Submitted:
20 October 2023
Posted:
21 October 2023
You are already at the latest version
Abstract
Inflammation is natural protective response of the body tissue caused by various stimuli. Macrophages start and control the inflammatory response. Microalgae have high value of ingredients including carotenoids which are well known for anti-inflammatory properties and used for various products in functional food industries. A green microalgal, Tetraselmis species contains a various type of carotenoids such as lutein, violaxanthin, astaxanthin, fucoxanthin, α-carotene and β-carotene. Therefore, we studied whether carotenoids isolated from Tetraselmis sp. with different culture conditions in Raw 264.7 cells and zebrafish (Danio rerio) have an anti -inflammatory effect. Carotenoid extracts of natural seawater (NS) and magma seawater (MS) showed inhibitory activity of nitric oxide (NO) overproduction without toxicity. In addition, carotenoid extracts of NS and MS inhibited pro-inflammatory cytokines including tumor necrosis factor (TNF)-α, interleukin (IL)- 1β and IL-6. Inducible nitric oxide synthase (iNOS) and cyclooxygenase (COX)-2, which are inflammatory mediators also decreased by pretreatment of NS and MS. Carotenoid extracts of NS and MS pretreatment also reduced phosphorylation of mitogen-activated protein kinases (MAPKs) and nuclear factor-kB (NF-kB). In conclusion, the carotenoid extract of Tetraselmis sp. has an anti -inflammatory effect and can be potential industrial ingredient.
Keywords:
Tetraselmis sp. Carotenoid
; anti-inflammatory
; zebrafish
; inflammation
1. Introduction
Inflammation response typically occur as a natural physiological response against injurious stimuli such as pathogens infection and toxics invasion, and it can be lead many disease such as diabetes arthritis, asthma, rheumatoid arthritis, atherosclerosis, asthma, chronic hepatitis, septic shock, and inflammatory neurodegenerative diseases and the other multiple disease [1,2,3,4,5,6,7,8], as well as it is a hallmarks of stress condition such as oxidative stress [9] and senescence [10].
When the inflammation occurred, macrophages initiate and regulate inflammatory responses, the activated macrophages secrete high chemokines, cytokines such as tumor necrosis factor (TNF) -α, interleukin (IL)-1α, IL-1β, IL-6, and pro-inflammatory mediators such as nitric oxide (NO), inducible nitric oxide synthase (iNOS), cyclooxygenase (COX)-2 [8].
Lipopolysaccharide (LPS) is a component of the outer membrane of Gram-negative bacteria and have been widely used to study inflammatory models of macrophages [11]. LPS regulates the expression of pro-inflammatory cytokines including TNF-α, IL-6 and IL-1β [12]. Moreover, it induces NO and prostaglandin E2 (PGE2) that are products of iNOS and COX-2. LPS lead to activate important pro-inflammatory factors like nuclear factor (NF)-kB and mitogen-activated protein kinases (MAPKs) including extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK) and p38 [13,14].
Carotenoids are well known tetraterpene pigments and are the most widely distributed pigments in nature and are present in photosynthetic bacteria, some species of archaea and fungi, plants, animals and algae. Macro- and microalgae are the potentially valuable source of bioactive compounds applied in the various industries and human usage in different fields of pharmaceutical, nutraceutical, and cosmetic disciplines. One of the interesting aspects is their application as the anti-inflammatory agents for prevention and/or treatment of inflammation.
Microalgae constitute rich source of high-value compounds like proteins, carbohydrates, minerals and diverse functional pigments including carotenoids [15]. Therefore, Microalgae has a variety of bioactive substances, and many studies have found that anti-tumors, anti-inflammation and immune regulatory properties [16,17,18]. Therefore, microalgae compounds are of great importance in the treatment of inflammations to reduce the reaction of immune system against pathogens, toxic compounds and damaged cells.
Tetraselmis species, a member of the Chlorodendrophyceae, are green algae and are commonly found in estuaries, tide pools, and brackish ponds where the environment frequently changes [19,20,21] and studies have shown that they can survive long-term outdoor culture conditions [22,23], is industrially important because it is microalgae that have been successfully mass-cultured. Moreover, they have functional compound include pigment such as carotenoid that are a diverse and widespread class of bioactive compounds. Carotenoids are separated into carotenes including lutein, violaxanthin, astaxanthin, fucoxanthin, α-carotene and β-carotene [24]. Carotenoids have characteristic structure, and they exert various bioactive properties including antioxidant and anti-inflammation [25].
In the study, therefore, natural seawater (NS) and magma seawater (MS) were utilized instead of culture medium vitamins to increase the carotenoid content and confirmed whether the culture environment of Tetraselmis sp. affects carotenoid content. In addition, it was determined that the carotenoids derived from Tetraselmis sp., have an anti-inflammatory effect against the LPS-stimulating inflammatory response in RAW 264.7 cells and zebrafish embryos.
2. Results
2.1. Carotenoid Extract from Natural Seawater (NS) and Magma Seawater (MS) Inhibits Lipopolysaccharide (LPS)-Induced Toxicity and Nitric Oxide Production
First, to check whether the carotenoid extract of NS and MS have toxicity, it was tested with RAW 264.7 cells and zebrafish embryos. In the cells, the carotenoid derived from NS and MS were pretreated at the various concentrations in the range of 50 to 200 µg/mL and then treated with 1 µg/mL LPS. Neither NS nor MS showed cytotoxicity at the tested concentrations compared to LPS only treated cells (Figure 1A). Heartbeats was measured to test toxicity of the carotenoid derived from NS and MS in vivo, 1 μg/mL of the extracts pretreated to embryos for 30 min and then treated with 20 μg/mL LPS. Treatment of LPS significantly increased heartbeats, indicating toxicity, and the elevated heartbeat was decreased by pretreatment of the extracts (Figure 1C). These results suggesting that the extracts (NS and MS) attenuate LPS-induced toxicity. LPS stimulates immune responses by inducing cytokine production [26], and immune myeloid cells such as macrophages induced nitric oxide (NO) overproduction in response to inflammatory signals [27]. LPS treatment on the cells occurred NO overproduction compared to that of the control, and the extracts (NS and MS) pretreatment significantly inhibited NO production caused by LPS in dose-dependent manners (Figure 1B). To evaluate the effects of the extracts (NS and MS) against LPS in vivo, DAF-DA prove used to quantify NO concentration in embryos. NO overproduction was occurred caused by LPS and this reduced pretreatment of the extracts (NS and MS) treatments significantly (Figure 1D, E). These results suggest that pretreatment of the extracts (NS or MS) inhibits LPS-induced NO overproduction without toxicity in both RAW 264.7 cells and zebrafish embryos.
2.2. Carotenoid Extract from Natural (NS) and Magma Seawater (MS) Inhibits Lipopolysaccharide (LPS)-Induced Pro-Inflammatory Cytokines Secretion
Lipopolysaccharide (LPS) is a potent proinflammatory activator of macrophages in vivo and in vitro can directly elicit cytokine responses [26,28]. The extracts (NS and MS) pretreatment inhibited LPS-induced NO overproduction in both RAW 264.7 cells and embryos. So, it was confirmed whether the extracts (NS and MS) pretreatment could reduce the increase in cytokines caused by LPS. The secretion of cytokines including tumor necrosis factor (TNF)-α, interleukin (IL)-1β, and IL-6 was induced, and the induction was significantly decreased by pretreatment of the extracts (NS and MS) dose-dependently in the cells (Figure 2A-C), and although not significant in the embryos, the extracts (NS and MS) pretreatment prevented the increase in IL-1β (Figure 2D). These results suggest that the extracts (NS and MS) expected to contribute to reducing secretion of pro-inflammatory cytokines.
2.3. Carotenoid Extract from Natural Seawater (NS) and Magma Seawater (MS) Inhibits Lipopolysaccharide (LPS)-Induced Inflammatory Mediators
Nitric oxide (NO) is produced by immunocompetent cells such as macrophages that serves as a messenger molecule for various physiological and pathological processes and it is synthesized from L-arginine by nitric oxide synthase (NOS) present in tissues, and of the three genes found mainly on different chromosomes, inducible NOS (iNOS) is triggered by various inflammatory stimuli such as LPS and cytokines [29]. In addition, LPS activates the production of pro-inflammatory cytokines such as IL-1β, IL-6, and TNF-α in macrophages to release cyclooxygenase (COX)-2 and iNOS, and COX-2 can be overexpressed after LPS stimulation. Prostaglandin E2 (PGE2) is mainly synthesized by COX-2, which also leads to inflammatory response [30]. In the present study, LPS treatment induced pro-inflammatory cytokines and the induction was decreased by pretreatment of the extracts (NS and MS). So, the protein expression level of iNOS and COX-2 were determined in the cells. As the result of Western blotting in Figure 3 (A, B), both the protein expression level was increased caused by LPS treatment and significantly decreased by pretreatment of the extracts (NS and MS). LPS treatment significantly increased the protein expression levels of both iNOS and COX-2, and compared to LPS treated only, NS pretreatment did not reduce iNOS expression at a concentration of 100 µg/mL, but significantly decreased at 200 µg/mL concentration, and COX-2 expression significantly decreased at both 100 and 200 µg/mL concentrations (Figure 3A). On the other hand, MS pretreatment significantly reduced the protein expression levels of iNOS and COX-2 compared to LPS treated only in both 100 and 200 µg/mL (Figure 3B). However, neither NS and MS inhibit the production of LPS-induced PGE2, the principal mediator of inflammation (Figure 3C) [31]. Also, there was no change in cox-2a mRNA expression level in the extracts (NS and MS) treatment compared to LPS in zebrafish embryo (Figure 3D). These results indicate that NS and MS contributed to inflammatory effects by inhibiting the production of LPS-induced iNOS and COX-2 in in vitro but did not suppress the production of PGE2.
2.4. Carotenoid Extract from Natural Seawater (NS) and Magma Seawater (MS) Suppress Lipopolysaccharide (LPS)-Stimulated Mitogen Activated Protein Kinases (MAPKs) and Nuclear Factor (NF)-kB Signaling Pathway
Lipopolysaccharide (LPS) has been presented to initiate multiple intracellular signaling events, including the stimulation of pathways that lead to the activation of nuclear factor (NF)-kB and three distinct mitogen-activated protein kinases (MAPKs), the ERK (extracellular signal-regulated kinase), p38, and JNK (c-Jun N-terminal kinase). Next, so, the MAPKs and NF-kB signaling was determined in the cells. LPS treatment induced the phosphorylation of ERK, p38, and JNK, and this highly elevated phosphorylation was reduced by treatment of PD98059 (ERK1/2 signaling inhibitor), SP600125 (JNK inhibitor), and SB203580 (p38 MAPK inhibitor), respectively. The extracts (NS and MS) treatment also decreased LPS-stimulated phosphorylation of ERK, p38, and JNK (Figure 4). Interestingly, the phosphorylation of ERK and JNK was reduced a similar level as the inhibitor in the 200 µg/mL concentration NS pretreated cells (Figure 4A, C), and phosphorylation of p38 also slightly reduced by NS pretreatment (Figure 4E). As well, the phosphorylation of p38 protein expression was decreased a similar level as the inhibitor in MS pretreated cells, in both 100 and 200 µg/mL concentrations (Figure 4F), and phosphorylation of ERK and JNK protein expression level was decreased as a similar to that of the inhibitor at the 200 µg/mL concentrations MS (Figure 4B, D). On the other hands, LPS treatment downregulated IkB protein expression level, whereas upregulated p-65 phosphorylation in the cytoplasm, and induced p65 protein expression levels in the nucleus. Treatment with NF-kB inhibitor, PDTC which ammonium pyrrolidinedithiocarbamate reversed the LPS-induced protein expression levels of IkB, p-p65, and p65, as well as the protein expression levels was reduced by pretreatment of the both extracts mL (Figure 4G-L). These results indicating that NS and MS improve inflammatory responses by inhibiting MAPKs and NF-kB signaling pathways in RAW 264.7 cells stimulated by LPS.
3. Discussion
Carotenoids are well-known plant chemicals since the early 19th century. In particular, β-carotene is well known as a precursor for producing vitamin A. And also, Carotenoids are well known as fundamental elements of the photosynthetic process in photosynthetic bacteria, some fungi, plants and algae. In particular, in the photosynthetic organ, there is a removing process system of enzyme that produces xanthophyll and lutein, which are popular as functional materials. Carotenoid is one of most rich ingredients in microalgae, and has excellent biological activity, and research on its derived carotenoids in various industries [32,33]. Microalgae origin carotenoids prevent diabetes by suppressing blood sugar growth [34], and is reported to have beneficial effects through anti-inflammatory and anti-oxidant activity in various diseases such as obesity [35,36], brain disease [37,38], and cardiovascular disease [39,40]. In addition, microalgae are known to synthesize the new molecular structure according to the seawater composition [41]. Although the culture medium containing vitamins is generally used for mass production of microalgae, it is necessary to reduce cost production and to study the culture medium for the content of growth and active ingredients [42,43]. Therefore, we confirmed whether Tetraselmis sp. has a beneficial effect and increased carotenoid content when cultured using natural sea water (NS) and magma sea water (MS) instead of vitamin-containing media. Our analysis of the carotenoid content of Tetraselmis sp. extracts cultured in NS and MS showed no significant difference, with MS yielding about 8% higher than NS.
One of the main physiological activity of carotenoids is their anti-inflammatory effects while eliminating reactive nitrogen species (RNSs) [41]. A significant overproduction of RNS in cells/tissues can occur as a result of excessive exposure to harmful external chemical or physical factors. An important group of compounds with unpaired electrons characterized by high chemical activity are the reactive forms of nitrogen such as nitric oxide (NO) can result of metabolic changes to nitrosonium cation (NO+), niitroxylanion (NO−) and peroxynitrite (ONOO−) are play a protective effect for the body against microbes in general, however, excessive increase these levels, leading to the occurrence of nitrosation stress, finally causing inflammation [42]. Recent research has shown that if inflammation is not properly controlled, it is a major factor in the development of various chronic diseases/disorders, including diabetes [43,44], cancer [45], cardiovascular disease [46], eye disease [47,48], arthritis [49], obesity [50], autoimmune disease [51] and inflammatory bowel disease [52]. In this study confirmed that the carotenoids isolated from Tetraselmis sp. showed significant NO scavenging ability in both RAW 264.7 cells and zebrafish embryos.
Macrophages such as RAW 264.7 cells are the initial response cells in the inflammatory response, are immune cells in most body tissues, which are directly effective from external substances, responds, and macrophages participate in various stages of inflammation [53,54], as well as the cells are activated by LPS to increase inflammatory cytokines such as TNF-α and IL-6 and NO production to initiate an inflammatory response [55,56]. And, lipopolysaccharide (LPS) is one of the effective macrophage activators that activate inflammatory mediators such as NO and PGE2 and induce overproduction [57]. In addition, iNOS is involved in NO production, and inhibition of iNOS is the target of anti-inflammatory, and COX-2 produces PGE2 and is associated with the progression of chronic inflammatory diseases [55,56]. Therefore, the inhibitory of iNOS and COX-2 expression can be involved in the decrease in NO and PGE2 production, which can contribute to the treatment of inflammatory diseases. Our results confirmed that NS and MS carotenoids inhibited NO production in a dose-dependent manner in LPS-stimulated RAW 264.7 cells, but the protein expression levels of iNOS and COX-2 are reduced along the concentration of NS and MS, although levels of PGE2 remained constant. This suggests that the production level of PGE2 remains the same, but the NS and MS carotenoids reduce iNOS and COX-2 to contribute to inflammation attenuating.
Activation of MAPK and NF-kB pathways in LPS-stimulated macrophages can increase the secretion of pro-inflammatory cytokines such as TNF-α, IL-6, and IL-1β, resulting in increased expression of iNOS and COX-2 [30,58]. Our result, LPS increased the phosphorylation of ERK, JNK, p38, and p65, and NS and MS carotenoids inhibited the increase in this phosphorylation. In addition, NS and MS carotenoids inhibited the production of pro-inflammatory cytokines TNF-α, IL-6, and IL-1β increased by LPS in a concentration-dependent manner. This proves that in LPS-stimulated RAW 264.7 cells, NS and MS carotenoids inhibit the MAPKs and NF-kB pathways, thereby inhibiting the production of pro-inflammatory cytokines, resulting in anti-inflammatory action.
Zebrafish (Danio rerio) is one of the in vivo models used in various biomedical research fields, including immunology, toxicology, cancer, and behavioral biology, with advantages such as high fertility, various transgenic, and human tissue and genomic similarity [59,60]. The LPS-stimulated zebrafish model has been used to evaluate anti-inflammatory materials because a response similar to that observed in mammals has been observed [60,61]. In this study, the zebrafish embryo showed an increase in heartbeats, is an indicator of toxicity, by LPS treatment, whereas this increase was reduced by NS and MS carotenoids pretreatment. In addition, the induction il-1β and cox-2a mRNA expression level caused by LPS was reduced by NS and MS carotenoids pretreatment. Overall, the current results, NS and MS carotenoids are found to have anti -inflammatory activity by effectively reducing NO production in both in vitro and in vivo. However, NS and MS carotenoids in zebrafish embryos do not significantly reduce the expression level of pro-inflammatory cytokine genes caused by LPS, so further research is needed to identify mechanisms related to anti-inflammatory efficacy in vivo.
4. Materials and Methods
4.1. Microalgal Culture
The marine microalga, Tetraselmis sp. LIM-PS-1293, was obtained from the Library of Marine Samples (LIMS) of the Korea Institute of Ocean Science and Technology (KIOST, Geoje, Korea). Tetraselmis biomass was produced in Jeju Bio Research Center of KIOST and cultivated in a vertical rounded photobioreactor (named ROSEMAX) containing 200 L of culture medium. Two types of seawater, natural seawater (NS) collected at the surface of the Jeju coast and magma seawater (MS) from a depth of 140 m underground of Jeju, were used instead of vitamins to modify Guillard’s f/2 medium as the culture media. For experiment, each culture medium was sterilized under dark conditions (at night) by adding 13% sodium hypochlorite (NaOCl)to 1% of culture volume, and then the NaOCl was neutralized with sodium thiosulfate (1 mol/L) after 24 h. Culture conditions were maintained at 26.1 ± 1.9℃ of water temperature and 33.0 ± 0.5 psu of salinity under a natural light/dark cycle, the pH value increased from 7.6 to 8.1 as the cells grew. During the culture periods, the air was continuously imposed at 0.4 vvm (air volume per culture volume per minute) to prevent cell sedimentation. Microalga cultured for 9 days was harvested using a tubular separator (GQLY series, Hanil S.M.E, Anyang, Korea) at 8,000 rpm for 30 min. Harvested wet biomass was stored at −50℃ in a deep freezer and then lyophilized for 2 days in a freeze-dryer (FDTA-45, Operon, Gimpo, Korea).
4.2. Carotenoid Extract And Analysis the Content
Carotenoid of Tetraselmis sp. was extracts with acetone and the carotenoid including the astaxanthin, fucoxanthin, lutein, capsanthin, zeaxanthin and canthaxanthin contents were analyzed [62].
4.3. Measurement of Cytotoxicity
First, we have test cytotoxicity of the extracts, to do this, the murine macrophage cell line RAW264.7 (KCLB NO 40071; Seoul, Republic of Korea) was cultured in DMEM (11995-065, Gibco, Waltham, MA, USA) containing 10% FBS (Gibco, Waltham, MA, USA), 1% of mixture of streptomycin (100 µg/mL)/penicillin (100 unit/mL) (Gibco). The cells were maintained at 37℃ in an incubator with a humidified atmosphere of 5% CO2. The cells were seeded on 24-well plates at a concentration of 1.5×105 cells/mL. After 16 h, cells were treated with 100, 200 µg/mL concentrations of the extracts, then incubated for 1 h, followed by incubation with 1 µg/mL lipopolysaccharide (LPS, Sigma-Aldrish) for 24 h. The cytotoxicity of the extracts was assessed using the 3-(4,5-Dimethyl-2-thiazolyl)-2,5-diphenyltetrazolium Bromide (MTT, Sigma Aldrich) assay, MTT solution (10 µg/mL) was added at 25 µL per well, after 3 h formazan crystals in each well were dissolved in dimethyl sulfoxide (DMSO, Sigma Aldrich). The intensity of purple formazan was determined by measuring absorbance at 570 nm using a microplate reader (Bio Tek Instruments, lnc, Winooski, VT, USA).
4.4. Measurement of and Nitric Oxide (NO), Prostaglandin E2 (PGE2) and Pro-Inflammatory Cytokines of RAW 264.7 Cells
In order to check whether the extracts have anti-inflammatory effect, first we have determined the production of NO which is a basic indicator of inflammation occurrence. The cells were seeded on 24-well plates at a concentration of 1.5×105 cells/mL. After 16 h, cells were treated with 100, 200 µg/mL concentrations of the extracts, then incubated for 1 h, followed by incubation with 1 µg/mL LPS (Sigma Aldrich) for 24 h. NO production was evaluated using the Griess assay. A cell culture medium volume of 100 µL was mixed with 100 µL Griess reagent (1% sulfanilamide and 0.1% naphthylethylenediamide dihydrochloride in 2.5% phosphoric acid; Sigma Aldrich, St. Louis, MO, USA) and the mixture was incubated for 10 min at room temperature in the dark. Reactants were measured by absorbance at 540 nm using a microplate reader. The PGE2 (Minneapolis, MN, USA) and pro-inflammatory cytokines (TNF-α, IL-6, IL-1β; BD Biosciences, Franklin Lakes, NJ, USA) of the culture supernatant were measured using a mouse ELISA kit (R&D Systems, Minneapolis, MN, USA) following the manufacturer’s instructions.
4.5. Western Blots
The cells were seeded on 6-well plates at a concentration of 1.5×105 cells/mL. After 16 h, the cells were treated with 100, and 200 µg/mL concentrations of the extracts then incubated for 1 h, and then LPS (1 µg/mL) was added for 24 h or for 15 min at 37℃ in an incubator with a humidified atmosphere of 5% CO2. After incubation, the cells were harvested and washed twice with cold-PBS (Welgene, Gyeongsan, Korea). Protein concentrations of the cell lysates were measured using a BCATM protein assay kit (Thermo Scientific, Waltham, MA, USA) according to the manufacturer’s instructions. Blocked membranes with 3% bovine serum albumin and 2% skim milk were incubated with primary (1:1000 dilution, Cell Signaling Technology, Danvers, MA, USA) and goat anti-mouse or anti-rabbit secondary (1:3000 dilution, Santa Cruz Biotechnology, Dallas, TX, USA) antibodies. The protein bands were detected using FUSION SOLO (Vilber Lourmat, Marne La Vallée, France), and the intensity quantification of the Western blot results was measured using ImageJ software (version 1.46r, www.nih.gov).
4.6. Zebrafish Maintenance and Embryo Harvesting
Adult zebrafish were maintained in the zebrafish housing system (Aqua Blue, CHUNG FU TECHNICAL DEVELOPMENT CO., Taipei, Taiwan). The zebrafish maintained optimum growth conditions under a 14/10 light cycle, and the housing temperature was maintained at 28.5℃. The zebrafish were fed with Color Charasin PREMIUM (JAQNO®, Gwangmyeong, Korea). For collecting embryos, zebrafish were set up in breeding tanks with male to female ratio of 2:1. Next morning, collect the embryos using a stain sieve and move to a petri dish. Collected embryos were treated with methylene blue (0.5 ppm) containing Egg H2O (0.003% sea salt, 0.0075% calcium sulfate) to bleaching, and incubated in the incubator at 28℃. Two hours after incubation, the media was changed with fresh Egg H2O and the embryos were maintained at 28℃ for the subsequent experiment.
4.7. Extract Treatment on Zebrafish Embryos
The extract was dissolved in DMSO to make 50 mg/mL stock concentration and 20 μg/mL LPS (Sigma-Aldrich) were dissolved in Egg H2O and stored at -20℃ for further experiments. A 3 dpf (day post fertilization) stage 15 embryos were transferred of 1 mL Egg H2O in a 24-well plate. The extracts (1 µg/mL, NS and MS) were treated to the embryos for 30 min, and added 20 µg/mL LPS (Sigma-Aldrich) for 15-30 min. And then, the media were changed with fresh Egg H2O for further experiments.
4.8. Heartbeat Measurement
The heartbeat was measured to check the toxicity of the extracts. Heartbeat was measured under the standard microscope for 3 min and the results present mean values were calculated for 1 min on the result.
4.9. NO Measurement of the Embryos
To determine the NO production of zebrafish embryos, fluorescent probe dye of 10 µg/mL DAF-FMDA (Sigma Aldrich) was added for 1 h. After the prior experiment, the experiment embryos were washed with fresh Egg H2O and anesthetized with 0.003% Tricaine (Sigma-Aldrich), and observed under a fluorescence microscope (SZX16, Olympus Corporation, Tokyo, Japan). The fluorescence intensity of the image was quantified by ImageJ software (https://imagej.nih.gov/ij). The triplicated mean value was represented by a bar graph.
4.10. RT-qPCR
To determine proinflammatory cytokines of the embryos, total mRNA was extracted from zebrafish embryos using RNAiso plus (Takara Bio Inc., Kusatsu, Japan), and cDNA was synthesized with the PrimeScriptTM cDNA synthesis kit (Takara Bio lnc., Kusatsu, Japan) following the manufacturer’s protocol. cDNA was analyzed using SYBR® Premix TaqTM, ROX plus (Takara Bio Inc., Kusatsu, Japan) on Bio-Rad cycler (Hercules, CA, USA). The β-actin gene was used as an internal reference gene and was not affected by LPS. Relative expression of fold difference was calculated with the ΔΔCT method, in triplicate for each group. The sequences of RT-qPCR primer pairs were presented in Table 1.
il-1β: Interleukin (il)-1β; cox-2a: cyclooxygenase-2a.
4.11. Statistics
All experiments were performed in triplicate and data were statistically analyzed using GraphPad Prism 7.0. Tukey test was used to determine the p value in GraphPad Prism 7.0. The result was considered statistically at p<0.05. All values are expressed as mean ± SE.
5. Conclusions
In summary, in this study, pretreatment of carotenoids extracted from Tetraselmis sp. cultured with natural sea water (NS) and magma sea water (MS) inhibited NO production in LPS-treated RAW 264.7 cells and zebrafish embryos. In addition, protein expression of iNOS, COX-2 in LPS-treated RAW 264.7 cells were down-regulated by NS and MS carotenoid pretreatment, and the production of pro-inflammatory cytokines TNF-α, IL-6, and IL-1β increased by LPS was also inhibited. This is the result of NS and MS carotenoid pretreatment inhibiting the MAPK and NF-kB pathways in LPS-treated RAW264.7 cells. The use of magma sea water (MS) has a higher overall yield of carotenoids than natural sea water (NS). However, in our study, both NS and MS use confirmed the effective anti-inflammatory effects of carotenoids isolated from Tetraselmis sp. Therefore, Tetraselmis SP. can be a potential treatment for inflammatory diseases in the pharmaceutical and food industries.
Acknowledgments
This work was supported by the KOREA INNOVATION CLUSTER grant funded by the Ministry of Science and ICT (No.2023-DD-UP-0007-01-203) and the Korea Institute of Ocean Science and Technology (grant. No. PEA0125 and PEA0126).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jo, W. S.; Choi, Y. J.; Kim, H. J.; Nam, B. H.; Hong, S. H.; Lee, G. A.; Lee, S. W.; Seo, S. Y.; Jeong, M. H. Anti-inflammatory effect of microalgal extracts from Tetraselmis suecica. Food Science and Biotechnology 2010, 19, 1519–1528. [Google Scholar] [CrossRef]
- Moriya, J. Critical roles of inflammation in atherosclerosis. Journal of cardiology 2019, 73, 22–27. [Google Scholar] [CrossRef]
- Hansson, G. K. Inflammation, atherosclerosis, and coronary artery disease. The New England journal of medicine 2005, 352, 1685–1695. [Google Scholar] [CrossRef]
- KleinJan, A. Airway inflammation in asthma: Key players beyond the Th2 pathway. Current opinion in pulmonary medicine 2016, 22, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Zeremski, M.; Petrovic, L. M.; Talal, A. H. The role of chemokines as inflammatory mediators in chronic hepatitis C virus infection. Journal of viral hepatitis 2007, 14, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Liu, S. F.; Malik, A. B. NF-kappa B activation as a pathological mechanism of septic shock and inflammation. American journal of physiology. Lung cellular and molecular physiology 2006, 290, L622–l645. [Google Scholar] [CrossRef] [PubMed]
- Zipp, F.; Aktas, O. The brain as a target of inflammation: Common pathways link inflammatory and neurodegenerative diseases. Trends in neurosciences 2006, 29, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Abdulkhaleq, L. A.; Assi, M. A.; Abdullah, R.; Zamri-Saad, M.; Taufiq-Yap, Y. H.; Hezmee, M. N. M. The crucial roles of inflammatory mediators in inflammation: A review. Veterinary world 2018, 11, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M. C.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxidative medicine and cellular longevity 2016, 2016, 7432797. [Google Scholar] [CrossRef] [PubMed]
- Lasry, A.; Ben-Neriah, Y. Senescence-associated inflammatory responses: Aging and cancer perspectives. Trends in immunology 2015, 36, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, R. F.; Sá-Correia, I.; Valvano, M. A. Lipopolysaccharide modification in Gram-negative bacteria during chronic infection. FEMS microbiology reviews 2016, 40, 480–493. [Google Scholar] [CrossRef]
- Han, Y.; Jung, H. W.; Lee, D. H.; Kwon, S. Y.; Son, K. H.; Park, Y. K. Anti-inflammatory effects of prosapogenin III from the dried roots of Liriope platyphylla in LPS-stimulated RAW264.7 cells. Journal of Asian natural products research 2013, 15, 1038–1049. [Google Scholar] [CrossRef]
- Xu, R.; Ma, L.; Chen, T.; Wang, J. Sophorolipid Suppresses LPS-Induced Inflammation in RAW264.7 Cells through the NF-κB Signaling Pathway. Molecules (Basel, Switzerland) 2022, 27. [Google Scholar] [CrossRef]
- Park, M. Y.; Ha, S. E.; Kim, H. H.; Bhosale, P. B.; Abusaliya, A.; Jeong, S. H.; Park, J. S.; Heo, J. D.; Kim, G. S. Scutellarein Inhibits LPS-Induced Inflammation through NF-κB/MAPKs Signaling Pathway in RAW264.7 Cells. Molecules (Basel, Switzerland) 2022, 27. [Google Scholar] [CrossRef]
- Patel, A. K.; Singhania, R. R.; Awasthi, M. K.; Varjani, S.; Bhatia, S. K.; Tsai, M. L.; Hsieh, S. L.; Chen, C. W.; Dong, C. D. Emerging prospects of macro- and microalgae as prebiotic. Microb Cell Fact 2021, 20, 112. [Google Scholar] [CrossRef] [PubMed]
- Martínez Andrade, K. A.; Lauritano, C.; Romano, G.; Ianora, A. Marine Microalgae with Anti-Cancer Properties. Mar Drugs 2018, 16. [Google Scholar] [CrossRef] [PubMed]
- Ávila-Román, J.; García-Gil, S.; Rodríguez-Luna, A.; Motilva, V.; Talero, E. Anti-Inflammatory and Anticancer Effects of Microalgal Carotenoids. Mar Drugs 2021, 19. [Google Scholar] [CrossRef] [PubMed]
- Riccio, G.; Lauritano, C. Microalgae with Immunomodulatory Activities. Mar Drugs 2019, 18. [Google Scholar] [CrossRef] [PubMed]
- Hyung, J.-H.; Kim, E.-J.; Moon, S.-J.; Kang, N. S.; Park, J. Tetraselmis jejuensis sp. nov. (Chlorodendrophyceae), a Euryhaline Microalga Found in Supralittoral Tide Pools at Jeju Island, Korea. 2021, 10, 1289. [Google Scholar]
- Isdepsky, A.; Borowitzka, M. A. In-pond strain selection of euryhaline Tetraselmis sp. strains for reliable long-term outdoor culture as potential sources of biofuel and other products. Journal of Applied Phycology 2019, 31, 3359–3370. [Google Scholar] [CrossRef]
- Fon-Sing, S.; Borowitzka, M. A. Isolation and screening of euryhaline Tetraselmis spp. suitable for large-scale outdoor culture in hypersaline media for biofuels. Journal of Applied Phycology 2016, 28, 1–14. [Google Scholar] [CrossRef]
- Ahmad, I.; Hellebust, J. A. Osmoregulation in the Extremely Euryhaline Marine Micro-Alga Chlorella autotrophica. Plant Physiol 1984, 74, 1010–1015. [Google Scholar] [CrossRef] [PubMed]
- Garrity, S. D. Some Adaptations of Gastropods to Physical Stress on a Tropical Rocky Shore. Ecology 1984, 65, 559–574. [Google Scholar] [CrossRef]
- Cardoso, C.; Pereira, H.; Franca, J.; Matos, J.; Monteiro, I.; Pousão-Ferreira, P.; Gomes, A.; Barreira, L.; Varela, J.; Neng, N.; Nogueira, J. M.; Afonso, C.; Bandarra, N. M. Lipid composition and some bioactivities of 3 newly isolated microalgae (Tetraselmis sp. IMP3, Tetraselmis sp. CTP4, and Skeletonema sp.). Aquaculture International 2020, 28, 711–727. [Google Scholar] [CrossRef]
- Kabir, M. T.; Rahman, M. H.; Shah, M.; Jamiruddin, M. R.; Basak, D.; Al-Harrasi, A.; Bhatia, S.; Ashraf, G. M.; Najda, A.; El-kott, A. F.; Mohamed, H. R. H.; Al-malky, H. S.; Germoush, M. O.; Altyar, A. E.; Alwafai, E. B.; Ghaboura, N.; Abdel-Daim, M. M. Therapeutic promise of carotenoids as antioxidants and anti-inflammatory agents in neurodegenerative disorders. Biomedicine & Pharmacotherapy 2022, 146, 112610. [Google Scholar]
- Meng, F.; Lowell, C. A. Lipopolysaccharide (LPS)-induced macrophage activation and signal transduction in the absence of Src-family kinases Hck, Fgr, and Lyn. J Exp Med 1997, 185, 1661–1670. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, E. M.; McGinity, C.; Wink, D. A.; McVicar, D. W. Nitric Oxide in Macrophage Immunometabolism: Hiding in Plain Sight. Metabolites 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Rostkowska-Nadolska, B.; Pośpiech, L.; Fortuna, W.; Szymaniec, S.; Miedzybrodzki, R.; Gawron, W.; Latocha, M. [RANTES expression in nasal polyps fibroblasts; spontaneously and after stimulation with lipopolisaccharides (LPS) and phytohemagglutinin (PHA)]. Otolaryngologia polska = The Polish otolaryngology 2006, 60, 143–147. [Google Scholar]
- Ozleyen, A.; Yilmaz, Y. B.; Tumer, T. B. Dataset on the differentiation of THP-1 monocytes to LPS inducible adherent macrophages and their capacity for NO/iNOS signaling. Data Brief 2021, 35, 106786. [Google Scholar] [CrossRef]
- Lee, Y. M.; Kim, M.; Yuk, H. J.; Kim, S. H.; Kim, D. S. Siraitia grosvenorii Residual Extract Inhibits Inflammation in RAW264.7 Macrophages and Attenuates Osteoarthritis Progression in a Rat Model. Nutrients 2023, 15. [Google Scholar] [CrossRef]
- Park, J. Y.; Pillinger, M. H.; Abramson, S. B. Prostaglandin E2 synthesis and secretion: The role of PGE2 synthases. Clin Immunol 2006, 119, 229–240. [Google Scholar] [CrossRef]
- Gómez-Zorita, S.; Trepiana, J.; González-Arceo, M.; Aguirre, L.; Milton-Laskibar, I.; González, M.; Eseberri, I.; Fernández-Quintela, A.; Portillo, M. P. Anti-Obesity Effects of Microalgae. Int J Mol Sci 2019, 21. [Google Scholar] [CrossRef]
- Khavari, F.; Saidijam, M.; Taheri, M.; Nouri, F. Microalgae: Therapeutic potentials and applications. Mol Biol Rep 2021, 48, 4757–4765. [Google Scholar] [CrossRef] [PubMed]
- Zhuge, F.; Ni, Y.; Wan, C.; Liu, F.; Fu, Z. Anti-diabetic effects of astaxanthin on an STZ-induced diabetic model in rats. Endocr J 2021, 68, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Gammone, M. A.; D'Orazio, N. Anti-obesity activity of the marine carotenoid fucoxanthin. Mar Drugs 2015, 13, 2196–2214. [Google Scholar] [CrossRef] [PubMed]
- Pappalardo, I.; Santarsiero, A.; Radice, R. P.; Martelli, G.; Grassi, G.; de Oliveira, M. R.; Infantino, V.; Todisco, S. Effects of Extracts of Two Selected Strains of Haematococcus pluvialis on Adipocyte Function. 2023, 23, 1737. [Google Scholar] [CrossRef] [PubMed]
- El-Baz, F. K.; Salama, A.; Ali, S. I.; Elgohary, R. Lutein isolated from Scenedesmus obliquus microalga boosts immunity against cyclophosphamide-induced brain injury in rats. Scientific Reports 2022, 12, 22601. [Google Scholar] [CrossRef] [PubMed]
- Gallego, R.; Valdés, A.; Sánchez-Martínez, J. D.; Suárez-Montenegro, Z. J.; Ibáñez, E.; Cifuentes, A.; Herrero, M. Study of the potential neuroprotective effect of Dunaliella salina extract in SH-SY5Y cell model. Anal Bioanal Chem 2022, 414, 5357–5371. [Google Scholar] [CrossRef] [PubMed]
- Cui, S.; Wu, H.; He, Q.; Wang, L.; Yi, X.; Feng, G.; Wu, Q.; Tao, B.; Han, D.; Hu, Q.; Xia, H.; Xu, L. Fucoxanthin alleviated atherosclerosis by regulating PI3K/AKT and TLR4/NFκB mediated pyroptosis in endothelial cells. Int Immunopharmacol 2023, 120, 110370. [Google Scholar] [CrossRef]
- El-Baz, F. K.; Hussein, R. A.; Saleh, D. O.; Abdel Jaleel, G. A. R. Zeaxanthin Isolated from Dunaliella salina Microalgae Ameliorates Age Associated Cardiac Dysfunction in Rats through Stimulation of Retinoid Receptors. Mar Drugs 2019, 17. [Google Scholar] [CrossRef]
- Mucha, P.; Skoczyńska, A.; Małecka, M.; Hikisz, P.; Budzisz, E. Overview of the Antioxidant and Anti-Inflammatory Activities of Selected Plant Compounds and Their Metal Ions Complexes. 2021, 26, 4886. [Google Scholar] [CrossRef]
- Medzhitov, R. Inflammation 2010: New adventures of an old flame. Cell 2010, 140, 771–776. [Google Scholar] [CrossRef]
- Böni-Schnetzler, M.; Meier, D. T. Islet inflammation in type 2 diabetes. Semin Immunopathol 2019, 41, 501–513. [Google Scholar] [CrossRef]
- Diedisheim, M.; Carcarino, E.; Vandiedonck, C.; Roussel, R.; Gautier, J. F.; Venteclef, N. Regulation of inflammation in diabetes: From genetics to epigenomics evidence. Mol Metab 2020, 41, 101041. [Google Scholar] [CrossRef]
- Singh, N.; Baby, D.; Rajguru, J. P.; Patil, P. B.; Thakkannavar, S. S.; Pujari, V. B. Inflammation and cancer. Ann Afr Med 2019, 18, 121–126. [Google Scholar] [CrossRef]
- Henein, M. Y.; Vancheri, S.; Longo, G.; Vancheri, F. The Role of Inflammation in Cardiovascular Disease. 2022, 23, 12906. [Google Scholar] [CrossRef] [PubMed]
- Perez, V. L.; Stern, M. E.; Pflugfelder, S. C. Inflammatory basis for dry eye disease flares. Exp Eye Res 2020, 201, 108294. [Google Scholar] [CrossRef] [PubMed]
- Baudouin, C.; Kolko, M.; Melik-Parsadaniantz, S.; Messmer, E. M. Inflammation in Glaucoma: From the back to the front of the eye, and beyond. Prog Retin Eye Res 2021, 83, 100916. [Google Scholar] [CrossRef]
- Jutley, G. S.; Sahota, K.; Sahbudin, I.; Filer, A.; Arayssi, T.; Young, S. P.; Raza, K. Relationship Between Inflammation and Metabolism in Patients With Newly Presenting Rheumatoid Arthritis. Front Immunol 2021, 12, 676105. [Google Scholar] [CrossRef]
- Kawai, T.; Autieri, M. V.; Scalia, R. Adipose tissue inflammation and metabolic dysfunction in obesity. Am J Physiol Cell Physiol 2021, 320, C375–c391. [Google Scholar] [CrossRef] [PubMed]
- Duan, L.; Rao, X.; Sigdel, K. R. Regulation of Inflammation in Autoimmune Disease. J Immunol Res 2019, 2019, 7403796. [Google Scholar] [CrossRef]
- Saez, A.; Herrero-Fernandez, B.; Gomez-Bris, R.; Sánchez-Martinez, H.; Gonzalez-Granado, J. M. Pathophysiology of Inflammatory Bowel Disease: Innate Immune System. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Rodriguez, P.; Chen, H.; Erndt-Marino, J. D.; Liu, F.; Totsingan, F.; Gross, R. A.; Hahn, M. S. Impact of Select Sophorolipid Derivatives on Macrophage Polarization and Viability. ACS applied bio materials 2019, 2, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Yeom, M.; Park, J.; Lim, C.; Sur, B.; Lee, B.; Han, J. J.; Choi, H. D.; Lee, H.; Hahm, D. H. Glucosylceramide attenuates the inflammatory mediator expression in lipopolysaccharide-stimulated RAW264.7 cells. Nutrition research (New York, N.Y.) 2015, 35, 241–250. [Google Scholar] [CrossRef]
- Alam, M. B.; Chowdhury, N. S.; Sohrab, M. H.; Rana, M. S.; Hasan, C. M.; Lee, S. H. Cerevisterol Alleviates Inflammation via Suppression of MAPK/NF-κB/AP-1 and Activation of the Nrf2/HO-1 Signaling Cascade. Biomolecules 2020, 10. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, Y.; Chen, S.; Hu, Y.; Zhu, Z.; Wang, Y.; Du, N.; Song, T.; Yang, Y.; Guo, A.; Wang, Y. Macrophage migration inhibitory factor facilitates prostaglandin E(2) production of astrocytes to tune inflammatory milieu following spinal cord injury. Journal of neuroinflammation 2019, 16, 85. [Google Scholar] [CrossRef]
- Park, C. M.; Song, Y. S. Luteolin and luteolin-7-O-glucoside inhibit lipopolysaccharide-induced inflammatory responses through modulation of NF-κB/AP-1/PI3K-Akt signaling cascades in RAW 264.7 cells. Nutrition research and practice 2013, 7, 423–429. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, W.; Zheng, F.; Yu, H.; Wei, K. Xanthatin Alleviates LPS-Induced Inflammatory Response in RAW264.7 Macrophages by Inhibiting NF-κB, MAPK and STATs Activation. Molecules (Basel, Switzerland) 2022, 27. [Google Scholar] [CrossRef]
- Patton, E. E.; Tobin, D. M. Spotlight on zebrafish: The next wave of translational research. Disease models & mechanisms 2019, 12. [Google Scholar]
- Kim, E. A.; Kang, N.; Kim, J.; Yang, H. W.; Ahn, G.; Heo, S. J. Anti-Inflammatory Effect of Turbo cornutus Viscera Ethanolic Extract against Lipopolysaccharide-Stimulated Inflammatory Response via the Regulation of the JNK/NF-kB Signaling Pathway in Murine Macrophage RAW 264.7 Cells and a Zebrafish Model: A Preliminary Study. Foods (Basel, Switzerland) 2022, 11. [Google Scholar]
- Forn-Cuní, G.; Varela, M.; Pereiro, P.; Novoa, B.; Figueras, A. Conserved gene regulation during acute inflammation between zebrafish and mammals. Sci Rep 2017, 7, 41905. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.-A.; Kang, N.; Heo, S.-Y.; Oh, J.-Y.; Lee, S.-H.; Cha, S.-H.; Kim, W.-K.; Heo, S.-J. Antioxidant, Antiviral, and Anti-Inflammatory Activities of Lutein-Enriched Extract of Tetraselmis Species. 2023, 21, 369. 2023, 21, 369. [Google Scholar] [PubMed]
Figure 1.
Carotenoid extract from natural seawater (NS) and magma seawater (MS) inhibits LPS-induced toxicity and nitric oxide (NO) production in both RAW 264.7 cells and zebrafish embryos. The cells were treated with 25, 50, 100 and 200 μg/mL concentration of NS and MS extract for 1 h, and then treated with or without LPS (1 μg/mL) for 24 h. (A) The cells viability was measured by MTT assay. (B) The cells culture medium was evaluated NO production using Griess assay. NO production was read by microplate reader at absorbance at 540 nm. Control, vehicle only. Data represent the means ± SD. SD: standard deviation, ns: not significant, *p<0.05, ***p<0.001 vs. the control; ###p<0.001 vs. LPS. LPS: lipopolysaccharide.
Figure 1.
Carotenoid extract from natural seawater (NS) and magma seawater (MS) inhibits LPS-induced toxicity and nitric oxide (NO) production in both RAW 264.7 cells and zebrafish embryos. The cells were treated with 25, 50, 100 and 200 μg/mL concentration of NS and MS extract for 1 h, and then treated with or without LPS (1 μg/mL) for 24 h. (A) The cells viability was measured by MTT assay. (B) The cells culture medium was evaluated NO production using Griess assay. NO production was read by microplate reader at absorbance at 540 nm. Control, vehicle only. Data represent the means ± SD. SD: standard deviation, ns: not significant, *p<0.05, ***p<0.001 vs. the control; ###p<0.001 vs. LPS. LPS: lipopolysaccharide.
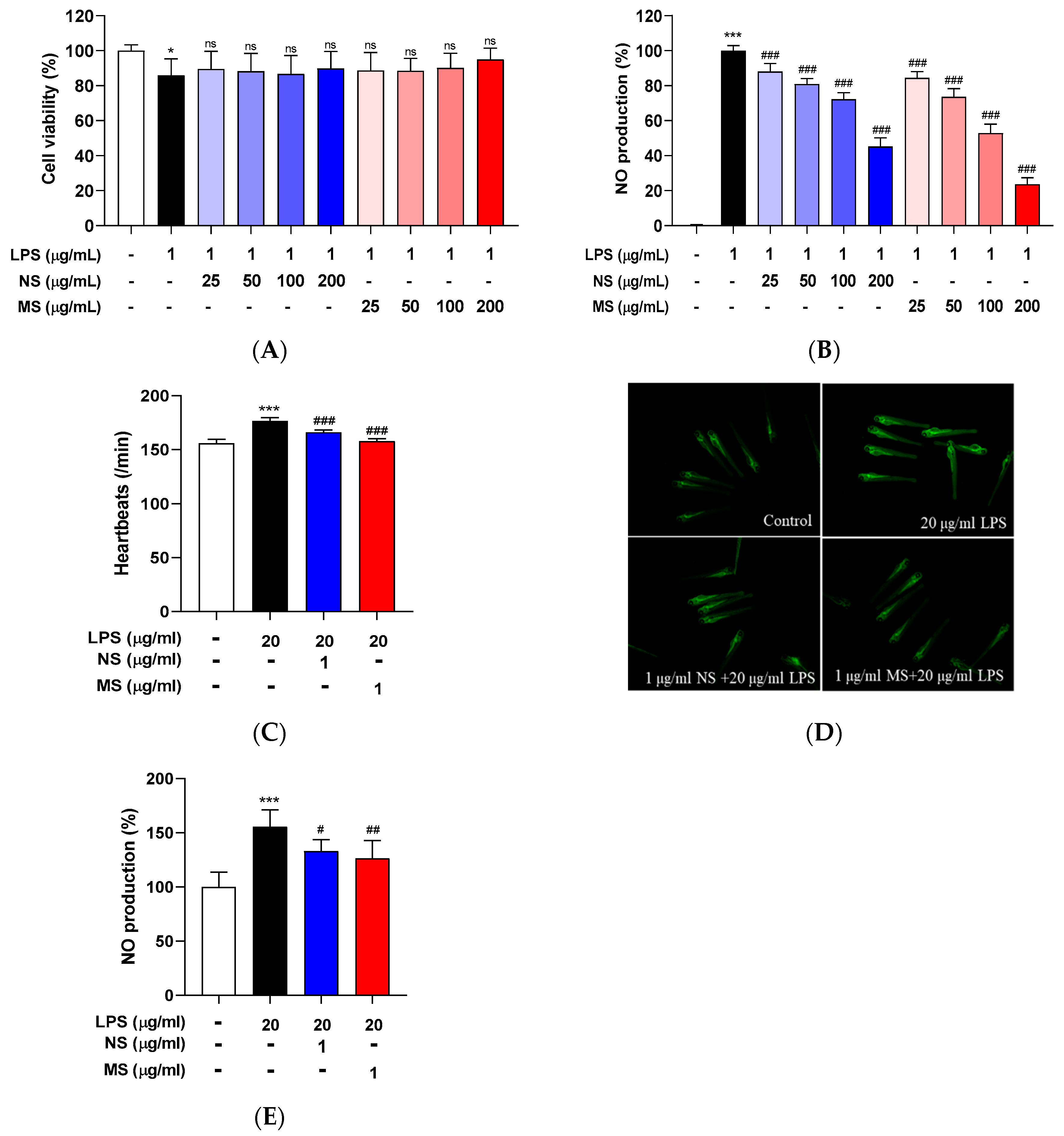
Figure 2.
Carotenoid extract from natural seawater (NS) and magma seawater (MS) inhibits LPS-induced mRNA expression of pro-inflammatory cytokines in both RAW 264.7 cells and zebrafish embryos. The cells were treated with 100, 200 μg/mL concentrations of NS and MS extract for 1 h and then treated with or without LPS (1 μg/mL) for 24 h. (A) tumor necrosis factor (TNF)-α, (B) interleukin (IL)- 1β, and (C) IL-6 were measured using a mouse ELISA kit. Wild type zebrafish embryos (at 72 hpf, hour post fertilization) were pretreated with 1 µg/ml concentration of NS and MS extract for 30 min, and then treated with 20 µg/mL LPS for 15-30 min. The mRNA level of pro-inflammatory cytokine (D) interleukin (il)-1β was measured by RT-qPCR. Control, vehicle only. Data represent the means ± SD. SD: standard deviation, ns: not significant, ***p<0.001 vs. the control; ##p<0.01, ###p<0.001 vs. LPS. LPS: lipopolysaccharide.
Figure 2.
Carotenoid extract from natural seawater (NS) and magma seawater (MS) inhibits LPS-induced mRNA expression of pro-inflammatory cytokines in both RAW 264.7 cells and zebrafish embryos. The cells were treated with 100, 200 μg/mL concentrations of NS and MS extract for 1 h and then treated with or without LPS (1 μg/mL) for 24 h. (A) tumor necrosis factor (TNF)-α, (B) interleukin (IL)- 1β, and (C) IL-6 were measured using a mouse ELISA kit. Wild type zebrafish embryos (at 72 hpf, hour post fertilization) were pretreated with 1 µg/ml concentration of NS and MS extract for 30 min, and then treated with 20 µg/mL LPS for 15-30 min. The mRNA level of pro-inflammatory cytokine (D) interleukin (il)-1β was measured by RT-qPCR. Control, vehicle only. Data represent the means ± SD. SD: standard deviation, ns: not significant, ***p<0.001 vs. the control; ##p<0.01, ###p<0.001 vs. LPS. LPS: lipopolysaccharide.
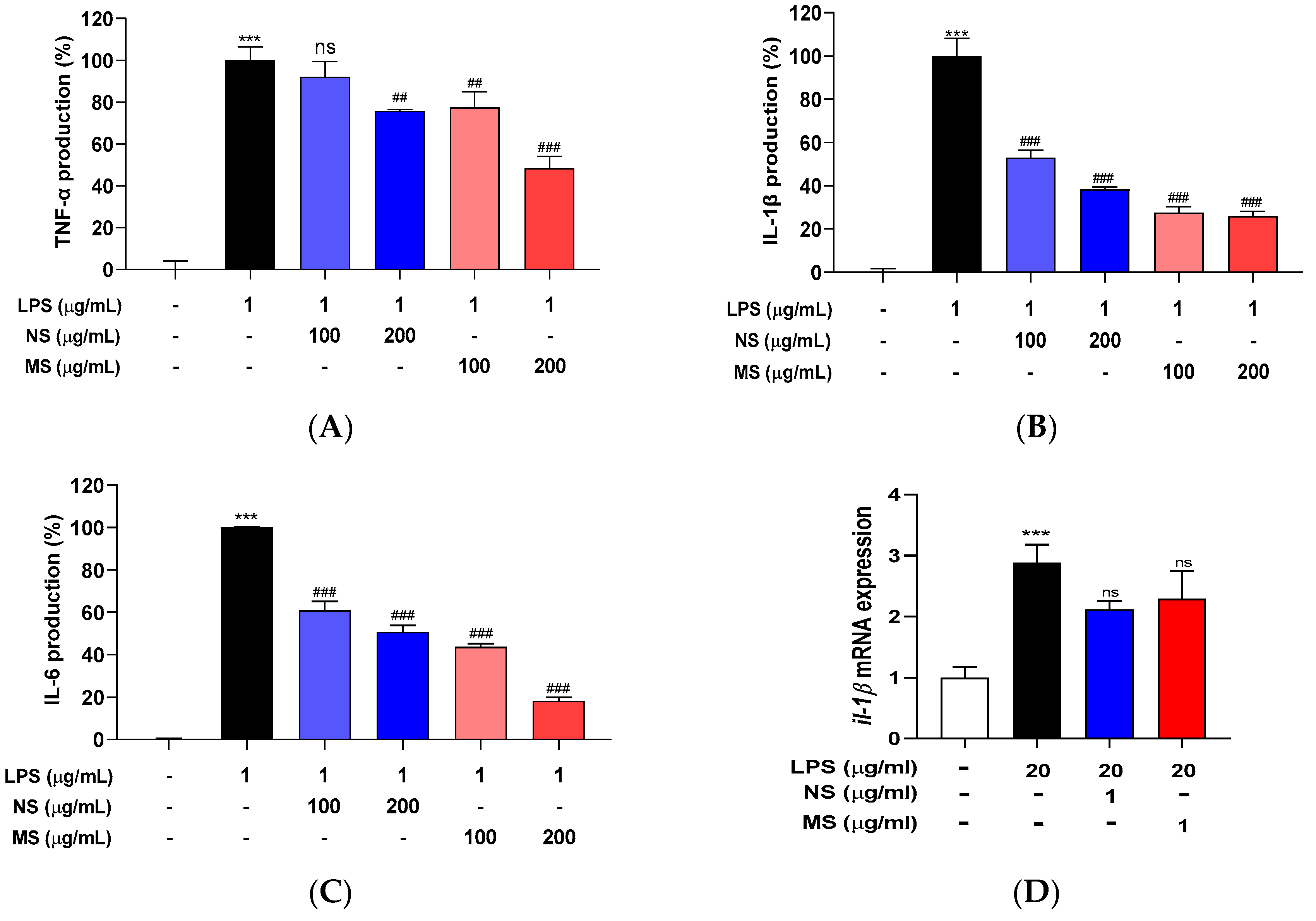
Figure 3.
Carotenoid extract from natural seawater (NS) and magma seawater (MS) inhibits LPS-induced protein expression of inflammatory mediators in both RAW 264.7 cells and zebrafish embryos. The cells were treated with 100, 200 μg/mL concentrations of NS and MS extract, respectively for 1 h and then treated with or without LPS (1 μg/mL) for 24 h. Inducible nitric oxide synthase (iNOS) and cyclooxygenase (COX)-2 protein expression levels of (A) NS and (B) MS were measured by Western blotting. The expression was normalized to that of internal control, β-actin, respectively, and quantified using ImageJ software. (C) Prostaglandin E2 (PGE2) was measured using a mouse ELISA kit. Wild type zebrafish embryos (at 72 hpf, hour post fertilization) were pretreated with 1 µg/ml concentration of NS and MS extract for 30 min, and then treated with 20 µg/mL LPS for 15-30 min. (D) The mRNA expression level of cyclooxygenase (cox)-2a was measured by RT-qPCR. Control, vehicle only. Data represent the means ± SD. SD: standard deviation, ns: not significant, ***p<0.001 vs. the control; ## p<0.01, ###p<0.001 vs. LPS. LPS: lipopolysaccharide.
Figure 3.
Carotenoid extract from natural seawater (NS) and magma seawater (MS) inhibits LPS-induced protein expression of inflammatory mediators in both RAW 264.7 cells and zebrafish embryos. The cells were treated with 100, 200 μg/mL concentrations of NS and MS extract, respectively for 1 h and then treated with or without LPS (1 μg/mL) for 24 h. Inducible nitric oxide synthase (iNOS) and cyclooxygenase (COX)-2 protein expression levels of (A) NS and (B) MS were measured by Western blotting. The expression was normalized to that of internal control, β-actin, respectively, and quantified using ImageJ software. (C) Prostaglandin E2 (PGE2) was measured using a mouse ELISA kit. Wild type zebrafish embryos (at 72 hpf, hour post fertilization) were pretreated with 1 µg/ml concentration of NS and MS extract for 30 min, and then treated with 20 µg/mL LPS for 15-30 min. (D) The mRNA expression level of cyclooxygenase (cox)-2a was measured by RT-qPCR. Control, vehicle only. Data represent the means ± SD. SD: standard deviation, ns: not significant, ***p<0.001 vs. the control; ## p<0.01, ###p<0.001 vs. LPS. LPS: lipopolysaccharide.
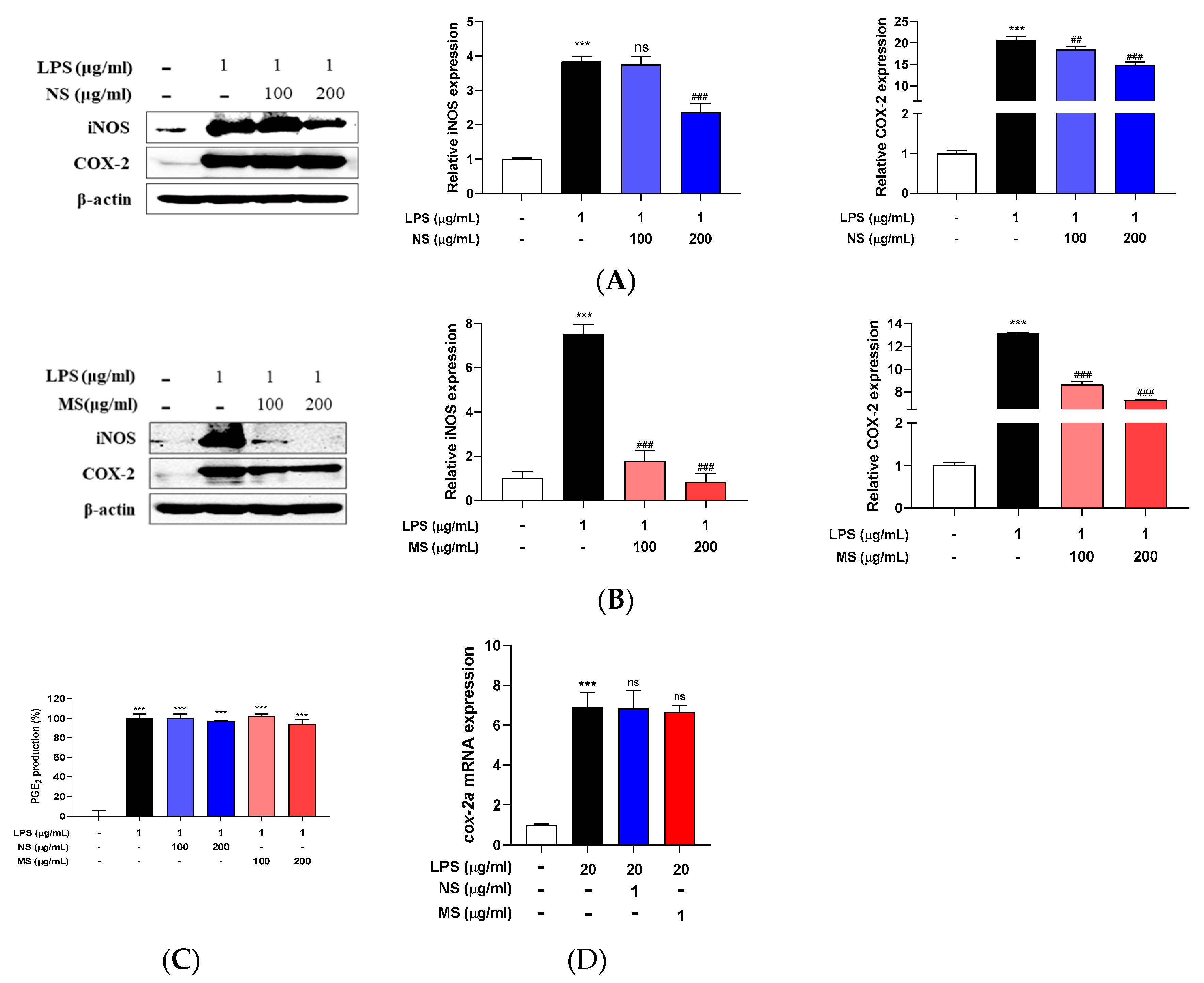
Figure 4.
Carotenoid extract from natural seawater (NS) and magma seawater (MS) inhibits LPS-induced mitogen activated protein kinase and nuclear factor-kB signaling pathways. The cells were treated with 100, 200 μg/mL concentrations of NS and MS extract, respectively for 1 h and then treated with or without LPS (1 μg/mL) for 24 h. The protein expression levels of (A, B) phosphorylation (p)-extracellular signal-regulated kinase (ERK), (C, D) p-c-JUN N-terminal kinase (JNK), (E, F) p-p38 and (G-N) IkB, p-p65 and p65 were measured by Western blotting. The expression was normalized to that of internal control, actin, Lamin B, respectively, and quantified using ImageJ Software. Control, vehicle only. Data represent the means ± SD. SD: standard deviation, ns: not significant, ***p<0.01, ***p<0.001 vs. the control; #p<0.05, ##p<0.01, ###p<0.001 vs. LPS. LPS: lipopolysaccharide, PD: PD98059, SP: SP600125, SB: SB203580.
Figure 4.
Carotenoid extract from natural seawater (NS) and magma seawater (MS) inhibits LPS-induced mitogen activated protein kinase and nuclear factor-kB signaling pathways. The cells were treated with 100, 200 μg/mL concentrations of NS and MS extract, respectively for 1 h and then treated with or without LPS (1 μg/mL) for 24 h. The protein expression levels of (A, B) phosphorylation (p)-extracellular signal-regulated kinase (ERK), (C, D) p-c-JUN N-terminal kinase (JNK), (E, F) p-p38 and (G-N) IkB, p-p65 and p65 were measured by Western blotting. The expression was normalized to that of internal control, actin, Lamin B, respectively, and quantified using ImageJ Software. Control, vehicle only. Data represent the means ± SD. SD: standard deviation, ns: not significant, ***p<0.01, ***p<0.001 vs. the control; #p<0.05, ##p<0.01, ###p<0.001 vs. LPS. LPS: lipopolysaccharide, PD: PD98059, SP: SP600125, SB: SB203580.
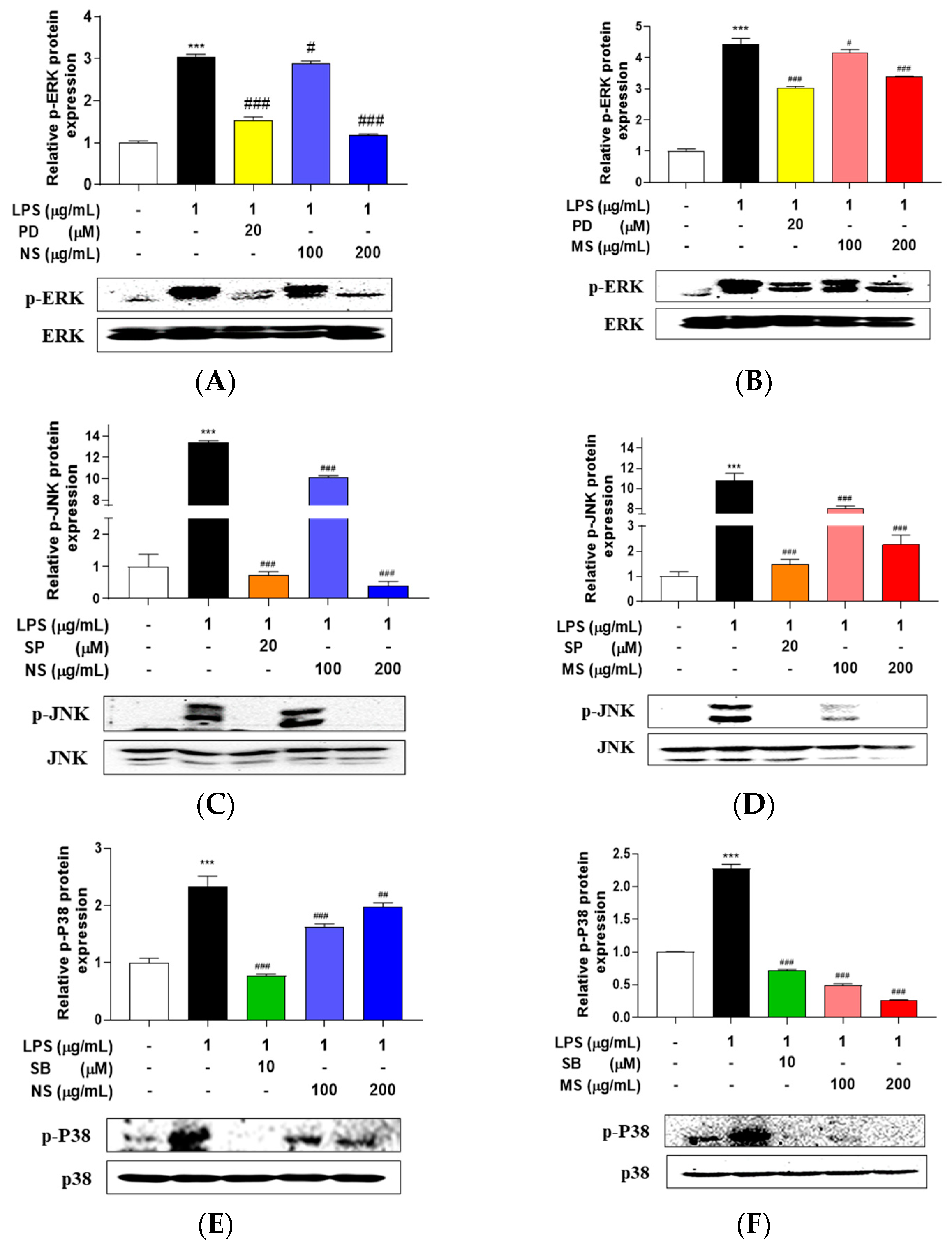
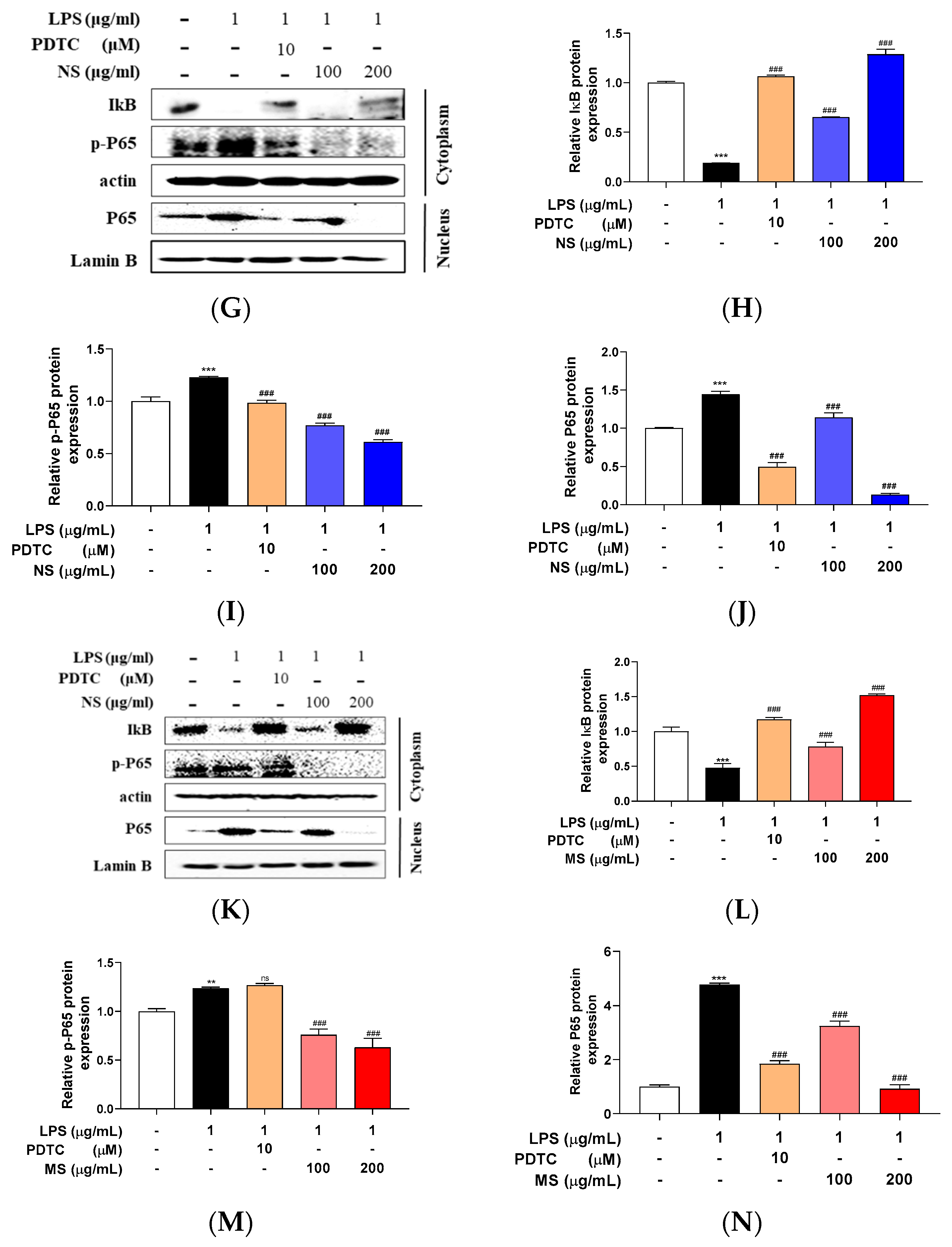

Table 1.
Primer Sequence.
| Gene name | Sequence 5' - 3' |
|---|---|
| il-1β | 5` - TCAAACCCCAATCCACAGAG- 3` |
| 5` - TCACTTCACGCTCTTGGATG- 3` | |
| cox-2a | 5` - AGCCCTACTCATCCTTTGAGG - 3` |
| 5` - TCAACCTTGTCTACGTGACCATA - 3` | |
| β-actin | 5` - AATCTTGCGGTATCCACGAGACCA - 3` |
| 5` - TCTCCTTCTGCATCCTGTCAGCAA - 3` |
il-1β: Interleukin (il)-1β; cox-2a: cyclooxygenase-2a.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



