
Submitted:
31 October 2023
Posted:
01 November 2023
You are already at the latest version
Abstract
Current food production methods and consumption behaviours are unsustainable and contribute to environmental harm. One example is food waste around 38% of food produced is wasted each year. Here, we show that two common food waste products, wheat bran and brewer’s spent grain, can successfully be upcycled via miso fermentation. During the fermentation process, kokumi g-glutamyl peptides, known to increase mouthfulness, are produced; these include g-ECG (oxidized), g-EVG, g-EV, g-EE, g-EF, and g-EL. The profiles of kokumi peptides and volatile aroma compounds are correlated with koji substrate, pH, and enzymatic activity, offering straightforward parameters that can be manipulated for increasing the abundance of kokumi peptides during the fermentation process. Correlation analysis demonstrates that some volatile aroma compounds, such as fatty acid ethyl esters, are correlated with kokumi peptide abundance and may be responsible for the fatty, greasy, and buttery aroma. Consumer sensory analysis conveys that the bitter taste of vegetables, such as that in endives, can be dampened when miso extract containing kokumi peptides is added. This suggests that kokumi peptides, along with aroma volatile compounds, can enhance the overall flavor in plant-based products. This study opens new opportunities for cereal processing by-product upcycling via fermentation, ultimately having the potential to promote a plant-based diet.
Keywords:
kokumi
; fermentation
; upcycled miso
; sustainability
; aroma
; side stream utilization
; bitterness
1. Introduction
The planet is facing a global climate crisis. This is primarily attributed to unsustainable consumption and production patterns, of which the global food system is one of the major contributors [1,2]. Current industrialised meat production is a key driver of climate change [3], largely due to the high feed to food conversion ratios, resulting in systems that are resource intensive with low production efficiency compared to plant-based foods [4]. Another inefficiency of the current food system is its wastefulness, with 38% of food produced being wasted annually world-wide [5]. Only small amounts are currently being repurposed as animal feed or for other purposes [6]. Reducing meat consumption, shifting to more plant-based diets, and reducing food waste have therefore been proposed as key strategies for transitioning to more sustainable food systems [2].
To facilitate the necessary rapid transition in food consumption behaviours, taste is key [7]. However, one of the challenges with plant-based raw materials is that they often lack the umami, sweetness, mouthfeel and aroma compounds characteristic of animal products [8]. “Green” or “beany” aromas such as hexanal, (E)-2-nonenal, and (E,E)-2,4-decadienal, among others, present a challenge in plant-based ingredients [9].
Another challenge with many plant ingredients, for example vegetables belonging to the Brassica and Cichorium genera, is that they have an inherent bitterness due to the presence of various bitter compounds such as glucosinolates [10,11], which are often less favoured by consumers [12].
Fermentation has been used to preserve food but also to improve the flavour and the bioavailability of nutrients from raw materials [13]. In particular, γ-glutamyl peptides have been shown to increase ‘mouthfeel’ amongst other desirable qualities and has been proposed as a flavour enhancer in food [14]. Additionally, γ-glutamylization is being researched as a means of reducing bitterness by synthesis of peptides such as γ-Glu-Phe, γ-Glu-Leu or γ-Glu-Val, from bitter amino acids like Phe, Leu, and Val through GGT (gamma-glutamyl transferase) catalysis [15]. These γ-glutamyl peptides cause a sensation called koku, a Japanese word describing mouthfeels of thickness, richness, complexity, and continuity, and have been shown to be present in different fermented products like beer, cheese, soy sauce, sake, and miso [15,16] . Additionally, a concept called the “umamification” of plant-based materials suggests that fermentation can increase the concentration of amino acids and simple sugars to improve flavour [7], and kokumi peptides have been shown to increase umami [17], suggesting fermentation as a promising strategy for enhancing the favourable qualities of plant-based foods.
Previous studies have shown that kokumi peptides are present in miso prepared with alternative grains [16]. We were therefore interested in whether this concept can be expanded to additional substrates, especially cereal processing by-products, as a strategy to revalorise and reduce food waste. Wheat bran, a side product of milling of wheat into white flour, and brewer’s spent grains (BSG), a by-product of beer filtration, are amongst the most common food production by-products; respectively around 90 million tons and 32.5 tons are produced annually world-wide [6,7,8,9,10,11,12,13,14,15,16,17,18].Combined with the high protein and other nutritional concentrations remaining after processing, both by-products are promising potential candidates for producing kokumi peptides [19,20], as well as improving the bioavailability of nutrients following fermentation [13].
In this study, we aimed to repurpose cereal processing by-products through miso fermentation as an approach to create a flavouring product that can be added to plant-based ingredients to enhance their desirability and favourable taste attributes. We particularly focussed on the generation of kokumi peptides through miso fermentation and investigated their relation to physicochemical properties and aroma compounds, as well as the potential to reduce bitterness – while increasing umami and mouthfulness – by performing sensory analysis. The intention of the study is to propose new strategies through exploring the role of taste to enable the necessary transitions towards more sustainable future food systems and diets.
2. Materials and Methods
2.1. Preparation of Koji and Miso
Dried pearl barley (1 kg, Aurion, Denmark) was rinsed under cold water and then soaked in 4 L of tap water overnight at 7 ⁰C [21]. Wheat bran (Kornby Mølle, Lynge, Denmark) and brewer’s spent grains (BSG) (small brewery, Copenhagen, Denmark) were mixed with distilled water at a 1:1 ratio. Raw materials (Table 1) were steamed at 100 ⁰C for 90 min and allowed to cool to 30 ⁰C before inoculating with 0.3 % (w/w) koji spores. Two A. oryzae strains were used: S1, which is traditionally used for soybean miso (BF-3; Higuchi Matsunosuke Shoten Co. Osaka, Japan), and S2, which is traditionally used for barley koji (mugi senyo ki; Bio’c CO., Ltd Uchida, Murocho, Toyohashi-shi, Aichi, Japan). The temperature was kept at 31 ⁰C, and the relative humidity around 95 % during the koji fermentation. Miso was prepared as shown in Table 1, with a ratio of 1 koji: 2 raw material, 4 % (w/w) of salt, then kept at 28 ⁰C for 12 weeks. The samples were taken at 0, 4, 8 and 12 weeks and held at -20 ⁰C until analysis.
2.2. Physico-Chemical Parameters
To follow the miso evolution, physicochemical analyses were conducted after 0, 4, 8, and 12 weeks of fermentation. The physicochemical qualities of moisture, colour, pH, REDOX, and reducing sugar content were analysed according to previous studies [22,23]. All measurements were conducted in triplicate, except colour which was measured 10 times.
2.2.1. Colour Measurement
The colour was measured using a colour analyser CR400 (CR400, Konica Minolta, Inc, Tokyo, Japan) and analysed with SpectrMagic NX software, using the CIE L*a*b* colour space, illuminant D65, and a 10° observer as a reference. Calibration was done with a white standard board (L*=100.5, a*=–2.4, b*=5.6). L* indicates lightness (range 0-100), and a* and b* are the chromatic coordinates; a* has positive values for reddish colours and negative values for greenish colours, whereas b* is positive for yellowish and negative for bluish. Data were expressed with the L*a*b* values. Processing of the miso in a Thermomix® contributed to a uniform colour [16].
2.2.2. Water Content, pH, and REDOX
Water content (%) was measured using a moisture analyser (XM-124-60 124/0.001 g, COBOS®, Barcelona, Spain) [24]. The pH was analysed on 10 % (w/v) miso samples diluted in distilled water using a pH meter (Elite pH pocket testers, ThermoFisher Scientific, Waltham, MA, USA). The oxidation-reduction potential (REDOX) was measured by ORP/Redox tester (ORPTestr® 10BNC, Oakton, Cole-Parmer, Cambs, UK) on a solution of 10 % (w/v) miso samples diluted in distilled water [16].
2.2.3. Reducing Sugar content with Dinitrosalicylic Acid (DNS)
Reducing sugar concentration was measured on 1% (w/v) miso samples diluted in distilled water and filtered through Whatman no.2 filter paper (Whatman International Ltd., Maidstone, Kent, UK). One mL of dinitrosalicylic acid (DNS) (Sigma-Aldrich, Madrid, Spain) and 1 mL of miso were mixed and heated at 100 ºC for 5 min while stirring (MSH-300i, bioSan, Riga, Latvia), then immediately cooled on ice [25]. The absorbance was recorded at 540 nm with a UV spectrophotometer (Cary 100 Series 2 Scan, Varian, Madrid, Spain), and the standard curve and results obtained in terms of ACS grade anhydrous D (+)-Glucose content (mg/g) PA-ACS (PanReac AppliChem GmbH, Darmstadt, Germany) [16−22].
2.3. Protease Activity
Protease activity was measured following a modified previously described method [16, 22−23]. Two grams of miso were mixed with 40 mL distilled water and filtered with Whatman no. 1 filter paper (Whatman International Ltd., Maidstone, Kent, U.K). One mL of miso solution was mixed with 3 mL of 0.6 % casein solution (Alfa Aesar, Ward Hill, MA, USA) and kept in a shaking water bath at 37 °C for 10 min. Then, 5 mL of 0.4 M trichloroacetic acid (VWR Chemicals, Barcelona, Spain) was added to the miso sample and incubated for 20 min at 37°C to stop the enzymatic reaction. The solution was centrifuged at 1,147xg for 5 min (VWR Mega Star 600, Radnor, PA, USA), and 2 mL of the supernatant was mixed with one mL of Folin-Ciocalteu reagent (VWR Chemicals, Barcelona, Spain), followed by addition of 5 mL of 0.4 M Na2CO3 (VWR Chemicals, Barcelona, Spain). After 30 min, absorbance was measured at 660 nm using a UV-visible spectrophotometer (UH5300 Spectrophotometer, Hitachi, Ltd, Tokyo, Japan). For the standard curve, tyrosine (L-Tyrosine 99%, Alfa Aesar, Ward Hill, MA, USA) was used, and one unit of protease activity was expressed as the amount of tyrosine produced from 1 mL of the koji solution after one minute. All measurements were taken in triplicate.
2.4. Extraction and Identification of Kokumi Peptides by HPLC/Ms-Ms
Six different peptides were analysed based on suggestions from the literature: γ-Glu-Cys-Cly (analysed in its reduced and oxidized forms) (Sigma Aldrich, St. Louis, MO, USA), γ-Glu-Val-Gly (Trichem, Skanderborg, Denmark), γ-Glu-Val [26] , γ-Glu-Glu, γ-Glu-Leu and γ-Glu-Phe (Bachem, Frechen, Germany) [27]. Identification was carried out by targeted detection of peptides with standards. Quantifications are based on the chromatography areas.
Samples were prepared by adding 80% (v/v) of methanol (EMD OmniSolv® LCMS Methanol, Sigma Aldrich, St. Louis, MO, USA) to 800 mg of sample. Samples were homogenised by vortex for two minutes and centrifuged at 9,000 rpm for 20 min (Eppendorf® Centrifuge 5424 RG, Eppendorf North America, Enfield, CT, USA). The upper solid fat layer and the protein pellet were discarded, and the liquid layer retained, membrane-filtered (SPARTAN HPLC Syringe Filter, 13 mm, Regenerated Cellulose, 0.45 µm, Whatman International Ltd., Madstone, Kent, U.K), and transferred to a chromatography vial. All samples were kept at -20 °C until analysis [16].
Samples were analysed by LC-MS on a Vanquish LC coupled to an ID-X MS (ThermoFisher Scientific, Waltham, MA, USA). Five µL of sample or standard was injected on a ZIC-pHILIC peek-coated column (150 mm x 2.1 mm, 5-micron particles, maintained at 40 °C, SigmaAldrich). Buffer A was 20 mM Ammonium Carbonate and 0.1% Ammonium hydroxide in water, and Buffer B was Acetonitrile 97% in water. The LC program was as follows: starting at 93% B, to 40% B in 19 min, then to 0% B in 9 min, maintained at 0% B for 5 min, then back to 93% B in 3 min and re-equilibrated at 93% B for 9 min. The flow rate was maintained at 0.15 mL min-1, except for the first 30 seconds, where the flow rate was uniformly ramped from 0.05 to 0.15 mL min-1. Data was acquired on the ID-X in switching polarities at 120000 resolution, with a RF lens at 30%, normalized AGC target at 25%, max IT at 50 ms, and m/z range: 120-500. For each peptide target, a targeted MS2 with isolation 1.6 m/z, HCD 20, 30,000 resolution in positive mode was also acquired (Supplemental Materials Table S1). For each peptide, a unique MS2 fragment was selected for integration. The fragments were selected based on the pure standards MS2 spectra. These unique fragments, in addition to retention time differences, allowed for the differentiation between isobaric species [16].
2.5. Extraction and Quantification of Volatile Aroma Compounds by HS-SPME-GC/MS
For each sample, 20.0 g of miso was homogenised with 44.0 g of ultrapure water (Arium® Pro Ultrapure Water System, Sartorius, Goettingen, Germany) and shaken at 25⁰C for 2 h. The homogenate was filtered with folded filter paper (grade MN 615 1/4, 185 mm), and 4 mL were transferred to 10 mL gas-tight vials. Ten microliters of 2-methyl-3-heptanone (Sigma Aldrich, St. Louis, MO, USA) (27.2 mg/L in methanol) was added to the homogenate as an internal standard [21]. The negative control was prepared with 20.0 g steamed barley, 4% (w/w) salt, 44.0 g distilled water, and 10.0 µL of internal standard. This volatile compound extraction method has been previously used to analyse different miso and soy sauces [16−21].
Solid-phase microextraction (SPME) fibre, polydimethylsiloxane/divinylbenzene (PDMS/DVB), 65 µm, was used to extract volatile compounds. Following extraction, the samples were incubated at 45°C for 15 min, after which the SPME fibre was inserted into the headspace at 45 °C for 40 min and then inserted into the GC injection port. The GC-MS analysis was performed on a Thermo Scientific TRACE 1310 Gas Chromatograph equipped with a Thermo Scientific Q Exactive Orbitrap mass spectrometry system with a Thermo fused-silica capillary column of cross-linked TG-5SILMS (30 m x 0.25 mm x 0.25 µm) (ThermoFisher Scientific, Waltham, MA, USA). The GC conditions were as follows: inlet and transfer line temperatures, 250˚C; oven temperature program, 40˚C for 2 min, 5˚C/min to 120˚C for 2 min, 7˚C/min to 220˚C for 5 min, 50˚C/min to 325˚C for 3 min; inlet helium carrier gas flow rate, 1 mL/min; split ratio, 20:1. The electron impact (EI)-MS conditions were as follows: ionisation energy, 70 eV; ion source temperature, 250˚C; full scan m/z range, 30 - 350 Da; resolution, 60,000; AGC target, 1e6; maximum IT, 200ms [16−21]. Retention index based on Thermo TG-5SILMS column using C7-C27 as external references. Concentration is expressed as 2-mehtyl-3-heptanone equivalent (ug/L). Method of identification: A, by comparison of the MS spectra with the NIST library; B, by comparison of RI (Kovat indices). All the compounds were identified by A and B methods. Data were acquired and analyzed with Thermo TraceFinder 4.1 software package (ThermoFisher Scientific, Waltham, MA, USA) [28].
2.6. Sensory Analysis
A kokumi rich water-soluble extract was prepared from each miso following the Water-Soluble Extract protocol (WSE) [29]. Each of the three miso samples (Table 1, 50 g) were mixed with filtered water at a ratio of 1:1 (w/w), autoclaved for 120 min at 40 kPa and 110 ºC, and filtered through Whatman no.2 filter paper (Whatman International Ltd., Maidstone, Kent, UK), with the aqueous phase obtained. The residue on the filter paper was re-extracted with water (42 mL) at room temperature and re-filtered. Both water extractions were combined and centrifuged at 9338xg for 15 min, and the liquid phase reserved. Endive purée (prepared at a ratio of 2:1 endive:olive oil) was cooked at 180 ºC for 20 min in the oven in vacuum bags and blended with a Thermomix® (full speed, for 4 min). Extracts from each of the three miso samples were added to three independent samples of endive puree (20 % w/w). A control sample was prepared with endive pure and 20 % (w/w) of tap water instead of miso extract.
For the consumer sensory analysis each sample (30g) was served at room temperature to a panel of 60 consumers in a room with controlled temperature and relative humidity (21 ºC ± 2; 35 ± 5 % RH). Consumers tasted each sample and rated the intensity of bitterness, coating, aftertaste, and thickness using a 100-point general Labelled Magnitude Scale (gLMS) (0 = barely detectable, 100 = strongest imaginable) [30]. Aftertaste intensity was rated 20 seconds after placing the sample in the mouth and thickness after 5 seconds. Answers to the questionnaire were collected by RedJade Sensory Software (RedJade Sensory Solutions, LLC, 2023). The samples were randomly coded with 3-digit numbers and offered to the consumers in random order. Consumers rinsed their mouths with water and unsalted crackers between samples.
The sensory analysis followed Regulation (EU) 20216/679 on the protection of personal data. The regulation and the experimental procedure were explained to the consumers and consent for voluntary participation was given by consumer signature. The study was conducted in accordance with the Declaration of Helsinki, and the protocol was approved by the Ethics Committee of IEB-20220927-I.
2.7. Data Analysis
A one-way ANOVA test was conducted to find significant differences between samples taken at different times. A post-hoc test was carried out using Tukey’s HSD. All data analysis was performed by the statistical package XLSTAT Version 2020.4.01 (Lumivero, Denver, CO, USA) [31]. Results were considered significant for p < 0.05. The correlation plot, heat map and cluster analysis were produced with RStudio Desktop 2022.07.1+554 (RStudio, Boston, MA, USA) [32], with 95 % confidence level and 0.001, 0.01, and 0.05 significance levels for the correlation analysis.
3. Results and Discussion
3.1. Miso Fermentation of Cereal Processing by-Products Generates γ-Glutamyl Peptides
Kokumi peptides have been shown to enhance the palatability and perceived mouthfulness of foods [33]. In a previous study, we have demonstrated that miso fermentation of regional cereal grains results in the generation of γ-glutamyl peptides [16], we were therefore interested in whether miso fermentation of cereal byproducts is able to produce kokumi peptides as well. We selected two main cereal production and processing by-products, wheat bran and brewer’s spent grain (BSG), and introduced them at different stages of the miso fermentation. Wheat bran was used as the key substrate for preparation of koji in this study based on evidence of wheat bran being utilised in traditional and industrial koji making practices [34,35,36,37], and BSG was used as the miso substrate based on its high residual protein and nutrient contents that is comparable to other common miso substrates [6−20]. We also included barley, a substrate that was demonstrated to produce γ-glutamyl peptides through miso fermentation [16], and tested different combinations of miso fermentation as detailed in Table 1. To investigate the potential effect of the genetic influence of the koji fungus, A. oryzae, on the fermentation process, we also tested different strains of A. oryzae (S1 and S2) for specific combinations (Table 1).
Our results show that all but one of the γ-glutamyl peptides that were identified in grain-based miso fermentation [16] were also present in the miso samples prepared using cereal-processing by-products, including γ-ECG (oxidized), γ-EE, γ-EL, γ-EF, γ-EV, and γ-EVG. Likewise, the reduced form of γ-ECG was not detected, most likely due to natural oxidation during the fermentation process (Figure 1) [16].
Cluster analysis of the different kokumi peptides and miso shows that the miso made with wheat bran koji are clustered into one group, and the one made with barley koji clustered into another (Figure 1A). This observation demonstrates that koji substrate is an important parameter for the abundance and identity of kokumi peptides in subsequent miso fermentations as suggested previously [16].
Looking more closely at the individual γ-glutamyl peptides identified, γ-ECG oxidize was present in all of the samples after 12 weeks of fermentation (Figure 1A). Notably, the abundance of γ-ECG is inverse to the abundance of γ-EVG and γ-EV in the different samples (Figure 1A). This trend might be explained by the mechanism suggested by Sofyanovich et al. [38] , where γ-ECG is the precursor of γ-EV and γ-EVG. The synthesis of γ-EV and γ-EVG is mediated through the transfer of the γ-glutamyl residue from γ-ECG to valine via the GGT (glutamyl transferase) pathway or the dipeptide Val-Gly (VG), respectively. The abundance of γ-EVG is promising because it is the most potent kokumi peptide found and studied in a wide range of fermented products [33].
Interestingly γ-EE was identified in all samples at the end of the fermentation process (Figure 1A). In our previous study on alternative miso, this peptide was identified in barley koji, so this confirming the relation between koji substrate and the generation of kokumi peptides [16]. The mechanism of γ-glutamyl dipeptides is led by γ-glutamyl transfer reaction via GCL and GCS. Thus, free amino acids such as glutamic acid are a limiting factor to synthesise γ-EE [39].
Peptide γ-EL was found in all miso samples, with higher abundance observed in the two misos that were made using BSG as the miso substrate (Figure 1A). This could be explained by the higher concentration of free leucine in BSG (6.1 w/w %) than in in barley (0.3 w/w %) [40]. This peptide has also been reported to be present in various fermented foods, including soy sauce, sourdough, parma dry-cured hams, gouda, and parmesan cheese, as well as occurring naturally in various edible beans [33].
By contrast, γ-EF was only detected in wheat bran S1/BSG in the last stage of the miso fermentation (Figure 1D). The abundance of this dipeptide is slightly decreased during the fermentation time in the remaining samples until it reaches zero (Figure 1B–D). It has been found in various fermented products, such as miso and soy sauce [33].
3.2. Physicochemical Changes in Fermentation Processes Influence the Generation of γ-Glutamyl Peptides
It is evident from this study as well as previous studies, that components in the raw materials used to prepare koji contribute to the formation of precursors such as γ-ECG and the release of different amino acids [16]. Our previous results have also shown that physicochemical changes occurring during the fermentation process, including pH, REDOX, reducing sugar and color, are factors that are correlated with differential kokumi peptide formation [16]. Since physicochemical changes can both influence and be influenced by related enzymatic activities during the fermentation process, monitoring the physical and chemical parameters (Supplemental Materials Table 2) not only provides a better understanding of the production of kokumi γ-glutamyl peptides, but can also become a useful tool for tracking the progress of fermentation, especially for unconventional substrates like by-products.
In this study, we measured the changes in pH levels, protease activity, reducing sugar, water content, and colour, across all samples throughout the 12-week fermentation period, taking samples every 4 weeks. During the fermentation process, the pH levels of 3 of the 4 misos (wheat bran S1/barley, wheat bran S2/barley, and barley S1/BSG) followed a similar pattern, whereby the pH level initially decreased from 6 ±0.06 to 4 ±0.06 in the first eight weeks, then increased slightly back to around 6 in the final stage of the fermentation (Figure 2A). This pattern of modulation in pH profile is consistent with what we observed in our previous studies of miso fermentation [40−16] In contrast, the pH level of the miso made from wheat bran S1/BSG showed an unusual pH profile, where a gradual increase from 6.63 ±0.06 to 9.43 ±0.06 (Figure 2B) was observed over the 12-weeks, without the pH ever reaching below 6, indicating an unusual fermentation process. This unexpected change in pH seem to have an impact on the abundance of the γ-EV peptide in particular (Figure 2B). This pattern and difference suggest that the combination of substrates impact the pH behaviour, and consequently kokumi peptide generation. In addition to being a good indicator for monitoring the fermentation process, pH changes may also serve a functional role for the synthesis of γ-dipeptides such as γ-EE, γ-EF and γ-EL, as it influences the ionic form of amino acids, like glutamic acid to glutamate. Glutamate is also an important amino acid for the γ-glutamylation reaction and have been implicated in dampening bitterness from phenylalanine, leucine, and valine [41], a taste attribute that we will further explore in Section 3.4.
We observe in this study that pH levels are strongly correlated with the changes in enzymatic activities during fermentation (p<0.01), as shown in Supplemental materials Figure 1. The results show the lowest activity for barley S1/BSG (0.80±0.01 U/g), followed by wheat bran S2/ barley (1.09±0.02 U/g), wheat bran S1/BSG (1.23±0.01 U/g) and the highest for wheat bran S1/barley (1.70±0.02 U/g) (Supplementary Table 1) for the final time point. We also found that the koji and miso substrates are not the only determing factors for the fermentation process. Based on results from the two misos that used the same koji and miso substrates (wheat bran S1/barley and wheat bran S2/barley), we observed very different enzymatic activities (Figure 2C) . This suggests that the A. oryzae strain also has an influence on the enzymatic activity during fermentaion. According to a study by Lin et al. in [42], protease activity reaches its highest point at pH levels ranging from 6-7, depending on the specific A. strain. Meanwhile, the synthesis of γ-glutamyl peptides involves three distinct enzymes, including GGT (γ-glutamyl transferase), glutaminase, and either GCS (γ-glutamylcysteine synthetase) or GCL (glutamate-cysteine ligase). In vitro, these enzymes catalyse the creation of γ-glutamyl dipeptides from glutamate and amino acids with an optimal pH above 7.5 [43]. These observations suggest that the strain of A. oryzae used could also have an influence on γ-glutamyl peptide formation and remains to be further explored.
Another physicochemical property we investigated were the parameters in relation to colour (a*, b* and L*). Our previous study showed that a* and b* were also correlated with kokumi peptides, particularly for γ-EL and γ-EV [16]. According to a study by Wang et al. in [44], the impact of colour on consumer preferences in product appearance was examined, where on average, high a* (reddish) was associated with higher miso quality, whereas b* (blueish) and L* (luminosity) showed the opposite effect. Luminosity (L*) affects the darkness/reddish characteristic of miso and is closely related to its palatability and acceptance. The study described in this paper, found that L* was negatively correlated with γ-EE, γ-EF (p<0.05), and γ-EL (p<0.01) (Supplemental Materials Figure S1). We also observed that L* is negatively correlated with pH (Supplemental Materials Figure S1) and there is an especially pronounced decrease in L* accompanying the increase in pH for the wheat bran S1/BSG miso (Figure 2D). Based on our earlier conclusion of pH as a potential indicator for fermentation progress (Figure 2D), as well as subsequent characterisations of volatile aroma profile in section 3.3, this suggests that luminosity could serve as a potential indicator for predicting the desirable characteristics for miso-based fermentation.
3.3. Volatile Aroma Compound Profiles of Upcycled Miso Reveal Correlation with g-Glutamyl Peptides
Aroma plays an important role for food acceptance. Some interactions of plant-based protein and aromas have been studied, suggesting that compounds such hexanal, (E)-2-nonenal, and (E,E)-2,4-decadienal, among others, can lead to consumer rejection. However, fermentation is able to reduce these compounds, making plant-based ingredients more palatable [9].
To explore this further, we conducted an HS-SPME-GC/MS analysis and found a total of 133 volatile organic compounds across the different miso samples (Supplemental Materials Table S3). We first performed a correlation analysis between the kokumi peptides and volatile aroma compounds and identified 67 compounds that showed correlative association with the γ-glutamyl peptides (Supplemental Materials Figure S2), and 63 in particular that were present in the last stage of the miso fermentation (Figure 3). These compounds included three acids, four alcohols, six aldehydes, 31 esters, ten ketones, five phenols, and four pyrazines. These results corresponded with our previous study on alternative grain-based miso, where the most represented volatile aroma group present was esters [16].
Cluster analysis of the volatile aroma compound profiles revealed that 3 of the misos (wheat bran S1/barley, wheat bran S2/barley and barley S1/BSG) belong to the same cluster, whereas the miso made of wheat bran S1/BSG belongs to a separate second cluster (Figure 3). This observation may be related to the unexpected differences in physicochemical changes that were exhibited during the fermentation process, especially the non-acidic pH levels for the wheat bran S1/BSG miso (Figure 2B), which could serve as a potential explanation for the unusual aromatic attributes of this sample, a point that we will return to later in this section.
First focusing our analysis on the volatile aroma profile of the main clusters, we observe that the esters were amongst the group of volatile compounds most strongly associated with kokumi peptides, especially high fatty acid ethyl esters like hexadecanoic acid methyl ester, hexadecanoic acid ethyl ester, 9,12-octadecadienoic acid (Z, Z)-methyl ester, elaidic acid methyl ester, linoleic acid ethyl ester, and ethyl oleate (Figure 3). This observation is in line with our previous study on alternative grain-based miso, where we also observed high concentrations of ester aroma compounds due to the degradation of fatty acids [16]. These molecules are known to have waxy, fatty, oily and fruity aromas. Most of them are produced during long-term fermentation processes lasting 3-6 months [45,46]. They might be involved in the relationship between the koku sensation and fat, contributing to a mouthfeel of coating, lingering and aftertaste, – as has also been suggested in a previous study that showed that kokumi peptides can enhance the fatty taste [26].
Compounds that cause fruity aromas are also present prominently in the main cluster; these include compounds from the ester group, such as isobutyl acetate, 2-methyl-butanoic acid- ethyl ester, 3-methyl-butanoic acid- ethyl ester and isoamyl lactate, as well as phenylethyl alcohol (Figure 3); the latter has been associated with a rose-honey-like aroma [44]. Phenylethyl alcohol is particularly noteworthy, as it is found to be present in all 3 misos in the cluster (Figure 3) and has been reported to be produced by degrading the amino acid phenylalanine [46], which could be a possible explanation for why these misos do not contain γ-EF (Figure 1A.).
Within the main cluster, the 3 misos can be further sub-clustered into two different groups based on their volatile aroma profiles. Interestingly, wheat bran S1/ barley and wheat bran S2/barley did not cluster into the same group, suggesting that the strain of A. oryzae also has a notable influence on volatile aroma profile of the final miso product, which may be influenced by the differences in inherent enzymatic activities of the strain of fungi, as observed in Figure 2C. Nonetheless, we still observe a few aromatic compounds in common, including 2-methoxy-4-vinylphenol, which is often found in soy sauce, and is associated with a burnt scent [47], and trimethyl-pyrazine, which is associated with roasted and chocolate aromas [48]. These compounds can likely be attributed to the presence of lignin in wheat bran and provides evidence for koji and miso substrates still contributing towards the final aromatic profile of the fermented miso.
In the miso made from wheat bran S1/barley, we also detected other aroma compounds that are found in many fermented products, including red sufu, cacao beans, and soybean products, and are known for their sweet, fruity, and floral notes [49]. In particular, benzaldehyde, α-ethylidene, are formed through the degradation of phenylalanine [49] and cyclohexanol, 5-methyl-2-(1-methylethyl)-, (1α,2α,5α)-, creates a menthol aroma [50].
In terms of the other cluster, the miso made up of wheat bran S1/BSG exhibited a high concentration of phenol compounds, including 2,3-dimethyl, which is not typically associated with miso flavour. This phenomenon may be attributed to the breakdown of the high concentration of lignin in cereal bran and BSG, producing phenol compounds [44]. Other phenolic compounds detected include, 4-ethyl-2-methoxy, which imparts smoky and spicy notes [51], and 2-phenylpropenal, a compound responsible for the green, honey, alcoholic, sweet, caramel, bread, and coffee aromas found in fermented rice bran [45]. Compounds belonging to the pyrazine group, such as trimethyl-pyrazine, which is associated with a roasted aroma [49] were detected in higher concentrations than in the other two misos made from wheat bran koji (Figure 3). This high concentration of pyrazine compounds may be attributed to the high pH during the fermentation of this miso, which could accelerate the formation of pyrazines through the Maillard reaction between reducing sugars and amino acids [52].
Overall, we observe a strong negative correlation between volatile aroma compounds and γ-glutamyl peptides (Figure 4). This observation may be explained by the cabalism of amino acids; for instance, aldehydes and alcohols are the primary compounds produced through the Ehrlich pathway by various fungal enzymes during fermentation [21] and Strecker degradation [53]. Therefore, the synthesis of kokumi peptides may be hindered as a result of the degradation of amino acids.
Volatile aroma compounds and kokumi peptides may intensify the experience of the koku sensation, a Japanese term used to describe the presence of kokumi peptides. The link between high levels of fatty acid ethyl esters and kokumi peptides could be a possible explanation for these characteristic associations with the koku sensation, such as aftertaste, lingering and coating, and remains a question to be further explored.
3.4. Addition of Kokumi Peptides Reduces Bitter Taste
Bitterness is not a preferred taste for many people, despite the bitter causing compounds, for example isothiocyanates or glucosinolates, being present in many foods like vegetables or cereals, which are nutritious foods contributing to a healthy diet [54]. Similarly, the new generation of upcycling products often run into a similar problem of less desirable tastes including bitterness [55]. The taste of bitterness, common in many plant-based food, is one of the factors preventing wider acceptance and changes in dietary habits when it comes to consumer choices towards more ‘planetary healthy’ food options [8].
Kokumi peptides have been studied to increased umami, salty, sweet taste, and fatty texture [17]. In particular, bitterness has been implicated to be reduced by fermentation through γ-glutamylation reactions [39]. Based on this observation, we tested whether the γ-glutamyl peptides produced during the fermentation of our upcycled miso was able to reduce bitterness when added to plant-based ingredients. We selected an endive (Cichorium intybus var. foliosum) as the vegetable for our sensory analysis due to its distinct bitter taste produced by glucosinolates [12]. Using a method of water extraction, we obtained an extract from each of our upcycled misos that was rich in kokumi peptides, and after adding them to the endive purée in the same concentrations, performed sensory analysis for each of the samples (for details, see Materials and Methods). It should be noted that extraction and sensory analysis were only performed for 3 of the 4 misos (barley S1/BSG, wheat bran S1/barley, and wheat bran S2/barley). The wheat bran S1/BSG miso was excluded due to the undesirable aroma profile and unexpectedly high pH observed throughout the fermentation. For the sensory analysis, an intensity scale was used to evaluate the bitterness, aftertaste, coating and thickness. The questionnaire also included questions about the participants’ attitudes to vegetables, in order to establish whether their preferences and previous experiences might affect their perception of bitterness.
Results from the sensory analysis confirmed our hypothesis that the addition of kokumi rich miso extracts could reduce the perceived bitterness in the endive samples; we observed that this was the case for two of the three samples tested. The endive sample containing extract from the barley S1/BSG miso was perceived to be the least bitter, followed by wheat bran S1/barley, control (water instead of extract added), and wheat bran S2/barley (Figure 5). This result may partly be explained by the total abundance of γ-glutamyl peptides in each sample (Figure 5B), where barley S1/BSG miso has the highest amount, followed by wheat bran S2/barley and wheat bran S1/barley. Interestingly, despite wheat bran S1/barley having a lower abundance of γ-glutamyl peptides overall it shows a higher absolute quantity of γ-EVG than wheat bran S2/BSG (Figure 5B). γ-EVG has been reported as a potent kokumi peptide with a sensory activity that is 12.8-fold stronger than GSH [56]. The higher abundance of γ-EVG in the wheat bran S2/BSG could therefore be a possible explanation for why its addition to endive purée results in lower perceptions of bitterness than the wheat bran S1/BSG sample.
In terms of the other attributes assessed in the sensory analysis, aftertaste was the only attribute other than bitterness that significantly differed between samples (Table 2). This could be the result of the lingering bitterness, especially for the sample containing the extract from the wheat bran S2/barley, which was perceived to be the most bitter and was ranked the highest for aftertaste (Figure 5A). Other studies have shown that the extent and impact between the bitter taste and aftertaste may be related to the regional distribution of bitter taste receptors on our tongue [57]. Future experiments should investigate whether γ-glutamyl peptides have differential effects on different bitter taste receptors.
Finally, we found that study participants who reported liking vegetables perceived the bitterness as being lower than those who did not. Furthermore, the perceived reduction in bitterness was also greater amongst those participants who reported eating vegetables more often (Supplemental Materials Table S4). These observations are in agreement with the study by Vecchio et al. [12], where positive preferences toward bitterness was correlated with greater compensatory health beliefs. This opens up a promising area of research for influencing consumer preferences towards more plant-based diet options by reducing the less desirably flavour attributes of plant-based ingredients.
4. Conclusions
In conclusion, this study demonstrates the possibility of producing γ-glutamyl peptides through miso fermentation of two common cereal processing by-products, wheat bran and BSG. Overall, we observed that the choice of koji and miso substrates played a crucial role in the total abundance of kokumi peptides at the end of the 12-week fermentation, with the combination of using barley as the koji substrate and BSG as the miso substrate generating the greatest absolute concentration of peptides overall. Physicochemical characterisation during the fermentation process revealed that the optimum pH for the production of the peptides is between 5 and 6. Analysis of the volatile aroma profiles of the respective misos demonstrated that particular volatile compounds, specifically high fatty acid ethyl esters, are correlated to the abundance of kokumi peptides. Given these observations, the choice of fermentation substrates, as well as monitoring physicochemical changes like pH, are suggested to be important factors for creating favourable conditions to generate more desirable flavour attributes like kokumi peptides and the volatile aroma compounds when fermenting plant-based ingredients. Furthermore, we showed that extracts from the misos that were rich in kokumi peptides reduced the bitter taste in vegetables. To our knowledge this is the first time this has been demonstrated. These results show great potential for revalorising food production by-products through fermentation by creating flavouring products that can enhance the desirability and favourable taste attributes of plant-based ingredients. Such strategies can contribute towards shifting current food consumption patterns and transforming our food production systems to more planetary healthy and sustainable futures.
Supplementary Materials
The following supporting information can be downloaded at Supplemental Materials.
Acknowledgement
We thank Dr. Jennifer Wang and Dr. Charles Vidoudez at the Harvard Center for Mass Spectrometry for supporting technical aroma and peptide analysis. We also thank Dr. Caroline Isabel Kothe at the Novo Nordisk Foundation Center for Biosustainability at the Technical University of Denmark for helpful discussions and Dr. Olaia Estrada at BCC Innovation for lending access to the BCC Innovation colour analyser.
References
- Ravindran, R.; Jaiswal, A.K. Exploitation of Food Industry Waste for High-Value Products. Trends in Biotechnology 2016, 34(1), 58–69. [Google Scholar] [CrossRef] [PubMed]
- The Eat-Lancet Commission. 2019; Food Planet Health: Healthy Diets From Sustainable Food Systems. The Lancet.
- Agudelo Higuita, N.I.; LaRocque, R.; McGushin, A. Climate change, industrial animal agriculture, and the role of physicians – Time to act. Journal of Climate Change and Health 2023, 13(9), 2–6. [Google Scholar] [CrossRef]
- Garnett, T.; Little, D.; Röös, E. 2015; Lean, green, mean, obscene…? What is efficiency? And is it sustainable? Animal production and consumption reconsidered.
- Capanoglu, E.; Nemli, E.; Tomas-Barberan, F. Novel Approaches in the Valorization of Agricultural Wastes and Their Applications. Journal of Agricultural and Food Chemistry 2022, 70(2), 6787–6804. [Google Scholar] [CrossRef] [PubMed]
- Lynch, K.M.; Steffen, E.J.; Arendt, E.K. Brewers’ spent grain: a review with an emphasis on food and health. Journal of the Institute of Brewing 2016, 122(4), 553–568. [Google Scholar] [CrossRef]
- Mouritsen, O.G.; Styrbæk, K. Design and ‘umamification’ of vegetable dishes for sustainable eating. International Journal of Food Design 2020, 5(2), 9–42. [Google Scholar] [CrossRef]
- Mouritsen, O.G. Umamification of food facilitates the green transition. Soil Ecology Letters 2023, 5(1), 1–3. [Google Scholar] [CrossRef]
- Leonard, W.; Zhang, P.; Ying, D.; Fang, Z. Surmounting the off-flavor challenge in plant-based foods. Critical Reviews in Food Science and Nutrition 2022, 5(5), 1–22. [Google Scholar] [CrossRef]
- Hance, P.; Martin, Y.; Vasseur, J.; Hilbert, J.L.; Trotin, F. Quantification of chicory root bitterness by an ELISA for 11β,13-dihydrolactucin. Food Chemistry 2007, 105(2), 742–748. [Google Scholar] [CrossRef]
- Wieczorek, M.N.; Walczak, M.; Skrzypczak-Zielińska, M.; Jeleń, H.H. Bitter taste of Brassica vegetables: The role of genetic factors, receptors, isothiocyanates, glucosinolates, and flavor context. Critical Reviews in Food Science and Nutrition 2018, 58(8), 3130–3140. [Google Scholar] [CrossRef]
- Vecchio, R.; Cavallo, C.; Cicia, G.; Del Giudice, T. Are (All) consumers averse to bitter taste? Nutrients 2019, 11(2), 323–337. [Google Scholar] [CrossRef]
- Bertsch, A.; Roy, D.; LaPointe, G. Fermentation of Wheat Bran and Whey Permeate by Mono-Cultures of Lacticaseibacillus rhamnosus Strains and Co-culture With Yeast Enhances Bioactive Properties. Frontiers in Bioengineering and Biotechnology 2020, 8(8), 1–18. [Google Scholar] [CrossRef]
- Lu, Y.; Wang, J.; Soladoye, O.P.; Aluko, R.E.; Fu, Y.; Zhang, Y. Preparation, receptors, bioactivity and bioavailability of γ-glutamyl peptides: A comprehensive review. Trends in Food Science and Technology 2021, 113(9), 301–314. [Google Scholar] [CrossRef]
- Yang, J.; Bai, W.; Zeng, X.; Cui, C. Gamma glutamyl peptides: The food source, enzymatic synthesis, kokumi-active and the potential functional properties – A review. Trends in Food Science and Technology 2019, 91(5), 339–346. [Google Scholar] [CrossRef]
- Rodríguez Valerón, N.; Mak, T.; Jahn, L.J.; Arboleya, J.C.; Sörensen, P.M. Characterization of kokumi γ-glutamyl peptides and volatile aroma compounds in alternative grain miso fermentations. LWT Food Science and Technology 2023, 188(10), 1–9. [Google Scholar] [CrossRef]
- Nishimura, T.; Kuroda, M. Koku in Food Science and Physiology. Springer. 2019; Singapure. [Google Scholar] [CrossRef]
- Onipe, O.O.; Jideani AI, O.; Beswa, D. (2015). Composition and functionality of wheat bran and its application in some cereal food products. International Journal of Food Science and Technology 2015, 50(12), 2509–2518. [Google Scholar] [CrossRef]
- Balandrán-Quintana, R.R.; Mercado-Ruiz, J.N.; Mendoza-Wilson, A.M. (2015). Wheat Bran Proteins: A Review of Their Uses and Potential. Food Reviews International 2015, 31(3), 279–293. [Google Scholar] [CrossRef]
- Wen, C.; Zhang, J.; Duan, Y.; Zhang, H.; Ma, H. A Mini-Review on Brewer’s Spent Grain Protein: Isolation, Physicochemical Properties, Application of Protein, and Functional Properties of Hydrolysates. Journal of Food Science 2019, 84(12), 3330–3340. [Google Scholar] [CrossRef]
- Feng, Y.; Cui, C.; Zhao, H.; Gao, X.; Zhao, M.; Sun, W. (2012). Effect of koji fermentation on generation of volatile compounds in soy sauce production. International Journal of Food Science and Technology 2012, 48(3), 609–619. [Google Scholar] [CrossRef]
- Hong, H.H.; Kim, M.K. Physiochemical quality and sensory characteristics of koji made with soybean, rice, and wheat for commercial doenjang production. Foods 2020, 9(8), 1–10. [Google Scholar] [CrossRef]
- Kim, M.K.; Chung, H.J.; Bang, W.S. Correlating physiochemical quality characteristics to consumer hedonic perception of traditional Doenjang (fermented soybean paste) in Korea. Journal of Sensory Studies 2018, 33(6), 1–10. [Google Scholar] [CrossRef]
- Bechman, A.; Phillips, R.D.; Chen, J. Changes in Selected Physical Property and Enzyme Activity of Rice and Barley Koji during Fermentation and Storage. Journal of Food Science 2012, 77(6), 1–5. [Google Scholar] [CrossRef]
- Deshavath, N.N.; Mukherjee, G.; Goud, V.V.; Veeranki, V.D.; Sastri, C.V. Pitfalls in the 3, 5-dinitrosalicylic acid (DNS) assay for the reducing sugars: Interference of furfural and 5-hydroxymethylfurfural. International Journal of Biological Macromolecules 2020, 156(8), 180–185. [Google Scholar] [CrossRef] [PubMed]
- Miyamura, N.; Jo, S.; Kuroda, M.; Kouda, T. Flavour improvement of reduced-fat peanut butter by addition of a kokumi peptide, γ-glutamyl-valyl-glycine. Flavour 2015, 4(1), 1–4. [Google Scholar] [CrossRef]
- Kuroda, M.; Kato, Y.; Yamazaki, J.; Kai, Y.; Mizukoshi, T.; Miyano, H.; Eto, Y. Determination and quantification of the kokumi peptide, γ-glutamyl-valyl-glycine, in commercial soy sauces. Food Chemistry 2013, 141(2), 823–828. [Google Scholar] [CrossRef]
- ThermoFisher Scientific. 2016. TraceFinder Software. Waltham, MA, USA.
- Feng, T.; Wu, Y.; Zhang, Z.; Song, S.; Zhuang, H.; Xu, Z.; Yao, L.; Sun, M. Purification, identification, and sensory evaluation of kokumi peptides from agaricus bisporus mushroom. Foods 2019, 8(2), 1–12. [Google Scholar] [CrossRef]
- Deba-Rementeria, S.; Zugazua-Ganado, M.; Estrada, O.; Regefalk, J.; Vázquez-Araújo, L. Characterization of salt-preserved orange peel using physico-chemical, microbiological, and sensory analyses. Lwt 2021, 148(5), 1–11. [Google Scholar] [CrossRef]
- Lumivero. 2022. XLSTAT statistical and data analysis solution. Denver, CO. Retrieved from https://www.xlstat.com/en/.
- Team, Rs. 2022. RStudio: Integrated Development for R. RStudio. Boston, MA. Retrieved from http://www.rstudio.com/.
- Wang, H.; Suo, R.; Liu, X.; Wang, Y.; Sun, J.; Liu, Y.; Wang, W.; Wang, J. Kokumi γ-glutamyl peptides: Some insight into their evaluation and detection, biosynthetic pathways, contribution and changes in food processing. Food Chemistry Advances 2022, 1(7), 1–9. [Google Scholar] [CrossRef]
- Amano-enzyme. 2021. How the Amano-style Koji-Making Process Changed Microbial Enzymes for Food Industry. Retrieved October 23, 2023. from https://www.amano-enzyme.com/uk/introduction-to-the-amano-style-aerated-koji-making-process/.
- Chancharoonpong, C.; Hsieh, P.C.; Sheu, S.C. Effect of Different Combinations of Soybean and Wheat Bran on Enzyme Production from Aspergillus oryzae S. APCBEE Procedia 2012, 2(4), 68–72. [Google Scholar] [CrossRef]
- Kusumoto, K.I.; Yamagata, Y.; Tazawa, R.; Kitagawa, M.; Kato, T.; Isobe, K.; Kashiwagi, Y. Japanese traditional miso and Koji making. Journal of Fungi 2021, 7(7), 1–17. [Google Scholar] [CrossRef]
- Nakadai, T.; Nasuno, S. Enzyme preparation from extract of wheat bran koji by alcohol precipitation. Journal of Fermentation and Bioengineering 1989, 67(4), 253–257. [Google Scholar] [CrossRef]
- Sofyanovich, O.A.; Nishiuchi, H.; Yamagishi, K.; Matrosova, E.V.; Serebrianyi, V. Multiple pathways for the formation of the γ-glutamyl peptides γ-glutamyl-valine and γ-glutamyl-valyl-glycine in Saccharomyces cerevisiae. PLoS ONE 2019, 14(5), 1–18. [Google Scholar] [CrossRef]
- Guha, S.; Majumder, K. Comprehensive Review of γ-Glutamyl Peptides (γ-GPs) and Their Effect on Inflammation Concerning Cardiovascular Health. Journal of Agricultural and Food Chemistry 2022, 70(6), 7851–7870. [Google Scholar] [CrossRef]
- Egg, P.C. 2023; Safety of Fermented Foods.
- Saini, M.; Kashyap, A.; Bindal, S.; Saini, K.; Gupta, R. Bacterial Gamma-Glutamyl Transpeptidase, an Emerging Biocatalyst: Insights Into Structure–Function Relationship and Its Biotechnological Applications. Frontiers in Microbiology 2021, 12(4), 1–30. [Google Scholar] [CrossRef]
- Lin, H.; Zhou, B.; Zhao, J.; Liao, S.; Han, J.; Fang, J.; Liu, P.; Ding, W.; Che, Z.; Xu, M. Insight into the protein degradation during the broad bean fermentation process. Food Science and Nutrition 2022, 10(8), 2760–2772. [Google Scholar] [CrossRef]
- Xie, J.; Gänzle, M.G. Characterization of γ-glutamyl cysteine ligases from Limosilactobacillus reuteri producing kokumi-active γ-glutamyl dipeptides. Applied Microbiology and Biotechnology 2021, 105(13), 5503–5515. [Google Scholar] [CrossRef]
- Wang, S.; Tamura, T.; Kyouno, N.; Liu, X.; Zhang, H.; Akiyama, Y.; Yu Chen, J. Effect of the Chemical Composition of Miso (Japanese Fermented Soybean Paste) Upon the Sensory Evaluation. Analytical Letters 2021, 52(11), 1813–1827. [Google Scholar] [CrossRef]
- Astuti, R.D.; Fibri DL, N.; Handoko, D.D.; David, W.; Budijanto, S.; Shirakawa, H.; Ardiansyah. The Volatile Compounds and Aroma Description in Various Rhizopus oligosporus Solid-State Fermented and Nonfermented Rice Bran. Fermentation 2022, 8(120), 1–18. [Google Scholar] [CrossRef]
- Mao, Z.; Liu, L.; Zhang, Y.; Yuan, J. Efficient Synthesis of Phenylacetate and 2-Phenylethanol by Modular Cascade Biocatalysis. ChemBioChem 2020, 21(8), 2676–2679. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Imamura, M.; Katayama, H.; Obata, A.; Sugawara, E. Key compounds contributing to the fruity aroma characterization in Japanese raw soy sauce. Bioscience, Biotechnology and Biochemistry 2017, 81(10), 1984–1989. [Google Scholar] [CrossRef] [PubMed]
- Mortzfeld, F.B.; Hashem, C.; Vranková, K.; Winkler, M.; Rudroff, F. Pyrazines: Synthesis and Industrial Application of these Valuable Flavor and Fragrance Compounds. Biotechnology Journal 2020, 15(11). [Google Scholar] [CrossRef]
- Wang, P.; Ma, X.; Wang, W.; Xu, D.; Zhang, X.; Zhang, J.; Sun, Y. Characterization of flavor fingerprinting of red sufu during fermentation and the comparison of volatiles of typical products. Food Science and Human Wellness 2019, 8(4), 375–384. [Google Scholar] [CrossRef]
- Galata, M.; Mahmoud, S. Bioactive plant isoprenoids: Biosynthetic and biotechnological approaches. Studies in Natural Products Chemistry 2012. (Vol. 37, pp. 135–171). Elsevier. [Google Scholar] [CrossRef]
- Martorell, N.; Martí, M.P.; Mestres, M.; Busto, O.; Guasch, J. Determination of 4-ethylguaiacol and 4-ethylphenol in red wines using headspace-solid-phase microextraction-gas chromatography. Journal of Chromatography A 2002, 975(2), 349–354. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.N.; Tan, Z.W.; Shi, B.A. Influence of the pH on the formation of pyrazine compounds by the Maillard reaction of L-ascobic acid with acidic, basic and neutral amino acids. Asia-Pacific Journal of Chemical Engineering 2011, 7(7), 743–753. [Google Scholar] [CrossRef]
- Giri, A.; Ohshima, T. Dynamics of Aroma-Active Volatiles in Miso Prepared from Lizardfish Meat and Soy during Fermentation: A Comparative Analysis. International Journal of Nutrition and Food Sciences 2012, 1(1), 1–12. [Google Scholar] [CrossRef]
- Wooding, S.P.; Ramirez, V.A.; Behrens, M. Bitter taste receptors, Genes, evolution and health. Evolution, Medicine and Public Health 2021, 9(1), 431–447. [Google Scholar] [CrossRef]
- Moshtaghian, H.; Bolton, K.; Rousta, K. Challenges for upcycled foods: Definition, inclusion in the food waste management hierarchy and public acceptability. Foods 2021, 10(11), 1–17. [Google Scholar] [CrossRef]
- Ohsu, T.; Amino, Y.; Nagasaki, H.; Yamanaka, T.; Takeshita, S.; Hatanaka, T.; Maruyama, Y.; Miyamura, N.; Eto, Y. Involvement of the calcium-sensing receptor in human taste perception. Journal of Biological Chemistry 2010, 285(2), 1016–1022. [Google Scholar] [CrossRef] [PubMed]
- Higgins, M.J.; Hayes, J.E. Regional variation of bitter taste and aftertaste in humans. Chemical Senses 2019, 44(9), 721–732. [Google Scholar] [CrossRef]
Figure 1.
A) γ-Glutamyl peptides detected in barley, BSG and wheat bran miso after 12 weeks of fermentation, including cluster analysis for samples. The abundance of γ-glutamyl peptides is based on chromatography areas, which are unitless. Relative abundance can be compared for the same peptide but not across different peptides. B–E) Evolution of γ-glutamyl peptides over time: B) barley S1/BSG, C) wheat bran S2/ barley, D) wheat bran S1/ barley, E) wheat bran S1/BSG. Supplemental Materials Table S1 shows MS information.
Figure 1.
A) γ-Glutamyl peptides detected in barley, BSG and wheat bran miso after 12 weeks of fermentation, including cluster analysis for samples. The abundance of γ-glutamyl peptides is based on chromatography areas, which are unitless. Relative abundance can be compared for the same peptide but not across different peptides. B–E) Evolution of γ-glutamyl peptides over time: B) barley S1/BSG, C) wheat bran S2/ barley, D) wheat bran S1/ barley, E) wheat bran S1/BSG. Supplemental Materials Table S1 shows MS information.
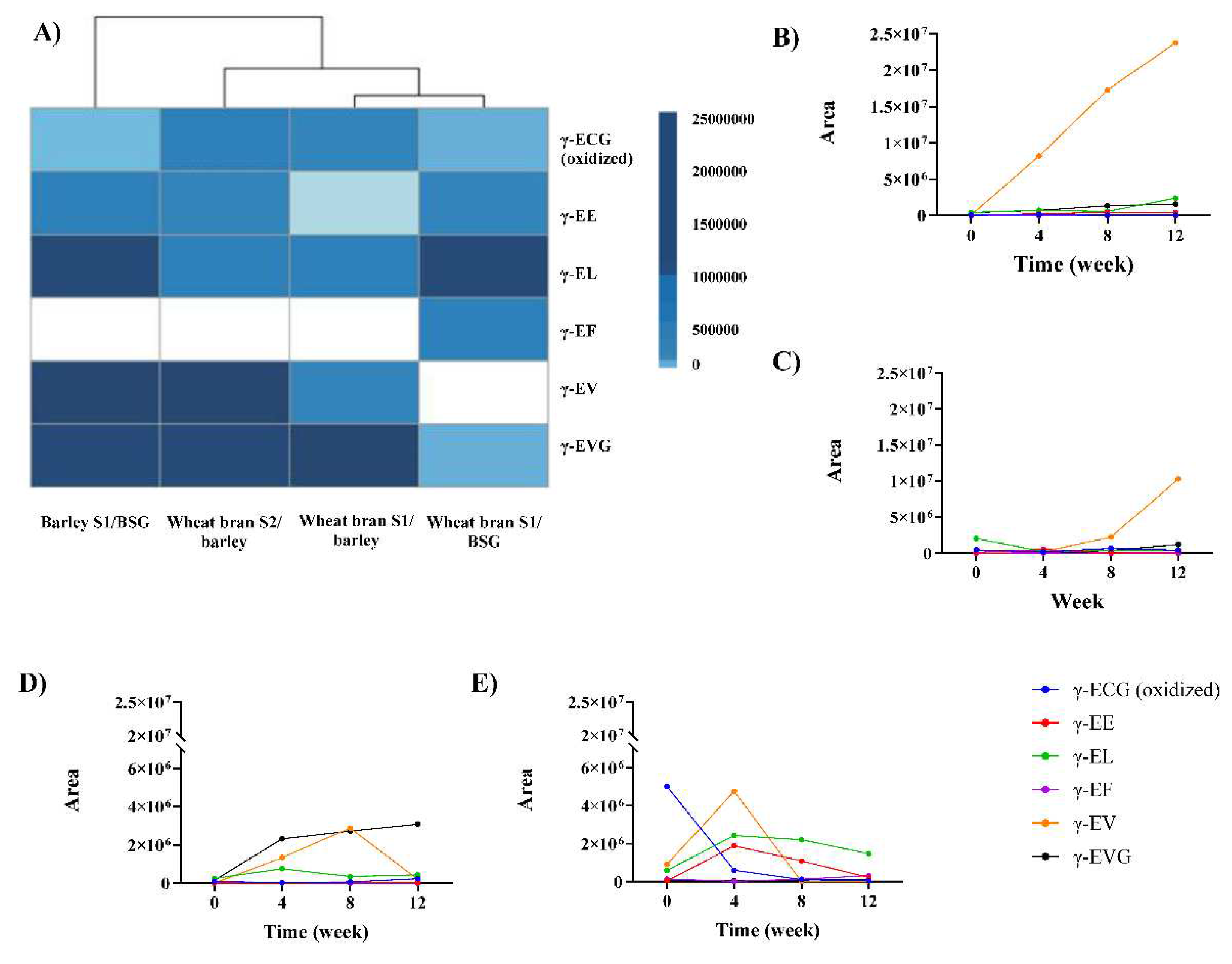
Figure 2.
Physicochemical characteristics during the miso fermentation. A) pH and γ-EV for the first cluster, B) pH and γ-EV for the second cluster, C) proteases activity (U/g) D) L* luminosity vs pH. One-way ANOVA and post-hoc analysis (Tukey test) with 95% confidence levels are shown in Supplemental Materials Table S1. All parameters were measured in triplicate, excluding colour where n=10. Legend: pH: dotted line, peptides: continuous line.
Figure 2.
Physicochemical characteristics during the miso fermentation. A) pH and γ-EV for the first cluster, B) pH and γ-EV for the second cluster, C) proteases activity (U/g) D) L* luminosity vs pH. One-way ANOVA and post-hoc analysis (Tukey test) with 95% confidence levels are shown in Supplemental Materials Table S1. All parameters were measured in triplicate, excluding colour where n=10. Legend: pH: dotted line, peptides: continuous line.
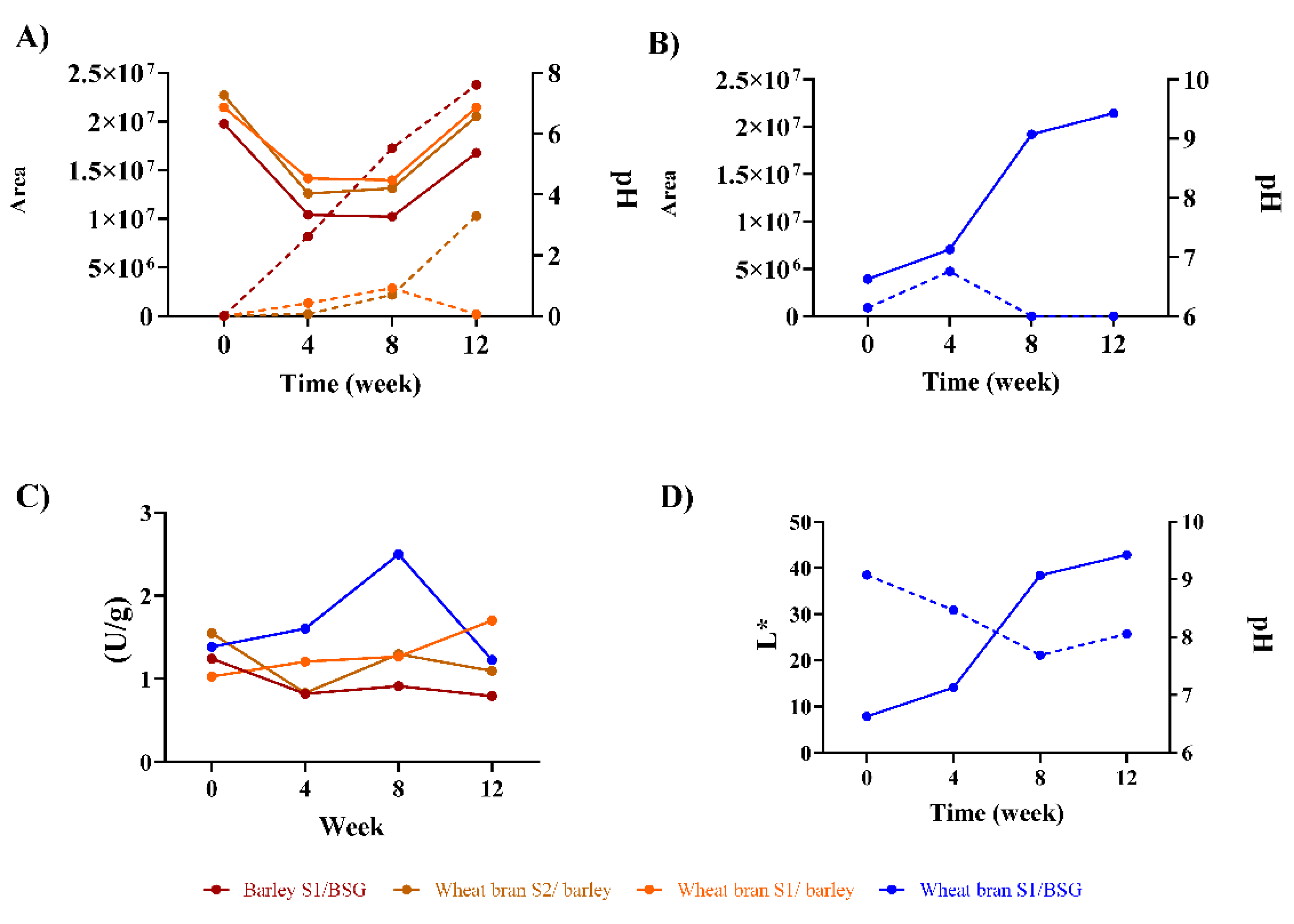
Figure 3.
Volatile aroma compounds were identified in the miso after 12 weeks of fermentation. All the volatile compounds shown have a significant correlation with the kokumi peptides (Pearson’s coefficient, 95 % confidence level). Kokumi correlation plot and statistical analysis are available in Supplemental Materials Figure S2 and Table S3, respectively. Compounds in bold are present in the highest concentrations and are mentioned in the manuscript.
Figure 3.
Volatile aroma compounds were identified in the miso after 12 weeks of fermentation. All the volatile compounds shown have a significant correlation with the kokumi peptides (Pearson’s coefficient, 95 % confidence level). Kokumi correlation plot and statistical analysis are available in Supplemental Materials Figure S2 and Table S3, respectively. Compounds in bold are present in the highest concentrations and are mentioned in the manuscript.
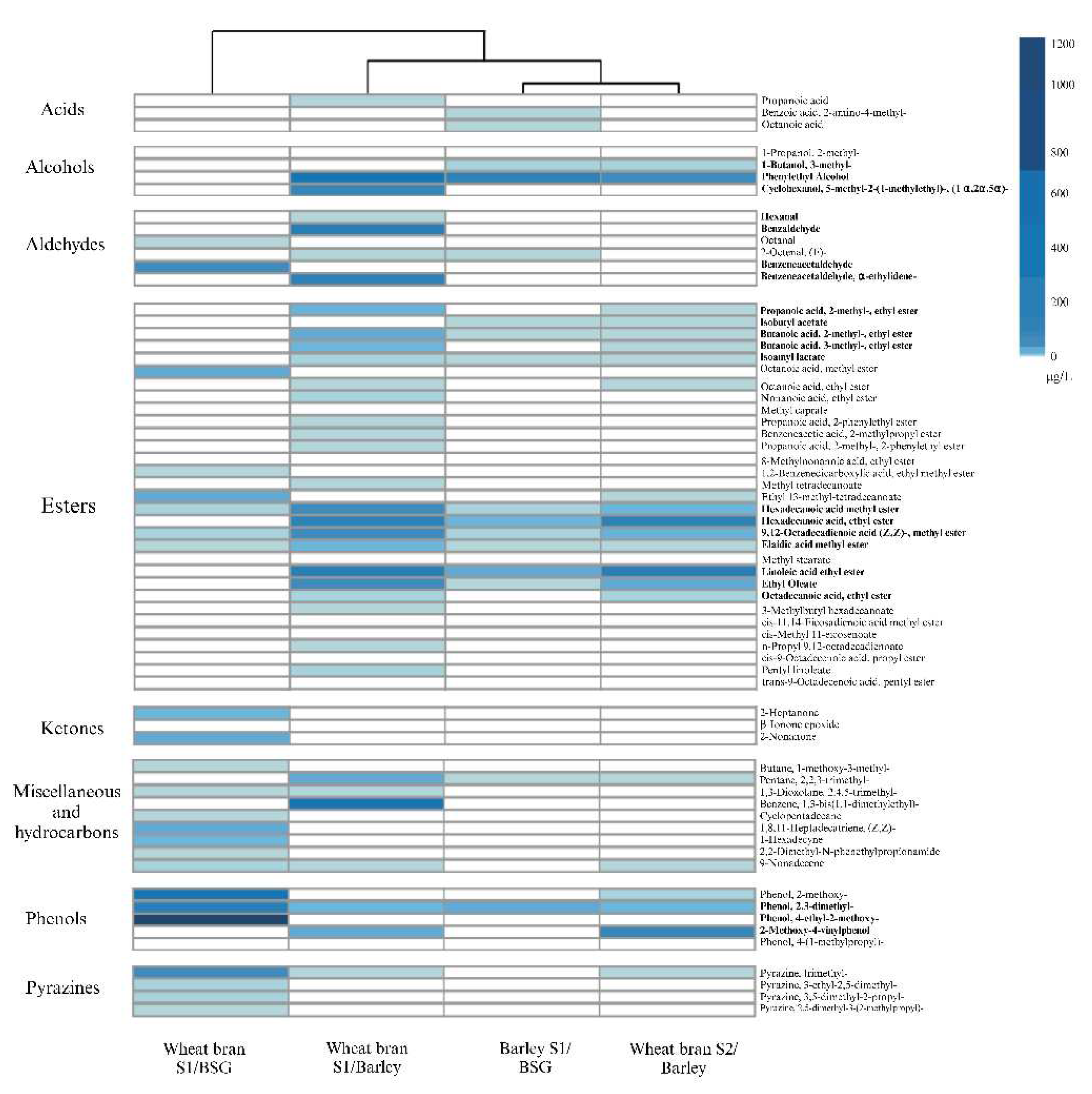
Figure 4.
A) Total abundance of γ-glutamyl peptides in each sample after 12 weeks of fermentation as based on chromatography areas. B) Total concentrations of volatile aroma compounds in each sample after 12 weeks of fermentation.
Figure 4.
A) Total abundance of γ-glutamyl peptides in each sample after 12 weeks of fermentation as based on chromatography areas. B) Total concentrations of volatile aroma compounds in each sample after 12 weeks of fermentation.
Figure 5.
A) Results fromthe consumer study: Intensity perception of aftertaste, coating, thickness, and bitterness when a kokumi rich extract from each of the misos were added to endive purée. Legend: Red: control; Orange: barley S1/BSG; Blue: wheat bran S2/ barley; Green: wheat bran S1/ barley. Attributes followed by * means significant differences (p<0.05). B) Sum of γ-glutamyl peptides at 12 weeks.
Figure 5.
A) Results fromthe consumer study: Intensity perception of aftertaste, coating, thickness, and bitterness when a kokumi rich extract from each of the misos were added to endive purée. Legend: Red: control; Orange: barley S1/BSG; Blue: wheat bran S2/ barley; Green: wheat bran S1/ barley. Attributes followed by * means significant differences (p<0.05). B) Sum of γ-glutamyl peptides at 12 weeks.
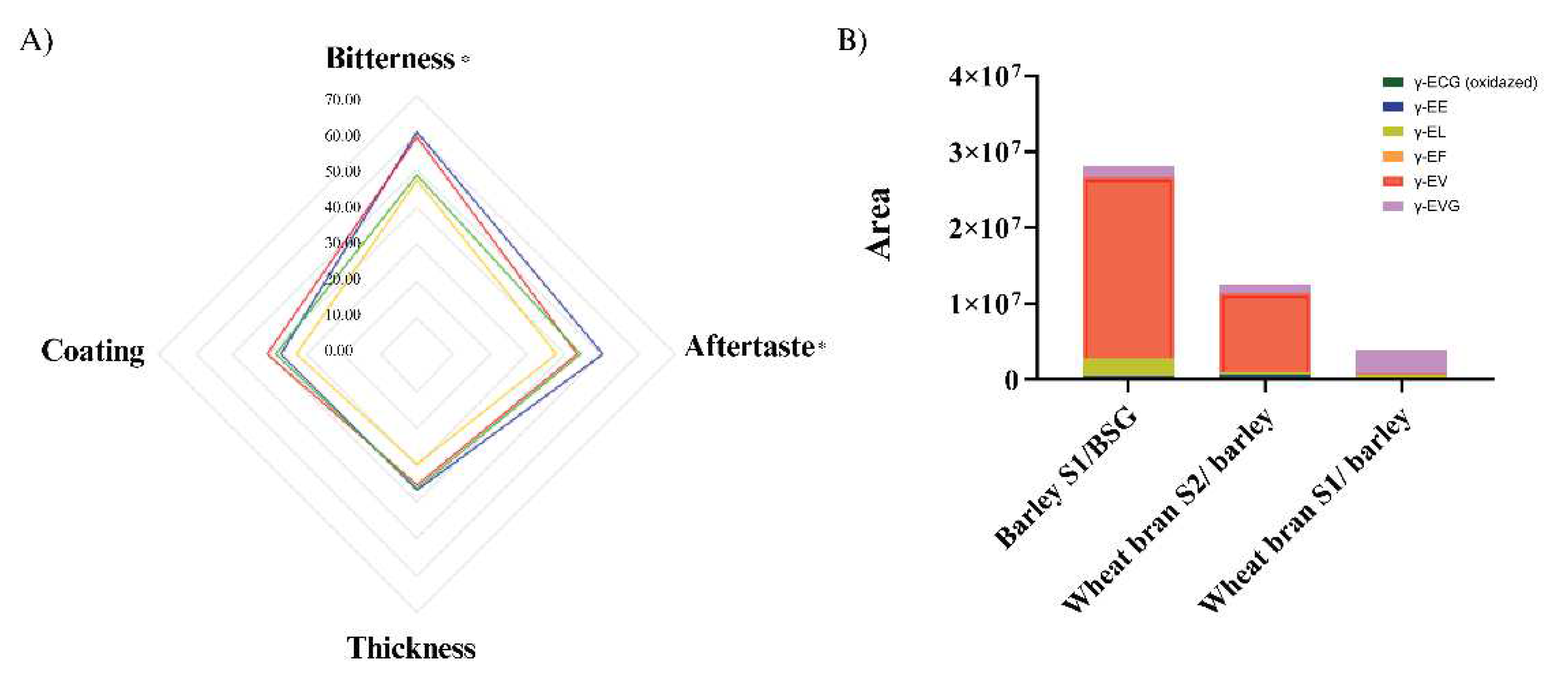

Table 1.
Ingredients for miso preparation in terms of koji substrate, miso substrate, the strain of A. oryzae, and sample code. The combinations of substrate and koji was based on a previous study (Rodríguez et al., 2023).
Table 1.
Ingredients for miso preparation in terms of koji substrate, miso substrate, the strain of A. oryzae, and sample code. The combinations of substrate and koji was based on a previous study (Rodríguez et al., 2023).
| Sample code | Koji substrate | Miso substrate | A. oryzae strain |
|---|---|---|---|
| Barley S1/BSG | Barley | BSG | S1 |
| Wheat bran S2/barley | Wheat bran | Barley | S2 |
| Wheat bran S1/barley | Wheat bran | Barley | S1 |
| Wheat bran S1/BSG | Wheat bran | BSG | S1 |
Table 2.
Results of one-way ANOVA and post-hoc analysis (Tukey HSD) of intensity perception in a gLMS for bitterness, aftertaste, thickness and coating. Samples within the same column followed by different letters were significantly different; bold font indicate significant p-values (p<0.05).
Table 2.
Results of one-way ANOVA and post-hoc analysis (Tukey HSD) of intensity perception in a gLMS for bitterness, aftertaste, thickness and coating. Samples within the same column followed by different letters were significantly different; bold font indicate significant p-values (p<0.05).
| Sample | Bitterness intensity |
Aftertaste intensity |
Thickness intensity |
Coating intensity |
| Control | 59.00 ab | 43.350ab | 35.52 a | 40.45 a |
| Miso Barley S1/BSG | 47.10 c | 37.87 b | 29.95 a | 32.72 a |
| Miso wheat bran S2/ barley | 60.45 a | 50.62 a | 36.53 a | 36.93 a |
| Miso wheat bran S1/ barley | 48.75 bc | 44.27 ab | 36.07 a | 38.22 a |
| p-value | 0.003 | 0.042 | 0.339 | 0.360 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



