Submitted:
12 November 2023
Posted:
13 November 2023
Read the latest preprint version here
Abstract
The Galápagos archipelago is a vast reservoir of terrestrial and marine biodiversity, owing in large part to its relatively recent volcanic genesis and colonization by humans. This unique ecological system is particularly susceptible to human, animal, and environmental impacts. Climate change, globalization, and the blurring of human-domestic animal-wildlife interfaces are poised to bring new threats and challenges to the region. A One Health perspective that simultaneously considers human, animal, and environmental health is imperative in assessing and mitigating the challenges facing the Galápagos Islands.In Part I of this review, we provide the historical context for biodiversity in the archipelago; discuss the role of invasive species in habitat destruction, fragmentation, and competition with endemic species; and summarize the established and emerging infectious disease threats. We also discuss the imperative to implement research, surveillance, and preventative measures to identify and manage future threats from a One Health perspective, with a specific emphasis on implications for wildlife health. In Part II, we outline the socioeconomic context of life in the Galápagos Islands, evaluate the current and predicted effects of climate change, and discuss direct anthropogenic factors affecting Galápagos biodiversity, such as tourism, fishing, pollution, and the illegal wildlife trade. We also examine the impact of the COVID-19 pandemic in the region. Throughout this two-part review, we build a cohesive picture of One Health in the Galápagos Islands by integrating past work, current needs, and emerging threats. We also consider overarching goals for conservation, ecosystem management, and socioeconomic sustainability that have been previously defined by both governmental and non-governmental stakeholders, and we identify discrete, implementable, and interdisciplinary recommendations that will facilitate achievement of those goals.
Keywords:
Galapagos
; one health
; planetary health
; wildlife
; conservation
; endemic species
; invasive species
1. Introduction
1.1. Historical Context of Galapagos Biodiversity
Located 960 kilometers west of Ecuador, the Galápagos Islands are a vast reservoir of terrestrial and marine biodiversity, home to over 6,000 species, 1,870 of which are endemic (CDF, Species Checklist). “Endemic” species are those limited to a small geographic region: in layman’s terms, “found nowhere else.” The Galápagos Islands have one of the highest rates of endemism in the world, including 22% of birds, 72% of reptiles, 38% of terrestrial mammals, 7% of marine mammals, and 12% of fish (CDF, Species Checklist). The origin and conservation status of Galápagos vertebrates are summarized in Table 1.
The archipelago’s relatively recent volcanic genesis—60,000 years young in the case of Fernandina Island (Tye et al., 2002)—has influenced the composition of its flora and fauna. Because the land mass was never attached to a continent, it did not inherit complex soil communities. The ancestors of all Galápagos wildlife arrived by flying, swimming, or floating, or evolved from those that did. Isolated habitats and diverse ecosystems also provided selective pressures for species diversification. The Galápagos mockingbird (Mimus parvulus), for example, diverged into four subspecies, and their striking morphological differences are thought to have inspired Charles Darwin’s theory of evolution (Arbogast et al., 2006).
The Galápagos Islands are also unique, even among oceanic islands, in that there has been relatively little time for humans to impart change. The archipelago was first discovered in 1535, but apart from serving as an occasional mooring for pirates and whalers since the mid-1700s, was not colonized until 1832 (Smith, 1979). “The Galápagos are a rare remnant of a prehistorical pattern of global biological diversity where great proportions of the world’s distinctive and often bizarre species occurred on islands,” wrote Charles Darwin Foundation (CDF) researchers in 2002. “Man has destroyed much of that pattern. The biological diversity of the Galápagos is one of the best examples of that pattern because it remains,” (Tye et al., 2022).
Nonetheless, human exploitation of endemic wildlife and extractive use of natural resources have long been a part of the archipelago’s history. Whalers touted the Galápagos giant tortoise as a source of fresh meat, taking aboard an estimated 200,000 tortoises between 1800-1870 (Townsend, 1925). Mariners also intentionally introduced goats, pigs, and donkeys. Black rats and house mice reached the archipelago as stowaways on ships. In 1832, Ecuador claimed the Galápagos Islands from Spain and the first settlers, a prison colony, arrived on Floreana Island (Smith, 1979). Between 1860 and 1930, further colonies were established for sugarcane farming, coffee crops, and salt mining. By that time, horses, cattle, dogs, and cats had also been introduced. These invasive species destroyed vegetation, predated nests, and competed with endemic species for resources (MacFarland et al., 1974). The decimation of the giant tortoise population through hunting and nest predation, combined with slow reproduction, led to extinction of two subspecies and near extinction of the other twelve (MacFarland et al., 1974), from which modern populations have never fully recovered.
In considering the historical narrative of the Galápagos Islands, the long and complicated relationship with foreign scientists cannot be ignored. In 1835, Charles Darwin documented biodiversity in the region through 1,500 biologic specimens. For the next century, researchers were fascinated by the novel flora and fauna and even began to recognize threats imposed by whalers and invasive species (Dumbacher & West, 2010). However, their solution was preservation of endemic species not in their natural habitat, but in the form of museum specimens. Between 1897 and 1905, American scientist Rollo Beck collected over 13,000 insects, 11,500 birds, 10,000 plants, and 5,000 reptiles, including the last remaining giant tortoise of Fernandina Island (Gifford, 1908; Dumbacher & West, 2010). G. T. Corley Smith, former secretary-general of the CDF, wrote in 1979, “Conservation was a concept virtually unknown to their generation. Scientists simply accepted that the Galápagos fauna was doomed to extinction and that their duty to posterity was to preserve as much as they could in museums.” Beck’s collection was instrumental in characterizing unique endemic species (Dumbacher & West, 2010), and contributes towards studies evolution even a century later (Tonnis et al., 2005). But despite the research value of these specimens, their method of acquisition—collecting nesting individuals and eggs, or in some cases, capture of every single individual seen—doubtlessly contributed to population declines still faced by these species today. Thankfully, scientists in the 1930s began to recognize the importance of conservation and shifted their attitudes and practices. Today, the Galápagos National Park Directorate (GNPD) requires scientists to acquire research permits prior to specimen collection, with stringent review of the scientific justification and soundness of proposed research methodology, to ensure that any use of biological materials is likely to result in meaningful research outcomes.
The Galápagos Islands are still faced with the challenge of “helicopter research,” in which foreign scientists with considerable resources enter developing regions, collect samples, publish data without inclusion of local scientists, and then leave (Adame, 2021; Minasny et al., 2020; Chin et al., 2019). Too often, research methods and results are not applicable or reproducible for local stakeholders, hindering long-term benefits. Conversely, there still exist barriers to dissemination of results of locally performed research. For instance, Barnett and Rudd first documented canine heartworm (Dirofilaria immitis) microfilariae in Galápagos sea lions (Zalophus wollebaeki) on Floreana Island in the 1980s (Barnett & Rudd, 1983; Barnett, 1985a). This publication, written in Spanish and held in print at the Charles Darwin Research Station, is largely inaccessible to the larger scientific community, and thus, a 2023 publication claims itself to be the first report of D. immitis infection in Galápagos sea lions (Gregory et al., 2023).
Ultimately, research efforts by international scientists must strive to produce tangible and direct regional benefits, and integrate solutions for preservation of biodiversity while prioritizing participation by residents and supporting local development. Scientists must prioritize ethical research practices that consider the ecological impacts of specimen collection, appropriately compensate and acknowledge local collaborators and participants, utilize methods that can be replicated using local resources, and involve reciprocal exchange of knowledge. Barriers to information-sharing must be addressed by expanding scientific journal access to avoid information gatekeeping, and establishing digital repositories and translations of print literature to prevent their loss, inaccessibility, or unintentional disregard.
1.2. Recognition of threats to Galápagos ecosystems
In 1959, in response to rising tourism and immigration and concern over exploitation of natural resources, the Galápagos National Park (GNP) was established to protect 96% of the land mass, with the remaining 4% designated as human-inhabited areas across the four populated islands: Santa Cruz, San Cristóbal, Floreana, and Isabela. In 1978, the Galápagos Islands and surrounding waters were designated as a UNESCO World Heritage site. Under the Special Law for Galápagos in 1998, 133,000 square-kilometers of marine, coastal, and inland waters were designated as the Galápagos Marine Reserve (GMR).
Prompted by ongoing and emerging threats to biodiversity, in 1999, the CDF and the World Wildlife Fund (WWF) conducted a joint biological analysis of the region. The resulting Biodiversity Vision for Galápagos summarized the ecological status of the islands, identified species most threatened by anthropogenic factors, and outlined a vision for conservation—including recommendations to mitigate biodiversity threats, and benchmarks by which to measure progress (Bensted-Smith, 2002). At that time, the archipelago retained 95% of its terrestrial biodiversity as compared to pre-human colonization. However, pressures from immigration, tourism, and industrial fishing were mounting, and along with them, the prospect of ecological degradation and consequent decline in biodiversity. Biologists feared that Galápagos flora and fauna might be at a precipice, and that failing to intervene could be disastrous, so much so that the workshop goals were summarized as: “Back to Eden—one last chance.”
The terrestrial and marine environments of the Galápagos Islands are subject to distinct, but sometimes overlapping, pressures. As defined by Snell et al. (2002), the primary threat to marine biodiversity is the exploitation of key species through fishing, poaching, or accidental killing, compounded by the effects of climate change and invasive species. Terrestrial ecosystems are most threatened by invasive species and associated predation, competition, and habitat alteration (Snell et al., 2002). Natural and anthropogenic changes were summarized by Snell et al. (2002) as reproduced in Table 2.
1.3. Impetus for the current review
Over the past 25 years, the Galápagos Islands have continued to experience a steady rise in tourism, illegal immigration, introduction of invasive species, and overfishing, as well as insufficient resources for local agencies to effectively manage these issues, leading to lack of implementation of quarantine, inspection, and biosecurity regulations (UNESCO, 2006). The GNPD has also documented tremendous pressures on ecosystem services and increasing demand for resources, making current conditions unsustainable (DPNG, 2014). Factors currently affecting ecosystem services include changes in land use, introduction of invasive species, loss of biodiversity, habitat fragmentation and degradation, overfishing, and stone materials overexploitation. Water resource degradation is a compounding factor, especially in wetland ecosystems, as well as a marine pollution and resource deterioration due to the demand for infrastructure for fishing, tourism, transportation, and maritime activities (DPNG, 2014).
These concerns led to the inclusion of the Galápagos Islands on the UNESCO List of World Heritage in Danger from 2007-2010 (UNESCO, 2006; UNESCO, 2007). In October 2012, the Agency for the Regulation and Control of Biosecurity and Quarantine for Galápagos (ABG) was established to prevent phyto/zoo-sanitary threats, and to control and reduce the risk of invasive species in the Galápagos Islands. In general, the import of non-native plants and animals are prohibited. ABG oversees prevention and early detection of invasive species and infectious disease threats through various surveillance mechanisms, and where necessary, develops strategies for control and eradication. Previously, these responsibilities were distributed among various national and regional institutions (Bensted-Smith et al., 2002); centralizing of these responsibilities under ABG has improved the efficiency of management, surveillance, and research. However, optimal implementation and maintenance of these systems is still hindered by periodic lapses in funding, personnel, and equipment (Rogg et al., 2005).
Only one extinction has been documented in the Galápagos Islands in the past 30 years: the Pinta Island tortoise (Chelonoidis abingdonii), of which the famous final member, Lonesome George, passed away in 2012. The most recent prior extinctions were the San Cristóbal Vermilion Flycatcher (Pyrocephalus dubius), last sighted in 1987, and the Floreana Island tortoise (C. niger), hunted to extinction around 1850. In total, nine Galápagos species are extinct, all formerly endemic (Table 3). There is clear evidence that population dynamics of many endemic Galápagos species have also changed profoundly over the past decades. Declines in multiple finch species on Floreana Island have been documented by several groups since 1979 (Grant et al., 2005; O’Connor et al., 2010; Dvorak et al., 2017; Dvorak et al., 2011). Per on a 2015-16 survey of endemic birds on Floreana Island, the Galápagos rail (Laterallus spilonota), vegetarian finch (Platyspiza crassirostris), and gray warbler finch (Certhidea fusca) were extirpated (locally extinct), and the Galápagos dove (Zenaida galapagoensis) was at risk (Dvorak et al., 2017). Populations of the endangered Galápagos sea lion have declined approximately 50% in the past 30 years (Riofrío-Lazo et al., 2017) due to human activities (Denkinger et al., 2014; Denkinger et al., 2015). As Galápagos ecosystems continue to suffer the pressures of climate change, invasive species, and resource limitation, further shifts in the relative composition of endemic species can be expected to occur. In addition, marine species, particularly invertebrates, are understudied.
Both the International Union for Conservation of Nature (IUCN) and the CDF monitor the population status of native and endemic species in the Galápagos archipelago. Overall, CDF recognizes 82 species as Critically Endangered (CR), 63 as Endangered (EN), 193 as Vulnerable (VU), and 87 has Near-Threatened (NT), while the IUCN Red List assesses only 61 Galápagos species as CR, 32 as EN, 95 as VU, and 25 as NT (IUCN, 2022). The at-risk Galápagos vertebrate species and their IUCN Red List statuses from 1994-2020 are summarized in Figure 1; the species included are listed in the Supplemental Tables.
The essentiality of the link between human, domestic animal, wildlife, and ecosystem health must be the lens through which we approach biodiversity preservation in the Galápagos Islands. As defined by the One Health High-Level Expert Panel, One Health “recognizes the health of humans, domestic and wild animals, plants, and the wider environment (including ecosystems) are closely linked and interdependent” (OHHLEP et al., 2022). One Health has often been approached through an anthropocentric lens, aiming to improve animal health in order to mitigate the effects that poor animal health can have on human populations, such as the emergence and transmission of zoonotic diseases (Fauci, 2006), or compromised food security. However, to truly apply a One Health perspective, we must also recognize the intrinsic importance of safeguarding animal and environmental health (Stephen et al., 2023). “Planetary Health” was proposed in 2015 as a field focused on how human activities, ranging from industrialization to food security, can compromise environmental health, and how these effects are then further reflected in public health consequences. Addressing interactions across these sectors, such as infectious disease, food and water sustainability, clean air and energy, and climate change, requires a sustainable balance between human, animal, and ecosystem health. The timeliness of this conversation could not be more apt in the wake of both the COVID-19 pandemic, with its emergence and propagation associated with human activities (Tounta et al., 2022), and the emergence of Highly Pathogenic Avian Influenza (HPAI) strain H5N1.
Here, we discuss the complex historical and socioeconomic contexts surrounding Galápagos biodiversity, revisit the pressures previously identified by Snell et al. (2002), and describe threats to biodiversity that have emerged over the past twenty-five years. Despite the large amount of work that has been accomplished in Galápagos conservation, we hold that there remains an imperative for further change that necessitates a One Health perspective.
2. Threats to Galápagos Biodiversity
Part I of this review describes the impacts of introduced plants, invertebrates, and vertebrates on native Galápagos species, and reviews established, emerging, and future infectious disease threats. Part II of this review will address climate change, ocean acidification, and direct human activities such as tourism, fishing, pollution, agriculture, and human-wildlife conflict.
2.1. Introduced species
Introduced species are non-native species that have been introduced to a given environment. While introductions can occur secondary to natural events, anthropogenic introductions are by far more common. Between 1990 and 2007, the documented introduced species in the archipelago increased ten-fold (Watkins & Cruz, 2007). As of 2017, 1,579 species were estimated to have been introduced to the Galápagos Islands, with 93% having become established (Toral-Granda et al., 2017). Pizzitutti et al. (2016) reported that in 2012, 2 of every 10 species observed were introduced, and predicted that by 2033, under a model that simulates continued high rates of growth, the proportion of introduced species could rise to 50% (Pizzitutti et al., 2016).
Invasive species are those introduced species that spread, often rapidly, beyond the area of introduction and have detrimental effects on human, animal, or environmental health. Introduced and invasive species can then become naturalized (integrated into the ecosystem and capable of maintaining their populations). Invasive species continue to be one of the most significant threats to endemic Galápagos flora and fauna.
2.1.1. Invasive plants
Nearly half of the species introduced to the Galápagos Islands were intentional introductions of plants, primarily for agricultural purposes; non-native plants now outnumber endemic flora (Toral-Granda et al., 2017; Guézou et al., 2010; Tye et al., 2006). Invasive plants can alter soil composition, water and light availability, nutrient cycling, and pollinator populations, impacting native plants as well as animals that rely on the ecosystem for food and shelter. In a review of IUCN Extinct species, alien plants were implicated as a cause for 25% of plant and 33% of animal extinctions (Blackburn et al., 2019). The blackberry shrub (Rubus niveus) is of the most widespread invasive plants in the Galápagos Islands and serves as a prime example of the vast economic and biodiversity impacts of introduced flora (Rentería et al., 2012). Originally introduced for agriculture on Santa Cruz and San Cristóbal Islands in the 1960s and 1970s, R. niveus has since spread to other islands and overgrown into vast thickets, displacing native flora and diminishing the suitability of the land for agriculture (Rentería et al., 2012). R. niveus has been associated with diminished richness of native plants and implicated in the altered structure of the Scalesia pedunculata forest, a key Galápagos ecosystem (Rentería et al., 2012).
The most common strategy for invasive plant eradication in the Galápagos Islands is mechanical removal followed by chemical control. Mechanical removal is labor-intensive, requires removal of the roots to prevent regrowth, and does not account for recurrence due to residual seeds or wind dispersal. Chemical control can deteriorate soil quality and have off-target effects on insects and vertebrates. Biological control of invasive species involves the release of a living organism—typically bacteria, fungi, or insects—that results in selective depopulation of the invasive species. The Australian ladybird beetle (Novius cardinalis), for instance, was introduced to the Galápagos Islands in 2002 to control the invasive cotton scale insect (Icerya purchasi), successfully reducing their populations (Calderón Alvaréz et al., 2012). The fungus Puccinia lantanae has been suggested as a potential biocontrol agent of Lantana camara, an invasive perennial shrub, with no documented effects on the related endemic Lantana pedicularis (Rentería & Ellison, 2004; Thomas et al., 2021). However, due to their often irreversible nature, stringent risk assessments must be conducted before implementing a biological control agents.
2.1.2. Invasive invertebrates
Unintentional introduction of invertebrates occurs primarily via hitchhiking on imported plants or produce, or on transport vehicles (e.g., boats or planes) (Toral-Granda et al., 2017). As of 2006, almost 500 insects and arthropods had been introduced to the archipelago (Causton et al., 2006). The southern house mosquito (Culex quinquefasciatus) and the yellow fever mosquito (Aedes aegypti) were introduced via airplanes (Bataille et al., 2009a; Bataille et al., 2009b) and both have become naturalized (Whiteman et al., 2005; Sinclair, 2017). These mosquitoes are competent vectors of a number of human and animal diseases, including West Nile Virus (WNV) (Kilpatrick et al., 2006; Sardelis et al., 2001), canine heartworm (Barnett, 1985a; Barnett, 1985b; Hendrix et al., 1986), and avian malaria (Plasmodium spp.) (van Riper et al., 1996; Harvey-Samuel et al., 2021) and thus pose risks for infectious disease introduction and establishment (Causton et al., 2006; Nishida & Evenhuis, 2000). A survey of Galápagos penguins (Spheniscus mendiculus) in 1996, shortly after C. quinquefasciatus is thought to have been introduced, identified no positive cases of avian malaria (Miller et al., 2001). However, in 2013, avian malaria was identified in Galápagos penguins (Palmer et al., 2013; Levin et al., 2013) and yellow warblers (Setophaga petechia) (Levin et al., 2013), likely transmitted to the archipelago’s mosquitos via migratory birds (Levin et al., 2013). Control of C. quinquefasciatus in the Galápagos Islands is primarily via fumigation of airplanes with insecticides and reduction of mosquito-attracting light sources in tourist areas (Harvey-Samuel et al., 2021).
The invasive parasitic bot fly, Philornis downsi, was introduced to the Galápagos Islands in 1964 (Causton et al. 2006) and has since become widespread (Fessl et al., 2018). Larvae laid in bird nests feed on hatchlings and cause severe morbidity and mortality (Causton et al., 2013). P. downsi has been associated with severe population declines in at least 16 species of Galápagos land birds, including the critically endangered mangrove finch (Fessl et al., 2018). The CDF and GNPD have ongoing multi-center collaborations to continue researching the biology and impacts of P. downsi and implement appropriate control strategies.
Several groups have reported molecular surveillance protocols to identify invasive arthropods from mixed-species samples on insect traps (Butterworth et al., 2022; Mee et al., 2021). In addition, eastern equine encephalitis virus, WNV, yellow fever virus, and dengue virus can be identified from mosquitoes via PCR (Hadfield et al., 2001; Ali et al., 2022). These strategies may prove useful in surveilling for new introduction events at airports or selected environmental sites.
2.1.3. Invasive vertebrates
Invasive vertebrates in the Galápagos Islands are responsible for habitat alteration, decline in vegetation diversity, predation, and resource competition with endemic species. Goats, pigs, donkeys, cattle, horses, rats, and mice were introduced by mariners and early settlers, and without natural predators, established large feral colonies. Donkeys were introduced in 1834 for transporting cargo; a half-century later, feral donkeys were widespread on multiple islands (Carrion et al., 2007). Feral goats, introduced in the 1920s-40s, are now responsible for significant habitat destruction via grazing, decimating native plants and accelerating erosion, incurring an estimated cost of 20 million US dollars between 1983 and 2017 (Ballesteros-Meija et al., 2021). Cattle and horses had similar impacts on vegetation, resulting in extirpation of native plant species (Bush et al., 2022), a factor which also contributes to takeover by invasive plants.
Following recognition of these impacts, conservation programs were implemented with the goal of eradicating invasive animals. In 1974, Santiago Island had an estimated 100,000 feral goats and 20,000 feral pigs (deVries & Black, 1983). Through a combination of land and aerial depopulation methods, goats and pigs were completely eradicated from Santiago Island by 2005 (Cruz et al., 2009; Cruz et al., 2005). Project Isabela was responsible for the depopulation of over 140,000 goats from Isabela Island (Carrion et al., 2011). Intensive conservation efforts also resulted in presumptive eradication of feral donkeys from Santiago Island and the Alcedo Volcano area of Isabela Island by 2005 (Carrion et al., 2007); pigeons from Santa Cruz, San Cristóbal, and Isabela Islands by 2005 (Phillips et al., 2012a); cats from Balta Island in 2003 (Phillips et al., 2012b); and fire ants (Wasmannia auropunctata) from Marchena Island in 2002 (Causton et al., 2005). In each case, negative surveillance for 1-2 years following the last known sighting led to conclusions that eradication was successful. However, care should be taken when interpreting a lack of sightings as conclusive evidence of eradication, particularly in the case of insects. Additionally, until a given feral species is entirely eradicated from the archipelago, there is an ever-present risk of reintroduction through anthropogenic interisland movement, and thus a need for ongoing surveillance and prevention.
While depopulation has mitigated the destruction caused by invasive vertebrates, much work remains. In particular, domestic dog overpopulation is a significant problem for endemic Galápagos species. Domestic dogs were first introduced to Floreana Island in 1832 and to San Cristóbal Island approximately a decade later (Barnett, 1986). Large feral dog colonies persisted on Isabela Island through the 1980s (Barnett and Rudd, 1983; Reponen et al., 2014). Canine nest predation of giant tortoises and land iguanas resulted in population declines that ultimately prompted nest conservation programs (Barnett, 1986). Canine predation upon the marine iguana, Galápagos penguin, Galápagos sea lion, Galápagos fur seal, blue pelican, blue-footed booby, and Audubon shearwater has also been reported (Barnett and Rudd, 1983; Kruuk and Snell, 1981). Depopulation efforts by the GNPD and CDF, principally via bait laced with toxic sodium monofluoroacetate (Compound 1080), led to drastic reduction in feral dog populations (Barnett and Rudd, 1983; Barnett, 1986). Nevertheless, free-roaming behavior in pet dogs is still common (Jimenez et al., 2020; Diaz et al., 2018; Gingrich et al., 2010). Free-roaming dogs may encounter wildlife, leading to predation, traumatic injury, or disease transmission (Barnett, 1985b). Controlling populations of domestic dogs thus remains a key focus. Spay and neuter campaigns have been ongoing over the past decade, but at their current level are still insufficient to mitigate population growth (Hernandez et al., 2020). As of 2018, the human:dog ratio was estimated to be 4:1 on Santa Cruz Island, a 55% increase in the dog population since 2014 (Hernandez et al., 2020). Re-establishment of truly feral canine colonies in remote regions is a low but present risk (Reponen et al., 2014).
Another prominent example of the persistent impacts of introduced vertebrates is that of the smooth-billed ani (Crotophaga ani), intentionally introduced in the 1960s as a form of agricultural control to predate upon cattle ticks (Cooke et al., 2019). The smooth-billed ani is considered the most damaging invasive bird in the Galápagos Islands, owing to its rapid population growth and predation of native plants and animals, particularly endemic birds (Connett et al., 2019), snakes (Cooke et al., 2020), and the endemic Galápagos carpenter bee, a major pollinator. The bird also propagates invasive plants by ingesting and spreading seeds (Cooke et al., 2019). Since 1980, the CDF has studied the impacts of this species and evaluated eradication techniques, but no large-scale control plan has been implemented to date, and the bird remains widespread.
2.1.4. Invasive marine species
Marine traffic from tourism, fishing, and cargo shipping is likely the major route by which marine invasive species enter the GMR (Keith et al., 2016; Carlton et al., 2019). Carlton et al. (2019) hypothesized that shoreline structures such as docks and buoys also facilitate colonization by marine species introduced via the hulls of ships. According to the CDF, at least 59 invasive marine species have been documented in the archipelago, including the green algae Caulerpa chemnitzia (CDF, 2022). Fast-growing algae can outcompete corals, particularly in already compromised areas; C. chemnitzia is thus a threat to Darwin Island’s Wellington Reef (CDF, 2022). Recent work by Keith et al. (2022) reported that C. chemnitzia populations grow and contract in response to climate change, calling for an early detection rapid response system to monitor marine invasive species.
Overall, marine invasive species, particularly invertebrates, have been understudied (Keith et al., 2016; Baert, 1994; Carlton et al., 2019). A baseline of the species in the GMR and modes of entry for invasive species must be defined to inform prevention and eradication strategies. In addition, the marine environment poses obvious challenges for endemic species definition. Edgar et al. (2004) identified three biogeographical areas with diverse compositions of fish and invertebrates and different levels of endemism, highlighting that relative species composition varies considerably between marine ecosystems.
2.2. Infectious diseases
Non-native hosts and vectors contribute to the introduction and establishment of infectious diseases in susceptible endemic populations. Introduced pathogens can establish natural reservoirs, circulating within wildlife populations without requiring reintroduction from a domestic animal host. Once wildlife reservoirs are established, disease eradication becomes increasingly complex, as strategies used in domestic animals—such as mass vaccination or depopulation—are more economically challenging, labor-intensive, and ethically controversial to implement in wildlife. Many of the pathogens described in this section remain under-researched in the Galápagos Islands; therefore, in many cases, the presence of wildlife reservoirs has yet to be confirmed.
Domestic dogs are one of the most prominent potential reservoirs of infectious diseases that could affect Galápagos wildlife. The dog population remains under-vaccinated and thus susceptible to infectious disease outbreaks. Canine vaccination was prohibited in the Galápagos Islands until 2017 (Levy et al., 2008; Vega-Mariño et al., 2023). While ABG has since promoted vaccination campaigns, efforts have not yet resulted in adequate herd immunity against common canine pathogens, such as Canine Distemper Virus (CDV) (Vega-Mariño et al., 2023) and canine parvovirus. For instance, an outbreak of CDV in 2001 resulted in the death of over 600 dogs and documented exposure of Galápagos sea lions (Levy et al., 2008). Due to lack of education and poor product availability, the use of adequate ectoparasite preventatives is also uncommon, leading domestic dogs to carry a high tick burden (Jimenez et al., 2020).
The poultry industry also presents a risk to the diverse endemic bird population. Food security and economic sustainability for human populations of oceanic islands require stable sources of locally-produced food. As of 2014, approximately 100,000 chickens and over 3,500 pigs were present within the archipelago, in backyard and commercial farms (Puente-Rodríguez et al., 2019). Four laying hen farms produce most of the archipelago’s egg supply (Puente-Rodríguez et al., 2019). Importation of unvaccinated day-old chicks, a practice common in the growing broiler industry and a key exemption to the prohibition on animal importation, could serve as a route of introduction of pathogens (Puente-Rodríguez et al., 2019). Pathogens such as Newcastle Disease Virus (NDV), Marek’s Disease Virus (MDV), Infectious Bronchitis Virus (IBV) and Mycoplasma spp. have been detected in poultry within the Galápagos Islands, and could spread to wildlife, particularly from backyard flocks with environmental and direct contact with wild birds (Gottdenker et al., 2005; Whitehead et al., 2018; Wikelski et al., 2004).
In Table 4, we report selected bacterial, viral, fungal, and parasitic pathogens of One Health importance to the Galápagos Islands, the risks of which are summarized in the following sections. We focus on pathogens that have been documented in domestic animal reservoirs and/or in endemic wildlife, and/or that have zoonotic potential. For those pathogens without confirmed cases in Galápagos wildlife, such as Coxiella burnetii, we assess risk based on the competency of these pathogens in related species in other geographic regions. In addition, we include several pathogens that have not been identified in the Galápagos Islands, such as WNV, but which are still considered high risk. We also assign each pathogen a Risk Level, based on a consideration of the potential for introduction, global status of the disease, susceptibility of Galápagos species, presence of wildlife reservoirs and vectors, and zoonotic potential. It should be noted that this list is not comprehensive. For example, ubiquitous species such as Escherichia coli and Salmonella enterica can be commensal or may cause clinical disease in mammals, birds, and reptiles, depending on various factors, such as pathogen strain and host immune status.
2.2.1. Viral pathogens of importance to the Galápagos Islands
Canine Distemper Virus (CDV) is a paramyxovirus that affects wild and domestic carnivores, but is not zoonotic (Martinez-Gutierrez & Ruiz-Saenz, 2016; Beineke et al., 2015). CDV is globally distributed and remains one of the leading causes of death in domestic dogs. Transmission of CDV from domestic dogs to wild carnivores, including the black-footed ferret (Williams et al., 1988), Amur tiger (Gilbert et al., 2020), and African wild dog (van de Bildt et al., 2002), has posed a major threat to conservation. CDV is also a documented emerging threat to the Galápagos sea lion. Outbreaks of CDV have occurred in domestic dogs in the Galápagos Islands (Vega-Mariño et al., 2023), and while an outbreak of CDV has not occurred in Galápagos sea lions to date, positive pups and adults having been identified (Levy et al., 2008; Denkinger at al., 2017), and CDV can cause morbidity and mortality in pinnipeds (Review: Kennedy et al., 2019). CDV can become established in wild pinniped populations, as previously reported in Antarctic seals (Bengtson et al., 1991), posing a barrier to disease eradication. Control of canine overpopulation and limitation of domestic animal-wildlife contact, vaccination campaigns, and continued surveillance of wildlife populations is necessary to prevent the spread of CDV to Galápagos wildlife (Vega-Mariño et al., 2023).
Phocine distemper virus (PDV) emerged in 1988, likely derived from CDV following contact between domestic dogs and seals. Several outbreaks of PDV decimated seal populations in Arctic and North Atlantic waters between 1980 and 2006 (Duignan et al., 2014; Härkönen et al., 2006; Earle et al., 2006; Kennedy et al., 2019). PDV was first identified in the Northern Pacific Ocean in 2004 in association with sea ice reduction, with evidence to suggest that marine mammals in that region were previously naïve to this virus (Goldstein et al., 2009; Kennedy et al., 2019; VanWormer et al., 2019). While PDV has yet to be identified in tropical or subtropical climates, including the Galápagos Islands, its continued spread and high mortality rate are concerning. Cross-reactivity between antibodies against CDV and PDV has been documented, therefore RT-PCR is the gold standard for differentiating these related viruses (Stanton et al., 2004).
Viral pathogens of domestic poultry can affect economic stability and threaten endemic birds. MDV, NDV, IBV, and Infectious Bursal Disease Virus (IBDV) are highly infectious viruses, for which seropositive poultry have been identified on multiple islands (Soos et al., 2008; Whitehead et al., 2018; Deem et al., 2012a; Gottdenker et al., 2005; Wikelski et al., 2004). In a 2008 survey of broiler and backyard poultry on Santa Cruz Island (Soos et al., 2008), backyard flocks were more likely to show clinical disease compared to broilers and had higher rates of seropositivity for infectious laryngotracheitis virus, IBR, avian reovirus, and MDV. Spillover of pathogens from backyard poultry to wild birds is a significant concern, given that backyard poultry are more likely to encounter wildlife, directly or through a shared environment (Review: Ayala et al., 2020). Serologic evaluation of multiple species of wild birds on Santa Cruz Island showed no evidence of these pathogens, suggesting that poultry-to-wildlife transmission was at the time contained, and that the wild birds were not serving as a reservoir for transmission to poultry (Soos et al., 2008). Similarly, Galápagos penguins, flightless cormorants (Nannopterum harrisi), and waved albatross (Phoebastria irrorata) were serologically negative for NDV, IBV, IBDV, and MDV (Travis et al., 2006a, Travis et al., 2006b, Padilla et al., 2003). In two surveys on Floreana Island, Floreana mockingbirds (Mimus trifasciatus) had no evidence of exposure to NDV or avian adenovirus II (AAV-II) (Deem et al., 2011); however, but wild birds of other species tested seropositive for NDV, avian poxvirus, and AAV-II (Deem et al., 2012a). Avian poxvirus was introduced to the Galápagos Islands over a century ago via anthropogenic transport (Parker et al., 2011) and has been associated with nestling mortality in the critically endangered waved albatross (Tompkins et al., 2017). Different finch species in the Galápagos Islands may have varying levels of susceptibility to avian poxvirus (McNew et al., 2022). Taken together, these surveys of poultry and wild birds suggest that there are differences in susceptibility or risk factors between endemic species. It is important to consider that seropositivity does not indicate active infection, and even infected birds may be subclinical for disease. Continued surveillance of poultry and wild birds is necessary to fully assess the threat of these avian viruses for endemic birds.
2.2.2. Bacterial pathogens of importance to the Galápagos Islands
Leptospirosis is a re-emerging zoonotic bacterial disease caused by the spirochete Leptospira interrogans that affects multiple mammalian species, including domestic dogs and humans. Rodents, particularly rats, serve as reservoirs. Leptospira is shed in urine and then contaminates water and soil, remaining infective for months. Leptospirosis has been documented in Galápagos sea lions on San Cristóbal Island (Denkinger et al., 2017) and in the California sea lion (Lloyd-Smith et al., 2007), South American fur seal (Arctocephalus australis) and South American sea lion (Otaria flavescens, O. byronia) (Sepúlveda et al., 2015, Katz et al., 2022). Whether sea lions are only sporadic hosts is unclear. No human cases of leptospirosis were reported from the Galápagos Islands between 2000 and 2020 (Calvopiña et al., 2022).
Mycoplasma gallisepticum is a pathogen of poultry with significant implications for wild birds. In North America, M. gallisepticum established wildlife reservoirs in house finches before becoming endemic in wild songbirds (Ley et al., 2016; Delaney et al., 2012; Sawicka et al., 2020). M. gallisepticum causes severe conjunctivitis, affecting sight, flight, and resource acquisition. M. gallisepticum has been documented in backyard poultry and broilers on Santa Cruz (Soos et al., 2008) and Floreana Island (Deem et al., 2012). Although it has not yet been identified in surveyed wild birds (Soos et al., 2008; Deem et al., 2011), the potential remains for it to be transmitted from poultry to wildlife and then establish an endemic reservoir.
Other bacterial pathogens of poultry, such as Mycoplasma synoviae, Bordetella avium, Pasteurella multocida, and Chlamydia psittaci, are also threats to endemic birds. C. psittaci has been identified in flightless cormorants (Travis et al., 2006b) and Galápagos doves (Padilla et al., 2004). Padilla et al. (2003) reported no waved albatross seropositive for C. psittaci. In a survey of several vulnerable species of endemic Galápagos birds, Aaziz et al. (2023) identified C. abortus in 35.6% of waved albatross from Española Island. C. abortus is a causative agent of abortion in ruminants, but its potential as an avian pathogen, or the role of birds in its transmission to livestock, is unknown (Szymanska-Czerwinska et al., 2017).
2.2.3. Parasitic diseases of importance to the Galápagos Islands
D. immitis, the causative agent of canine heartworm disease, is a filarial nematode transmitted by infected mosquitoes. Canids are the natural hosts, in which nematodes grow to maturity within the pulmonary arteries, then mate to produce circulating microfilariae that are then transmitted by mosquitoes to the next host. Heartworm disease affects a variety of mammals, including humans, causing pulmonary disease, heart failure, and rarely, neurologic signs. Treatment is prolonged and physiologically taxing, making it unlikely that infected wildlife could be treated without being brought into captivity.
D. immitis was first documented in domestic dogs on Floreana Island in the 1980s, with microfilaria detected in blood samples from 77% (24/31) of domestic dogs sampled (Barnett & Rudd, 1983; Barnett, 1985a). D. immitis has since been identified in dogs on Isabela Island (Levy et al., 2008) and Santa Cruz Island, more commonly around brackish water lagoons that serve as a mosquito breeding site (Jimenez et al., 2020). In 1985, Barnett (1985a) documented circulating microfilariae in 19% (7/36) of Galápagos sea lions sampled, demonstrating a direct and long-standing risk to this iconic species. A newer report confirmed the presence of intracardiac worms in a Galápagos sea lion (Gregory et al., 2023), demonstrating that D. immitis could become established in a wildlife reservoir. D. immitis has also been reported in South African fur seals (Arctocephalus pusillus pusillus) and common seals (Phoca vitulina) (Alho et al., 2017), as well as a Humboldt penguin (S. humboldti) (Sano et al., 2005), suggesting that the Galápagos fur seal and Galápagos penguin are also at risk. In addition, Barnett (1985a) documented 84% seropositivity for antibodies against D. immitis among humans on Floreana Island. However, no studies in the past 40 years have evaluated the prevalence of this disease in humans in the Galápagos Islands. Serologic testing of humans would provide information both about public health and the regional risk to sea lions.
2.2.4. Emerging pathogens of One Health importance for the Galápagos Islands
Emerging pathogens discussed in this section include those recently documented among humans, domestic animals, and/or wildlife in the Galápagos Islands. This section also includes pathogens that have been documented in humans or domestic animals in the archipelago but have either not yet been documented in Galápagos wildlife or are in the early stages of diagnosis in wildlife.
2.2.4.1. Avian influenza
Avian influenza is an emerging pathogen in the Galápagos Islands. This virus is a respiratory and gastrointestinal pathogen of birds, with both Low Pathogenic (LPAI) and Highly Pathogenic (HPAI) forms, that also has zoonotic potential. LPAI strains circulate naturally in wild birds, particularly waterfowl, and can spread to domestic birds (e.g., poultry) via fecal contamination. LPAI typically causes mild or subclinical disease in birds and is not considered a major public health threat. Wild waterfowl can carry multiple LPAI strains and remain subclinical. The emergence of HPAI is intrinsically linked to anthropogenic activities through the maintenance of poultry at high stocking densities in intensified agricultural conditions. Transmission of LPAI strains to poultry promotes the development of HPAI strains that then spill back over into wild birds; migrating birds can then spread HPAI to new locales along migration routes. HPAI strains cause severe disease and high mortality, with outbreaks in poultry leading to severe economic losses. Because control measures of positive flocks involve depopulation, an HPAI outbreak can decimate the poultry industry. HPAI is also a threat to wild birds and mammals in the Galápagos Islands (Kaplan & Webby, 2013).
While outbreaks of HPAI have periodically cycled through Eurasia and North America, South America has historically remained geographically insulated from this pathogen. However, in 2021, a new strain of HPAI, H5N1, emerged in Eurasia and rapidly spread to North America. This strain is both highly transmissible and carries a high risk of mortality for poultry, wild birds, and mammals. Genetic evidence suggests that H5N1 was transmitted to Peru via a single introduction event from North America, presumably through wild bird migration. In 2022, H5N1 was linked to mortality in harbor seals and gray seals in Maine (Puryear et al., 2023) and dolphins (Delphinus delphis) and South American sea lions (Otaria flavescens) in Peru (Leguia et al., 2023). Several mutations concerning for mammalian host adaptation have been identified in samples from the Peruvian outbreak, with direct mammal-to-mammal transmission suspected to play a role in sea lion die-offs (Leguia et al., 2023). These findings have important implications for other wildlife species at risk, such as the Galápagos sea lion and fur seal, and for transmission to humans. In previous HPAI outbreaks, the individuals most at risk for zoonotic infection were those in close contact with birds, such as poultry farmers and veterinarians. However, these recent findings suggest that H5N1 could have the potential to cause a larger-scale outbreak in humans.
The first outbreak of HPAI in Ecuador occurred in November of 2022, with high mortality in poultry (Bruno et al., 2023). In response, ABG issued an emergency resolution to prohibit importation of day-old chicks and poultry products, including meat and eggs, and to suspend interisland movement of poultry. Over the past two decades, several studies have surveilled endemic Galápagos birds for HPAI, with no positive samples identified (Travis et al., 2006a, Travis et al., 2006b, Padilla et al., 2003, Deem et al., 2012). With the recent spread of H5N1 to South America, ABG has been engaged in active surveillance for HPAI in poultry farms, for which all samples to date have been negative.
Unfortunately, in September of 2023, HPAI was identified in the Galápagos Islands for the first time, following reports of mortality and clinical signs in wild birds on the northern islands of Wolf and Genovesa (Gobierno del Ecuador, 2023). Two dead frigate birds and one red-footed booby were confirmed positive for HPAI by molecular testing performed on mainland Ecuador (Stokstad, 2023). Since the initial detection, ABG has rapidly established the capability to perform on-site molecular testing and intensive surveillance is underway. ABG also plans to conduct genomic testing on any HPAI strains identified to assess its origin, virulence, and transmissibility, informing risk assessment, management and containment strategies. As a precautionary measure, several visitor sites across the GNP have been closed pending further investigation. Education of farmers, tour guides, and GNPD staff is also needed to increase the capacity for visual surveillance of wild birds and poultry, and to warn tourists to report—but not to approach—any birds with concerning signs. HPAI can cause respiratory, gastrointestinal, and neurologic signs, including uncoordinated walking or flying, abnormal head position, circling, or sudden death. Given the evolving global status of the H5N1 outbreak, HPAI remains a transboundary disease with high risk and dire potential consequences for both humans and animals of the Galápagos Islands.
2.2.4.2. Toxoplasma gondii and intestinal parasites
Toxoplasma gondii is a zoonotic apicomplexan parasite for which felids are the definitive host. T. gondii has been identified in domestic cats in the Galápagos Islands (Levy et al., 2008). This parasite is shed in the feces and sporulates to the infective form within 48 hours. Oocysts subsequently contaminate groundwater and reach aquatic environments (VanWormer et al., 2014; Shapiro et al., 2019; Dubey et al., 2003). Ingestion of oocysts by mammals or birds results in formation of cysts within muscle or migration to the brain, causing neurologic signs. Pet and feral domestic cats contribute a large proportion of environment-contaminating oocysts (VanWormer et al., 2013).
Seropositivity for T. gondii has been reported in the Galapagos hawk (Deem et al., 2012b), Galápagos penguin, and flightless cormorant (Deem et al., 2010). Ongoing ABG research efforts include characterization of T. gondii seropositivity rates in cattle and canines from the four populated islands of the archipelago. ABG has also established a protocol for circumstantial monitoring of the Galapagos sea lion and Galapagos fur seal for this parasite, with preliminary results identifying positive individuals of both species (unpublished data). T. gondii has also been reported in the California sea lion (Zalophus californianus) (Migaki et al., 1977; Carlson-Bremer et al., 2015) and other sea lion species (Michael et al., 2016; Sepúlveda et al., 2015, Alvarado-Esquivel et al., 2012).
The canine hookworm, Ancylostoma caninum, and the canine roundworm, Toxocara canis, have been identified in dogs in the Galápagos Islands (Diaz et al., 2016; Gingrich et al., 2010). These parasites can cause cutaneous and visceral larva migrans in humans, respectively. Neither species has been reported in pinnipeds. Another hookworm species, Uncinaria spp., has been identified in otariids (Seguel & Gottdenker, 2017), including the Galápagos sea lion (Herbert, 2014). The potential remains for marine mammals to serve as aberrant hosts of canine intestinal parasites.
Given that the primary mode of transmission for these parasites involves fecal contamination, several strategies to mitigate their spread can be implemented. Preventing domestic cats and dogs from roaming freely would decrease environmental contamination and eliminate the possibility of predation and other interactions with wildlife. Pet owners should be diligent about collecting waste to reduce the risk of groundwater contamination. For instance, T. gondii takes 48 hours to sporulate in the environment, thus discarding cat feces daily will prevent household exposure to infectious stages of the parasite. At the regional level, development of appropriate wastewater treatment and solid waste handling strategies is essential, such that feces discarded in municipal trash does not ultimately end up contaminating water sources or marine environments.
2.2.4.3. Vector-borne pathogens
The naturalized yellow fever mosquito (A. aegypti) is the vector of dengue virus, chikungunya virus, and Zika virus, all of which are emerging pathogens in the Galápagos Islands, with the first human cases identified in 2002, 2015, and 2016, respectively (Ryan et al., 2019). During epidemics of these viruses, humans serve as the primary host and the source of infection of new mosquitoes and thus, disease transmission. Concern for enzootic maintenance of these viruses in wildlife populations has been raised, with seropositivity has been documented in rodents and birds; however, there is scant evidence that these species develop clinical disease or are capable of infecting new mosquitoes (Silva & Dermody, 2017; Bueno et al., 2016; Gwee et al., 2021; Bosco-Laugh et al., 2016). Serologic and genetic evidence of dengue, Zika, chikungunya, and other arboviruses have been identified in various bats, although the epidemiologic significance of these findings requires further study (Review: Fagre & Kading, 2019; Review: Gwee et al., 2021). Bats are frequently perceived as threats to public health as hosts of emerging pathogens, particularly viruses, although these concerns may be overstated (Review: Weinberg & Yovel, 2022). Galápagos is home to the endemic Galápagos red bat (Laciurus borealis brachyotis) and the native hoary bat (Lasiurus cinereus spp. villosissimus). Few studies have been conducted on these bats (Key & Sangoquiza, 2008; McCracken et al., 2009), and to the authors’ knowledge, no studies have evaluated these bats in the context of infectious disease.
In March 2023, 100,000 sterile A. aegypti mosquitos were released on Santa Cruz Island as part of a collaborative campaign to eradicate the mosquito, spearheaded by ABG and the National Institute for Public Health Research (INSPI) and supported by the Galápagos Conservancy. Similar biological control methods have been successful elsewhere, such as the eradication of the American screwworm (Cochliomyia hominivorax) from several countries in the Americas (Wyss, 2000).
The brown dog tick, Rhipicephalus sanguineus, is a common ectoparasite of dogs in the Galápagos Islands (Jimenez et al., 2020) and will also rarely bite humans. R. sanguineus is a vector for several zoonotic diseases identified in Galápagos dogs, including Ehrlichia canis, E. ewingii, and Anaplasma phagocytophilum (Diaz et al., 2016; Jimenez et al., 2020). Several species of Bartonella have been identified in cats on Isabela Island (Levy et al., 2008). Bartonella henselae, the zoonotic causative agent of cat scratch fever, utilizes fleas as an intermediate host and vector. Bartonellosis has also been documented in seals in other regions, transmitted by seal lice (Morick et al., 2009).
Leishmania donovani, a zoonotic pathogen, has also been reported in dogs on Isabela Island (Levy et al., 2008), although its New World vector, the phlebotomine sand fly (Lutzomyia spp.), has yet to be identified in the archipelago. Further surveillance is therefore warranted to identify the arthropod vector in the Galápagos Islands and assess its risk to the human population.
2.2.4.4. Mycoplasma spp.
Mycoplasma spp. are commensal organisms common to the respiratory tract of many reptiles, birds, and mammals, including pinnipeds (Greig et al., 2005). In the context of concurrent respiratory disease, however, Mycoplasma spp. can complicate and exacerbate clinical signs. Mycoplasma spp. have been isolated from Galápagos sea lions with concurrent respiratory signs and appear to be distinct species from those commonly found in cats and dogs (Sarzosa et al., 2021). While Mycoplasma spp. can also affect humans, zoonotic transmission from infected animals is unlikely. Further research should be conducted regarding potentially novel bacterial species identified in Galápagos wildlife.
2.2.4.5. Novel reptile adenoviruses and herpesviruses
In 2021, researchers identified novel adenoviruses and herpesviruses among five species of giant tortoises on Santa Cruz, Isabela, and Española Islands (Nieto-Claudin et al., 2021a). The pathogenic potential of these viruses is unknown and should be further studied. Herpesviruses in other species are often latent, long-standing infections with the potential to exacerbate clinical signs in the context of concurrent infection or immunosuppression.
2.2.5. Future infectious disease risks
This section includes pathogens that have yet to be documented in the Galápagos Islands, but for which competent vectors and/or reservoirs are present and for which the risk of introduction is high. Continued surveillance of both domestic animals and endemic wildlife is necessary to monitor these pathogens as potential future threats to the Galápagos Islands.
2.2.5.1. West Nile Virus and other mosquito-transmitted arboviruses
WNV is a mosquito-transmitted flavivirus with avian reservoirs that causes disease in birds and mammals, including humans. WNV was first introduced from Europe to North America in 1999, resulting in significant mortality of wild birds and febrile encephalitis in humans and horses, both dead-end hosts (Sejvar, 2003). WNV has not been detected in the Galápagos Islands, with surveys of wild birds on multiple islands finding no seropositive individuals (Eastwood et al., 2014). However, WNV is present in mainland Ecuador (Coello-Peralta et al., 2019) and is at risk for introduction to the Galápagos Islands (Kilpatrick et al., 2006). The naturalized mosquitoes C. quinquefasciatus and A. aegypti are competent vectors of WNV (Eastwood et al., 2011; Eastwood et al., 2013; Eastwood et al., 2019). Given repeated introductions of C. quinquefasciatus to the archipelago via airplanes (Bataille et al., 2009b), continued upward trajectory of tourism, and rising global temperatures, the introduction and establishment of WNV are ongoing risks. While WNV causes only sporadic disease among birds in Europe, North American birds exhibit high morbidity and mortality (McLean, 2006), likely due to being immunologically naïve. We expect that Galápagos birds would similarly be susceptible.
Other mosquito-transmitted arboviruses such as Eastern equine encephalitis virus (EEEV), Western equine encephalitis virus (WEEV), Venezuelan equine encephalitis virus (VEEV), and Japanese encephalitis virus (JEV) also pose risks as emerging zoonotic threats, in the context of climate change, urbanization, and global tourism and immigration (Go et al., 2014; Mackenzie et al., 2016). To date, only one study has evaluated the seroprevalence of these viruses in Galápagos wildlife, with Travis et al. (2006a) finding no Galápagos penguins seropositive for these viruses.
2.2.5.2. Coxiella burnetii
Coxiella burnetii is an intracellular bacterial pathogen and the causative agent of Q fever in humans and abortions in ruminants. Many infected humans and animals are asymptomatic. However, clinical disease can manifest as acute, severe fever, headache, respiratory signs, and muscle and joint pain. Chronically infected individuals can also develop hepatitis and endocarditis, with higher mortality than acute cases. C. burnetii naturally cycles within wildlife populations, with ticks as the primary vector. In addition, a distinct domestic cycle exists within domestic ruminants, with transmission to humans via direct contact with contaminated milk, urine, feces, or reproductive tissues and fluids from periparturient goats, sheep, or cattle (Welch, 2016). Transmission can also occur via inhalation of aerosolized dust particles containing C. burnetii spores. Human-to-human transmission is possible but uncommon. Due to its hardiness in the environment, potential for aerosolization, and high infectivity, C. burnetii has been identified as a potential bioterrorism agent (Kagawa et al., 2003).
C. burnetii has been identified in many ruminant herds worldwide, although with variable prevalence depending on the region (Bauer et al., 2022; Bwatota et al., 2022; El-Mahallawy et al., 2014; Epelboin et al., 2023; Georgiev et al., 2013; Kim et al., 2005). Overall, this pathogen remains understudied in Latin America (Epelboin et al., 2023). C. burnetii is endemic in Ecuador, with a seroprevalence of 43-53% in dairy cattle and 34% in farm workers (Carbonero et al., 2015; Changoluisa et al., 2019; Echeverría et al., 2019); occupational exposure is likely underreported.
Two studies to date have investigated the prevalence of C. burnetii in the Galápagos Islands. In one study, 500 bovine serum samples were analyzed by ELISA, with no positive cases (Chalan and Omar, 2021). In the second study, 5 milk samples from dairy cattle were negative on molecular testing (Rojas et al., 2013). While the archipelago may currently be considered free of C. burnetii, more comprehensive surveys should be conducted of livestock serum samples and bulk milk tank samples. In addition, occupational surveillance and education on disease recognition should be provided for farmers. Surveillance of feral goats, which could serve as reservoirs, is desirable. C. burnetii has been identified in Australian fur seals (Arctocephalus pusillus doriferus), although its significance as a causative agent of abortion in this species remains unclear (Gardner et al., 2022); this pathogen should thus be assumed to pose a potential risk to the Galápagos fur seal.
2.2.5.3. Mycobacteria
Tuberculosis is an important pathogen of humans, non-human primates, and cattle worldwide. The Mycobacterium tuberculosis complex includes M. tuberculosis, M. bovis, and M. pinnipedii, among many others. Tuberculosis classically presents as respiratory disease with granulomatous pulmonary lesions. M. tuberculosis, the principal cause of tuberculosis in humans, was long thought to have originated in the Old World via cross-species transmission from M. bovis-infected cattle during domestication, and then introduced to humans in the Americas by early explorers. However, comparative genomics has since demonstrated that human strains predate M. bovis and other animal-adapted Mycobacteria (Brosch et al., 2002). Recent archaeological evidence from ancient Peruvians also suggests that M. tuberculosis was present in the New World long before European colonization (Bos et al., 2014). In fact, these researchers posited that it was the hunting of marine mammals infected with M. pinnipedii that served as the original source of New World tuberculosis.
Infections with the M. tuberculosis complex have been documented in multiple species of fur seals and sea lions (Forshaw & Phelps, 1991; Katz et al., 2022), and M. bovis has been isolated from a grey seal pup (Barnett et al., 2013), suggesting that multiple species of Mycobacteria may pose risks for Galápagos pinnipeds. The first confirmed case of animal-to-human transmission of M. pinnipedii was reported in 2019 (Macedo et al., 2020). Tuberculosis remains a public health concern in mainland Ecuador, and has been described in one case report from a patient in the Galápagos Islands (Garzon-Chavez et al., 2020).
2.2.5.4. Other pathogens of pinnipeds
Many pathogens have been characterized in the California sea lion which likely have the potential to affect the Galápagos sea lion, and thus should be targets for future surveillance in Galápagos wildlife. For example, otarine adenovirus 1, related to canine adenovirus 1 and 2, causes viral hepatitis in sea lions (Goldstein et al., 2011). California sea lions are also host to otariine herpesvirus-1 (Gulland et al., 2020) and their own caliciviruses (Smith et al., 1990), the respiratory agent Mycoplasma zalophi (Sarzosa et al., 2021, Haulena et al., 2006), and the hemoplasm Mycoplasma haemozalophi (Volokhov et al., 2011).
2.2.5.5. Livestock-transmitted zoonoses
Surveillance of infectious diseases in cattle, goats, pigs, and horses in the Galápagos Islands has been limited. Livestock may transmit zoonotic diseases (Review: McDaniel et al., 2014) and transmit pathogens to wildlife, with particular concern for pinnipeds. For instance, Vesicular Exanthema of Swine, which causes clinical signs that mimic Foot and Mouth Disease, is genetically indistinguishable from San Miguel Sea Lion Virus. ABG conducted extensive surveillance of cattle between 2014 and 2015, with no evidence of Brucella abortus (Gioia et al., 2018), Foot and Mouth Disease, or bovine leukosis virus. Nonetheless, ongoing surveillance is necessary.
2.2.5.6. Fungal pathogens
With climate change and rising global temperatures, fungal agents are poised to emerge as important pathogens (Nnadi & Carter, 2021), yet fungi remain understudied globally. No vaccines currently target fungi, and a limited spectrum of antifungals are available, often requiring long courses of therapy. Fungal spores are also notoriously resilient to degradation. Several fungal organisms have emerged as wildlife pathogens in the past decades, including chytrid fungus (Batrachochytrium dendrobatidis) of amphibians and white-nose syndrome (Pseudogymnoascus destructans) of bats. Cryptococcus gattii, a systemic fungal pathogen of humans and animals, has caused outbreaks of pulmonary and neurologic disease in humans, marine mammals, and penguins (Rosenberg et al., 2016; Fenton et al., 2017; Huckabone et al., 2015; Venn-Watson et al., 2014; Brito Devoto et al., 2022). Candida auris has also recently emerged as a human pathogen of public health importance.
Several studies have surveyed Galápagos fungi in association with soil, trees, vegetation, and insects (Ajello & Padhye, 1974; Schoenborn et al., 2023; Nelder et al., 2004; James et al., 2015; Freitas et al., 2013; Guamán-Burneo et al., 2015), but few studies focus on fungi as pathogens in the region (Carvajal Barriga et al., 2014). Sutton et al. (2013) and Christman et al. (2020) identified two novel fungal species in Galápagos tortoises, from carapace and pulmonary lesions, respectively. Fungal pathogen surveillance of both environmental and wildlife samples is therefore imperative for risk assessment and development of a comprehensive surveillance and monitoring plan.
2.3. Antimicrobial resistance
In 2013, the United States Centers for Disease Control and Prevention (CDC) highlighted the global public health implications of antimicrobial resistance (AMR) via the first Antibiotic Resistance Threats report (CDC, 2013), bringing more widespread attention to the growing threat. Factors contributing to the development of AMR include overuse of broad-spectrum antibiotics public and veterinary health, prophylactic use of antibiotics, and subtherapeutic antibiotics used to promote growth in livestock (CDC, 2013), a practice which has been discontinued in the United States for medically important antibiotics under the 2017 Veterinary Feed Directive (FDA, 2017). Antibiotics exert selective pressure for emergence or persistence of antimicrobial resistance genes (ARGs), which can then be horizontally transferred between bacteria. Despite efforts to safeguard public health through judicious use of antibiotics, AMR remains a significant threat to healthcare worldwide. Infections with AMR bacteria result in prolonged hospitalization, economic hardship, failure of treatment and increased morbidity and mortality (Review: Dadgostar, 2019). In early 2022, the CDC reported that the SARS-CoV-2 pandemic prompted a significant increase in antibiotic use and AMR-associated infections worldwide, in part due to enormous pressure placed on global healthcare networks (CDC, 2022). AMR affects human, domestic animal, and wildlife health, therefore a comprehensive assessment of the impacts of AMR requires a One Heath perspective.
Several studies have evaluated AMR in the Galápagos Islands, all supporting a link between human-wildlife contact and the presence of bacteria with ARGs. Thaller et al. (2010) screened colonies of land iguanas (Conolophus pallidus) in remote areas of Santa Fe Island, finding that acquired AMR traits were exceedingly rare (~1%); the authors concluded that absence of humans was protective against AMR acquisition. Wheeler et al. (2012) documented a higher rate of AMR in E. coli isolated from reptiles in closer proximity to human areas compared to those from remote research areas (Wheeler et al., 2012). Nieto-Claudin et al. (2021) also found that giant tortoises in urban and agricultural zones contained more fecal ARGs compared to samples from remote areas. Sewage discharge at beaches on San Cristóbal Island was associated with higher concentrations of Enterococcus and more AMR in E. coli (Overbey et al., 2015), providing one mechanism by which proximity to human activities can promote AMR strains and lead to zooanthroponotic transmission. In addition, antibiotics carried in tourist luggage with inappropriate use or disposal could contribute to environmental contamination, with downstream effects for human and wildlife health.
While the presence of AMR bacteria does not necessarily confirm a pathogenic role, their presence is concerning for two main reasons: 1) many commensal or environmental microbes may become opportunistic pathogens in the context of concurrent stress, disease, or immune compromise; and 2) if an infectious disease outbreak were to occur in endemic wildlife, the presence of AMR would complicate treatment, particularly in a region where access to antibiotics is limited even in the public health sector. Further research should evaluate AMR in the Galápagos Islands in the context of domestic and wild species to develop guidelines for antibiotic stewardship in the public health, agriculture, and regulatory sectors.
3. Regulations and Surveillance
3.1. Institutions with a role in building the “One Health” strategy for the Galápagos Islands
At the national level, several agencies overlap in responsibilities to protect public health. INSPI is the national surveillance and reference laboratory that provides specialized public health services for Ecuador (Gobierno del Ecuador, 2022a). The Phytosanitary and Zoosanitary Regulation and Control Agency (AGROCALIDAD), associated with the Ministry of Agriculture and Livestock is responsible for the regulation, protection, and improvement of animal and plant health, and food safety (Gobierno del Ecuador, 2022b). The National Agency for Regulation, Control and Sanitary Surveillance (ARCSA) regulates the quality, safety, and efficacy of medicines, processed foods, and cosmetics, and conducts sanitary surveillance (Gobierno del Ecuador, 2022c). Notably, these institutions act primarily at the continental level, but have agreements with the agencies in charge of health of the Galápagos Islands.
In 2003, the Ecuadorian government enacted the Regulation for the Total Control of Introduced Species in Galapagos Province (RCTEI), which further defined regulations on the transport of food products to and within the archipelago, and established a Provincial Agricultural Health and Quarantine Committee, responsible for reviews and recommendations every 5 years. RCTEI also outlines biosecurity procedures, such as disinfection procedures for vessels and a system of vessel certification, with reported compliance of 95% (Brewington et al., 2012). Additional recommendations have been made to reduce the risk of contaminated maritime equipment and ballast water (Brewington et al., 2012).
The institutions that govern sustainability of the Galápagos Islands at the local level are the Government Council (CREG), focused on public policy; the GNPD, centered on management of protected areas and wildlife; and ABG, safeguarding animal, vegetal, and human health. ABG is a technical-public entity attached to the Ministry of the Environment. ABG’s lines of action were framed to comply with the Invasive Species Management Plan for Galápagos, summarized in Table 5 (Espinosa & Cedeño, 2022). ABG carries out surveillance, diagnostics, and research for timely identification of threats in urban and wild fauna. ABG is also responsible for emergency response plans, disease control programs, and epidemiological research. To monitor the food production sector, ABG institutes and biosecurity and containment measures for livestock, including quarantine procedures and slaughterhouse inspections, and certifies the movement of animals and animal products between islands (Gobierno del Ecuador, 2022d).

Table 5.
ABG Strategic Plan for Management of Invasive Species.
| Biosecurity Strategy | Health Strategy | Priority Invasive Species Management Strategy |
| C1: Prevention C2: Early Detection C3: Rapid Response |
C1: Baseline of diseases C2: Phyto-zoo-sanitary Epidemiological Surveillance C3: Health of domestic and feral animals |
C1: Integral Management C2: Innovation |
| Transversal Strategy of the Information Management System for Invasive Species in Galápagos. | Transversal Strategy for Institutional Strengthening for the Management of Invasive Species | Transversal Strategy for Communication, Environmental Education and Participation for the Management of Invasive Species |
| C1: Technological Development C2: Baseline C3: Follow-up C4: Research C5: Prioritization |
C1: Coordination C2: Financing C3: Training C4: Legal Framework |
C1: Communication C2: Education C3: Participation |
C: components.
Given the unique ecological status of the Galápagos Islands, international and intergovernmental organizations play key roles in setting conservation goals and evaluating progress. Approximately every 8 years, the UNESCO World Heritage Committee evaluates the status of World Heritage sites based on voluntary updates on legislative and administrative progress provided by the State Party (UNESCO, 2023a). New Action Plans are then formulated with discrete goals to address the most urgent challenges. The most recent state of conservation report for the Galápagos Islands was presented on November 28, 2022, highlighting threats of tourism, land use, unregulated or illegal fishing, and invasive species (UNESCO, 2023b). The corresponding conclusions from the World Heritage Centre and Advisory Bodies will be further discussed in Part II of this review.
The IUCN Red List monitors wildlife populations and is an invaluable monitoring and assessment tool. However, several caveats should be considered when using the IUCN Red List to assess the conservation status of Galápagos species. The interval between IUCN assessments is variable, thus the most recent IUCN classification may not accurately reflect a species’ current conservation status. Accurate assessments rely on available scientific data, which may be scarce. For instance, the San Cristóbal giant tortoise (C. chathamensis) was first assessed in 1996 as Vulnerable and not re-assessed until 2017, when it was classified as Endangered (Caccone et al., 2017). Thus, the IUCN Red List should not be used to exclude certain species from conservation efforts simply because they have not yet been classified as threatened or endangered. In addition, many species have yet to be evaluated by the IUCN. Most importantly, the global status of a species may not fully reflect the conservation status of local or regional populations. The IUCN states that the Red List was “developed for assessing extinction risk at the global level … If the criteria are used on their own to assess non-endemic species at regional or national levels, this could result in incorrect or even misleading assessments” (IUCN, 2022). Particularly on oceanic islands, isolated populations have diversified into subspecies and are subject to distinct pressures compared to the parent species. For instance, the blue-footed booby (Sula nebouxii) is native to the Galápagos Islands but is also found throughout the Gulf of California and western Central and South America. The Galápagos endemic subspecies, S. n. excisa, has declined by over 50% in the past two decades (Anchundia et al., 2014) and is considered Endangered by the Charles Darwin Foundation, yet the parent species remains listed as Least Concern by the IUCN as of 2020 (CDF Species Checklist, 2023; BirdLife International, 2021). Similarly, endemic subspecies of the short-eared owl (Asio flammeus ssp. galapagoensis), great blue heron (Ardea herodias ssp. cognata), and yellow-crowned night heron (Nyctanassa violacea ssp. pauper) are all threatened by regional pressures, despite global populations remaining of Least Concern. These considerations may in part explain differences between the IUCN and CDF assessments of risk status of endemic species in the archipelago.
Multidisciplinary collaboration between governmental institutions, non-profit and non-governmental organizations, and private entities or private research centers are key in the control and maintenance of biosafety in the Galápagos Islands, as well as for identifying conservation strategies to mitigate existing threats. Organizations such as the CDF, Galápagos Science Center (GSC), and the Galápagos Conservancy promote sustainability through conservation and research, providing personnel, funding, and expertise to drive key projects forward. These organizations also conduct global campaigns to increase public awareness of threats facing the Galápagos, leading to a valuable source of external funding.
3.2. Current ABG activities and avenues for regulatory improvement
ABG is a bold institution due to its administrative autonomy and sanctioning/regulatory power, and its acceptance as a regulatory unit by the local, national, and international community. On some Islands, ABG sites have the most advanced infrastructure, laboratory equipment, and technical and managerial capacity. ABG also has the scope to collaborate with inter-institutional and international organizations that provide technical assistance. As a relatively young entity, however, ABG can still strive to implement improvements at an institutional and operational level.
Because of the large volume of tourist activity and the reliance on imported products, both air and sea are potential routes for introduction of invasive species or pathogens, and require rigorous and active surveillance. For the purposes of inspection and quarantine, ABG maintains a list of approved and prohibited organic products (ABG, 2013), devised by balancing the need to supply internal markets while considering the risks associated with potential introduction of invasive species or biologic threats (Rogg et al., 2005). Import of animals has been prohibited since 1999, with limited exceptions. Until recent restrictions associated with HPAI surveillance, unvaccinated chicks could be imported to supply the poultry industry. Case-by-case exemptions are granted by the board of ABG; for example, in 2014, trained dogs were imported to detect invasive giant African snails (Lissachatina fulica), which destroy crops and native vegetation and threaten critically endangered endemic Galápagos land snails (Bulimulus ochsneri).
Currently, the list of prohibited products is only published in Spanish, which limits full accessibility to tourists. Providing lists in English to tour companies and cruise ships would increase awareness and improve compliance. Additionally, having these lists is not entirely beneficial if baggage or import inspections are not rigorous. Gaps in current surveillance could allow for introduction of non-native plants, animals, or microbes by passenger or cargo vessels. X-ray scanning of crates and baggage is performed on the mainland and upon arrival at official ports of entry. However, there is often a delay between scanning of cargo on the mainland, loading of crates onto the transport vessel, and vessel departure, during which time the crates are stored for days in areas with open air access. Improving the efficiency of cargo surveillance and loading—thus decreasing the duration of time spent in storage, on the dock, or in the hold of vessels—would decrease exposures to sources of contamination or rodent infestation. Cargo could be tagged with designated ship dates and X-ray scanning and visual inspection conducted just prior to loading. Additionally, where pre-shipment storage is necessary, cargo should be stored on raised shipping pallets in clean, dry, and vermin-proof indoor areas. Cargo crates are randomly selected for opening and visual inspection, but it is not currently possible to open every crate. Passenger baggage arriving to Baltra and Cristóbal airports is already surveilled by sniffer dogs trained to detect organic materials. We recommend the incorporation of sniffer dogs for cargo, allowing more targeted inspections.
Since ABG is considered a public investment project, activities may also be constrained by budget availability and delays in accessing external funding. There are also limitations in equipment, reagents, or infrastructure to support advanced diagnostic techniques. ABG requires personnel with specialized training in microbiology, epidemiology, biosecurity surveillance, and related fields. Current legislation, meant to promote local economic growth, restricts the contracts available to non-Galapagueños. In practice, however, there is a lack of investment in education opportunities for Galápagos residents, thus these positions may remain unfilled. It is necessary to expand the acquisition of human talent to ensure that ABG has the expertise and bandwidth to carry out all necessary surveillance and monitoring.
Under the current infrastructure, there is also a barrier to communication, impeding the efficiency of surveillance programs for emerging threats. ABG is responsible for monitoring diseases of public health and veterinary importance. A central electronic database for sharing diagnostic, epidemiological, and geographic reports would enhance information transfer and transparency. This database would be of most utility in the context of a One Health monitoring network, to consolidate information generated by research institutions for access by policymakers, regulatory agencies, and scientists, enabling efficient and informed decision-making.
Most of the Galápagos Islands is a protected area (PA). Research in Southeast Asia has shown that effectively managed PAs are an asset to vertebrate conservation, with increased wildlife diversity inside reserves and in adjacent unprotected areas, suggesting the United Nations’ goal of achieving 30% PA coverage by 2030 will benefit global biodiversity (Brodie et al., 2023). However, many large PAs suffer from lack of resources and infrastructure for efficient management, thus creating “paper parks.” It is imperative that the Galápagos Islands do not fall prey to this pitfall, particularly where human presence and wildlife conservation are so closely intertwined and managed visitation of protected areas are a core economic component. Continued assessment of the biodiversity landscape is necessary to ensure that GNP protections are functioning as intended.
4. Foreword to Part II
In Part II of this review, we provide an overview of prior and predicted effects of climate change on Galápagos biodiversity and highlight the widespread ecosystem impacts of anthropogenic activities including tourism, fishing, and agriculture from a One Health Perspective. We also review factors governing socioeconomic stability in the region and discuss further recommendations towards a sustainable future.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
IAJ, PAVM, and EH contributed to conception and design of the review. All authors contributed to review of the literature and writing of the manuscript. IAJ utilized CDF and IUCN databases to compile tables on wildlife conservation status. All authors contributed to manuscript revision and have read and approved of the submitted version.
Conflicts of Interest
PAVM is a paid employee of ABG. The other authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Charles Darwin Foundation. (2023). Galápagos Species Checklist. https://www.darwinfoundation.org/en/datazone/checklist [Accessed 28 Jan, 2023].
- Tye, A., Snell, H. L., Peck, S. B., Adsersen, H. (2002). “Outstanding terrestrial features of the Galápagos archipelago,” in A biodiversity vision for the Galápagos Islands. (Puerto Ayora, Galápagos: Charles Darwin Foundation and World Wildlife Fund), pages 12-23.
- Arbogast, B. S., Drovetski, S. V., Curry, R. L., Boag, P. T., Seutin, G., Grant, P. R., et al. (2006). The origin and diversification of Galapagos mockingbirds. Evolution. 60(2):370-82. PMID: 16610327. [CrossRef]
- Smith, G. T. C. (1979). “Looking Back on Twenty Years of the Charles Darwin Foundation.” Charles Darwin Foundation. Noticias De Galapagos. 30:5-13.
- Townsend, C. H. (1925). The Galápagos tortoises in their relation to the whaling industry: A study of old logbooks. Zoologica. 4:55-135. [CrossRef]
- MacFarland, C. G., Villa, J., Toro, B. (1974). The Galápagos Giant Tortoises (Geochelone elephantopus) Part I: Status of the Surviving Populations. Biological Conservation. 6(2):118-133. [CrossRef]
- Dumbacher, J. P., and West, B. (2010). Collecting the Galápagos and the Pacific: How Rollo Howard Beck Shaped our Understanding of Evolution. Proceedings of the California Academy of Sciences. 4(61) Suppl. II, No.13. pp211-243.
- Gifford, E. W. (1908). The rehabilitation of the California Academy of Sciences. The Condor. 10:95–96.
- Tonnis, B., Grant, P. R., Grant, B. R., Petren, K. (2005). Habitat selection and ecological speciation in Galápagos warbler finches (Certhidea olivacea and Certhidea fusca). Proc Biol Sci. 22;272(1565):819-26. [CrossRef]
- Adame, F. (2021). Meaningful collaborations can end “helicopter research.” Nature Career Columns. [Accessed 8 June, 2023]. [CrossRef]
- Minasny, B., Fiantis, D., Mulyanto, B., Sulaeman, Y., Widyatmanti, W. (2020). Global soil science research collaboration in the 21st century: Time to end helicopter research. Geoderma. 373:114299. [CrossRef]
- Chin, A., Baje, L., Donaldson, T., Gerhardt, K., Jabado, R. W., Kyne, P. M., et al. (2019). The scientist abroad: Maximizing research impact and effectiveness when working as a visiting scientist. Biological Conservation. 238:108231.
- Barnett, B. D., Rudd, R. L. (1983). Feral dogs of the Galápagos Islands: impact and control. Int. J. Stud. Anim. Prob. 4, 44–58. [CrossRef]
- Barnett, B. D. (1985a). El gusano del corazon del perro (Dirofilaria immitis) en las Galápagos. Biblioteca Charles Darwin.
- Bensted-Smith, R. (2002). A Biodiversity Vision for the Galapagos Islands: Based on an International Workshop of Conservation Biologists in Galapagos in May 1999. Puerto Ayora, Galapagos: Charles Darwin Foundation.
- Snell, H. L., Powell, G., Tye, A., Bensted-Smith, R., Bustamante, R. H., Branch, G. M. (2002). “Approach to projecting the future of Galápagos biodiversity,” in A biodiversity vision for the Galápagos Islands. (Puerto Ayora, Galápagos: Charles Darwin Foundation and World Wildlife Fund).
- Dirección del Parque Nacional Galápagos. (2014). “Plan de Manejo de las Áreas Protegidas de Galápagos para el Buen Vivir.” Puerto Ayora, Galápagos, Ecuador.
- UNESCO. (2006). “World Heritage Committee: Thirtieth session.” https://whc.unesco.org/en/sessions/30COM/documents/[Accessed 24 March, 2023], P 70-71.
- UNESCO. (2007). “World Heritage Committee: Thirty-first session.” https://whc.unesco.org/en/sessions/31COM/documents/[Accessed 24 March, 2023], P 68-69.
- UNESCO. (2010). “World Heritage Committee: Thirty-fourth session.” https://whc.unesco.org/en/sessions/34COM/documents/[Accessed 24 March, 2023], P 34-35.
- Bensted-Smith, R., Powell, G., Dinerstein, E. (2002). “Planning for the ecoregion,” in A biodiversity vision for the Galápagos Islands. (Puerto Ayora, Galápagos: Charles Darwin Foundation and World Wildlife Fund), pages 1-5.
- Rogg, H., Buddenhagen C., Causton, C. (2005). “Experiences and limitations with pest risk analysis in the Galápagos Islands.” In: Identification of risks and management of invasive alien species using the IPPC framework. Proceedings of the workshop on invasive alien species and the International Plant Protection Convention, Braunschweig, Germany, 22-26 September 2003. Rome, Italy, FAO. https://www.fao.org/3/y5968e/y5968e0m.htm [Accessed 10 September, 2023].
- Grant, P., Grant, B., Petren, K., Keller, L. (2005). Extinction behind our backs: the possible fate of one of the Darwin’s finch species on Isla Floreana, Galapagos. Biological Conservation 122: 499–503.
- O’Connor, J., Sulloway, F., Kleindorfer, S. (2010). Avian population survey in the Floreana highlands: is Darwin’s Medium Tree Finch declining in remnant patches of Scalesia forest? Bird Conservation International 20: 1–11. [CrossRef]
- Dvorak, M., Nemeth, E., Wendelin, B., Herrera, P., Mosquera, D., Anchundia, D., et al. (2017). Conservation Status of Landbirds on Floreana: The Smallest Inhabited Galápagos Island. Journal of Field Ornithology. 88(2):132–45. [CrossRef]
- Dvorak, M., Fessl, B., Nemeth, E., Kleindorfer, S., Tebbich, S. (2011). Distribution and abundance of Darwin’s finches and other land birds on Santa Cruz Island, Galapagos: evidence for declining populations. Oryx 46: 1–9. [CrossRef]
- Riofrío-Lazo, M., Arreguín-Sánchez, F., Páez-Rosas, D. (2017). Population Abundance of the Endangered Galapagos Sea Lion Zalophus wollebaeki in the Southeastern Galapagos Archipelago. PLoS One. 12(1):e0168829. [CrossRef]
- Denkinger, J., Quiroga, D., Murillo, JC. (2014). “Assessing human–wildlife conflicts and benefits of Galápagos sea lions on San Cristobal Island, Galápagos,” in The Galápagos Marine Reserve 2014 (pp. 285-305). Springer, Cham. [CrossRef]
- Denkinger, J., Gordillo, L., Montero-Serra, I., Murillo, J. C., Guevara, N., Hirschfeld, M., Fietz, K., Rubianes, F., Dan, M. (2015). Urban life of Galápagos sea lions (Zalophus wollebaeki) on San Cristobal Island, Ecuador: colony trends and threats. Journal of Sea Research. 1;105:10-4. [CrossRef]
- One Health High-Level Expert Panel (OHHLEP); Adisasmito, W. B., Almuhairi, S., Behravesh, C. B., Bilivogui, P., Bukachi, S. A. et al. (2022). One Health: A new definition for a sustainable and healthy future. PLoS Pathogens. 18(6):e1010537. [CrossRef]
- Fauci, A. S. (2006). Emerging and re-emerging infectious diseases: influenza as a prototype of the host-pathogen balancing act. Cell. 124(4):665-70. [CrossRef]
- Stephen, C., Wilcox, A., Sine, S., Provencher, J. (2023). A reimagined One Health framework for wildlife conservation. Research Directions: One Health, 1, E12. [CrossRef]
- Tounta, D. D., Panagiotis, T. T., Tesseromatis, C. (2022). Human Activities and Zoonotic Epidemics: A Two-Way Relationship. The Case of the COVID-19 Pandemic. Global Sustainability 5 (January): e19. [CrossRef]
- Watkins, G., Cruz, F. (2007). Galapagos at Risk: A Socioeconomic Analysis of the Situation in the Archipelago. Puerto Ayora, Province of Galapagos, Ecuador, Charles Darwin Foundation.
- Toral-Granda, M. V., Causton, C. E., Jäger, H., Trueman, M., Izurieta, J. C., Araujo, E., et al. (2017). Alien species pathways to the Galápagos Islands, Ecuador. PLoS One. 12(9):e0184379. [CrossRef]
- Pizzitutti, F., Walsh, S. J., Rindfuss, R. R., Gunter, R., Quiroga, D., Tippett, R., et al. (2016). Scenario planning for tourism management: a participatory and system dynamics model applied to the Galapagos Islands of Ecuador. Journal of Sustainable Tourism. 25(8):1117-1137. [CrossRef]
- Guézou, A., Trueman, M., Buddenhagen, C. E., Chamorro, S., Guerrero, A. M., Pozo, P. et al. (2010). An extensive alien plant inventory from the inhabited areas of Galápagos. PLoS ONE. 5:1-8. [CrossRef]
- Tye, A. (2006). Can we infer island introduction and naturalization rates from inventory data? Evidence from introduced plants in Galápagos. Biol Invasions. 8:201-215. [CrossRef]
- Blackburn, T. M., Bellard, C., Ricciardi, A. (2019). Alien versus native species as drivers of recent extinctions. Frontiers in Ecology and the Environment. 17(4):203-207. [CrossRef]
- Rentería, J. L., Gardener, M. R., Panetta, F. D., Atkinson, R., Crawley, M. J. (2012). Possible impacts of the invasive plant Rubus niveus on the native vegetation of the Scalesia forest in the Galapagos islands. PLoS One. (10):e48106. [CrossRef]
- Calderón Alvarez, C., Causton, C. E., Hoddle, M. S., Hoddle, C., Van Driesche, R., Stanek III, E. (2012). Monitoring the effects of Rodolia cardinalis on Icerya purchasi populations on the Galapagos Islands. BioControl 57:167-179. [CrossRef]
- Rentería, J. L., Ellison, C. (2004). Potential biological control of Lantana camara in the Galapagos using the rust Puccinia lantanae. SIDA, Contributions to Botany. 21(2):1009-1017.
- Thomas, S. E., Evans, H. C., Cortat, G., Koutsidou, C., Day, M. D., & Ellison, C. A. (2021). Assessment of the microcyclic rust Puccinia lantanae as a classical biological control agent of the pantropical weed Lantana camara. Biological Control, 160, 104688. [CrossRef]
- Causton, C. E., Peck, S. B., Sinclair, B. J., Roque-Albelo, L., Hodgson, C. J., Landry, B. (2006). Alien insects: threats and implications for conservation of Galápagos Islands. Ann. Entomol. Soc. Am. 99, 121–143. [CrossRef]
- Bataille, A., Cunningham, A. A., Cedeno, V., Cruz, M., Eastwood, G., Fonseca, D. M., et al. (2009a). Evidence for regular ongoing introductions of mosquito disease vectors into the Galápagos Islands. Proc. R. Soc. Lond. Biol. 276, 3769–3775.
- Bataille, A., Cunningham, A. A., Cedeno, V., Patino, L., Constantinou, A., Kramer, L. D., et al. (2009b). Natural colonization and adaptation of a mosquito species in Galápagos and its implications for disease threats to endemic wildlife. Proc. Natl. Acad. Sci. 106, 10230–10235. [CrossRef]
- Whiteman, N. K., Goodman, S. J., Sinclair, B. J., Walsh, T., Cunningham, A. A., Kramer, L. D., et al. (2005). Establishment of the avian disease vector Culex quinquefasciatus SAY, 1823 (Diptera: Culicidae) on the Galápagos Islands, Ecuador. Ibis. 147, 844–847. [CrossRef]
- Sinclair, B. J. (2017). “CDF checklist of Galápagos flies,” in Charles Darwin Foundation Galápagos Species Checklist, eds. F. Bungartz, H. Herrera, P. Jaramillo, N. Tirado, G. Jiménez-Uzcátegui, D. Ruiz, A. Guézou, F. Ziemmeck. Charles Darwin Foundation, Puerto Ayora, Galápagos, 45.
- Kilpatrick, A. M., Daszak, P., Goodman, S. J., Rogg, H., Kramer, L. D., Cedeño, V. et al. (2006). Predicting pathogen introduction: West Nile virus spread to Galápagos. Conserv Biol. 20(4):1224-31. [CrossRef]
- Sardelis, M. R., Turell, M. J., Dohm, D. J., O’Guinn, M. L. (2001). Vector competence of selected North American Culex and Coquillettidia mosquitoes for West Nile virus. Emerg. Infect. Dis. 7: 1018D1022. [CrossRef]
- Barnett, B.D. (1985b). Impact of domestic dog populations in the galápagos: prevalence and transmission of canine heartworm. In: Dogs of the Galápagos Islands: Evolution, Ecology, Impact and Control. University of California, Davis Doctoral dissertation. (Retrieved from University Microfilms International Dissertation Abstracts database. Accession No. 852195).
- Hendrix, C., Brunner, C., Bellamy, L. (1986). Natural transmission of Dirofilaria immitis by Aedes aegypti. J. Am. Mosq. Control Assoc. 2, 48–51.
- Van Riper, C., van Riper, S. C., Goff, M. L., Laird, M. C. (1986). The epizootiology and ecological significance of malaria on the birds of Hawaii. Ecol. Monogr. 56: 327Ð344. [CrossRef]
- Harvey-Samuel, T., Ant, T., Sutton, J. Niebuhr, C. N., Asigau, S., Parker, P. et al. (2021). Culex quinquefasciatus: status as a threat to island avifauna and options for genetic control. CABI Agric Biosci 2, 9. [CrossRef]
- Nishida, G. M., Evenhus, N. L. (2000). “Arthropod pests of conservation significance in the Pacific: a preliminary assessment of selected groups,” in Invasive species in the Pacific: a technical review and draft regional strategy, ed. G. Sherley. South Pacific Regional Environment Programme, Samoa, 115D142.
- Miller, G. D., Hofkin, B. V., Snell, H., Hahn, A., Miller, R. D. (2001). Avian malaria and Marek’s disease: potential threats to Galápagos penguins Spheniscus mendiculus. Marine Ornithology. 29:43-46.
- Palmer, J. L., McCutchan, T. F., Vargas, F. H., Deem, S. L., Cruz, M., Hartman, D. A., et al. (2013). Seroprevalence of malarial antibodies in Galapagos penguins (Spheniscus mendiculus). J Parasitol. 99(5):770-6. [CrossRef]
- Levin, I. I., Zwiers, P., Deem, S. L., Geest, E. A., Higashiguchi, J. M., Iezhova, T. A., et al. (2013). Multiple lineages of Avian malaria parasites (Plasmodium) in the Galapagos Islands and evidence for arrival via migratory birds. Conserv Biol. 27(6):1366-77. [CrossRef]
- Fessl, B., Heimpel, G. E., Causton, C. E. (2018). “Chapter 9: Invasion of an Avian Nest Parasite, Philornis downsi, to the Galapagos Islands: Colonization History, Adaptations to Novel Ecosystems, and Conservation Challenges,” in Disease Ecology, Social and Ecological Interactions in the Galapagos Islands, ed. P.G. Parker. [CrossRef]
- Causton C., Cunninghame F., Tapia W. (2013). “Management of the avian parasite Philornis downsi in the Galapagos Islands: A collaborative and strategic action plan,” in Galapagos Report 2011-2012. GNPS, GCREG, CDF and GC. Puerto Ayora, Galapagos, Ecuador, 167-173.
- Butterwort, V., Dansby, H., Zink, F. A., Tembrock, L. R., Gilligan, T. M., Godoy, A., et al. (2022). A DNA extraction method for insects from sticky traps: targeting a low abundance pest, Phthorimaea absoluta (Lepidoptera: Gelechiidae) in mixed species communities. Journal of Economic Entomology. 115(3):844-851. [CrossRef]
- Mee, P. T., Wong, S., Brown, K., Lynch, S. E. (2021). Quantitative PCR assay for the detection of Aedes vigilax on mosquito trap collections containing large numbers of morphologically similar species and phylogenetic analysis of specimens collected in Victoria, Australia. Parasites & Vectors. 14(434). [CrossRef]
- Hadfield, T. L., Turell, M., Dempsey, M. P., David, J., Park, E. J. (2001). Detection of West Nile virus in mosquitoes by RT-PCR. Mol Cell Probes. 15(3):147-50. [CrossRef]
- Ali, E. O. M., Babalghith, A. O., Bahathig, A. O. S., Dafalla, O. M., Al-Maghamsi, I. W., Mustafa, N. E. A. G., et al. (2022). Detection of Dengue Virus From Aedes aegypti (Diptera, Culicidae) in Field-Caught Samples From Makkah Al-Mokarramah, Kingdom of Saudi Arabia, Using RT-PCR. Front Public Health. 10:850851. [CrossRef]
- Carrion, V., Donlan, C. J., Campbell, K., Lavoie, C., Cruz, F. (2007). Feral donkey (Equus asinus) eradications in the Galápagos. Biodiversity and Conservation. 16:437-445. [CrossRef]
- Ballesteros-Mejia, L., Angulo, E., Diagne, C., Cooke, B., Nuñez M. A., Courchamp, F. (2021). Economic costs of biological invasions in Ecuador: the importance of the Galapagos Islands. NeoBiota, 67: 375–400. [CrossRef]
- Bush, M. B., Conrad, S., Restrepo, A., Thompson, D. M., Lofverstrom, M., Conroy, J. L. (2022). Human-induced ecological cascades: extinction, restoration, and rewilding in the Galápagos highlands. 119(24):e2203752119. [CrossRef]
- deVries, T., Black, J. (1983). Of Men, Goats, & Guava—Problems Caused by Introduced Species in the Galápagos. Noticias de Galápagos. 38.
- Cruz, F., Carrion, V., Campbell, K. J., Lavoie, C., Donlan, C. J. (2009) Bio-economics of large-scale eradication of feral goats from Santiago Island, Galápagos. Journal of Wildlife Management 73: 191–200. [CrossRef]
- Cruz, F., Donlan, C. J., Campbell, K., Carrion, V. (2005). Conservation action in the Galápagos: feral pig (Sus scrofa) eradication from Santiago Island. Biol Conserv 121:473–478. [CrossRef]
- Carrion, V., Donlan, C. J., Campbell, K. J., Lavoie, C., Cruz, F. (2011). Archipelago-wide island restoration in the Galápagos Islands: Reducing Costs of Invasive Mammal Eradication Programs and reinvasion risk. PLoS ONE. 6(5):18835. [CrossRef]
- Phillips, R. B., Cooke, B. D., Carrión, V., Snell, H. L. (2012a). Eradication of rock pigeons, Columba livia, from the Galápagos Islands. Biological Conservation. 147(1):264-269. [CrossRef]
- Phillips, R. B., Cooke, B. D., Campbell, K., Carrion, V., Marquez, C., Snell, H. L. (2012b). Eradicating feral cats to protect Galapagos land iguanas: methods and strategies. Pacific Conservation Biology. 11(4):257-267. [CrossRef]
- Causton, C. E., Sevilla, C. R., Porter, S. D. (2005). Eradication of the Little Fire Ant, Wasmannia Auropunctata (Hymenoptera: Formicidae), from Marchena Island, Galápagos: On the Edge of Success? The Florida Entomologist. 88 (2):159–68. [CrossRef]
- Barnett, B. D. (1986). “Eradication and control of feral and free-ranging dogs in the Galápagos Islands,” in Proc. 12th Vertebr. Pest C., ed. T. Salmon. University of California, Davis, pp. 358–368.
- Reponen, S. E., Brown, S. K., Barnett, B. D., Sacks, B. N. (2014). Genetic and morphometric evidence on a Galápagos Island exposes founder effects and diversification in the first-known (truly) feral western dog population. Mol Ecol. 23(2):269-83. [CrossRef]
- Kruuk, H., and Snell., H. (1981). Prey selection by feral dogs from a population of marine iguanas (Amblyrhynchus cristatus). J Appl Ecol. 18:197-204. [CrossRef]
- Jimenez, I. A., Vega Mariño, P. A., Stapleton, G. S., Prieto, J. B., Bowman, D. D. (2020). Canine vector-borne disease in domestic dogs on Isla Santa Cruz, Galápagos. Vet Parasitol Reg Stud Reports. 19:100373. [CrossRef]
- Diaz, N. M., Mendez, G. S., Grijalva, C. J., Walden, H. S., Cruz, M., Aragon, E., et al. (2016). Dog overpopulation and burden of exposure to canine distemper virus and other pathogens on Santa Cruz Island, Galapagos. Prev Vet Med. 123:128-137. [CrossRef]
- Gingrich, E. N., Scorza, A. V., Clifford, E. L., Olea-Popelka, F. J., Lappin, M. R. (2010). Intestinal parasites of dogs on the Galapagos Islands. Vet Parasitol. 169(3-4):404-7. [CrossRef]
- Hernandez, J. A., Yoak, A. J., Walden, H. S., Thompson, N., Zuniga, D., Criollo, R., Duque, V., Cruz, M. (2020). Dog overpopulation on Santa Cruz Island, Galapagos 2018. Conservation Science and Practice. e201. [CrossRef]
- Cooke, S. C., Haskell, L. E., van Rees, C. B., Fessl, B. (2019). A review of the introduced smooth-billed ani Crotophaga ani in Galápagos. Biol Conserv. 229:38-49. [CrossRef]
- Connett, L., Guézou, A., Herrera, H. W., Carrión, V., Parker, P. G., Deem, S. L. (2016). Gizzard contents of the smooth-billed ani Crotophaga ani in Santa Cruz, Galápagos Islands, Ecuador. Galápagos Res 68:43-48.
- Cooke, S. C., Anchundia, D., Caton, E., Haskell, L. E., Jäger, H., Kalki, Y., et al. (2020). Endemic species predation by the introduced smooth-billed ani in Galápagos. Biol Invasions 22:2113–2120. [CrossRef]
- Keith, I., Dawson, T. P., Collins, K. J., Campbell, M. L. (2016). Marine invasive species: establishing pathways, their presence and potential threats in the Galapagos Marine Reserve, Pacific Conservation Biology, 22 (4), 377-385. [CrossRef]
- Carlton, J. T., Keith I., Ruiz, G. M. (2019). Assessing marine bioinvasions in the Galápagos Islands: Implications for conservation biology and marine protected areas. Aquatic Invasions 14(1), 1-20. [CrossRef]
- Charles Darwin Foundation. (2023). “2022 Impact Report.” 52nd General Assembly of the Charles Darwin Foundation. https://www.darwinfoundation.org/en/publications/annual-report/impact-report-2022 [Accessed 1 July 2023].
- Keith, I., Bensted-Smith, W., Banks, S., Suarez, J., Riegl, B. (2022). Caulerpa chemnitzia in Darwin threatening Galapagos coral reefs. PLoS One. 17(8):e0272581. [CrossRef]
- Baert, L. (1994). Notes on the Status of Terrestrial Arthropods in Galápagos. Noticias de Galápagos. 15-21.
- Edgar, G. J., Banks, S., Fariña, J. M., Calvopiña, M., Martínez, C. (2004) Regional biogeography of shallow reef fish and macro-invertebrate communities in the Galápagos archipelago. Journal of Biogeography. 31(7):1107-1124. [CrossRef]
- Levy, J. K., Crawford, P. C., Lappin, M. R., Dubovi, E. J., Levy, M. G., Alleman, R., et al. (2008). Infectious diseases of dogs and cats on Isabela Island, Galapagos. J Vet Intern Med 22:60–65. [CrossRef]
- Vega-Mariño, P., Olson, J., Howitt, B., Criollo, R., Figueroa, L., Orlando, S. A. et al. (2023). A recent distemper virus outbreak in the growing canine populations of Galapagos Islands: a persistent threat for the endangered Galapagos Sea Lion. Front Vet Sci. 10:1154625. [CrossRef]
- Puente-Rodríguez, D., Bos, A. P. B., Koerkamp, P. W. G. G. (2019). Rethinking livestock production systems on the Galápagos Islands: Organizing knowledge-practice interfaces through reflexive interactive design. Environmental Science & Policy. 101:166-174.
- Gottdenker, N. L., Walsh, T., Vargas, H., Merkel, J., Jiménez, G. U., Miller, R. E., et al. (2005). Assessing the risks of introduced chickens and their pathogens to native birds in the Galápagos Archipelago. Biol Conserv. 126(3):429-439. [CrossRef]
- Whitehead, A. B. R., Butcher, G. D., Walden, H. S., Duque, V., Cruz, M., Hernandez, J. A. (2018). Burden of exposure to infectious bursal disease virus, infectious bronchitis virus, Newcastle disease virus, Mycoplasma gallisepticum, and intestinal parasites in introduced broiler chickens on the Galapagos. PLoS One. 13(9):e0203658. [CrossRef]
- Wikelski, M., Foufopoulos, J., Vargas, H., Snell, H. (2004) Galápagos Birds and Diseases: Invasive Pathogens as Threats for Island Species. Ecology and Society 9(1): 5. http://www.ecologyandsociety.org/vol9/iss1/art5/[Accessed 5 May, 2023].
- Martinez-Gutierrez, M., Ruiz-Saenz, J. (2016). Diversity of susceptible hosts in canine distemper virus infection: a systematic review and data synthesis. BMC Vet Res. 12:78. [CrossRef]
- Beineke, A., Baumgärtner, W., Wohlsein, P. (2015). Cross-species transmission of canine distemper virus-an update. One Health. 1:49-59. [CrossRef]
- Williams, E. S., Thome, E. T., Appel, M. J. G., Belitsky, D. W. (1988). Canine Distemper in black-footed ferrets (Mustela nigripes) from Wyoming. Journal of Wildlife Diseases. 24(3):385-398. [CrossRef]
- Gilbert, M., Sulikhan, N., Uphyrkina, O., Goncharuk, M., Kerley, L., Castro, E. H., et al. (2020). Distemper, extinction, and vaccination of the Amur tiger. Proc Natl Acad Sci USA. 117(50):31954-31962. [CrossRef]
- van de Bildt, M. W., Kuiken, T., Visee, A. M., Lema, S., Fitzjohn, T. R., Osterhaus, A. D. (2002). Distemper outbreak and its effect on African wild dog conservation. Emerg Infect Dis. 8(2):211-3. [CrossRef]
- Denkinger, J., Guevara, N., Ayala, S., Murillo, J. C., Hirschfeld, M., Montero-Serra, I., et al. (2017). Pup mortality and evidence for pathogen exposure in Galapagos sea lions (Zalophus wollebaeki) on San Cristobal Island, Galapagos, Ecuador. J Wildl Dis. 53(3):491-498. [CrossRef]
- Kennedy, J. M., Earle, J. A. P., Omar, S., Abdullah, H., Nielsen, O., Roelke-Parker, M. E., et al. (2019). Canine and Phocine Distemper Viruses: Global Spread and Genetic Basis of Jumping Species Barriers. Viruses. 11(10):944. [CrossRef]
- Bengtson, J. L., Boveng, P., Franzen, U., Have, P., Heidejorgensen, M-P., Harkönen, T. J. (1991). Antibodies to canine distemper virus in Antarctic seals. Marine Mammal Science 7, 85-87. [CrossRef]
- Duignan, P. J., Van Bressem, M. F., Baker, J. D., Barbieri, M., Colegrove, K. M., De Guise, S., et al. (2014). Phocine distemper virus: current knowledge and future directions. Viruses. 6(12):5093-134. [CrossRef]
- Härkönen, T., Dietz, R., Reijnders, P., Teilmann, J., Harding, K., Hall, A., et al. (2006). The 1988 and 2002 phocine distemper virus epidemics in European harbour seals. Dis Aquat Organ. 68(2):115-30. [CrossRef]
- Earle, J. A. P., Melia, M. M., Doherty, N. V., Nielsen, O., Cosby, S. L. (2006). Phocine distemper virus in seals, east coast, United States, Emerg. Infect. Dis. 17:215–220. [CrossRef]
- Goldstein, T., Mazet, J. A., Gill, V. A., Doroff, A. M., Burek, K. A., Hammond, J. A. (2009). Phocine distemper virus in Northern sea otters in the Pacific Ocean, 690 Alaska, USA. Emerg. Infect. Dis. 15:925–927. [CrossRef]
- VanWormer, E., Mazet, J. A. K., Hall, A., Gill, V. A., Boveng, P. L., London, J. M., et al. (2019). Viral emergence in marine mammals in the North Pacific may be linked to Arctic sea ice reduction. Sci Rep. 9(1):15569. [CrossRef]
- Stanton, J. B., Brown, C. C., Poet, S., Lipscomb, T. P., Saliki, J., Frasca, S. Jr. (2004). Retrospective differentiation of canine distemper virus and phocine distemper virus in phocids. J Wildl Dis. 40(1):53-9. [CrossRef]
- Soos, C., Padilla, L., Iglesias, A., Gottdenker, N., Bédon, M. C., Rios, A., et al. (2008). Comparison of pathogens in broiler and backyard chickens on the Galápagos Islands: implications for transmission to wildlife. The Auk. 125(2):445-455. [CrossRef]
- Deem, S. L., Cruz, M. B., Higashiguchi, J. M., Parker, G. P. (2012a). Diseases of poultry and endemic birds in Galapagos: implications for the reintroduction of native species. Anim Conserv. 15:73–82. [CrossRef]
- Ayala, A. J., Yabsley, M. J., Hernandez, S. M. (2020). A Review of Pathogen Transmission at the Backyard Chicken-Wild Bird Interface. Front Vet Sci. 7:539925. [CrossRef]
- Travis, E. K., Vargas, F. H., Merkel, J., Gottdenker, N., Miller, R. E., Parker, P. G. (2006a). Hematology, serum chemistry, and serology of Galápagos penguins (Spheniscus mendiculus) in the Galápagos Islands, Ecuador. J Wildl Dis. 42(3):625-32. [CrossRef]
- Travis, E. K., Vargas, F. H., Merkel, J., Gottdenker, N., Miller, R. E., Parker, P. G. (2006b). Hematology, plasma chemistry, and serology of the flightless cormorant (Phalacrocorax harrisi) in the Galapagos Islands, Ecuador. J Wildl Dis. 42(1):133-41. [CrossRef]
- Padilla, L. R., Huyvaert, K. P., Merkel, J., Miller, R. E., Parker, P. G. (2003). Hematology, plasma chemistry, serology, and Chlamydophila status of the waved albatross (Phoebastria irrorata) on the Galapagos Islands. J Zoo Wildl Med. 34(3):278-83. [CrossRef]
- Deem, S. L., Parker, P. G., Cruz, M. B., Merkel, J., Hoeck, P. E. A. (2011). Comparison of blood values and health status of Floreana Mockingbirds (Mimus trifasciatus) on the islands of Champion and Gardner-by-Floreana, Galápagos Islands. J Wildl Dis. 47: 94–106. [CrossRef]
- Parker, P. G., Buckles, E. L., Farrington, H., Petren, K., Whiteman, N. K., Ricklefs, R. E., et al. (2011). 110 years of Avipoxvirus in the Galapagos Islands. PLoS One. 6(1):e15989. [CrossRef]
- Tompkins, E. M., Anderson, D. J., Pabilonia, K. L., Huyvaert, K. P. (2017). Avian Pox Discovered in the Critically Endangered Waved Albatross (Phoebastria irrorata) from the Galápagos Islands, Ecuador. J Wildl Dis. 53(4):891-895. [CrossRef]
- McNew, S. M., Loyola, D. C., Yepez, J., Andreadis, C., Gotanda, K., Saulsberry, A., et al. (2022). Transcriptomic responses of Galápagos finches to avian poxvirus infection. Mol Ecol. 31(21):5552-5567. [CrossRef]
- Lloyd-Smith, J. O., Greig, D. J., Hietala, S. Ghneim, G. S., Palmer, L., St. Leger, J., et al. (2007). Cyclical changes in seroprevalence of leptospirosis in California sea lions: endemic and epidemic disease in one host species?. BMC Infect Dis. 7,125. [CrossRef]
- Sepúlveda, M. A., Seguel, M., Alvarado-Rybak, M., Verdugo, C., Muñoz-Zanzi, C., Tamayo, R. 2015. Postmortem findings in four south American sea lions (Otaria byronia) from an urban colony in Valdivia, Chile. J Wildl Dis. 51(1):279-82. [CrossRef]
- Katz, H., Schelotto, F., Bakker, D., Castro-Ramos, M., Gutiérrez-Expósito, D., Panzera, Y., et al. (2022). Survey of selected pathogens in free-ranging pinnipeds in Uruguay. Dis Aquat Organ. 150:69-83. [CrossRef]
- Calvopiña, M., Vásconez, E., Coral-Almeida, M., Romero-Alvarez, D., Garcia-Bereguiain, M. A., Orlando, A. (2022). Leptospirosis: Morbidity, mortality, and spatial distribution of hospitalized cases in Ecuador. A nationwide study 2000-2020. PLoS Negl Trop Dis. 16(5):e0010430. [CrossRef]
- Ley, D. H., Hawley, D. M., Geary, S. J., Dhondt, A. A. (2016). House finch (Haemorhous mexicanus) conjunctivitis, Mycoplasma spp. Isolated from north American wild birds, 1994-2015. J Wildlife Dis. 52:669–73. [CrossRef]
- Delaney, N. F., Balenger, S., Bonneaud, C., Marx, C. J., Hill, G. E., Ferguson-Noel N, et al. (2012). Ultrafast evolution and loss of CRISPRs following a host shift in a novel wildlife pathogen, Mycoplasma gallisepticum. PLoS Genet. 8:e1002511. [CrossRef]
- Sawicka, A., Durkalec, M., Tomczyk, G., Kursa O. (2020). Occurrence of Mycoplasma gallisepticum in wild birds: A systematic review and meta-analysis. PLoS One. 15(4):e0231545. [CrossRef]
- Padilla, L. R., Santiago-Alarcon, D., Merkel, J., Miller, R. E., Parker, P. G. (2004). Survey for Haemoproteus spp., Trichomonas gallinae, Chlamydophila psittaci, and Salmonella spp. in Galapagos Islands columbiformes. J Zoo Wildl Med. 35(1):60-4. [CrossRef]
- Aaziz, R., Vinueza, R. L., Vorimore, F., Schnee, C., Jiménez-Uzcátegui G., Zanella, G., et al. (2023). Avian Chlamydia abortus Strains Detected in Galápagos Waved Albatross (Phoebastria irrorata). J Wildl Dis. 59(1):143-148. [CrossRef]
- Szymańska-Czerwińska, M., Mitura, A., Niemczuk, K., Zaręba, K., Jodełko, A., Pluta, A., et al. (2017). Dissemination and genetic diversity of chlamydial agents in Polish wildfowl: isolation and molecular characterisation of avian Chlamydia abortus strains. PLoS ONE. 12:e0174599. [CrossRef]
- Gregory, T. M., Livingston, I., Hawkins, E. C., Loyola, A., Cave, A., Vaden, S. L., et al. (2023). Dirofilaria immitis identified in Galapagos sea lions (Zalophus wollebaeki): a wildlife health and conservation concern. J Wildl Dis. 59(3):487-494. [CrossRef]
- Alho, A. M., Marcelino, I., Colella, V., Flanagan, C., Silva, N., Correia, J. J., et al. (2017). Dirofilaria immitis in pinnipeds and a new host record. Parasit Vectors. 10(1):142. [CrossRef]
- Sano, Y., Aoki, M., Takahashi, H., Miura, M., Komatsu, M., Abe, Y., et al. (2005). The first record of Dirofilaria immitis infection in a Humboldt penguin, Spheniscus humboldti. J Parasitol. 91(5):1235-7. [CrossRef]
- Kaplan, B. S., and Webby, R. J. (2013). The avian and mammalian host range of highly pathogenic avian H5N1 influenza. Virus Res. 178(1):3-11. [CrossRef]
- Puryear, W., Sawatzki, K., Hill, N., Foss, A., Stone, J. J., Doughty, L., et al. (2023). Highly Pathogenic Avian Influenza A(H5N1) Virus Outbreak in New England Seals, United States. Emerging Infectious Diseases. 2023;29(4):786-791. [CrossRef]
- Leguia, M., Garcia-Glaessner, A., Muñoz-Saavedra, B., Juarez, D., Barrera, P., Calvo-Mac, C., et al. (2023). Highly pathogenic avian influenza A (H5N1) in marine mammals and seabirds in Peru. Nature Communications. 14:5489. [CrossRef]
- Bruno, A., Alfaro-Núñez, A., de Mora, D., Armas, R., Olmedo, M., Garcés, J., et al. (2023). Phylogenetic analysis reveals that the H5N1 avian influenza A outbreak in poultry in Ecuador in November 2022 is associated with the highly pathogenic clade 2.3.4.4b. Int J Infect Dis. 133:27-30. [CrossRef]
- Gobierno del Ecuador. (2023) “Comunicado Oficial: Activación de protocolos ante possible afectación de aves en Galápagos.” Ministerio del Ambiente, Agua y Transición Ecológica. Parque Nacional Galápagos. [Accessed 18 September, 2023].
- Stokstad, E. (2023). “Deadly Avian Flu Reaches Galápagos Islands.” Science. https://www.science.org/content/article/deadly-avian-flu-reaches-galapagos-islands [Accessed 20 October, 2023].
- VanWormer, E., Miller, M. A., Conrad, P. A., Grigg, M. E., Rejmanek, D., Carpenter, T. E., et al. (2014). Using molecular epidemiology to track Toxoplasma gondii from terrestrial carnivores to marine hosts: implications for public health and conservation. PLoS Negl Trop Dis. 8(5):e2852. [CrossRef]
- Shapiro, K., Bahia-Oliveira, L., Dixon, B., Dumètre, A., de Wit, L. A., VanWormer, E., et al. (2019). Environmental transmission of Toxoplasma gondii: Oocysts in water, soil and food. Food Waterborne Parasitol. 15:e00049. [CrossRef]
- Dubey, J. P., Zarnke, R., Thomas, N. J., Wong, S. K., Van Bonn, W., Davis, J., et al. (2003b). Toxoplasma gondii, Neospora caninum, Sarcocystis neurona, and Sarcocystis canis-like infections in marine mammals. Vet. Parasitol. 116, 275–296. [CrossRef]
- VanWormer, E., Conrad, P. A., Miller, M. A., Melli, A. C., Carpenter, T. E., Mazet, J. A. (2013). Toxoplasma gondii, source to sea: higher contribution of domestic felids to terrestrial parasite loading despite lower infection prevalence. Ecohealth. 10(3):277-89. [CrossRef]
- Deem, S. L., Rivera-Parra, J. L., Parker, P. G. (2012b). Health evaluation of Galapagos Hawks (Buteo galapagoensis) on Santiago Island, Galapagos. J Wildl Dis. 48(1):39-46. [CrossRef]
- 145. Deem SL, Merkel J, Ballweber L, Vargas FH, Cruz MB, Parker PG. (2010). Exposure to Toxoplasma gondii in Galapagos Penguins (Spheniscus mendiculus) and flightless cormorants (Phalacrocorax harrisi) in the Galapagos Islands, Ecuador. J Wildl Dis. 46(3):1005-11. [CrossRef] [PubMed]
- Migaki, G., Allen, J. F., Casey, H. W. (1977). Toxoplasmosis in a California sea lion (Zalophus californianus). Am J Vet Res. 38(1):135-6. [PubMed]
- Carlson-Bremer, D., Colegrove, K. M., Gulland, F. M., Conrad, P. A., Mazet, J. A., Johnson, C. K. (2015). Epidemiology and pathology of Toxoplasma gondii in free-ranging California sea lions (Zalophus californianus). J Wildl Dis. 51(2):362-73. [CrossRef]
- Michael, S. A., Howe, L., Chilvers, B. L., Morel, P., Roe, W. D. (2016). Seroprevalence of Toxoplasma gondii in mainland and sub-Antarctic New Zealand sea lion (Phocarctos hookeri) populations. N Z Vet J. 64(5):293-7. [CrossRef]
- Alvarado-Esquivel, C., Sánchez-Okrucky, R., Dubey, J. P. (2012). Serological evidence of Toxoplasma gondii infection in captive marine mammals in Mexico. Vet Parasitol. 184(2-4):321-4. [CrossRef]
- Seguel, M., Gottdenker, N. (2017). The diversity and impact of hookworm infections in wildlife. Int J Parasitol Parasites Wildl. 6(3):177-194. [CrossRef]
- Herbert, A. (2014). Caracterización Molecular, Morfológica y Ultra Estructural Del Nematodo Hematófago Uncinaria Sp. en el Lobo Marino de Galapagos. BSc Thesis Univ. Queretaro, Queretaro, Mexico. P 71.
- Ryan, S. J., Lippi, C. A., Nightingale, R., Hamerlinck, G., Borbor-Cordova, M. J., Cruz, B. M., et al. (2019). Socio-ecological factors associated with dengue risk and Aedes aegypti presence in the Galápagos Islands, Ecuador. Int J Environ Res Public Health. 16(5):682. [CrossRef]
- Silva, L. A., Dermody, T. S. (2017). Chikungunya virus: epidemiology, replication, disease mechanisms, and prospective intervention strategies. J Clin Invest. 127(3):737-749. [CrossRef]
- Bueno, M. G., Martinez, N., Abdalla, L., Duarte Dos Santos, C. N., Chame, M. (2016). Animals in the Zika Virus Life Cycle: What to Expect from Megadiverse Latin American Countries. PLoS Negl Trop Dis. 10(12):e0005073. [CrossRef]
- Gwee, S. X. W., St. John, A. L., Gray, G. C., Pang, J. (2021). Animals as potential reservoirs for dengue transmission: A systematic review. One Health. 12:100216. [CrossRef]
- Bosco-Lauth, A. M., Nemeth, N. M., Kohler, D. J., Bowen, R. A. (2016). Viremia in North American Mammals and Birds After Experimental Infection with Chikungunya Viruses. Am J Trop Med Hyg. 94(3):504-6. [CrossRef]
- Fagre, A. C., and Kading, R. C. (2019). Can Bats Serve as Reservoirs for Arboviruses? Viruses. 11(3):215. [CrossRef]
- Weinberg, M., and Yovel, Y. (2022). Revising the paradigm: Are bats really pathogen reservoirs or do they possess an efficient immune system? iScience. 25(8):104782. [CrossRef]
- Key, G., and Sangoquiza, M. (2008). Activity Patterns and Distribution of Galapagos Bats. Galapagos Research. 65:20-24. http://hdl.handle.net/1834/35978.
- McCracken, G. F., Hayes, J. P., Cevallos, J., Guffey, S. Z., Carlos Romero, F. (1997) Observations on the distribution, ecology, and behaviour of bats on the Galápagos Islands. Journal of Zoology. 243(4):757-770. [CrossRef]
- Wyss, J. H. (2000) Screwworm eradication in the Americas. Ann N Y Acad Sci. 916:186-93. [CrossRef]
- Morick, D., Osinga, N., Gruys, E., Harrus, S. (2009). Identification of a Bartonella species in the harbor seal (Phoca vitulina) and in seal lice (Echinophtirius horridus). Vector Borne Zoonotic Dis. 9(6):751-3. [CrossRef]
- Greig, D. J., Gulland, F. M., Kreuder, C. (2005). A decade of live California sea lion (Zalophus californianus) strandings along the central California coast: Causes and trends, 1991–2000. Aquat Mamm 31:11–22. [CrossRef]
- Sarzosa, M. S., Duignan, P., DeRango, E. J., Field, C., Ríos, C., Sanchez, S., et al. (2021). Occurrence of Mycoplasmas in Galapagos Sea Lions (Zalophus wollebaeki) and their Association with Other Respiratory Pathogens. J Wildl Dis. 57(3):623-627. [CrossRef]
- Nieto-Claudin, A., Esperón, F., Apakupakul, K., Peña, I., Deem, S. L. (2022a). Health assessments uncover novel viral sequences in five species of Galapagos tortoises. Transbound Emerg Dis. 69(4):e1079-e1089. [CrossRef]
- Sejvar, J. J. (2003). West Nile virus: an historical overview. Ochsner J. 5(3):6-10. PMID: 21765761.
- Eastwood, G., Goodman, S. J., Hilgert, N., Cruz, M., Kramer, L. D., Cunningham, A. A. (2014). Using avian surveillance in Ecuador to assess the imminence of West Nile virus incursion to Galápagos. Ecohealth. 11(1):53-62. [CrossRef]
- Coello-Peralta, R., González-González, M., Martínez-Cepeda, G. (2019). West Nile Virus in Ecuador. Journal MVZ Cordoba. 24(1):7151-7156.
- Kilpatrick, A. M., Daszak, P., Goodman, S. J., Rogg, H., Kramer, L. D., Cedeño, V., Cunningham, A. A. (2006). Predicting pathogen introduction: West Nile virus spread to Galáipagos. Conserv Biol. 20(4):1224-31. [CrossRef]
- Eastwood, G., Kramer, L. D., Goodman, S. J., Cunningham, A. A. (2011). West Nile virus vector competency of Culex quinquefasciatus mosquitoes in the Galapagos Islands. Am J Trop Med Hyg. 85(3):426-33. [CrossRef]
- Eastwood, G., Cunningham, A. A., Kramer, L. D., Goodman, S. J. (2019). The vector ecology of introduced Culex quinquefasciatus populations, and implications for future risk of West Nile virus emergence in the Galápagos archipelago. Med Vet Entomol. 33(1):44-55. [CrossRef]
- Eastwood, G., Goodman, S. J., Cunningham, A. A., Kramer, L. D. (2013). Aedes taeniorhynchus vectorial capacity informs a pre-emptive Assessment of West Nile Virus establishment in Galápagos. Sci Rep. 3,1519. [CrossRef]
- McLean, R. G. (2006). West Nile Virus in North American Birds. Ornithological Monographs, 60, 44–64. [CrossRef]
- Go, Y. Y., Balasuriya, U. B., Lee, C. K. (2014). Zoonotic encephalitides caused by arboviruses: transmission and epidemiology of alphaviruses and flaviviruses. Clin Exp Vaccine Res. 3(1):58-77. [CrossRef]
- Mackenzie, J. S., Childs, J. E., Field, H. E., Wang, L. F., Breed, A. C. (2016). The Role of Bats as Reservoir Hosts of Emerging Neuroviruses. Neurotropic Viral Infections. Apr 8:403–54. [CrossRef]
- Welch, D. (2016). “Coxiella burnetii.” In: Sentinel Level Clinical Laboratory Guidelines for Suspected Agents of Bioterrorism and Emerging Infectious Diseases. American Society for Microbiology. https://www.asm.org/Articles/Policy/Laboratory-Response-Network-LRN-Sentinel-Level-C [Accessed 10 September 2023].
- Kagawa, F. T., Wehner, J. H., Mohindra V. (2003). Q fever as a biological weapon. Semin Respir Infect. 18(3):183-95.
- Bauer, B. U., Schoneberg, C., Herms, T. L., Runge, M., Ganter, M. (2022). Surveillance of Coxiella burnetii Shedding in Three Naturally Infected Dairy Goat Herds after Vaccination, Focusing on Bulk Tank Milk and Dust Swabs. Vet Sci. 9(3):102. [CrossRef]
- Bwatota, S. F., Shirima, G. M., Hernandez-Castro, L. E., Bronsvoort, B. M. dC., Wheelhouse, N., Mengele, I. J., Motto, S. K., Komwihangilo, D. M., Lyatuu, E., Cook, E. A. J. (2022). Seroprevalence and Risk Factors for Q fever (Coxiella burnetii) Exposure in Smallholder Dairy Cattle in Tanzania, Veterinary Sciences, 9, 12, (662). [CrossRef]
- El-Mahallawy, H., Lu, G., Kelly, P., Xu, D., Li, Y., Fan, W., Wang, C. (2015). Q fever in China: A systematic review, 1989–2013. Epidemiology & Infection, 143(4), 673-681. [CrossRef]
- Epelboin, L., De Souza Ribeiro Mioni, M., Couesnon, A. et al. (2023). Coxiella burnetii Infection in Livestock, Pets, Wildlife, and Ticks in Latin America and the Caribbean: a Comprehensive Review of the Literature. Curr Trop Med Rep 10, 94–13. [CrossRef]
- Georgiev, M., Afonso, A., Neubauer, H., Needham, H., Thiery, R., Rodolakis, A., Roest, H., Stark, K., Stegeman, J., Vellema, P., van der Hoek, W., More, S. (2013). Q fever in humans and farm animals in four European countries, 1982 to 2010. Euro Surveill. 18(8):20407.
- Kim, S. G., Kim, E. H., Lafferty, C. J., Dubovi, E. (2005). Coxiella burnetii in bulk tank milk samples, United States. Emerg Infect Dis. 11(4):619-21. [CrossRef]
- Carbonero, A., Guzmán, L. T., Montaño, K., Torralbo, A., Arenas-Montes, A., Saa, L. R. (2015). Coxiella burnetii seroprevalence and associated risk factors in dairy and mixed cattle farms from Ecuador. Prev Vet Med. 118(4):427-35. [CrossRef]
- Changoluisa, D., Rivera-Olivero, I.A., Echeverria, G. et al. (2019). Serology for Neosporosis, Q fever and Brucellosis to assess the cause of abortion in two dairy cattle herds in Ecuador. BMC Vet Res 15, 194. [CrossRef]
- Echeverría, G., Reyna-Bello, A., Minda-Aluisa, E., Celi-Erazo, M., Olmedo, L., García, H. A., Garcia-Bereguiain, M. A., de Waard, J. H., UNU/BIOLAC network for infectious cattle diseases. (2019). Serological evidence of Coxiella burnetii infection in cattle and farm workers: is Q fever an underreported zoonotic disease in Ecuador? Infect Drug Resist. 12:701-706. [CrossRef]
- Chalan M. and Omar, D. (2021). Seroprevalencia y factores de riesgo asociados a Fiebre Q Coxiella burnetii en bovinos en las Islas Galápagos Ecuador. http://dspace.utpl.edu.ec/jspui/handle/20.500.11962/27785 [Accessed 10 September, 2023].
- Rojas, M. I., Barragán, V., Trueba, G., Hornstra, H., Pearson, T., Keim, P. (2013). Detección de Coxiella burnetii en leche de bovinos domésticos del Ecuador. Avances. 5(1):B5-B9. [CrossRef]
- Gardner, B. R., Stenos, J., Hufschmid, J., Arnould, J. P. Y., McIntosh, R. R., Tadepalli, M., et al. (2022) An Old Pathogen in a New Environment–Implications of Coxiella burnetii in Australian Fur Seals (Arctocephalus pusillus doriferus). Front. Mar. Sci. 9:809075. [CrossRef]
- Brosch, R., Gordon, S. V., Marmiesse, M., Brodin, P., Buchrieser, C., Eiglmeier, et al. (2002). A new evolutionary scenario for the Mycobacterium tuberculosis complex. Proc Natl Acad Sci USA. 99(6):3684-9. [CrossRef]
- Bos, K. I., Harkins, K. M., Herbig, A., Coscolla, M., Weber, N., Comas, I., et al. (2014). Pre-Columbian mycobacterial genomes reveal seals as a source of New World human tuberculosis. Nature 514, 494–497. [CrossRef]
- Forshaw, D., and Phelps, G. R. (1991). Tuberculosis in a captive colony of pinnipeds. J Wildl Dis. 27(2):288-95. [CrossRef]
- Barnett, J. E., Booth, P., Brewer, J. I., Chanter, J., Cooper, T., Crawshaw, T., et al. (2013). Mycobacterium bovis infection in a grey seal pup (Halichoerus grypus). Vet Rec. 173(7):168. [CrossRef]
- Macedo, R., Isidro, J., Gomes, M. C., Botelho, A., Albuquerque, T., Sogorb, A., et al. (2020). Animal-to-human transmission of Mycobacterium pinnipedii. Eur Respir J. 56(6):2000371. [CrossRef]
- Garzon-Chavez, D., Garcia-Bereguiain, M.A., Mora-Pinargote, C., Granda-Pardo, J. C., Leon-Benitez, M., Franco-Sotomayor, G., et al. (2020). Population structure and genetic diversity of Mycobacterium tuberculosis in Ecuador. Sci. Rep. 10, 6237. [CrossRef]
- Goldstein, T., Colegrove, K. M., Hanson, M., Gulland, F. M. (2011). Isolation of a novel adenovirus from California sea lions Zalophus californianus. Dis Aquat Organ. 94(3):243-8. [CrossRef]
- Gulland, F. M. D., Hall, A. J., Ylitalo, G. M., Colegrove, K. M., Norris, T., Duignan, P. J., et al. (2020). Persistent contaminants and Herpesvirus OtHV1 are positively associated with Cancer in wild California Sea Lions (Zalophus californianus). Front. Mar. Sci. 7:602565. [CrossRef]
- Smith, A. W., and Boyt, P. M. (1990). Caliciviruses of ocean origin: a review. J. Zoo Wildl. Med. 21(1):3-23. [CrossRef]
- Haulena, M., Gulland, F. M., Lawrence, J. A., Fauquier, D. A., Jang, S., Aldridge, et al. (2006). Lesions associated with a novel Mycoplasma sp. in California sea lions (Zalophus californianus) undergoing rehabilitation. J Wildl Dis. 42(1):40-5. [CrossRef]
- Volokhov, D. V., Norris, T., Rios, C., Davidson, M. K., Messick, J. B., Gulland, F. M., Chizhikov, V. E. (2011). Novel hemotrophic mycoplasma identified in naturally infected California sea lions (Zalophus californianus). Vet Microbiol. 149(1-2):262-8. [CrossRef]
- McDaniel, C. J., Cardwell, D. M., Moeller, R. B. Jr, Gray, G. C. (2014). Humans and cattle: a review of bovine zoonoses. Vector Borne Zoonotic Dis. 14(1):1-19. [CrossRef]
- Gioia, G., Vinueza, R. L., Cruz, M., Jay, M., Corde, Y., Marsot, M., Zanella, G. (2018). Estimating the probability of freedom from bovine brucellosis in the Galapagos Islands. Epidemiol Infect. 147:e9. [CrossRef]
- Nnadi, N. E., and Carter, D. A. (2021). Climate change and the emergence of fungal pathogens. PLoS Pathog. 17(4):e1009503. [CrossRef]
- Rosenberg, J. F., Haulena, M., Hoang, L. M., Morshed, M., Zabek, E., Raverty, S. A. (2016). Cryptococcus gattii Type VGIIa Infection in Harbor Seals (Phoca vitulina) in British Columbia, Canada. J Wildl Dis. 52(3):677-81. [CrossRef]
- Fenton, H., Daoust, P. Y., Forzán, M. J., Vanderstichel, R. V., Ford, J. K. B., Spaven, L., et al. (2017). Causes of mortality of harbor porpoises Phocoena phocoena along the Atlantic and Pacific coasts of Canada. Dis Aquat Organ. 122(3):171-183. [CrossRef]
- Huckabone, S. E., Gulland, F. M., Johnson, S. M., Colegrove, K. M., Dodd, E. M., Pappagianis, D., et al. (2015). Coccidioidomycosis and other systemic mycoses of marine mammals stranding along the central California, USA coast: 1998-2012. J. Wildl. Dis. 51(2):295-308. [CrossRef]
- Venn-Watson, S., Daniels, R., Smith, C. (2012). Thirty year retrospective evaluation of pneumonia in a bottlenose dolphin Tursiops truncatus population. Dis Aquat Organ. 99(3):237-42.
- Brito Devoto, T., Toscanini, M. A., Hermida Alava, K., Etchecopaz, A. N., Pola, S. J., Martorell, M. M., et al. (2022). Exploring fungal diversity in Antarctic wildlife: isolation and molecular identification of culturable fungi from penguins and pinnipeds. N Z Vet J. 70(5):263-272. [CrossRef]
- Ajello, L., and Padhye, A. (1974). Keratinophilic fungi of the Galapagos Islands. Mykosen. 17(10):239-43. [CrossRef]
- Schoenborn, A. A., Yannarell, S. M., MacVicar, C. T., Barriga-Medina, N. N., Bonham, K. S., Leon-Reyes, A., et al. (2023). Microclimate is a strong predictor of the native and invasive plant-associated soil microbiome on San Cristóbal Island, Galápagos archipelago. Environ Microbiol. Epub ahead of pri. [CrossRef]
- Nelder, M. P., McCreadie, J. W., Coscarón, C., Brockhouse, C. L. (2004). First report of a trichomycete fungus (Zygomycota: Trichomycetes) inhabiting larvae of Simulium ochraceum sensu lato Walker (Diptera: Simuliidae) from the Galapagos Islands. J. Invertebr. Pathol. 87(1):39-44. [CrossRef]
- James, S. A., Carvajal-Barriga, E. J., Barahona, P. P., Nueno-Palop, C., Cross, K., Bond, C. J., et al. (2015). Kazachstania yasuniensis sp. nov., an ascomycetous yeast species found in mainland Ecuador and on the Galápagos. Int J Syst Evol Microbiol. 65(Pt 4):1304-1309. [CrossRef]
- Freitas, L. F. D., Carvajal-Barriga, E. J., Barahona, P. P., Lachance, M. A., Rosa, C. A. (2013). Kodamaea transpacifica f.a., sp. nov., a yeast species isolated from ephemeral flowers and insects in the Galapagos Islands and Malaysia: further evidence for ancient human transpacific contacts. Int J Syst Evol Microbiol. 63(Pt 11):4324-4329. [CrossRef]
- Guamán-Burneo, M. C., Dussán, K. J., Cadete, R. M., Cheab, M. A. M., Portero, P., Carvajal-Barriga, E. J., et al. (2015). Xylitol production by yeasts isolated from rotting wood in the Galápagos Islands, Ecuador, and description of Cyberlindnera galapagoensis f.a., sp. nov. Antonie Van Leeuwenhoek. 108(4):919-31. [CrossRef]
- Carvajal Barriga, E. J., Barahona, P. P., Tufiño, C., Bastidas, B., Guamán-Burneo, C., Freitas, L., et al. (2014). An Overview of the Yeast Biodiversity in the Galápagos Islands and Other Ecuadorian Regions. InTech. [CrossRef]
- Sutton, D. A., Marín, Y., Thompson, E. H., Wickes, B. L., Fu, J., García, D., et al. (2013). Isolation and characterization of a new fungal genus and species, Aphanoascella galapagosensis, from carapace keratitis of a Galapagos tortoise (Chelonoidis nigra microphyes). Med Mycol. 51(2):113-20. [CrossRef]
- Christman, J. E., Alexander, A. B., Donnelly, K. A., Ossiboff, R. J., Stacy, N. I., Richardson, R. L., et al. (2020). Clinical manifestation and molecular characterization of a novel member of the Nannizziopsiaceae in a pulmonary granuloma from a Galapagos Tortoise (Chelonoidis nigra). Front Vet Sci. 7:24. [CrossRef]
- Centers for Disease Control and Prevention. (2013). Antibiotic Resistance Threats in the United States, 2013. https://www.cdc.gov/drugresistance/pdf/ar-threats-2013-508.pdf [Accessed 9 May, 2023].
- Food and Drug Administration. (2017). Veterinary Feed Directive. https://www.fda.gov/animal-veterinary/development-approval-process/veterinary-feed-directive-vfd [Accessed 9 May, 2023].
- Dadgostar, P. (2019). Antimicrobial Resistance: Implications and Costs. Infect Drug Resist. 12:3903-3910. [CrossRef]
- Centers for Disease Control and Prevention. (2022). COVID-19: U.S. Impact on Antimicrobial Resistance, Special Report 2022. Atlanta, GA: U.S. Department of Health and Human Services. https://www.cdc.gov/drugresistance/covid19.html [Accessed 9 May, 2023].
- Thaller, M. C., Migliore, L., Marquez, C., Tapia, W., Cedeño, V., Rossolini, G. M., et al. (2010). Tracking acquired antibiotic resistance in commensal bacteria of Galápagos land iguanas: no man, no resistance. PLoS One. 5(2):e8989. [CrossRef]
- Wheeler, E., Hong, P. Y., Bedon, L. C., Mackie, R. I. (2012). Carriage of antibiotic-resistant enteric bacteria varies among sites in Galapagos reptiles. J. Wildl. Dis. 48(1):56-67. [CrossRef]
- Nieto-Claudin, A., Deem, S. L., Rodríguez, C., Cano, S., Moity, N., Cabrera, F., et al. (2021b). Antimicrobial resistance in Galapagos tortoises as an indicator of the growing human footprint. Environ Pollut. 284:117453. [CrossRef]
- Overbey, K. N., Hatcher, S. M., Stewart, J. R. (2015). Water quality and antibiotic resistance at beaches of the Galápagos Islands. Front. Environ. Sci. 3. [CrossRef]
- Gobierno del Ecuador. (2022a). Instituto Nacional de Investigación en Salud Pública-INSPI-Dr. Leopoldo Izquieta Pérez: Misión-Visión-Objetivos. http://www.investigacionsalud.gob.ec/mision-vision-objetivos/[Accessed 9 May, 2023].
- Gobierno del Ecuador. (2022b). AGROCALIDAD: Misión y visión. https://www.agrocalidad.gob.ec/mision-vision/[Accessed 9 May, 2023].
- Gobierno del Ecuador. (2022c). ARCSA: Misivón,visión y valores https://www.controlsanitario.gob.ec/valores/[Accessed 9 May, 2023].
- Brewington, L., Rosero, O., Bigue, M., & Cervantes, K. (2012). The quarantine chain: establishing an effective biosecurity system to prevent the introduction of invasive species into the Galápagos Islands. Wildaid, San Francisco. https://wildaid.org/wp-content/uploads/2017/09/QuarantineChain.pdf [Accessed 10 September, 2023].
- Espinosa, M., & Cedeño, M. (2022). Plan estratégico isntitucional de la Agencia de regulación y control del Bioseguridad y Cuarente de Galápagos 2019-2022 L. https://www.fao.org/faolex/results/details/es/c/LEX-FAOC197011/[Accessed 9 May, 2023].
- Gobierno del Ecuador. (2022d). ABG: Vigilancia Sanitaria. https://bioseguridadgalapagos.gob.ec/vigilancia-zoosanitaria/[Accessed 9 May, 2023].
- UNESCO. (2023a). Periodic Reporting. https://whc.unesco.org/en/periodicreporting. [Accessed 20 October, 2023].
- UNESCO. (2023b). State of conservation reports. https://whc.unesco.org/en/soc/4511. [Accessed 20 October, 2023].
- Caccone, A., Cayot, L. J., Gibbs, J. P., Tapia, W. (2017). Chelonoidis chathamensis. The IUCN Red List of Threatened Species. doi: 10.2305/IUCN.UK.2017 3.RLTS.T9019A82688009.en [Accessed on 10 June, 2023]. [CrossRef]
- IUCN. (2022). The IUCN Red List of Threatened Species. Version 2022-2. https://www.iucnredlist.org [Accessed on 8 May, 2023].
- Anchundia, D., Huyvaert, K. P., Anderson, D. J. (2014). Chronic lack of breeding by Galápagos Blue-footed Boobies and associated population decline. Avian Conservation and Ecology 9(1):6. [CrossRef]
- BirdLife International. (2021). Sula nebouxii. The IUCN Red List of Threatened Species. [CrossRef]
- ABG. (2013). “Subproductos y derivados de origen vegetal y animal reglamentados para su ingreso a la provincial de Galápagos.” https://bioseguridadgalapagos.gob.ec/lista-de-productos/[Accessed 18 September, 2023].
- Agencia de Regulación y Control de la Bioseguridad y Cuarentena para Galápagos. (2021). Informe de Gestión 2020: Rendición de Cuentas. 26. https://bioseguridadgalapagos.gob.ec/wp-content/uploads/2021/06/Informe-de-Gestión-2020-vs-final.pdf/[Accessed 9 May, 2023].
- Brodie, J. F., Mohd-Azlan, J., Chen, C., Wearn, O. R., Deith, M. C. M., et al. (2023). Landscape-scale benefits of protected areas for tropical biodiversity. Nature. (620):807-812. [CrossRef]
Figure 1.
IUCN Red List status of at-risk vertebrate species of the Galápagos Islands, 1994-2020.
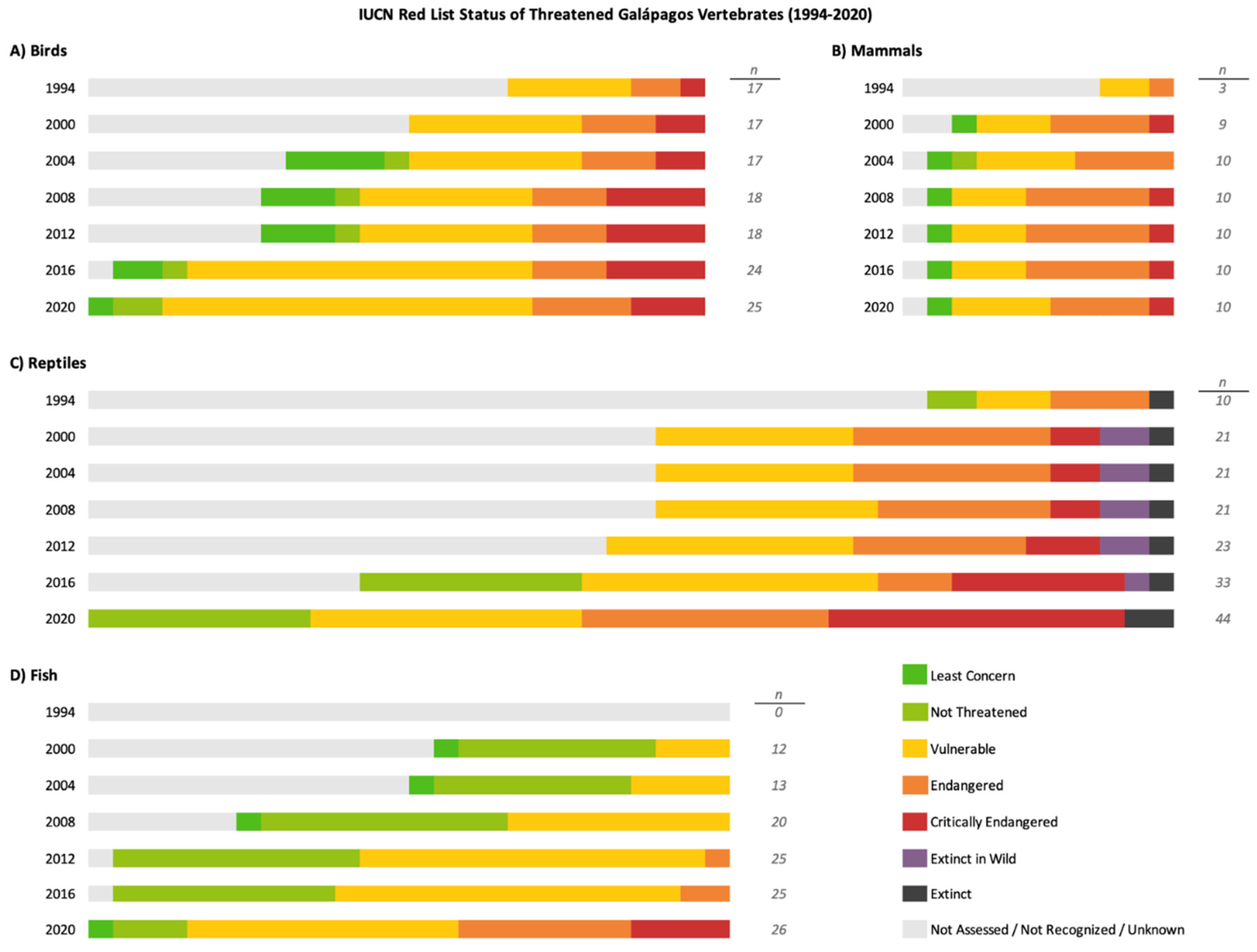
Table 1.
Vertebrates Species of the Galápagos Islands: Origin and CDF Conservation Status.
| Taxon | Endemic | Native | Migrant | Vagrant | Introduced | Other | Total |
| Birds | 47 (22%) | 27 (13%) | 31 (15%) | 65 (31%) | 12 (6%) | 30 (14%) | 212 |
| Extinct | 1 | 1 | |||||
| Critically endangered | 4 | 4 | |||||
| Endangered | 5 | 0 | 1 | 6 | |||
| Vulnerable | 15 | 7 | 0 | 3 | 25 | ||
| Near Threatened | 5 | 3 | 0 | 11 | 1 | 20 | |
| Least Concern | 16 | 14 | 31 | 49 | 9 | 119 | |
| Terrestrial mammals | 9 (38%) | 0 | 0 | 0 | 15 (63%) | 0 | 24 |
| Extinct | 3 | 3 | |||||
| Critically endangered | |||||||
| Endangered | |||||||
| Vulnerable | 5 | 5 | |||||
| Near Threatened | |||||||
| Least Concern | 3 | 3 | |||||
| Marine mammals | 2 (7%) | 2 (7%) | 13 (46%) | 11 (39%) | 0 | 2 (7%) | 28 |
| Extinct | |||||||
| Critically endangered | |||||||
| Endangered | 2 | 2 | 1 | 5 | |||
| Vulnerable | 1 | 1 | |||||
| Near Threatened | |||||||
| Least Concern | 6 | 5 | 11 | ||||
| Reptiles | 37 (9%) | 2 (4%) | 4 (8%) | 0 | 5 (10%) | 4 (8%) | 52 |
| Extinct | 2 | 2 | |||||
| Critically endangered | 7 | 1 | 8 | ||||
| Endangered | 3 | 1 | 4 | ||||
| Vulnerable | 7 | 1 | 2 | 9 | |||
| Near Threatened | 10 | 10 | |||||
| Least Concern | 6 | 1 | 5 | 12 | |||
| Fish | 65 (12%) | 396 (75%) | 0 | 52 (10%) | 1 (<1%) | 14 (3%) | 528 |
| Extinct | |||||||
| Critically endangered | |||||||
| Endangered | 1 | 1 | |||||
| Vulnerable | 3 | 8 | 11 | ||||
| Near Threatened | 10 | 1 | 11 | ||||
| Least Concern | 1 | 16 | 17 | ||||
| Amphibians | 0 | 0 | 0 | 0 | 1 (50%) | 1 (50%) | 2 |
Table 2.
Historical causes of natural and anthropogenic changes in the Galápagos Islands (Snell et al., 2002).
Table 2.
Historical causes of natural and anthropogenic changes in the Galápagos Islands (Snell et al., 2002).
| Causes of natural change | Causes of anthropogenic change |
| Climate change and weather | Invasive species |
| Volcanism | Introduction of infectious diseases |
| Ecological succession | Extractive use of natural resources |
| Competition | Habitat alteration |
| Predation | |
| Dispersal |
Table 3.
Extinct Species of the Galápagos Islands.
| Scientific name | Common name | First Described | Approximate Year of Extinction | Threats/Likely Drivers of Extinction | References |
| Sicyos vollosus | Darwin’s Galápagos gourd | 1835 | 1835 | Grazed to extinction or target of a cucumber virus | CDF Species Checklist Sebastian et al., 2010 |
| Delilia inelegans | N/A | 1835 | 1835 | Unknown; possibly competition by invasive plants or grazed to extinction | CDF Species Checklist Delprete, 1995 |
| Chelonoidis niger | Floreana giant tortoise | 1827 | 1850 | Human exploitation (whalers/mariners) Impacts of introduced species (donkeys, dogs, pigs, black rats, mice, cats, cattle, goats) |
van Dijk et al., 2017 CDF Species Checklist |
| Gomphrena rigida | Galápagos amaranth | 1835 | 1906 | Unknown; possibly competition by invasive plants or grazed to extinction | CDF Species Checklist Lawesson, 1987 |
| Megaoryzomys curioi | Galápagos giant rat | 1964 | 1930 | Unknown; possibly competition, predation, or disease from introduced species (pigs, dogs, black rats, cats) | CDF Species Checklist Weksler & Tirira, 2019 Lange, 2015 |
| Nesoryzomys darwini | Darwin’s Galápagos mouse | 1906 | 1930 | Competition and disease introduction by introduced black rats Predation by feral cats |
Tirira & Weksler, 2019a CDF Species Checklist Dowler et al., 2000 |
| Nesoryzomys indefessus | Indefatigable Galápagos mouse | 1898 | 1934 | Competition and disease introduction by introduced black rats, Norway rats, and house mice Predation by feral cats |
CDF Species Checklist Dowler et al., 2000 Tirira & Weksler, 2019b |
| Pyrocephalus dubius | San Cristóbal Vermilion Flycatcher | 1839 | 1987 | Competition and predation by introduced rats Introduction of avian pox virus Invasive bot fly Philornis downsi |
CDF Species Checklist Carmi et al., 2016 Vargas, 1996 BirdLife International, 2017 |
| Chelonoidis abingdonii | Pinta giant tortoise | 1877 | 2012 | Human exploitation (whalers/mariners) Resource competition and habitat destruction by introduced species (goats) |
Snow, 1964 Cayot et al., 2022 CDF Species Checklist |
Table 4.
Selected Pathogens of One Health Importance for the Galápagos Islands.
| Pathogen | Primary Animal Reservoir (s) | Zoonotic Potential | Documented in Galápagos? | Wildlife at risk | Risk Level | References |
| Avian influenza | Poultry Wild birds |
Yes | Yes | Birds Pinnipeds |
3 | Kaplan & Webby, 2013 Puryear et al., 2023 Leguia et al., 2023 Bruno et al., 2023 |
| Canine distemper virus | Dog | No | Yes | Pinnipeds | 3 | Levy et al., 2008 Diaz et al., 2016 Denkinger at al., 2017 Vega-Mariño et al., 2023 |
| West Nile Virus | Birds | Yes | No | Birds | 3 | Sejvar, 2003 Coello-Peralta et al., 2019 Kilpatrick et al., 2006 Eastwood et al., 2011 McLean, 2006 |
| Coxiella burnetii | Cattle, goats, sheep | Yes | No | Fur seals | 2 | Chalan and Omar, 2021 Gardner et al., 2022 Rojas et al., 2013 |
| Dengue virus | NHP | Yes | Yes | Unlikely | 2 | Gwee et al., 2021 |
| Chikungunya virus | Rodents NHP |
Yes | Yes | Unlikely | 2 | Bosco-Lauth et al., 2016 |
| Dirofilaria immitis | Dog | Yes | Yes | Pinnipeds Galápagos penguins |
2 | Levy et al., 2008 Barnett and Rudd, 1983 Barnett, 1985 Culda et al., 2022 Jimenez et al., 2020 |
| Leptospira interrogans | Rat, Dog, Cattle, Swine | Yes | Yes | Pinnipeds | 2 | Denkinger et al., 2017 Sepúlveda et al., 2015 |
| Phocine Distemper Virus | Seals | No | No | Pinnipeds | 2 | Duignan et al., 2014 Härkönen et al., 2006 Earle et al., 2006 Kennedy et al., 2019 Goldstein et al., 2009 |
| Mycobacterium tuberculosis | Humans | Yes | Yes | Pinnipeds | 2 | Forshaw & Phelps, 1991 Katz et al., 2022 Garzon-Chavez et al., 2020 |
| Mycoplasma gallisepticum | Poultry Passerines Columbiformes |
No | Passerines Galápagos doves |
2 | Soos et al., 2008 Deem et al., 2011 |
|
| Plasmodium spp. (avian malaria) | Birds | No | Birds | 2 | Levin et al., 2009 Levin et al., 2013 Palmer et al., 2013 |
|
| Newcastle Disease Virus (avian paramyxovirus I) | Poultry Waterfowl Other birds |
No | Birds | 2 | Soos et al., 2008 Whitehead et al., 2018 Deem et al., 2011 |
|
| Cryptococcus gattii | N/A | Yes | No | Pinnipeds Galápagos penguins |
2 | Rosenberg et al., 2016 Fenton et al., 2017 Huckabone et al., 2015 Venn-Watson et al., 2014 Brito Devoto et al., 2022 |
| Infectious Bronchitis Virus | Chicken Pigeons (sporadic) |
No | Yes | Galápagos doves | 2 | Soos et al., 2008 Whitehead et al., 2018 Deem et al., 2011 Barr et al., 1988 |
| Infectious Bursal Disease Virus | Poultry Waterfowl |
No | Yes | Flightless cormorant Lava gull |
2 | Soos et al., 2008 Whitehead et al., 2018 Deem et al., 2011 |
| Toxoplasma gondii | Cats | Yes | Yes | Pinnipeds Birds |
2 | Levy et al., 2008 Deem et al., 2012b Deem et al., 2010 |
| Marek’s Disease Virus | Poultry | No | Yes | Unlikely | 1 | Soos et al., 2008 Deem et al., 2012a Gottdenker et al., 2005 Wikelski et al., 2004 |
| Zika virus | NHP | Yes | Yes | Unlikely | 1 | Bueno et al., 2016 |
| Ancylostoma caninum | Dog | Yes | Yes | Unlikely |
1 | Gingrich et al., 2010 Diaz et al., 2016 |
| Toxocara canis | Dog | Yes | Yes | Unlikely | 1 | Gingrich et al., 2010 Diaz et al., 2016 |
| Bartonella spp. | Cats Dogs |
Yes | Yes | Pinnipeds | 1 | Levy et al., 2008 Morick et al., 2009 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



