautomatic system [26,27]. Recent studies have reported the use of the PFA-100® for the evaluation of primary haemostasis also in horses [28]. The device simulates the process of primary haemostasis by triggering platelet adhesion and hemostatic plug formation under high shear flow conditions. Whole blood is aspirated through an aperture through a collagen membrane coated with plate let agonists (collagen and adenosine diphosphate (C-ADP) or col lagen and epinephrine (C-EPI)). This design mimics in vivo endothelial damage and platelet plug formation [27]. Time until plug formation is measured in seconds and is defined closure time [24]. The test was performed ac cording to the manufacturer’s instructions.
2.5. Nitrite-Nitrate (NOx)
As NO˙ has a very short half-life, the plasma concentrations of oxidized NO˙ metabolites (NOx) nitrate (NO3-) and nitrite (NO2-), is frequently used to assess NO bioavailability in vivo. Nitrite-nitrate (NOx) plasma concentration, an indirect measurement of NO generation in biological fluids, were measured by a colorimetric assay kit (R&D system Minneapolis, USA), according to the manufacturer’s instructions as previously described [29,30] .
2.6. RT-qPCR Analyses
We examined gene expression profiles in peripheral blood leukocytes of horses during the training program. mRNA expression values were normalized with three reference genes (SDHA, B2M and HPRT). Total RNA was extracted from buffy coat recovered from 10 mL of total blood as previously described [22,31]. The resulting pellets were resuspended in 2 ml of TriZol reagent at room temperature (Thermo Fisher Scientific, Waltham, MA, USA) and RNA isolated using TriZol Plus RNA purification kit (Thermoficher Scientific, Waltham, MA, USA), as previously described from T-30, T0, T30 and T90 samples. The RNA was cheeked for quantity using the NanoDrop2000 spectrophotometer (Thermoficher Scientific, Waltham, MA, USA) and for quality through electrophoresis in a denaturing 1.2% agarose gel.
One microgram (1 ug) of total RNA of each sample was reverse-transcribed using the SuperScript®VILO IVTM Master Mix (Thermoficher Scientific, Waltham, MA, USA) according to the manufacturer’s recommendations. Primers for reference genes Succinate dehydrogenase complex, subunit A (SDHA) and Beta-2 microglobulin (B2M) optimal housekeeping for blood cells in horses, as well as IL-1β genes of interest were previously determined [32], while for genes of interest SELP, ENTPD1, PTGIS and NOS3 primers were designed on available sequences using the Primer-BLAST software (https://www.ncbi.nlm.nih.gov/tools/primer-blast/), trying to place them in different exons or at exon–exon junctions to avoid biases due to genomic DNA amplification. Primer sequences and accession numbers for tested genes are listed in Table 2. The relative normalized expression was calculated through the 2−∆∆Ct method, using T0 as control through the CFX Maestro software (ver. 4.1- BioRad, Hercules, CA, USA).
2.7. Statistical Analysis
All results are expressed as mean ± standard error (SEM). Differences among time points were analyzed by the one-way repeated measures ANOVA. A p value <0.05 was considered statistically significant.
RT-qPCR samples were divided into groups (T-30, T0, T30 and T90) and the Shapiro-Wilk test was used to determine the likelihood that the expression values of the samples in a biological group were obtained from a normally distributed population. Then, modifications in the relative gene expression between groups were evaluated by one-way ANOVA. Compact letter displays (CLD) was used to highlight significance (p < 0.05). Data are expressed as means of fold change with the standard error using CFX maestro software (ver. 4.1- BioRad, Hercules, CA, USA).
3. Results
3.1. Effect of Training on Platelet Activation
Platelet aggregation induced by low concentrations of ADP, by collagen and by A23187 increased significantly after 30 or 60 days of training program compared with baseline (T0) (Table 3 and Figure 1). Exercise had no significant effect instead on platelet responses to higher concentrations of ADP (Table 3). Concomitantly, a shorter closure time of the C-ADP cartridge (70.0±4.2 vs. 89.8±4.1 sec) and of C-EPI cartridge (190.0±21 vs. 242.4±21 sec) with the PFA-100®analyzer was observed at T30 compared to baseline (Table 4 and Figure 2), confirming platelet hyperreactivity (Table 4). Most of these parameters tended to return to the basal values at the end of training period.
3.2. NO Metabolism
Compared to the values measured at T0, physical exercise significantly increased NOx concentrations in plasma at T30 (Figure 3).
3.3. RT-qPCR Analyses
Reference genes showed high stability, with an M value below the accepted threshold [33]. Differences in gene expression for target genes were assessed setting T0 as control time and carrying out pairwise comparisons between all the different sampling times. All the tested genes (SELP, ENTPD1, PTGIS and NOS3) resulted modulated (Figure 4), with PTGIS and NOS3 showing a decrease in T0 and T30 compared to T-30 and both increasing again in T90. Indeed, NOS3 returned to the same level of T-30 while PTGIS, although up-regulated, maintained lower expression levels with respect to T-30. For ENTPD1 and SELP an opposite trend with respect to the first two genes was assessed. ENTPD1 showed a marked increase in T0 compared to T-30 and a progressive reduction in T30 and T90, although the expression levels for these two time points were kept higher compared to T-30. SELP was characterized by an increased expression in T0 compared to T-30 which decreased again in T30 and further in T90, returning at initial expression levels for the last time point.
4. Discussion
Our study shows that the first long-term training period in untrained Thoroughbreds induces significant changes in platelet activation which become apparent already after 30 days. In fact, a transiently increased platelet aggregation induced by collagen, low dose of ADP and calcium ionophore A23187 associated with a shortening of the PFA-100® closure time were observed. These effects of training on platelet activation tended to disappear after 90 days, indicating an adaptation of the primary hemostasis system to long-term exercise training.
Only a few studies have assessed the influence of training on platelet aggregation in thoroughbred race horses and most of these were focused on the acute effects of one single strenuous exercise which seems to transiently inhibit platelet reactivity, possibly due to a protective activation of the endothelium with the production of nitric oxide in response to physical exercise [4,10]. Nevertheless, the results of these studies are conflicting due to differences in the anticoagulants used for blood sampling, analytical methods used, and the effect of different training programs (exercise intensity, duration, and fitness status) [2,3,34,35].
Here we show for the first time that young Thoroughbreds during their first long incremental training season display an enhanced platelet activation. We demonstrate an increased aggregation of platelets after 30 days of training with a subsequent tendency to return to basal concentrations with the continuation of training. These results suggest the development of a condition of transient platelet hyperactivity soon after the start of intensive training which tends to subside with the habituation to physical exercise. Our data are in accordance with data reported in sportsmen undergoing strenuous long training [2], but they differ from previous studies in Thoroughbreds [12] and in jumping horses evaluated during 5 weeks of training [3], probably due to different analytical methods used and fitness status of the animals. To our knowledge, most of the published studies evaluating platelet aggregation in horses used impedance aggregometry [3,12,36,37,38] and only rarely light transmission aggregometry [39,40], in some cases with poorly validated devices. Light transmission aggregometry is considered the gold standard test for the assessment of platelet function and for the monitoring of bleeding risk [27] while impedance aggregometry did not appear to have sufficient sensitivity or specificity to detect platelet function alterations. Finally, almost all previous studies used a single agonist, e.g. ADP [3,12,37,38] or collagen [41] while we evaluated various pathways of platelet activation by using a panel of agonists and, for the first time in the horse, by the calcium ionophore A23187 that seems to be active on platelets in this species as well as ADP and collagen. More studies are needed to standardize platelet agonists for horse platelet studies and to identify reference values for the gold standard light transmission aggregometry.
Over the last decades, assessment of platelet function in vitro has been used in human medicine to study primary hemostasis. The PFA-100® analyzer evaluates platelet function in vitro in whole blood by simulating platelet adhesion, secretion and aggregation after a vascular injury. Moreover, PFA-100®, differently from LTA, is a test that relies on high shear rates. In veterinary medicine this method has been recently shown to be sensitive and accurate in the diagnosis of platelet dysfunction in pigs, dogs and horses using species-specific reference values [28]. We analyzed, for the first time, platelet function in young untrained Thoroughbreds during the first long term training period by using the PFA-100®. In our study, a shorter C-ADP and C-EPI closure time at T30 confirmed increased platelet reactivity, which tended then to return to basal concentrations at the subsequent time points. The mean CT identified in Thoroughbreds at T0 was similar to that previously identified (85.1±13.1 s) in normal horses [28].
Our results also show increased plasma concentrations of the NO degradation products nitrite and nitrate after 30 days of incremental training simulating competitions. Previous human and animal studies have reported increased plasma concentrations of the NO degradation products nitrite and nitrate after acute exercise [42]. As an increased generation of NO might be counterbalanced by its inactivation, for example, by reactive oxygen species (ROS), measured concentrations of NO metabolites do not always indicate biologically active NO. In fact, during physical activity, muscles spasm, accompanied by damage to the muscle fibers, initiates an inflammatory response to exercise and pro-inflammatory cytokines, like IL-1β, activate neutrophils and nicotinamide adenine dinucleotide phosphate (NADPH) oxidase enzymes, which in turn enhance the production of ROS and free radicals. Our results suggest that studies on NO bioavailability in horses during exercise training are warranted. Unfortunately, we did not evaluate ROS and free radicals in the horses included in this study.
We also assessed the expression of some genes known to be related to hemostasis and inflammation in humans (SELP-selectin P, ENTPD1-ectonucleotidase CD39, PTGIS-prostaglandin I2 synthase, NOS3-endothelial nitric oxide synthase 3) to deeply investigate the observed exercise-induced changes in platelet activation.
SELP encodes a 140 kDa protein (P-selectin) constitutively expressed by platelets and endothelial cells, essential for the interactions between blood cells and the endothelium and involved in the inflammation response. Our data show that after 30 days of exercise SELP expression increased return to baseline values after long term exercise (T90), suggesting adaptation to exercise training. Endothelium-derived inhibitory mechanisms play an important role in regulation of platelet activation. These mainly include nitric oxide (NO), prostacyclin PGI2, and adenosine, which are synthesized by endothelial NO synthases (eNOS), prostacyclin synthase (PTGHS), and CD39/CD73, respectively. Vascular laminar shear stress increases during exercise and is associated with a rapid up-regulation of endothelial nitric oxide synthase (eNOS) mRNA and protein expression levels [43]. Previous studies in rats showed that exercise-training increased endothelial NO synthesis suggesting that increased production of endothelial NO constitutes an initial phase in the adaptive response to exercise training [44]. Our results show an increase in eNOS transcription between initial phases of training (T0 and T30) and the end of the monitored training period (T90) whereas eNOS expression was unchanged between T-30 and T90.
Prostacyclin synthase catalyzes the last step of prostaglandin I2 synthesis, a potent inhibitor of platelet aggregation. Interestingly the gene encoding for prostacyclin synthase, PTGIS, was downregulated at all-time points compared to T-30 suggesting a possible reduced prostaglandin I2 -mediated inhibition of platelet aggregation. Meanwhile, a partial attempt to increase PTGIS expression at T90 compared to T0 and T30, probably indicating an adaptation to training and a effort to restoration of homeostasis.
CD39/CD73 constitutes a further pathway that regulates platelet function. ENTPD1 gene coding for CD39 a protein implicated in the prevention of platelet aggregation by hydrolyzing platelet-activating ADP to AMP. In platelets, adenosine triphosphate (ATP) and adenosine diphosphate (ADP) are stored in dense granules and released upon activation. ATP and ADP interact with specific receptors on platelets (P2Y1 and P2Y12) to activate and recruit additional platelets to the site of vascular injury to form a thrombus. Within the vasculature, ATP and ADP concentrations are regulated by transmembrane nucleotidases that rapidly degrade ATP and ADP to adenosine thus diminishing platelet activation and preventing thrombotic events. There is evidence that in an inflammatory environment, the loss of CD39 activity from endothelium sustains platelet aggregation and thrombogenesis [45]. In our white blood cell samples, an increase in ENTPD1 for all the time period compared to T-30 was observed, indicating a mechanism facilitating platelet activation by exercise. Indeed, vigorous exercise acutely elevates the expression of CD39 on lymphocytes, promoting the degradation of ATP to adenosine via the CD39/CD73 pathway [46]. However, a reduction of ENTPD1 expression characterized T30 and even more T90, indicating a gradual return to the basal situation.
Our study includes the assessment of numerous factors and genes involved in platelet function, with the purpose of fully understanding the dynamics of this system in the athletic horse in response to long-term training programs. It seems that during the first exercise season most parameters had the maximum changes after 1 month (T30) of training in never trained horses and that an adaptive response to conditioning probably began after two months (T60) of incremental training and continues after three months (T90). This agrees with what was found in previous studies regarding platelet count and clotting parameters [13,47]. As in our study, clotting parameters showed increased activity (decreased Prothrombin Time and Thrombin time times, increased Fibrinogen concentrations) already 30 days after the beginning of training but they tended to came back to basal concentrations as an adaptation to the training program solely 3 months later showing a later recovery compared to platelet parameters [13].
Main limitations of this study are the lack of control group not entering training even if each animal has been evaluated 1 month before the training period (T-30) to state the starting point, and the subjective reporting of workloads by trainers. In fact, because of it’s impossible to have a control group at the same age of untrained in Thoroughbreds, we sampling a group of horses that had never been trained before to have a control of untrained subjects. To also reduce the environmental effects, we waited 30 days form housing in which the animals arrived in the same training center which standardized management and diet and begin light exercise (T-30; T0).
The evaluation of platelet function with light transmission aggregometry and the assessment of platelet function in vitro with the use of the PFA-100® in thoroughbreds seems to be promising tools to study primary hemostasis in Thoroughbred and further studies are needed to standardize these gold standard methods for horse platelet studies and to identify reference values.
5. Conclusions
The first long-term exercise period was found to induce platelet hyperactivity after 30 days of training in never trained Thoroughbreds. Regular physical training attenuates the negative consequences of acute effort on platelets activation. This effect may be attributed to training status. It seems probable that the platelet adaptive response to conditioning occurs after two months of training in young untrained Thoroughbred horses during the first training season.
Author Contributions
A. Miglio contributed to conceptualisation, methodology, investigation, formal analysis, writing of the original draft manuscript and visualisation. E. Falcinelli contributed to conceptualisation, methodology, investigation, writing–review and editing. A. Mezzasoma, K. Cappelli and S. Mecocci contributed to formal analysis, writing–review and editing. M. Antognoni and P. Gresele contributed to methodology, supervision, funding acquisition, writing–review and editing. All authors had full access to all study data and they have given final approval to the manuscript.
Funding
This study did not receive any specific institutional, private and corporate financial support
Institutional Review Board Statement
Research ethics committee oversight is not currently required by this journal: The study was performed on material collected during clinical procedures. Blood sampling was approved by the Italian Horse Racing Board and performed by the authorized veterinary practitioner during routine controls to assess the health of the animals in the course of the training season
Informed Consent Statement
Prior to sample collection, written owner consent was obtained for all animals included in the study. Animal care procedures were compliant with the European recommendations (Directive 2010/63/EU) for protecting animals used for scientific purposes
Acknowledgments
We wish to thank Dr. Gianluca Alunni for technical assistance.
Conflicts of Interest
The authors declare no conflict of interest
Abbreviations
SLs, storage lesions; RBC, red blood cells; LR, leukoreduction; pRBC, packed red blood cells; LR-pRBC, leukoreduced pRBC; nLR-pRBC, non-leukoreduced pRBC; CBC, complete blood count; WB, whole blood; CPD-SAGM, Citrate Phosphate Dextrose-Saline Adenine Glucose Mannitol; P, pellet; S, supernatant.
References
- Assenza, A.; Tosto, F.; Casella, S.; Fazio, F.; Giannetto, C.; Piccione, G. Changes in Blood Coagulation Induced by Exercise Training in Young Athletic Horses. Research in Veterinary Science 2013, 95, 1151–1154. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, M.S.; Ali, N.; El-Sayed Ali, Z. Aggregation and Activation of Blood Platelets in Exercise and Training. Sports Med 2005, 35, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Giannetto, C.; Arfuso, F.; Fazio, F.; Giudice, E.; Pietro, S.D.; Bruschetta, D.; Piccione, G. Different Training Schedules Influence Platelet Aggregation in Show Jumping Horses. Polish Journal of Veterinary Sciences 2017, 20, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Kingston, J.K.; Sampson, S.N.; Beard, L.A.; Meyers, K.M.; Sellon, D.C.; Bayly, W.M. The Effect of Supramaximal Exercise on Equine Platelet Function. Equine Vet J Suppl 1999, 181–183. [Google Scholar] [CrossRef] [PubMed]
- Piccione, G.; Bazzano, M.; Giannetto, C.; Marafioti, S.; Fazio, F. Training-Induced Changes in Clotting Parameters of Athletic Horses. J Vet Sci 2014, 15, 45. [Google Scholar] [CrossRef] [PubMed]
- Piccione, G.; Grasso, F.; Fazio, F.; Giudice, E. The Effect of Physical Exercise on the Daily Rhythm of Platelet Aggregation and Body Temperature in Horses. The Veterinary Journal 2008, 176, 216–220. [Google Scholar] [CrossRef]
- Piccione, G.; Fazio, F.; Giudice, E.; Grasso, F.; Caola, G. Exercise-Induced Changes in the Clotting Times and Fibrinolytic Activity during Official 1600 and 2 000 Meters Trot Races in the Standardbred Horses. Acta Vet. Brno 2005, 74, 509–514. [Google Scholar] [CrossRef]
- McKeever, K.H.; Hinchcliff, K.W.; Kociba, G.J.; Reed, S.M.; Muir, W.W. Changes in Coagulation and Fibrinolysis in Horses during Exercise. Am. J. Vet. Res. 1990, 51, 1335–1339. [Google Scholar] [CrossRef] [PubMed]
- Monreal, L.; Angles, A.M.; Monreal, M.; Espada, Y.; Monasterio, J. Changes in Haemostasis in Endurance Horses: Detection by Highly Sensitive ELISA-Tests. Equine Veterinary Journal 1995, 27, 120–123. [Google Scholar] [CrossRef]
- Piccione, G.; Assenza, A.; Casella, S.; Giannetto, C.; Tosto, F.; Caola, G. Modifications of Platelet Aggregation during Treadmill Section and Obstacle Course in Athletic Horse. Acta vet. (Beogr.) 2010, 60, 165–172. [Google Scholar] [CrossRef]
- Karampour, S.; Gaeini, A.A. Response of Coagulation and Anti-Coagulant Factors of Elite Athletes Following Acute Resistance and High-Intensity Interval Training. J Sports Med Phys Fitness 2018, 58, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Piccione, G.; Marafioti, S.; Giannetto, C.; Panzera, M.; Fazio, F. Effect of Dietary Supplementation with Omega 3 on Clotting Time, Fibrinogen Concentration and Platelet Aggregation in the Athletic Horse. Livestock Science 2014, 161, 109–113. [Google Scholar] [CrossRef]
- Miglio, A.; Falcinelli, E.; Mezzasoma, A.M.; Cappelli, K.; Mecocci, S.; Gresele, P.; Antognoni, M.T. Effect of First Long-Term Training on Whole Blood Count and Blood Clotting Parameters in Thoroughbreds. Animals (Basel) 2021, 11, 447. [Google Scholar] [CrossRef]
- Lundberg, J.O.; Carlström, M.; Weitzberg, E. Metabolic Effects of Dietary Nitrate in Health and Disease. Cell Metabolism 2018, 28, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Piknova, B.; Nghiem, K.; Lozier, J.N.; Schechter, A.N. Inhibitory Effect of Nitrite on Coagulation Processes Demonstrated by Thrombelastography. Nitric Oxide 2014, 40, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Barrionuevo, N.; Gatica, S.; Olivares, P.; Cabello-Verrugio, C.; Simon, F. Endothelial Cells Exhibit Two Waves of P-Selectin Surface Aggregation Under Endotoxic and Oxidative Conditions. Protein J 2019, 38, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.; Chi, L.; Zhao, R.; Tourdot, B.E.; Yalavarthi, S.; Jacobs, B.N.; Banka, A.; Liao, H.; Koonse, S.; Anyanwu, A.C.; et al. Ectonucleotidase Tri(Di)Phosphohydrolase-1 (ENTPD-1) Disrupts Inflammasome/Interleukin 1β-Driven Venous Thrombosis. J Clin Invest 2019, 129, 2872–2877. [Google Scholar] [CrossRef] [PubMed]
- Boudreaux, M.K.; Koehler, J.; Habecker, P.L.; Piero, F.D. Evaluation of the Genes Encoding CD39/NTPDase-1 and CD39L1/NTPDase-2 in Horses with and without Abnormal Hemorrhage and in Horses with Pathologic Evidence of Exercise-Induced Pulmonary Hemorrhage. Veterinary Clinical Pathology 2015, 44, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, C.; Yabuki, T.; Inoue, H.; Tone, Y.; Hara, S.; Hatae, T.; Nagata, M.; Takahashi, E.I.; Tanabe, T. Human Gene Encoding Prostacyclin Synthase (PTGIS): Genomic Organization, Chromosomal Localization, and Promoter Activity. Genomics 1996, 36, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Dao, V.T.-V.; Elbatreek, M.H.; Fuchß, T.; Grädler, U.; Schmidt, H.H.H.W.; Shah, A.M.; Wallace, A.; Knowles, R. Nitric Oxide Synthase Inhibitors into the Clinic at Last. Handb Exp Pharmacol 2020. [CrossRef]
- Vedmedovska, N.; Bokucava, D.; Kivite-Urtane, A.; Rovite, V.; Zake-Nikitina, L.; Klovins, J.; Fodina, V.; Donders, G.G.G. The Correlation Between Abnormal Uterine Artery Flow in the First Trimester and Genetic Thrombophilic Alteration: A Prospective Case-Controlled Pilot Study. Diagnostics (Basel) 2020, 10. [Google Scholar] [CrossRef]
- Miglio, A.; Cappelli, K.; Capomaccio, S.; Mecocci, S.; Silvestrelli, M.; Antognoni, M.T. Metabolic and Biomolecular Changes Induced by Incremental Long-Term Training in Young Thoroughbred Racehorses during First Workout Season. Animals (Basel) 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Segura, D.; Monreal, L.; Espada, Y.; Pastor, J.; Mayós, I.; Homedes, J. Assessment of a Platelet Function Analyser in Horses: Reference Range and Influence of a Platelet Aggregation Inhibitor. The Veterinary Journal 2005, 170, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Falcinelli, E.; Francisci, D.; Schiaroli, E.; Minuz, P.; Orsini, S.; Malincarne, L.; Sebastiano, M.; Mezzasoma, A.M.; Pasticci, M.B.; Guglielmini, G.; et al. Effect of Aspirin Treatment on Abacavir-Associated Platelet Hyperreactivity in HIV-Infected Patients. International Journal of Cardiology 2018, 263, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Falcinelli, E.; Guglielmini, G.; Torti, M.; Gresele, P. Intraplatelet Signaling Mechanisms of the Priming Effect of Matrix Metalloproteinase-2 on Platelet Aggregation. J Thromb Haemost 2005, 3, 2526–2535. [Google Scholar] [CrossRef] [PubMed]
- Kundu, S.K.; Heilmann, E.J.; Sio, R.; Garcia, C.; Davidson, R.M.; Ostgaard, R.A. Description of an in Vitro Platelet Function Analyzer--PFA-100. Semin Thromb Hemost 1995, 21 Suppl 2, 106–112. [Google Scholar] [CrossRef]
- Gresele, P.; Bury, L.; Mezzasoma, A.M.; Falcinelli, E. Platelet Function Assays in Diagnosis: An Update. Expert Rev Hematol 2019, 12, 29–46. [Google Scholar] [CrossRef] [PubMed]
- Segura, D.; Monreal, L.; Espada, Y.; Pastor, J.; Mayós, I.; Homedes, J. Assessment of a Platelet Function Analyser in Horses: Reference Range and Influence of a Platelet Aggregation Inhibitor. Vet. J. 2005, 170, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Momi, S.; Caracchini, R.; Falcinelli, E.; Evangelista, S.; Gresele, P. Stimulation of Platelet Nitric Oxide Production by Nebivolol Prevents Thrombosis. Arterioscler Thromb Vasc Biol 2014, 34, 820–829. [Google Scholar] [CrossRef]
- Marañón, G.; Muñoz-Escassi, B.; Manley, W.; García, C.; Cayado, P.; de la Muela, M.S.; Olábarri, B.; León, R.; Vara, E. The Effect of Methyl Sulphonyl Methane Supplementation on Biomarkers of Oxidative Stress in Sport Horses Following Jumping Exercise. Acta Veterinaria Scandinavica 2008, 50, 45. [Google Scholar] [CrossRef]
- Cappelli, K.; Amadori, M.; Mecocci, S.; Miglio, A.; Antognoni, M.T.; Razzuoli, E. Immune Response in Young Thoroughbred Racehorses under Training. Animals 2020, 10, 1809. [Google Scholar] [CrossRef] [PubMed]
- Cappelli, K.; Felicetti, M.; Capomaccio, S.; Nocelli, C.; Silvestrelli, M.; Verini-Supplizi, A. Effect of Training Status on Immune Defence Related Gene Expression in Thoroughbred: Are Genes Ready for the Sprint? The Veterinary Journal 2013, 195, 373–376. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate Normalization of Real-Time Quantitative RT-PCR Data by Geometric Averaging of Multiple Internal Control Genes. Genome Biol 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef] [PubMed]
- Giordano, A.; Meazza, C.; Salvadori, M.; Paltrinieri, S. Thromboelastometric Profiles of Horses Affected by Exercise-Induced Pulmonary Hemorrhages. Veterinary Medicine International 2010, 2010, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kingston, J.K.; Bayly, W.M.; Meyers, K.M.; Sellon, D.C.; Wardrop, K.J. Evaluation of Binding of Fibrinogen and Annexin V to Equine Platelets in Response to Supramaximal Treadmill Exercise. Equine Vet J Suppl 2002, 502–505. [Google Scholar] [CrossRef] [PubMed]
- Kornreich, B.; Enyeart, M.; Jesty, S.A.; Nydam, D.V.; Divers, T. The Effects of Pentoxifylline on Equine Platelet Aggregation. J Vet Intern Med 2010, 24, 1196–1202. [Google Scholar] [CrossRef] [PubMed]
- Lanier, C.J.; Taintor, J.S.; Christopherson, P.W.; Spangler, E.A. Effect of Lactic Acid Addition to Equine Whole Blood on Platelet Aggregation Measured by Impedance Aggregometry. Veterinary Clinical Pathology 2022, 51, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Casella, S.; Giannetto, C.; Fazio, F.; Assenza, A.; Piccione, G. Nictemeral Profile of Platelet Aggregation and Clotting Parameters in Horses during Training. Bulletin of the Veterinary Institute in Puławy 2009, 53, 801–806. [Google Scholar]
- Norris, J.W.; Watson, J.L.; Tablin, F.; Kozikowski, T.A.; Knych, H.K. Pharmacokinetics and Competitive Pharmacodynamics of ADP-Induced Platelet Activation after Oral Administration of Clopidogrel to Horses. Am J Vet Res 2019, 80, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Brooks, M.B.; Divers, T.J.; Watts, A.E.; Ness, S.L.; Frye, A.H.; Stokol, T.; Fubini, S.L. Effects of Clopidogrel on the Platelet Activation Response in Horses. American Journal of Veterinary Research 2013, 74, 1212–1222. [Google Scholar] [CrossRef] [PubMed]
- Roscher, K.A.; Failing, K.; Moritz, A. Inhibition of Platelet Function with Clopidogrel, as Measured with a Novel Whole Blood Impedance Aggregometer in Horses. Vet J 2015, 203, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Heber, S.; Volf, I. Effects of Physical (In)Activity on Platelet Function. Biomed Res Int 2015, 2015, 165078. [Google Scholar] [CrossRef] [PubMed]
- Di Francescomarino, S.; Sciartilli, A.; Di Valerio, V.; Di Baldassarre, A.; Gallina, S. The Effect of Physical Exercise on Endothelial Function. Sports Med 2009, 39, 797–812. [Google Scholar] [CrossRef]
- Green, D.J.; Maiorana, A.; O’Driscoll, G.; Taylor, R. Effect of Exercise Training on Endothelium-Derived Nitric Oxide Function in Humans. The Journal of Physiology 2004, 561, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, B.; Dwyer, K.; Enjyoji, K.; Robson, S.C. Ecto-Nucleotidases of the CD39/NTPDase Family Modulate Platelet Activation and Thrombus Formation: Potential as Therapeutic Targets. Blood Cells Mol Dis 2006, 36, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Lindemann, U.; Moggio, A.; Dutsch, A.; Kessler, T.; Sager, H.B. The Impact of Exercise on Immunity, Metabolism, and Atherosclerosis. International Journal of Molecular Sciences 2023, 24, 3394. [Google Scholar] [CrossRef]
- Rivero, J.-L.L.; Ruz, A.; Martí-Korff, S.; Estepa, J.-C.; Aguilera-Tejero, E.; Werkman, J.; Sobotta, M.; Lindner, A. Effects of Intensity and Duration of Exercise on Muscular Responses to Training of Thoroughbred Racehorses. Journal of Applied Physiology 2007, 102, 1871–1882. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Light transmission aggregometry, expressed as maximal amplitude (MA%), induced by collagen (4Ug/ml) and ADP (1uM) at baseline and during the training program. * p<0.05.
Figure 1.
Light transmission aggregometry, expressed as maximal amplitude (MA%), induced by collagen (4Ug/ml) and ADP (1uM) at baseline and during the training program. * p<0.05.
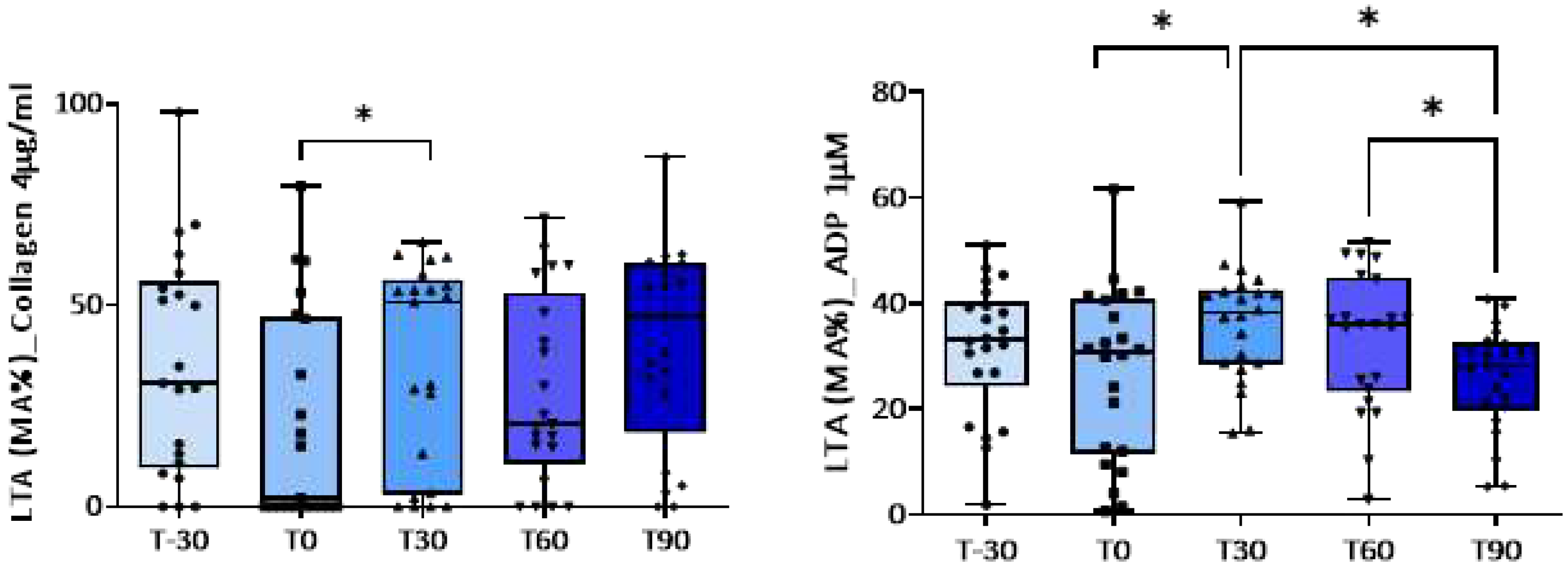
Figure 2.
PFA-100® closure time (sec) with collagen/ADP (C/ADP) and collagen/EPI (C/EPI) cartridges at baseline and during the training program. * p<0.05; **p<0.01; ***p<0.001.
Figure 2.
PFA-100® closure time (sec) with collagen/ADP (C/ADP) and collagen/EPI (C/EPI) cartridges at baseline and during the training program. * p<0.05; **p<0.01; ***p<0.001.
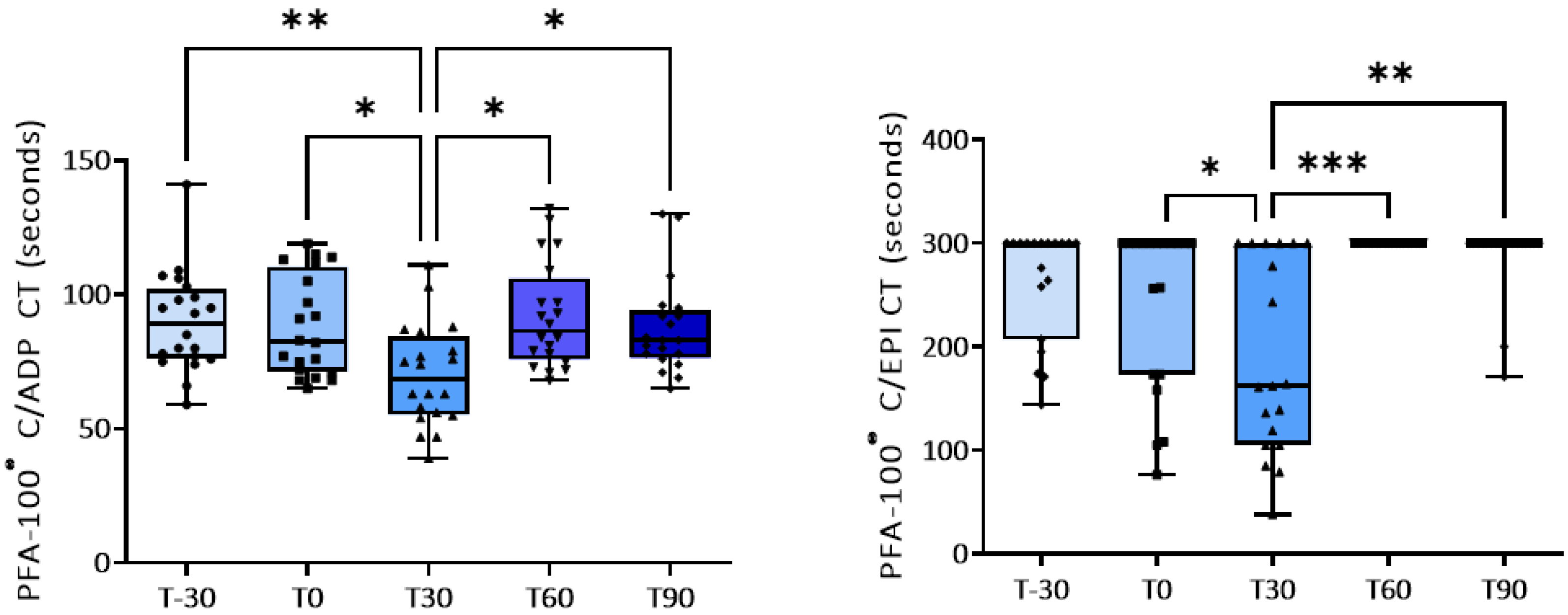
Figure 3.
Plasma concentrations (uM) of nitrite and nitrate (NOx) at baseline and at 30 days of training. * p<0.05.
Figure 3.
Plasma concentrations (uM) of nitrite and nitrate (NOx) at baseline and at 30 days of training. * p<0.05.
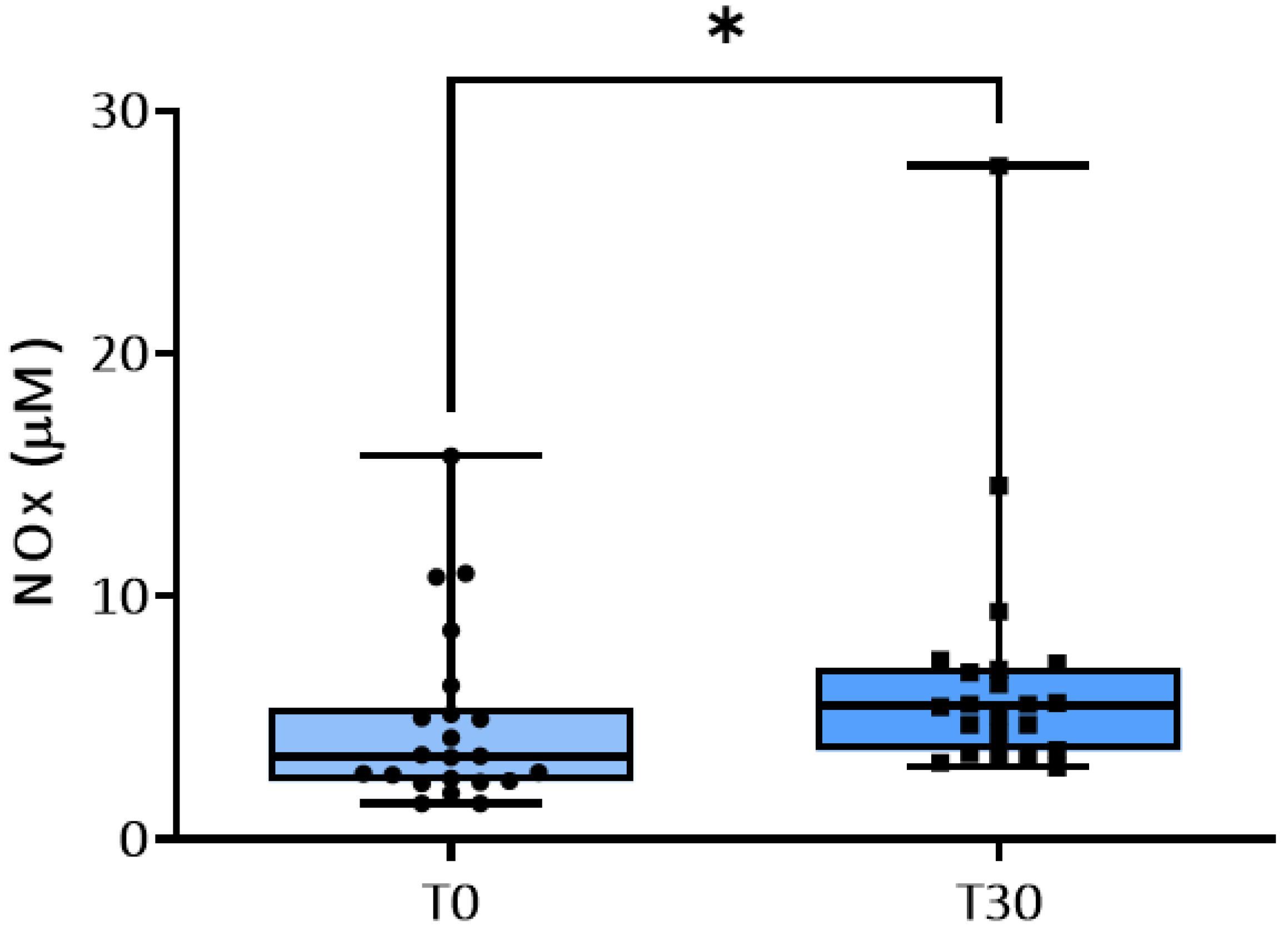
Figure 4.
Histograms representing the relative normalized expression values for the investigated genes at T-30, T0, T30 and T90. Data are expressed as ΔΔCq ± 1 standard error setting as T0 as control group. Statistical significance (p < 0.05) tested pairwise comparisons for all the training times through compact letter displays (CLD). .
Figure 4.
Histograms representing the relative normalized expression values for the investigated genes at T-30, T0, T30 and T90. Data are expressed as ΔΔCq ± 1 standard error setting as T0 as control group. Statistical significance (p < 0.05) tested pairwise comparisons for all the training times through compact letter displays (CLD). .
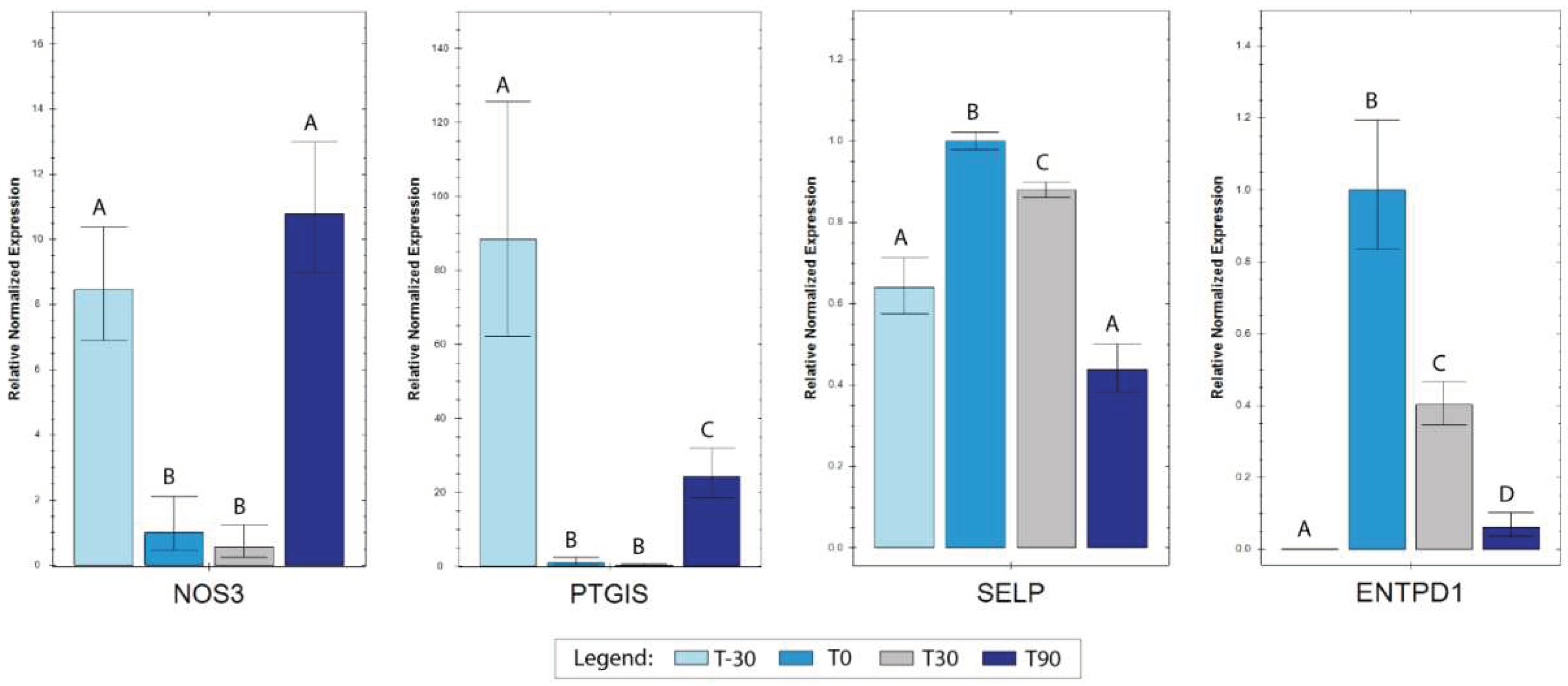

Table 1.
Standard daily training program completed by each horse involved in the study. Speeds: Walk: 100 m/min, Trot: 200 m/min, Canter: 350 m/min, and Gallop: 1000 m/min (min: minutes).
Table 1.
Standard daily training program completed by each horse involved in the study. Speeds: Walk: 100 m/min, Trot: 200 m/min, Canter: 350 m/min, and Gallop: 1000 m/min (min: minutes).
| March (T-30) | April (T0) | May (T30) | June (T60) | July (T90) |
|---|---|---|---|---|
| Untrained horses | Start of training | Incremental training | Incremental training | Incremental training |
| 15 min Walk 10 min Trot Rest 10 min Trot Walk |
15 min Walk 10 min Trot 6 min Canter every Tuesday: 1 min Gallop |
15 min Walk 10 min Trot 6 min Canter every Tuesday: 2 min Gallop |
15 min Walk 10 min Trot 6 min Canter every Tuesday: 3 min Gallop |
15 min Walk 10 min Trot 6 min Canter every Tuesday: 4 min Gallop |
Table 2.
Genes evaluated in this study and relative primer pairs used.
| Gene | Primer Forward | Primer Reverse | Amp. Length |
Accession |
| SELP | GCTGACAATCCAGGAAGCCC | CGCTTTGAGCAGTCAAGGGA | 146 | NM_001081792 |
| ENTPD1 | TTGAGCCACCAAGACCAGAAG | ATTCTGGGTCAACCCCACAG | 125 | XM_001500628 |
| PTGIS | TTCCTGAGTCCGCAGAAGGA | TCGCTTCCCGTCCTTGTAAA | 117 | XM_001501166 |
| NOS3 | TTCGGGAGAGTGAGCTGGTA | CAATCCCGCGCATCAAAGAC | 109 | XM_001504650 |
| B2M | TCCTGCTCGGGCTACTCTC | TGCTGGGTGACGTGAGTAAA | 83 | NM_001082502 |
| SDHA | GCGCGCTTCAGACGATTTAT | CCAGTGCTCCTCAAATGGCT | 146 | XM_014734954 |
Table 3.
Maximal amplitude (MA) of platelet aggregation induced by several agonists at baseline and during the training program. Data are reported as mean±SEM. Significant differences are expressed with p values (p<0.05).
Table 3.
Maximal amplitude (MA) of platelet aggregation induced by several agonists at baseline and during the training program. Data are reported as mean±SEM. Significant differences are expressed with p values (p<0.05).
| Maximal amplitude (%) | T-30 | T0 | T30 | T60 | T90 |
|---|---|---|---|---|---|
| Collagen (4g/ml) | 35.4±6.0 | 21.0±5.7 | 35.0±5.0 p=0.05 vs. T0 |
28.8±5.1 | 40.4±5.4 |
| ADP (1M) | 31.5±2.7 | 26.9±3.4 | 35.9±2.2 p=0.008 vs. T0; p=0.013 vs. T90 |
33.1±2.8 p=0.029 vs. T90 |
25.7±2.1 |
| ADP (5M) | 58.4±3.7 | 57.8±1.9 | 61.0±3.0 | 60.8±4.6 | 60.5±3.6 |
| ADP (10M) | 63.5±2.3 | 63.3±1.6 | 64.4±2.2 | 59.3±2.2 | 62.3±2.1 |
| A23187 (5M) | Not done | 24.2±5.1 | 30.9±5.2 | 41.8±5.3 p=0.03 vs. T0; p=0.04 vs. T90 |
22.0±4.9 |
Table 4.
PFA-100® closure times (sec) with collagen(C)-ADP and collagen(C)-EPI cartridges at baseline and during the training program. Data are reported as mean±SEM. Significant differences are expressed with p values (p<0.05).
Table 4.
PFA-100® closure times (sec) with collagen(C)-ADP and collagen(C)-EPI cartridges at baseline and during the training program. Data are reported as mean±SEM. Significant differences are expressed with p values (p<0.05).
| PFA-100® (sec) | T-30 | T0 | T30 | T60 | T90 |
|---|---|---|---|---|---|
| C-ADP | 89.8±4.1 | 88.2±4.1 | 70.0±4.2 p=0.03 vs. T0; p=0.0021 vs. T-30 |
92.0±4.4 p=0.011 vs. T30 |
87.6±3.9 p=0.019 vs. T30 |
| C-EPI | 262.0±12.5 | 242.4±18.0 | 190.0±21.0 p=0.045 vs. T0 |
300±0.0 p=0.0006 vs. T30 |
287.0±8.3 p=0.0021 vs. T30 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



