
Submitted:
11 October 2023
Posted:
12 October 2023
You are already at the latest version
Abstract
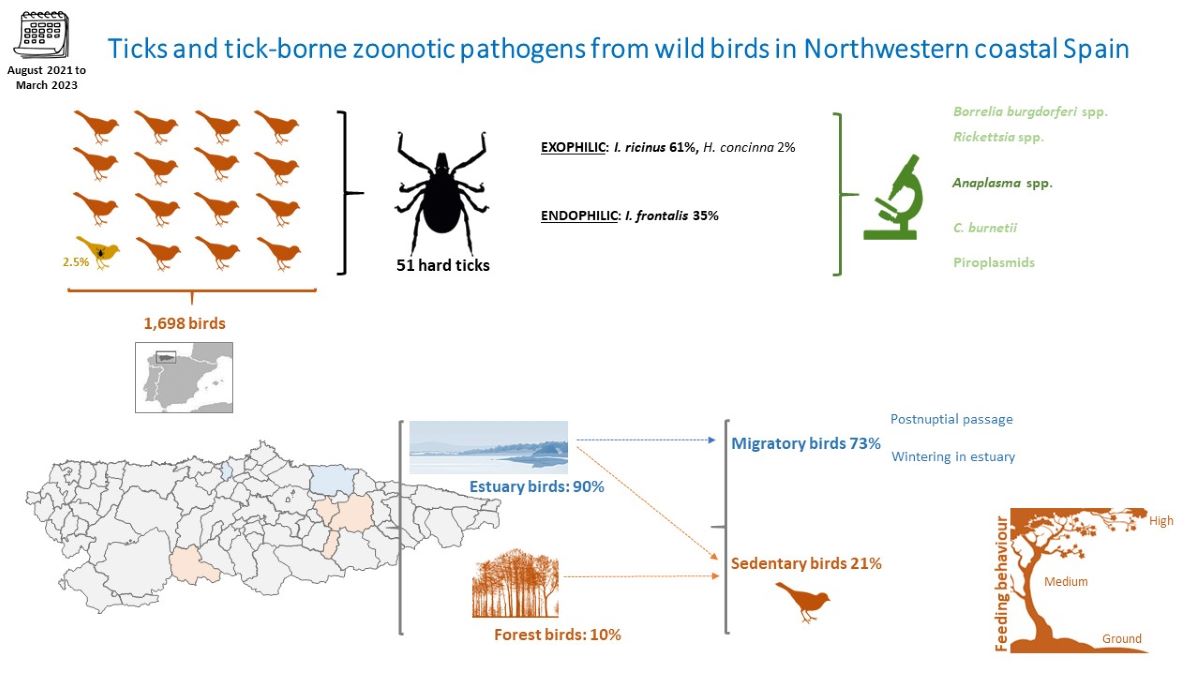
Migratory and local birds may disperse ticks and their associated pathogens. The aim of this study was to provide information regarding tick infesting birds in Asturias, region that accounts for most of the Lyme disease hospitalizations in Spain. From September 2021 and April 2023 trained and experienced bird- banders collected ticks from birds in six estuary and forest locations. A total of 1,698 birds (52 species, 38 genera and 26 families) were captured. A total 51 ticks (28 larvae, 20 nymphs and 3 females) were collected from 43 birds, belonging to three species: Ixodes ricinus (31), I. frontalis (18) and Haemaphysalis concinna (1). The average prevalence of tick infestation was 2.5% and the average tick burden was 1.2 ticks per host. The bird species Turdus merula, Parus major, Luscinia svecica and Anthus pratensis were among the most infested. Anaplasma phagocytophilum was detected from one Turdus merula. We have not detected B. burgdorferi s.l., Rickettsia spp., Coxiella burnetii or piroplasmids in any of the 51 analyzed ticks. The results of the study suggest low infestation rate in migratory/estuary birds and a higher rate in forest/sedentary ones. Despite this, the detection of pathogens, although with low prevalence, can pose a risk to public health.
Keywords:
wild birds
; ticks
; Ixodes spp.
; A. phagocytophilum
; Borrelia spp.
; Rickettsia spp.
; piroplasms
; Spain
1. Introduction
It is known that birds can be infested by ectoparasites such as mites, ticks, fleas and lice. These ectoparasites, especially ticks, can carry several pathogens, and both ticks and pathogens can be dispersed by the birds over short, medium and long distances, posing a constant risk to local host populations in different geographic regions [1,2,3,4].
The annual cycle of migratory birds is marked by their seasonal movements between breeding and non-breeding sites. In addition, one of the best documented responses to recent climate change is the altered migratory behaviour of birds [5]. Besides, the Convention on the Conservation of Migratory Species of Wild Animals (CMS) launched in 2022 the first interactive “Eurasian-African Bird Migration Atlas” [6], a very useful tool not only for conservation but also for the study of the epidemiology of many diseases, as has been demonstrated in the case of avian influenza. In this context, it is essential to know the role of the ticks that infest birds from different locations of the migratory route that goes from Africa to Northern Europe, since the risks vary according to latitudes but there are epidemiological links between the situation in each of those locations.
The Iberian Peninsula, due to its strategic location, is a vital connection point for migratory birds between Europe and Africa. It is the main migratory corridor in Western Europe and hundreds of thousands of birds cross the Strait of Gibraltar twice a year on their migratory journey. The wetlands of the Cantabrian coast, where Asturias is located, play a fundamental role in the provision of essential resources during pre-nuptial migrations (March-April) to northern Europe and post-nuptial migrations (July-September) to southern Spain and Africa. In addition, the temperate climate of this region offers refuge to various species of breeding birds from the north, where winter conditions are adverse. A large group of species that spend the winter in Africa migrate exclusively to the Iberian Peninsula to breed [7,8].
In Spain, several tick-borne microorganisms such as Anaplasma, Borrelia, Rickettsia and Crimean-Congo hemorrhagic fever virus (CCHFV) were detected in ticks taken from birds, mainly in the inland areas of the country, confirming that birds can disperse vectors and microorganisms [9,10]. However, to our knowledge, no data are available on tick species associated with avian hosts in Asturias, a region located on the northern coast of Spain within the southern limit of distribution of Ixodes ricinus and other hygrophilous ticks and which accounts for most of the hospitalizations for Lyme disease in Spain [11].
This study aims to provide information on the avian tick burden on the migratory and sedentary birds that cross the Spanish Cantabrian coast to better understand their role in the tick- transmission cycles of tick-borne zoonotic diseases, especially Lyme borreliosis.
2. Materials and Methods
2.1. Ethical Statement
All birds were trapped during the normal trapping activities of staff members at the GIA Asturias-Torquilla ringing group (G00037), under a general ringing license from the Aranzadi Ringing Office, in accordance with Spanish regulations. Sampling of birds was approved by the Ministry of Rural Environment and Territorial Cohesion of the Principality of Asturias (ref. AUTO/2020/20871, AUTO/2020/20935, AUTO/2020/20937, AUTO/2020/20938 y AUTO/2020/20939 DECO/2021/15449 Y DICO/2022/16272).
2.2. Site Selection
Wild birds were surveyed on three sampling stations belonging to two types of habi-tats in Northern Spain: postnuptial passage in estuary, wintering in estuary and forest. Six ringing locations conformed these samplings (Figure 1): Villaviciosa estuary – Villaviciosa - 43°30’44” -5°23´40” and Nalón estuary - Soto del Barco - 43°32´11” -6º53´22” (postnuptial passage and wintering in estuary); and Fuensanta - Nava - 43°30´44” -5°23´40”, Les Prae-res - Nava- 43°20´45” -5°28´57”, Somiedo - Sta. María del Puerto 43°1´24.05” -6°13´29.06” and Sobrescobio - Ladines- 43°12´48.57” -5º25´48.57” (Forest).
Villaviciosa estuary showed vegetation of maritime reeds (Juncus maritimus), amphibious reedbeds of maritime cirpus (Scirpus maritimus), of reeds (Phagmites australis) and grasslands of marsh ballast (Elymus pycnanthus). There are also marshmallow spots (Althea officinalis). Two invasive plants have thrived in the area: Paspalum vaginatum and Cotula coronopifolia. Around these formations that border the Sordo River, there is a wide area in which there are hardly any arboreal elements, except on the banks of some of the drainage channels and in the few existing boundaries.
Nalon estuary was bordered by a meander of the Nalón River, in the internal marsh area. It has plots of agricultural use and areas of reed (P. australis) and rush (J. maritimus). In the surroundings there are also trees, mainly willows (Salix sp.) and alders (Alnus glutinosa). The cultivation regime consists of planting maize for feed.
Forests were located at low and medium altitude in mountain areas with predominantly oak and beech arboreal vegetation.
Samplings took place from August 2021 to March 2023. Postnuptial passage sampling comprised fourteen weekly sessions between mid-July and mid-October. The wintering samplings comprised six weekly sessions between November and March. Forest station occurred between mid-March and April.
2.3. Bird Trapping
Bird captures were made with mist nets, also known as Japanese nets (Figure 2). They are used worldwide and are the preferred trapping method, especially to catch live birds. Mist nets are very fine nets that go unnoticed by birds, which, when passing where they are installed, get trapped in the net pockets that are formed with the tensioners, which are horizontal threads that run through them. With this system the trapped animal does not suffer any harm. Their use is regulated and they can only be used for scientific purposes [12].
The constant effort station methodology, known by the acronym CES, has been followed: the same number of nets are always available at the same locations and at regular time intervals, which facilitates long-term studies in which biological and demographic parameters can be compared [13].
Table S1 compiles all available information for each of the ringed birds including date, type of migration, location, habitat, ring number, species, age, sex and tick parasitization data. Bird species were classified as ground, low, medium low, medium, medium high and high feeders. The group of migratory birds captured includes migratory in passage, summer migratory and wintering migratory birds. The sedentary birds are those that carry out their entire life cycle in these habitats, despite almost all of these species make short movements due to lack or abundance of feeding.
2.4. Ticks Collection
The captured birds were identified following Harrison and Svensson bird field guides [14,15,16], ringed for individual identification, inspected for attached ticks and quickly released back in the same area (Figure 3).
Each bird was examined for ticks paying special attention to the areas around the head of eyes, ears, beak, crown and neck (Figure 4). All attached ticks were removed using fine tweezers and stored in tubes with ethanol (70%) until specific morphological identification in the laboratory. In addition, other ectoparasites occasionally found on birds, such as lice or trombiculid mites, were also collected. Prevalence (percent infested) and mean intensity (mean number per infested host) [17] were calculated for each tick species on each infested bird species (Table 1).
2.5. Tick Identification
Each tick was morphologically identified using a stereomicroscopy (NIKON© SMZ 1270) and the taxonomic keys of Manilla and Estrada-Peña & Mihalca [18,19]. Regarding the genus Ixodes, larvae were identified according to Heylen et al. [20]. Ticks were photographed dorsally and ventrally using the software NIS-elements (NIKON©).
The degree of blood ingested by each tick was estimated as unfed (U), little feed (LF), half feed (HF) or fully feed (FF) following Sandelin et al. [21].
In order to confirm morphologically identification, especially when the ticks had been damaged during removal from the birds, they were subjected to molecular identification using primers targeting the mitochondrial 16S rRNA gene as previously described [22].
2.6. DNA Extraction and PCR Amplification and Sequencing Analysis
Total DNA was extracted from ticks using the DNeasy Blood and Tissue kit (Qiagen, Hilden, Germany) according to the manufacturer’s protocol. Ticks were analyzed individually or in pools according to bird host, tick species and tick stage. Five µl of tick DNA were used for the detection of B. burgdorferi s.l., Rickettsia spp., Anaplasma spp., C. burnetii and piroplasmids by previously reported real-time and conventional PCR assays [23]. Pathogen detection was confirmed by sequencing both strands of the PCR products of expected size (Eurofins Genomics, Köln, Germany) and obtained sequences were compared to those in the GenBank® database by using the Basic Local Alignment Search Tool (BLASTN) at the NCBI. Nucleotide sequence obtained in this study were submitted to GenBank® under accession number OR623250.
2.7. Statistical Analyses
Differences in total bird abundance and relative abundances of each bird family and species in each sampling station were assessed by Kruskal-Wallis tests. This test was also used to assess differences in overall parasitization and parasitization per tick species among bird species and sampling stations. Then, Mann-Whitney U-tests with Bonferroni adjustment for multiple tests (α = 0.05/number of pairwise comparisons) were used to establish differences between pairs of bird species.
We grouped species of birds by their foraging behaviour and also assigned to a migratory or sedentary group. Tick prevalence and intensity of infestation were compared among these groups.
Statistical analyses were performed with SPSS (IBM SPSS statistics version 20.0).
3. Results
3.1. Bird Species
A total of 1,698 birds belonging to 26 families were captured during ringing seasons between September 2021 and April 2023 (Table 1 and Table S1). Of these birds, a total of 90.3% (1,533/1,698) were captured in estuaries and 9.7% (165/1,698) in forested areas, with significant differences in bird abundance between three sampling stations (H’ = 494.31, p < 0.01). Overall, most frequent families were Acrocephalidae (n=334), Phylloscopidae (n=317), Emberizidae (n=295) and Motacillidae (n=183), although family diversity notably varied among sampling stations (Figure 5). We identified 52 different species belonging to 38 genera, the most frequent being the following in general Emberiza schoeniclus (n=294), P. collybita (n=283), A. schoenobaenus (n=233), A. pratensis (n=146) and A. scirpaceus (n=107). Most frequent bird species were R. ignicapilla (30), A. schoenobaenus (233) and E. schoeniclus (291) in forest, postnuptial passage and wintering estuary respectively.
Out of the 52 different bird species, 27 (1,009/1,698 birds; 59% of the total captured) were classified as ground feeders, 12 (265/1,698; 16%) as medium feeders, 12 (424/1,698; 25%) as high feeders and one not included in the classification (Alcedo atthis) (Table 1). A total of 22 species (1,233/1,698; 72.6%) were classified as migratory, 24 (363/1,698; 21.4%) as sedentary and 5 (102/1,698; 6.0%) can be migratory or sedentary (Table 1).
3.2. Tick Infestations
A total of 51 bird-attached ticks were removed from 43 individual birds (Table 1 and Figure 6), tick burden ranging from 1 to 4 ticks per bird.
The prevalence of ticks in birds was 2.5% (43/1,698) and the average tick burden was 1.2 ticks per bird (51/43). We found 28 larvae, 20 nymphs and 3 female adult ticks in the bird species included in analyses.
Ticks collected belonged to three species: I. ricinus (14 larvae, 15 nymphs and 2 adult females; from 31 birds), I. frontalis (14 larvae, 3 nymphs and 1 adult female; from 18 birds) and H. concinna nymph on a sedge warbler (A. schoenobaenus).
The number of ticks for each of the established degree of engorgement was 0 unfed, 9 little feed (3 adults, 3 nymphs and 3 larvae), 12 half feed (6 nymphs and 6 larvae) and 24 fully feed (5 nymphs and 19 larvae).
Considering all bird species, no differences in overall parasitization were observed between different ringing locations within any of the three sampling stations (p > 0.05 in all cases). The same occurred in the case of parasitization by I. ricinus (wintering estuary: H’ = 0.00 p = 1.00; Forest: H’ = 6.25. p = 0.28) and by I. frontalis (wintering estuary: H’ = 0.00 p = 1.00). However, significant differences in overall (H’ = 19.96, p < 0.01) and in I. ricinus parasitization (H’ = 6.54, p < 0.05) (Figure 6) but no in I. frontalis ((H’ = 2.45, p = 0.29) were observed between different sampling stations. In these cases, forest birds showed higher parasitization than both estuarian stations (Figure 7).
Significant differences in overall parasitization were observed between families from each sampling station (postnuptial estuary: H’ = 33.19, p < 0.01; Forest: H’ = 26.26. p < 0.01) except for the wintering estuary (H’ = 8.64, p = 0.73). Indeed, significant differences in overall parasitization were also observed between bird species captured from each sampling station (postnuptial estuary: H’ = 36.51, p < 0.05; Forest: H’ = 42.29. p < 0.01) except for the wintering estuary (H’ = 21.08, p = 0.39) (Figure 7). The species T. merula showed the highest parasitization (average ± SD) in wintering estuary (1.50 ± 1.91), postnuptial estuary (0.50 ± 0.71) and forest (0.09 ± 0.30) (Figure 8). Nevertheless, the overall parasitization did not vary for each bird species between different sampling stations (p > 0.05 in all cases). No differences in I. ricinus and I. frontalis parasitization were observed between families or species from each sampling station (p > 0.05 in all cases).
The level at which each species feeds did not influence global or specific parasitization in any of the sampling stations (p > 0.05 in all cases). No differences in global parasitization were observed between migratory behaviour (Forest: H’ = 1.63, p = 0.44; wintering estuary: H’ = 0.87, p = 0.83) except in birds from postnuptial estuary (H’ = 28.23, p < 0.01) (Figure 9). Overwintering and Sedentary/migratory birds showed higher global parasitization.
Birds, as expected, were mostly parasitized by immature ticks (94.1% of all ticks). Out of the total 51 collected ticks, only two I. ricinus and one I. frontalis adults were identified on a common blackbird (T. merula), marsh bunting (E. schoeniclus) and common chiffchaff (P. collibyta).
The ticks we collected from the birds were mostly located around the eye (13/18) or the ear hole (3/18) and also on the beak (2/18) (Figure 4), although we could not have this information for all the birds.
Lice were found on 2 birds (Tringa totanus and Actitis hypoleucos) and trombiculid mites on a dunlin (Calidris alpina).
3.3. Pathogen Surveillance
A total of 51 (3 adults, 20 nymphs and 28 larvae) belonging to I. ricinus (31), I. frontalis (18) and H. concinna (1), were analyzed in 39 pools for B. burgdorferi s.l., Rickettsia spp., Anaplasma spp., C. burnetii and piroplasmids detection. Anaplasma phagocytophilum was detected in one sample collected from one T. merula. Sequencing analysis of a fragment of the 16S rRNA gene (546 bp) revealed the presence of the A. phagocytophilum genetic variant “I”.
None of the samples tested positive for B. burgdorferi s.l., Rickettsia spp., C. burnetii or piroplasmids.
4. Discussion
Previous studies carried out in northern Europe have shown that the tick exotic species are mostly those inhabiting the Southern and Central parts of Europe, rather than those native to Africa or Western Asia, because ticks from those areas seldom stay attached to birds for long enough [24,25]. In Southern Europe, the arrival of infected ticks transported by migratory birds from Africa is a fact that has already been confirmed by previous studies [9]. Cantabrian regions of Spain are located in an intermediate geographical position, without adequate environmental conditions for the settlement and survival of xerophytic ticks of the genus Hyalomma. By contrast, this area is the Southern limit of distribution of hygrophilous species such as I. ricinus [26], a recognized vector of Lyme borreliosis, a highly prevalent human disease in Asturias [11].
Among the 1,698 ringed birds we found significant differences in bird relative abundance between the three sampling stations belonging to two types of habitats: postnuptial passage in estuary, wintering in estuary and forest. During the post-nuptial passage in the estuary the most abundant family of ringed birds was the Acrocephalidae, while during the wintering, Paridae in the estuary and Phylloscopidae and Emberizidae in the forest were most frequent families. This highlights that the different proportion of families and species of birds examined due to geographical but also temporal factors, would explain an important part of the variability of the results obtained in different studies.
The bird species in our study, mostly passerines, showed a remarkable 2.5% tick prevalence, consistent with other studies across Europe, such as the prevalence of 1,7% reported in Greece [27], 3.1% in Germany [28], 4.4% in Poland [29] or 2.0% in Sweden-Denmark [30] but far from the 58.0% reported in a Lyme endemic area in Switzerland [31], 41.1% in Czechia [32], 36.7 in Slovakia & Czechia [33] or 32.4 (Mafra -Lisboa) and 16.7 % (Coimbra) reported in two forested areas in western Portugal [34]. A detailed analysis of the epidemiological studies carried out in Europe in the last 30 years (Table S2) revealed great methodological differences between them, both in geographical and climatic characteristics of the studied areas, diversity and abundance of bird species and other tick host animals, or in the capture period. In this regard, it should be noted that in our study we also found significant differences in overall and in I. ricinus parasitization (but no in I. frontalis) between different sampling stations. In these cases, forest birds showed higher parasitization than both estuarian stations. We must keep in mind that many of the studied bird species have a clear habitat preference. In fact, of the 52 species identified in our study, 32 were captured exclusively in estuaries and 7 exclusively in forests.
Many studies suggest that the level of parasitism in birds is a consequence of their feeding behaviour, which brings them often into contact with subadult ticks but also of the diversity and dominant tick species at each study area [31,32,33,34,35,36,37,38,39]. However, in our study the level at which each species feeds did not influence global or specific parasitization in any of the sampling stations. The difficulty in fit each bird species into a specific feeding behaviour category could be one of the possible causes. The most common is to divide the birds into groups of ground-feeders and non-ground-feeders, with more or less intermediate groups according to the different authors. We consider that a categorization reduced to two groups is not realistic and that, on the contrary, the establishment of an excessive number of groups does not allow us to appreciate trends in terms of tick prevalence and mean tick infestation. Just as an example, indicate that the same species (common chiffchaff – P. collybita) is grouped as a “ground-feeder” (two groups) by Klaus et al. [28] in Germany and as a “high foraging level” (four groups) by Norte et al. in [34] in Portugal.
No differences in global parasitization were observed between migratory behaviour except in birds from postnuptial estuary. Something similar to what we have mentioned for feeding behaviour occurs with the division of bird species into “migratory” or “sedentary”, given that in the same species in Asturias (F. coelebs, C. chloris, E. rubecula, M. alba and S. atricapilla) we find populations that migrate and others that remain in the region.
Tick burden, considered an important indicator of reservoir competence, had a low range of 0–4 ticks per bird. The mean tick abundance or tick load was 1.2 (51/43) ticks per infected bird. Our results are very close to those obtained in other European countries such as the 1.9 (335/173) ticks per infected bird in Poland [29], 1.8 (1,335/748) in Sweden [21]and 0.4 (212/107) in Russia [40] but lower than the 5.1 (2,240/562) described in Czechia [32], 3.8 (3,195/838) in Germany [28], 2.3 (417/180) and 1.9 (33/17) in two areas at different altitudes in Switzerland [41] or 2.1 (967/465) in Sweden & Denmark [30].
Ixodid tick species in Europe represent five genera: Ixodes, Haemaphysalis, Hyalomma, Dermacentor and Rhipicephalus but the last two usually do not parasitize birds [35]. In our study the species of ticks identified in the captured birds were almost exclusively belonging to the genus Ixodes (98%), with the only exception of one H. concinna nymph found in a common warbler (A. schoenobaenus). Although H. concinna larvae and nymphs occurred more frequently on arboreal birds, its relatively high questing height on the vegetation can explain our finding on a ground-feeding bird species, similar to that described in other central European countries [36]. The genus Hyalomma has not been described for the moment in our region due to its marked xerophytic character, although it has been identified in bordering regions to the south of the Cantabrian Mountains.
In published studies on tick species that parasitize birds [28,29,42], both endophilic (I. frontalis, I. acuminatus, I. canisuga, I. arboricola, I. lividus, I. trianguliceps) and exophilic species (I. ricinus, H. punctata, Hyalomma spp.) are described, although the latter vary much more according to the study areas. We also found endophilic (I. frontalis 35.3%, 18/51) and exophilic species (I. ricinus 60.8%, 31/51 and H. concinna 2.0%, 1/51). Among exophilic species, I. ricinus predominance is consistent with their abundance in the vegetation of Asturias where is by far the most abundant species for all stages as well as the one with a longer period of activity all around the year [26]. Among endophilic species, I. frontalis also showed notable prevalence values, which is striking if we take into account that almost all birds in our study area need to build a new nest every year due to the winter conditions, whereas the larvae of endophilic tick species are assumed to overwintering in the nest burrows until birds return. Despite the reduced variety of tick species found infesting birds in Asturias, our results agree with those of Heylen et al. [43], who also found almost exclusively the same two species: I. ricinus and I. frontalis in seven locations in the Netherlands.
Birds, according to many previous studies, were predominantly infested by immature I. ricinus stages and rarely by adult females. On the contrary, all I. frontalis developmental stages feed on birds therefore, a greater proportion of adult specimens would be expected for I. frontalis compared to I. ricinus. Our results do not reflect these differences but rather very similar relative proportions (5.6 and 6.4% respectively) of adults for both species. Although the number of ticks available does not allow us to deep into this topic, the 3 adult specimens of ticks came from 3 different bird families captured in the estuaries at the end of autumn.
I. ricinus is not only the predominant species in forest birds in Asturias, but also shows an infestation prevalence of 9.1% (15/165), well above the 1.0% (16/1,533) of this same species in estuary birds. Norte et al. [34] showed that the seasonal pattern of tick infestation on birds was similar to that of questing ticks, at least as regards the immature stages of the species I. ricinus. In that sense, I. ricinus is the predominant species in the vegetation of Asturias, showing much higher abundances in forest areas than in coastal areas. Regarding I. frontalis, this species showed lower prevalence of infestation in both forests areas (0.6%, 1/165) and estuaries (1.1%, 17/1,533).
Regarding occurrence of tick-borne pathogens in ticks infesting birds, the only zoonotic bacteria detected was A. phagocytophilum. In a previous work carried out in our region [44], we already detected high prevalence of this pathogen (61.0% in roe deer and 80.8% in red deer) suggesting the relevance of deer as reservoir host of A. phagocytophilum that could act as source of infection for vector ticks. The genetic variant “I” has been previously identified in Asturias in a questing adult I. ricinus and in a wolf [45]. The variant detected is not pathogenic and has already been described in I. ricinus ticks collected from roe deer and tissues as well as in the vegetation in the autonomous community of Galicia [46], which borders Asturias.
The detection of B. burgdorferi s.l. among questing ticks and small mammals in Asturias [23,26], as well as the abundance of ticks and of large wild and domestic mammals, indicate a high risk of this infection in the region, where Lyme borreliosis is highly prevalent [11]. Despite this, we have not detected B. burgdorferi s.l. in any of the 51 analyzed ticks. The reasons that may explain this are, first of all, the small number of ticks examined despite the high number of birds captured, due to the low prevalence of infestation. Also, 28 of the 51 analyzed ticks (51.9%) were larvae that are not usually carriers of B. burgdorferi s.l., since vertical transmission of this pathogen is very rare, although in some cases they can be infected by co-feeding. Furthermore, only 17 of the remaining nymphs and adults were I. ricinus. In a previous study in the same area we detected B. burgdorferi s.l. in 1.4% (12/845) of I. ricinus questing nymphs and 9.1% (2/33) of questing adults [26] which, despite being a relevant percentage, makes it improbable to detect this pathogen in a sample of only 15 nymphs and 2 adults of this species. Finally, we must also assess that most of the birds were captured in coastal estuaries, 1,533 birds compared to only 165 in forest areas and the abundance of ticks is much lower in these coastal areas than in more wooded inland zones. This fact, which we verified in many tick dragging carried out in the vegetation of the region, is also reflected in the lower prevalence of infestation of birds caught in seashore compared to those in forested areas.
5. Conclusions
Ixodes ricinus and I. frontalis are the most common tick infesting birds of several species. Turdus merula, P. major, L. svecica and A. pratensis were some of the bird species that presented the highest prevalence of infesting ticks. Anaplasma phagocytophilum was detected in one I. ricinus collected from one T. merula. None of the rest of the pathogens tested were detected in any of the analyzed ticks. Updated knowledge of tick bird species and tick-borne pathogens that covers as many areas as possible is essential for understanding their possible impact on public health.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: List of all the ringed birds and data collected from them prepared by the members of the “Torquilla” ringing group. Table S2: Epidemiological studies of tick infestation in birds in Europe.
Author Contributions
Conceptualization, A.E., A.S. and A.C.; methodology, P.P.-T., JV.G.-E., A.E. and A.C.; formal analysis, A.S.; data curation, P.P.-T., JV.G.-E. and A.E.; writing—original draft preparation, A.E.; writing—review and editing, A.E., A.S., A.C., P.P.-T. and JV.G.-E.; funding acquisition, A.C., A.E. and A.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by PCTI 2021-2023 (GRUPIN: IDI2021-000102).
Institutional Review Board Statement
Ethical review and approval were waived for this study due to the fact that birds were captured during the normal trapping activities of members of the “Torquilla” ringing group, belonging to GIA (Iberian Ringing Group). See also chapter 2.1. Ethical statemen for more details.
Data Availability Statement
data supporting reported results can be found in the supplementary materials cited above
Acknowledgments
We would like to thank all the volunteers bird-ringers for their collaboration, pivotal in this study.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Brinkerhoff, R.J.; Folsom-O’Keefe, C.M.; Tsao, K.; Diuk-Wasser, M.A. Do birds affect Lyme Disease risk? Range expansion of the vector-borne pathogen Borrelia burgdorferi. Front. Ecol. Environ. 2011, 9, 103–110. [Google Scholar] [CrossRef]
- Hornok, S.; Karcza, Z.; Csörgo, T. Birds as disseminators of ixodid ticks and tick- borne pathogens: note on the relevance to migratory routes. Ornis. Hung. 2012, 20, 86–89. [Google Scholar] [CrossRef]
- Vuong, H.B.; Canham, C.D.; Fonseca, D.M.; Brisson, D.; Morin, P.J.; Smouse, P.E.; Ostfeld, R.S. Occurrence and transmission efficiencies of Borrelia burgdorferi OspC types in avian and mammalian wildlife. Infect. Genet. Evol. 2014, 27, 594–600. [Google Scholar] [CrossRef]
- Sparagano, O.; George, D.; Giangaspero, A.; Spitalska, E. Arthropods and associated arthropod-borne diseases transmitted by migrating birds. The case of ticks and tick-borne pathogens. Vet. Parasitol. 2015, 213, 61–6. [Google Scholar] [CrossRef]
- Jones, T.; Cresswell, W. The phenology mismatch hypothesis: Are declines of migrant birds linked to uneven global climate change? J. Anim. Ecol. 2010, 79, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Convention on Migratory Species (CMS). Eurasian-African Bird Migration Atlas. 2023. Available online: https://migrationatlas.org/ (accessed on 03 October 2023).
- Gordo, O.; Sanz, J.J.; Lobo, J.M. La migración de las aves sobre la península Ibérica. Quercus 2009, 280, 14–22. [Google Scholar]
- Moncada-Lorén, M. España, uno de los principales corredores migratorios del mundo por su ubicación geográfica. National Geographic España 2018, 11 mayo. https://www.nationalgeographic.es/animales/2018/05/espana-uno-de-los-principales-corredores-migratorios-del-mundo-por-su-ubicacion-geografica.
- Palomar, A. M.; Portillo, A.; Santibáñez, P.; Mazuelas, D.; Arizaga, J.; Crespo, A.; Gutiérrez, Ó.; Cuadrado, J. F.; Oteo, J. A. Crimean-Congo hemorrhagic fever virus in ticks from migratory birds, Morocco. Emerg. Infect. Dis. 2013, 19, 260–263. [Google Scholar] [CrossRef] [PubMed]
- Palomar, A.M.; Santibáñez, P.; Mazuelas, D.; Roncero, L.; Santibáñez, S.; Portillo, A. Oteo; J.A. Role of birds in dispersal of etiologic agents of tick-borne zoonoses, Spain. Emerg. Infect. Dis. 2012, 18, 1188–1191. [Google Scholar] [CrossRef]
- Alguacil, M. A. Carga hospitalaria de enfermedad de Lyme en España (2005-2019). In Boletín Epidemiológico Semanal (ISCIII) 2022, 30. Available in: https://revista.isciii.es/index.php/bes/article/view/1214.
- Sutherland, W.J.; Newton, I.; Green, R. Bird Ecology and Conservation: A Handbook of Techniques, Techniques in Ecology & Conservation. Oxford, 2004; online Edn, Oxford Academic, 1 Sept. 2007. Accessed 19 July 2023. [CrossRef]
- Robinson, R.A.; Julliard, R.; Saracco, J.F. Constant effort: Studying avian population processes using standardised ringing. Ringing & Migration 2009, 24, 199–204. [Google Scholar] [CrossRef]
- Harrison, C. Guía de Campo de los nidos, huevos y polluelos de las aves de España y de Europa. Ed. Omega. Barcelona, 1991.
- Svensson, L. Identification Guide to European Passerines. Stockholm, 1992.
- Svensson, L.; Mullarney, K.; Zetterstrom, D. Guía de Aves de España, Europa y Región Mediterránea. Ed. Omega. Barcelona, 2010.
- Durden, L. A.; Oliver, J. H.; Kinsey, A.A. Ticks (Acari: Ixodidae) and spirochetes (Spirochaetaceae: Spirochaetales) recovered from birds on a Georgia Barrier Island. J. Med. Entomol. 2001, 38, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Manilla, G. Fauna D’Italia: Acari. Ixodida. Edizioni Calderini. Bologna, 1998.
- Estrada-Peña, A.; Mihalca, A.D.; Petney, T.N. Editors. Ticks of Europe and North Africa. A guide to species identification. Springer International Publishing. New York, 2017.
- Heylen, D.; de Coninck, E.; Jansen, F.; Madder, M. Differential diagnosis of three common Ixodes spp. ticks infesting songbirds of Western Europe: Ixodes arboricola, I. frontalis and I. ricinus. Ticks and Tick-Borne Dis. 2014, 5, 693–700. [Google Scholar] [CrossRef] [PubMed]
- Sandelin, L. L.; Tolf, C.; Larsson, S.; Wilhelmsson, P.; Salaneck, E.; Jaenson, T. G. T.; Lindgren, P. E.; Olsen, B.; Waldenström, J. Candidatus Neoehrlichia mikurensis in ticks from migrating birds in Sweden. PLoS ONE 2015, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Black, W.C.; Piesman, J. Phylogeny of hard- and soft-tick taxa (Acari: ixodida) based on mitochondrial 16S rDNA sequences. In Proc. Natl. Acad. Sci. USA 1994, 91, 10034–10038. [Google Scholar] [CrossRef] [PubMed]
- Del Cerro, A.; Oleaga, A.; Somoano, A.; Barandika, J.F.; García-Pérez, A.L.; Espí, A. Molecular identification of tick-borne pathogens (Rickettsia spp., Anaplasma phagocytophilum, Borrelia burgdorferi sensu lato, Coxiella burnetii and piroplasms) in questing and feeding hard ticks from North-Western Spain. Ticks Tick Borne Dis. 2022, 13, 101961. [Google Scholar] [CrossRef] [PubMed]
- Capligina, V.; Salmane, I.; Keišs, O.; Vilks, K.; Japina, K.; Baumanis, V.; Ranka, R. Prevalence of tick- borne pathogens in ticks collected from migratory birds in Latvia. Ticks Tick Borne Dis 2014, 5, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Ciebiera, O.; Jerzak, L.; Nowak-Chmura, M.; Bocheński, M. Ticks (Acari: Ixodida) on birds (Aves) migrating through the Polish Baltic coast. Exp Appl Acarol 2019, 77, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Espí, A.; del Cerro, A.; Somoano, A.; García, V.; Prieto, J.M.; Barandika, J.F.; García-Pérez, A.L. Borrelia burgdorferi sensu lato prevalence and diversity in ticks and small mammals in a Lyme borreliosis endemic Nature Reserve in North-Western Spain. Incidence in surrounding human populations. Enferm. Infecc. Microbiol. Clin. 2017, 35, 563–568. [Google Scholar] [CrossRef]
- Diakou, A.; Norte, A.C.; Lopes de Carvalho, I.; Núncio, S.; Nováková, M.; Kautman, M.; Alivizatos, H.; Kazantzidis, S.; Sychra, O.; Literák, I. Ticks and tick-borne pathogens in wild birds in Greece. Parasitol. Res. 2016, 115, 2011–2016. [Google Scholar] [CrossRef]
- Klaus, C.; Gethmann, J.; Hoffmann, B.; Ziegler, U.; Heller, M.; Beer, M. Tick infestation in birds and prevalence of pathogens in ticks collected from different places in Germany. Parasitol. Res. 2016, 115, 2729–2740. [Google Scholar] [CrossRef]
- Zając, Z.; Kulisz, J.; Kunc-Kozioł, R.; Woźniak, A.; Filipiuk, M.; Rudolf, R.; Bartosik, K.; Cabezas-Cruz, A. Tick Infestation in Migratory Birds of the Vistula River Valley, Poland. Int. J. Environ. Res. Public Health 2022, 19, 13781. [Google Scholar] [CrossRef]
- Olsen, B.; Jaenson, T. G. T.; Bergstrom, S. Prevalence of Borrelia burgdorferi sensu lato-infected ticks on migrating birds. Appl. Environ. Microbiol. 1995, 61, 3082–3087. [Google Scholar] [CrossRef]
- Humair, P.F.; Turrian, N.; Aeschlimann, A.; Gern, L. Ixodes ricinus immatures on birds in a focus of Lyme borreliosis. Folia Parasitol. 1993, 40, 237–242. [Google Scholar]
- Dubska, L.; Literak, I.; Kocianova, E.; Taragelova, V.; Sychra, O. Differential role of passerine birds in distribution of Borrelia Spirochetes, based on data from ticks collected from birds during the postbreeding migration period in central Europe. Appl Env Microbiol. 2009, 75, 596–602. [Google Scholar] [CrossRef]
- Taragelová, V.; Koči, J.; Hanincová, K.; Kurtenbach, K.; Derdáková, M.; Ogden, N.H.; Literák, I.; Kocianová, E.; Labuda, M. Blackbirds and song thrushes constitute a key reservoir of Borrelia garinii, the causative agent of Borreliosis in central Europe. Appl Environ Microbiol. 2008, 74, 1289–1293. [Google Scholar] [CrossRef] [PubMed]
- Norte, A. C.; de Carvalho, I. L.; Ramos, J. A.; Gonçalves, M.; Gern, L.; Núncio, M. S. Diversity and seasonal patterns of ticks parasitizing wild birds in western Portugal. Exp. Appl. Acarol. 2012, 58, 327–339. [Google Scholar] [CrossRef]
- Hasle, G. Transport of ixodid ticks and tick-borne pathogens by migratory birds. Front. Cell. Infect. Microbiol. 2013, 10, 48. [Google Scholar] [CrossRef] [PubMed]
- Hornok, S.; Flaisz, B.; Takács, N.; Kontschán, J.; Csörgo, T.; Csipak, A.; Jaksa, B. R.; Kováts, D. Bird ticks in Hungary reflect western, southern, eastern flyway connections and two genetic lineages of Ixodes frontalis and Haemaphysalis concinna. Parasit. Vectors. 2016, 9, 1–9. [Google Scholar] [CrossRef]
- Papadopoulos, B.; Humair, P.F.; Aeschlimann, A.; Vaucher, C.; Buttiker, W. Ticks on birds in Switzerland. Acarologia 2001, 42, 3–19. [Google Scholar]
- Movila, A.; Gatewood, A.; Toderas, I.; Duca, M.; Papero, M.; Uspenskaia, I.; Conovalov, J.; Fish, D. Prevalence of Borrelia burgdorferi sensu lato in Ixodes ricinus and I. lividus ticks collected from wild birds in the Republic of Moldova. Int. J. Med. Microbiol. 2008, 298, 149–153. [Google Scholar] [CrossRef]
- James, M.C.; Furness, R.W.; Bowman, A.S.; Forbes, K.J.; Gilbert, L. The importance of passerine birds as tick hosts and in the transmission of Borrelia burgdorferi, the agent of Lyme disease: a case study from Scotland. Ibis 2011, 153, 293–302. [Google Scholar] [CrossRef]
- Movila, A. , Alekseev, A. N., Dubinina, H. V., & Toderas, I. Detection of tick-borne pathogens in ticks from migratory birds in the Baltic region of Russia. Med. Vet. Entomol. 2013, 27, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Poupon, M.A.; Lommano, E.; Humair, P.F.; Douet, V.; Rais, O.; Schaad, M.; Jenni, L.; Gern, L. Prevalence of Borrelia burgdorferi sensu lato in ticks collected from migratory birds in Switzerland. Appl. Environ. Microbiol. 2006, 72, 976–979. [Google Scholar] [CrossRef] [PubMed]
- Norte, A. C.; da Silva, L. P.; Tenreiro, P. J. Q.; Felgueiras, M. S.; Araújo, P. M.; Lopes, P. B.; Matos, C.; Rosa, A.; Ferreira, P. J. S. G.; Encarnação, P.; Rocha, A.; Escudero, R.; Anda, P.; Núncio, M. S.; Lopes de Carvalho, I. Patterns of tick infestation and their Borrelia burgdorferi s.l. infection in wild birds in Portugal. Ticks and Tick-Borne Dis. 2015, 6, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Heylen, D.; Fonville, M.; Docters Van Leeuwen, A.; Stroo, A.; Duisterwinkel, M.; Van Wieren, S.; Diuk-Wasser, M.; De Bruin, A.; Sprong, H. Pathogen communities of songbird-derived ticks in Europe’s low countries. Parasit. Vectors. 2017, 10, 1–12. [Google Scholar] [CrossRef]
- García-Pérez, A. L.; Oporto, B.; Espí, A.; del Cerro, A.; Barral, M.; Povedano, I.; Barandika, J. F.; Hurtado, A. Anaplasmataceae in wild ungulates and carnivores in northern Spain. Ticks and Tick-Borne Diseases 2016, 7, 264–269. [Google Scholar] [CrossRef]
- Del Cerro, A.; Oleaga, A.; Somoano, A.; Balseiro, A.M.; Royo, L.; Espí, A. Phylogenetic analysis of Anaplasma species in wild animals in North Western Spain. In Proceedings of the International congress on Rickettsiae (ESCCAR), Lausanne, Switzerland, 23-26 August 2022. [Google Scholar]
- Remesar, S.; Díaz, P.; Prieto, A.; García-Dios, D.; Fernández, G.; López, C.M.; Panadero, R.; Díez-Baños, P.; Morrondo, P. Prevalence and molecular characterization of Anaplasma phagocytophilum in roe deer (Capreolus capreolus) from Spain. Ticks Tick Borne Dis. 2020, 11, 101351. [Google Scholar] [CrossRef]
Figure 1.
Bird ringing locations in Asturias. Numbers in parentheses represent the number of birds captured from corresponding locations. Blue and red dots represent respectively estuary and forest areas (the size of the dots in the figure is proportional to the number of birds captured). The text boxes show the prevalence of infestation in the two differentiated areas and for each identified tick species.
Figure 1.
Bird ringing locations in Asturias. Numbers in parentheses represent the number of birds captured from corresponding locations. Blue and red dots represent respectively estuary and forest areas (the size of the dots in the figure is proportional to the number of birds captured). The text boxes show the prevalence of infestation in the two differentiated areas and for each identified tick species.
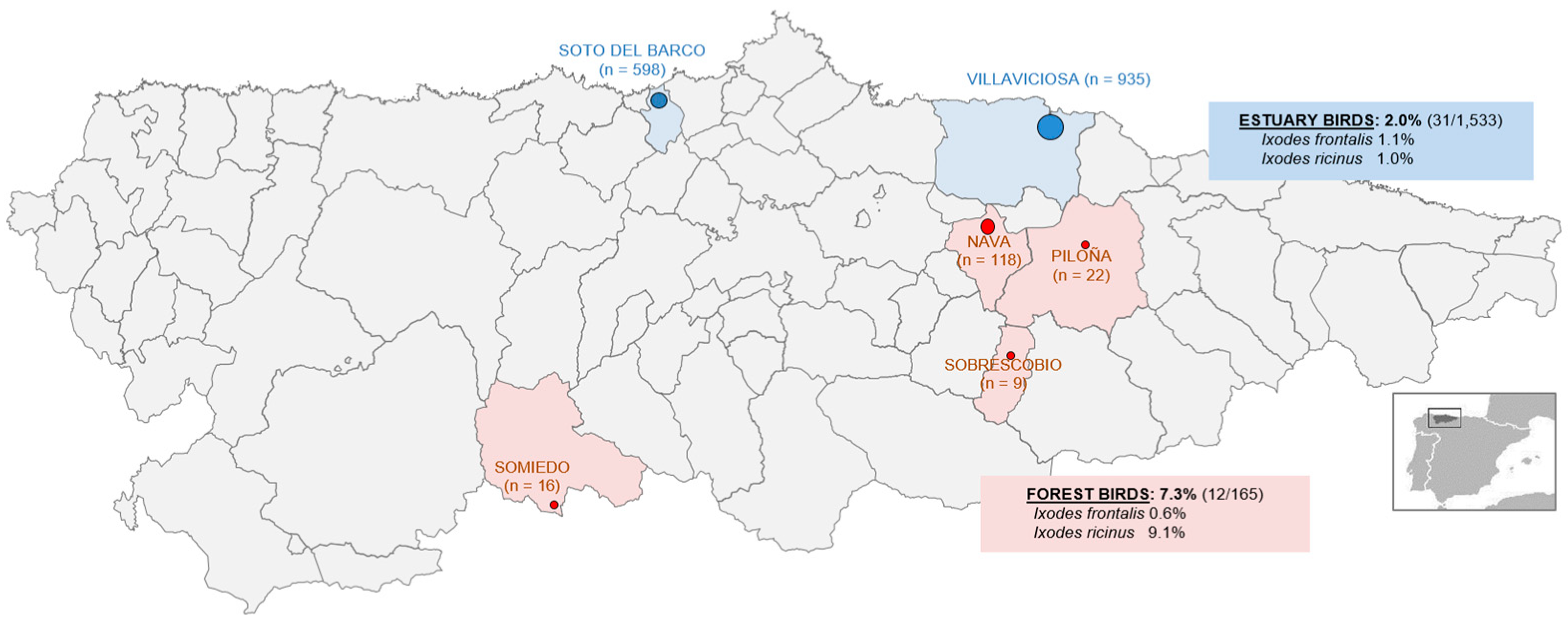
Figure 2.
Members of the GIA-Asturias-Torquilla Ringing Group releasing the small birds caught in the Japanese nets placed in the Villaviciosa estuary (photo on the right: common reed warbler, Acrocephalus scirpaceus).
Figure 2.
Members of the GIA-Asturias-Torquilla Ringing Group releasing the small birds caught in the Japanese nets placed in the Villaviciosa estuary (photo on the right: common reed warbler, Acrocephalus scirpaceus).

Figure 3.
Members of the GIA-Asturias-Torquilla Ringing Group banding, identifying and measuring the birds in the Villaviciosa estuary.
Figure 3.
Members of the GIA-Asturias-Torquilla Ringing Group banding, identifying and measuring the birds in the Villaviciosa estuary.

Figure 4.
Tick found attached to a common reed warbler (Acrocephalus scirpaceus) during post-banding examination.
Figure 4.
Tick found attached to a common reed warbler (Acrocephalus scirpaceus) during post-banding examination.

Figure 5.
Relative abundances of the families for each sampling.
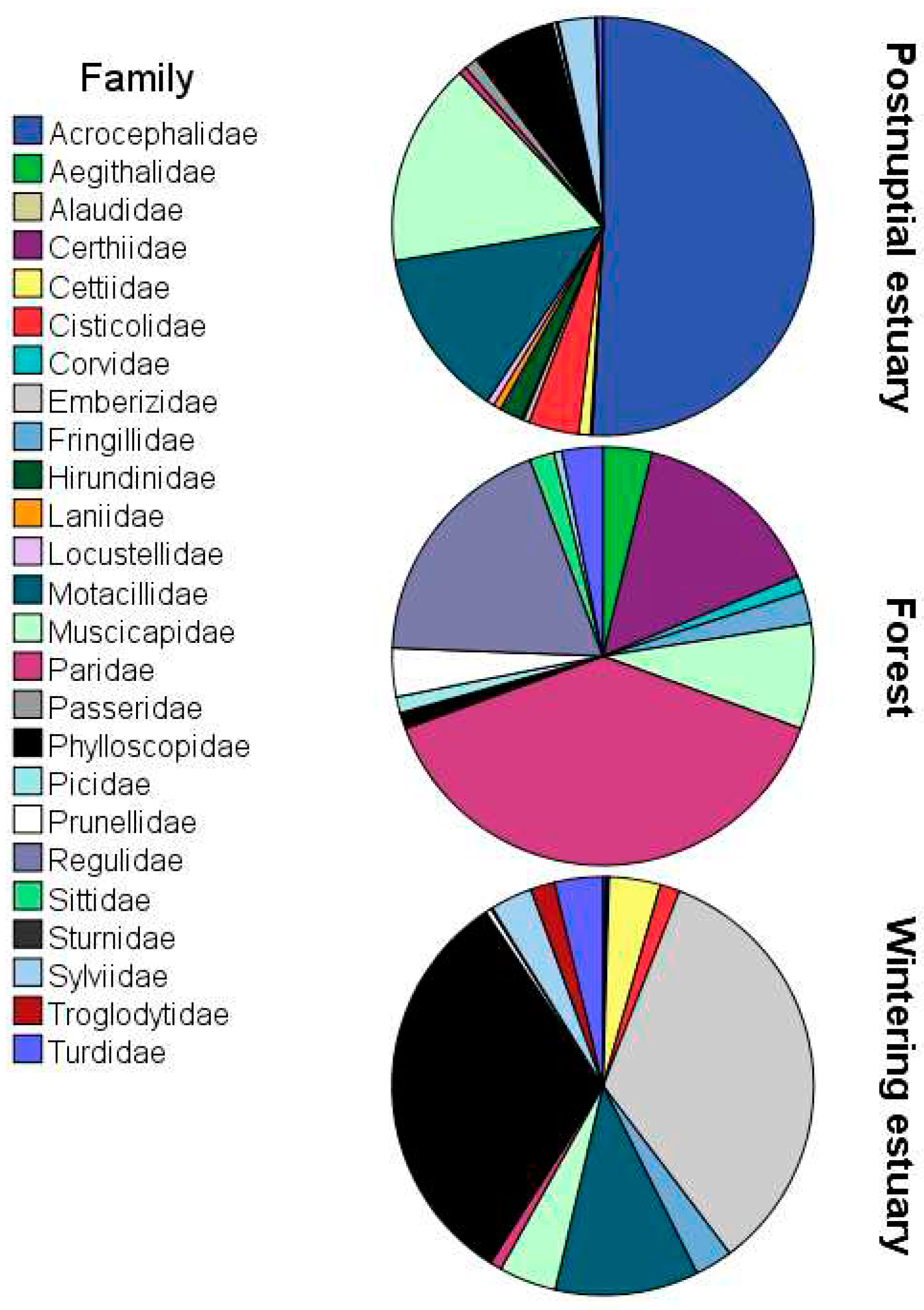
Figure 6.
Morphology of some tick species identified on birds in Asturias. (a) I. ricinus female from a T. merula captured in Villaviciosa estuary in December 2022; (b) I. frontalis female from a P. collybyta, captured in Navia estuary in November 2022; (c) H. concinna nymph from a A. schoenobaenus captured in Villaviciosa estuary in July 2022.
Figure 6.
Morphology of some tick species identified on birds in Asturias. (a) I. ricinus female from a T. merula captured in Villaviciosa estuary in December 2022; (b) I. frontalis female from a P. collybyta, captured in Navia estuary in November 2022; (c) H. concinna nymph from a A. schoenobaenus captured in Villaviciosa estuary in July 2022.

Figure 7.
Number of ticks (average ± SD) for each sampling station. Significant differences in number of ticks between sampling stations are indicated by different letters.
Figure 7.
Number of ticks (average ± SD) for each sampling station. Significant differences in number of ticks between sampling stations are indicated by different letters.
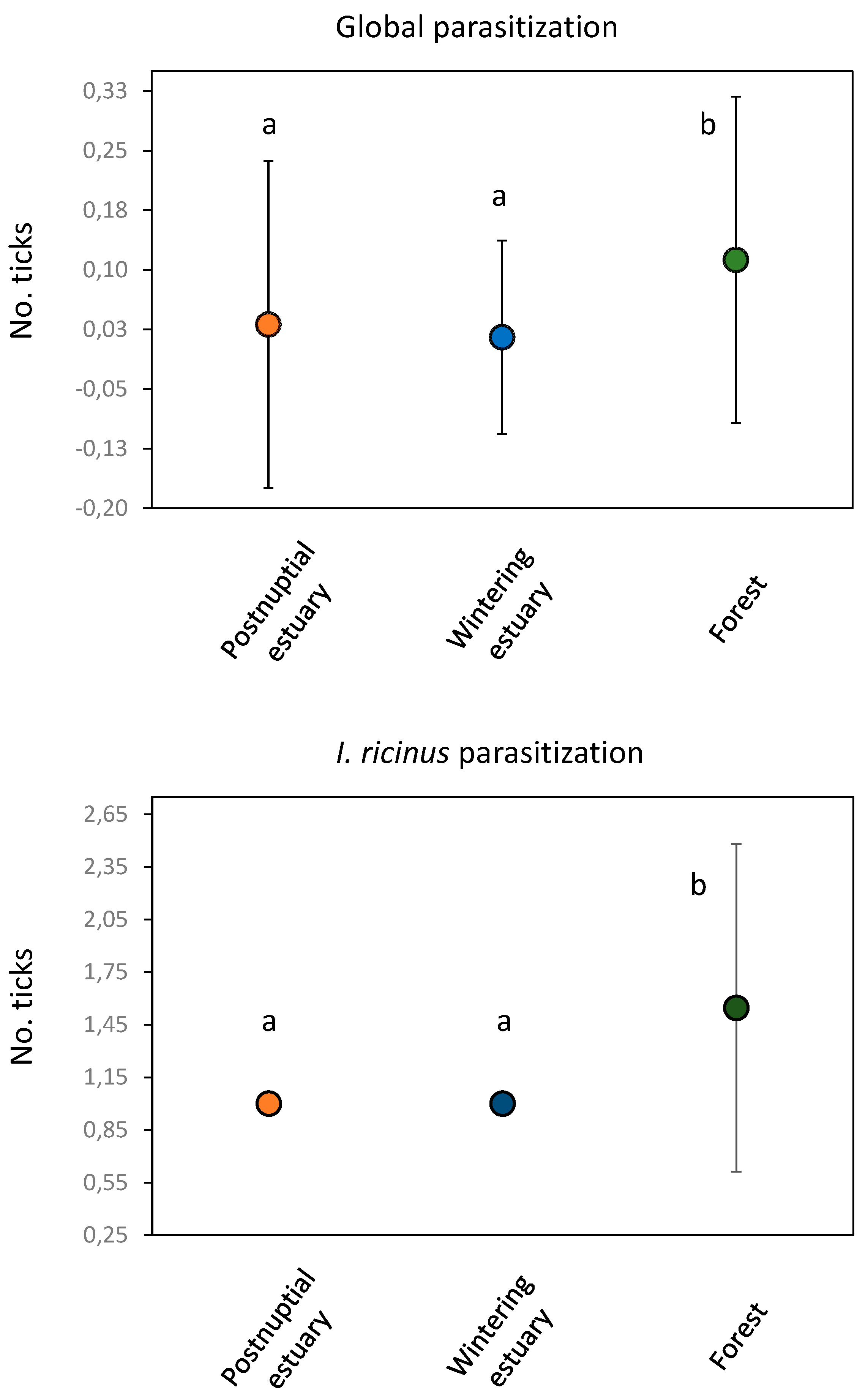
Figure 8.
Number of ticks (average ± SD) in each bird species captured in each sampling station. Significant differences in tick number between bird species are indicated by different letters.
Figure 8.
Number of ticks (average ± SD) in each bird species captured in each sampling station. Significant differences in tick number between bird species are indicated by different letters.
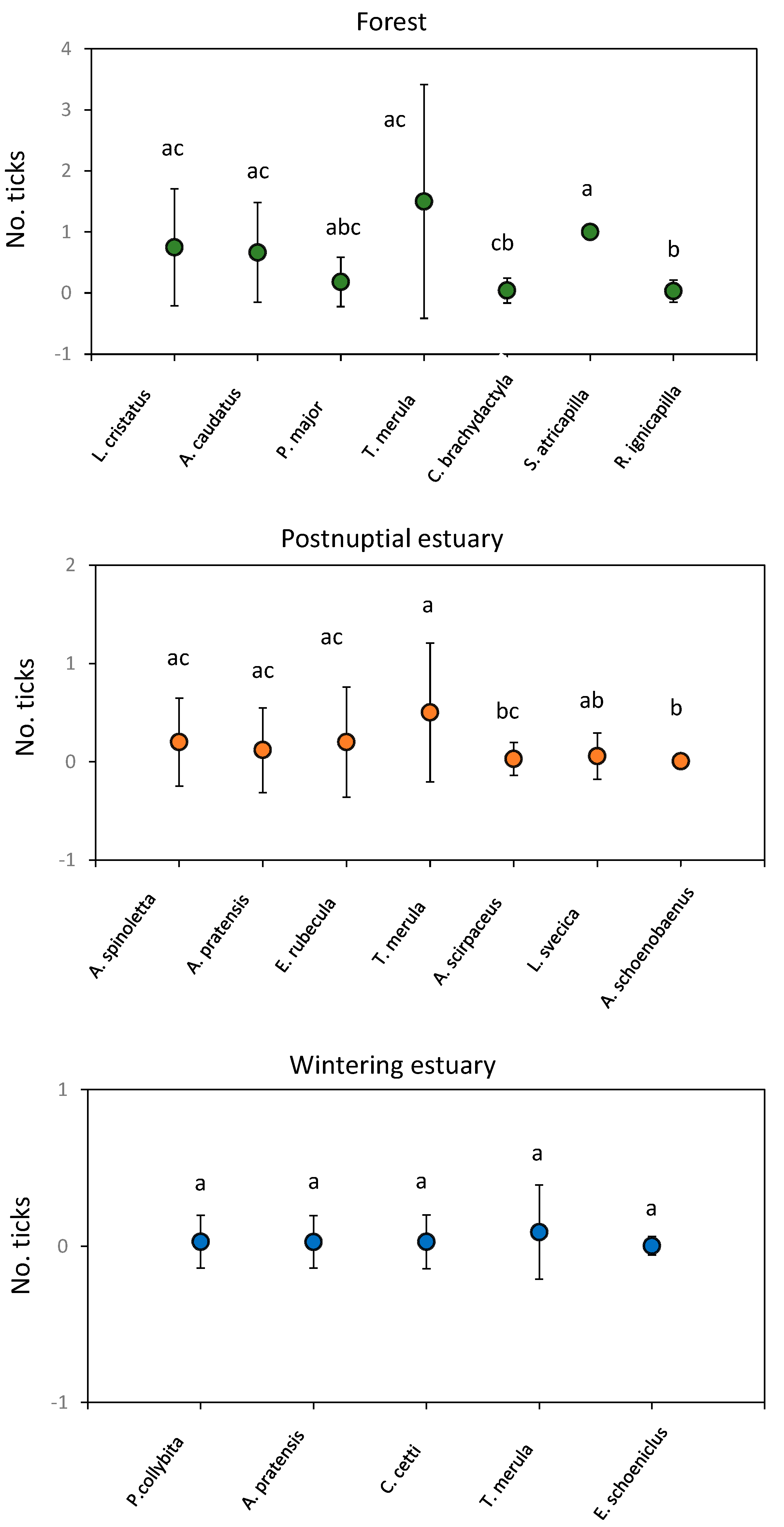
Figure 9.
Number of ticks (average ± SD) in birds for each migratory behaviour captured in postnuptial estuary. Significant differences in tick number between different migratory behaviour are indicated by different letters.
Figure 9.
Number of ticks (average ± SD) in birds for each migratory behaviour captured in postnuptial estuary. Significant differences in tick number between different migratory behaviour are indicated by different letters.
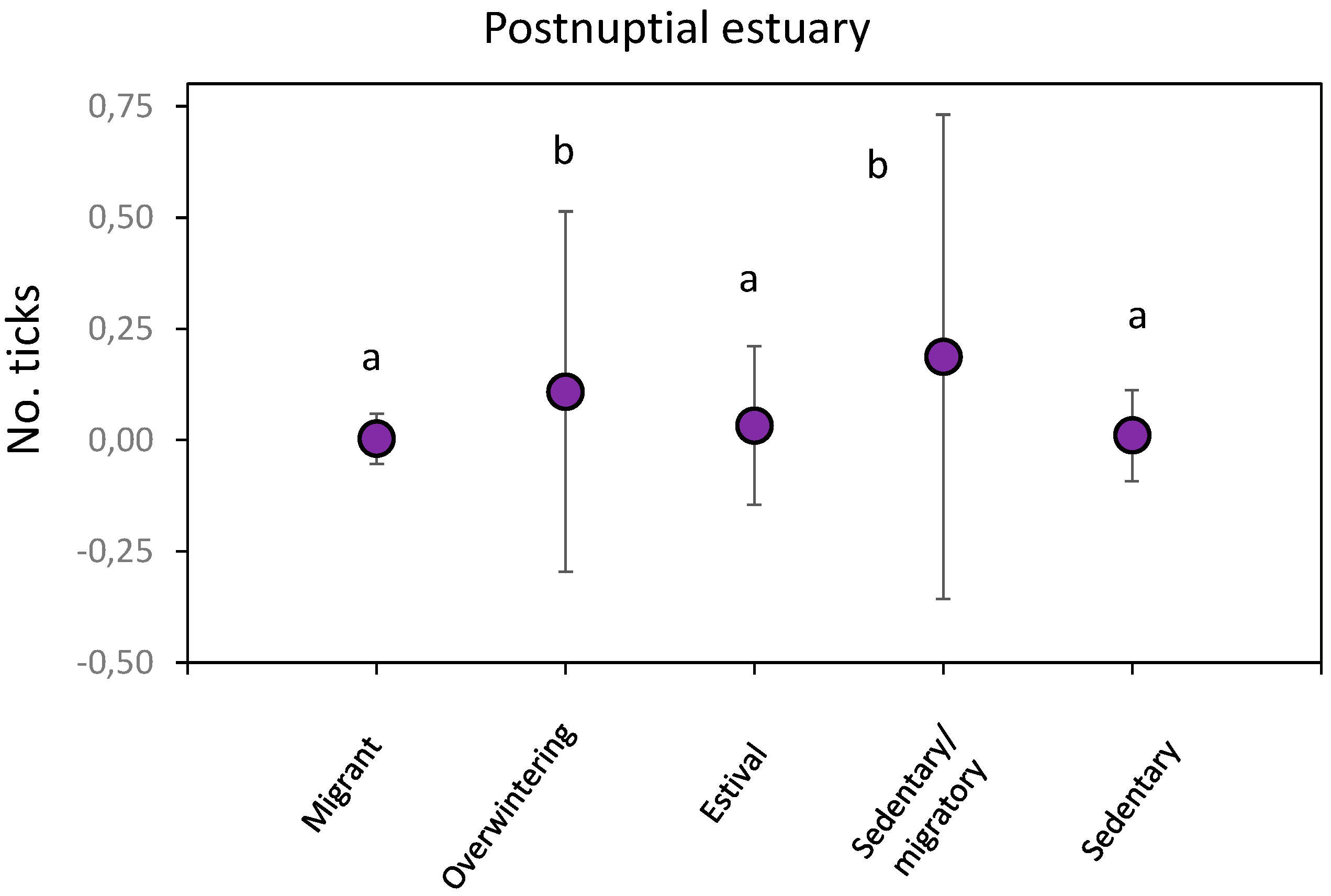

Table 1.
Birds species captured and infested in Asturias (2021-2023). Birds behaviour and tick species.
Table 1.
Birds species captured and infested in Asturias (2021-2023). Birds behaviour and tick species.
| Bird family |
Bird species |
MB | FB | C. bird No. |
Tick No. |
BI % |
Tick stage | Tick species | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ad | Nf | Lv | I.r | I. f | I. | H.c | |||||||
| Acrocephalidae | A. paludicola | m | g | 4 | 0 | 0.0 | |||||||
| A. schoenobaenus | m | g | 233 | 1 | 0.4 | 1 | 1 | ||||||
| A. scirpaceus | m | m | 107 | 3 | 2.8 | 3 | 1 | 2 | |||||
| Aegithalidae | Aegithalos caudatus | s | m | 7 | 3 | 42.9 | 2 | 1 | 3 | ||||
| Alaudidae | Alauda arvensis | m(s) | g | 1 | 0 | 0.0 | |||||||
| Alcedinidae | Alcedo atthis | s | - | 1 | 0 | 0.0 | |||||||
| Certhiidae | Certhia brachydactyla | s | m | 25 | 1 | 4.0 | 1 | 1 | |||||
| Cettiidae | Cettia cetti | s | g | 40 | 1 | 2.5 | 1 | 1 | |||||
| Cisticolidae | Cisticola juncidis | s | m | 39 | 0 | 0.0 | |||||||
| Corvidae | Garrulus glandarius | s | g | 2 | 0 | 0.0 | |||||||
| Emberizidae | Emberiza schoeniclus | m (h) | g | 294 | 1 | 0.3 | 1 | 1 | |||||
| Emberiza pusilla | m (h) | m | 1 | 0 | 0.0 | ||||||||
| Fringillidae | Pyrrhula pyrrhula | s | m | 3 | 0 | 0.0 | |||||||
| Fringilla coelebs | s/m(h) | l | 14 | 0 | 0.0 | ||||||||
| Chloris Chloris | s/m(h) | l | 7 | 0 | 0.0 | ||||||||
| Carduelis carduelis | s | g | 6 | 0 | 0.0 | ||||||||
| Serinus serinus | s | h | 1 | 0 | 0.0 | ||||||||
| Hirundinidae | Hirundo rustica | m | h | 12 | 0 | 0,0 | |||||||
| Laniidae | Lanius collurio | m(s) | g | 4 | 0 | 0,0 | |||||||
| Locustellidae | Locustella naevia | m(s) | g | 4 | 0 | 0.0 | |||||||
| Motacillidae | Anthus spinoletta | m(h) | g | 29 | 1 | 3.4 | 1 | 1 | |||||
| Motacilla flava | m(s) | g | 4 | 1 | 25.0 | 1 | 1 | ||||||
| Anthus petrosus | m(h) | g | 1 | 0 | 0.0 | ||||||||
| Anthus pratensis | m(h) | g | 146 | 9 | 6.2 | 2 | 9 | 7 | 4 | ||||
| Anthus trivialis | m(s) | m | 1 | 0 | 0.0 | ||||||||
| Motacilla alba | s/m(h) | g | 2 | 0 | 0.0 | ||||||||
| Muscicapidae | Luscinia svecica | m(s) | g | 39 | 2 | 5.1 | 2 | 2 | |||||
| Erithacus rubecula | s/m(h) | g | 52 | 2 | 3.8 | 1 | 2 | 1 | 2 | ||||
| Saxicola rubicola | s | g | 46 | 0 | 0.0 | ||||||||
| Saxicola rubetra | m | g | 20 | 0 | 0.0 | ||||||||
| Paridae | Cyanistes caeruleus | s | h | 15 | 0 | 0.0 | |||||||
| Lophophanes cristatus | s | h | 5 | 2 | 40.0 | 1 | 2 | 3 | |||||
| Parus major | s | m | 16 | 2 | 12.5 | 2 | 2 | ||||||
| Poecile palustris | s | h | 23 | 0 | 0.0 | ||||||||
| Periparus ater | s | h | 15 | 0 | 0.0 | ||||||||
| Passeridae | Passer domesticus | s | g | 8 | 0 | 0.0 | |||||||
| Phylloscopidae | Phylloscopus collybita | m(h) | h | 283 | 8 | 2.8 | 1 | 1 | 6 | 2 | 6 | ||
| Phylloscopus ibericus | m(s) | h | 3 | 0 | 0.0 | ||||||||
| Phylloscopus trochilus | m | h | 31 | 0 | 0.0 | ||||||||
| Picidae | Dendrocopos major | s | h | 2 | 0 | 0.0 | |||||||
| Jynx torquilla | m | g | 2 | 0 | 0.0 | ||||||||
| Prunellidae | Prunella modularis | s | g | 10 | 0 | 0.0 | |||||||
| Regulidae | Regulus ignicapilla | s | h | 31 | 1 | 3.2 | 1 | 1 | |||||
| Sittidae | Sitta europaea | s | h | 3 | 0 | 0.0 | |||||||
| Sturnidae | Sturnus unicolor | s | g | 1 | 0 | 0.0 | |||||||
| Sylviidae | Sylvia communis | m | m | 12 | 0 | 0.0 | |||||||
| Sylvia atricapilla | s/m(h) | m | 27 | 1 | 3.7 | 1 | 1 | ||||||
| Sylvia melanocephala | s | m | 10 | 0 | 0.0 | ||||||||
| Troglodytidae | Troglodytes troglodytes | s | m | 16 | 0 | 0.0 | |||||||
| Turdidae | Turdus merula | s | g | 17 | 4 | 23.5 | 1 | 7 | 6 | 1 | 1 | ||
| Turdus iliacus | m(h) | g | 2 | 0 | 0.0 | ||||||||
| Turdus philomelos | s | g | 21 | 0 | 0.0 | ||||||||
| 26 families | 52 species | Total | 1,698 | 43 | 2.53 | 3 | 20 | 28 | 31 | 18 | 1 | 1 | |
* MB-migratory behaviour (m-migratory, s-sedentary, s/m-sedentary and migratory, (h)-hibernating, (s)-summer), FB-feeding behaviour (g-ground, m-medium, h-high), not included in the classification the common kingfisher (Alcedo atthis) that hunts mainly fish from a perch, C.bird-number of captured birds of particular species, BI- prevalence of tick infestation among particular bird species, tick stages (Ad-adult, N-nymph, L-larvae), tick species (I.r-I. ricinus, I.f-I. frontalis, I-Ixodes spp., H.c-H. concinna).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



