Submitted:
10 October 2023
Posted:
11 October 2023
You are already at the latest version
Abstract
Supplements in fish nutrition can help alleviate pressure on the wild stock; however, there is limited information on the benefits of rooibos tea extract in aquafeed. This study aimed to determine the effect of rooibos tea extract inclusion in the commercial diet on the growth performance of larval fish. Fish were reared on three diets: a commercial diet with 30% fermented or green rooibos extract and a commercial tilapia diet (control). Fish (initial weight of 0.54 ± 0.008 g and length of 2.22 ± 0.01 cm) were randomly distributed into recirculating aquaculture systems in triplicates at 100 fish per tank. Fish was hand-fed thrice daily for eight weeks at 2% body weight. After that, samples were withdrawn to measure growth parameters. Fish-fed fermented, and green tea extracts exhibited a significantly (p ˂ 0.05) higher weight gain than the controls. The feed conversion ratio was higher in the control group, fermented (1.50 ± 0.25 cm) and green tea (1.41 ± 0.07 cm). Similarly, the control group exhibited the highest condition factor (K=13.14 ± 4.87). The two treatment groups were comparable (p >0.05) in growth performance. Overall, the evidence indicates that adding rooibos tea extracts to the basal diet can improve fish growth.
Keywords:
Aspalathus linearis
; Oreochromis mossambicus
; feed supplements
; growth performance
; survival rate
1. Introduction
Tilapia, mainly endemic to the African continent, is one of the most popular cultured species in the global aquaculture industry, according to the United Nations Food and Agriculture Organization [1]. Recently, world production of both farmed and wild tilapia has increased by 7.6% annually, from 900,000 tonnes in 1990 to nearly 7 million tonnes in 2018 [2], and aquaculture has been identified as a significant driving force behind the impressive growth. This growth further consolidated the position of tilapia as the second-most important farmed finfish species group. The estimated value of farmed tilapia reached 11.2 billion US dollars, accounting for 4.5% of the total value of all farmed aquatic animals, with a 2.8% increase in 1998, but lower than the peak increase of 5.2% achieved in 2012 [3,4]. Nevertheless, tilapia production is highly imbalanced [5], with China being the largest tilapia farming country, producing over 30% of the global output. In China, for example, while it is common for a fish farmer to harvest 15 tonnes/ha per crop through intensive pond culture, Africa produces less than 5 tonnes/ha per crop [6]. As the global demand for fish products surges from 154 million tonnes in 2011 to the projected 186 million tonnes by 2030 [7], it is apparent that the quality of aquafeeds used in fish farming will be a crucial driver in the blue economy. Unfortunately, many subsistent fish farmers in low and medium-income countries cannot afford many of the high-quality aquafeeds because of their huge cost of production and purchase. Thus, there is a need to seek cost-effective, high-quality aquafeeds to meet the increasing demand. It is well established that global fisheries’ landing has remained constant at about 90 million tonnes per annum, whereas conventional aquaculture production has increased and supplied approximately 78 million tonnes [8]. On the other hand, consumers’ demand for fish products is growing by more than 2.5% per year, while wild fish stocks are declining drastically . According to the FAO, over 40% of global fish stocks are fully exploited, and an additional 18% are over-exploited, with no possibility for further expansion [9] while, food and nutrition security remains a global challenge in many low- and medium-income countries, as aquaculture's importance as a source of protein continues to grow [10]. Therefore, reliance on wild fishery resources is not feasible soon [11]. Accordingly, the global shift to intensive fish farming has increased demand for nutritionally complete feed to promote optimal growth and expand production capacity with minimal mortality or reduced susceptibility to infectious diseases [12]. This trend has intensified the application of various growth promoters and hormones to enhance feed conversion efficiency and productivity [13]. Some of these interventions have negative implications for human and environmental health.
The shortage of wild fish and the increasing demand for farmed fish have necessitated research interest in quality and safe aquafeed devoid of pollutants and antibiotics [14]. The aquafeed industry has turned to plant-based ingredients due to their availability, low costs, evidence-based ethnomedicinal properties and local use as ingredients in livestock feed. Therefore, in the recent past, aquaculture nutrition research has focused on developing feed substitution strategies such as the use of components and supplements of plant origin [15,16,17,18,19,20,21]. Besides, plants and their extracts have been used in animal nutrition as appetizers, flavourants, digestive and physiological stimulants, and antioxidants to prevent or treat specific pathological conditions and promote growth [22]. The benefits of applying phytochemicals and herbal products to enhance productivity in the aquaculture industry are well established [23,24,25,26]. Phytochemicals are commonly found in fruits, vegetables, and plant-based beverages such as wine and tea [27].
Tea, Camellia sinensis, considered native to South China [28], is one of the most consumed beverages worldwide. The main constituents in tea leaves are catechins, normally found in unfermented green tea [29], while theaflavins and thearubigins are the major polyphenolic compounds in black tea [30]. Rooibos is one of the most popular herbal teas in South Africa [31]. It is prepared from a leguminous shrub, Aspalathus linearis, an indigenous herbal plant well known for its many aesthetic health benefits associated with the antioxidant properties of tea polyphenols [32,33,34]. According to the various manufacturing processes and properties such as colour, components, and taste, rooibos tea is branded as fermented (FRT) or unfermented (green) tea (GRT) [35]. Some of the class of compounds in FRT are tannins, flavonoids (aspalathin and aspalalinin), and phenolic acids [36] and this study). Aspalathin is the main flavonoid of GRT and is one of the main constituents of FRT water extract [37,38]. Polyphenols are well known for their antioxidative action and regulation of several protein functions, including the prevention of oxidative stress-related diseases in animals and humans [39,40,41,42,43]. Due to the high polyphenolic contents of rooibos extracts, they can scavenge free radicals and prevent cellular oxidative damage [34] (Joubert et al., 2008). However, the health benefits of rooibos tea extracts have never been evaluated in fish growth studies. Therefore, the current study assessed the effects of FRT and GRT tea extracts as feed potential supplements in O. mossambicus production.
2. Materials and Methods
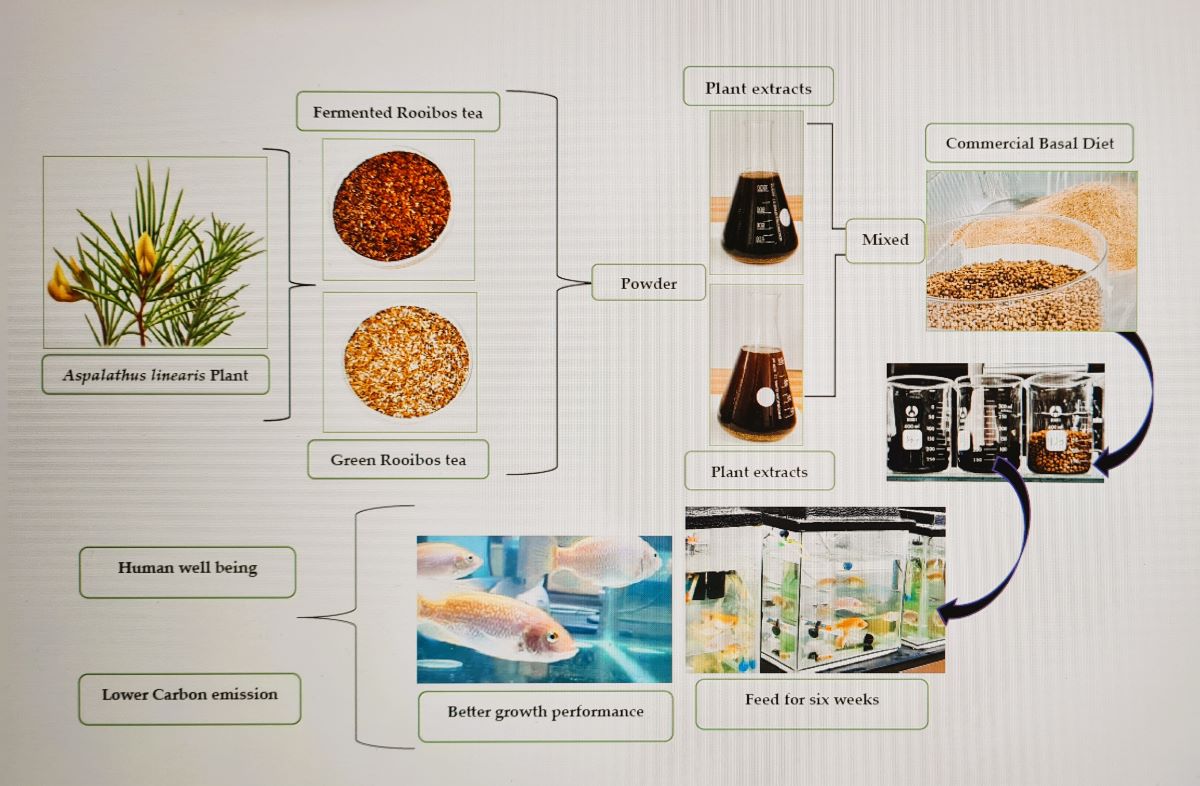
2.1. Commercial Diet and Tea Composition
The ingredients and chemical composition of the commercial feed and the teas (fermented and green rooibos tea) are shown in Table 1. The Commercial feed was sourced from a commercial feed company (Avi Feed Co., South Africa). Green (GRT) and fermented (FRT) rooibos tea were purchased from Rooibos Ltd. (Clanwilliam, South Africa).
2.2. General Experimental Procedures
All experiments were performed at the freshwater biology laboratory of the Department of Biological and Environmental Sciences, Walter Sisulu University (WSU). O. mossambicus larvae were sourced from a registered supplier (Aquaculture Innovations, Grahamstown, South Africa). Larval fish were acclimatised for two weeks before the onset of the experiments and fed a commercial tilapia diet twice daily. Before feeding trials, fish were starved for 24 h, size-sorted by hand, and randomly stocked into each recirculating aquaculture system (RAS). The RAS consisted of a PVC circular cage connected to a biological filter, a water pump, a UV steriliser, and an electric water heater. Tanks were filled with de-chlorinated tap water two weeks before fish stocking. The water flow rate into each tank was 600 L/h. Supplemental aeration was provided in each tank for optimal oxygen dissolution using a regenerative blower and a submerged air diffuser. The water temperature was maintained at 28 ± 1 °C. Temperature and dissolved oxygen readings were recorded daily using a handheld Hanna portable Model HI198198 dissolved oxygen (DO) meter (Hanna Instruments, South Africa). Total ammonia nitrogen (TAN), nitrite nitrogen, and pH were measured weekly using a Freshwater Aquaculture Test Kit (Hanna Instruments, South Africa). The photoperiod was maintained at 14:10 h (light: dark), and water in each system topped up in each system weekly. Fish were cultured on three diets: a control, basal commercial tilapia diet (Avi Feed Co., South Africa) - (CBD) with 0.0% (w/v) rooibos tea extract inclusion, a basal diet with 30% (w/v) fermented rooibos tea (FRT) extract inclusion and basal diet with 30% (w/v) green rooibos tea (GRT) extract inclusion. A total of nine hundred fish (initial weight of (0.54 ± 0.08 g) and length (2.22 ± 0.01 cm) were randomly distributed into the RAS in triplicates at hundred fish per tank. Each diet was assigned to triplicate tanks of fish and fish hand-fed to satiation three times daily for eight weeks at 2% body weight. At the end of eight weeks, samples were withdrawn to measure different growth parameters.
2.3. Preparation of Experimental Feed
Aqueous extracts of fermented (FRT) and green rooibos tea (GRT) were separately prepared by adding 300 g of plant material into a clean glass beaker with 1 L of deionised water and brewed for 30 min. The tea extracts were allowed to cool at room temperature (≈25 ⁰C) before filtering. The tea extracts were then refrigerated at 4 ⁰C until further use. The control diet was prepared by mixing dry pellets of tilapia feed (7.5 g) with 30 mL of deionised water. Similarly, equal amounts of pellet feed were mixed with either 30 mL each of FRT and GRT aqueous tea extracts to form a ball-shaped dough. These three diets were kept in labelled containers and stored at -20 ⁰C until feeding.
2.4. General Experimental Procedures
All experiments were performed at the freshwater biology laboratory of the Department of Biological and Environmental Sciences, Walter Sisulu University (WSU). O. mossambicus larvae were sourced from a registered supplier (Aquaculture Innovations, Grahamstown, South Africa). Larval fish were acclimatised for two weeks before the onset of the experiments and fed a commercial tilapia diet twice daily. Before feeding trials, fish were starved for 24 h, size-sorted by hand, and randomly stocked into each recirculating aquaculture system (RAS). The RAS consisted of a PVC circular cage connected to a biological filter, a water pump, a UV steriliser, and an electric water heater. Tanks were filled with de-chlorinated tap water two weeks before fish stocking. The water flow rate into each tank was 600 L/h. Supplemental aeration was provided in each tank for optimal oxygen dissolution using a regenerative blower and a submerged air diffuser. The water temperature was maintained at 28 ± 1 °C. Temperature and dissolved oxygen readings were recorded daily using a handheld Hanna portable Model HI198198 dissolved oxygen (DO) meter (Hanna Instruments, South Africa). Total ammonia nitrogen (TAN), nitrite nitrogen, and pH were measured weekly using a Freshwater Aquaculture Test Kit (Hanna Instruments, South Africa). Photoperiod was maintained at 14:10 h (light: dark), and water in each system topped up in each system weekly.
Fish were cultured on three diets: a control, basal commercial tilapia diet (Avi Feed Co., South Africa) - (CBD) with 0.0% (w/v) rooibos tea extract inclusion, a basal diet with 30% (w/v) fermented rooibos tea (FRT) extract inclusion and basal diet with 30% (w/v) green rooibos tea (GRT) extract inclusion. A total of nine hundred fish (initial weight of (0.54 ± 0.08 g) and length (2.22 ± 0.01 cm) were randomly distributed into the RAS in triplicates at hundred fish per tank. Each diet was assigned to triplicate tanks of fish and fish hand-fed to satiation three times daily for eight weeks at 2% body weight. At the end of eight weeks, samples were withdrawn to measure different growth parameters.
2.5. Measurement of Growth Performance Parameters
The initial body weight (IBW) (g), initial body length (IBL) (cm), final body weight (FBW) (g), and final body length (FBL) (cm), were measured at the beginning and end of the feeding trial. Specific growth rate (SGR) was calculated using equation 1:
SGR (%)=(100 ×(FBW – IBW))/(time (d))
The rate of weight gain (WGR) was calculated using equation 2:
WGR=100%× ((FBW – IBW))/IBW
The condition factor (CF) was calculated using Equation 3:
CF=100%× FBW/FBL
The survival rate (SR) was calculated using Equation 4:
SR=100%× NF/NI
NF is the number of fish at the end of the feeding trial and NI is the number of fish at the beginning of the feeding trial.
2.6. Tissue fixation and processing
The method of Okuthe (2013) was adopted. Briefly, liver and spleen samples were fixed in Histochoice fixative (Sigma-Aldrich, St. Louis, MO, USA) overnight at room temperature, rinsed in running tap water, and then in 70 % ethanol. Tissues were preserved in 70 % ethanol until further processing. Fixed tissues sufficient for analysis were dehydrated through an ethanol series and cleared in a xylene solution. Tissues were embedded in paraffin wax (Paraplast®, Merck, Darmstadt Germany) at approximately 58 ⁰C in the embedding machine (Thermo Scientific Microm EC 350). Sections were manually cut at 5 - 7 μm thickness, using Leica manual microtome (Leica RM2235: Leica Biosystems), floated in a water bath at 37 ⁰C, and then positioned on Poly-Prep glass slides (Sigma-Aldrich, St. Louis, MO, USA). Tissue sections were dewaxed in xylene, hydrated in ethanol series (Okuthe, 2016), stained with Harrison’s modified haematoxylin, counterstained with eosin, and mounted in DPX. Images of stained tissues were examined and captured using a Leica DM 750 microscope fluorescent (Leica Microsystems, GmbH, Germany), attached to a DFX 310 FX digital camera, and analyzed using Leica LAS imaging software version 4.5.
2.7. The micronucleus assay (MN)
Blood samples were collected from sampled fish by cardiac puncture, with a heparinized syringe (needle 0.5 mm thick) for micronucleus analysis. Blood drops from each fish were placed on a Poly-Prep glass slide (Sigma-Aldrich, St. Louis, MO, USA) with the use of a pipette. Glass coverslips were used to spread blood onto slides. Slides were air-dried for 2 h at room temperature and then fixed in methanol for 10 min. For each experimental group, ten slides were prepared. Five slides from each treatment group were stained with a fluorescent dye, acridine orange (AO) to provide information about the physiologic or pathologic state of fish during the feeding trial. Images of the stained material were viewed and captured using a Leica DM 750 microscope fluorescent microscope (Leica Microsystems, GmbH, Germany. Units similar and next to the main nucleus were interpreted as micronuclei (Fig. 3), (Minissi et al., 1996).
2.8. Statistical analysis
2.8.1. Growth Performance Parameters
Statistical Package for Social Sciences (IBM, SPSS, V. 5) was used to analyse fish growth data, and one-way analysis of variance (ANOVA, Table 5) was performed to test the differences between growth parameter means of different treatments. Duncan’s post hoc test for multiple comparisons was performed to determine significant differences among groups. A significant difference was considered when P ≤ 0.05.
2.8.2. Plant Metabolite Analysis
The concentration (mg/L) was measured for each of the five herbal samples (clients) for each of the compounds (IDs). There were three replicates for each measurement. The hypothesis that the vectors of the client means were all equal was to be tested. The multivariate permutations test (PERMANOVA) was performed by using the Adonis function in the vegan package of R.
3. Results
3.1. Analysis of plant metabolites
3.1.1. Sugars in GRT and FRT
Rooibos tea extracts contain three significant groups of sugar: the monosaccharides (D-glucose and D-fructose), the disaccharides (sucrose and α-lactose), and a trisaccharide (raffinose). The FRT and GRT extracts have the same fruit sugar concentration (sucrose = 90 mg/mL). It is worth mentioning that the FRT extract had a higher glucose concentration (≈78 mg/mL) when compared to the GRT, with 40 mg/mL. On the other hand, the α-lactose concentration was lower in the FRT extract (8 mg/mL) compared to the GRT extract, which gave 28 mg/mL (Figure 1).
Both FRT and GRT extracts have 4-hydroxybenzoic acid, vanillic acid, protocatechuic acid, p-coumaric acid, syringic acid, ferulic acid and caffeic acid. The mean concentrations of phenolic acid profiles are shown in Table 1. The results of a PERMANOVA test show that FRT mean concentrations were much greater than the corresponding GRT ones. This also applies to mean concentrations of protocatechuic acid. The molecular structures of the identified sugar and phenolic compounds in the FRT and GRT extracts are shown in Figure 2.
Experimental fish groups fed a diet containing rooibos tea extracts (FRT and GRE) showed an improvement in the growth indices compared to the group fed with the CBD but with a significant increase in the Viscerosomatic Index (VSI). Fish groups fed GRT extracts had the highest VSI compared to other test groups. Weight gain and the specific growth rate of fish fed with FRT and GRT were also higher than the control (those fed the CBD), showing the role natural plant products (metabolites) played in fish growth. The feed conversion ratio (FRC) was higher in fish fed the BCD (2.32 ± 0.57), followed by FRT (1.50 ± 0.25) and GRT (1.41 ± 0.07). Similarly, fish-fed BCD had higher Condition factor (CF): CBD (13.14±4.87), GRT (6.70 ± 2.53), and FRT (6.29 ± 2.45). A higher feed conversion ratio and condition factor in the control than FRT and GRT reaffirms the aesthetic value of rooibos tea metabolites. There was no significant difference between the FRT and GRT treatment groups (ANOVA, P ˂ 0.05) Table 3.
Values mean ± SD of three replicates. Values with the different letters in the same row are significantly different (n = 15). (a, b & c) in a row indicates, that similar superscripts do not differ (P>0.05), and different superscripts indicate a significant difference (P<0.05). *FRT– Fermented Rooibos tea-supplemented diet. *GRT – Green Rooibos tea-supplemented diet. Basal diet – non-supplemented diet
3.2.2. Liver and spleen histopathology
The potential effects of rooibos tea extracts on the liver and splenic functions were evaluated. The spleen of fish-fed CBD showed a higher degree of loosely dispersed macrophages, specifically in the white pulp, with a reduced number of blood cells (Fig. 3A). At low magnification, the boundary between the white and red pulp was blurry, the red pulp being more diffuse. The FRT group exhibited a balance of red and white pulp and few blood cells, with the moderate occurrence of scattered macrophages mainly restricted to the white pulp (Fig 3B). The GRT group indicated fewer macrophages within the diffuse white pulp and a prominent red pulp surrounding the white pulp (Figure 3D).
The spleen was full of red blood cells and had an apparent boundary between the white and the red pulp. Normal polygonal hepatocytes with normal blood vessels were observed in fish-fed CBD with a moderate degree of cytoplasmic vacuolisation, and hepatocytes showed clear boundaries (Fig. 3D). Fat vacuoles and large hepatocytes with displaced nuclei characterised the liver of fish fed FRT (E) and GRT extracts (F) (Fig. 3 E and F respectively).
3.2.3. The Micronucleus (MN) Assay
The mutagenic potential of the tea extracts was examined using the MN assay in blood samples to evaluate different chromosomal alterations in blood cells. No signs of toxicity, such as loss of appetite, were noted during the feeding trial in all groups. Contrary to the expected results, MN were seen in the control group blood samples and rarely in experimental groups (Figure 4), affirming the absence of a cytotoxic effects diet. Supplementing the commercial basal diet (CBD) with rooibos tea extracts did not enhance the frequencies of MN frequencies inferring the non-toxicity of the tea extracts. There were no differences between the GRT and FRT treatment groups. These effects might be related to the natural compounds present in the extracts, which did not interfere with the DNA synthesis process.
4. Discussion
Feed is one of the foremost expenses of intensive fish farming, and its availability in quality and quantity is significant for the appropriate growth and reproduction of fish [39]. It is, unfortunately, one of the limiting factors in aquaculture production in low and middle-income countries. Similarly, diet supplementation is a major consideration in intensive aquaculture during fish culture, specifically during the early stages of growth [40]. Thus, the application of functional feed supplements of plant origin, which provide safer and sustainable alternatives to antibiotics, is highly recommended to increase cultured fish's growth performance and fish health status and for food security.
4.1. Larval fish growth response to FRT- and GRT extract-supplemented diets
It is well known that fish modifies feed intake to satisfy their energy demand [41,42]. Thus, an elevation in feed intake usually occurs to meet nutrient demand in deficient diets. In this present study, the effects of fermented (FRT) and green rooibos tea (GRT) extracts inclusion in the basal diet (CBD) on the growth and survival of larval O. mossambicus were evaluated. The study revealed that FRT and GRT extract inclusion in the CBD improved feed utilisation and growth in larval fish, which may indicate efficient feed utilisation, resulting in an increased organo-somatic index (VSI%). The GRT-supplemented diet seemed to be more suitable for growth and FCR than that of the FRT. Thus, adding rooibos tea extracts, specifically GRT, to the CBD may have influenced feeding behaviour in the larval fish to favoured growth performance.
Tilapia fish generally accepts a wide variety of plant diets. Therefore, adding GRT and FRT extracts as supplements to the basal diet might have been useful, as reported in feeding trials with other fish species [43]. In similar studies, positive effects of green tea (Camellia sinensis L) have been demonstrated, wherein dietary green tea's negative effects of dietary oxidised fish oil were reversed. Green tea also improved growth performance, lipid metabolism, and antioxidant defense in the sturgeon hybrid of Sterlet [44]. Similarly, positive effects of green tea on the antioxidant defense and immune parameters of rainbow trout, Oncorhynchus mykiss at the molecular level have been reported [45], while its ability to positively modulate myogenic regulatory factors and stress-related genes in juvenile grass carp (Ctenopharyngodon idellus) has also been reported [46].
As with other tea types, rooibos' potential health benefits and bioactivity have been linked to its naturally occurring polyphenolic contents [32,47]. The high polyphenolic contents of rooibos extracts confer considerable antioxidant properties, with scavenging effects on free radicals, which may prevent cellular oxidative damage [35]. Two classes of phenolic acids, hydroxybenzoic and hydroxycinnamic acids, were found in the two rooibos tea extracts analysed. Hydroxybenzoic acids (C6-C1) included p-hydroxybenzoic, vanillic, syringic, and protocatechuic acids. The hydroxycinnamic acids, known as phenylpropanoids [48], (C6-C3), included p-coumaric, caffeic, and ferulic acids. Protocatechuic acid (PCA) was the most abundant in GRT, followed by ferulic, vanillic, and syringic acids. PCA is a major metabolite of complex polyphenols and has been effective in several physiological functions due to its anti-inflammatory properties [49,50]. Also, the addition of PCA was reported to be effective in improving the growth of freshwater algae, Euglena gracilaris at 800g/L [51], while it also improved meat quality and body weight in broilers [52]. The authors argue that applying PCA may induce an active cellular immune response, resulting in good health and higher body weight, which agrees with the current study's findings. Therefore, the high content of PCA in GRT extracts explains the growth performance and increased body weight seen in fish fed GRT extracts compared to other treatment groups. The major phenolic acids in the FRT extract were ferulic, syringic, protocatechuic, and vanillic. Ferulic acid (FA), like PCA, is a secondary metabolite which can enhance growth performance, carcass characteristics, and meat quality in ruminants by reducing lipid peroxidation and increasing the average daily gain and carcass weight [53].
Overall, phenolic acids are generally regarded as strong natural antioxidants, having key roles in various biological and pharmacological properties, such as anti-
inflammatory, antimicrobial, antiviral, food additive, signalling molecules, and many more [52,54]. The benefits of these naturally occurring organic acids in the dietary supplements of farm animals have been well documented. In farm animal production, for example, phenolic acids act as growth promotors by stimulating digestive enzyme secretions, thus decreasing pathogenic bacteria in the gastrointestinal tract (GIT) or modulating gut morphology [54]. Studies in farm animals have also shown improved FCR in poultry feed supplemented with natural extracts containing phenolic compounds, wherein improvement in FCR values was associated with the alteration of intestinal surface area, resulting in better nutrient absorption [55,56]. Furthermore, phenolic compounds are known to balance the useful and pathogenic bacteria in the GIT, which helps maintain sound gut health and ultimately enhances growth [52]. They may also improve the flavour and palatability of feed, thus promoting feed intake and growth performance [57]. However, aquafeed rarely exploits phenolic acid-rich natural extracts for the same purpose [54]. Phenolics are associated with high growth rates, probably attributed to their potential antioxidants and ability to avert cell damage from free-radical oxidation reactions. Thus, our study findings have provided insight into the usefulness of natural products, such as the phenolic acid-rich rooibos extracts, as a potential aquafeed supplement.
Although the effects of FRT and GRT extracts on gut health were not evaluated in the current study, it may be deduced that the presence of a variety of phenolic compounds in rooibos tea extracts may have improved flavour and palatability of feed leading to better growth performance. Alternatively, a higher growth rate and improved FCR in the GRT treatment group could be due to alterations of the intestinal surface area and digestive enzyme activities, which resulted in better food absorption, increased body weight, and less wastage of feed. Furthermore, the observed variations in the concentrations of glucose and lactose for the two rooibos tea extracts could only have been possible because of the breaking down of α-lactose in the GRT extract upon fermentation process to afford glucose and galactose units [58], thus accounting for the observed increase in the concentration of glucose in the FRT extract and a consequential decrease in its α-lactose concentration. Even though sugars are regarded as non-essential dietary nutrients in aquafeed, they still represent an inexpensive source of valuable dietary energy while also giving integrity to feed pellets in the form of less density, more stability, and optimal binding activity during the feed manufacturing process [59,60].
4.2. Micronucleus (MN) assay
In this study, the potential health benefits of rooibos tea extracts were corroborated by the micronucleus (MN) assay results. The MN assay is a delicate biomarker commonly used to investigate the genotoxicity effects of chemicals isolated or in mixtures present in the aquatic ecosystem. The method validated by the World Health Organization, the United Nations Environmental Program, and the International Program on Chemical Safety (IPCS, OMS) for the evaluation of environmental samples, natural products, including biological agents [61] and reveals chromosome damage or mitotic spindle dysfunction [61,62]. The assay has been demonstrated to be sensitive to various environmental pollutants, such as polycyclic aromatic hydrocarbons (PAHs) [63], and pesticides [64].
In fish, cytogenetic damage of erythrocytes is best indicated by MN assay [65], which can be influenced by diet or water quality [66]. Here, the assay was used to provide information about the physiologic or pathologic state of fish during the feeding trial. It well known that plants tend to synthesize these bioactive polyphenols as a defense mechanism, to protect their vital metabolic functions against free radicals and survive in different highly oxidative environments [67]. The low occurrence of micronuclei counts in treatment group following the acridine orange (AO) staining procedure can be attributed to the free radical scavenging (antioxidant) properties conferred by the phenolic compounds in rooibos tea extracts to prevent cellular oxidative damage [33]. This has been reported in laboratory animals, including humans [68,69,70,71,72].
4.3. Histopathology
In teleost fish, the spleen is a main hematopoietic and peripheral lymphoid organ [67]. It plays a critical role in the storage [73] and production of erythrocytes and the destruction of aged blood cells, endothelial cells, reticulocytes, macrophages, and melanin macrophages [74]. In addition, the teleost spleen contains antibody-producing cells [75,76]. It has a role in antigen presentation and initiation of the adaptive immune response [77,78], and also used as a biomarker for environmental pollution [79]. In the current study, the effects of FRT and GRT on splenic function were not obvious. There were no indications of histopathological lesions or degeneration in tissues. However, our study observed a few scattered splenic macrophage aggregates specifically in the CBD group, while an increase in the number of blood cells in the GRT treatment group may have indicated better blood flow.
The liver, on the other hand, is an accessory digestive organ and a good indicator of the nutritional condition of fish [80,81] and is one of the most important organs for digestion and absorption of nutrients from feed. Monitoring the histological structure of the fish liver is the method of choice in assessing the effects of nutrient mixtures or the use of raw materials of plant origin in feed. New compounds often negatively affect the hepatic function, fitness, health, and production characteristics of cultivated fish [81]. Notwithstanding, commercial feeds cause fat accumulation and degeneration of the liver, cell membrane, and vacuolation of hepatocytes. Such lesions may indicate circulatory anomalies [82,83]. In the present study, fish fed the experimental diets revealed morphological alterations in hepatic tissues, including increased cytoplasm nuclear ratio and changes in the nucleus, cytoplasm, and hepatocyte vacuolisation, especially in the FRT treatment group. Similar nutritional disorders have been previously reported [81], which corroborates the results of the current study, specifically in the FRT and GRT treatment groups, where changes in the tissue parenchyma were prevalent. Nonetheless, hepatic alteration due to excessive caloric ingestion is known to saturate the physiological competency of the liver and may lead to lipid accumulation [84].
Lipid accumulation, distorted nuclei, change in the cell membranes, and mild necrosis in fish liver are the main symptoms of liver toxicity. Liver toxicity due to FRT and GRT extract inclusion could not be determined with certainty in the current study. Nevertheless, cytoplasmic vacuolisation was the most frequent hepatocyte change and could have been caused by excessive caloric ingestion, and vacuolar degeneration due to various factors, making it difficult to establish a threshold of what was healthy and pathological in the liver. To facilitate the transition towards sustainable aquafeeds, we recommend that feeding trials using A. linearis extracts be conducted under appropriate production conditions to determine the optimal levels of inclusion and their effects on fillet quality and flavour. The micronucleus assay is herein reaffirmed as a bioindicator, which describes short-term feeding and environmental insults under laboratory conditions.
5. Conclusions
Our study focused on the effects of rooibos tea extracts (FRT and GRT) on growth performance in larval Oreochromis mossambicus. Rooibos tea extracts may have increased the aroma and palatability of diets, resulting in higher feed intake and body weight. The results support using rooibos tea extracts in larval O. mossambicus rearing. GRT-fed fish experienced significantly higher weight gain and conferred a feasible option of enriching tilapia feed with rooibos (A. linearis) tea extracts as a supplement. The herbal teas may have improved intestinal health and nutrient digestibility, enhanced the growth of beneficial bacteria and overall improved fish growth performance. It is hypothesised that protocatechuic-rich phenolic complexes in GRT extracts increased the aroma and palatability of diets, resulting in higher feed intake and body weight gain for fish. At the same time, their sugar contents may act as an inexpensive, valuable energy source with the potential to improve feed pellet integrity. The two tea brands are locally sourced and may benefit the aquaculture industry, creating avenues for their inclusion in aquafeed. However, further research to elucidate the various governing factors on the effectiveness of dietary inclusion of A. linearis tea extracts in aquafeed and how the functional components of GRT extracts exert their effects in fish is required.
6. Ethics statement
The animal study application was reviewed and approved by experimental procedures that followed all guidelines regarding animal care. The use of animals was approved by Walter Sisulu University’s Senate Research Ethics Committee (# WSU/FNS-AREC- 2021/01-11/A1).
References
- FAO, Cultured aquatic species information programme, Oreochromis niloticus (Linnaeus, 1758). Food and Agriculture Organization of the United Nations, 2010.
- Fisheries, F., et al., FAO Yearbook: FAO Annuaire. Statistiques Des Pêches Et de L'aquaculture. Fishery and aquaculture statistics. 2009: FAO.
- Miao, W. and W. Wang, Trends of aquaculture production and trade: Carp, tilapia, and shrimp. Asian Fisheries Science, 2020. 33(S1): p. 1-10.
- Stankus, A., State of world aquaculture 2020 and regional reviews: FAO webinar series. FAO aquaculture newsletter, 2021(63): p. 17-18.
- Cai, J., et al., Improving the performance of tilapia farming under climate variation: perspective from bioeconomic modelling. FAO Fisheries and Aquaculture Technical Paper, 2018(608): p. I-64.
- FAO, I., The state of food and agriculture 2019. Moving forward on food loss and waste reduction. FAO, Rome, 2019: p. 2-13.
- Kobayashi, M., et al., Fish to 2030: the role and opportunity for aquaculture. Aquaculture economics & management, 2015. 19(3): p. 282-300. [CrossRef]
- Pauly, D. and D. Zeller, Comments on FAOs state of world fisheries and aquaculture (SOFIA 2016). Marine Policy, 2017. 77: p. 176-181. [CrossRef]
- Department, A.O.o.t.U.N.F., The state of world fisheries and aquaculture. 2014: Food and Agriculture Organization of the United Nations.
- Waltz, E., First genetically engineered salmon sold in Canada. Nature, 2017. 548(7666): p. 148-148. [CrossRef]
- New, M.B. and U.N. Wijkström, Use of fishmeal and fish oil in aquafeeds: further thoughts on the fishmeal trap. FAO Fisheries Circular (FAO), 2002.
- Naylor, R.L., et al., A 20-year retrospective review of global aquaculture. Nature, 2021. 591(7851): p. 551-563. [CrossRef]
- Makkar, H., G. Francis, and K. Becker, Bioactivity of phytochemicals in some lesser-known plants and their effects and potential applications in livestock and aquaculture production systems. animal, 2007. 1(9): p. 1371-1391. [CrossRef]
- Okocha, R.C., I.O. Olatoye, and O.B. Adedeji, Food safety impacts of antimicrobial use and their residues in aquaculture. Public health reviews, 2018. 39(1): p. 1-22. [CrossRef]
- Chakraborty, S.B. and C. Hancz, Application of phytochemicals as immunostimulant, antipathogenic and antistress agents in finfish culture. Reviews in Aquaculture, 2011. 3(3): p. 103-119. [CrossRef]
- Citarasu, T., Herbal biomedicines: a new opportunity for aquaculture industry. Aquaculture International, 2010. 18(3): p. 403-414. [CrossRef]
- Faggio, C., et al., Oral administration of Gum Arabic: effects on haematological parameters and oxidative stress markers in Mugil cephalus. 2015.
- Lee, K.W., H.S. Jeong, and S.H. Cho, Dietary inclusion effect of yacon, ginger, and blueberry on growth, body composition, and disease resistance of juvenile black rockfish (Sebastes schlegeli) against Vibrio anguillarum. Fisheries and Aquatic Sciences, 2020. 23: p. 1-8. [CrossRef]
- Sutili, F.J., et al., Plant essential oils as fish diet additives: benefits on fish health and stability in feed. Reviews in Aquaculture, 2018. 10(3): p. 716-726. [CrossRef]
- Wang, J., et al., The effect of partial replacement of fish meal by soy protein concentrate on growth performance, immune responses, gut morphology and intestinal inflammation for juvenile hybrid grouper (Epinephelus fuscoguttatus♀× Epinephelus lanceolatus♂). Fish & shellfish immunology, 2020. 98: p. 619-631.
- Wang, W., et al., Application of immunostimulants in aquaculture: current knowledge and future perspectives. Aquaculture Research, 2017. 48(1): p. 1-23. [CrossRef]
- Wang, Y.-Z., et al., Effects of a natural antioxidant, polyphenol-rich rosemary (Rosmarinus officinalis L.) extract, on lipid stability of plant-derived omega-3 fatty-acid rich oil. LWT, 2018. 89: p. 210-216. [CrossRef]
- Chakraborty, S.B., P. Horn, and C. Hancz, Application of phytochemicals as growth-promoters and endocrine modulators in fish culture. Reviews in Aquaculture, 2014. 6(1): p. 1-19. [CrossRef]
- Rawling, M.D., D.L. Merrifield, and S.J. Davies, Preliminary assessment of dietary supplementation of Sangrovit® on red tilapia (Oreochromis niloticus) growth performance and health. Aquaculture, 2009. 294(1-2): p. 118-122. [CrossRef]
- Shaban, A. and R.P. Sahu, Pumpkin seed oil: an alternative medicine. International journal of pharmacognosy and phytochemical research, 2017. 9(2).
- Shaban, H.H., S.L. Nassef, and G.S. Elhadidy, Utilization of garden cress seeds, flour, and tangerine peel powder to prepare a high-nutrient cake. Egyptian Journal of Agricultural Research, 2023. 101(1): p. 131-142.
- Arts, I.C. and P.C. Hollman, Polyphenols and disease risk in epidemiologic studies. The American journal of clinical nutrition, 2005. 81(1): p. 317S-325S.
- Li, X., et al., The effects of five types of tea solutions on epiboly process, neural and cardiovascular development, and locomotor capacity of zebrafish. Cell biology and toxicology, 2019. 35: p. 205-217. [CrossRef]
- Musial, C., A. Kuban-Jankowska, and M. Gorska-Ponikowska, Beneficial properties of green tea catechins. International journal of molecular sciences, 2020. 21(5): p. 1744. [CrossRef]
- Koch, W., W. Kukula-Koch, and Ł. Komsta, Black tea samples origin discrimination using analytical investigations of secondary metabolites, antiradical scavenging activity and chemometric approach. Molecules, 2018. 23(3): p. 513.
- Chandrasekara, A. and F. Shahidi, Herbal beverages: Bioactive compounds and their role in disease risk reduction-A review. Journal of traditional and complementary medicine, 2018. 8(4): p. 451-458. [CrossRef]
- Joubert, E. and D. de Beer, Rooibos (Aspalathus linearis) beyond the farm gate: From herbal tea to potential phytopharmaceutical. South African Journal of Botany, 2011. 77(4): p. 869-886. [CrossRef]
- Joubert, E., et al., South African herbal teas: Aspalathus linearis, Cyclopia spp. and Athrixia phylicoides—A review. Journal of ethnopharmacology, 2008. 119(3): p. 376-412. [CrossRef]
- Van Wyk, B.-E., B.v. Oudtshoorn, and N. Gericke, Medicinal Plants of South Africa. 1997: Briza.
- Joubert, E., et al., Effect of species variation and processing on phenolic composition and in vitro antioxidant activity of aqueous extracts of Cyclopia spp.(honeybush tea). Journal of Agricultural and Food Chemistry, 2008. 56(3): p. 954-963.
- Shimamura, N., et al., Phytoestrogens from Aspalathus linearis. Biological and Pharmaceutical Bulletin, 2006. 29(6): p. 1271-1274. [CrossRef]
- Bramati, L., et al., Quantitative characterization of flavonoid compounds in rooibos tea (Aspalathus linearis) by LC− UV/DAD. Journal of agricultural and food chemistry, 2002. 50(20): p. 5513-5519. [CrossRef]
- Joubert, E., HPLC quantification of the dihydrochalcones, aspalathin and nothofagin in rooibos tea (Aspalathus linearis) as affected by processing. Food Chemistry, 1996. 55(4): p. 403-411. [CrossRef]
- Volkoff, H., L.J. Hoskins, and S.M. Tuziak, Influence of intrinsic signals and environmental cues on the endocrine control of feeding in fish: potential application in aquaculture. General and Comparative Endocrinology, 2010. 167(3): p. 352-359. [CrossRef]
- Syed, R., et al., Growth performance, haematological assessment and chemical composition of Nile tilapia, Oreochromis niloticus (Linnaeus, 1758) fed different levels of Aloe vera extract as feed additives in a closed aquaculture system. Saudi journal of biological sciences, 2022. 29(1): p. 296-303. [CrossRef]
- Bureau, D.P., J. Gibson, and A. El-Mowafi, Use of animal fats in aquaculture feeds. Avances en Nutrición Acuicola, 2002.
- Vivas, M., et al., Dietary self-selection in sharpsnout seabream (Diplodus puntazzo) fed paired macronutrient feeds and challenged with protein dilution. Aquaculture, 2006. 251(2-4): p. 430-437. [CrossRef]
- Fortes-Silva, R., I.G. Guimaraes, and E.d.F.F. Martíns, Feeding Practices and Their Determinants in Tilapia. Biology and Aquaculture of Tilapia, 2021: p. 86.
- Hasanpour, S., et al., Effects of dietary green tea (Camellia sinensis L.) supplementation on growth performance, lipid metabolism, and antioxidant status in a sturgeon hybrid of Sterlet (Huso huso♂× Acipenser ruthenus♀) fed oxidized fish oil. Fish Physiology and Biochemistry, 2017. 43: p. 1315-1323.
- Nootash, S. Nootash, S., et al., Green tea (Camellia sinensis) administration induces expression of immune relevant genes and biochemical parameters in rainbow trout (Oncorhynchus mykiss). Fish & shellfish immunology, 2013. 35(6): p. 1916-1923. [CrossRef]
- Zhou, J., et al., The effect of green tea waste on growth and health of grass carp (Ctenopharyngodon idellus). Turkish Journal of Fisheries and Aquatic Sciences, 2016. 16(3): p. 679-689.
- McKay, D.L. and J.B. Blumberg, A review of the bioactivity of South African herbal teas: rooibos (Aspalathus linearis) and honeybush (Cyclopia intermedia). Phytotherapy Research: An International Journal Devoted to Pharmacological and Toxicological Evaluation of Natural Product Derivatives, 2007. 21(1): p. 1-16.
- Shahidi, F., Antioxidant properties of food phenolics. Phenolics in food and nutraceuticals, 2004.
- Kay, C.D., P.A. Kroon, and A. Cassidy, The bioactivity of dietary anthocyanins is likely to be mediated by their degradation products. Molecular nutrition & food research, 2009. 53(S1): p. S92-S101. [CrossRef]
- Lin, C.-Y., et al., Antiglycative effects of protocatechuic acid in the kidneys of diabetic mice. Journal of agricultural and food chemistry, 2011. 59(9): p. 5117-5124. [CrossRef]
- Tan, X., J. Zhu, and M. Wakisaka, Effect of protocatechuic acid on Euglena gracilis growth and accumulation of metabolites. Sustainability, 2020. 12(21): p. 9158. [CrossRef]
- Mahfuz, S., Q. Shang, and X. Piao, Phenolic compounds as natural feed additives in poultry and swine diets: A review. Journal of Animal Science and Biotechnology, 2021. 12(1): p. 1-18. [CrossRef]
- Tánori-Lozano, A., et al., Influence of ferulic acid and clinoptilolite supplementation on growth performance, carcass, meat quality, and fatty acid profile of finished lambs. Journal of Animal Science and Technology, 2022. 64(2): p. 274. [CrossRef]
- Kumar, N. and N. Goel, Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnology reports, 2019. 24: p. e00370. [CrossRef]
- Mountzouris, K., et al., Assessment of a phytogenic feed additive effect on broiler growth performance, nutrient digestibility and caecal microflora composition. Animal Feed Science and Technology, 2011. 168(3-4): p. 223-231. [CrossRef]
- Viveros, A., et al., Effects of dietary polyphenol-rich grape products on intestinal microflora and gut morphology in broiler chicks. Poultry science, 2011. 90(3): p. 566-578. [CrossRef]
- Christaki, E., et al., Innovative uses of aromatic plants as natural supplements in nutrition, in Feed additives. 2020, Elsevier. p. 19-34.
- Shakerdi, L.A., et al., Determination of the lactose and galactose content of common foods: Relevance to galactosemia. Food Science & Nutrition, 2022. 10(11): p. 3789-3800. [CrossRef]
- Craig, S.R., et al., Understanding fish nutrition, feeds, and feeding. 2017.
- Tedesco, M., et al., Assessment of the antiproliferative and antigenotoxic activity and phytochemical screening of aqueous extracts of Sambucus australis Cham. & Schltdl.(ADOXACEAE). Anais da Academia Brasileira de Ciências, 2017. 89: p. 2141-2154. [CrossRef]
- Barka, S., et al., Monitoring genotoxicity in freshwater microcrustaceans: A new application of the micronucleus assay. Mutation Research/Genetic Toxicology and Environmental Mutagenesis, 2016. 803: p. 27-33. [CrossRef]
- Polard, T., et al., Giemsa versus acridine orange staining in the fish micronucleus assay and validation for use in water quality monitoring. Ecotoxicology and environmental safety, 2011. 74(1): p. 144-149. [CrossRef]
- Pacheco, M. and M. Santos, Induction of EROD Activity and Genotoxic Effects by Polycyclic Aromatic Hydrocarbons and Resin Acids on the Juvenile Eel (Anguilla anguillaL.). Ecotoxicology and environmental safety, 1997. 38(3): p. 252-259. [CrossRef]
- Grisolia, C.K., A comparison between mouse and fish micronucleus test using cyclophosphamide, mitomycin C and various pesticides. Mutation Research/Genetic Toxicology and Environmental Mutagenesis, 2002. 518(2): p. 145-150. [CrossRef]
- Minissi, S., E. Ciccotti, and M. Rizzoni, Micronucleus test in erythrocytes of Barbus plebejus (Teleostei, Pisces) from two natural environments: a bioassay for the in situ detection of mutagens in freshwater. Mutation Research/Genetic Toxicology, 1996. 367(4): p. 245-251. [CrossRef]
- Araldi, R.P., et al., Using the comet and micronucleus assays for genotoxicity studies: A review. Biomedicine & Pharmacotherapy, 2015. 72: p. 74-82. [CrossRef]
- Zapata, A., et al., Ontogeny of the immune system of fish. Fish & shellfish immunology, 2006. 20(2): p. 126-136. [CrossRef]
- Fraga, C.G., et al., Basic biochemical mechanisms behind the health benefits of polyphenols. Molecular aspects of medicine, 2010. 31(6): p. 435-445.
- Khan, A.-u. and A.H. Gilani, Selective bronchodilatory effect of Rooibos tea (Aspalathus linearis) and its flavonoid, chrysoeriol. European journal of nutrition, 2006. 45: p. 463-469. [CrossRef]
- Marnewick, J.L., et al., Effects of rooibos (Aspalathus linearis) on oxidative stress and biochemical parameters in adults at risk for cardiovascular disease. Journal of ethnopharmacology, 2011. 133(1): p. 46-52. [CrossRef]
- Opuwari, C. and T. Monsees, In vivo effects of A spalathus linearis (rooibos) on male rat reproductive functions. Andrologia, 2014. 46(8): p. 867-877.
- Sanderson, M., et al., Effects of fermented rooibos (Aspalathus linearis) on adipocyte differentiation. Phytomedicine, 2014. 21(2): p. 109-117. [CrossRef]
- Pearson, M. and E. Stevens, Size and hematological impact of the splenic erythrocyte reservoir in rainbow trout, Oncorhynchus mykiss. Fish Physiology and Biochemistry, 1991. 9: p. 39-50. [CrossRef]
- Graf, R. and J. Schlüns, Ultrastructural and histochemical investigation of the terminal capillaries in the spleen of the carp (Cyprinus carpio L.). Cell and Tissue Research, 1979. 196: p. 289-306. [CrossRef]
- Chiller, J., H. Hodgins, and R. Weiser, Antibody response in rainbow trout (Salmo gairdneri) II. Studies on the kinetics of development of antibody-producing cells and on complement and natural hemolysin. The Journal of Immunology, 1969. 102(5): p. 1202-1207.
- Whyte, S.K., The innate immune response of finfish–a review of current knowledge. Fish & shellfish immunology, 2007. 23(6): p. 1127-1151. [CrossRef]
- Espenes, A., et al., Immune-complex trapping in the splenic ellipsoids of rainbow trout (Oncorhynchus mykiss). Cell and tissue research, 1995. 282: p. 41-48. [CrossRef]
- Raida, M.K. and K. Buchmann, Development of adaptive immunity in rainbow trout, Oncorhynchus mykiss (Walbaum) surviving an infection with Yersinia ruckeri. Fish & shellfish immunology, 2008. 25(5): p. 533-541. [CrossRef]
- Torrealba, D., et al., Innate immunity provides biomarkers of health for teleosts exposed to nanoparticles. Frontiers in immunology, 2019. 9: p. 3074. [CrossRef]
- Lozano, A.R., et al., Effect of different dietary vitamin E levels on growth, fish composition, fillet quality and liver histology of meagre (Argyrosomus regius). Aquaculture, 2017. 468: p. 175-183.
- Rašković, B., et al., Histological methods in the assessment of different feed effects on liver and intestine of fish. Journal of Agricultural Sciences (Belgrade), 2011. 56(1): p. 87-100. [CrossRef]
- Bilen, A.M. and S. Bilen, Effect of diet on the fatty acids composition of cultured sea bass (Dicentrarchus labrax) liver tissues and histology compared with wild sea bass caught in Eagean Sea. Marine Science and Technology Bulletin, 2013. 2(1): p. 15-21.
- Coz-Rakovac, R., et al., Blood chemistry and histological properties of wild and cultured sea bass (Dicentrarchus labrax) in the North Adriatic Sea. Veterinary research communications, 2005. 29: p. 677-687. [CrossRef]
- Spisni, E., et al., Hepatic steatosis in artificially fed marine teleosts. Journal of Fish Diseases, 1998. 21(3): p. 177-184.
Figure 1.
Plots of mean sugar concentrations (mg/L) for the herbal compounds.
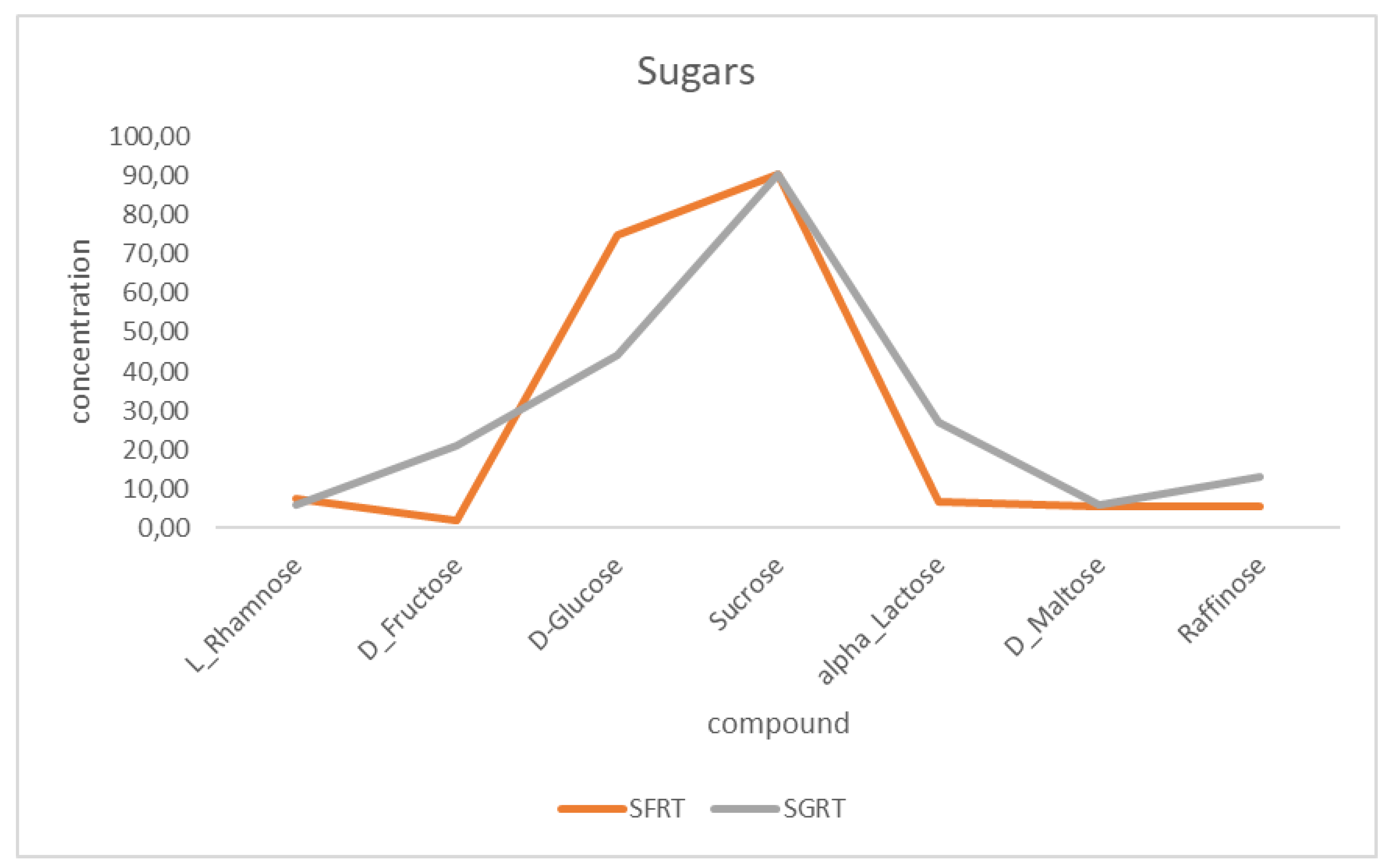
Figure 2.
Chemical structures of the sugar and phenolic compounds identified in the fermented and green rooibos tea extracts, with major functional groups highlighted in red and blue colour. Structures were prepared with ChemDraw Ultra Version 7 (CambridgeSoft Corporation, Cambridge, MA, USA).
Figure 2.
Chemical structures of the sugar and phenolic compounds identified in the fermented and green rooibos tea extracts, with major functional groups highlighted in red and blue colour. Structures were prepared with ChemDraw Ultra Version 7 (CambridgeSoft Corporation, Cambridge, MA, USA).
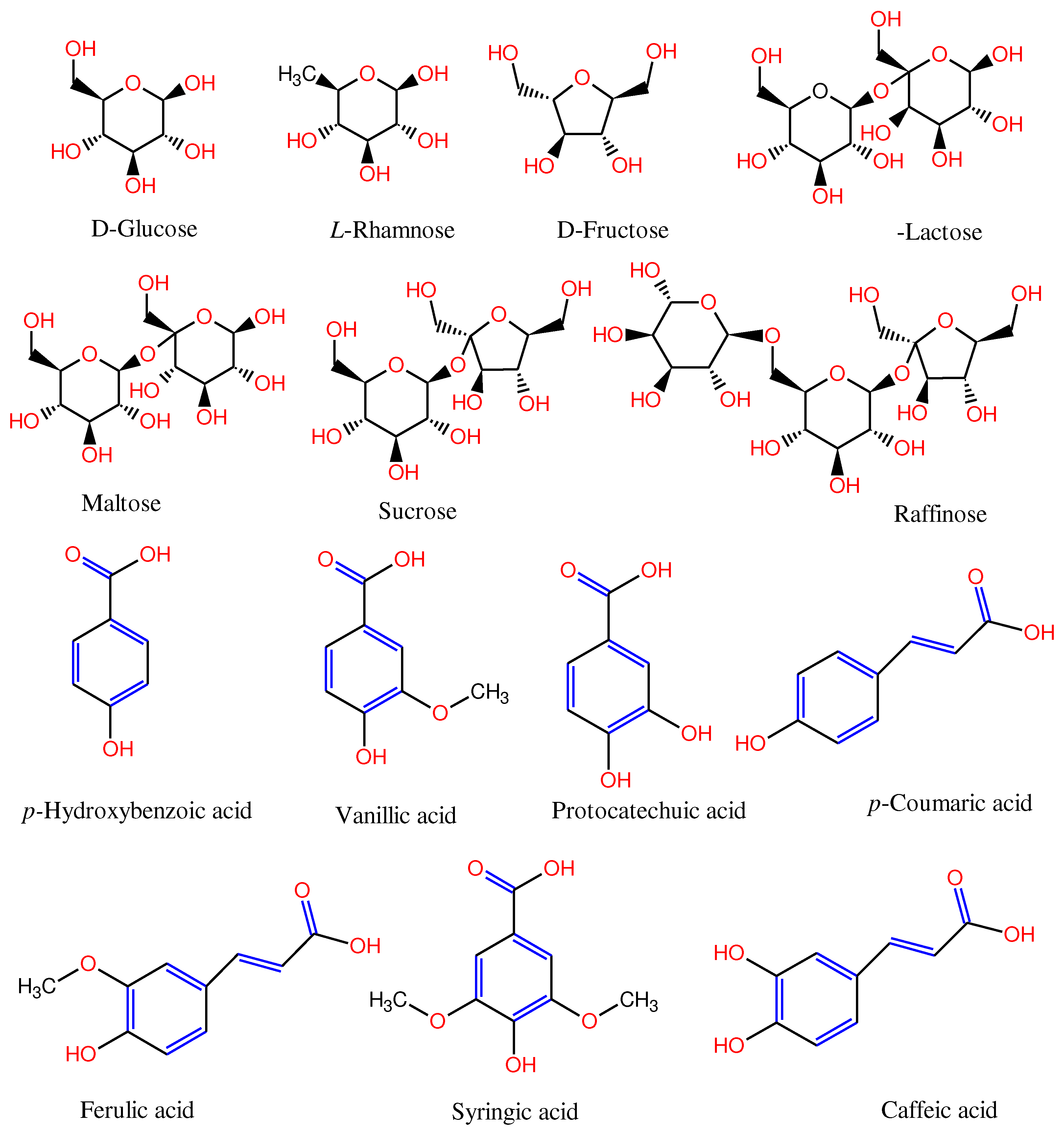
Figure 3.
Haematoxylin and eosin (H&E) stained sections of O. mossambicus. Spleen (A-C); A, CBD = control with melanoma-macrophages (MM) within the white pulp; B, FRT with fewer melanomacrophages and a few erythrocytes (Er); C, GRT with a distinct boundary between the white and red pulp and a few scattered melano-macrophages within both the red and white pulp. Liver (D – F).
Figure 3.
Haematoxylin and eosin (H&E) stained sections of O. mossambicus. Spleen (A-C); A, CBD = control with melanoma-macrophages (MM) within the white pulp; B, FRT with fewer melanomacrophages and a few erythrocytes (Er); C, GRT with a distinct boundary between the white and red pulp and a few scattered melano-macrophages within both the red and white pulp. Liver (D – F).
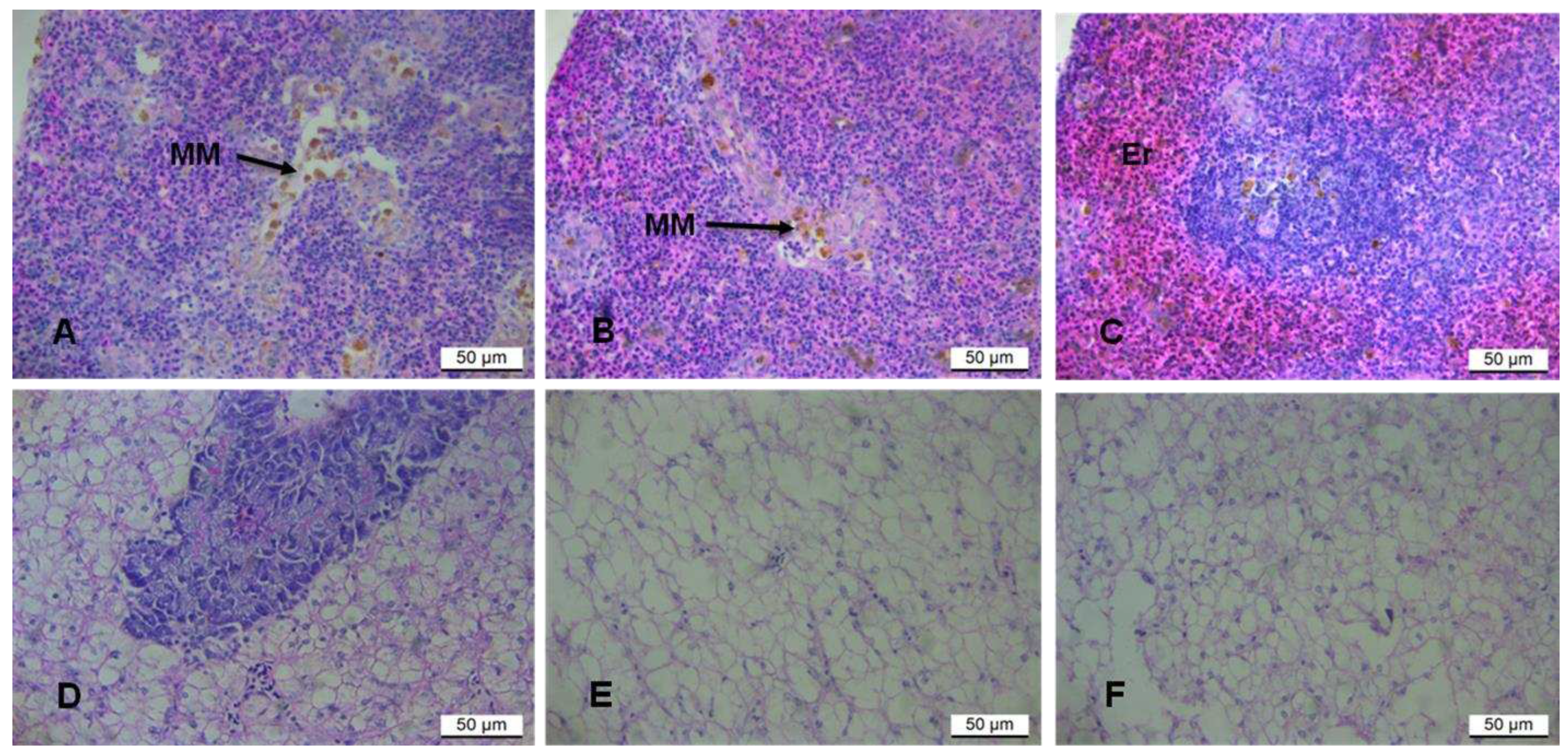
Figure 4.
Photomicrographs of Micronucleus (MN) assay in fish fed different rooibos tea extracts for 6 weeks, acridine orange (AO) stain. (A and B), Diet 1 CBD = (control); (C and D) Diet 2 (FRT), and (E and F), Diet 3 (GRT). Arrows in A,B and D indicate the micronucleus.
Figure 4.
Photomicrographs of Micronucleus (MN) assay in fish fed different rooibos tea extracts for 6 weeks, acridine orange (AO) stain. (A and B), Diet 1 CBD = (control); (C and D) Diet 2 (FRT), and (E and F), Diet 3 (GRT). Arrows in A,B and D indicate the micronucleus.
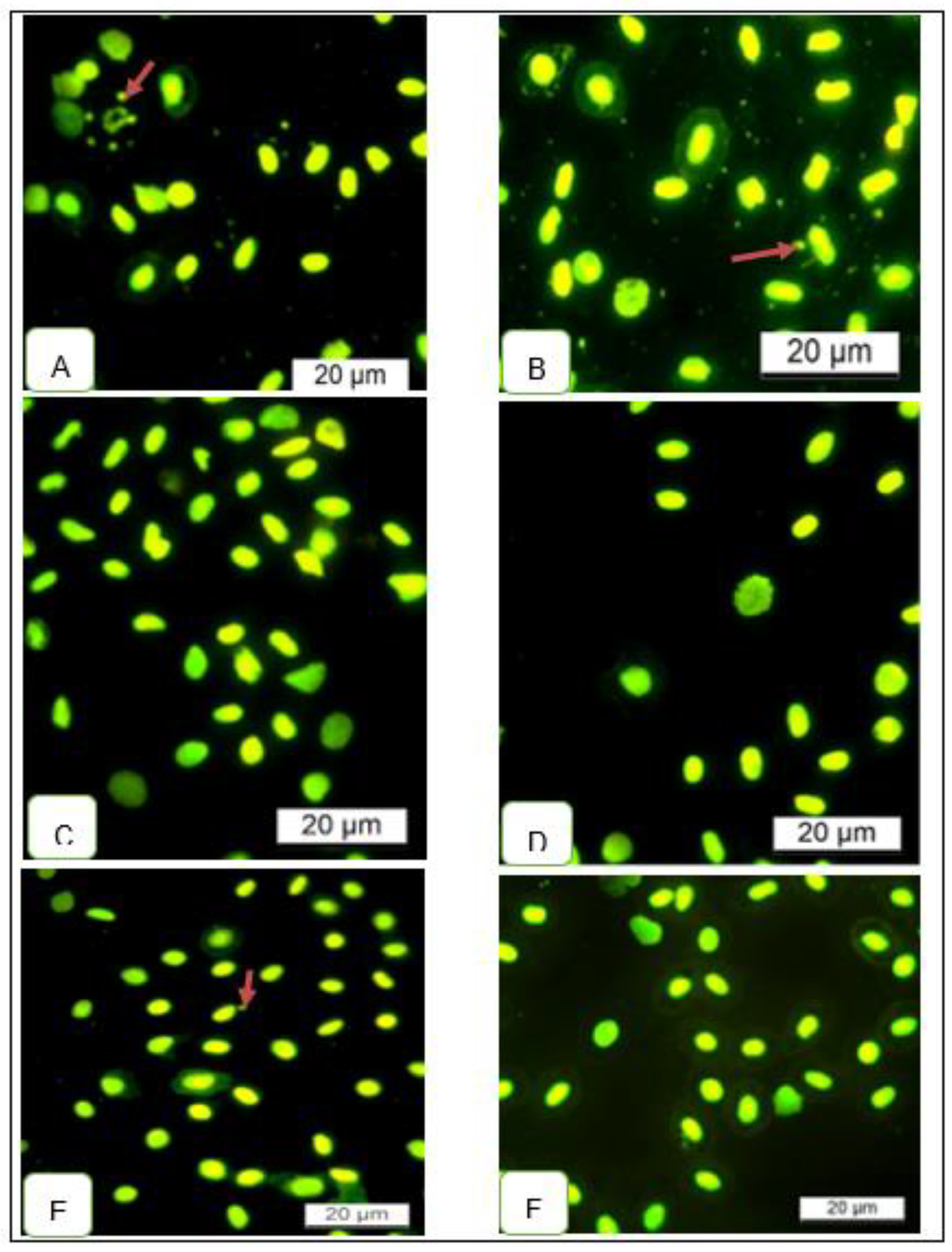

Table 1.
Amino acid and phenolic acids concentrations for the four (4) different teas, (-) = Absent/not detected.
Table 1.
Amino acid and phenolic acids concentrations for the four (4) different teas, (-) = Absent/not detected.
| FRT* | GRT* | |
|---|---|---|
| Essential amino acids (Concentration mg/L) | ||
| Alanine | 23.58 | 26.95 |
| Glycine | 16.6 | 19.69 |
| Valine | 15.25 | 17.64 |
| Leucine | 30.44 | 36.21 |
| Isoleucine | 11.9 | 13.89 |
| Proline | 24.98 | 31.02 |
| Methionine | 6.59 | 7.07 |
| Serine | 11.09 | 12.95 |
| Threonine | 22.79 | 25.16 |
| Phenylalanine | 17.28 | 20.48 |
| Aspartic acid | 30.21 | 44.27 |
| Glutamic acid | 41.43 | 47.95 |
| Asparagine | 10.64 | 12.43 |
| Tyrosine | 10.87 | 12.77 |
| Phenolic acids (Concentration ppb:µg/L) | ||
| FRT* | GRT* | |
| 4-Hydroxybenzoic_acid | 183.93 | 71.72 |
| Vanillic acid | 752.47 | 161.62 |
| Protocatechuic acid | 1243 | 920.2 |
| m-coumaric acid | − | − |
| p-coumaric acid | 129.43 | 77.75 |
| Syringic acid | 1404.98 | 135.83 |
| Ferulic acid | 3572.69 | 833.71 |
| Caffeic acid | 490.83 | 88.36 |
*FRT – Fermented rooibos tea, *GRT – Green rooibos tea.
Table 2.
Proximate Chemical Composition (wet basis) of the basal diet.
| Diets | CBD* | FRT* | GRT* |
|---|---|---|---|
| Proximate Composition % | |||
| Dry matter | 95.28 | 95.29 | 95.32 |
| Moisture | 4.72 | 4.71 | 4.68 |
| Protein | 32.99 | 32.8 | 33.17 |
| Fat | 4.94 | 4.75 | 4.81 |
| Ash | 7.03 | 6.98 | 7.03 |
| Fibre | 3.32 | 2.69 | 3.87 |
| Carbohydrates | 50.01 | 50.73 | 50.31 |
Table 3.
Growth performance and survival rate of larval Oreochromis mossambicus fed three different diets: commercial basal diet (CBD), and diets supplemented with either fermented (FRT) or green (GRT) rooibos tea extracts.
Table 3.
Growth performance and survival rate of larval Oreochromis mossambicus fed three different diets: commercial basal diet (CBD), and diets supplemented with either fermented (FRT) or green (GRT) rooibos tea extracts.
| Growth parameters | Basal diet | FRT* | GRT* |
|---|---|---|---|
| Initial number of fish - NI | 300 | 300 | 300 |
| Initial body weight (g) - IBL | 0.55 ± 0.02a | 0.55 ± 0.02a | 0.54 ± 0.03a |
| Initial body length (cm) - IBL | 2.22 ± 0.41a | 2.24 ± 0.30a | 2.21 ± 0.45a |
| Final body weight (cm) - FBW | 2.86 ± 0.60a | 4.01 ± 0.88b | 4.13 ± 0.75b |
| Final body length (cm) - FBL | 5.25 ± 0.51a | 5.51 ± 0.50ab | 5.84 ± 0.46bc |
| SGR (%) | 3.65 ± 0.40 a | 4.38 ± 0.44 b | 4.48 ± 0.42 b |
| Condition factor (%) - CF | 13.14 ± 4.87 a | 6.92 ± 2.45 b | 6.70 ± 2.53 b |
| Growth (%) | 5.15 ± 1.32 a | 7.69 ± 1.95 b | 7.98 ± 1.65 b |
| HIS (%) | 6.59 ± 1.91a | 6.75± 1.96a | 7.02 ± 2.10a |
| VSI (%) | 21.06 ± 7.10a | 22.91 ± 11.87b | 26.64 ± 12.52c * |
| Rate of weight gain (g) - RWG | 2.32 ± 0.59 a | 3.46 ± 0.88 b | 3.59 ± 0.74 b |
| Food conversion ratio (%) – FCR | 2.32± 0.57a | 1.50± 0.25b | 1.41± 0.07b |
| Survival Rate (%) - SR | 95.30 | 96.70 | 96.30 |
*FRT – Fermented rooibos tea, *GRT – Green rooibos tea.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



