
Submitted:
23 November 2023
Posted:
23 November 2023
You are already at the latest version
Abstract
Ureaplasma species (Ureaplasma spp.) are commonly found as commensals in the human urogenital tracts , but their overgrowth can lead to infection in the urogenital tract and at distal sites. Treatment of Ureaplasma spp. infections is difficult. The lack of a cell wall confers resistance to all β-lactam and glycopeptide antibiotics whereas the lack of de novo synthesis of folic acid makes them resistant to sulfonamides and diaminopyrimidines. Thus, there is a need to search for new drugs effective against these bacteria.
In the presented studies, the effect of N,N-diethyldithiocarbamate (DDC) at a concentration of 2 µg/ml, 20 µg/ml and 200 µg/ml on the growth and survival of Ureaplasma urealyticum and Ureaplasma parvum was tested. Our results indicated that all DDC concentrations used showed both bacteriostatic and bactericidal activity against both tested strains
In brief, for the first time we showed the bacteriostatic and bactericidal activity of DDC against Ureplasma urealyticum and Ureaplasma parvum. Based on literature data, we conclude that this DDC activity is related to the inhibition of urease activity, which is an indispensable enzyme for these bacteria. Further research is needed and the results obtained in the present study should be verified in in vivo models of infection.
Keywords:
Ureaplasma urealyticum
; Ureaplasma parvum
; N
; N-diethyldithiocarbamate
; disulfiram
; urease
1. Introduction
The Ureaplasma is the genus of bacteria belonging to the family Mycoplasmataceae in the order Mycoplasmatales. Members of the family Mycoplasmataceae include small pleomorphic bacteria that characteristically lack a cell wall. The genus Ureaplasma is biochemically unique in that all its members possess urease and therefore hydrolyze urea to produce ATP. Ureaplasma species (Ureaplasma spp.) are commonly found as commensals in the human urogenital tracts but their overgrowth can lead to infection in the urogenital tract and, rarely, at distal sites [1,2]. Ureaplasma urealyticum (U. urealyticum) and Ureaplasma parvum (U. parvum) formerly considered as two distinct biovars of U. urealyticum, are now classified as separate species and are the only ureaplasmas associated with disease in humans. U. urealyticum was first isolated in men with non-gonococcal urethritis (NGU) in 1954 [3]. Some authors have reported that U. urealyticum strains are detected more often in men with NGU than in men without NGU [4,5,6]. It has also been demonstrated that U. parvum strains are found more often than U. urealyticum strains in men without NGU [7,8]. It has also been shown that U. urealyticum may be the causative agent in some cases of chronic prostatitis [9].
However, due to the anatomical differences in the structure of the urogenital tract of women and men Ureaplasma spp. are more often identified in women than in men. Epidemiological data indicate that nearly 90% of sexually active healthy women are carriers of Ureaplasma spp. [10]. As already mentioned, Ureaplasma spp. are generally regarded as low-virulent commensals. It is known, however, that colonization of the genital tract with Ureaplasma spp. increases especially in the third trimester of pregnancy. Intraamniotic Ureaplasma spp. infections increase the risk for chorioamnionitis, premature rupture of membranes, and preterm birth. Moreover, these bacteria may be transmitted from mothers to neonates, either in utero or during passage through the infected birth canal. Vertical transmission rates are maximal among preterm infants with very low birthweight (<1500g) [11,12].. These microorganisms mainly colonize the respiratory tract of newborns, and their presence can cause numerous life-threatening disorders. There is no clear difference in the pathogenicity of both species of ureaplasmas. According to Katz et al. and Heggie et al. risk of development of bronchopulmonary dysplasia in neonates infected by U. parvum and U. urealyticum is similar [13,14]. However, other authors assess that the chance of developing bronchopulmonary dysplasia in children colonized by U. urealyticum is 56%, and in the case of U. parvum it reaches 25% [15].
Moreover, it has been shown that the presence of U. urealyticum in the mother’s reproductive tract affects also the central nervous system (CNS) of neonates and causes meningitis, intraventricular hemorrhage and hydrocephalus [16]. In addition, there is strong evidence from experimental animal studies and clinical trials indicating that genital ureaplasma infections cause female infertility, mainly because these bacteria can enter the upper genital tract and colonize the endometrium, fallopian tubes and ovaries. This condition known as pelvic inflammatory disease has serious long-term consequences, including infertility [17]. This problem also applies to men. It has been indicted that U. urealyticum produces a toxic factor that impairs sperm function [18,19].
Treatment of Ureaplasma infections is exceptionally difficult. The lack of a cell wall confers resistance to all β-lactam and glycopeptide antibiotics whereas the lack of de novo synthesis of folic acid makes them resistant to sulfonamides and diaminopyrimidines. Based on current knowledge, only four classes of antibiotics are considered effective in treating Ureaplasma infections. These are the antibiotics belonging to the fluoroquinolone, tetracycline, chloramphenicol and macrolide classes [20]. However, the management of Ureaplasma infections is possible only when dealing with antibiotic-susceptible strains. However, many authors point to the growing incidence of antibiotic-resistant strains of Ureaplasma in the population. Research by Khosropour et al. indicated that 57% of individuals with NGU who were infected with Ureaplasma spp. and received antimicrobial therapy with initially azithromycin followed by doxycycline for 7 days, or vice versa, remained still colonized with these bacteria after 6 weeks of therapy [21]. Biernat-Sudolska et al. showed that as many as 90% of U. parvum and 83% of U. urealyticum strains present in endotracheal aspirate from prematurely born neonates were resistant to ciprofloxacin belonging to the fluoroquinolone class [22].
Thus, there is a need to search for new drugs effective against these atypical bacteria. We chose N,N-diethyldithiocarbamate (DDC) for our research. DDC is the main metabolite of disulfiram (DSF) that is an irreversible inhibitor of aldehyde dehydrogenases (ALDHs), enzymes that catalyze the oxidation of aldehydes to carboxylic acids. For this reason, DSF has been used for over 70 years in the treatment of alcohol addiction. However, it is increasingly believed that the use of DSF as a treatment in patients with alcoholism is an unethical therapy because it is connected with the risk of poisoning and even a loss of life. Thus, at present the use of DSF in the treatment of alcoholism has declined. However, there have been reports indicating new areas of pharmacological activity of DSF and its metabolites. For this reason, DSF has been studied as a possible treatment for cancer, and viral, bacterial, parasitic and fungal infections [23,24,25,26,27].
In this study, we examined for the first time the bacteriostatic (effects on the cell division) and bactericidal activity of DDC against U. urealyticum and U. parvum.
2. Material and Methods
2.1. General
Ureaplasma strains isolated in the Chair of Microbiology of the Jagiellonian University Medical College in Kraków (Poland) from women with infections of the urogenital tract were tested in the study. The study involved 90 strains of Ureaplasma spp. including 36 U. urealyticum strains and 54 U. parvum strains.
2.2. Detection and identification of Ureaplasma spp
The vaginal swabs were collected from each woman and placed in BioMerieux transport media. Next, they were subcultured in liquid and solid PPLO media with 1% urea, prepared in-house according to the previously described procedure [Shepard 1986] and parallelly in Mycoplasma IST 2 kit BioMerieux medium (R2). Identification of ureaplasma species was done by PCR using the previously described two pairs of primers specific for the U. urealyticum and U. parvum genes [28]. Reference strains: U. urealyticum (ATCC 27816) and U. parvum (ATCC 27815) were used as the positive control. Negative control contained of distilled water instead of bacteria. The amplified products were visualized under ultraviolet light after agarose gel electrophoresis containing ethidium bromide (EtBr).
2.3. The effect of DDC on Ureaplasma spp
Baseline titers (as a control) were established for each strain by titration of an 18-24 hour culture in growth medium without the addition of DDC. The titer for each Ureaplasma strain was estimated using the color changing units (CCU) assay. Titration was performed in 96-well plates using a series of 10-fold dilutions (10–1 to 10–10). The titer was read after 24-hour incubation at 370C. The CCU is defined as the highest dilution at which there is a color change [29]. In parallel, the effect of DDC at a concentration of 2 µg/ml, 20 µg/ml and 200 µg/ml on the growth and survival of Ureaplasma strains was tested. The highest concentration of 200 μg/ml used in the studies was also the highest non-toxic concentration of DDC for cell culture of the RK13 line, which was previously determined in in vitro studies.
2.4. Bacteriostatic effect of DDC on Ureaplasma cultures
The cultures of each Ureaplasma strain were titrated by making 10-fold serial dilutions (10–1 to 10–10) in growth medium with the addition of the tested DDC concentrations. Titration was carried out in 96-well plates. The titers were read after 24 hours of incubation at 37°C and expressed as CCU/ml. The obtained values were compared with the control titer for a given strain.
2.5. Bactericidal activity of DDC
The culture of each Ureaplasma strain was suspended in medium with the tested concentrations of DDC and was incubated for 24 hours at 37°C. After incubation, the culture was centrifuged (Sigma 113 microcentrifuge; 14,000G/20,000 rpm) and washed twice with saline to remove DDC and then titration was performed by making 10-fold serial dilutions (10–1 to 10–10) with growth medium without the addition of DDC similarly as described above. The obtained values were compared with the control titer for a given strain.
2.6. Statistical analysis
Statistical calculations were carried out with the IBM® SPSS® Statistic 29 (Armonk, NY: IBM Corp, USA). Differences between the study groups were analyzed using Kruskal-Wallis non-parametric tests with Dunn's post-hoc test. Statistical significance was defined as p < 0.05 for all tests.
3. Results
3.1. Bacteriostatic effect of DDC on Ureaplasma cultures
The median value of log titre for control U. parvum cultures was 3 (2, 5) while the median values of log titre for U. parvum cultured in the presence of DDC at a concentration of 200 μg/ml, 20 μg/ml, and 2 μg/ml were 1 (0, 2), 1 (0, 1) and 2 (1, 3) respectively. The differences between U. parvum titers obtained in the control system (without DDC) and the tested system (in the presence of DDC) were statistically significant (p < 0.001) (Figure 1). The median value of log titre for control U. urealyticum cultures was 3 (2, 6) while the median values of log titre for U. urealyticum cultured in the presence of DDC at a concentration of 200 μg/ml, 20 μg/ml, and 2 μg/ml were 1 (0, 1), 1 (1, 2) and 2 (1, 5,75), respectively. The differences between U. urealyticum titers obtained in the control system (without DDC) and the tested system (in the presence of DDC) were statistically significant (p < 0.05) (Figure 1). Therefore, the obtained results showed that the presence of DDC affected the multiplication rate (cell division) of both tested strains.
The obtained results demonstrated that strains of the U. urealyticum are more sensitive to the bacteriostatic effect of DDC at doses of 20 and 200 µg/ml than U. parvum strains. Only the lowest (2 µg/ml) used concentration of DDC inhibited the cell division of a larger percentage of strains from the U. parvum species than from the U. urealyticum species. The bacteriostatic effect of DDC at a dose of 20 µg/ml was observed for 88.9% of U. parvum and 97.2% of U. urealyticum strains. It means that DDC at a dose of 20 µg/ml elicited the most effective bacteriostatic effect against both tested strains (Figure 2).
3.2. Bactericidal activity of DDC on Ureaplasma cultures
The median value of log titre for control U. parvum cultures was 3 (1, 5) while the mean values of log titre for U. parvum cultured in the presence of DDC at a concentration of 200 μg/ml, 20 μg/ml, and 2 μg/ml 1 (0, 2), 0 (0, 2) and 1 (0, 3), respectively (Figure 3). The median value of log titre for control U. urealyticum cultures was 3 (1, 5.75) while the median values of log titre for U. urealyticum cultured in the presence of DDC at a concentration of 200 μg/ml, 20 μg/ml, and 2 μg/ml were 0 (0, 1), 1 (0, 2) and 2 (0, 3.75), respectively (Figure 3).
It should be noted that although the decrease in bacterial titre correlates with the decrease in bacterial numbers, it does not clearly indicate the bactericidal properties of DDC. In order to test whether DDC has such properties, we conducted an experiment to check whether the tested strains cultured in the presence of DDC retained the ability to grow in growth medium without the addition of DDC. The percentage of strains against which DDC bactericidal activity has been demonstrated is shown in Figure 4.
The obtained results indicated that DDC at a dose of 200 μg/ml had a bactericidal effect against 46% of U. parvum strains and 59.4% of U. urealyticum strains. After incubation on the medium with DDC at a dose of 20 μg/ml and then transfer of the bacteria to the medium without DDC, no growth was observed for 54% of U. parvum strains and 46.9% of U. urealyticum strains. DDC at the lowest dose of 2 μg/ml has a bactericidal effect on 42 of U. parvum strains and and 28.1% of U. urealyticum strains.
Thus, the greatest bactericidal effect for U. urealyticum strains was observed at a DDC concentration of 20 µg/ml, and for U. parvum strains at 200 µg/ml.
4. Discussion
It is known that DDC is the major breakdown product of DSF in vivo. One molecule of DSF gives rise to two molecules of DDC by the reduction of intramolecular disulfide bond. In the blood of humans and other animals, DSF is rapidly and completely converted to two molecules of DDC by serum albumin and erythrocyte enzymes [30,31].
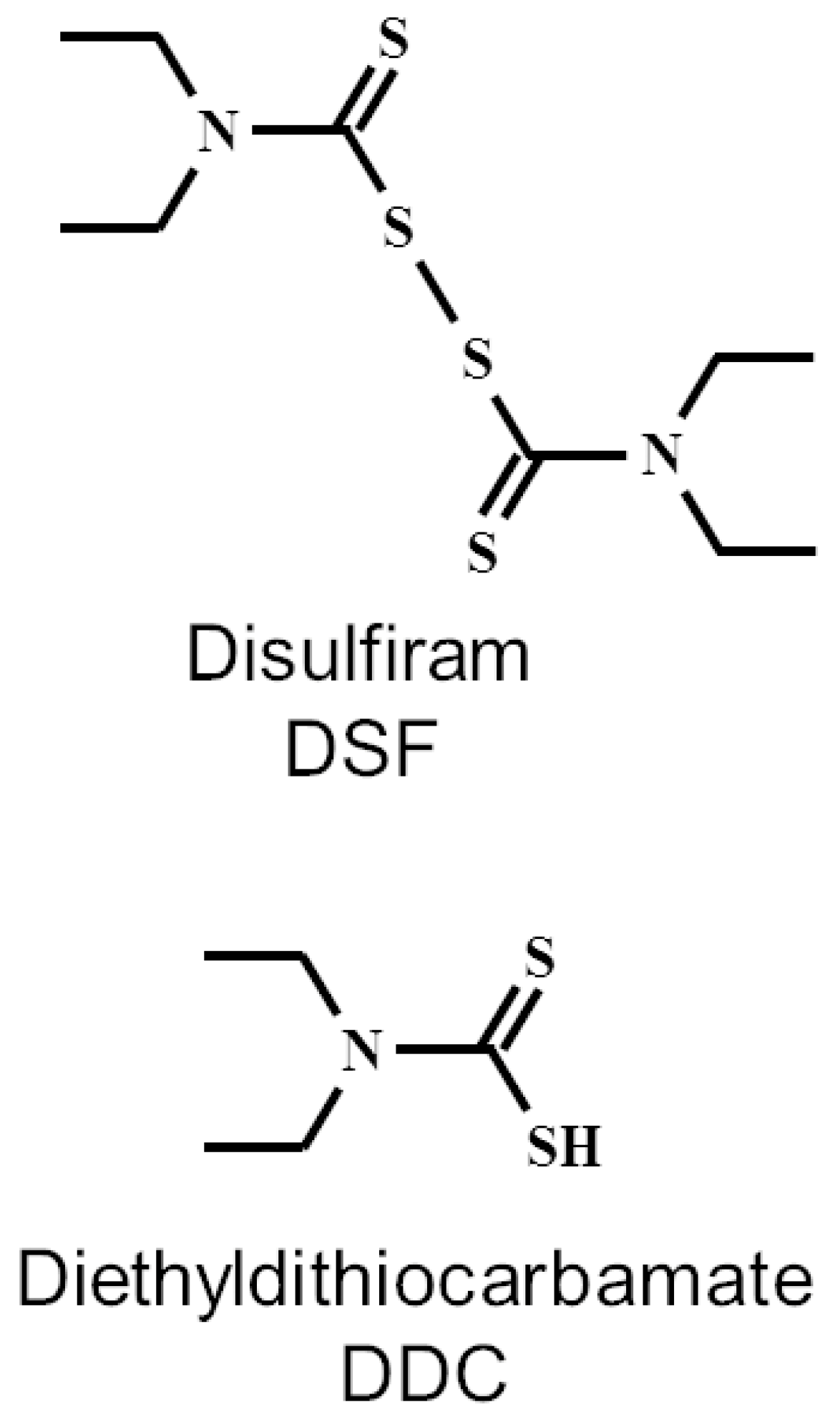
So, DDC is the main metabolite of DSF and is generally believed to be responsible for most of the observed pharmacological effects of DSF. The chemical structure of DSF and DDC is shown in Figure 5.
Our results indicated that all DDC concentrations used in these experiments showed both bacteriostatic and bactericidal activity against both tested Ureaplasma strains. The antimicrobial effect of DSF and its main metabolite DDC has been known for a long time. In 1974 Marshall et al. indicated that DDC had activity against Malassezia pachydermatis (formerly known as Pityrosporum canis) that has been found to cause skin and ear infections, most often occurring in canines. The same authors showed that in addition to having antifungal activity, DDC killed ear mites (Otodectes cynotis) in cats [32]. It is worth recalling that otodectosis is the most commonly diagnosed ectoparasitic disease in cats. In 1979 Scheibel et al. showed that DSF inhibited in vitro the growth of the human malaria parasite Plasmodium falciparum [33].
The studies conducted in the 21st century confirm the antifungal and antiparasitic potential of DSF and DDC. Khan et al. indicated that DSF showed fungicidal activity on Aspergillus fumigatus (A. fumigatus), Aspergillus flavus (A. flavus) and Aspergillus niger (A. niger) [27]. It has also been indicated that DDC showed high antifungal activity against Candida albicans (C. albicans) and Candida tropicalis (C. tropicalis) biofilms [34]. Khouri et al. demonstrated that DDC dose-dependently induced parasite killing and was able to "sterilize" Leishmania amazonensis (L. amazonensis) infection at 2 mM in human macrophage in vitro cultures [35]. Celes et al. tested a topical formulation of bacterial cellulose (BC) membranes containing DDC. The obtained results indicated that exposure of Leishmania braziliensis (L. braziliensis)-infected cells to BC-DDC decreased the parasite load in a significant manner [36]. In 1987 Taylor et. al. demonstrated that DDC inhibited the growth of methicillin-resistant Staphylococcus aureus (S. aureus) (MRSA) in vitro [37]. Later research showed that DSF also inhibited the growth of MRSA [38]. Multidrug-resistant (MDR) S. aureus is associated with high rates of serious disease and treatment failures. It has recently been shown that DSF in vitro potentiates vancomycin (VAN) susceptibility in vancomycin-resistant S. aureus (VRSA) strains [39]. It has also recently been shown that the combination of DDC and copper ions is active against S. aureus and Staphylococcus epidermidis (S. epidermidis) biofilms in vitro and in vivo [40]. Kobatake et al. recently showed that DSF had a bactericidal effect on Helicobacter pylori (H. pylori) at a low concentration (1 μg/mL) suggesting that it can be used as a supplement to current H. pylori eradication drugs. Moreover, the authors showed that DSF reduced the expression levels of proteins, such as urease, VacA (vacuolating cytotoxin) and CagA (cytotoxin-associated gene A) proteins that are the virulence proteins of H. pylori [41].
Thus, the position of DSF and its metabolites as antibiotics should not be contested. Unfortunately, most studies on the antibacterial properties of DSF and its metabolites focus on various Gram-positive and Gram-negative pathogens, with little attention to the genus Ureaplasma. In various bibliographic databases we were able to find only one paper on the effect of DSF on bacteria from the Mollicutes class which also includes the genus Ureaplasma. The results obtained by the authors showed that DSF exhibited copper-dependent antimicrobial activity against Mycoplasma hominis (M. hominis) [42].
In our previous research on bacteria from the Mollicutes class, we observed that lipoic acid (LA) had a statistically significant inhibitory effect on cell division of U. urealyticum and U.parvum cells in vitro compared to the control [43], while it did not affect the growth of M. hominis cells in vitro [44].
We recall these studies because LA, like DSF, is characterized by the presence of disulfide bonds. However, dihydrolipoic acid (DHLA), which is a reduced form of LA, is a thiol, like DDC. So, both LA and DSF are electrophiles that readily form disulfides with thiol-bearing substances. Bacteria contain many intracellular compounds containing thiophilic residues (for example cysteine (Cys), coenzyme A (CoA), glutathione (GSH), thioredoxin (Trx)) can be potentially modified by LA and DSF via thiol-disulfide exchange, which can lead to antimicrobial effects [39]. In addition, the reaction of intracellular thiols with DSF results in the generation of DDC that have metal (e.g. copper, iron, zinc)-chelating properties. As a result, DDC is able to inhibit the activity of metalloenzymes in the bacterial cells [45]. On the other hand, DDC or DHLA that are thiol-bearing substances can potentially modify compounds containing disulfide bonds present in the bacterial cell, for example oxidized glutathione (GSSG) or oxidized cystine (Cys-S-S-Cys) by thiol-disulfide exchange. „The Thiol World is an extremely interesting and challenging world from which many mysteries are still to be discovered” [46].The thiol-disulfide exchange is especially expected to occur with cellular GSH/GSSG. This causes a decrease in GSH levels in bacterial cells which can lead to antimicrobial effects. In studies on S.aureus, Long showed that the introduction of GSH in the growth medium caused an increase in the minimum inhibitory concentrations in μg/mL (MICs) of DSF and DDC [39]. In our opinion, the most likely mechanism by which DDC produces a bactericidal and bacteriostatic effect on U. urealyticum and U. parvum cells is related to the inhibition of urease (urea amidohydrolase, EC 3.5.1.5) activity. It is a nickel-containing enzyme that catalyzes the hydrolysis of urea to form carbonic acid and two molecules of ammonia. Urease activity has been detected in a large variety of bacteria but the ureaplasmas are the only organisms known to depend on urea for growth [47]. Ureaplasmas lack the conventional mechanisms for ATP generation, such as glycolysis or arginine breakdown, present in other mycoplasmas. It has been indicated that ATP is generated in these organisms through the formation of an ion gradient coupled to urea hydrolysis that is catalyzed by urease [48,49,50].. Thiol compounds are urease inhibitors, which was first demonstrated in 1980. The authors indicated then that ß-mercaptoethanol (ß-ME) acted as a competitive inhibitor of jack bean urease (JBU) [51]. Benini et al. demonstrated that ß-ME inhibited Bacillus pasteurii (B. pasteurii) urease by bridging the two nickel ions in the enzyme active site through the sulfur atom and chelation one Ni through the OH functionality. The authors also showed that ß-ME was involved in a mixed disulfide bond with Cysα322, a residue which plays a significant role in the urease-catalyzed process of urea hydrolysis [52]. There have been no studies on the effect of thiol compounds, including DDC on urease activity in Ureaplasma genus cells, so far. However, it is known that urease is very highly conserved in sequence and three-dimensional structure [53,54]. So, it can be assumed that the mechanism of the bacteriostatic and bactericidal effect of DDC against U. urealyticum and U. parvum, proven in the presented studies, is related to the inhibition of urease activity, which is an essential enzyme for these bacteria. In our opinion, it is a plausible proposal.
The problem we investigated certainly requires more research, but it is worth considering DDC as a drug in the treatment of patients with infections caused by Ureaplasma genus. An additional advantage of DDC is a lack of teratogenic, mutagenic or carcinogenic effects in animal models [55].
5. Conclusions
The clinical consequences of Ureaplasma infections can be serious and their treatment is often difficult. Thus, there is a need to search for new drugs effective against these atypical bacteria.
In this study, we showed for the first time the bacteriostatic (effects on the cell division) and bactericidal activity of DDC against U. urealyticum and U. parvum. Based on literature studies, we conclude that this DDC activity is related to the inhibition of urease activity, which is an essential enzyme for these bacteria. An additional advantage of DDC is a lack of teratogenic, mutagenic or carcinogenic effects in animal models.
In conclusion, DDC showed considerable antimicrobial activity against U. urealyticum and U. parvum in in vitro cultures. Further research is needed and the results obtained in the present study should be verified in in vivo models of infection.
Acknowledgments
This work was supported by statutory funds N41/DBS/001016 of the Faculty of Medicine, Jagiellonian University Medical College, Cracow, Poland.
Conflicts of Interest
The authors declare that there are no conflicts of interest.
References
- Deetjen:, P.; Maurer, C.; Rank, A.; Berlis, A.; Schubert, S.; Hoffmann, R. Brain abscess caused by Ureaplasma urealyticum in an adult patient. J Clin Microbiol 2014, 52, 695–698. [Google Scholar] [CrossRef]
- Vittecoq, O.; Schaeverbeke, T.; Favre, S.; Daragon, A.; Biga, N.; Cambon-Michot, C.; Bébéar, C.; Le Loët, X. Molecular diagnosis of Ureaplasma urealyticum in an immunocompetent patient with destructive reactive polyarthritis. Arthritis Rheum 1997, 40, 2084–2089. [Google Scholar] [CrossRef]
- Shepard, M.C. The recovery of pleuropneumonia-like organisms from Negro men with and without nongonococcal urethritis. Am J Syph Gonorrhea Vener Dis 1954, 38, 113–124. [Google Scholar]
- Crăcea, E.; Constantinescu, S.; Lazar, M. Serotypes of Ureaplasma urealyticum isolated from patients with nongonococcal urethritis and gonorrhea and from asymptomatic urethral carriers. Sex Transm Dis. 1985, 12, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Shepard, M.C.; Lunceford, C. D. Serological typing of Ureaplasma urealyticum isolates from urethritis patients by an agar growth inhibition method. J Clin Microbiol 1978, 8, 566–574. [Google Scholar] [CrossRef]
- Maeda, S.; Deguchi, T.; Ishiko, H.; Matsumoto, T.; Naito, S.; Kumon, H.; Tsukamoto, T.; Onodera, S.; Kamidono, S. Detection of Mycoplasma genitalium, Mycoplasma hominis, Ureaplasma parvum (biovar 1) and Ureaplasma urealyticum (biovar 2) in patients with non-gonococcal urethritis using polymerase chain reaction-microtiter plate hybridization. Int J Urol 2004, 11, 750–754. [Google Scholar] [CrossRef]
- Povlsen, K.; Bjørnelius, E.; Lidbrink, P.; Lind, I. Relationship of Ureaplasma urealyticum biovar 2 to nongonococcal urethritis. Eur J Clin Microbiol Infect Dis 2002, 21, 97–101. [Google Scholar] [CrossRef]
- Deguchi, T.; Yoshida, T.; Miyazawa, T.; Yasuda, M.; Tamaki, M.; Ishiko, H.; Maeda, S. Association of Ureaplasma urealyticum (biovar 2) with nongonococcal urethritis. Sex Transm Dis 2004, 31, 192–195. [Google Scholar] [CrossRef]
- Weidner, W.; Brunner, H.; Krause, W. Quantitative culture of ureaplasma urealyticum in patients with chronic prostatitis or prostatosis. J Urol 1980, 124, 622–625. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, J.; Lee, K.A. Prevalence of sexually transmitted infections among healthy Korean women: implications of multiplex PCR pathogen detection on antibiotic therapy. J Infect Chemother 2014, 20, 74–76. [Google Scholar] [CrossRef]
- Kafetzis, D.A.; Skevaki, C.; Skouteri, V.; Gavrili, S.; Peppa, K.; Kostalos, C.; Petrochilou, V.; Michalas, S. Maternal genital colonization with Ureaplasma urealyticum promotes preterm delivery. Association of the respiratory colonization of the premature infants with chronic lung disease and increased mortality. Clin Infect Dis 2004, 39, 1113–1122. [Google Scholar] [CrossRef]
- Sanchez, P.J.; Regan, J.A. Vertical transmission of Ureaplasma urealyticum from mothers to preterm infants. Pediatr Infect Dis J 1990, 9, 398–401. [Google Scholar] [CrossRef]
- Katz, B.; Patel, P.; Duffy, L.; Schelonka, R.L.; Dimmitt, R.A.; Waites, K.B. Characterization of ureaplasmas isolated from preterm infants with and without bronchopulmonary dysplasia. J Clin Microbiol 2005, 43, 4852–4854. [Google Scholar] [CrossRef]
- Heggie, A.D.; Bar-Shain, D.; Boxerbaum, B.; Fanaroff, A.A.; O'Riordan, M.A. , Robertson, J.A. Identification and quantification of ureaplasmas colonizing the respiratory tract and assessment of their role in the development of chronic lung disease in preterm infants. Pediatr Infect Dis J 2001, 20, 854–859. [Google Scholar] [CrossRef] [PubMed]
- Abele-Horn, M.; Wolff, C.; Dressel, P.; Pfaff, F.; Zimmermann, A. Association of Ureaplasma urealyticum biovars with clinical outcome for neonates, obstetric patients, and gynecological patients with pelvic inflammatory disease. J Clin Microbiol 1997, 35, 1199–1202. [Google Scholar] [CrossRef]
- Pinna, G.S.; Skevaki, C.L.; Kafetzis, D.A. The significance of Ureaplasma urealyticum as a pathogenic agent in the paediatric population. Curr Opin Infect Dis 2006, 19, 283–289. [Google Scholar] [CrossRef]
- Tantengco, O.A.G.; de Castro Silva, M.; Velayo, C.L. The role of genital mycoplasma infection in female infertility: A systematic review and meta-analysis. Am J Reprod Immunol 2021, 85, e13390. [Google Scholar] [CrossRef]
- Busolo, F.; Zanchetta, R. The effect of Mycoplasma hominis and Ureaplasma urealyticum on hamster egg in vitro penetration by human spermatozoa. Fertil Steril 1985, 43, 110–114. [Google Scholar] [CrossRef]
- Abdulrazzak, A.A.; Bakr, S.S. Role of mycoplasma in male infertility. East Mediterr Health J 2000, 6, 149–155. [Google Scholar] [CrossRef]
- Beeton, M.L.; Spiller, O.B. Antibiotic resistance among Ureaplasma spp. isolates: cause for concern? J Antimicrob Chemother 2017, 72, 330–337. [Google Scholar] [CrossRef]
- Khosropour, C.M.; Manhart, L.E.; Gillespie, C.W.; Lowens, M.S.; Golden, M.R.; Jensen, N.L.; Kenny, G.E.; Totten, P.A. Efficacy of standard therapies against Ureaplasma species and persistence among men with non-gonococcal urethritis enrolled in a randomised controlled trial. Sex Transm Infect 2015, 91, 308–313. [Google Scholar] [CrossRef]
- Biernat-Sudolska, M.; Rojek-Zakrzewska, D.; Drzewiecki, A.; Lauterbach, R. Antimicrobial susceptibility of Ureaplasma urealyticum and ureaplasma parvum isolated from premature infants with respiratory disorders. Przegl Epidemiol 2007, 61, 371–376, Polish. [Google Scholar]
- Ekinci, E.; Rohondia, S.; Khan, R.; Dou, Q.P. Repurposing Disulfiram as An Anti-Cancer Agent: Updated Review on Literature and Patents. Recent Pat Anti-Cancer Drug Discov 2019, 14, 113–132. [Google Scholar] [CrossRef]
- Iciek, M.; Bilska-Wilkosz, A.; Kozdrowicki, M.; Górny, M. Reactive Sulfur Compounds in the Fight against COVID-19. Antioxidants (Basel) 2022, 11, 1053. [Google Scholar] [CrossRef]
- Lajarin-Reinares, M.; Pena-Rodríguez, E.; Cañellas-Santos, M.; Rosell-Vives, E.; Cortés, P.; Casas, M.L.; Calvo, M.À.; Fernandez-Campos, F. Repurposing Disulfiram as an Antimicrobial Agent in Topical Infections. Antibiotics (Basel) 2022, 11, 1752. [Google Scholar] [CrossRef]
- Shirley, D.-A.; Sharma, I.; Warren, C.A.; Moonah, S. Drug Repurposing of the Alcohol Abuse Medication Disulfiram as an Anti-Parasitic Agent. Front Cell Infect Microbiol 2021, 11, 165. [Google Scholar] [CrossRef]
- Khan, S.; Singhal, S.; Mathur, T.; Upadhyay, D.J.; Rattan, A. Antifungal potential of disulfiram. Nihon Ishinkin Gakkai Zasshi 2007, 48, 109–113. [Google Scholar] [CrossRef]
- Biernat-Sudolska, M.; Rojek-Zakrzewska, D.; Lauterbach, R. Assesment of various diagnostic methods of ureaplasma respiratory tract infections in newborns. Acta Biochim Pol 2006, 53, 609–611. [Google Scholar] [CrossRef]
- Poveda, J.B.; Nicholas, R. Serological identification of mycoplasmas by growth and metabolic inhibition tests. Methods Mol Biol 1998, 104, 105–111. [Google Scholar] [CrossRef]
- Agarwal, R.P.; McPherson, R.A.; Phillips, M. Rapid degradation of disulfiram by serum albumin. Res Commun Chem Pathol Pharmacol 1983, 42, 293–310. [Google Scholar]
- Cobby, J.; Mayersohn, M.; Selliah, S. The rapid reduction of disulfiram in blood and plasma. J Pharmacol Exp Ther 1977, 202, 724–731. [Google Scholar] [PubMed]
- Marshall, M.J.; Harris, A.M.; Horne, J.E. The bacteriological and clinical assessment of a new preparation for the treatment of otitis extema in dogs and cats. J Small Anim Pract 1974, 15, 401–410. [Google Scholar] [CrossRef]
- Scheibel, L.W.; Adler, A.; Trager, W. Tetraethylthiuram disulfide (Antabuse) inhibits the human malaria parasite Plasmodium falciparum. Proc Natl Acad Sci USA 1979, 76, 5303–5307. [Google Scholar] [CrossRef]
- Harrison, J.J.; Turner, R.J.; Ceri, H. A subpopulation of Candida albicans and Candida tropicalis biofilm cells are highly tolerant to chelating agents. FEMS Microbiol Lett 2007, 272, 172–181. [Google Scholar] [CrossRef]
- Khouri, R.; Novais, F.; Santana, G.; de Oliveira, C.I.; Vannier dos Santos, M.A.; Barral, A.; Barral-Netto, M.; Van Weyenbergh, J. DETC induces Leishmania parasite killing in human in vitro and murine in vivo models: a promising therapeutic alternative in Leishmaniasis. PLoS One 2010, 5, e14394. [Google Scholar] [CrossRef]
- Celes, F.S.; Trovatti, E.; Khouri, R.; Van Weyenbergh, J.; Ribeiro, S.J.; Borges, V.M.; Barud, H.S.; de Oliveira, C.I. DETC-based bacterial cellulose biocuratives for topical treatment of cutaneous leishmaniasis. Sci Rep 2016, 6, 38330. [Google Scholar] [CrossRef]
- Taylor, E.H.; Walker, E.M. Jr.; Bartelt, M.; Day, S.; Pappas, A.A. In-vitro antimicrobial activity of diethyldithiocarbamate and dimethyldithiocarbamate against methicillin-resistant staphylococcus. Ann Clin Lab Sci 1987, 17, 171–177. [Google Scholar] [PubMed]
- Phillips, M.; Malloy, G.; Nedunchezian, D.; Lukrec, A.; Howard, R.G. Disulfiram inhibits the in vitro growth of methicillin-resistant staphylococcus aureus. Antimicrob Agents Chemother 1991, 35, 785–787. [Google Scholar] [CrossRef]
- Long, T.E. Repurposing Thiram and Disulfiram as Antibacterial Agents for Multidrug-Resistant Staphylococcus aureus Infections. Antimicrob Agents Chemother 2017, 61, e00898–17. [Google Scholar] [CrossRef]
- Kaul, L.; Abdo, A.I.; Coenye, T.; Krom, B.P.; Hoogenkamp, M.A.; Zannettino, A.C.W.; Süss, R.; Richter, K. The combination of diethyldithiocarbamate and copper ions is active against Staphylococcus aureus and Staphylococcus epidermidis biofilms in vitro and in vivo. Front Microbiol 2022, 13, 999893. [Google Scholar] [CrossRef]
- Kobatake, T.; Ogino, K.; Sakae, H.; Gotoh, K.; Watanabe, A.; Matsushita, O.; Okada, H.; Yokota, K. Antibacterial Effects of Disulfiram in Helicobacter pylori. Infect Drug Resist 2021, 14, 1757–1764. [Google Scholar] [CrossRef]
- Totten, A.H.; Crawford, C.L.; Dalecki, A.G.; Xiao, L.; Wolschendorf, F.; Atkinson, T.P. Differential Susceptibility of Mycoplasma and Ureaplasma Species to Compound-Enhanced Copper Toxicity. Front Microbiol 2019, 10, 1720. [Google Scholar] [CrossRef]
- Biernat-Sudolska, M.; Rojek-Zakrzewska, D.; Bilska-Wilkosz, A. In-vitro activity of lipoic acid against Ureaplasma urealyticum and Ureaplasma parvum isolated from women with infections of the urogenital tract. A pilot study. Acta Biochim Pol 2020, 67, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Biernat-Sudolska, M.; Rojek-Zakrzewska, D.; Gajda, P.; Bilska-Wilkosz, A. Lipoic Acid Does Not Affect The Growth of Mycoplasma hominis Cells In Vitro. Pol J Microbiol 2021, 70, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Lewis, A.D.; Riedel, T.M.; Kesler, M.B.A.; Varney, M.E.; Long, T.E. Pharmacological evaluation of disulfiram analogs as antimicrobial agents and their application as inhibitors of fosB-mediated fosfomycin resistance. J Antibiot (Tokyo) 2022, 75, 146–154. [Google Scholar] [CrossRef]
- Messens, J.; Collet, J.F. Thiol-disulfide exchange in signaling: disulfide bonds as a switch. Antioxid Redox Signal 2013, 18, 1594–1596. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, A.; Razin, S.; Kenny. G.E.; Barile, M.F. Characteristics of Ureaplasma urealyticum urease. J Bacteriol 1988, 170, 2692–2697. [Google Scholar] [CrossRef]
- Masover, G. K.; Hayflick, L. Growth of T-strain mycoplasmas in media without added urea. Ann N Y Acad Sci 1973, 225, 118–130. [Google Scholar] [CrossRef]
- Romano, N.; La Licata, R.; Russo Alesi, D. Energy production in Ureaplasma urealyticum. Pediatr Infect Dis 1986, 5, S308–S312. [Google Scholar] [CrossRef]
- Smith, D.G.; Russell, W.C.; Ingledew, W.J.; Thirkell, D. Hydrolysis of urea by Ureaplasma urealyticum generates a transmembrane potential with resultant ATP synthesis. J Bacteriol 1993, 175, 3253–3258. [Google Scholar] [CrossRef]
- Dixon, N.E.; Blakeley, R.L.; Zerner, B. Jack bean urease (EC 3.5.1.5). III. The involvement of active-site nickel ion in inhibition by beta-mercaptoethanol, phosphoramidate, and fluoride. Can J Biochem 1980, 58, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Benini, S.; Rypniewski, W. R.; Wilson, K. S.; Ciurli, S.; Mangani, S. The complex of Bacillus pasteurii urease with β-mercaptoethanol from X-ray data at 1.65-Å resolution. J Biol Inorg Chem 1998, 3, 268–273. [Google Scholar] [CrossRef]
- Kappaun, K.; Piovesan, A.R.; Carlini, C.R.; Ligabue-Braun, R. Ureases: Historical aspects, catalytic, and non-catalytic properties - A review. J Adv Res 2018, 2018 13, 3–17. [Google Scholar] [CrossRef]
- Zhang, L.; Mulrooney, S.B.; Leung, A.F.; Zeng., Y.; Ko, B.B.; Hausinger, R.P.; Sun, H. Inhibition of urease by bismuth(III): implications for the mechanism of action of bismuth drugs. Biometals 2006, 19, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Kaul, L.; Abdo, A.I.; Coenye, T.; Krom, B.P.; Hoogenkamp, M.A.; Zannettino, A.C.W.; Süss, R.; Richter, K. The combination of diethyldithiocarbamate and copper ions is active against Staphylococcus aureus and Staphylococcus epidermidis biofilms in vitro and in vivo. Front Microbiol 2022, 13, 999893. [Google Scholar] [CrossRef]
Figure 1.
The bacteriostatic effect of DDC on the cells of Ureaplasma spp. The graph shows the dependence of log titre for U. parvum (Up) and U. urealyticum (Uu) on DDC dose. Data are shown as the median value (red rectangle) with upper and lower quartyl. The whiskers represent the minimum and maximum values. Groups connected by a line are statistically significant at p < 0.05.
Figure 1.
The bacteriostatic effect of DDC on the cells of Ureaplasma spp. The graph shows the dependence of log titre for U. parvum (Up) and U. urealyticum (Uu) on DDC dose. Data are shown as the median value (red rectangle) with upper and lower quartyl. The whiskers represent the minimum and maximum values. Groups connected by a line are statistically significant at p < 0.05.
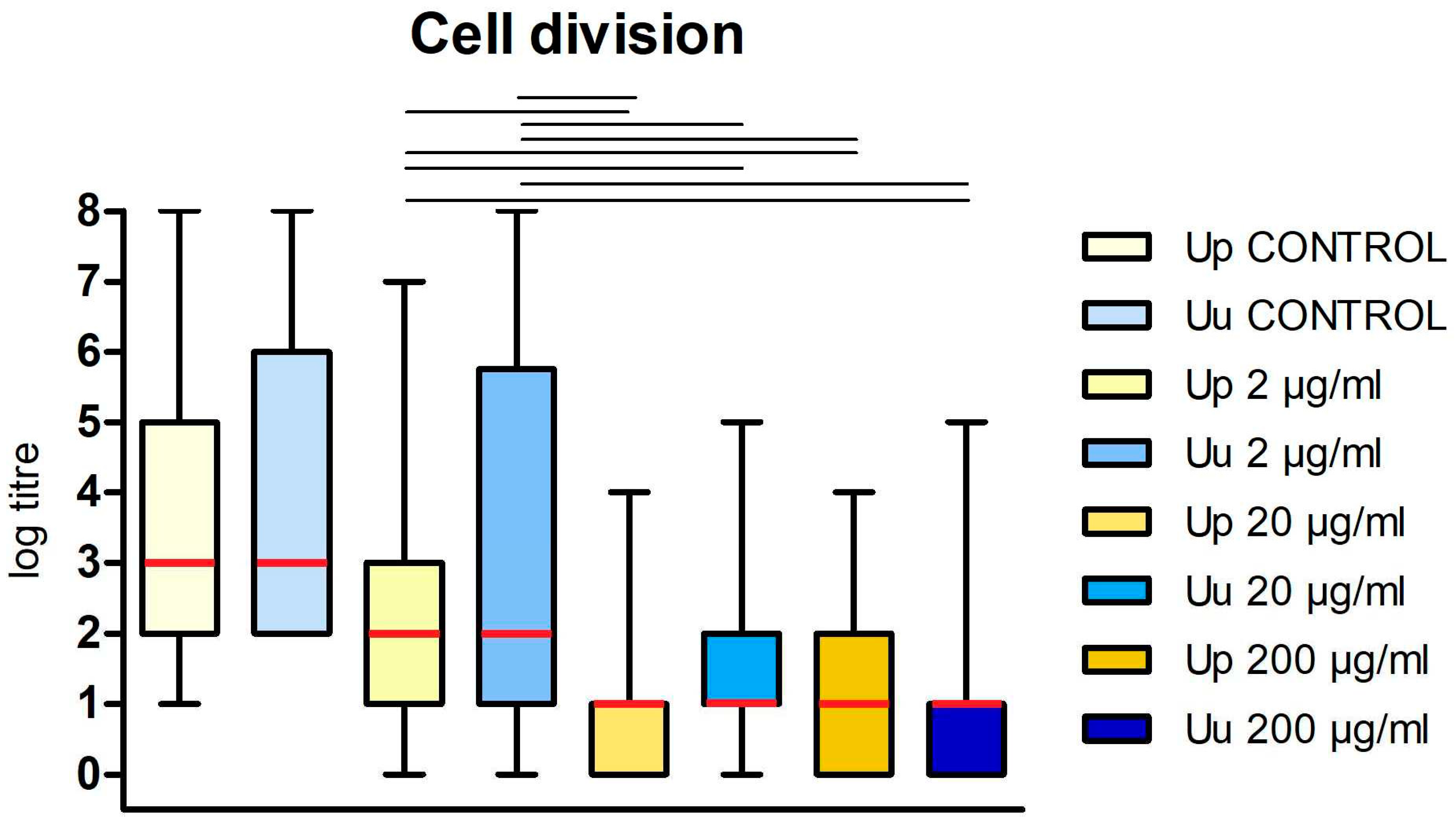
Figure 2.
Percentage of U. parvum (Up) and U. urealyticum (Uu) strains for which inhibition of cell division (bacteriostatic activity) was found at different DDC concentrations.
Figure 2.
Percentage of U. parvum (Up) and U. urealyticum (Uu) strains for which inhibition of cell division (bacteriostatic activity) was found at different DDC concentrations.
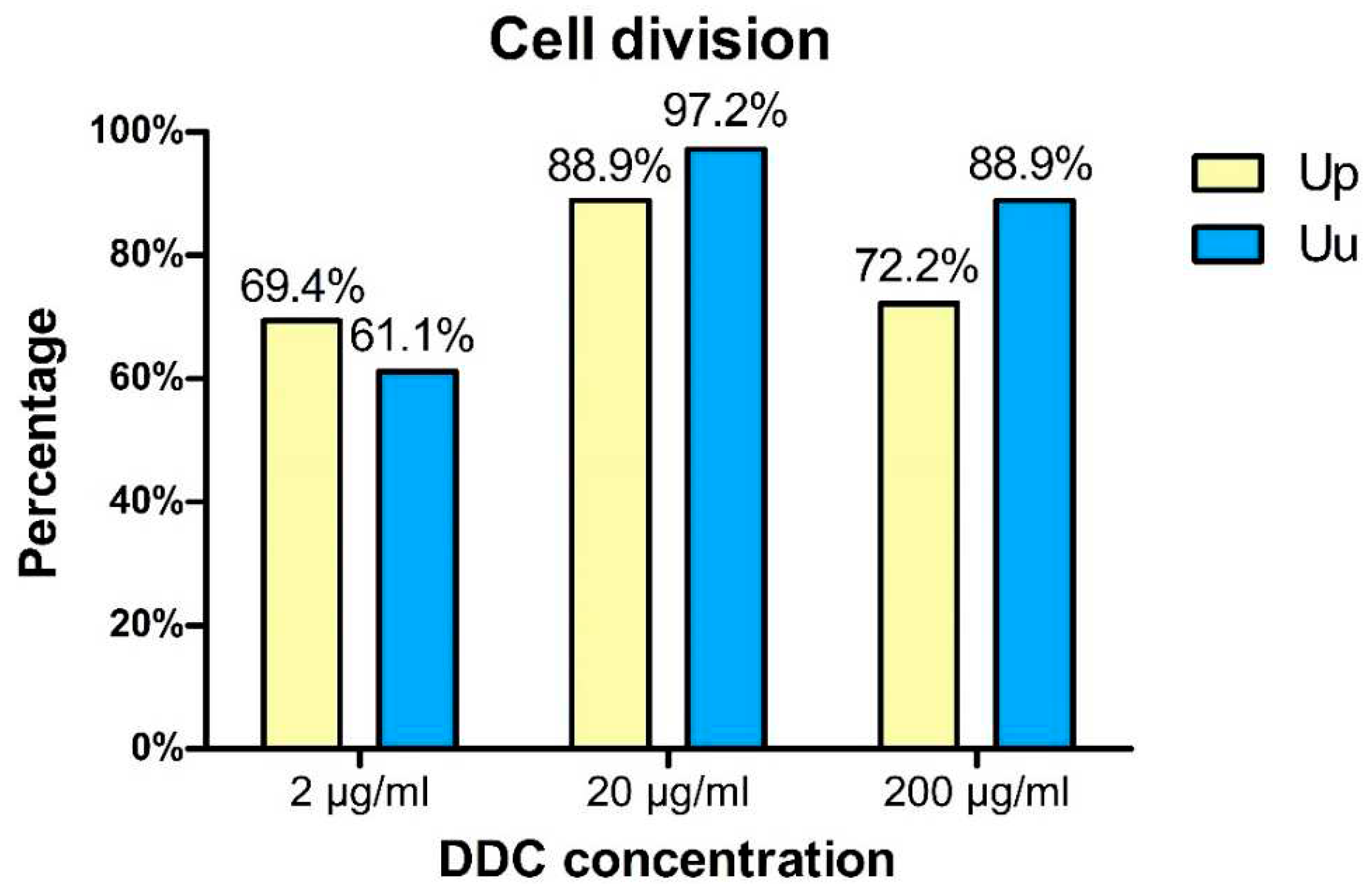
Figure 3.
The bactercidial effect of DDC on the cells of Ureaplasma spp. The graph shows the dependence of log titre for U. parvum (Up) and U. urealyticum (Uu) on DDC dose. Data are shown as the median value (red rectangle) with upper and lower quartyl. The whiskers represent the minimum and maximum values. Groups connected by a line are statistically significantly different at p < 0.05 level.
Figure 3.
The bactercidial effect of DDC on the cells of Ureaplasma spp. The graph shows the dependence of log titre for U. parvum (Up) and U. urealyticum (Uu) on DDC dose. Data are shown as the median value (red rectangle) with upper and lower quartyl. The whiskers represent the minimum and maximum values. Groups connected by a line are statistically significantly different at p < 0.05 level.
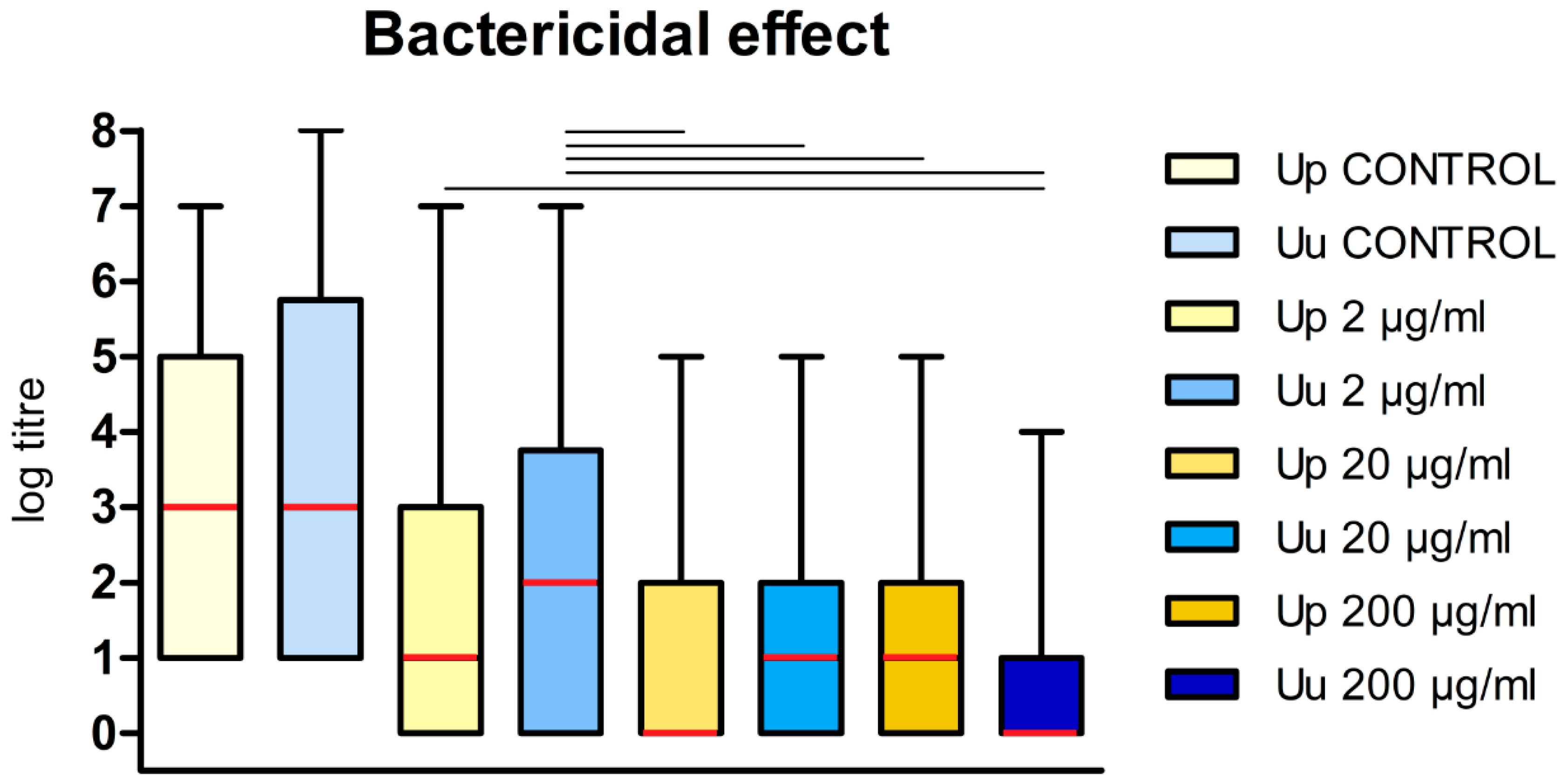
Figure 4.
Percentage of U. parvum (Up) and U. urealyticum (Uu) strains for which bactericidal effects of DDC were demonstrated.
Figure 4.
Percentage of U. parvum (Up) and U. urealyticum (Uu) strains for which bactericidal effects of DDC were demonstrated.
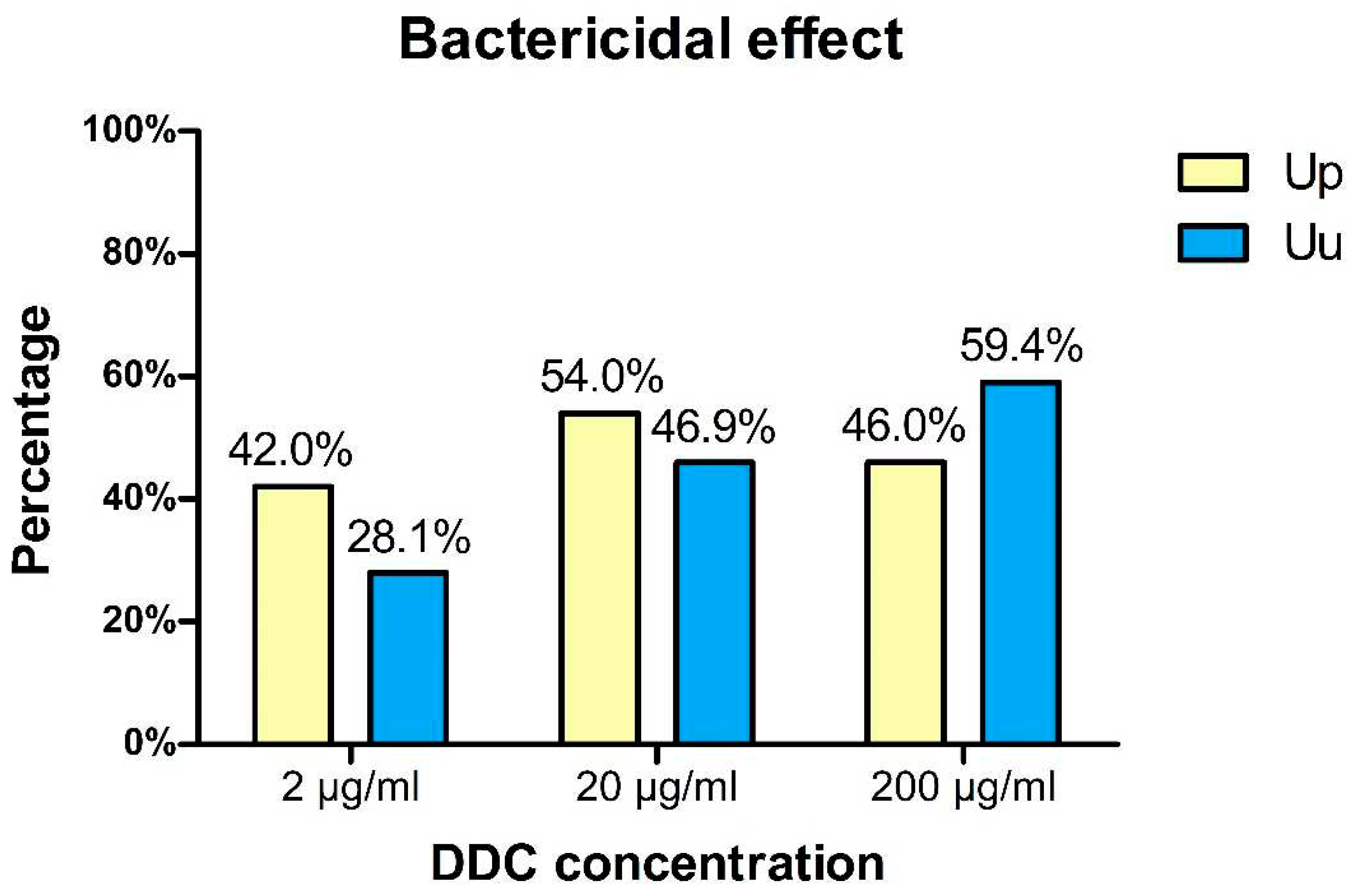
Figure 5.
Chemical structures of disulfiram and diethyldithiocarbamate.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



