
Submitted:
15 November 2023
Posted:
16 November 2023
You are already at the latest version
Abstract
Toxoplasma gondii, a protozoan parasite with the ability to infect various warm-blooded vertebrates, including humans, is the causative agent of toxoplasmosis. This infection poses significant risks, leading to severe complications in immunocompromised individuals and potentially affecting the fetus through congenital transmission. A comprehensive understanding of the intricate molecular interactions between T. gondii and its host is pivotal for the development of effective therapeutic strategies. This review emphasizes the crucial role of proteomics in T. gondii research, with a specific focus on host-parasite interactions, post-translational modifications (PTMs), PTM crosstalk, and ongoing efforts in drug discovery. Additionally, we provide an overview of recent advancements in proteomics techniques, encompassing interactome sample preparation methods such as BioID, APEX, and Y2H, as well as various proteomics approaches, including single-cell analysis, DIA, targeted, top-down, and plasma proteomics. Furthermore, we discuss the integration of proteomics with other omics technologies, highlighting its potential in unraveling the intricate mechanisms of T. gondii pathogenesis and identifying novel therapeutic targets.
Keywords:
Toxoplasma gondii
; proteomics
; host-parasite interactions
; PTMs
; drug discovery
1. Introduction
T. gondii is the causative agent of toxoplasmosis, one of the most common parasitic infections worldwide. The World Health Organization (WHO) estimates that about one-third of the world’s population has been infected with T. gondii. Among them, over 800,000 persons have been exposed and an estimated 750 deaths are caused by toxoplasmosis in the United States each year [1].
T. gondii engages in both sexual and asexual reproduction, utilizing felines as definitive hosts and other animals as intermediate hosts. Intermediate hosts become infected through consuming oocysts or tissues containing T. gondii cysts. Within the host, the parasite progresses through developmental stages—tachyzoites, bradyzoites, and sporozoites—facilitating survival and dissemination.
T. gondii employs a range of mechanisms to invade, replicate, manipulate host metabolism, and establish virulence. It invades host cells, through a process called gliding motility, which involves the coordinated action of proteins like myosin A, actin filaments, gliding-associated proteins (GAP), microneme proteins, rhoptry neck proteins (RONs), apical membrane antigen 1 (AMA1), and rhoptry proteins (ROPs). These proteins work together to facilitate attachment to host cells, form a moving junction, and ultimately, the invasion and egress of host cells [2]. T. gondii also exhibits varying levels of virulence, with different strains having distinct characteristics. One of the virulence factors is ROP18, which is critical for the parasite’s ability to evade the host’s immune response [3,4]. Virulent strains can cause severe disease in immunocompromised individuals, while less virulent strains may result in milder symptoms or even asymptomatic infections in healthy hosts. Once inside host cells, T. gondii replicates quickly and divides within a host cell to form daughter parasites, which are then released to infect new cells. By manipulating the host’s metabolic pathways, T. gondii can downregulate the host’s metabolism of fatty acids, lipids, and energy, often through hijacking host-signaling pathways such as peroxisome proliferator-activated receptors (PPAR) [5]. Additionally, the parasite can influence the host’s xenobiotic metabolism, affecting the breakdown of various substances. Understanding these metabolic alterations is crucial for developing targeted therapies.
Clinical outcomes of T. gondii infection vary based on the host’s immunity. Immunocompetent individuals typically experience mild symptoms or remain asymptomatic, while immunocompromised cases, like those with HIV/AIDS or organ transplants, face severe complications—encephalitis, retinochoroiditis, and organ damage. Congenital infection can affect fetuses. Existing clinical treatments are limited in scope so far.
Mass spectrometry (MS)-based Proteomics, which involves the large-scale study of proteins and their functions, has become a cornerstone of biological and medical research [6,7,8,9,10,11]. Through investigating the dynamic changes in protein expression, PTMs, and protein-protein interactions (PPIs) that occur during T. gondii infection, host immune responses and disease progression, scientists have acquired valuable insights into its biology, host interactions, virulence factors, and potential therapeutic targets [12,13,14,15,16,17,18].
After many years of progress, proteomics has advanced to the point where it can identify and measure around 10,000 proteins from human tissues simultaneously in an experiment [19]. It has played a crucial role in identifying potential biomarkers for diseases caused by parasites and therapeutic targets for treatments. This review aims to spotlight proteomic methodologies applied within these areas and summarize the important findings about key proteins (Table 1) engaged in T. gondii invasion and the interactions between the host and the parasite. To enhance the understanding of T. gondii biology, we also carried out an in-depth analysis of the proteomes, including stage-specific, PTMs, PTM crosstalk, and subcellular proteome data from the ToxoDB database.
2. Proteomics Approaches in T. gondii Research
Proteomics has evolved significantly over the past few decades (Figure 1), with the advancements in technology and methodology, playing a crucial role in our understanding of the parasite’s biology, host-parasite interactions, and potential drug targets. Here is an overview of the development of proteomics techniques in T. gondii research.
2.1. Early Studies (late 1990s to early 2000s)
Proteomics studies on T. gondii began in the late 1990s and early 2000s, primarily using two-dimensional gel electrophoresis (2DGE) combined with matrix-assisted laser desorption/ionization (MALDI) and electrospray ionization (ESI) to separate and analyze proteins. These early studies helped identify and characterize some of the parasite’s proteins [20,21], enabling researchers to analyze thousands of proteins from a biological sample. However, they had limitations concerning sensitivity and the ability to analyze low-abundance proteins.
Shotgun proteomics, also known as bottom-up proteomics, gained popularity in the early 2000s. It involved digesting proteins into peptides and then analyzing those peptides using MS. Several databases, such as UniProt, and searching algorithms, like SEQUEST and Mascot, were established to annotate and search protein sequences and information. These resources have become essential for proteomics research. Liquid chromatography-tandem MS (LC-MS/MS) techniques began to gain prominence in T. gondii research due to their improved accuracy in protein identification and quantification [22]. During this stage, researchers could identify a few hundred to a few thousand proteins in a single experiment using techniques like LC-MS/MS coupled with 2DGE. However, this process often required multiple injections of LC-MS/MS, contributing to the overall cost and time investment associated with the research endeavor.
2.2. Mid-2000s to the Present
Several key developments and technologies have been established and applied to T. gondii research, including:
2.2.1. Comparative Proteomics
Comparative proteomics involves comparing the proteomes of T. gondii and its infected host cells/tissues under different conditions to identify differentially expressed proteins and reveal molecular changes associated with infection. This approach allows researchers to identify and quantify parasite-specific proteins, host factors targeted by the parasite, and alterations in host protein expression induced by T. gondii infection [23,24,25,26,27,28,29]. During this stage, more than 6000 proteins can be identified and quantified using iTRAQ-labeling and strong cation exchange (SCX) fractionation techniques [16].
2.2.2. Subcellular Proteomics
Subcellular proteomics focuses on studying the proteome of specific cellular compartments or organelles involved in host-parasite interactions. By isolating and analyzing proteins from particular subcellular fractions, researchers can identify organelle-specific proteins and elucidate the role of specific compartments in T. gondii infection and host response [30,31,32].
2.2.3. Time-resolved Proteomics
Time-resolved proteomics approach involves collecting samples at different time points during a biological process, infection, or treatment, and then analyzing the proteomic changes that occur over those time intervals. It is a powerful tool for deciphering temporal changes in disease progression [33], cellular responses [34], biological processes [35], and drug action [36]. These applications in T. gondii provide valuable knowledge about the dynamics of complex systems and molecular mechanisms.
2.2.4. Post-translational Modifications (PTMs)
PTMs such as phosphorylation [37], glycosylation [38], and acetylation [39,40], play crucial roles in regulating various cellular processes in apicomplexan parasites, including T. gondii. These modifications affect protein activity, localization, interactions, and contribute to the complexity of the proteome, thereby likely influencing developmental transitions, biology, and pathogenesis of these parasites.
2.3. Recent Advancements in Proteomics Techniques
The advancements in MS have significantly improved its sensitivity, resolution, and throughput. High-resolution MS instruments like orbitrap and Q-TOF systems provide enhanced accuracy and dynamic range, enabling the identification and quantification of a wider range of proteins and PTMs. The recently developed Orbitrap/Astral dual analyzer mass spectrometer can handle five separate ion packets in parallel, leading to acquiring rates of up to 200 Hz [41]. This innovation substantially increases MS/MS scan speed, resulting in faster throughput, broader coverage, and higher sensitivity, along with accurate and precise quantitation in proteomics analysis. Here are some current hot topics in proteomics research, either currently in progress or poised for upcoming applications in T. gondii research.
2.3.1. Single-cell Proteomics
With the advance of MS, single-cell proteomics is experiencing rapid technological advancements and taking center stage [42]. These improvements include increased proteome coverage, which has grown from analyzing around 200 protein groups to over 1000 protein groups within individual mammalian cells [43]. Tandem Mass Tag (TMT) is typically ideal for studies requiring high multiplexing, compatibility with various sample types, and higher throughput. Junho et al. optimized the real-time search MS3-based TMT quantification method for the 126 single cells study; about 1000 out of the 1200 identified proteins were quantifiable [44]. In the 2023 ASMS meeting, Bruker highlighted the timsTOF Ultra’s capacity for single-cell proteomics, showing that the instrument can identify roughly 5,000 proteins at the single-cell level and quantify more than 4,800 proteins (https://www.nsmedicaldevices.com/news/bruker-introduces-timstof-ultra-mass-spectrometer-and-vistascan/#).
2.3.2. Data-independent Acquisition (DIA) and High-throughput Proteomics
DIA represents an emerging MS-based approach that effectively addresses the limitations associated with traditional data-dependent acquisition (DDA) methods. Through systematically fragmenting all discernible ions within a defined mass range, DIA offers several notable advantages, including expanded coverage of the proteome, enhanced precision in quantification, and the ability to detect proteins and PTMs at low abundance levels. Srinivasan et al. compared DIA with DDA methods for phosphoproteomics from nocodazole-treated and untreated U2OS cells. They found 15,548 unique site-localized phosphopeptides using DDA and 6,817 using DIA. While DDA excels in identifying more unique analytes, DIA demonstrated better reproducibility, with approximately 66% of localized peptides consistently identified in at least 5 out of 10 replicates compared to 32% in DDA [45]. It anticipates that novel data analysis methods, such as machine learning, will leverage advanced MS instrumentation to enhance the capabilities of DIA-MS, enabling more comprehensive and accurate measurements of PTMs [46]. Furthermore, it supports high-throughput proteomics applications and allows for identifying, quantifying, and characterizing thousands of proteins within a single experiment. Combining offline high-pH reversed-phase peptide fractionation with short online LC gradients (180 samples per day) and DIA-MS, enables near-complete coverage of the human proteome in just 4.5 hours of LC-MS/MS analysis [47].
2.3.3. Targeted Proteomics
Targeted proteomics, which includes techniques like selected reaction monitoring (SRM) and parallel reaction monitoring (PRM), is an active and rapidly advancing field in proteomics research. With advances in high-resolution mass spectrometers, targeted proteomics is gaining prominence in clinical and translational research [48]. It plays a crucial role in biomarker discovery and validation for various diseases. Researchers are developing targeted assays for specific proteins and PTMs relevant to disease. Nguyen et al. demonstrated the successful development of a workflow for the quantification of low abundance proteins involved in the UPR pathway, ranging from 4 to 103 amol of the LOQs for the targeted proteins were determined [49]. DIA approaches are also applied for the simultaneous quantification of multiple peptides within a predefined mass range in complex samples. Multiplexed targeted proteomics assays, such as TMT, are integrated with targeted proteomics for quantification [50]. Targeted proteomics is also applied in food safety and environmental monitoring, where it is used to quantify specific proteins and peptides relevant to these fields [51].
2.3.4. Plasma Proteome
While fractionation and depletion techniques have made notable progress in improving coverage of the plasma proteome, the considerable 10 orders of magnitude dynamic range present in analyzed plasma samples remains a substantial challenge. Using an innovative sample-enrichment kit called Mag-Net, a magnetic bead-centered procedure developed by Wu and colleagues [52], this study quantified more than 5000 plasma proteins from an extracellular vesicle-enriched sample in a single one-hour LC-MS/MS run, representing a significant advancement in the field [53].
2.3.5. Top-down Proteomics
Top-down proteomics is a powerful approach for studying intact proteins, including the characterization of protein isoforms and PTMs. With advancements in High-resolution mass spectrometers, particularly those equipped with Orbitrap or FT-ICR and data analysis tools, such as ProSight and TopPIC, top-down proteomics is well-suited for the characterization of complex PTMs, including glycosylation, phosphorylation, and acetylation, at the protein level. Top-down proteomics also plays a crucial role in structural proteomics, which enhances our understanding of protein folding, conformational changes, and the study of PPIs at the intact protein level. There is growing interest in applying top-down proteomics to clinical research and diagnostics since it has the potential to identify disease-specific protein isoforms, truncated forms, and PTMs for biomarker discovery. Recently, Melby JA et al. highlighted the effectiveness of the top-down proteomics method in characterizing protein isoforms (>200 kDa) diversity within single muscle cells and suggested its broader utility in understanding cellular heterogeneity and advancing precision medicine efforts [54]. It is also possible to integrate the results of hydrogen-deuterium exchange MS (HDX-MS) with top-down proteomics to gain a more comprehensive understanding of a protein’s structure, function, and dynamics. For example, HDX-MS can help us understand better how a protein’s conformational changes upon ligand binding, and these changes can be further analyzed using top-down proteomics to identify and characterize the modified protein forms.
2.3.6. Multi-omics Integration
Multi-omics integration has also been applied to investigate host-pathogen interactions and infectious diseases. Jean Beltran et al. discussed proteomic methods and their application in studying host-pathogen interactions, highlighting how multi-omics approaches, including proteomics, genomics, transcriptomics, and metabolomics, contribute to a systems-level understanding of infectious diseases. [55]. It has paved the way for the development of potential therapies and vaccines against toxoplasmosis and continues to be an active area of research. Leo et al. employed proteomics, transcriptomics, and pharmacoproteomics to analyze childhood acute lymphoblastic leukemia cell lines and their responses to oncology drugs, identifying correlations between molecular phenotypes and drug responses [56].
In T. gondii research, multi-omics integration has played a pivotal role. For instance, Kloehn et al. utilized multi-omics analysis to explore the distinct functions of subcellular acetyl-CoA pools in T. gondii. This approach unveiled a deeper understanding of the parasite’s physiological and metabolic adjustments [57]. Similarly, the study by Nie et al. employed global proteomic profiling and transcriptomics to understand the proteome-transcriptome correlation during T. gondii infection in cats. Their work shed light on the complex proteotranscriptomic reprogramming that mediates the dynamic interplays between T. gondii and feline tissues [58].
Machine learning approaches have been employed for multi-omics data analysis to uncover a deeper understanding of biological systems. Reel et al. reviewed different integrative machine-learning methods used to analyze multi-omics data, which can aid in disease prediction, patient stratification, and precision medicine [59]. Analyzing these diverse data sources simultaneously can enable researchers to identify complex interactions and relationships that may not be apparent when analyzing each data type separately.
3. Interactome Analysis
Interactome analysis aims to map and characterize PPIs between T. gondii proteins and host proteins, which can identify key protein complexes and pathways involved in host-parasite interactions [60,61,62]. Xia J et al. investigated the human interactome of the T. gondii rhoptry protein ROP18 (TgROP18) and identified the targets of ROP18I and ROP18II, highlighting their association with immune response and apoptosis [3]. Matthew et al. conducted a proteomic analysis of host cells infected with different types of parasites expressing MAF1b, MAF1a, and a non-functional mutant of MAF1b. Among the 13 proteins uniquely enriched in the MAF1b pull-down experiments, two host proteins—TOM70 and HSPA9—were identified as essential mediators of the Host Mitochondrial Association (HMA) phenomenon driven by MAF1b [63].
Below are several common sample preparation techniques and approaches employed for studying PPIs.
3.1. Affinity Purification
In this approach, T. gondii proteins are tagged with a specific affinity tag (such as FLAG or HA tags) and expressed in the parasite. The tagged proteins and their interacting partners can then be selectively isolated using affinity purification techniques, followed by identification using MS. Yakubu et al. utilized affinity purification combined with proteomics to reveal that Protein Arginine Methyltransferase 1 (PRMT1) significantly contributes to arginine monomethylation in T. gondii [28]. Anghel et al. applied differential affinity chromatography followed by MS to investigate cellular and molecular targets of nucleotide-tagged ruthenium complexes in T. gondii and Trypanosoma brucei [64]. Müller et al. employed differential affinity chromatography coupled with MS to identify common binding proteins for an antimicrobial peptide in T. gondii [65]. Li et al. conducted a global proteome analysis of lysine crotonylation (Kcr) in T. gondii using LC-MS/MS and an immune-affinity method. They identified thousands of Kcr sites on diverse proteins, suggesting a widespread involvement of Kcr in various biological processes [66]. Sun et al. demonstrated the significance of immunoproteomics and MALDI-TOF MS in identifying antigenic proteins of T. gondii RH strain that 18 immunoreactive proteins had been identified and recognized by human immunoglobulin G (IgG). Notably, these proteins did not show reactivity with negative-control sera from healthy individuals without Toxoplasma infection [67].
3.2. Proximity Labeling Techniques
By labeling molecules that are close to a specific protein or molecular complex of interest, the labeled proteins can be isolated and identified using MS. Proximity labeling techniques offer a high-throughput approach to studying various biomolecular interactions with spatial and temporal precision. Shkel et al. utilized enzymes to covalently label nearby biomolecules, enabling their identification using MS. Their advantage lies in capturing weak or transient interactions, making them crucial for investigating organelle interactomes and macromolecular complexes [68]. Back et al. used Proximity labeling to identify novel IMC proteins enriched in daughter buds and revealed that IMC29 plays an important role in Toxoplasma endodyogeny [69]. Cygan et al. employed proximity labeling to identify proteins at the host-cytosolic side of the PVM. The study identified specific parasites (GRA61, GRA62, GRA63) and host proteins (PDCD6IP/ALIX, PDCD6, CC2D1A, MOSPD2) in PVM [70].
3.3. BirA*-mediated Proximity-dependent Biotin Identification (BioID)
BioID is a specific proximity labeling technique that uses a mutant form of the biotin ligase enzyme (BirA*) as the labeling enzyme. The protein of interest is fused to BirA*, and when the fusion protein is expressed in cells, BirA* biotinylates lysine residues on proteins within its spatial proximity. These biotinylated proteins can be isolated using streptavidin-based purification and then identified using MS [71]. Engelberg et al. used BioID technology and identified four architecturally distinct basal complexBC subcomplexes, and key proteins including BCC0, BCC4, and MORN1, highlighting their roles in different aspects of cell division [71]. Proteins associated with coiled-coil structures and involved in signaling functions have been detected in the apical annuli and are linked to inner membrane complex (IMC) sutures [72]. Nadipuram et al. applied BioID technology to uncover novel dense granule proteins secreted by T. gondii bradyzoites and identify previously unknown proteins involved in chronic infection and cyst formation [73]. Tu et al. employed BioID to identify clusters of proteins associated with dense granules, cyst matrix, and the cyst wall, enhancing the understanding of cyst wall composition [74]. Song et al. utilized BioID to identify more than 300 proximal interacting proteins of calmodulin (CaM) in T. gondii. CaM was found to play essential roles in tachyzoite proliferation, invasion, and egress, with potential functions related to ion binding and oxidation-reduction [75].
3.4. Ascorbate Peroxidase-mediated Proximity Labeling (APEX)
One notable benefit of APEX compared to traditional BioID is its considerably faster labeling rate, operating within minutes as opposed to hours. When combined with quantitative proteomic methods, this helps detect the rapid alterations in protein interactions over time or as a reaction to cellular disturbances [76,77]. With BioID and APEX techniques, Pan M et al. identified 46 proteins, including 20 known and 26 new GRAs. These GRAs, mainly in coccidian parasites, might not be vital for in vitro growth but could play roles in animal infections [78].
3.5. Two-hybrid (Y2H)
Yeast two-hybrid (Y2H) assays are useful to identify PPIs in cells. Two proteins, one "bait" and one "prey," are put into yeast cells. If there is an interaction, it is proven by the activation of reporter genes. Using Y2H assays, Lai et al. discovered significant interactions between T. gondii surface antigens (SAG2 and SAG1) and specific human proteins. They identified 20 and 39 positive clones interacting with SAG2 and SAG1, respectively. Notably, Homo sapiens zinc finger protein strongly interacted with SAG2, while Homo sapiens lysine-rich coil-coiled protein showed a strong interaction with SAG1 [79,80].
Interactome analysis using the abovementioned techniques has significantly contributed to our understanding of host-parasite interaction networks, virulence factors, and intracellular signaling pathways, and facilitated the identification of potential therapeutic targets for combating toxoplasmosis.
4. ToxoDB in T. gondii Proteome
The ToxoDB database (https://toxodb.org/toxo/app/) is a valuable resource for the proteome study of T. gondii, allowing researchers to access experimental data related to T. gondii proteins, which include MS data, protein expression profiles, and post-translational modification information [81]. According to the current release, the ToxoDB-65_TgondiiME49_AnnotatedProteins database, which contains 8322 protein-coding genes, can be used for proteomics database searching. Researchers can also cross-reference proteomic data with other omics data to gain a more comprehensive understanding of T. gondii biology. There are 96, 1521, and 104 records about T. gondii research for MS, PTMs, and quantitative MS respectively in ToxoDB (Figure 2A). We also list the search results by inputting the keywords of different biological processes such as invasion, virulence, replication, metabolism, and gliding (Figure 2B). Comparing identified proteins and expression levels with those available in ToxoDB, researchers can understand when and where the proteins of interest are expressed in different stages of the parasite’s life cycle. The visualization tools and bioinformatics resources on ToxoDB can help identify integrated data patterns, correlations, and potential functional perceptions. For example, ROP18, an essential virulence factor, plays a pivotal role in the parasite’s capacity to initiate infection and induce illness in the host. This protein possesses serine/threonine kinase activity, enabling it to phosphorylate host cell proteins. Consequently, ROP18 can manipulate host cell functions and immune responses. ToxoDB contains 36 records related to ROP18 in the context of MS, 5 records concerning PTMs, and 4 records for Quantitative MS in studies involving T. gondii. Researchers can utilize these records to construct protein/peptide libraries or seamlessly integrate them into other omics projects. With more data imputed into the database, ToxoDB will play many more roles in the integration of proteomics with genomics data, providing various bioinformatics tools and resources to assist researchers in data analysis.
4.1. Stage-specific Proteomes
The first T. gondii proteome was generated from the tachyzoite stage [82]. In that work, three proteomic platforms were designed: two-dimensional electrophoresis, one-dimensional electrophoresis gel coupled with LC-MS/MS, and MudPIT analysis. Combining all of them the authors identified 2,252 non-redundant expressed proteins, approximately 29% of the annotated genes. An interesting aspect is that 57% of the genes exhibiting evidence of transcription through the Expressed Sequence Tag method, failed to yield detectable peptides. This observation raises the possibility of low translation levels, protein instability, transient protein expression, or even the existence of non-coding RNAs. In contrast, when comparing the proteins identified in this study with mRNA expression levels determined by microarray analysis, it was observed that a substantial number of proteins were detected even in cases where mRNA expression levels were negligible or minimal, with 204 and 632 instances, respectively. This could suggest highly stable proteins over time. Finally, peptide identification helped to define or confirm the exon-intron predictions of the annotated genes. The incorporation of proteome data from several laboratories over time, with different objectives and strategies, allowed to raise the number of proteins identified in the tachyzoite stage to 5,325 [83]. Another proteomic study in EuPathDB identified ∼35% of the total protein-coding genes and ∼50% of the total proteins for ME49 [84]. Interestingly, this analysis added 289 loci by RNA-seq and proteomics that were not previously identified.
In 2013, the proteome of partially sporulated oocysts was published [83]. The generation of the oocyst stage proteins was of high relevance since there were very few studies carried out on this stage, and the lack of identification of specific stage proteins hindered progress on this important stage for environmental dissemination of T. gondii. The analysis compared the oocyst proteome with transcriptomic data obtained in unsporulated (day 0) and sporulated (day 10) oocysts. At day 0, the analysis could identify proteins from upregulated genes associated with metabolism, cellular transport, and cell fate, while on day 10 the proteome was enriched in proteins associated with protein synthesis, cell rescue, defense, and virulence. On the other hand, the comparison of this proteome with that of tachyzoites allowed the identification of 154 oocyst-specific proteins. A large number of these proteins is associated with hypothetical genes. In the case of oocyst-specific functional genes, they tend to be enriched in metabolic functions, comprising the largest category. This is followed by proteins involved in processes related to cell rescue, defense, virulence, energy production, and protein fate. Compared to tachyzoites, oocysts have a greater capability of de novo amino acid biosynthesis and are well equipped to fuel the glycolysis/gluconeogenesis and tricarboxylic cycle (TCA), including the expression of Eno1 (enolase 1 gene) in the unsporulated oocyst, enolase also expressed in bradyzoite.
The generation of a bradyzoite stage proteome was more challenging. In the in vitro differentiation process, only less than 80% of T. gondii tachyzoites differentiate into bradyzoites. For this reason, Garfoot et al. [85] conducted experimental infections in CBA/J mice using the Me49 strain. In the in vivo model, by contrast, 100% of T. gondii differentiated into bradyzoites were obtained, however, their extraction was done from brains (brain bradyzoite) of infected mice at 21-90 days post-infection (dpi), requiring a process of purification in dextran to obtain larger amounts of peptide reads. In total, 1,683 T. gondii proteins were mapped in the bradyzoite brain proteome, and a total of 893 proteins if duplicates were taken into account. The number of identified proteins common to all time points analyzed was only 366. Part of the sample was used to generate a transcriptome. Most of the genes that encode the 366 proteins have high levels of expression. However, 100 of the 366 proteins presented a low level of gene expression during chronic infection. Again, that could be due to short-lived transcripts but long-lived proteins. An interesting fact from the RNA-seq analysis is that the transcriptomic profile remains constant in the period from 28 to 120 days of the analysis.
4.2. PTM Proteomes
The proteomes of tachyzoites, bradyzoites, and oocysts can offer information about the genes of interest that potentially provide prior knowledge about their expression levels in the different stages. However, proteomes that investigate various PTMs can provide valuable insights for researchers interested in the study of a specific gene. There are more than 400 protein PTMs, which in turn present crosstalk with each other. In T. gondii, information from a PTM could help formulate a hypothesis regarding the function and/or regulation of the protein of interest. Knowing which residues have incorporated specific PTMs provides essential information for generating site-specific mutants. In ToxoDB there is data obtained from three PTM proteomes: phosphorylation, acetylation, and ubiquitinylation. Beyond the scope of ToxoDB, other more specific proteomes analyze PTMs in various contexts, including histone PTMs [86], acetylation in T. gondii deficient in acetyl-coA [57], crotonylation [66], phosphorylation in kinase mutants or different stages [37,87,88,89,90,91,92], lipidation [93,94], Cysteine S-nitrosylation [95]. In this section, we will only mention what was obtained in the proteomes of PTMs in the ToxoDB.
4.2.1. Phosphoproteome
In 2011, Treeck et al. [96] generated the first phosphoproteome of T. gondii and Plasmodium falciparum. The objective was to obtain information about which proteins could regulate their activity through this PTM. Although it is common to both parasites, T. gondii possesses several kinases, including protein kinase A, protein kinase G, rhoptries, calcium-dependent kinases, etc., which play important roles in various biological processes, such as invasion, egress, DNA and parasite replication, pathogenesis, differentiation, and energy metabolism [97,98,99,100,101,102]. The T. gondii phosphoproteome was generated based on two sources: intracellular tachyzoites and tachyzoites purified from cell cultures in which secreted parasite proteins were discarded [96]. In total, 2,793 (intracellular) and 3,506 (purified) phosphorylated proteins were identified, detecting 12,793 and 24,298 phosphorylated sites, respectively. By the comparison of the intracellular and purified parasites, about 50 proteins were detected that are phosphorylated within the host cell, in turn, suggesting that most of the exportable proteins (with signal peptide) are phosphorylated in the tachyzoite.
4.2.2. Acetylome
Acetylation is a PTM that was initially studied in histones and its role in chromatin modulation. However, it was observed that acetylation regulates the activity and function of many non-histone cytoplasmic and nuclear proteins [103]. The acetylation mainly happens on the lysine residues, and this PTM can be reversible (Nε-acetylation and O-acetylation) or irreversible (Nα-acetylation). Other residues including serine, threonine, and histidine also can undergo acetylation [104,105]. In T. gondii, it has been shown that the reversible acetylation process, mediated by lysine acetyltransferases (KAT) and lysine deacetyl transferases (KDAC), can be excellent therapeutic targets [106]. Among the KDAC inhibitors, also called histone deacetyl transferases, identified are apicidin, FR235222, hydroxamate-based compounds, panobinostat, JF363, tubastatin, SAHA, and MC1742 [107,108,109,110,111,112,113]. Additionally, a KDAC activator, resveratrol, has been described [114].
The T. gondii acetylome identified 411 lysine acetylation sites across 274 tachyzoite proteins [115], data present in the ToxoDB. The vast majority of acetylated proteins are associated with translation, energy metabolism, chromatin-associated proteins, and stress response. The study detected 25 proteins associated with cellular metabolism in addition to five histones. In the acetylation process, the donor of the acetyl group is acetyl-CoA. In a subsequent study, the acetylome of a T. gondii double mutant was analyzed. The double mutant where the ATP citrate lyase gene was deleted (ΔACL) together with an inducible knockdown (KD) of the acetyl-CoA synthetase [57] is inviable when KD is induced. The acetylome of these tachyzoites shows hypoacetylation of proteins involved in metabolism and histones, among others.
An acetylome of three T. gondii strains was also generated for comparative analysis [116]: the virulent strains RH, PYS (Chinese 1), and the avirulent PRU. Based on the analysis of the two replicates for each case, 457 acetylated proteins for RH and 188 acetylated proteins for PYS were identified. In the case of the RH strain, the top 6 of the KEGG pathways were metabolic pathways, biosynthesis of secondary metabolites, biosynthesis of antibiotics, microbial metabolism in diverse environments, ribosome and carbon metabolism. In the PYS strain, the top six of the enriched KEGG pathways were metabolic pathways, biosynthesis of secondary metabolites, biosynthesis of antibiotics, carbon metabolism, microbial metabolism in diverse environments, and the ribosome. It is noticed that both acetylomes are very similar. The PRU acetylome yielded 134 acetylated proteins, with a KEGG pathway profile very similar to that of RH and PYS. However, acetylated proteins from RH and PYS were more enriched in the pyruvate metabolism pathway compared to acetylated proteins from the PRU strain. Between RH and PYS, 26 differentially acetylated proteins (DAP) were observed: 2 upregulated and 24 downregulated in PYS, including histone-acetyl-transferase and glycyl-tRNA synthase. Both enzymes play roles in stress tolerance and proliferation, key features in parasite virulence.
4.2.3. Ubiquitin Proteome
Ubiquitinylation is a process where one or several molecules are conjugated to a protein. Ubiquitin is made up of 76 amino acids and has a molecular mass of approximately 8.6 kDa. Its structure is highly conserved in the eukaryotic lineage. If the binding to the target protein occurs primarily through lysine 48 of ubiquitin, with lysines 6 and 33 playing a minor role, this protein will undergo degradation. On the other hand, binding by lysine 63 of the ubiquitin leads the target protein to different biological roles such as DNA repair, chromatin silencing, signaling activation, protein trafficking, and receptor endocytosis [117]. The ubiquitinoma of intracellular tachyzoites of T. gondii showed 454 ubiquitinylated proteins and 800 sites [118]. Ubiquitinylated proteins correspond to different subcellular locations and functions, but this PTM was mainly detected in proteins of the cytoskeleton, inner membrane complex (IMC), and the nucleus, including histones. One interesting aspect of the study was the regulation of ubiquitinylation in T. gondii proteins throughout the cell cycle. The ubiquitinoma of the extracellular tachyzoite comprised 346 proteins, of which 51 were typical of the extracellular stage, among them glycolytic enzymes, matching the metabolic differences between the extracellular and intracellular stages [119].
4.3. PTM Crosstalk and Proteome
A study on combined methylation, phosphorylation, and acetylation of proteins done in a lung cancer model showed a large number of proteins that presented the three PTMs [120,121]. Taking a functional perspective, PTMs increase the diversity of functional units of protein origin within a cell. PTMs are still a field with great potential for future exploration in T. gondii. A crosstalk study with ubiquitination and the other PTMs showed that less than 10% of the proteins overlapped between ubiquitin and arginine methylation, 21% combined acetylation with ubiquitin, 25% of the SUMO proteome was ubiquitinated, while 78% of the phosphorylated proteins are also ubiquitinated [118]. Among the proteins detected that present combinations of different PTMs, the histone acetyltransferase EP300 and the chaperone Hsp90 [120] stand out.
4.3.1. Histones
Over several decades of research, it has become evident that various PTMs on histone proteins engage in intricate crosstalk, ultimately influencing the regulation of gene expression. This intricate interplay of histone modifications has been characterized as a "histone code," which specific proteins can interpret, providing in conjunction with the substitution of different histone variants, a means for epigenetic regulation [122]. Understanding the roles of PTMs is also complicated by the fact that the same writers and readers of marks also write and read PTMs on other proteins [123].
Protozoan parasites, such as T. gondii, are no exception to this trend. The use of high-throughput mass spectrometry is enabling the systematic dissection of histone modifications and core markers involved in epigenetic regulation [124]. Numerous residues have been identified for modification, particularly within the histone tails. However, it is worth noting that various residues within the histone globular domains can also undergo diverse modifications, encompassing processes such as acetylation, phosphorylation, ubiquitination, methylation, and lately crotonylation and 2-hydroxyisobutyrylation [124]. These last newly described marks have demonstrated their crucial significance in regulating global transcription in mammalian cells. Consequently, in T. gondii, they may exert a noteworthy influence on the transcription process [124].
There is a high frequency of lysine residues in histones, and these residues are often modified by PTMs [86]. Among the histones, H3 and H4 PTMs are widely distributed not only in unicellular organisms, like protozoan parasites, but also in multicellular eukaryotes, and many of the amino acid residues that carry certain PTMs are highly conserved [123]. Figure 3A shows the different histone marks and their crosstalk in the mammal model as described. Furthermore, histone marks identified in T. gondii PTM proteomic analysis were highlighted to compare (Figure 3B). In the figure, it can also see the conservation of possible crosstalk similar to those observed in mammals in punctuated lines, although some new marks are also detected and the crosstalks are marked as questions. As it can be observed, almost all the marks are observed, and although not much research has been conducted in this area in the parasite, their crosstalk can be inferred. As shown in the figure, in T. gondii, the N-terminal tails of H4 and H3 are highly acetylated and methylated regions [86]. Interestingly, some of the residues that are acetylated can also be methylated, like lysine 12 (H4K12me) or lysine 16 when not acetylated, was found trimethylated (H4K16me3) [86]. Another example is H3K9, which was also found methylated and acetylated. While mono methylation and acetylation are active marks, di or tri methylation is often repressive and found in subtelomeres and centromeres [125]. Acetylation on H4K31 has also been reported in T. gondii and P. falciparum and this residue can also be mono-methylated in a mutually exclusive manner [115,126]. In T. gondii, this residue was found to be acetylated at the promoter of nearby active genes associated with H3K4me3 and H3K9ac marks (Figure 3B). At the same time, it was mono-methylated in the core body of the gene, and it inversely correlated with gene expression, it would be a repressive mark as opposed to H4K31ac [126]. Some PTMs not found in other species for this histone were found, like H4R23me, in the globular domain and methylation, both di and monomethylation on the C-terminal tail, on R78 [86].
Another important modification on lysine residues is ubiquitination, but few PTMs occur on the same lysine residues as ubiquitin. One exception is H3K24, which is modified by ubiquitin, acetylation, mono- and trimethylation, implicating it as an important regulatory site [118]. Apart from this residue, only H3K116 was found to be ubiquitinylated, although other modifications occur at substoichiometric levels only detectable using an enrichment strategy [118].
There are also few PTMs in H2A.X, but two were found in the C-terminal tail, acetylation of K128 that has not been described, and the phosphorylated S132 [86], which corresponds to S139p in humans with a conserved function for TgH2A.X in response to DNA damage [127]. By contrast, H2A.Z displays several PTMs, of which many acetylations in the N-terminal tail stand out. Within the first 40 amino acids, there are 10 lysines (K6, K10, K14, K18, K24, K27, K29, K34, K36, and K37), all of which are acetylated. Additionally, lysine K18 could also be methylated, leading to various PTM combinations with acetylated residues [86].
A similar situation arises for H2B histones, as canonical H2B possesses only one acetylation on the N-terminus at K4, whereas H2B.Z exhibits multiple modifications in the N-terminal tail, including acetylation on lysines 3, 8, 13, 14, and 18 [86]. Another study using H2BK120 ubiquitin-specific antibody detected reactivity with T. gondii H2B and H2B.Z supporting the presence of ubiquitin-histone conjugates in T. gondii chromatin that could have conserved functions [118]. Histone H2B.Z is unique to apicomplexan parasites and has been described to conform to a double variant nucleosome (DVN), hyperacetylated in the N-terminal tail and associated with transcriptional activation (Figure 3C) [39,125,127,128,129,130]. Recent research has examined acetylations in the H2B.Z N-terminal tail lysines and their significance in various biological processes in T. gondii [39]. This study revealed that the inability to regulate this N-tail positive charge patch generated when no acetylation is possible, produced a reduced in vitro replication, a heightened differentiation rate, an increased sensitivity to DNA damage, and in an in vivo model produced a complete loss of T. gondii virulence [39].
As mentioned above, not much research has been conducted to date on histone crosstalks in T. gondii. Nevertheless, some conclusions have been made, taking into account results when one mark can be sufficient to elicit a specific biological output while in other cases, multiple marks are required [131] (Figure 3B). For example, it has been proposed that gene activation needs the dual signature of H3R17me2 and H3K18 [132]. In this work, the T. gondii arginine methyltransferase TgCARM1 was shown to work in concert with the acetylase TgGCN5-A, whose substrate preference is H3K18. Another example is the redundancy or multiplicity of sumoylation sites in the parasite histones suggested by Bougdour et al. [131].
Also, the transcription-associated PTMs H3K4ac, H3K9ac, and H3K27ac were found to be associated with H4K31ac in euchromatic regions and opposed to the heterochromatic regions revealed by the repressive marks H3K9me3 and H4K20me3 [126] (Figure 3B). In another study, the DVN conformed by H2A.Z and H2B.Z, which are both hyperacetylated in the N-terminal tails, was found in association with the activatory mark H3K4me3 [129]. In turn, H3K4me3 was found along with H3K9ac in most of the T. gondii genes that are expressed (Figure 3C) [131].
Another well-characterized example first in yeast but then in other eukaryotes is the requirement of histone H2B monoubiquitination for proper H3K4 and H3K79 methylation [122,133,134]. Although ubiquitination was detected on TgH2AK119Ub and TgH2BK120Ub [118], to date it has only been speculated that acetylation and ubiquitination may regulate the differential localization of H2A.Z and H2B.Z on active and silent genes in T. gondii [125] and so it is represented as a question in Figure 3B.
4.3.2. Hsp90 PTM Crosstalk
The Hsp90 chaperone was shown to be simultaneously acetylated and phosphorylated in the lung cancer model [120]. This protein is central since its main functions are to assist important proteins in biological processes such as DNA replication, regulation of gene expression, proliferation, etc.; in both stressed and non-stressed cells. The Hsp90 protein network includes hundreds of proteins, mainly transcription factors, and kinases, but also proteins associated with chromatin, metabolism, translation, and DNA damage [135,136,137]. Hsp90 can form different complexes, one of them called the Hsp70/Hsp90 cycle. It may include other co-chaperones, the hsp40 type I being the one that initiates the recruitment of the client protein. The Hsp70/Hsp90 cycle is conserved in T. gondii [137]. Recently, the proteome of Hsp90 and Hsp40 type I in T. gondii was analyzed and referred to as Tgj1 [62]. This study allowed defining a putative PPI network for both chaperones as part of the Hsp70/Hsp90 cycle and in their independent functions between them. Within the Hsp70/Hsp90 cycle, the enriched pathways were translation, cell redox homeostasis, and protein folding. The PPI associated with Hsp90 not related to the Tgj1-Hsp90 axis showed mainly interactors related to protein folding, RNA processing, cell signaling, and transcription. This shows the important role of this chaperone in a wide diversity of biological processes of T. gondii. The proteomes related to PTMs show that T. gondii Hsp90 presents phosphorylations, acetylations, ubiquitinylation, and even a monomethyl arginine modification (ToxoDB_TGME49_288380). Backe et al. describe the role of the different PTMs in Hsp90 in the review [138]. Based on the role of the PTMs present in Hsp90 of other organisms, a proposed role for the T. gondii Hsp90 PTMs counterpart can be deduced [61]. Briefly, phosphorylation of Hsp90 in yeast and other species may affect its binding to certain client proteins or even its affinity for ATP. In other cases (e.g., Hsp90 T101), it may promote kinase client activation. Hsp90 phosphorylation in turn is associated with the cell cycle and in some cases promotes cell proliferation [139,140]. Therefore, these PTMs could similarly affect T. gondii Hsp90. Phosphorylation of threonine 5 and 7 on Hsp90 are associated with double-strand break repair [141], but these modifications were not detected in T. gondii. The phosphorylation of residues S231 (T220 in T. gondii) and S263 is related to telomerase activity [142]. Similarly, acetylation of Hsp90 leads to a loss of affinity for ATP and a decrease in its binding to the client protein and co-chaperones [138]. T. gondii presents two acetylations in the acetylome (K384 and K559). Acetylation of lysine K384 in human Hsp90 affects the interaction with its client protein, the receptor tyrosine kinase ErbB2 [143]. Acetylation at K558 (TgK559) could be associated with the export of Hsp90 to the extracellular space [144]. The ubiquitinylation of Hsp90 seems to be associated only with its degradation. CHIP is one of the co-chaperones associated with the Hsp70/Hsp90 cycle, which negatively regulates the pathway. This CHIP protein is an E3 ubiquitin ligase and binds to Hsp90. However, phosphorylated forms of Hsp90 decrease their association with CHIP, suggesting a crosstalk between both PTMs [145].
5. Subcellular Localization Associated Proteome
For researchers delving into an uncharted protein of interest, one of the key considerations for its characterization and the potential revelation of its function lies in determining its subcellular localization. This becomes even more crucial in omics studies, where a multitude of genes or proteins are involved, and establishing associations among them often requires them to inhabit the same cellular compartments. Beyond proteomes focused on subcellular locations, some proteomes analyze the presence of T. gondii proteins within specific cellular structures. For example, T. gondii conoid proteome was performed [146]. This is a structure relevant for invasion in T. gondii and nearby related parasites, but absent in other Apicomplexas such as Plasmodium spp. In this work, 200 proteins located in the cytoskeleton conoid and the apical region of T. gondii were identified. Another work that provides very interesting data is the proteome of extracellular proteins and vesicles (exosomes/ectosomes) [147]. In this work, 512 proteins were detected in the excretory/secretory, 210 in the ectosome, and 285 in the exosome fraction. The authors also analyzed the supernatant obtained from the purification of vesicles, identifying 421 proteins in that fraction. Another study, in which data has been uploaded to ToxoDB, detected 346 proteins in extracellular vesicles [148]. As expected, proteins from the secretory organelles micronemes, rhoptries, and dense granules were detected. However, kinases, heat shock proteins, and even nuclear proteins, including histones, were also identified, showing the wide diversity of proteins released by some pathways, which could have an implication not yet studied in toxoplasmic infection. Another particular proteomic study is the analysis of the proteins present in the wall of the T. gondii cyst [149]. In this study, numerous anticipated proteins such as CST1, BPK1, MCP4, MAG1, GRA2, GRA3, and GRA5 were identified, and new components were also discovered. In one case (CST2), its null deletion altered the virulence and lost the cyst formation. Based on the importance of rhoptry in the invasion process a proteomic analysis of rhoptries was performed [150]. The authors identified 38 novel rhoptry proteins.
Toxoplasma has single mitochondria that is very relevant during the tachyzoite stage. Researchers observed that bradyzoites lack a functional TCA and respiratory chain [151]. The mitochondrial proteome was generated based on two proximal labeling systems already described above; one by mBirA and another modified for mitochondria using the plant ascorbate peroxidase (APEX) system [152]. The proteome data were uploaded to ToxoDB. In total, researchers identified 421 proteins: 213 in the APEX samples and 369 in the mBirA system. Notably, 36% were hypothetical and 33% were never defined in a role or location associated with mitochondria. Although some of them presented another non-mitochondrial location by other techniques, in the majority their location was confirmed. Among the new proteins identified, the authors focused on one called TgApiCox25, which was shown to be a component of the cytochrome c oxidase complex.
In the ToxoDB, two T. gondii proteomes analyze the components through subcellular fractionation. One was generated by the Laboratory of Dr. Silvia Moreno (Center for Tropical and Emerging Global Diseases, Department of Cellular Biology, University of Georgia, Athens, GA, USA). Another is based on a fractionation called hyperlexed localization of organelle proteins by isotope tagging (hyperLOPIT). LOPIT is based on protein correlation profiling, in which all proteins from a subcellular location can be marked by tandem mass tags followed by mass spectrometry [153]. In the case of T. gondii HyperLOPIT, an iodixanol density gradient was used to separate the subcellular compartments [32]. Tagged peptides from the different fractions of the gradient were analyzed. A subcellular localization map was generated based on the following compartments: apical 1, apical 2, micronemes, rhoptries 1, rhoptries 2, dense granules, IMC, tubulin cytoskeleton, Plasma Membrane (PM) − peripheral 1, endomembrane/vesicles, PM − peripheral 2, PM – integral, Golgi, endoplasmic reticulum (ER) 1, ER 2, apicoplast, mitochondria – membrane, mitochondria – soluble, nucleus – chromatin, nucleus − non-chromatin, nucleolus, 40S ribosome, 60S ribosome, cytosol, 19S proteasome, 20S proteasome and unassigned. This level of detail shows the strength of this technique in identifying proteins from different organelles and subcellular compartments. In total, the authors were able to identify and quantify 3,832 proteins of which they assigned 2,634 proteins to distinct subcellular niches mentioned above. The study also addresses issues related to the evolution of Apicomplexa, allowing the identification of evolutionary aspects of the different compartments in comparison with other species. Among them, dense granules stand out as the most recent organelles. In other cases, accelerated evolution is observed, such as IMC or nuclear-chromatin cluster proteins, the latter potentially having an impact on the machinery for regulating gene expression. Most rapidly changing proteins are enriched in rhoptries, dense granules, and at the parasite surface, which could be associated with host-parasite coevolution.
6. Proteomics Approach in Drug Discovery Efforts
Current drugs for toxoplasmosis primarily target acute infection but are less effective against chronic toxoplasmosis. Identifying new drug targets is crucial to develop improved treatment options that can overcome the limitations of existing drugs. These targets should ideally address both acute and chronic stages of the infection, particularly within the brain and skeletal muscles [154].
Proteomics has provided crucial understandings of T. gondii’s biology, identifying potential drug targets, repurposing certain drugs, and contributing to the development of innovative therapeutics, diagnostics, and vaccines against this parasitic pathogen [6,154,155,156,157]. The identification of functional proteins within structures like micronemes and rhoptries holds promise for advancing diagnostics and vaccine development against toxoplasmosis [155,158]. Initially, drugs were discovered through whole organism screening, which involved testing compounds in in vitro and in vivo models. However, recent advances in genomics and proteomics have shifted the focus towards target-based drug design [159]. Integrating multi-omics data, including proteomics, genomics, and transcriptomics, is crucial for understanding parasite biology and identifying new drug and vaccine targets. Effective data management strategies are essential to harness the potential of "big data" in parasitology [160].
6.1. Drug Target Identification and Therapeutics
Currently, there are only a few drugs available to treat T. gondii infections and some of them have limitations such as toxicity and the potential for parasite resistance. By comparing protein expression profiles between different stages of the parasite’s life cycle and between parasite and host cells, researchers can pinpoint proteins that are specific to the parasite, making them potential targets for drug development. Proteomics has identified key drug targets, such as histone variant H2B.Z acetylation, essential for T. gondii’s fitness [39]. Innovative methods like CRISPR-based oligo recombineering (CORe) have been employed to identify chemically reactive sites for drug targeting [161]. Protein phosphatases are promising drug targets against Apicomplexan parasites [162]. N-myristoyl transferase (NMT) is considered a potential target [163], and the spider peptide XYP1 demonstrates anti-T.gondii effects by interacting with membrane-associated proteins [164]. Identification of unique membrane proteins in T. gondii suggests potential candidates for therapeutic development [165].
6.2. Insights into Protein Modification and Pathway
Understanding the complex interaction between the parasite and the host is crucial for drug development, as the host’s immune response can influence the efficacy of treatment. PTMs, such as crotonylation and 2-hydroxyisobutyrylation, affect critical enzymes in energy-related pathways. Histone modifications like acetylation play essential roles in gene regulation, and parasite fitness [39]. It is not limited to histone proteins but also extends to nonhistone proteins involved in diverse cellular functions. The presence of acetylation as a regulatory mechanism in protozoan parasites offers an opportunity to explore this machinery as a target for drug development [166]. Disrupting specific protein pathways, such as plastidic iron-sulfur cluster biogenesis, affects parasite viability and cellular functions [167]. It was found that blocking palmitoylation enhances the release of invasion-related proteins, including AMA1, from apical secretory organelles. This observation suggests that AMA1 plays a role in controlling the secretion process of these proteins [94]. That information is crucial for designing drugs that disrupt key pathways or processes in the parasite’s biology while minimizing harm to the host.
6.3. Advancements in Diagnostics and Vaccines
Proteomics contributes to diagnostic tests and vaccine development against T. gondii. Multiepitope antigens and computational pipelines enhance diagnostic and vaccine strategies. Identifying immunodominant epitopes and functional proteins through proteomics aids in developing comprehensive protection strategies against toxoplasmosis [155]. A significant homology was observed in the antigenic proteome profiles between N. caninum and T. gondii, which has important implications. It suggests the feasibility of designing multicomponent vaccines that target common antigens shared by both parasites [168]. Proteomics can also play a role in vaccine development by identifying potential protein antigens that can stimulate an immune response [169].
6.4. Screening and Testing Drug Candidates
Discovering new drug candidates with anti-toxoplasma activity is a significant challenge. Proteomics facilitates the screening of potential drug compounds by assessing their impact on the parasite’s proteome. Researchers can examine how candidate drugs alter protein expression patterns, helping to identify compounds that effectively inhibit the parasite’s growth or disrupt essential pathways. The investigation of essential proteins of T. gondii as potential drug targets has been ongoing for the past two decades. While these efforts have shown promise in vitro, translating these findings into effective treatments for toxoplasmosis in humans has been challenging. The utilization of proteomics-based investigations to examine the interactions between drug candidates and proteins from both the parasite and the host can serve as an effective approach for characterizing drug targets [157]. In another study, in silico screening of compounds was conducted based on the proteome of T. gondii. They selected thirteen compounds and evaluated them in vitro against the parasite using a cell-based assay. This approach identified several compounds, including almitrine, bortezomib, and fludarabine, with in vitro anti-T. gondii activity. Almitrine, in particular, demonstrated a high selectivity index and showed interactions with specific transporters, suggesting a potential mechanism of action [170].
6.5. Characterizing Drug Resistance
T. gondii has the potential to develop resistance to existing drugs, reducing their effectiveness. The presence of drug-resistant T. gondii strains poses challenges to treatment efficacy, especially in immunocompromised patients. This necessitates a better understanding of resistance mechanisms and the implementation of monitoring programs to address this public health issue [171]. Proteomics can shed light on mechanisms of drug resistance in T. gondii. By comparing protein and PTM profiles of drug-sensitive and drug-resistant strains, researchers can identify changes in protein expression that contribute to resistance, informing the design of new drugs or treatment strategies. Proteomic analysis revealed the down- or up-regulation of various proteins, with a particular focus on some key proteins, both actin and MIC8, that play important roles in invasion capability [172]. Doliwa et al. provided valuable proteomic insights into sulfadiazine resistance in T. gondii strains isolated from clinical cases, contributing to our understanding of the mechanisms underlying drug resistance in this parasite [29]. Drug resistance in T. gondii can also develop when parasite strains are exposed to increasing concentrations of antiparasitic drugs, such as artemisone and artemiside. The development of resistance is associated with alterations in the parasite’s proteome, leading to changes in protein expression, particularly those involved in reactive oxygen species (ROS) regulation. This resistance mechanism differs from that observed in other parasites like Plasmodium, highlighting the complexity of drug resistance in T. gondii [173].
In summary, recent advances in high-throughput proteomics techniques show potential for identifying parasite-specific markers, especially very low abundance proteins and proteins that have small differences in protein structure but have significant functional consequences in increasing drug specificity and reducing toxicity. Utilizing high-throughput proteomics techniques can significantly reduce the expenses associated with screening and validating drug targets. This cost reduction is particularly advantageous for neglected diseases with limited market potential, such as toxoplasmosis. With the applications of these cutting-edge proteomic technologies in T. gondii research, the resolution of these challenges in drug development appears to be on the horizon.
7. Future Perspectives
The future of proteomics lies in the accurate and precise measurement of thousands of proteins across thousands of samples. Ongoing progress in proteomic techniques, data analysis, and sample preparation offers exciting prospects for enhancing our understanding of intricate biological systems. Over the past few decades, researchers have encountered challenges in discovering biomarkers within plasma due to the broad dynamic range of proteins. The introduction of groundbreaking MS technologies and innovative sample preparation methods, as discussed earlier, is poised to facilitate the discovery of new biomarkers and enable precision medicine at scale. Furthermore, as 90% of the human proteome is mapped [174], future proteomics research, including T. gondii studies will emphasize integrating proteomics with other multi-omics approaches. Artificial intelligence and machine learning will play a critical role in the analysis and interpretation of “big data”. These advancements not only hold promise for identifying novel biomarkers but also offer valuable insights into the invasion mechanisms of T. gondii and potential therapeutic targets.
Author Contributions
Funding
The Vermont Biomedical Research Network Proteomics Facility (RRID: SCR_018667) is supported through NIH grant P20GM103449. This work is supported by grants from ANPCyT: PICT 2021-00331 and PICT 2021-0169; and from CONICET: PIP 0572.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All the raw data generated are available upon reasonable request to the corresponding authors.
Acknowledgments
SO Angel and L. Vanagas are members of Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) and Professor of Universidad Nacional General San Martin (UNSAM). B Deng is a research professor at the University of Vermont.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jones JL, K.-M.D., Wilson M, McQuillan G, Navin T, McAuley JB, Toxoplasma gondii infection in the United States: seroprevalence and risk factors.Am J Epidemiol. Am J Epidemiol, 2001(154): p. 357-365. [CrossRef]
- Frénal, K.; Dubremetz, J.-F.; Lebrun, M.; Soldati-Favre, D. Gliding motility powers invasion and egress in Apicomplexa. Nat. Rev. Microbiol. 2017, 15, 645–660. [Google Scholar] [CrossRef]
- Xia, J.; Kong, L.; Zhou, L.-J.; Wu, S.-Z.; Yao, L.-J.; He, C.; He, C.Y.; Peng, H.-J. Genome-Wide Bimolecular Fluorescence Complementation-Based Proteomic Analysis of Toxoplasma gondii ROP18’s Human Interactome Shows Its Key Role in Regulation of Cell Immunity and Apoptosis. Front. Immunol. 2018, 9, 61. [Google Scholar] [CrossRef]
- Yang, Z.; Hou, Y.; Hao, T.; Rho, H.-S.; Wan, J.; Luan, Y.; Gao, X.; Yao, J.; Pan, A.; Xie, Z.; et al. A Human Proteome Array Approach to Identifying Key Host Proteins Targeted by Toxoplasma Kinase ROP18. Mol. Cell. Proteom. 2017, 16, 469–484. [Google Scholar] [CrossRef] [PubMed]
- Gallego-Lopez, G.M., et al., Metabolic changes to host cells with Toxoplasma gondii infection. bioRxiv, 2023. [CrossRef]
- Katsarou, E.I., et al., Applied Proteomics in ‘One Health’. Proteomes, 2021. 9(3). [CrossRef]
- Bautista, J.M.; Marín-García, P.; Diez, A.; Azcárate, I.G.; Puyet, A. Malaria proteomics: Insights into the parasite–host interactions in the pathogenic space. J. Proteom. 2014, 97, 107–125. [Google Scholar] [CrossRef]
- Muselius, B.; Durand, S.-L.; Geddes-McAlister, J. Proteomics of Cryptococcus neoformans: From the Lab to the Clinic. Int. J. Mol. Sci. 2021, 22, 12390. [Google Scholar] [CrossRef]
- Christopher, J.A., et al., Subcellular proteomics. Nat Rev Methods Primers, 2021. 1. [CrossRef]
- Jiao, F.; Zhang, D.; Jiang, M.; Mi, J.; Liu, X.; Zhang, H.; Hu, Z.; Xu, X.; Hu, X. Label-free proteomic analysis of placental proteins during Toxoplasma gondii infection. J. Proteom. 2017, 150, 31–39. [Google Scholar] [CrossRef]
- Zhang, D.; Sun, X.; Ren, L.; Yang, C.; Liu, X.; Zhang, H.; Jiang, Y.; Hu, X. Proteomic profiling of human decidual immune proteins during Toxoplasma gondii infection. J. Proteom. 2018, 186, 28–37. [Google Scholar] [CrossRef]
- Weiss, L.M., et al., Toxoplasma gondii proteomics. Expert Rev Proteomics, 2009. 6(3): p. 303-13. [CrossRef]
- Yakubu, R.R., E. Nieves, and L.M. Weiss, The Methods Employed in Mass Spectrometric Analysis of Posttranslational Modifications (PTMs) and Protein-Protein Interactions (PPIs). Adv Exp Med Biol, 2019. 1140: p. 169-198. [CrossRef]
- Morlon-Guyot, J.; El Hajj, H.; Martin, K.; Fois, A.; Carrillo, A.; Berry, L.; Burchmore, R.; Meissner, M.; Lebrun, M.; Daher, W. A proteomic analysis unravels novel CORVET and HOPS proteins involved in Toxoplasma gondii secretory organelles biogenesis. Cell. Microbiol. 2018, 20, e12870. [Google Scholar] [CrossRef] [PubMed]
- McMurtrey, C., et al., Toxoplasma gondii peptide ligands open the gate of the HLA class I binding groove. Elife, 2016. 5. [CrossRef]
- Wang, Z.X., et al., Proteomic Differences between Developmental Stages of Toxoplasma gondii Revealed by iTRAQ-Based Quantitative Proteomics. Frontiers in Microbiology, 2017. 8. [CrossRef]
- Xie H, S.H., Dong H, Dai L, Xu H, Zhang L, Wang Q, Zhang J, Zhao G, Xu C, Yin K., Label-free quantitative proteomic analyses of mouse astrocytes provides insight into the host response mechanism at different developmental stages of Toxoplasma gondii. PLoS Negl Trop Dis. 2023. 2023 Sep 18;17(9). [CrossRef]
- Nelson, M.M.; Jones, A.R.; Carmen, J.C.; Sinai, A.P.; Burchmore, R.; Wastling, J.M. Modulation of the Host Cell Proteome by the Intracellular Apicomplexan Parasite Toxoplasma gondii. Infect. Immun. 2008, 76, 828–844. [Google Scholar] [CrossRef] [PubMed]
- Muntel, J.; Gandhi, T.; Verbeke, L.; Bernhardt, O.M.; Treiber, T.; Bruderer, R.; Reiter, L. Surpassing 10 000 identified and quantified proteins in a single run by optimizing current LC-MS instrumentation and data analysis strategy. Mol. Omics 2019, 15, 348–360. [Google Scholar] [CrossRef] [PubMed]
- Jungblut, P.R., et al., Proteomics in human disease: cancer, heart and infectious diseases. Electrophoresis, 1999. 20(10): p. 2100-10. [CrossRef]
- Beckers, C.J.; Roos, D.S.; Donald, R.G.; Luft, B.J.; Schwab, J.C.; Cao, Y.; A Joiner, K. Inhibition of cytoplasmic and organellar protein synthesis in Toxoplasma gondii. Implications for the target of macrolide antibiotics. J. Clin. Investig. 1995, 95, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.W.; Kafsack, B.F.C.; Cole, R.N.; Beckett, P.; Shen, R.F.; Carruthers, V.B. The Opportunistic Pathogen Toxoplasma gondii Deploys a Diverse Legion of Invasion and Survival Proteins. J. Biol. Chem. 2005, 280, 34233–34244. [Google Scholar] [CrossRef] [PubMed]
- Fauquenoy, S.; Morelle, W.; Hovasse, A.; Bednarczyk, A.; Slomianny, C.; Schaeffer, C.; Van Dorsselaer, A.; Tomavo, S. Proteomics and Glycomics Analyses of N-Glycosylated Structures Involved in Toxoplasma gondii-Host Cell Interactions. Mol. Cell. Proteom. 2008, 7, 891–910. [Google Scholar] [CrossRef] [PubMed]
- Al-Bajalan, M.M.M.; Xia, D.; Armstrong, S.; Randle, N.; Wastling, J.M. Toxoplasma gondii and Neospora caninum induce different host cell responses at proteome-wide phosphorylation events; a step forward for uncovering the biological differences between these closely related parasites. Parasitol. Res. 2017, 116, 2707–2719. [Google Scholar] [CrossRef]
- Zhou, C.-X.; Gao, M.; Han, B.; Cong, H.; Zhu, X.-Q.; Zhou, H.-Y. Quantitative Peptidomics of Mouse Brain After Infection With Cyst-Forming Toxoplasma gondii. Front. Immunol. 2021, 12, 681242. [Google Scholar] [CrossRef]
- Sun, H.; Li, J.; Wang, L.; Yin, K.; Xu, C.; Liu, G.; Xiao, T.; Huang, B.; Wei, Q.; Gong, M.; et al. Comparative Proteomics Analysis for Elucidating the Interaction Between Host Cells and Toxoplasma gondii. Front. Cell. Infect. Microbiol. 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Hänggeli, K.P.A.; Hemphill, A.; Müller, N.; Heller, M.; Uldry, A.-C.; Braga-Lagache, S.; Müller, J.; Boubaker, G. Comparative Proteomic Analysis of Toxoplasma gondii RH Wild-Type and Four SRS29B (SAG1) Knock-Out Clones Reveals Significant Differences between Individual Strains. Int. J. Mol. Sci. 2023, 24, 10454. [Google Scholar] [CrossRef]
- Yakubu, R.R.; de Monerri, N.C.S.; Nieves, E.; Kim, K.; Weiss, L.M. Comparative Monomethylarginine Proteomics Suggests that Protein Arginine Methyltransferase 1 (PRMT1) is a Significant Contributor to Arginine Monomethylation in Toxoplasma gondii. Mol. Cell. Proteom. 2017, 16, 567–580. [Google Scholar] [CrossRef]
- Doliwa, C.; Xia, D.; Escotte-Binet, S.; Newsham, E.L.; J., S.S.; Aubert, D.; Randle, N.; Wastling, J.M.; Villena, I. Identification of differentially expressed proteins in sulfadiazine resistant and sensitive strains of Toxoplasma gondii using difference-gel electrophoresis (DIGE). Int. J. Parasitol. Drugs Drug Resist. 2013, 3, 35–44. [CrossRef]
- Marugán-Hernández, V.; Álvarez-García, G.; Tomley, F.; Hemphill, A.; Regidor-Cerrillo, J.; Ortega-Mora, L. Identification of novel rhoptry proteins in Neospora caninum by LC/MS-MS analysis of subcellular fractions. J. Proteom. 2011, 74, 629–642. [Google Scholar] [CrossRef]
- Rashidi, S.; Vieira, C.; Mansouri, R.; Ali-Hassanzadeh, M.; Ghani, E.; Karimazar, M.; Nguewa, P.; Manzano-Román, R. Host cell proteins modulated upon Toxoplasma infection identified using proteomic approaches: a molecular rationale. Parasitol. Res. 2022, 121, 1853–1865. [Google Scholar] [CrossRef]
- Barylyuk, K.; Koreny, L.; Ke, H.; Butterworth, S.; Crook, O.M.; Lassadi, I.; Gupta, V.; Tromer, E.; Mourier, T.; Stevens, T.J.; et al. A Comprehensive Subcellular Atlas of the Toxoplasma Proteome via hyperLOPIT Provides Spatial Context for Protein Functions. Cell Host Microbe 2020, 28, 752–766. [Google Scholar] [CrossRef] [PubMed]
- Demichev, V.; Tober-Lau, P.; Lemke, O.; Nazarenko, T.; Thibeault, C.; Whitwell, H.; Röhl, A.; Freiwald, A.; Szyrwiel, L.; Ludwig, D.; et al. A time-resolved proteomic and prognostic map of COVID-19. Cell Syst. 2021, 12, 780–794. [Google Scholar] [CrossRef]
- Herneisen, A.L., et al., Temporal and thermal profiling of the Toxoplasma proteome implicates parasite Protein Phosphatase 1 in the regulation of Ca2+-responsive pathways. Elife, 2022. 11. [CrossRef]
- Bernard, C.; Locard-Paulet, M.; Noël, C.; Duchateau, M.; Gianetto, Q.G.; Moumen, B.; Rattei, T.; Hechard, Y.; Jensen, L.J.; Matondo, M.; et al. A time-resolved multi-omics atlas of Acanthamoeba castellanii encystment. Nat. Commun. 2022, 13, 1–14. [Google Scholar] [CrossRef]
- Zecha, J.; Bayer, F.P.; Wiechmann, S.; Woortman, J.; Berner, N.; Müller, J.; Schneider, A.; Kramer, K.; Abril-Gil, M.; Hopf, T.; et al. Decrypting drug actions and protein modifications by dose- and time-resolved proteomics. Science 2023, 380, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-X.; Che, L.; Hu, R.-S.; Sun, X.-L. Comparative Phosphoproteomic Analysis of Sporulated Oocysts and Tachyzoites of Toxoplasma gondii Reveals Stage-Specific Patterns. Molecules 2022, 27, 1022. [Google Scholar] [CrossRef] [PubMed]
- Yakubu, R.R.; Weiss, L.M.; de Monerri, N.C.S. Post-translational modifications as key regulators of apicomplexan biology: insights from proteome-wide studies. Mol. Microbiol. 2017, 107, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Laura Vanagas, D.M., Constanza Cristaldi, et al., Histone variant H2B.Z acetylation is necessary for maintenance of Toxoplasma gondii biological fitness. Biochim Biophys Acta Gene Regul Mech, 2023. 2023 Sep;1866(3):194943. [CrossRef]
- Bouchut, A.; Chawla, A.R.; Jeffers, V.; Hudmon, A.; Sullivan, W.J. Proteome-Wide Lysine Acetylation in Cortical Astrocytes and Alterations That Occur during Infection with Brain Parasite Toxoplasma gondii. PLOS ONE 2015, 10, e0117966. [Google Scholar] [CrossRef] [PubMed]
- Stewart, H.I.; Grinfeld, D.; Giannakopulos, A.; Petzoldt, J.; Shanley, T.; Garland, M.; Denisov, E.; Peterson, A.C.; Damoc, E.; Zeller, M.; et al. Parallelized Acquisition of Orbitrap and Astral Analyzers Enables High-Throughput Quantitative Analysis. Anal. Chem. 2023, 95, 15656–15664. [Google Scholar] [CrossRef] [PubMed]
- Perkel, J.M. Single-cell proteomics takes centre stage. Nature 2021, 597, 580–582. [Google Scholar] [CrossRef]
- Kelly, R.T. Single-cell Proteomics: Progress and Prospects. Mol. Cell. Proteom. 2020, 19, 1739–1748. [Google Scholar] [CrossRef]
- Park, J.; Yu, F.; Fulcher, J.M.; Williams, S.M.; Engbrecht, K.; Moore, R.J.; Clair, G.C.; Petyuk, V.; Nesvizhskii, A.I.; Zhu, Y. Evaluating Linear Ion Trap for MS3-Based Multiplexed Single-Cell Proteomics. Anal. Chem. 2023, 95, 1888–1898. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, A.; Sing, J.C.; Gingras, A.-C.; Röst, H.L. Improving Phosphoproteomics Profiling Using Data-Independent Mass Spectrometry. J. Proteome Res. 2022, 21, 1789–1799. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Qiao, L. Data-independent acquisition proteomics methods for analyzing post-translational modifications. Proteomics 2022, 23, e2200046. [Google Scholar] [CrossRef] [PubMed]
- Ulises H Guzman, A.M.D.V., Zilu Ye, Eugen Damoc, et al., Narrow-window DIA: Ultra-fast quantitative analysis of comprehensive proteomes with high sequencing depth. bioRxiv, 2023. [CrossRef]
- Birhanu, A.G. Mass spectrometry-based proteomics as an emerging tool in clinical laboratories. Clin. Proteom. 2023, 20, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.D.L.; Malchow, S.; Reich, S.; Steltgens, S.; Shuvaev, K.V.; Loroch, S.; Lorenz, C.; Sickmann, A.; Knobbe-Thomsen, C.B.; Tews, B.; et al. A sensitive and simple targeted proteomics approach to quantify transcription factor and membrane proteins of the unfolded protein response pathway in glioblastoma cells. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- van Bentum, M. and M. Selbach, An Introduction to Advanced Targeted Acquisition Methods. Mol Cell Proteomics, 2021. 20: p. 100165. [CrossRef]
- Carrera, M.; Piñeiro, C.; Martinez, I. Proteomic Strategies to Evaluate the Impact of Farming Conditions on Food Quality and Safety in Aquaculture Products. Foods 2020, 9, 1050. [Google Scholar] [CrossRef] [PubMed]
- Wu C. C. Tsantilas K. A. Park J. Plubell D. L. Naicker P. Govender I et al. Mag-Net: Rapid Enrichment of Membrane-Bound Particles Enables High Coverage Quantitative Analysis of the Plasma Proteome. bioRxiv, 2023. p 2023.06.10.544439. [CrossRef]
- Heil, L.R.; Damoc, E.; Arrey, T.N.; Pashkova, A.; Denisov, E.; Petzoldt, J.; Peterson, A.C.; Hsu, C.; Searle, B.C.; Shulman, N.; et al. Evaluating the Performance of the Astral Mass Analyzer for Quantitative Proteomics Using Data-Independent Acquisition. J. Proteome Res. 2023, 22, 3290–3300. [Google Scholar] [CrossRef] [PubMed]
- Melby, J.A.; Brown, K.A.; Gregorich, Z.R.; Roberts, D.S.; Chapman, E.A.; Ehlers, L.E.; Gao, Z.; Larson, E.J.; Jin, Y.; Lopez, J.R.; et al. High sensitivity top–down proteomics captures single muscle cell heterogeneity in large proteoforms. Proc. Natl. Acad. Sci. 2023, 120, e2222081120. [Google Scholar] [CrossRef]
- Beltran, P.M.J.; Federspiel, J.D.; Sheng, X.; Cristea, I.M. Proteomics and integrative omic approaches for understanding host–pathogen interactions and infectious diseases. Mol. Syst. Biol. 2017, 13, 922. [Google Scholar] [CrossRef]
- Leo, I.R.; Aswad, L.; Stahl, M.; Kunold, E.; Post, F.; Erkers, T.; Struyf, N.; Mermelekas, G.; Joshi, R.N.; Gracia-Villacampa, E.; et al. Integrative multi-omics and drug response profiling of childhood acute lymphoblastic leukemia cell lines. Nat. Commun. 2022, 13, 1–19. [Google Scholar] [CrossRef]
- Kloehn, J., et al., Multi-omics analysis delineates the distinct functions of sub-cellular acetyl-CoA pools in Toxoplasma gondii. BMC Biol, 2020. 18(1): p. 67. [CrossRef]
- Nie, L.-B.; Cong, W.; He, J.-J.; Zheng, W.-B.; Zhu, X.-Q. Global proteomic profiling of multiple organs of cats (Felis catus) and proteome-transcriptome correlation during acute Toxoplasma gondii infection. Infect. Dis. Poverty 2022, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Reel, P.S.; Reel, S.; Pearson, E.; Trucco, E.; Jefferson, E. Using machine learning approaches for multi-omics data analysis: A review. Biotechnol. Adv. 2021, 49, 107739. [Google Scholar] [CrossRef] [PubMed]
- Sangaré, L.O.; Alayi, T.D.; Westermann, B.; Hovasse, A.; Sindikubwabo, F.; Callebaut, I.; Werkmeister, E.; Lafont, F.; Slomianny, C.; Hakimi, M.-A.; et al. Unconventional endosome-like compartment and retromer complex in Toxoplasma gondii govern parasite integrity and host infection. Nat. Commun. 2016, 7, 11191. [Google Scholar] [CrossRef] [PubMed]
- Angel, S.O.; Figueras, M.J.; Alomar, M.L.; Echeverria, P.C.; Deng, B. Toxoplasma gondii Hsp90: potential roles in essential cellular processes of the parasite. Parasitology 2014, 141, 1138–1147. [Google Scholar] [CrossRef] [PubMed]
- Munera López J, A.A., Figueras MJ, Saldarriaga Cartagena AM, Hortua Triana MA, Diambra L, Vanagas L, Deng B, Moreno SNJ, Angel SO, Analysis of the Interactome of the Toxoplasma gondii Tgj1 HSP40 Chaperone.Proteomes. Proteomes, 2023. 2023 Mar 1;11(1):9. [CrossRef]
- Blank, M.L.; Xia, J.; Morcos, M.M.; Sun, M.; Cantrell, P.S.; Liu, Y.; Zeng, X.; Powell, C.J.; Yates, N.; Boulanger, M.J.; et al. Toxoplasma gondii association with host mitochondria requires key mitochondrial protein import machinery. Proc. Natl. Acad. Sci. 2021, 118. [Google Scholar] [CrossRef] [PubMed]
- Anghel, N.; Müller, J.; Serricchio, M.; Jelk, J.; Bütikofer, P.; Boubaker, G.; Imhof, D.; Ramseier, J.; Desiatkina, O.; Păunescu, E.; et al. Cellular and Molecular Targets of Nucleotide-Tagged Trithiolato-Bridged Arene Ruthenium Complexes in the Protozoan Parasites Toxoplasma gondii and Trypanosoma brucei. Int. J. Mol. Sci. 2021, 22, 10787. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.; Boubaker, G.; Imhof, D.; Hänggeli, K.; Haudenschild, N.; Uldry, A.-C.; Braga-Lagache, S.; Heller, M.; Ortega-Mora, L.-M.; Hemphill, A. Differential Affinity Chromatography Coupled to Mass Spectrometry: A Suitable Tool to Identify Common Binding Proteins of a Broad-Range Antimicrobial Peptide Derived from Leucinostatin. Biomedicines 2022, 10, 2675. [Google Scholar] [CrossRef] [PubMed]
- Li, F.-C.; Nie, L.-B.; Elsheikha, H.M.; Yin, F.-Y.; Zhu, X.-Q. Lysine crotonylation is widespread on proteins of diverse functions and localizations in Toxoplasma gondii. Parasitol. Res. 2021, 120, 1617–1626. [Google Scholar] [CrossRef]
- Sun, X.-M.; Ji, Y.-S.; Elashram, S.A.; Lu, Z.-M.; Liu, X.-Y.; Suo, X.; Chen, Q.-J.; Wang, H. Identification of antigenic proteins of Toxoplasma gondii RH strain recognized by human immunoglobulin G using immunoproteomics. J. Proteom. 2012, 77, 423–432. [Google Scholar] [CrossRef]
- Shkel, O.; Kharkivska, Y.; Kim, Y.K.; Lee, J. Proximity Labeling Techniques: A Multi-Omics Toolbox. Chem. – Asian J. 2021, 17, e202101240. [Google Scholar] [CrossRef]
- Back, P.S.; Moon, A.S.; Pasquarelli, R.R.; Bell, H.N.; Torres, J.A.; Chen, A.L.; Sha, J.; Vashisht, A.A.; Wohlschlegel, J.A.; Bradley, P.J. IMC29 Plays an Important Role in Toxoplasma Endodyogeny and Reveals New Components of the Daughter-Enriched IMC Proteome. mBio 2023, 14, e0304222. [Google Scholar] [CrossRef] [PubMed]
- Cygan, A.M.; Beltran, P.M.J.; Mendoza, A.G.; Branon, T.C.; Ting, A.Y.; Carr, S.A.; Boothroyd, J.C. Proximity-Labeling Reveals Novel Host and Parasite Proteins at the Toxoplasma Parasitophorous Vacuole Membrane. mBio 2021, 12, e0026021. [Google Scholar] [CrossRef]
- Engelberg, K.; Bechtel, T.; Michaud, C.; Weerapana, E.; Gubbels, M.-J. Proteomic characterization of the Toxoplasma gondii cytokinesis machinery portrays an expanded hierarchy of its assembly and function. Nat. Commun. 2022, 13, 1–15. [Google Scholar] [CrossRef]
- Engelberg, K., et al., The apical annuli of Toxoplasma gondii are composed of coiled-coil and signalling proteins embedded in the inner membrane complex sutures. Cell Microbiol, 2020. 22(1): p. e13112. [CrossRef]
- Nadipuram, S.M.; Thind, A.C.; Rayatpisheh, S.; Wohlschlegel, J.A.; Bradley, P.J. Proximity biotinylation reveals novel secreted dense granule proteins of Toxoplasma gondii bradyzoites. PLOS ONE 2020, 15, e0232552. [Google Scholar] [CrossRef] [PubMed]
- Tu, V.; Tomita, T.; Sugi, T.; Mayoral, J.; Han, B.; Yakubu, R.R.; Williams, T.; Horta, A.; Ma, Y.; Weiss, L.M. The Toxoplasma gondii Cyst Wall Interactome. mBio 2020, 11. [Google Scholar] [CrossRef]
- Song, Y.; Li, L.; Mo, X.; Pan, M.; Shen, B.; Fang, R.; Hu, M.; Zhao, J.; Zhou, Y. Essential Functions of Calmodulin and Identification of Its Proximal Interacting Proteins in Tachyzoite-Stage Toxoplasma gondii via BioID Technology. Microbiol. Spectr. 2022, 10, e0136322. [Google Scholar] [CrossRef]
- Trinkle-Mulcahy, L. Recent advances in proximity-based labeling methods for interactome mapping. F1000Research 2019, 8, 135. [Google Scholar] [CrossRef]
- Samavarchi-Tehrani, P.; Samson, R.; Gingras, A.-C. Proximity Dependent Biotinylation: Key Enzymes and Adaptation to Proteomics Approaches. Mol. Cell. Proteom. 2020, 19, 757–773. [Google Scholar] [CrossRef] [PubMed]
- Pan, M., et al., Identification of Novel Dense-Granule Proteins in Toxoplasma gondii by Two Proximity-Based Biotinylation Approaches. J Proteome Res, 2019. 18(1): p. 319-330. [CrossRef]
- Lai, M.-Y.; Abdul-Majid, N.; Lau, Y.-L. Identification of Host Proteins Interacting with Toxoplasma gondii SAG1 by Yeast Two-Hybrid Assay. Acta Parasitol. 2019, 64, 575–581. [Google Scholar] [CrossRef]
- Lai, M.-Y.; Lau, Y.-L. Screening and identification of host proteins interacting with Toxoplasma gondii SAG2 by yeast two-hybrid assay. Parasites Vectors 2017, 10, 1–8. [Google Scholar] [CrossRef]
- Harb, O.S. and D.S. Roos, ToxoDB: Functional Genomics Resource for Toxoplasma and Related Organisms. Methods Mol Biol, 2020. 2071: p. 27-47. [CrossRef]
- Xia, D.; Sanderson, S.J.; Jones, A.R.; Prieto, J.H.; Yates, J.R.; Bromley, E.; Tomley, F.M.; Lal, K.; E Sinden, R.; Brunk, B.P.; et al. The proteome of Toxoplasma gondii: integration with the genome provides novel insights into gene expression and annotation. Genome Biol. 2008, 9, R116–R116. [Google Scholar] [CrossRef] [PubMed]
- Possenti, A., et al., Global proteomic analysis of the oocyst/sporozoite of Toxoplasma gondii reveals commitment to a host-independent lifestyle. BMC Genomics, 2013. 14: p. 183. [CrossRef]
- Krishna, R., et al., A large-scale proteogenomics study of apicomplexan pathogens-Toxoplasma gondii and Neospora caninum. Proteomics, 2015. 15(15): p. 2618-28. [CrossRef]
- Garfoot, A.L.; Wilson, G.M.; Coon, J.J.; Knoll, L.J. Proteomic and transcriptomic analyses of early and late-chronic Toxoplasma gondii infection shows novel and stage specific transcripts. BMC Genom. 2019, 20, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Nardelli, S.C.; Che, F.-Y.; de Monerri, N.C.S.; Xiao, H.; Nieves, E.; Madrid-Aliste, C.; Angel, S.O.; Sullivan, W.J.; Angeletti, R.H.; Kim, K.; et al. The Histone Code of Toxoplasma gondii Comprises Conserved and Unique Posttranslational Modifications. mBio 2013, 4, e00922–13. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, H.E.; Russo, P.; Angulo, N.; Ynocente, R.; Montoya, C.; Diestra, A.; Ferradas, C.; Schiaffino, F.; Florentini, E.; Jimenez, J.; et al. Toward detection of toxoplasmosis from urine in mice using hydro-gel nanoparticles concentration and parallel reaction monitoring mass spectrometry. Nanomedicine: Nanotechnology, Biol. Med. 2017, 14, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Bansal, P.; Antil, N.; Kumar, M.; Yamaryo-Botté, Y.; Rawat, R.S.; Pinto, S.; Datta, K.K.; Katris, N.J.; Botté, C.Y.; Prasad, T.S.K.; et al. Protein kinase TgCDPK7 regulates vesicular trafficking and phospholipid synthesis in Toxoplasma gondii. PLOS Pathog. 2021, 17, e1009325. [Google Scholar] [CrossRef] [PubMed]
- Montano, H., et al., TgTKL4 Is a Novel Kinase That Plays an Important Role in Toxoplasma Morphology and Fitness. Msphere, 2023. 8(2). [CrossRef]
- Yang, C.; Doud, E.H.; Sampson, E.; Arrizabalaga, G. The protein phosphatase PPKL is a key regulator of daughter parasite development in Toxoplasma gondii. mBio 2023, 14, e0225423. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-X.; Hu, R.-S.; Zhu, X.-Q.; Sun, X.-L.; Elsheikha, H.M. Global phosphoproteome analysis reveals significant differences between sporulated oocysts of virulent and avirulent strains of Toxoplasma gondii. Microb. Pathog. 2021, 161, 105240. [Google Scholar] [CrossRef] [PubMed]
- Treeck, M.; Sanders, J.L.; Gaji, R.Y.; LaFavers, K.A.; Child, M.A.; Arrizabalaga, G.; Elias, J.E.; Boothroyd, J.C. The Calcium-Dependent Protein Kinase 3 of Toxoplasma Influences Basal Calcium Levels and Functions beyond Egress as Revealed by Quantitative Phosphoproteome Analysis. PLOS Pathog. 2014, 10, e1004197. [Google Scholar] [CrossRef] [PubMed]
- Caballero, M.C.; Alonso, A.M.; Deng, B.; Attias, M.; de Souza, W.; Corvi, M.M. Identification of new palmitoylated proteins in Toxoplasma gondii. Biochim. et Biophys. Acta (BBA) - Proteins Proteom. 2016, 1864, 400–408. [Google Scholar] [CrossRef]
- Foe, I.T.; Child, M.A.; Majmudar, J.D.; Krishnamurthy, S.; van der Linden, W.A.; Ward, G.E.; Martin, B.R.; Bogyo, M. Global Analysis of Palmitoylated Proteins in Toxoplasma gondii. Cell Host Microbe 2015, 18, 501–511. [Google Scholar] [CrossRef]
- Zexiang Wang, J.L., Qianqian Yang and Xiaolin Sun, Global Proteome-Wide Analysis of Cysteine S-Nitrosylation in Toxoplasma gondii. molecules, 2023. [CrossRef]
- Treeck, M., et al., The Phosphoproteomes of Plasmmodium falciparum and Toxoplasma gondii Reveal Unusual Adaptations Within and Beyond the Parasites’ Boundaries. Cell Host & Microbe, 2011. 10(4): p. 410-419. [CrossRef]
- dos Santos, D.A., et al., Protein kinases on carbon metabolism: potential targets for alternative chemotherapies against toxoplasmosis. Frontiers in Cellular and Infection Microbiology, 2023. 13. [CrossRef]
- O’shaughnessy, W.J.; Dewangan, P.S.; Paiz, E.A.; Reese, M.L. Not your Mother’s MAPKs: Apicomplexan MAPK function in daughter cell budding. PLOS Pathog. 2022, 18, e1010849. [Google Scholar] [CrossRef] [PubMed]
- Gaji, R.Y.; Sharp, A.K.; Brown, A.M. Protein kinases in Toxoplasma gondii. Int. J. Parasitol. 2021, 51, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Ben Chaabene, R.; Lentini, G.; Soldati-Favre, D. Biogenesis and discharge of the rhoptries: Key organelles for entry and hijack of host cells by the Apicomplexa. Mol. Microbiol. 2020, 115, 453–465. [Google Scholar] [CrossRef] [PubMed]
- Triana, M.A.H.; Márquez-Nogueras, K.M.; Vella, S.A.; Moreno, S.N. Calcium signaling and the lytic cycle of the Apicomplexan parasite Toxoplasma gondii. Biochim. et Biophys. Acta (BBA) - Mol. Cell Res. 2018, 1865, 1846–1856. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.J.; Augusto, L.d.S.; Zhang, M.; Wek, R.C.; Sullivan, W.J. Translational Control in the Latency of Apicomplexan Parasites. Trends Parasitol. 2017, 33, 947–960. [Google Scholar] [CrossRef] [PubMed]
- Shukri AH, L.V., Charih F, Biggar KK, Unraveling the battle for lysine: A review of the competition among post-translational modifications. Biochim Biophys Acta Gene Regul Mech, 2023. 2023 Sep 24;1866(4):194990. [CrossRef]
- Mukherjee, S.; Hao, Y.-H.; Orth, K. A newly discovered post-translational modification – the acetylation of serine and threonine residues. Trends Biochem. Sci. 2007, 32, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Christensen, D.G.; Xie, X.; Basisty, N.; Byrnes, J.; McSweeney, S.; Schilling, B.; Wolfe, A.J. Post-translational Protein Acetylation: An Elegant Mechanism for Bacteria to Dynamically Regulate Metabolic Functions. Front. Microbiol. 2019, 10, 1604. [Google Scholar] [CrossRef] [PubMed]
- Vanagas, L., et al., Toxoplasma gondii histone acetylation remodelers as novel drug targets. Expert Review of Anti-Infective Therapy, 2012. 10(10): p. 1189-1201. [CrossRef]
- Bougdour, A.; Maubon, D.; Baldacci, P.; Ortet, P.; Bastien, O.; Bouillon, A.; Barale, J.-C.; Pelloux, H.; Ménard, R.; Hakimi, M.-A. Drug inhibition of HDAC3 and epigenetic control of differentiation in Apicomplexa parasites. J. Exp. Med. 2009, 206, 953–966. [Google Scholar] [CrossRef] [PubMed]
- Darkin-Rattray, S.J.; Gurnett, A.M.; Myers, R.W.; Dulski, P.M.; Crumley, T.M.; Allocco, J.J.; Cannova, C.; Meinke, P.T.; Colletti, S.L.; Bednarek, M.A.; et al. Apicidin: A novel antiprotozoal agent that inhibits parasite histone deacetylase. Proc. Natl. Acad. Sci. 1996, 93, 13143–13147. [Google Scholar] [CrossRef]
- Boissavy, T.; Rotili, D.; Mouveaux, T.; Roger, E.; Aliouat, E.M.; Pierrot, C.; Valente, S.; Mai, A.; Gissot, M. Hydroxamate-based compounds are potent inhibitors of Toxoplasma gondii HDAC biological activity. Antimicrob. Agents Chemother. 2023, 67, e0066123. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Q.; Li, H.; Cong, H.; Qu, Y. In vitro and in vivo anti−Toxoplasma activities of HDAC inhibitor Panobinostat on experimental acute ocular toxoplasmosis. Front. Cell. Infect. Microbiol. 2022, 12, 1002817. [Google Scholar] [CrossRef] [PubMed]
- Jublot, D.; Cavaillès, P.; Kamche, S.; Francisco, D.; Fontinha, D.; Prudêncio, M.; Guichou, J.-F.; Labesse, G.; Sereno, D.; Loeuillet, C. A Histone Deacetylase (HDAC) Inhibitor with Pleiotropic In Vitro Anti-Toxoplasma and Anti-Plasmodium Activities Controls Acute and Chronic Toxoplasma Infection in Mice. Int. J. Mol. Sci. 2022, 23, 3254. [Google Scholar] [CrossRef] [PubMed]
- Araujo-Silva, C.A.; De Souza, W.; Martins-Duarte, E.S.; Vommaro, R.C. HDAC inhibitors Tubastatin A and SAHA affect parasite cell division and are potential anti-Toxoplasma gondii chemotherapeutics. Int. J. Parasitol. Drugs Drug Resist. 2020, 15, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Mouveaux, T.; Rotili, D.; Boissavy, T.; Roger, E.; Pierrot, C.; Mai, A.; Gissot, M. A potent HDAC inhibitor blocks Toxoplasma gondii tachyzoite growth and profoundly disrupts parasite gene expression. Int. J. Antimicrob. Agents 2022, 59, 106526. [Google Scholar] [CrossRef] [PubMed]
- Contreras, S.M.; Ganuza, A.; Corvi, M.M.; Angel, S.O. Resveratrol induces H3 and H4K16 deacetylation and H2A.X phosphorylation in Toxoplasma gondii. BMC Res. Notes 2021, 14, 1–8. [Google Scholar] [CrossRef]
- Jeffers, V.; Sullivan, W.J. Lysine Acetylation Is Widespread on Proteins of Diverse Function and Localization in the Protozoan Parasite Toxoplasma gondii. Eukaryot. Cell 2012, 11, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-X.; Hu, R.-S.; Zhou, C.-X.; He, J.-J.; Elsheikha, H.M.; Zhu, X.-Q. Label-Free Quantitative Acetylome Analysis Reveals Toxoplasma gondii Genotype-Specific Acetylomic Signatures. Microorganisms 2019, 7, 510. [Google Scholar] [CrossRef]
- Trempe, J.F., Reading the ubiquitin postal code. Current Opinion in Structural Biology, 2011. 21(6): p. 792-801. [CrossRef]
- Silmon de Monerri, N.C., et al., The Ubiquitin Proteome of Toxoplasma gondii Reveals Roles for Protein Ubiquitination in Cell-Cycle Transitions. Cell Host Microbe, 2015. 18(5): p. 621-33. [CrossRef]
- MacRae, J.I.; Sheiner, L.; Nahid, A.; Tonkin, C.; Striepen, B.; McConville, M.J. Mitochondrial Metabolism of Glucose and Glutamine Is Required for Intracellular Growth of Toxoplasma gondii. Cell Host Microbe 2012, 12, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Grimes, M.; Hall, B.; Foltz, L.; Levy, T.; Rikova, K.; Gaiser, J.; Cook, W.; Smirnova, E.; Wheeler, T.; Clark, N.R.; et al. Integration of protein phosphorylation, acetylation, and methylation data sets to outline lung cancer signaling networks. Sci. Signal. 2018, 11. [Google Scholar] [CrossRef]
- Ross, K.E.; Zhang, G.; Akcora, C.; Lin, Y.; Fang, B.; Koomen, J.; Haura, E.B.; Grimes, M. Network models of protein phosphorylation, acetylation, and ubiquitination connect metabolic and cell signaling pathways in lung cancer. PLOS Comput. Biol. 2023, 19, e1010690. [Google Scholar] [CrossRef]
- Lee, J.-S.; Smith, E.; Shilatifard, A. The Language of Histone Crosstalk. Cell 2010, 142, 682–685. [Google Scholar] [CrossRef] [PubMed]
- Talbert, P.B.; Henikoff, S. The Yin and Yang of Histone Marks in Transcription. Annu. Rev. Genom. Hum. Genet. 2021, 22, 147–170. [Google Scholar] [CrossRef] [PubMed]
- Yin, D.; Jiang, N.; Zhang, Y.; Wang, D.; Sang, X.; Feng, Y.; Chen, R.; Wang, X.; Yang, N.; Chen, Q. Global Lysine Crotonylation and 2-Hydroxyisobutyrylation in Phenotypically Different Toxoplasma gondii Parasites. Mol. Cell. Proteom. 2019, 18, 2207–2224. [Google Scholar] [CrossRef] [PubMed]
- Contreras, S.M., et al., Architecture, Chromatin and Gene Organization of Toxoplasma gondii Subtelomeres. Epigenomes, 2022. 6(3). [CrossRef]
- Sindikubwabo, F., et al., Modifications at K31 on the lateral surface of histone H4 contribute to genome structure and expression in apicomplexan parasites. Elife, 2017. 6. [CrossRef]
- Dalmasso, M.C.; Onyango, D.O.; Naguleswaran, A.; Sullivan, W.J.; Angel, S.O. Toxoplasma H2A Variants Reveal Novel Insights into Nucleosome Composition and Functions for this Histone Family. J. Mol. Biol. 2009, 392, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Hoeijmakers, W.A., et al., H2A.Z/H2B.Z double-variant nucleosomes inhabit the AT-rich promoter regions of the Plasmodium falciparum genome. Mol Microbiol, 2013. 87(5): p. 1061-73. [CrossRef]
- Nardelli, S.C.; de Monerri, N.C.S.; Vanagas, L.; Wang, X.; Tampaki, Z.; Sullivan, W.J.; Angel, S.O.; Kim, K. Genome-wide localization of histone variants in Toxoplasma gondii implicates variant exchange in stage-specific gene expression. BMC Genom. 2022, 23, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Azizan S, S.S., Tang J, Jeninga MD, Schulz D, et al., The P. falciparum alternative histones Pf H2A.Z and Pf H2B.Z are dynamically acetylated and antagonized by PfSir2 histone deacetylases at heterochromatin boundaries. mBio, 2023. [CrossRef]
- Bougdour, A.; Braun, L.; Cannella, D.; Hakimi, M.-A. Chromatin modifications: implications in the regulation of gene expression in Toxoplasma gondii. Cell. Microbiol. 2010, 12, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Saksouk, N.; Bhatti, M.M.; Kieffer, S.; Smith, A.T.; Musset, K.; Garin, J.; Sullivan, W.J.; Cesbron-Delauw, M.-F.; Hakimi, M.-A. Histone-Modifying Complexes Regulate Gene Expression Pertinent to the Differentiation of the Protozoan Parasite Toxoplasma gondii. Mol. Cell. Biol. 2005, 25, 10379–10390. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-S.; Shukla, A.; Schneider, J.; Swanson, S.K.; Washburn, M.P.; Florens, L.; Bhaumik, S.R.; Shilatifard, A. Histone Crosstalk between H2B Monoubiquitination and H3 Methylation Mediated by COMPASS. Cell 2007, 131, 1084–1096. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, S.; Lee, J.S.; Gardner, K.E.; Gardner, J.M.; Takahashi, Y.-H.; Chandrasekharan, M.B.; Sun, Z.-W.; Osley, M.A.; Strahl, B.D.; Jaspersen, S.L.; et al. Histone H2BK123 monoubiquitination is the critical determinant for H3K4 and H3K79 trimethylation by COMPASS and Dot1. J. Cell Biol. 2009, 186, 371–377. [Google Scholar] [CrossRef]
- Zhao, R.; Davey, M.; Hsu, Y.-C.; Kaplanek, P.; Tong, A.; Parsons, A.B.; Krogan, N.; Cagney, G.; Mai, D.; Greenblatt, J.; et al. Navigating the Chaperone Network: An Integrative Map of Physical and Genetic Interactions Mediated by the Hsp90 Chaperone. Cell 2005, 120, 715–727. [Google Scholar] [CrossRef]
- Taipale, M.; Tucker, G.; Peng, J.; Krykbaeva, I.; Lin, Z.-Y.; Larsen, B.; Choi, H.; Berger, B.; Gingras, A.-C.; Lindquist, S. A Quantitative Chaperone Interaction Network Reveals the Architecture of Cellular Protein Homeostasis Pathways. Cell 2014, 158, 434–448. [Google Scholar] [CrossRef] [PubMed]
- Sawarkar, R.; Paro, R. Hsp90@chromatin.nucleus: an emerging hub of a networker. Trends Cell Biol. 2013, 23, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Backe, S.J.; Sager, R.A.; Woodford, M.R.; Makedon, A.M.; Mollapour, M. Post-translational modifications of Hsp90 and translating the chaperone code. J. Biol. Chem. 2020, 295, 11099–11117. [Google Scholar] [CrossRef] [PubMed]
- Dagar, M.; Singh, J.P.; Dagar, G.; Tyagi, R.K.; Bagchi, G. Phosphorylation of HSP90 by protein kinase A is essential for the nuclear translocation of androgen receptor. J. Biol. Chem. 2019, 294, 8699–8710. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lu, X.-A.; Song, X.; Zhuo, W.; Jia, L.; Jiang, Y.; Luo, Y. Thr90 phosphorylation of Hsp90α by protein kinase A regulates its chaperone machinery. Biochem. J. 2011, 441, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Solier, S.; Kohn, K.W.; Scroggins, B.; Xu, W.; Trepel, J.; Neckers, L.; Pommier, Y. Heat shock protein 90α (HSP90α), a substrate and chaperone of DNA-PK necessary for the apoptotic response. Proc. Natl. Acad. Sci. USA 2012, 109, 12866–12872. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.H.; An, S.; Lee, H.-C.; Jin, H.-O.; Seo, S.-K.; Yoo, D.-H.; Lee, K.-H.; Rhee, C.H.; Choi, E.-J.; Hong, S.-I.; et al. A Truncated Form of p23 Down-regulates Telomerase Activity via Disruption of Hsp90 Function. J. Biol. Chem. 2009, 284, 30871–30880. [Google Scholar] [CrossRef]
- Scroggins, B.T.; Robzyk, K.; Wang, D.; Marcu, M.G.; Tsutsumi, S.; Beebe, K.; Cotter, R.J.; Felts, S.; Toft, D.; Karnitz, L.; et al. An Acetylation Site in the Middle Domain of Hsp90 Regulates Chaperone Function. Mol. Cell 2007, 25, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Rao, R.; Shen, J.; Tang, Y.; Fiskus, W.; Nechtman, J.; Atadja, P.; Bhalla, K. Role of Acetylation and Extracellular Location of Heat Shock Protein 90α in Tumor Cell Invasion. Cancer Res 2008, 68, 4833–4842. [Google Scholar] [CrossRef]
- Muller, P.; Ruckova, E.; Halada, P.; Coates, P.J.; Hrstka, R.; Lane, D.P.; Vojtesek, B. C-terminal phosphorylation of Hsp70 and Hsp90 regulates alternate binding to co-chaperones CHIP and HOP to determine cellular protein folding/degradation balances. Oncogene 2012, 32, 3101–3110. [Google Scholar] [CrossRef]
- Hu, K.; Johnson, J.; Florens, L.; Fraunholz, M.; Suravajjala, S.; DiLullo, C.; Yates, J.; Roos, D.S.; Murray, J.M. Cytoskeletal Components of an Invasion Machine—The Apical Complex of Toxoplasma gondii. PLOS Pathog. 2006, 2, e13–e13. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Flores, C.J.; Cruz-Mirón, R.; Mondragón-Castelán, M.E.; González-Pozos, S.; Ríos-Castro, E.; Mondragón-Flores, R. Proteomic and structural characterization of self-assembled vesicles from excretion/secretion products of Toxoplasma gondii. J. Proteom. 2019, 208, 103490. [Google Scholar] [CrossRef] [PubMed]
- Wowk, P.F.; Zardo, M.L.; Miot, H.T.; Goldenberg, S.; Carvalho, P.C.; Mörking, P.A. Proteomic profiling of extracellular vesicles secreted from Toxoplasma gondii. Proteomics 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- Tu, V.; Mayoral, J.; Sugi, T.; Tomita, T.; Han, B.; Ma, Y.F.; Weiss, L.M. Enrichment and Proteomic Characterization of the Cyst Wall from In Vitro Toxoplasma gondii Cysts. mBio 2019, 10, e00469–19. [Google Scholar] [CrossRef] [PubMed]
- Bradley, P.J.; Ward, C.; Cheng, S.J.; Alexander, D.L.; Coller, S.; Coombs, G.H.; Dunn, J.D.; Ferguson, D.J.; Sanderson, S.J.; Wastling, J.M.; et al. Proteomic Analysis of Rhoptry Organelles Reveals Many Novel Constituents for Host-Parasite Interactions in Toxoplasma gondii. J. Biol. Chem. 2005, 280, 34245–34258. [Google Scholar] [CrossRef]
- Fleige, T.; Pfaff, N.; Gross, U.; Bohne, W. Localisation of gluconeogenesis and tricarboxylic acid (TCA)-cycle enzymes and first functional analysis of the TCA cycle in Toxoplasma gondii. Int. J. Parasitol. 2008, 38, 1121–1132. [Google Scholar] [CrossRef] [PubMed]
- Seidi, A., et al., Elucidating the mitochondrial proteome of Toxoplasma gondii reveals the presence of a divergent cytochrome c oxidase. Elife, 2018. 7. [CrossRef]
- Elzek, M.A.W., et al., Localization of Organelle Proteins by Isotope Tagging: Current status and potential applications in drug discovery research. Drug Discov Today Technol, 2021. 39: p. 57-67. [CrossRef]
- Hajj, R.E., et al., Toxoplasmosis: Current and Emerging Parasite Druggable Targets. Microorganisms, 2021. 9(12). [CrossRef]
- Rashidi, S.; Sánchez-Montejo, J.; Mansouri, R.; Ali-Hassanzadeh, M.; Savardashtaki, A.; Bahreini, M.S.; Karimazar, M.; Manzano-Román, R.; Nguewa, P. Mining the Proteome of Toxoplasma Parasites Seeking Vaccine and Diagnostic Candidates. Animals 2022, 12, 1098. [Google Scholar] [CrossRef] [PubMed]
- Stryiński, R.; Łopieńska-Biernat, E.; Carrera, M. Proteomic Insights into the Biology of the Most Important Foodborne Parasites in Europe. Foods 2020, 9, 1403. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.; Hemphill, A. Toxoplasma gondii infection: novel emerging therapeutic targets. Expert Opin. Ther. Targets 2023, 27, 293–304. [Google Scholar] [CrossRef]
- Sabou, M.; Doderer-Lang, C.; Leyer, C.; Konjic, A.; Kubina, S.; Lennon, S.; Rohr, O.; Viville, S.; Cianférani, S.; Candolfi, E.; et al. Toxoplasma gondii ROP16 kinase silences the cyclin B1 gene promoter by hijacking host cell UHRF1-dependent epigenetic pathways. Cell. Mol. Life Sci. 2019, 77, 2141–2156. [Google Scholar] [CrossRef]
- Müller, J.; Hemphill, A. Drug target identification in protozoan parasites. Expert Opin. Drug Discov. 2016, 11, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Kent, R.S.; Briggs, E.M.; Colon, B.L.; Alvarez, C.; Pereira, S.S.; De Niz, M. Paving the Way: Contributions of Big Data to Apicomplexan and Kinetoplastid Research. Front. Cell. Infect. Microbiol. 2022, 12, 900878. [Google Scholar] [CrossRef] [PubMed]
- Benns, H.J.; Storch, M.; Falco, J.A.; Fisher, F.R.; Tamaki, F.; Alves, E.; Wincott, C.J.; Milne, R.; Wiedemar, N.; Craven, G.; et al. CRISPR-based oligo recombineering prioritizes apicomplexan cysteines for drug discovery. Nat. Microbiol. 2022, 7, 1891–1905. [Google Scholar] [CrossRef] [PubMed]
- Freville, A., et al., Deciphering the Role of Protein Phosphatases in Apicomplexa: The Future of Innovative Therapeutics? Microorganisms, 2022. 10(3). [CrossRef]
- Broncel, M., et al., Profiling of myristoylation in Toxoplasma gondii reveals an N-myristoylated protein important for host cell penetration. Elife, 2020. 9. [CrossRef]
- Liu, Y.; Tang, Y.; Tang, X.; Wu, M.; Hou, S.; Liu, X.; Li, J.; Deng, M.; Huang, S.; Jiang, L. Anti-Toxoplasma gondii Effects of a Novel Spider Peptide XYP1 In Vitro and In Vivo. Biomedicines 2021, 9, 934. [Google Scholar] [CrossRef]
- Che, F.-Y.; Madrid-Aliste, C.; Burd, B.; Zhang, H.; Nieves, E.; Kim, K.; Fiser, A.; Angeletti, R.H.; Weiss, L.M. Comprehensive Proteomic Analysis of Membrane Proteins in Toxoplasma gondii. Mol. Cell. Proteom. 2011, 10. [Google Scholar] [CrossRef]
- Maran, S.R.; Fleck, K.; Monteiro-Teles, N.M.; Isebe, T.; Walrad, P.; Jeffers, V.; Cestari, I.; Vasconcelos, E.J.; Moretti, N. Protein acetylation in the critical biological processes in protozoan parasites. Trends Parasitol. 2021, 37, 815–830. [Google Scholar] [CrossRef]
- Renaud, E.A.; Pamukcu, S.; Cerutti, A.; Berry, L.; Lemaire-Vieille, C.; Yamaryo-Botté, Y.; Botté, C.Y.; Besteiro, S. Disrupting the plastidic iron-sulfur cluster biogenesis pathway in Toxoplasma gondii has pleiotropic effects irreversibly impacting parasite viability. J. Biol. Chem. 2022, 298, 102243. [Google Scholar] [CrossRef]
- Zhang, H.; Lee, E.-G.; Yu, L.; Kawano, S.; Huang, P.; Liao, M.; Kawase, O.; Zhang, G.; Zhou, J.; Fujisaki, K.; et al. Identification of the cross-reactive and species-specific antigens between Neospora caninum and Toxoplasma gondii tachyzoites by a proteomics approach. Parasitol. Res. 2011, 109, 899–911. [Google Scholar] [CrossRef]
- Sander, V.A.; López, E.F.S.; Morales, L.M.; Duarte, V.A.R.; Corigliano, M.G.; Clemente, M. Use of Veterinary Vaccines for Livestock as a Strategy to Control Foodborne Parasitic Diseases. Front. Cell. Infect. Microbiol. 2020, 10, 288. [Google Scholar] [CrossRef]
- Cajazeiro, D.C.; Toledo, P.P.M.; de Sousa, N.F.; Scotti, M.T.; Reimão, J.Q. Drug Repurposing Based on Protozoan Proteome: In Vitro Evaluation of In Silico Screened Compounds against Toxoplasma gondii. Pharmaceutics 2022, 14, 1634. [Google Scholar] [CrossRef]
- Montazeri, M.; Mehrzadi, S.; Sharif, M.; Sarvi, S.; Tanzifi, A.; Aghayan, S.A.; Daryani, A. Drug Resistance in Toxoplasma gondii. Front. Microbiol. 2018, 9, 2587. [Google Scholar] [CrossRef] [PubMed]
- Thabet, A., et al., Resistance towards monensin is proposed to be acquired in a Toxoplasma gondii model by reduced invasion and egress activities, in addition to increased intracellular replication. Parasitology, 2018. 145(3): p. 313-325. [CrossRef]
- Chen, T.; Ning, D.; Sun, H.; Li, R.; Shang, M.; Li, X.; Wang, X.; Chen, W.; Liang, C.; Li, W.; et al. Sequence Analysis and Molecular Characterization of Clonorchis sinensis Hexokinase, an Unusual Trimeric 50-kDa Glucose-6-Phosphate-Sensitive Allosteric Enzyme. PLOS ONE 2014, 9, e107940. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, S.; Nice, E.C.; Deutsch, E.W.; Lane, L.; Omenn, G.S.; Pennington, S.R.; Paik, Y.-K.; Overall, C.M.; Corrales, F.J.; Cristea, I.M.; et al. A high-stringency blueprint of the human proteome. Nat. Commun. 2020, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, C.; Li, X.; Ehsan, M.; Lu, M.; Li, K.; Xu, L.; Yan, R.; Song, X.; Li, X. Proteomics analysis reveals that the proto-oncogene eIF-5A indirectly influences the growth, invasion and replication of Toxoplasma gondii tachyzoite. Parasites Vectors 2021, 14, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.W.; Blackman, M.J.; Howell, S.A.; Carruthers, V.B. Proteomic Analysis of Cleavage Events Reveals a Dynamic Two-step Mechanism for Proteolysis of a Key Parasite Adhesive Complex. Mol. Cell. Proteom. 2004, 3, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.F., et al., Identification of a TNF-α inducer MIC3 originating from the microneme of non-cystogenic, virulent. Scientific Reports, 2016. 6. [CrossRef]
- Kawase, O.; Nishikawa, Y.; Bannai, H.; Zhang, H.; Zhang, G.; Jin, S.; Lee, E.-G.; Xuan, X. Proteomic analysis of calcium-dependent secretion in Toxoplasma gondii. Proteomics 2007, 7, 3718–3725. [Google Scholar] [CrossRef] [PubMed]
- Ghaffari, A.D., et al., Immunoinformatic analysis of immunogenic B- and T-cell epitopes of MIC4 protein to designing a vaccine candidate against Toxoplasma gondii through an in-silico approach. Clinical and Experimental Vaccine Research, 2021. 10(1): p. 59-77. [CrossRef]
- Dogga, S.K., et al., A druggable secretory protein maturase of Toxoplasma essential for invasion and egress. Elife, 2017. 6. [CrossRef]
- Ferra, B.; Holec-Gąsior, L.; Grąźlewska, W. Toxoplasma gondii Recombinant Antigens in the Serodiagnosis of Toxoplasmosis in Domestic and Farm Animals. Animals 2020, 10, 1245. [Google Scholar] [CrossRef]
- Bradley, P.J.; Sibley, L.D. Rhoptries: an arsenal of secreted virulence factors. Curr. Opin. Microbiol. 2007, 10, 582–587. [Google Scholar] [CrossRef] [PubMed]
- El Hajj, H.; Demey, E.; Poncet, J.; Lebrun, M.; Wu, B.; Galéotti, N.; Fourmaux, M.N.; Mercereau-Puijalon, O.; Vial, H.; Labesse, G.; et al. The ROP2 family ofToxoplasma gondii rhoptry proteins: Proteomic and genomic characterization and molecular modeling. Proteomics 2006, 6, 5773–5784. [Google Scholar] [CrossRef]
- Antil, N.; Kumar, M.; Behera, S.K.; Arefian, M.; Kotimoole, C.N.; Rex, D.A.B.; Prasad, T.S.K. Unraveling Toxoplasma gondii GT1 Strain Virulence and New Protein-Coding Genes with Proteogenomic Analyses. OMICS: A J. Integr. Biol. 2021, 25, 591–604. [Google Scholar] [CrossRef]
- Wang, Z.X., et al., iTRAQ-Based Global Phosphoproteomics Reveals Novel Molecular Differences Between Toxoplasma gondii Strains of Different Genotypes. Frontiers in Cellular and Infection Microbiology, 2019. 9. [CrossRef]
- Fukumoto, J.; Sakura, T.; Matsubara, R.; Tahara, M.; Matsuzaki, M.; Nagamune, K. Rhoptry kinase protein 39 (ROP39) is a novel factor that recruits host mitochondria to the parasitophorous vacuole of Toxoplasma gondii. Biol. Open 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Echeverria, P.C.; Figueras, M.J.; Vogler, M.; Kriehuber, T.; de Miguel, N.; Deng, B.; Dalmasso, M.C.; Matthews, D.E.; Matrajt, M.; Haslbeck, M.; et al. The Hsp90 co-chaperone p23 of Toxoplasma gondii: Identification, functional analysis and dynamic interactome determination. Mol. Biochem. Parasitol. 2010, 172, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Fritz, H.M.; Bowyer, P.W.; Bogyo, M.; Conrad, P.A.; Boothroyd, J.C. Proteomic Analysis of Fractionated Toxoplasma Oocysts Reveals Clues to Their Environmental Resistance. PLOS ONE 2012, 7, e29955. [Google Scholar] [CrossRef] [PubMed]
- Sangaré, L.O., et al., Toxoplasma GRA15 Activates the NF-κB Pathway through Interactions with TNF Receptor-Associated Factors. Mbio, 2019. 10(4). [CrossRef]
- Fox, B.A.; Rommereim, L.M.; Guevara, R.B.; Falla, A.; Triana, M.A.H.; Sun, Y.; Bzik, D.J. The Toxoplasma gondii Rhoptry Kinome Is Essential for Chronic Infection. mBio 2016, 7, e00193–16. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Mirón, R.; Ramírez-Flores, C.J.; Lagunas-Cortés, N.; Mondragón-Castelán, M.; Ríos-Castro, E.; González-Pozos, S.; Aguirre-García, M.M.; Mondragón-Flores, R. Proteomic characterization of the pellicle of Toxoplasma gondii. J. Proteom. 2021, 237, 104146. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.-Y.; Zhang, J.-Z.; Yin, G.-R.; Meng, X.-L.; Zhao, F. Toxoplasma gondii: Proteomic analysis of antigenicity of soluble tachyzoite antigen. Exp. Parasitol. 2009, 122, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.H., et al., Comparative proteomic analysis of different Toxoplasma gondii genotypes by two-dimensional fluorescence difference gel electrophoresis combined with mass spectrometry. Electrophoresis, 2014. 35(4): p. 533-45. [CrossRef]
- Leal-Sena, J.A., et al., Toxoplasma gondii antigen SAG2A differentially modulates IL-1beta expression in resistant and susceptible murine peritoneal cells. Appl Microbiol Biotechnol, 2018. 102(5): p. 2235-2249. [CrossRef]
- He, C., et al., iTRAQ-Based Phosphoproteomic Analysis of Toxoplasma gondii Tachyzoites Provides Insight Into the Role of Phosphorylation for its Invasion and Egress. Frontiers in Cellular and Infection Microbiology, 2020. 10. [CrossRef]
- Lockyer, E.J.; Torelli, F.; Butterworth, S.; Song, O.-R.; Howell, S.; Weston, A.; East, P.; Treeck, M. A heterotrimeric complex of Toxoplasma proteins promotes parasite survival in interferon gamma-stimulated human cells. PLOS Biol. 2023, 21, e3002202. [Google Scholar] [CrossRef] [PubMed]
- Coffey, M.J.; Dagley, L.F.; Seizova, S.; Kapp, E.A.; Infusini, G.; Roos, D.S.; Boddey, J.A.; Webb, A.I.; Tonkin, C.J. Aspartyl Protease 5 Matures Dense Granule Proteins That Reside at the Host-Parasite Interface in Toxoplasma gondii. mBio 2018, 9, e01796–18. [Google Scholar] [CrossRef] [PubMed]
- Mayoral, J.; Tomita, T.; Tu, V.; Aguilan, J.T.; Sidoli, S.; Weiss, L.M. Toxoplasma gondii PPM3C, a secreted protein phosphatase, affects parasitophorous vacuole effector export. PLOS Pathog. 2020, 16, e1008771. [Google Scholar] [CrossRef] [PubMed]
- Cygan, A.M., et al., Coimmunoprecipitation with MYR1 Identifies Three Additional Proteins within the Toxoplasma gondii Parasitophorous Vacuole Required for Translocation of Dense Granule Effectors into Host Cells. Msphere, 2020. 5(1). [CrossRef]
- Nadipuram, S.M.; Kim, E.W.; Vashisht, A.A.; Lin, A.H.; Bell, H.N.; Coppens, I.; Wohlschlegel, J.A.; Bradley, P.J. In Vivo Biotinylation of the Toxoplasma Parasitophorous Vacuole Reveals Novel Dense Granule Proteins Important for Parasite Growth and Pathogenesis. mBio 2016, 7, e00808–16. [Google Scholar] [CrossRef]
- Young, J.C.; Broncel, M.; Teague, H.; Russell, M.R.G.; McGovern, O.L.; Renshaw, M.; Frith, D.; Snijders, A.P.; Collinson, L.; Carruthers, V.B.; et al. Phosphorylation of Toxoplasma gondii Secreted Proteins during Acute and Chronic Stages of Infection. mSphere 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Mayoral, J.; Guevara, R.B.; Rivera-Cuevas, Y.; Tu, V.; Tomita, T.; Romano, J.D.; Gunther-Cummins, L.; Sidoli, S.; Coppens, I.; Carruthers, V.B.; et al. Dense Granule Protein GRA64 Interacts with Host Cell ESCRT Proteins during Toxoplasma gondii Infection. mBio 2022, 13, e0144222. [Google Scholar] [CrossRef] [PubMed]
- Mercer, H.L.; Snyder, L.M.; Doherty, C.M.; Fox, B.A.; Bzik, D.J.; Denkers, E.Y. Toxoplasma gondii dense granule protein GRA24 drives MyD88-independent p38 MAPK activation, IL-12 production and induction of protective immunity. PLOS Pathog. 2020, 16, e1008572. [Google Scholar] [CrossRef] [PubMed]
- Shastri, A.J.; Marino, N.D.; Franco, M.; Lodoen, M.B.; Boothroyd, J.C. GRA25 Is a Novel Virulence Factor of Toxoplasma gondii and Influences the Host Immune Response. Infect. Immun. 2014, 82, 2595–2605. [Google Scholar] [CrossRef] [PubMed]
- Bougdour, A.; Durandau, E.; Brenier-Pinchart, M.-P.; Ortet, P.; Barakat, M.; Kieffer, S.; Curt-Varesano, A.; Curt-Bertini, R.-L.; Bastien, O.; Coute, Y.; et al. Host Cell Subversion by Toxoplasma GRA16, an Exported Dense Granule Protein that Targets the Host Cell Nucleus and Alters Gene Expression. Cell Host Microbe 2013, 13, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Elizabeth, N. Rudzki, S.E.A., Rachel S. Coombs, et al., Toxoplasma gondiiGRA28 Is Required for Placenta-Specific Induction of the Regulatory Chemokine CCL22 in Human and Mouse. mBio, 2021. 2021 Nov-Dec; 12(6): e01591-21(2021 Nov-Dec; 12(6): e01591-21). [CrossRef]
- Takemae, H.; Sugi, T.; Kobayashi, K.; Gong, H.; Ishiwa, A.; Recuenco, F.C.; Murakoshi, F.; Iwanaga, T.; Inomata, A.; Horimoto, T.; et al. Characterization of the interaction between Toxoplasma gondii rhoptry neck protein 4 and host cellular β-tubulin. Sci. Rep. 2013, 3, 3199. [Google Scholar] [CrossRef] [PubMed]
- Poukchanski, A.; Fritz, H.M.; Tonkin, M.L.; Treeck, M.; Boulanger, M.J.; Boothroyd, J.C. Toxoplasma gondii Sporozoites Invade Host Cells Using Two Novel Paralogues of RON2 and AMA1. PLOS ONE 2013, 8, e70637. [Google Scholar] [CrossRef] [PubMed]
- de Leon, C.T.G., et al., Proteomic characterization of the subpellicular cytoskeleton of Toxoplasma gondii tachyzoites. Journal of Proteomics, 2014. 111: p. 86-99. [CrossRef]
- Vigetti, L.; Labouré, T.; Roumégous, C.; Cannella, D.; Touquet, B.; Mayer, C.; Couté, Y.; Frénal, K.; Tardieux, I.; Renesto, P. The BCC7 Protein Contributes to the Toxoplasma Basal Pole by Interfacing between the MyoC Motor and the IMC Membrane Network. Int. J. Mol. Sci. 2022, 23, 5995. [Google Scholar] [CrossRef]
- Roumégous, C.; Hammoud, A.A.; Fuster, D.; Dupuy, J.-W.; Blancard, C.; Salin, B.; Robinson, D.R.; Renesto, P.; Tardieux, I.; Frénal, K. Identification of new components of the basal pole of Toxoplasma gondii provides novel insights into its molecular organization and functions. Front. Cell. Infect. Microbiol. 2022, 12, 1010038. [Google Scholar] [CrossRef]
- He, C., et al., iTRAQ-based phosphoproteomic analysis reveals host cell’s specific responses to Toxoplasma gondii at the phases of invasion and prior to egress. Biochimica Et Biophysica Acta-Proteins and Proteomics, 2019. 1867(3): p. 202-212. [CrossRef]
- Rompikuntal, P.K.; Kent, R.S.; Foe, I.T.; Deng, B.; Bogyo, M.; Ward, G.E. Blocking Palmitoylation of Toxoplasma gondii Myosin Light Chain 1 Disrupts Glideosome Composition but Has Little Impact on Parasite Motility. mSphere 2021, 6. [Google Scholar] [CrossRef]
- Díaz-Martín, R.D.; Mercier, C.; de León, C.T.G.; González, R.M.; Pozos, S.G.; Ríos-Castro, E.; García, R.A.; Fox, B.A.; Bzik, D.J.; Flores, R.M. The dense granule protein 8 (GRA8) is a component of the sub-pellicular cytoskeleton in Toxoplasma gondii. Parasitol. Res. 2019, 118, 1899–1918. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.; Ge, C.-C.; Fan, Y.-M.; Jin, Q.-W.; Shen, B.; Huang, S.-Y. The determinants regulating Toxoplasma gondii bradyzoite development. Front. Microbiol. 2022, 13, 1027073. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-L.; Li, T.-T.; Elsheikha, H.M.; Liang, Q.-L.; Zhang, Z.-W.; Wang, M.; Sibley, L.D.; Zhu, X.-Q. The protein phosphatase 2A holoenzyme is a key regulator of starch metabolism and bradyzoite differentiation in Toxoplasma gondii. Nat. Commun. 2022, 13, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Pamukcu, S.; Cerutti, A.; Bordat, Y.; Hem, S.; Rofidal, V.; Besteiro, S. Differential contribution of two organelles of endosymbiotic origin to iron-sulfur cluster synthesis and overall fitness in Toxoplasma. PLOS Pathog. 2021, 17, e1010096. [Google Scholar] [CrossRef] [PubMed]
- Cerutti, A.; Blanchard, N.; Besteiro, S. The Bradyzoite: A Key Developmental Stage for the Persistence and Pathogenesis of Toxoplasmosis. Pathogens 2020, 9, 234. [Google Scholar] [CrossRef] [PubMed]
- Sandeep Srivastava, M.W.W., William J. Sullivan, Jr., Toxoplasma gondii AP2XII-2 Contributes to Proper Progression through S-Phase of the Cell Cycle. mSphere, 2020. 2020 Sep-Oct; 5(5): e00542-20.
- Chen, Y.; Yang, L.-N.; Cheng, L.; Tu, S.; Guo, S.-J.; Le, H.-Y.; Xiong, Q.; Mo, R.; Li, C.-Y.; Jeong, J.-S.; et al. Bcl2-associated Athanogene 3 Interactome Analysis Reveals a New Role in Modulating Proteasome Activity. Mol. Cell. Proteom. 2013, 12, 2804–2819. [Google Scholar] [CrossRef] [PubMed]
- Tomita, T.; Bzik, D.J.; Ma, Y.F.; Fox, B.A.; Markillie, L.M.; Taylor, R.C.; Kim, K.; Weiss, L.M. The Toxoplasma gondii Cyst Wall Protein CST1 Is Critical for Cyst Wall Integrity and Promotes Bradyzoite Persistence. PLOS Pathog. 2013, 9, e1003823. [Google Scholar] [CrossRef]
- Souza, R.O.O., et al., IMC10 and LMF1 mediate mitochondrial morphology through mitochondrion-pellicle contact sites in Toxoplasma gondii. Journal of Cell Science, 2022. 135(22). [CrossRef]
- Koreny, L., et al., Molecular characterization of the conoid complex in Toxoplasma gondii reveals its conservation in all apicomplexans, including Plasmodium species. Plos Biology, 2021. 19(3). [CrossRef]
- Nelson, M.M.; Jones, A.R.; Carmen, J.C.; Sinai, A.P.; Burchmore, R.; Wastling, J.M. Modulation of the Host Cell Proteome by the Intracellular Apicomplexan Parasite Toxoplasma gondii. Infect. Immun. 2008, 76, 828–844. [Google Scholar] [CrossRef]
- Michael, C. Gretes, L.B.P., P. Andrew Karplus. Peroxiredoxins in Parasites. Antioxid Redox Signal. Antioxid Redox Signal, 2012. 2012 Aug 15; 17(4): 608–633. [CrossRef]
- Tan, Q.; Wang, J.; Chen, J.; Liu, X.; Chen, X.; Xiao, Q.; Li, J.; Li, H.; Zhao, X.; Zhang, X. Involvement of Urm1, a Ubiquitin-Like Protein, in the Regulation of Oxidative Stress Response of Toxoplasma gondii. Microbiol. Spectr. 2022, 10, e0239421. [Google Scholar] [CrossRef]
- Müller, J.; Schlange, C.; Heller, M.; Uldry, A.-C.; Braga-Lagache, S.; Haynes, R.K.; Hemphill, A. Proteomic characterization of Toxoplasma gondii ME49 derived strains resistant to the artemisinin derivatives artemiside and artemisone implies potential mode of action independent of ROS formation. Int. J. Parasitol. Drugs Drug Resist. 2022, 21, 1–12. [Google Scholar] [CrossRef]
- Zhou, D.-H.; Wang, Z.-X.; Zhou, C.-X.; He, S.; Elsheikha, H.M.; Zhu, X.-Q. Comparative proteomic analysis of virulent and avirulent strains of Toxoplasma gondii reveals strain-specific patterns. Oncotarget 2017, 8, 80481–80491. [Google Scholar] [CrossRef] [PubMed]
- Harding, C.R.; Sidik, S.M.; Petrova, B.; Gnädig, N.F.; Okombo, J.; Herneisen, A.L.; Ward, K.E.; Markus, B.M.; Boydston, E.A.; Fidock, D.A.; et al. Genetic screens reveal a central role for heme metabolism in artemisinin susceptibility. Nat. Commun. 2020, 11, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.X., et al., Global iTRAQ-based proteomic profiling of Toxoplasma gondii oocysts during sporulation. Journal of Proteomics, 2016. 148: p. 12-19. [CrossRef]
- He, J.-J.; Ma, J.; Elsheikha, H.M.; Song, H.-Q.; Zhou, D.-H.; Zhu, X.-Q. Proteomic Profiling of Mouse Liver following Acute Toxoplasma gondii Infection. PLOS ONE 2016, 11, e0152022–e0152022. [Google Scholar] [CrossRef] [PubMed]
- Moog, D.; Przyborski, J.M.; Maier, U.G. Genomic and Proteomic Evidence for the Presence of a Peroxisome in the Apicomplexan Parasite Toxoplasma gondii and Other Coccidia. Genome Biol. Evol. 2017, 9, 3108–3121. [Google Scholar] [CrossRef] [PubMed]
- Nie, L.-B.; Liang, Q.-L.; Elsheikha, H.M.; Du, R.; Zhu, X.-Q.; Li, F.-C. Global profiling of lysine 2-hydroxyisobutyrylome in Toxoplasma gondii using affinity purification mass spectrometry. Parasitol. Res. 2020, 119, 4061–4071. [Google Scholar] [CrossRef] [PubMed]
- Xia, N.; Guo, X.; Guo, Q.; Gupta, N.; Ji, N.; Shen, B.; Xiao, L.; Feng, Y. Metabolic flexibilities and vulnerabilities in the pentose phosphate pathway of the zoonotic pathogen Toxoplasma gondii. PLOS Pathog. 2022, 18, e1010864. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, E.; Clark, M.; Goulart, C.; Steinhöfel, B.; Tjhin, E.T.; Gross, S.; Smith, N.C.; Kirk, K.; van Dooren, G.G. Substrate-mediated regulation of the arginine transporter of Toxoplasma gondii. PLOS Pathog. 2021, 17, e1009816. [Google Scholar] [CrossRef] [PubMed]
- Wallbank, B.A.; Dominicus, C.S.; Broncel, M.; Legrave, N.; Kelly, G.; MacRae, J.I.; Staines, H.M.; Treeck, M. Characterisation of the Toxoplasma gondii tyrosine transporter and its phosphorylation by the calcium-dependent protein kinase 3. Mol. Microbiol. 2018, 111, 1167–1181. [Google Scholar] [CrossRef] [PubMed]
- Lang, D.; Schott, B.H.; van Ham, M.; Morton, L.; Kulikovskaja, L.; Herrera-Molina, R.; Pielot, R.; Klawonn, F.; Montag, D.; Jänsch, L.; et al. Chronic Toxoplasma infection is associated with distinct alterations in the synaptic protein composition. J. Neuroinflammation 2018, 15, 1–19. [Google Scholar] [CrossRef]
- Daher, D.; Shaghlil, A.; Sobh, E.; Hamie, M.; Hassan, M.E.; Moumneh, M.B.; Itani, S.; El Hajj, R.; Tawk, L.; El Sabban, M.; et al. Comprehensive Overview of Toxoplasma gondii-Induced and Associated Diseases. Pathogens 2021, 10, 1351. [Google Scholar] [CrossRef]
- Mayoral, J., P. Shamamian, Jr., and L.M. Weiss, In Vitro Characterization of Protein Effector Export in the Bradyzoite Stage of Toxoplasma gondii. mBio, 2020. 11(2). [CrossRef]
- Gay, G., et al., Toxoplasma gondii TgIST co-opts host chromatin repressors dampening STAT1-dependent gene regulation and IFN-gamma-mediated host defenses. J Exp Med, 2016. 213(9): p. 1779-98. [CrossRef]
- Alex W Chan, M.B., Nicole Haseley, et al., Analysis of CDPK1 targets identifies a trafficking adaptor complex that regulates microneme exocytosis in Toxoplasma. bioRxiv, 2023. 2023 Jan 12. [CrossRef]
- Uboldi AD, M.J., Blume M, Gerlic M, Ferguson DJ, et al., Regulation of Starch Stores by a Ca(2+)-Dependent Protein Kinase Is Essential for Viable Cyst Development in Toxoplasma gondii. Cell Host Microbe, 2015. 18(6):670-81. [CrossRef]
- Nofal, S.D.; Dominicus, C.; Broncel, M.; Katris, N.J.; Flynn, H.R.; Arrizabalaga, G.; Botté, C.Y.; Invergo, B.M.; Treeck, M. A positive feedback loop mediates crosstalk between calcium, cyclic nucleotide and lipid signalling in calcium-induced Toxoplasma gondii egress. PLOS Pathog. 2022, 18, e1010901. [Google Scholar] [CrossRef]
- Tang, Q.; Andenmatten, N.; Triana, M.A.H.; Deng, B.; Meissner, M.; Moreno, S.N.J.; Ballif, B.A.; Ward, G.E. Calcium-dependent phosphorylation alters class XIVa myosin function in the protozoan parasite Toxoplasma gondii. Mol. Biol. Cell 2014, 25, 2579–2591. [Google Scholar] [CrossRef] [PubMed]
- Brossier, F.; Starnes, G.L.; Beatty, W.L.; Sibley, L.D. Microneme Rhomboid Protease TgROM1 Is Required for Efficient Intracellular Growth of Toxoplasma gondii. Eukaryot. Cell 2008, 7, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, S.; Deng, B.; del Rio, R.; Buchholz, K.R.; Treeck, M.; Urban, S.; Boothroyd, J.; Lam, Y.-W.; Ward, G.E. Not a Simple Tether: Binding of Toxoplasma gondii AMA1 to RON2 during Invasion Protects AMA1 from Rhomboid-Mediated Cleavage and Leads to Dephosphorylation of Its Cytosolic Tail. mBio 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Child, M.A.; Bogyo, M. Proteases as regulators of pathogenesis: Examples from the Apicomplexa. Biochim. et Biophys. Acta (BBA) - Proteins Proteom. 2012, 1824, 177–185. [Google Scholar] [CrossRef]
- Luo, Q.; Upadhya, R.; Zhang, H.; Madrid-Aliste, C.; Nieves, E.; Kim, K.; Angeletti, R.H.; Weiss, L.M. Analysis of the glycoproteome of Toxoplasma gondii using lectin affinity chromatography and tandem mass spectrometry. Microbes Infect. 2011, 13, 1199–1210. [Google Scholar] [CrossRef]
Figure 1.
Timeline of the Main Events Associated with Proteomic Studies. The vertical lines indicate early, middle, or late events depending on their position. Examples of some events: 1, Stable isotope labeling by amino acids (SILAC), Isotope-Coded Affinity Tag (ICAT); 2, SEQUEST, Mascot, and Scaffold; 3, UniProt, National Institutes of Health database; 4, phosphoproteome, acetylome, ubiqutinome, etc.; 5, Orbitrap, Q-TOF; 6, Hydrogen Deuterium Exchange (HDX)-MS; 7, Skyline, MaxQuant. iTRAQ: Isobaric Tags for Relative and Absolute Quantitation. TMT: Tandem Mass Tags. FAIMS: Field Asymmetric Ion Mobility Spectrometry. DIA: Data-independent acquisition of molecular structure determination. The decades are not in scale.
Figure 1.
Timeline of the Main Events Associated with Proteomic Studies. The vertical lines indicate early, middle, or late events depending on their position. Examples of some events: 1, Stable isotope labeling by amino acids (SILAC), Isotope-Coded Affinity Tag (ICAT); 2, SEQUEST, Mascot, and Scaffold; 3, UniProt, National Institutes of Health database; 4, phosphoproteome, acetylome, ubiqutinome, etc.; 5, Orbitrap, Q-TOF; 6, Hydrogen Deuterium Exchange (HDX)-MS; 7, Skyline, MaxQuant. iTRAQ: Isobaric Tags for Relative and Absolute Quantitation. TMT: Tandem Mass Tags. FAIMS: Field Asymmetric Ion Mobility Spectrometry. DIA: Data-independent acquisition of molecular structure determination. The decades are not in scale.
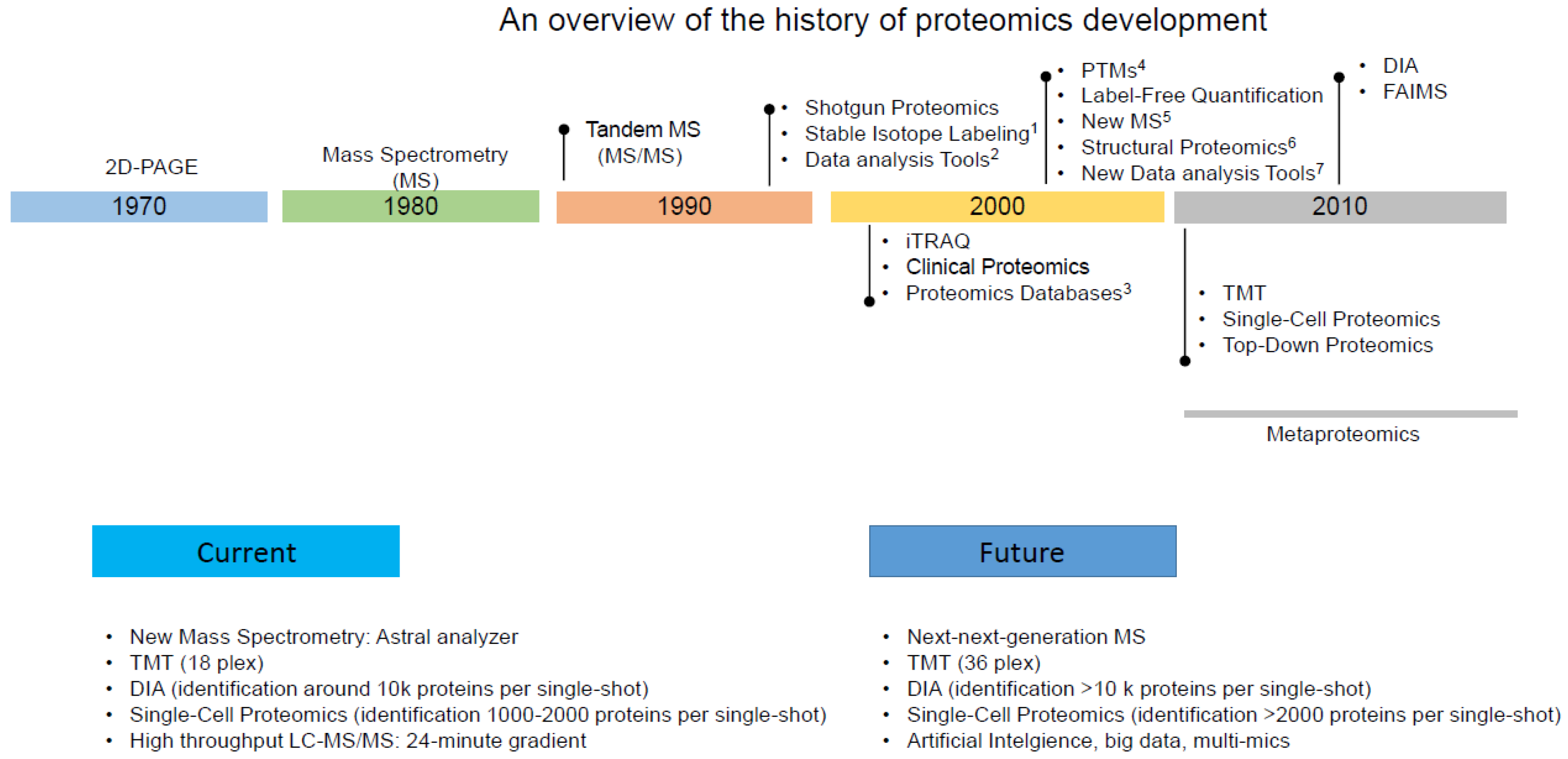
Figure 2.
Summary of Proteomics and Biological Processes Searching Results in ToxoDB. A) T.gondii proteomics research counts in ToxoDB; B) Search results of typical biological processes of T.gondii in ToxoDB.
Figure 2.
Summary of Proteomics and Biological Processes Searching Results in ToxoDB. A) T.gondii proteomics research counts in ToxoDB; B) Search results of typical biological processes of T.gondii in ToxoDB.
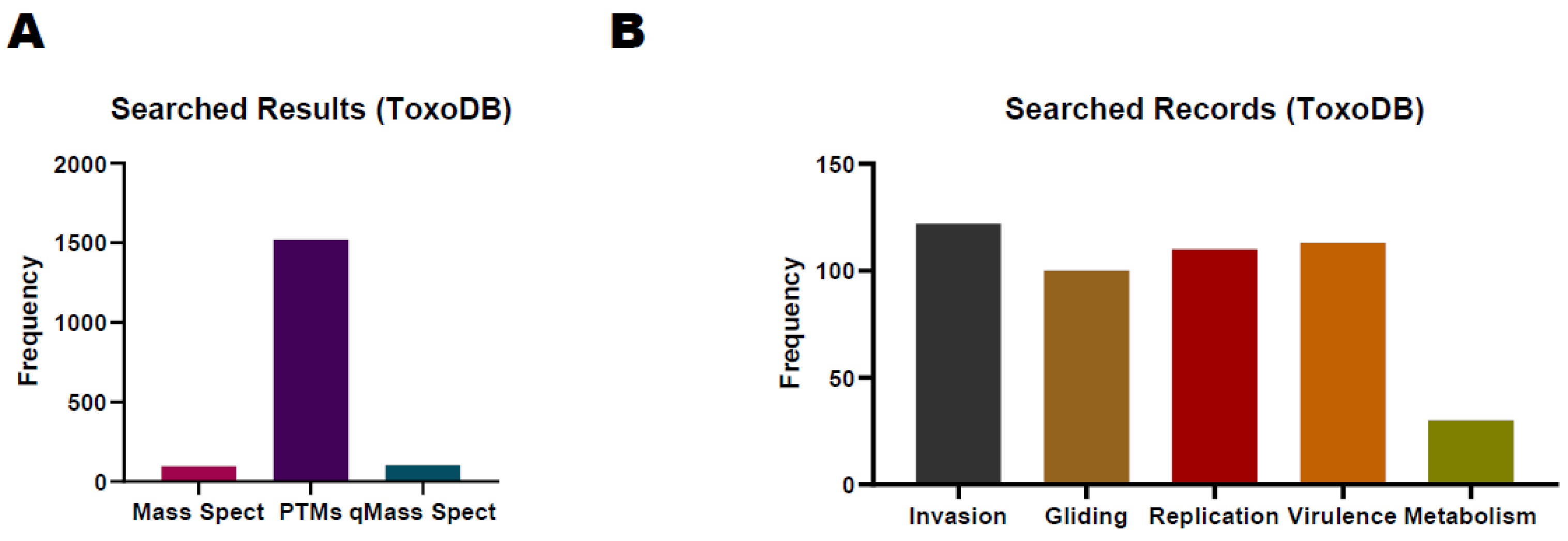
Figure 3.
Histone PTMs Crosstalk. A) Mammals and other eukaryotes: schematic representation of a nucleosome particle with the N-terminal tails of H3 and H4, and C-terminal tails of H2A and H2B histones. PTMs are represented in colored symbols as specified in the box. Crosstalks that have been studied are shown in blue arrows when one PTM activates another and in red lines when it is repressive. B) Schematic representation of a Toxoplasma gondii nucleosome particle, where canonical H3, H4, H2A, and H2B are shown as in A. H2A.Z and H2B.Z T. gondii C-terminal tails are represented in grey letters with the lysine residues, which were shown to be ubiquitinated, although H2A and H2B canonical ubiquitination are also conserved. Amino acidic residues are shown in black letters when conserved in T. gondii, while novel residues are shown in grey letters. The PTMs that are conserved in T. gondii follow the same color key as in A, while the differential Toxoplasma PTMs are shown in the box entitled “Toxo new PTMs” (the same for C). Although not much work has been conducted on T. gondii crosstalks, many of them can be inferred because of the conservation of the residues, and are marked in dotted lines. When a different residue may be involved in the crosstalk, but has not been proved, it is represented with dotted lines plus a question mark. New T. gondii crosstalks that have been observed are shown in green lines. C) Schematic representation of a Toxoplasma gondii nucleosome particle in which canonical H2A/H2B histones have been replaced by the H2A.Z/H2B.Z DVN, where the N-terminal tails with the PTMs are represented. Only the PTMs in H3 that have been observed to crosstalk with this DVN are shown.
Figure 3.
Histone PTMs Crosstalk. A) Mammals and other eukaryotes: schematic representation of a nucleosome particle with the N-terminal tails of H3 and H4, and C-terminal tails of H2A and H2B histones. PTMs are represented in colored symbols as specified in the box. Crosstalks that have been studied are shown in blue arrows when one PTM activates another and in red lines when it is repressive. B) Schematic representation of a Toxoplasma gondii nucleosome particle, where canonical H3, H4, H2A, and H2B are shown as in A. H2A.Z and H2B.Z T. gondii C-terminal tails are represented in grey letters with the lysine residues, which were shown to be ubiquitinated, although H2A and H2B canonical ubiquitination are also conserved. Amino acidic residues are shown in black letters when conserved in T. gondii, while novel residues are shown in grey letters. The PTMs that are conserved in T. gondii follow the same color key as in A, while the differential Toxoplasma PTMs are shown in the box entitled “Toxo new PTMs” (the same for C). Although not much work has been conducted on T. gondii crosstalks, many of them can be inferred because of the conservation of the residues, and are marked in dotted lines. When a different residue may be involved in the crosstalk, but has not been proved, it is represented with dotted lines plus a question mark. New T. gondii crosstalks that have been observed are shown in green lines. C) Schematic representation of a Toxoplasma gondii nucleosome particle in which canonical H2A/H2B histones have been replaced by the H2A.Z/H2B.Z DVN, where the N-terminal tails with the PTMs are represented. Only the PTMs in H3 that have been observed to crosstalk with this DVN are shown.
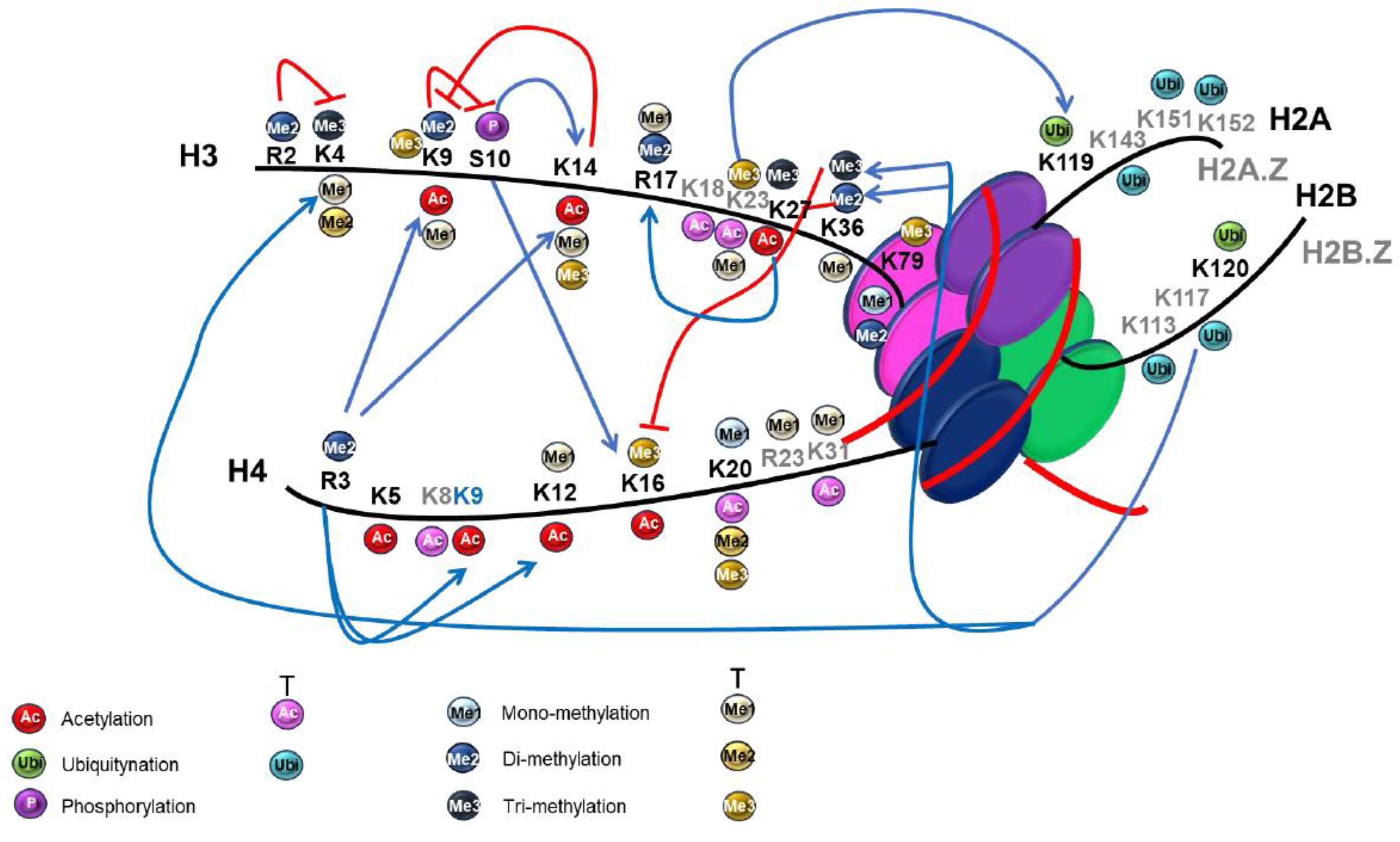

Table 1.
Proteomics Applications in T. gondii Research.
| Protein Group | Functions | Selected References |
|---|---|---|
| Microneme Protein (MIC) | Host cell adhesion and invasion are critical for the efficient invasion and attachment of T. gondii to host cells | 26, 29, 124, 163, 172, 175-181 |
| Rhoptry Proteins (ROP) | Host cell invasion and manipulation | 3, 4, 26, 29, 93, 124, 147, 175, 178, 182-190 |
| Surface Antigen (SAG) Proteins | Initial host cell invasion | 27, 87, 186, 191-194 |
| Dense Granule Antigens (GRA) | Modulation of host cell processes, immune evasion, and nutrient acquisition | 26, 29, 70, 73, 87, 147, 149, 175, 184, 195-206 |
| Rhoptry Neck (RON) Proteins | Manipulating host cell signaling pathways, and interacting with host cellular components during invasion | 180, 182, 207, 208 |
| Cytoskeletal Proteins | Facilitating the movement of the parasite and the rearrangement of the host cell’s cytoskeleton | 23, 71, 72, 172, 191, 209-214 |
| Bradyzoite-Specific Proteins | Parasite persistence | 28, 85, 181, 215-220 |
| Cyst Wall/Sporozoite-specific (CST) Proteins | Cyst wall of the parasite’s tissue-cyst stage | 74, 149, 221 |
| Inner Membrane Complex (IMC) Proteins | Host cell invasion | 69, 191, 222, 223 |
| Metabolic Enzymes | Parasite’s survival and replication | |
| · Antioxidant Enzymes | Counteracting oxidative stress generated by the host’s immune response | 224-226 |
| · Energy Production and Glycolysis | Parasite energy production | 124, 152, 173, 227 |
| · Krebs Cycle | Generating energy and providing essential metabolites for the parasite’s survival and growth | 124, 228-230 |
| · Fatty Acid Metabolism | Breakdown, synthesis, and regulation of fatty acids within cells | 57, 231, 232 |
| · Gluconeogenesis | Synthesis of glucose from non-carbohydrate precursors | 66, 193, 233 |
| · Pentose Phosphate Pathway | Generates nicotinamide adenine dinucleotide phosphate (NADPH) | 234 |
| · Amino Acid Metabolism | Protein synthesis and various metabolic processes | 83, 235, 236 |
| · Phosphatases | Catalyze the removal of phosphate groups from target proteins, counteracting the actions of kinases | 201, 150, 216 |
| Synaptic Proteins | Influence host neuronal function and potentially modulate synaptic activity | 237, 238 |
| T. gondiiIntraNuclear Shuttle Protein (TgIST) | Facilitating the movement of certain proteins into and out of the nucleus | 239, 240 |
| Ca2+-responsive Proteins | Translating Ca2+ signals into specific cellular responses | 34, 39, 241, 242, 243, 88, 244 |
| Rhomboid Proteases | Release of extracellular domains from the membrane and impact various signaling pathways | 245, 246, 247 |
| Heat Shock Proteins (HSP) | Maintenance of survival, growth, and adaptation of T. gondii within its host environment | 61, 62, 187, 193, 248 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



