
Submitted:
02 October 2023
Posted:
03 October 2023
You are already at the latest version
Abstract
Vasoreactivity is relatively well documented in terrestrial snakes, but has previously been investigated in only one semi-arboreal snake species. Consequently, the extent to which vasoreactivity is common across snake taxa, or varies by habitat is unclear. The Tokara habu (Protobothrops tokarensis) is a semi-arboreal snake endemic to only two small adjacent Japanese islands, and hence a useful species for further investigation of vasoreactivity. We evaluated responses to known vasoactive substances in thoracic aortas isolated from Tokara habu. Under resting tension, noradrenaline and angiotensin II induced concentration-dependent contraction, but acetylcholine, serotonin (5-hydroxytriptamine; 5-HT), and isoproterenol induced relaxation followed by contraction. Histamine and rattlesnake bradykinin had no effect. Experiments with receptor-specific antagonists suggest that M1 and M3 receptors are involved in the acetylcholine-induced response, 5-HT1, 5-HT2 and 5-HT7 receptors in the serotonin-induced response, and β1 and β2 receptors in isoproterenol-induced relaxation. NO may be involved in acetylcholine-induced relaxation, but not the responses to serotonin or isoproterenol. In contrast to the uniform vasoreactivity observed in terrestrial snakes, the vasoreactivity of semi-arboreal snakes may be governed by diverse regulatory mechanisms.
Keywords:
acetylcholine
; adrenoceptors
; environment
; habitat
; Tokara Islands
; serotonin
; snake
; vasoreactivity
1. Introduction
Snakes have colonized an environmentally diverse range of niches since they first evolved between 174 and 163 million years ago [1]. Their adaptations to these habitats present an interesting contrast to those seen in many other vertebrates [2]. Examples of habitat-adapted taxa include arboreal primates, various terrestrial quadrupeds, and cetaceans among mammals, and tree-nesting birds of flight, terrestrial birds such as ostriches, and semi-aquatic penguins among avian species. Snakes are distinctive as a group of taxa because they are found in terrestrial, arboreal, subterrain, and aquatic environments, and many of their basic characteristics (including tissue organization) are common at the order level in Linnean classification—specifically for the suborder Serpentes—unlike birds and mammals, which tend to show such characteristics at the class level. Snakes are thus useful animals for understanding environmental adaptations to a range of habitats. However, the mechanisms underlying these adaptations remain largely unknown [3], and their elucidation would be of great benefit for evolutionary studies.
Snakes vary in size, but they share a common shape. Their cardiovascular systems thus all operate within a similar cranial-caudal arrangement, but confront different habitat-related challenges in achieving systemic circulation. A major example of such a challenge is gravity, which theoretically exerts different effects on terrestrial, marine, and arboreal species, generating different levels of hydrostatic pressure, which the circulation must overcome [3,4,5]. A useful way to understand how the circulation achieves this is to investigate vasoreactivity, which will reveal how the vascular endothelium and smooth muscles react to a range of potentially vasoactive substances, and what receptor populations it possesses.
Vasoreactivity has been studied in the thoracic aorta in three terrestrial snakes [6,7,8,9], spanning the Natricinae, Colubridae and Viperidae families. Common features across these reports included noradrenaline-induced contractions in two of three species [7,9], angiotensin II-induced contractions in two of three species [6,7], and acetylcholine-induced relaxation in one of three species [8]. Vasoreactivity has been investigated in only one semi-arboreal snake, the Yellow rat snake (Pantherophis obsoletus) [10], which showed some similarities and some differences with the terrestrial snakes. Broadly this may suggest that basic vasoreactivity may be similar in species colonizing similar habitats; however, only one semi-arboreal snake has previously reported on. Further investigations in other semi-arboreal snakes are required to establish whether snakes in this habitat exhibit a similar pattern of vasoreactivity.
A semi-arboreal species of potential interest for evolutionary physiologists is the Tokara habu (Protobothrops tokarensis), a mildly venomous snake of the Viperidae family which is found in only two small, adjacent islands in south-western Japanese archipelago (Takarajima and Kodakarajima) [11]. This insular species is phylogenetically very close to the Habu (Protobothrops flavoviridis) [12], a terrestrial species, inhabiting other Japanese islands [13]. The Tokara habu is very interesting from the perspective of environmental adaptive capacity because it evolved as a separate species relatively shortly after the separation of the nearby Amami Islands from the Tokara Islands, and changed its habitat (terrestrial to semi-arboreal) [11].
Accordingly, in the present study, we aimed to characterize thoracic aortic vasoreactivity in the Tokara habu, as a representative semi-arboreal snake species by applying a range of vasoactive substances (including noradrenaline, angiotensin II, and acetylcholine), to aortic rings harvested from snake carcasses in Kodakarajima. Furthermore, we conducted a field survey to verify the semi-arboreal behavior of the Tokra habu.
2. Materials and Methods
2.1. Sampling location
We obtained Tokara habu carcasses from the island of Kodakarajima (latitude: 29° 22' east; longitude: 129° 32' north), which belongs to the Tokara archipelago, Kagoshima prefecture, Japan (Figure 1). Tokara habu are 0.6–1.1 m in length and have a snout-vent length of less than 0.9 m [11]. The snakes’ habitat consists of secondary forests (where they reportedly often climb trees), shrublands, and grasslands [11].
2.2. Ethics approval
Before the collection of snake carcasses, we submitted an experimental proposal to the Residential Affairs Division of Toshima Village Office, from which approval for the experiments (Approval Nos. 2260, 2930, and 2163) was obtained. All experiments were performed in accordance with the Guidelines for Animal Experiments of Kagoshima University. Since this experiment used arteries isolated from previously exterminated snakes, individual ethical and animal experimental approval was not required from Kagoshima University. AAALAC International has fully accredited our animal experimental facilities and programs since 2017.
2.3. Substances and Reagents
We used the following reagents at the final concentrations shown: noradrenaline (10−9–10−5 M), phentolamine mesylate (10−6 M), isoproterenol (10−9–10−5 M), propranolol (10−7 M), histamine hydrochloride (10−9–10−5 M), serotonin (5-hydroxytriptamine; 5-HT; 10−9−10−5 M), ketanserin tartrate (10−6 M), methiothepin maleate (10−8 M), SB269970 (I-3-(2-(2-(4-methylpiperidin-1-yl)-ethyl)pyrrolidine-1-sulphonyl) phenolhydrochloride), rattlesnake bradykinin ([Val1,Thr6]-bradykinin) (10−9–10−5 M), which was synthesized by Shimadzu Co. (Kyoto, Japan), atenolol (10−6 M, LKT Laboratories, Tokyo, Japan), butoxamine hydrochloride (10−6 M, Sigma-Aldrich, St. Louis, MO, USA), Nω-nitro-L-arginine (L-NNA; 10−4 M), sodium nitroprusside (SNP; 10−4 M, Nacalai Tesque, Kyoto, Japan), acetylcholine (10−9–10−5 M, Daiichi Sankyo, Tokyo, Japan), pirenzepine dihydrochloride (10−6 M; Santa Cruz Biotechnology, Santa Cruz, CA), methoctramine hydrate (10−6 M), hexahydro-sila-difenidol hydrochloride, p-fluoroanalog (pfHHSiD; 10−6 M ), angiotensin II (10−9–10−5 M). All reagents were employed in accordance with the relevant manufacturer’s guidelines. SB266970 was initially dissolved in dimethyl sulfoxide (DMSO) at 1 mM. The final DMSO concentration was 0.1% (v/v). All other drugs were dissolved in water. Each selected agent had previously been demonstrated to be an agonist in vertebrate taxa (including reptilian taxa) [15,16,17].
2.4. Functional study
Vasoreactivity was investigated in accordance with a previously published method, [18] with some modifications. Briefly, five rings each approximately 3 mm long were cut from the thoracic aorta of each exterminated snake. Each ring was mounted horizontally between two L-shaped stainless-steel holders, with one part fixed to an isometric force transducer, and immersed in a 5-mL water jacked organ bath (UMTB-a, Unique Medical Co. Ltd, Tokyo, Japan) containing oxygenated salt solution at 28.0 ± 0.5 °C (pH 7.4). The resting tension in each thoracic aorta was 2.4 mN.
Vascular tension strength was calculated based on percentage values for contraction (the contraction induced by 60 mM KCl was taken as 100%) and (the relaxation induced by 10−4 M sodium nitroprusside was taken as 100%).
2.5. Nitric Oxide Quantification Using Fluorescence
Nitric oxide (NO) was quantified using aorta specimens prepared in accordance with a previously described method [19], with some modifications. Briefly, each thoracic aorta (n=5 exterminated snakes; total: 1.20 ± 0.05 mg wet tissue per specimen) was cut into 4-mm segments and immersed in a 500-µL tube containing oxygenated physiological saline at 28 °C (pH 7.4). The segments were then treated with serotonin (10−6 M), acetylcholine (10−7 M), or isoproterenol (10−6 M) to achieve the relevant experimental condition. The involvement of endothelial NO was also investigated with comparator treatments involving pretreatment with L-NNA and incubation for 1 h.
2.6. Statistical analysis
Acquired data included the pEC50 values and Emax for each potentially vasoactive substance. The pEC50 value indicates the sensitivity of the substance to the corresponding receptor, and Emax indicates the strength of its effect on the vascular response. Higher pEC50 values thus indicate higher sensitivity to the binding receptor, and higher Emax indicates a greater impact on the vascular endothelial response.
Results are expressed as means ± SEM. Statistical analyses were performed by Student’s t-test or the Bonferroni test after one-way analysis of variance (Stat View J-4.5, Abacus Concepts Inc., Berkeley, CA, USA). Results were regarded as significant when the probability of rejecting a true null hypothesis was equal to or less than 5%.
2.7. Field research
To investigate the Tokara habu’s semi-arboreal behavior (how often and how high they climb), we carried out field observations to generate reference data. The field research was conducted over the entire area of Kodakarajima (0.98 km2), and involved searches to identify snakes over one day including periods of natural daylight and darkness, in September 2021 and October 2022. For each Tokara habu identified in these searches, we recorded the spatial position (on the ground or in a tree) and height from the ground (in meters).
3. Results
3.1. Responsiveness to noradrenaline, angiotensin II, histamine, rattlesnake bradykinin, isoproterenol, serotonin, and acetylcholine
We generated concentration-response curves for noradrenaline, angiotensin II, histamine, rattlesnake bradykinin, isoproterenol, serotonin, and acetylcholine based on data from thoracic aortas isolated from Tokara habu carcasses (Figure 2). Noradrenaline and angiotensin II induced contraction in a concentration-dependent manner. Isoproterenol, serotonin, and acetylcholine induced relaxation at a low concentration (10−9 M–10−7 or 10−6 M) followed by contraction at a high concentration (10−6 M–10−5 or only 10−5 M), in resting tension. Histamine and rattlesnake bradykinin had no effect on the resting tension or precontracted condition induced with noradrenaline (10−6 M). The maximum contractile responses for noradrenaline and angiotensin II, the maximal relaxation responses for isoproterenol, serotonin and acetylcholine, and the pEC50 (indicating sensitivity for the receptor) and Emax (indicating response strength) values for all vasoactive substances are shown in Table 1.
3.2. Effect of phentolamine on noradrenaline-induced contraction
We examined the effect of phentolamine (10−6 M, a non-selective α adrenoceptor antagonist) on noradrenaline-induced contraction. Phentolamine shifted the concentration-response curve for noradrenaline to the right in parallel (Figure 3).
3.3. Effects of β adrenoceptor antagonists on isoproterenol-induced relaxation
We examined the effects of propranolol (10−7 M, a non-selective β adrenoceptor antagonist), atenolol (10−6 M, a β1 adrenoceptor antagonist), and butoxamine (10−6 M, a β2 adrenoceptor antagonist), on isoproterenol-induced relaxation in resting tension (Figure 4). Propranolol completely abolished isoproterenol-induced relaxation and induced contraction. Atenolol and butoxamine also had effects in the same direction, changing isoproterenol-induced relaxation to contraction, but their effects were less strong than that of propranolol. Phentolamine shifted the isoproterenol-induced contraction to the right in parallel in the presence of propranolol.
3.4. Effects of serotonin receptor antagonists and L-NNA on serotonin-induced responses
We examined the effects of L-NNA (a NO synthase inhibitor), methiothepin (a 5-HT1 and 5-HT2 receptors antagonist), ketanserin (a 5-HT2 receptor antagonist), and SB269770 (a 5-HT7 receptor antagonist) on serotonin-induced relaxation. No antagonist or inhibitor significantly reduced serotonin-induced relaxation. We then examined the effect of these substances when applied in combinations, and found that a combination of L-NNA, SB269970, and methiothepin or ketanserin abolished serotonin-induced relaxation at 10−9 and 10−8 M, but had no significant effect at 10−7 or 10−6 M (Figure 5A). Serotonin had induced contraction at 10−5 M (indicated with the Δ symbol in Figure 2); therefore, we investigated the effects of methiothepin and ketanserin on this contraction, and the results are shown in Figure 5B. Methiothepin, but not ketanserin, significantly inhibited serotonin (10−5 M)-induced contraction (P < 0.01).
3.5. Effects of muscarine receptor antagonists on acetylcholine-induced relaxation
Acetylcholine induced relaxation at a low concentration of 10−9–10−7 M. To clarify the receptor subtype involved in this relaxation and the involvement of NO, we then applied several muscarinic (M) receptor antagonists and L-NNA. Relaxation induced by acetylcholine (10−7 M) was abolished by atropine (a non-selective M receptor antagonist), L-NNA, pirenzepine (an M1 receptor antagonist), and pfHHSiD (an M3 receptor antagonist) but not by methoctramine (an M2 receptor antagonist), as shown in Figure 6.
3.6. Effects of muscarine receptor antagonists on acetylcholine-induced contraction in the presence of L-NNA
Acetylcholine induced contraction at high concentrations of 10−6 and 10−5 M (● in Figure 2). Since acetylcholine-induced relaxation was abolished by L-NNA (Figure 6), we examined the effect of M receptor antagonists on acetylcholine-induced contraction in the presence of L-NNA (Figure 7). The treatment with atropine abolished acetylcholine (10−5 M)-induced contractions completely. The treatments with pirenzepine (10−6 M) and pfHHSiD (10−6 M) significantly inhibited acetylcholine-induced contractions (P < 0.01), whereas methoctramine (10−6 M) had no significant effect.
3.7. Nitric oxide production by serotonin, acetylcholine and isoproterenol
To determine whether NO was involved in the relaxation responses induced by serotonin, acetylcholine and isoproterenol, aorta specimens were treated with serotonin, acetylcholine and isoproterenol, and NO production was measured (Figure 8). The effect of L-NNA on any NO production was also examined; the amounts of NO production induced by serotonin (10−6 M) and isoproterenol (10−6 M) were not significantly different from that seen with control (the absence of any treatment). In contrast, a significant increase was observed with acetylcholine (10−7 M) versus control, and the increase in NO production was completely abolished by L-NNA treatment.
3.8. Result of field observation
We identified a total of 18 individual Tokara habu during the two field survey times. Fifteen of the 18 snakes were found in trees (83.3%), at a mean height of 1.3 ± 0.1 m (minimum: 0.5 m, maximum: 1.8 m).
4. Discussion
To the best of our knowledge, this is the first report on thoracic aortic responses to noradrenaline, angiotensin II, histamine, rattlesnake bradykinin, isoproterenol, serotonin, and acetylcholine in the Tokara habu. Interestingly, responses to isoproterenol, serotonin and acetylcholine involving initial relaxation followed by contraction. The response pattern not been previously reported in other snakes, and in particular, there is no previous report on aortic relaxation induced by serotonin and isoproterenol in any other snake species. We thus consider that our findings illustrate the characteristic pattern of vasoreactivity in the Tokara habu.
Tokara habu reportedly exhibit semi-arboreal behavior [11,20], and this was substantiated by the findings of our field study. We observed 83.3% (15/18) of identified snakes were in trees. There appear to be some physiological differences between the Tokara habu and the Yellow rat snake, despite both species exhibiting similar tree-climbing behavior. Histamine induces no response in the former, but contraction in the latter [10]. This phenomenon cannot yet be explained, and further studies are required. Multiple previous studies have focused on individual vasoactive substances in a single terrestrial snake species [6,7,8,9], investigating only either contraction or relaxation. However, vascular responses (encompassing both contraction and relaxation) appear more complex for semi-arboreal snakes (the Tokara habu investigated here, and the Yellow rat snake, reported elsewhere [10]) and may involve multiple vasoactive substances.
When compared with terrestrial snakes, the Tokara habu showed a more diverse and stronger relaxation response. The first major difference concerned acetylcholine-induced relaxation, a response that does not occur in terrestrial snakes in the absence of precontraction [8]. Previous studies have shown that the relaxation response usually cannot be observed or is very small without precontraction, for acetylcholine and other agents [18,21,22]. The second major difference concerned the isoproterenol-induced relaxation mediated via β adrenoceptors in the Tokara habu, which contrasts with isoproterenol inducing only contraction and not relaxation in a terrestrial snake, the Jararaca [9]. The Tokara habu and Jararaca both belong to Viperidae family, so we consider this physiological difference in vascular response may reflect the influence of some adaptation to a semi-arboreal habitat by the Tokara habu.
The Tokara habu may have developed a complex mechanism involving multiple responses to deal with the semi-arboreal nature of its habitat, which necessitates tree-climbing movements (and the concomitant large variations in the incline of the craniocaudal axis, especially with regard to the blood supply from the heart), and entails the need for flexible blood pressure. One species that appears to have dealt with possible gravitational effects on the cardiovascular system is the giraffe, which is reported to be naturally and healthily hypertensive due to the pumping of blood to their cranially elevated brain [23,24]; however, hypertension usually causes a range of disorders in most other animals [25]. Therefore, Tokara habu and Yellow rat snakes need mechanisms to counteract increases in blood pressure when they climb trees, and we postulate that such mechanisms may involve a relaxation response as part of their blood pressure regulation. However, a single mechanism may not suffice to achieve this regulation. Thus, a variety of relaxation mechanisms could have developed to deal with tree climbing, and these mechanisms could include NO-mediated responses such as those to acetylcholine and unmediated responses such as those to isoproterenol and serotonin. Blood pressure and the position of the heart in the body are known to vary in snakes depending on their habitat [2,26,27]. The biphasic thoracic aortic responses to acetylcholine, isoproterenol and serotonin we noted in the Tokara habu may reflect adaptations to large changes in blood pressure that occur during tree climbing as the snake’s head and heart become vertically aligned.
Focusing on the specific responses induced by vasoactive substances in this study, noradrenaline induced contraction in a concentration-dependent manner, and phentolamine shifted the concentration-response curve for noradrenaline to the right in parallel (Figure 3). The pEC50 value (indicating sensitivity for the relevant receptor) for noradrenaline in Tokara habu (6.04) was similar to that in bovine median caudal arteries (6.16), where responses are mediated via α1-adrenoceptors [28], as well as those in other snakes, such as the Indian cobra (Naja naja; 6.04) [7], Jararaca (Bothrops jararaca; 6.63) [9], and Yellow rat snake (Pantherophis obsoletus; 6.72) [10]. Phentolamine was also reported to have a similar effect in the Jararaca and Indian cobra [7,9]. These results suggest that noradrenaline-induced contraction in Tokara habu is mediated via α1-adrenoceptors.
Amino acid sequencing of angiotensin-like peptides in snakes previously revealed two types of angiotensin II in snakes, one identical to the corresponding human peptide and one with a sequence that differs by a single amino acid [29]. Angiotensin II induced contraction in the thoracic aorta of the Tokara habu. In other snakes, angiotensin II also induced aortic contraction [6,7,10]. Renin or renin-like substances are present in the kidneys of many vertebrates, from bony fishes to mammals, and are responsible for blood pressure regulation [30], and may play an important role in blood pressure regulation in snakes. The relevant pEC50 values in semi-arboreal snakes, the Tokara habu and Yellow rat snake, were 7.39 and 6.50 [10], respectively, and those in terrestrial snakes, the Indian cobra and Jararaca, were 8.73 and 6.76, respectively [6,7]. These figures suggest that sensitivity to the relevant receptor may be not related to habitat.
The β-adrenoceptor is classified into β1, β2 and β3 subtypes, all three of which are expressed in smooth muscle [31]. In this study, isoproterenol induced contraction at a high concentration, although it induced relaxation at a low concentration. Propranolol (10−7 M), abolished the relaxation response (Figure 4), suggesting that the relaxation had been mediated through β1 and/or β2 adrenoceptors. This is based on propranolol being a classic non-selective antagonist of β-adrenoceptors, known to block the β3 adrenoceptor only at high concentrations (10−6 or 10−5 M) [31,32,33]. We showed that pretreatment with atenolol and butoxamine significantly curtailed the relaxation response induced by isoproterenol. Therefore, we consider that β1 and β2 adrenoceptors are present in the thoracic aorta of the Tokara habu. As shown in Figure 8, isoproterenol-treated thoracic aortas did not produce NO, suggesting that the relaxation response induced by isoproterenol is not mediated via NO. Phentolamine shifted the contraction response curve yielded by isoproterenol treatment in the presence of propranolol parallel to the right (Figure 4), suggesting that the contraction was mediated via α adrenoceptors. In another snake species, the Jararaca (a member of the same family as the Tokara habu), isoproterenol induced only contraction mediated via α adrenoceptors, and not relaxation [9].
Multiple classes of serotonin (5-HT) receptors exist, from 1 to 7, with several additional subtypes [34]. In snakes, serotonin reportedly induced basilar arterial contraction via the 5-HT1 receptor in the Habu snake [18], and was involved in pulmonary vascular responses (a study with no receptor mediation) in the file snake, Acrochordus granulatus [35]. Many vascular responses to serotonin are constrictive, but reported responses also include relaxation in the rat jugular vein [36,37], pig pulmonary artery [38,39] and equine coronary artery [40]. Although serotonin-induced relaxation responses vary between animal species and blood vessels, 5-HT1, 5-HT2, and 5-HT7 receptors and endothelial cell-derived NO all appear to be involved [34,36,37,38,39,40]. In further experiments with antagonists and inhibitors that effectively yielded serotonin-induced relaxation in previous studies, the combination of L-NNA, SB269970, methiothepin or ketanserin inhibited serotonin-induced relaxation at low concentrations but not at high concentrations (Figure 5A). Also, NO production was not observed after treating thoracic aortas with serotonin (Figure 8). These results suggest that serotonin-induced relaxation in the thoracic aorta of the Tokara habu does not involve NO and may involve other relaxing factor(s) in addition to 5-HT2 and 5-HT7 receptors, but the details are unknown and require further investigation. The contraction induced by serotonin at 10−5 M was significantly inhibited by methiothepin, but not by ketanserin (Figure 5B). This result suggests contraction induced via the 5-HT1 receptor.
Both acetylcholine-induced relaxation and contraction responses were significantly inhibited by pirenzepine and pfHHSiD (Figure 6 and Figure 7). In addition, the relaxation response was also abolished by L-NNA and acetylcholine-induced NO production was significantly increased versus no treatment (Figure 6 and Figure 7). These results suggest that the acetylcholine-induced relaxation response was mediated via M1 and M3 muscarinic receptors, and production of NO in vascular endothelial cells. Furthermore, as discussed in antecedent sections, the acetylcholine-induced thoracic aortic relaxation response appeared to be stronger in Tokara habu than other snakes, which may be attributed to vascular responses in other snakes being induced by acetylcholine only after noradrenaline-induced precontraction [8]. To summarize, acetylcholine induced relaxation mediated by NO via M1 and M3 receptors in vascular endothelial cells at a low concentration, and contraction mediated via M1 and M3 receptors in vascular smooth muscle at a high concentration, in this study.
Histamine induces the strong tachycardia elicited via H1 and H2 receptors under anesthesia [41,42]. However, histamine may not be involved in the vascular response in Tokara habu thoracic aortas.
Bradykinin has previously been reported to show amino acid sequences that differ between humans and mammals at only two positions. In snakes, its vasoactive effect reportedly induces relaxation mediated by NO or an endothelium-derived relaxing factor [18,43]. In this study, rattlesnake bradykinin had no effect on resting tension or pre-contraction. Therefore, we consider that rattlesnake bradykinin may not be involved in the thoracic aortic vasoreactivity in the Tokara habu.
5. Conclusions
In the thoracic aorta of the Tokara habu, noradrenaline and angiotensin II induced contraction, whereas acetylcholine, isoproterenol and serotonin induced relaxation followed by contraction; histamine and rattlesnake bradykinin had no effect on resting tension. Unlike the monotonous vasoreactivity of terrestrial snakes, vasoreactivity in the Tokara habu may be governed by diverse regulatory mechanisms that are characteristic of snakes adapting to their semi-arboreal habitat.
Author Contributions
Conceptualization, T.O. and A.M.; methodology, M.Z.I. and H.T.T.N.; formal analysis, T.O. and A.M.; investigation, T.O., S.W. and R.S.; resources, A.M. and M.S.; data curation, T.O.; writing—original draft preparation, T.O.; writing—review and editing, H.S., A.M., Y.U., and M.S.; supervision, A.M.; project administration, A.M.; funding acquisition, A.M. and H.T.T.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded in part by JSPS KAKENHI Grant Number 16580242 and Vietnam National Foundation for Science and Technology Development (NAFOSTED; grant number 09/2019/ TN).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We thank Masashi Higo of the village mayors of Toshima and the residential affairs division of Toshima village. We also thank to Hideyuki Iwashita for their assistance with sample collection and Rieko Uchida of Kagoshima University for assisting with research procedures.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Caldwell, M.W.; Nydam, R.L.; Palci, A.; Apesteguía, S. The oldest known snakes from the Middle Jurassic-Lower Cretaceous provide insights on snake evolution. Nat. Comun. 2015, 6, 5996. [Google Scholar] [CrossRef] [PubMed]
- Perez, D.; Sheehy III, C.M.; Lillywhite, H.B. Variation of organ position in snakes. J. Morphol. 2019, 280, 1798–1807. [Google Scholar] [CrossRef]
- Adamopoulos, K.; Koutsouris, D.; Zaravinos, A.; Lambrou, G.I. Gravitational influence on human living systems and the evolution of species on earth. Molecules 2021, 26, 2784. [Google Scholar] [CrossRef] [PubMed]
- Seymour, R.S.; Hargens, A.R.; Pedley, T.J. The heart works against gravity. Am. J. Physiol.: Regul., Integr. Comp. Physiol. 1993, 265, R715–R720. [Google Scholar] [CrossRef] [PubMed]
- White, C.R.; Seymour, R.S. The role of gravity in the evolution of mammalian blood pressure. Evol. 2014, 68, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Esteves, C.A.; Burckhardt, P.L.; Breno, M.C. Presence of functional angiotensin II receptor and angiotensin converting enzyme in the aorta of the snake Bothrops jararaca. Life Sci. 2012, 91, 944–950. [Google Scholar] [CrossRef]
- Yung, W.; Chiu, K. Contractile response of the isolated dorsal aorta of the snake to angiotensin II and norepinephrine. Gen. Comp. Endocrinol. 1985, 60, 259–265. [Google Scholar] [CrossRef]
- Knight, G.E.; Burnstock, G. Acetylcholine induces relaxation via the release of nitric oxide from endothelial cells of the garter snake (Thamnophis sirtalis parietalis) aorta. Comp. Biochem. Physiol. Part C: Pharmacol., Toxicol. Endocrinol. 1993, 106, 383–388. [Google Scholar] [CrossRef]
- Yamanouye, N.; Picarelli, Z. Characterization of postjunctional alpha-adrenoceptors in the isolated aorta of the snake Bothrops jararaca. Comp. Biochem. Physiol. Part C: Pharmacol., Toxicol. Endocrinol. 1992, 615–618. [Google Scholar] [CrossRef]
- Conklin, D.J.; Lillywhite, H.B.; Olson, K.R.; Ballard, R.E.; Hargens, A.R. Blood vessel adaptation to gravity in a semi-arboreal snake. J. Comp. Physiol. B 1996, 165, 518–526. [Google Scholar] [CrossRef]
- Toda, M. Red Data Book 2014.-Threatened Wildlife of Japan -: Reptilia /Amphibia; Ministry of Environment (ed.); GYOSEI Corporation: Tokyo, Japan, 2014; p. 82. [Google Scholar]
- Shibata, H.; Chijiwa, T.; Hattori, S.; Terada, K.; Ohno, M.; Fukumaki, Y. The taxonomic position and the unexpected divergence of the Habu viper, Protobothrops among Japanese subtropical islands. Mol. Phylogenet. Evol. 2016, 101, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Tashiro, Y.; Goto, A.; Kunisue, T.; Tanabe, S. Contamination of habu (Protobothrops flavoviridis) in Okinawa, Japan by persistent organochlorine chemicals. Environ. Sci. Pollut. Res. 2021, 28, 1018–1028. [Google Scholar] [CrossRef] [PubMed]
- Geospatial Information Authority of Japan. Maps & Geospatial Information. 2023. Available online: https://www.gsi.go.jp/ENGLISH/page_e30031.html (accessed on 22 September 2023).
- Perry, S.F.; Capaldo, A. The autonomic nervous system and chromaffin tissue: neuroendocrine regulation of catecholamine secretion in non-mammalian vertebrates. Auton. Neurosci. 2011, 165, 54–66. [Google Scholar] [CrossRef]
- Ravhe, I.S.; Krishnan, A.; Manoj, N. Evolutionary history of histamine receptors: early vertebrate origin and expansion of the H3–H4 subtypes. Mol. Phylogent. Evol. 2021, 154, 106989. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Filogonio, R.; Joyce, W. Evolution of the cardiovascular autonomic nervous system in vertebrates. In Primer on the autonomic nervous system, 4th ed.; Biaggioni, I., Browning, K., Fink, G., Jordan, J., Low, P.A., Paton, J.F.R., Eds.; Elsevier Inc./Academic Press: San Diego, USA, 2023; pp. 3–9. [Google Scholar]
- Yoshinaga, N.; Okuno, T.; Watanabe, Y.; Matsumoto, T.; Shiraishi, M.; Obi, T.; Yabuki, A.; Miyamoto, A. Vasomotor effects of noradrenaline, acetylcholine, histamine, 5-hydroxytryptamine and bradykinin on snake (Trimeresurus flavoviridis) basilar arteries. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol. 2007, 146, 478–483. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Ootawa, T.; Sekio, R.; Smith, H.; Islam, M.Z.; Uno, Y.; Shiraishi, M.; Miyamoto, A. Involvement of beta3-adrenergic receptors in relaxation mediated by nitric oxide in chicken basilar artery. Poultry Sci. 2023, 102, 102633. [Google Scholar] [CrossRef]
- Harrington, S.M.; De Haan, J.M.; Shapiro, L.; Ruane, S. Habits and characteristics of arboreal snakes worldwide: arboreality constrains body size but does not affect lineage diversification. Biol. J. Linn. Soc. 2018, 125, 61–71. [Google Scholar] [CrossRef]
- Islam, M.Z.; Van Dao, C.; Miyamoto, A.; Shiraishi, M. Rho-kinase and the nitric oxide pathway modulate basilar arterial reactivity to acetylcholine and angiotensin II in streptozotocin-induced diabetic mice. Naunyn-Schmiedeb’rg's Arch. Pharmacol. 2017, 390, 929–938. [Google Scholar] [CrossRef]
- Matsumoto, F.; Watanabe, Y.; Obi, T.; Islam, M.; Yamazaki-Himeno, E.; Shiraishi, M.; Miyamoto, A. Characterization of 5-hydroxytryptamine-induced contraction and acetylcholine-induced relaxation in isolated chicken basilar artery. Poult. Sci. 2012, 91, 1158–1164. [Google Scholar] [CrossRef]
- Aalkjær, C.; Wang, T. The Remarkable Cardiovascular System of Giraffes. Annu. Rev. Physiol. 2021, 83, 1–15. [Google Scholar] [CrossRef]
- Mitchell, G.; Skinner, J.D. An allometric analysis of the giraffe cardiovascular system. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol. 2009, 154, 523–529. [Google Scholar] [CrossRef]
- Mills, K.T.; Stefanescu, A.; He, J. The global epidemiology of hypertension. Nat. Rev. Nephrol. 2020, 16, 223–237. [Google Scholar] [CrossRef]
- Lillywhite, H.B. Snakes, blood circulation and gravity. Sci. Am. 1988, 259, 92–99. [Google Scholar] [CrossRef]
- Lillywhite, H.B.; Albert, J.S.; Sheehy, C.M.; Seymour, R.S. Gravity and the evolution of cardiopulmonary morphology in snakes. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol. 2012, 161, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Hill, B.J.; Dyer, D.C. Pharmacological characterization of α-adrenoceptors in the bovine median caudal artery. Eur. J. Pharmacol. 1997, 339, 147–151. [Google Scholar] [CrossRef]
- Borgheresi, R.A.; Dalle Lucca, J.; Carmona, E.; Picarelli, Z.P. Isolation and identification of angiotensin-like peptides from the plasma of the snake Bothrops jararaca. Comp. Biochem. Physiol., Part B: Biochem. Mol. Biol. 1996, 113, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A. Comparative physiology of the renin-angiotensin system. In Federation Proceedings,; 1977; Volume 36, pp. 1776–1780. [Google Scholar]
- Tanaka, Y.; Horinouchi, T.; Koike, K. New insights into β-adrenoceptors in smooth muscle: Distribution of receptor subtypes and molecular mechanisms triggering muscle relaxation. Clin. Exp. Pharmacol. Physiol. 2005, 32, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Bardou, M.; Loustalot, C.; Cortijo, J.; Simon, B.; Naline, E.; Dumas, M.; Esteve, S.; Croci, T.; Chalon, P.; Frydman, R.; et al. Functional, biochemical and molecular biological evidence for a possible β3-adrenoceptor in human near-term myometrium. Br. J. Pharmacol. 2000, 130, 1960–1966. [Google Scholar] [CrossRef]
- Flacco, N.; Segura, V.; Perez-Aso, M.; Estrada, S.; Seller, J.; Jiménez-Altayó, F.; Noguera, M. ’ D'Ocon, P.; Vila, E.; Ivorra, M. Different β-adrenoceptor subtypes coupling to cAMP or NO/cGMP pathways: implications in the relaxant response of rat conductance and resistance vessels. Br. J. Pharmacol. 2013, 169, 413–425. [Google Scholar] [CrossRef]
- Hannon, J.; Hoyer, D. Molecular biology of 5-HT receptors. Behav. Brain Res. 2008, 195, 198–213. [Google Scholar] [CrossRef]
- Donald, J.; Lillywhite, H. Adrenergic nerves and 5-hydroxytryptamine-containing cells in the pulmonary vasculature of the aquatic file snake Acrochordus granulatus. Cell Tissue Res. 1989, 256. [Google Scholar] [CrossRef] [PubMed]
- Bodelsson, M.; Törnebrandt, K.; Arneklo-Nobin, B. Endothelial relaxing 5-hydroxytryptamine receptors in the rat jugular vein: similarity with the 5-hydroxytryptamine1C receptor. J. Pharmacol. Exp. Ther. 1993, 264, 709–716. [Google Scholar] [PubMed]
- Ellis, E.S. , Byrne, C., Murphy, O.E., Tilford, N.S., Baxter, G.S. Mediation by 5-hydroxytryptamine2B receptors of endothelium-dependent relaxation in rat jugular vein. Br. J. Pharmacol. 1995, 114, 400–404. [Google Scholar] [CrossRef] [PubMed]
- Glusa, E.; Pertz, H.H. Further evidence that 5-HT-induced relaxation of pig pulmonary artery is mediated by endothelial 5-HT2B receptors. Br. J. Pharmacol. 2000, 130, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Jähnichen, S.; Glusa, E.; Pertz, H.H. Evidence for 5-HT2B and 5-HT7 receptor-mediated relaxation in pulmonary arteries of weaned pigs. Naunyn-Schmiedeb’rg's Arch. Pharmacol. 2005, 371, 89–98. [Google Scholar] [CrossRef]
- Obit, T.; Kabeyama, A.; Nishio, A. Equine coronary artery responds to 5-hydroxytryptamine with relaxation in vitro. J. Vet. Pharmacol. Ther. 1994, 17, 218–225. [Google Scholar] [CrossRef]
- Skovgaard, N.; Møller, K.; Gesser, H.; Wang, T. Histamine induces postprandial tachycardia through a direct effect on cardiac H2-receptors in pythons. Am. J. Physiol.: Regul., Integr. Comp. Physiol. 2009, 296, R774–R785. [Google Scholar] [CrossRef]
- Skovgaard, N.; Abe, A.S.; Taylor, E.W.; Wang, T. Cardiovascular effects of histamine in three widely diverse species of reptiles. J. Comp. Physiol. B 2018, 188, 153–162. [Google Scholar] [CrossRef]
- Galli, G.L.; Skovgaard, N.; Abe, A.S.; Taylor, E.W.; Conlon, J.M.; Wang, T. Cardiovascular actions of rattlesnake bradykinin ([Val1, Thr6] bradykinin) in the anesthetized South American rattlesnake Crotalus durissus terrificus. Am. J. Physiol.-Regul., Integr. Comp. Physiol. 2005, 288, R456–R465. [Google Scholar] [CrossRef]
Figure 1.
(A) A Tokara habu, Protobothrops tokarensis, on a tree. (B) Locations of Takarajima and Kodakarajima within the Tokara islands, Japan. This figure was created by processing the relevant Geospatial Information Authority of Japan (GSI) tiles [14].
Figure 1.
(A) A Tokara habu, Protobothrops tokarensis, on a tree. (B) Locations of Takarajima and Kodakarajima within the Tokara islands, Japan. This figure was created by processing the relevant Geospatial Information Authority of Japan (GSI) tiles [14].
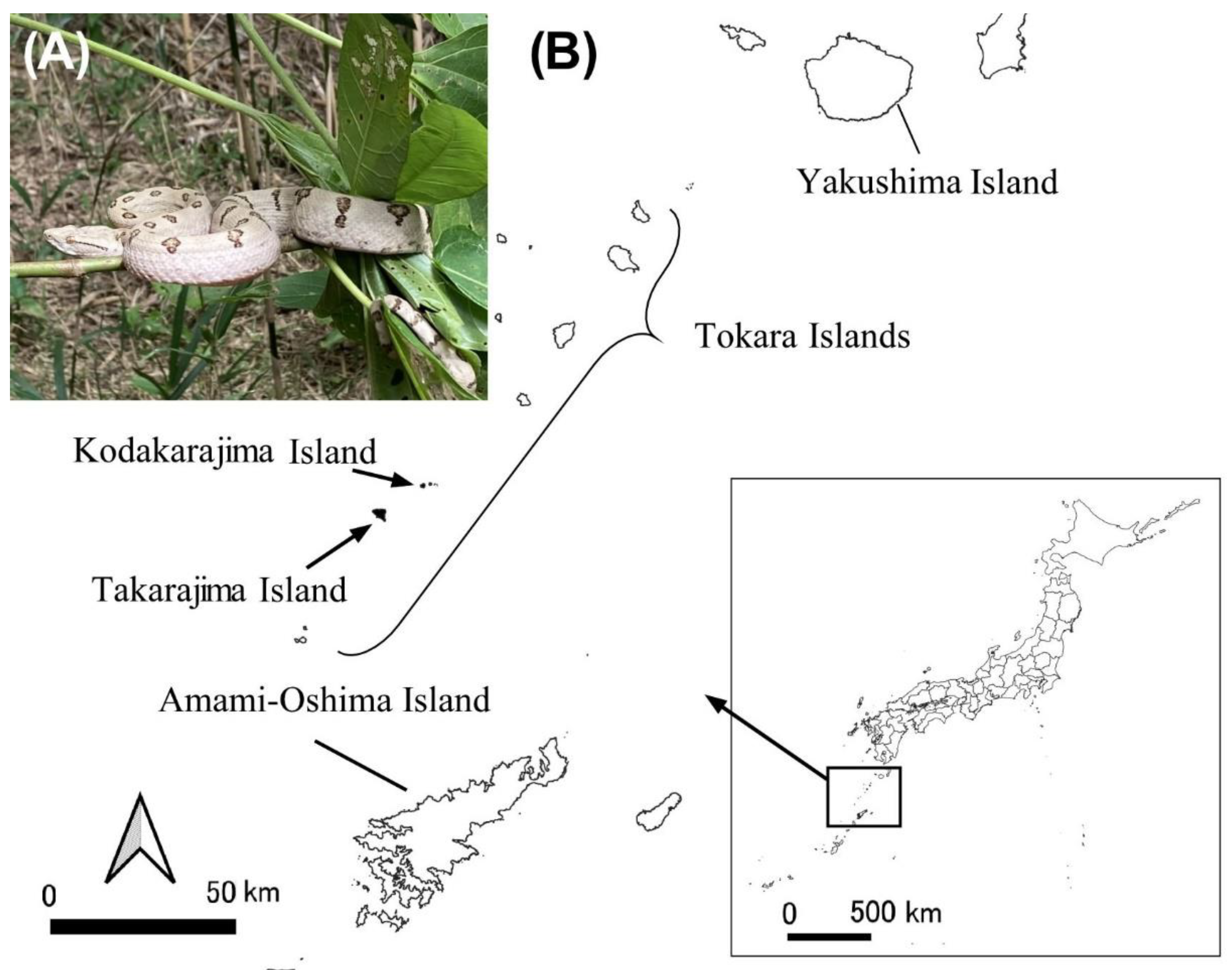
Figure 2.
Responsiveness to noradrenaline (○), angiotensin II (♦), histamine (▲), rattlesnake bradykinin (□), isoproterenol (×), serotonin (Δ), acetylcholine (●) in isolated Tokara habu thoracic aortas under resting tension. The contraction induced by 60 mM KCl was taken as 100%. The relaxation induced by sodium nitroprusside (10−4 M) was taken as 100%. Each point represents the mean ± SEM for 12 snakes.
Figure 2.
Responsiveness to noradrenaline (○), angiotensin II (♦), histamine (▲), rattlesnake bradykinin (□), isoproterenol (×), serotonin (Δ), acetylcholine (●) in isolated Tokara habu thoracic aortas under resting tension. The contraction induced by 60 mM KCl was taken as 100%. The relaxation induced by sodium nitroprusside (10−4 M) was taken as 100%. Each point represents the mean ± SEM for 12 snakes.
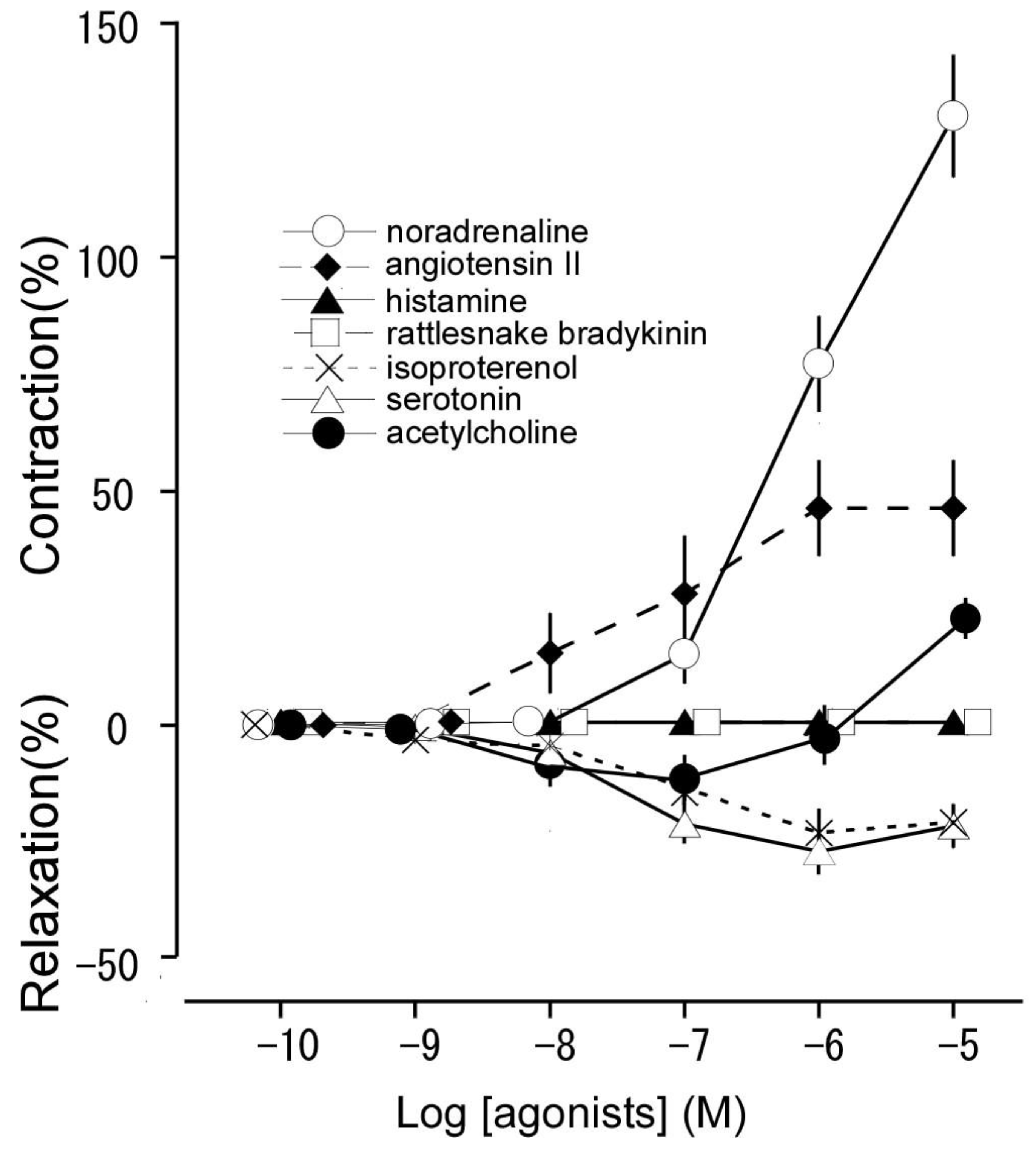
Figure 3.
Effect of phentolamine (○: a non-selective α adrenoceptor antagonist, 10−6 M) on noradrenaline-induced contraction (●: Control) in isolated thoracic aortas. The contraction induced by 60 mM KCl was taken as 100%. Each point represents the mean ± SEM for 6 snakes.
Figure 3.
Effect of phentolamine (○: a non-selective α adrenoceptor antagonist, 10−6 M) on noradrenaline-induced contraction (●: Control) in isolated thoracic aortas. The contraction induced by 60 mM KCl was taken as 100%. Each point represents the mean ± SEM for 6 snakes.
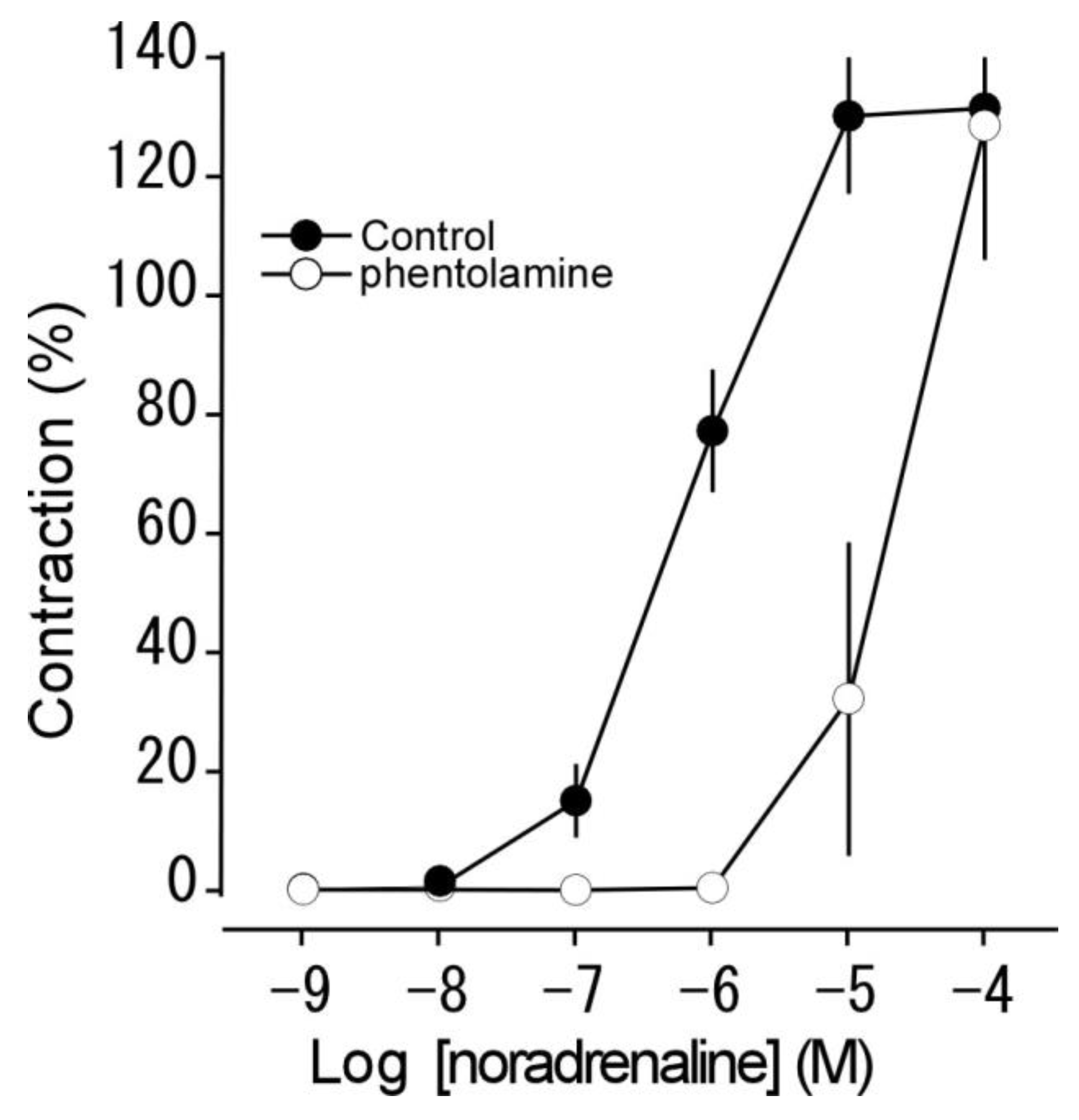
Figure 4.
Effects of propranolol (♦: 10−7 M, a β1 and β2 adrenoceptor antagonist), atenolol (Δ: 10−6 M, selective β1 receptor antagonist), butoxamine (■: 10−6 M, a selective β2 receptor antagonist), and propranolol (10−6 M) + phentolamine (10−6 M) on isoproterenol-induced relaxation and contraction (●: Control) in isolated thoracic aortas. The contraction induced by 60 mM KCl was taken as 100%. The relaxation induced by sodium nitroprusside (10−4 M) was taken as 100%. Each point represents the mean ± SEM for 6 snakes. (⁎P < 0.05 vs. control).
Figure 4.
Effects of propranolol (♦: 10−7 M, a β1 and β2 adrenoceptor antagonist), atenolol (Δ: 10−6 M, selective β1 receptor antagonist), butoxamine (■: 10−6 M, a selective β2 receptor antagonist), and propranolol (10−6 M) + phentolamine (10−6 M) on isoproterenol-induced relaxation and contraction (●: Control) in isolated thoracic aortas. The contraction induced by 60 mM KCl was taken as 100%. The relaxation induced by sodium nitroprusside (10−4 M) was taken as 100%. Each point represents the mean ± SEM for 6 snakes. (⁎P < 0.05 vs. control).
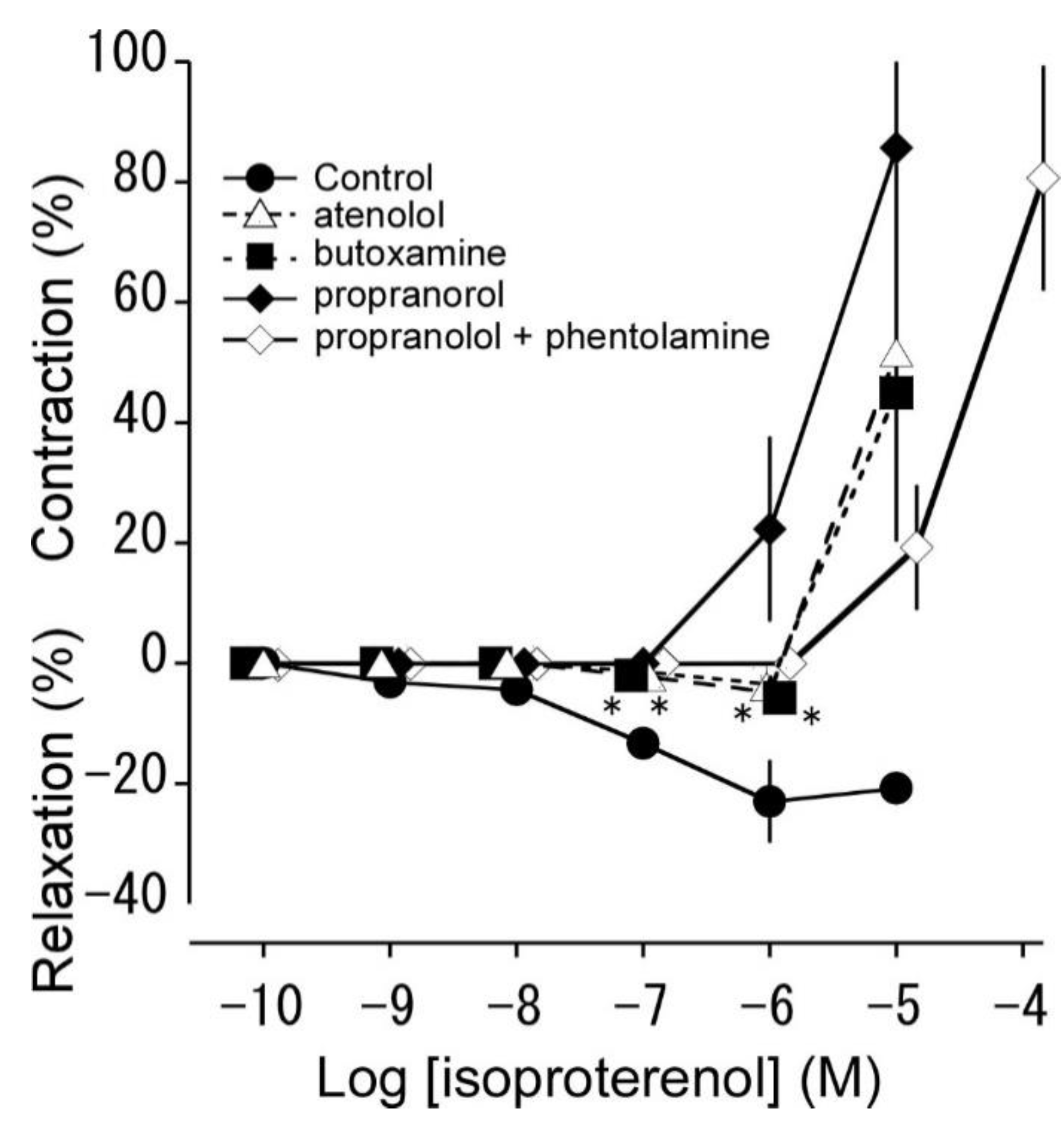
Figure 5.
Effects of SB269970, methiothepin, ketanserin and L-NNA on serotonin-induced relaxation or contraction. (A): Effect of SB269970 (◊: 10−7 M, n = 6), ketanserin (∇: 10−6 M, n = 6), methiothepin (×: 10−8 M, n = 6), L-NNA (□: 10−4 M, n = 6), L-NNA (10−4 M) + SB269970 (10−7 M) + methiothepin (10−8 M)) (Δ, n = 4), L-NNA(10−4 M) + SB269970 (10−7 M) + ketanserin (10−6 M) (○, n = 4) on serotonin-induced relaxation (●: Control, n = 12). The relaxation induced by 10−4 M sodium nitroprusside was taken as 100%. Each point represents the mean ± SEM. *P < 0.05 compared with the value of Control. (B): Effects of methiothepin (10−8 M) and ketanserin (10−6 M) on serotonin-induced contraction (black: 10−5 M, Control). The contraction induced by 60 mM KCl was taken as 100%. Each bar represents the mean ± SEM for 6 snakes. **P < 0.01 compared with the value of Control.
Figure 5.
Effects of SB269970, methiothepin, ketanserin and L-NNA on serotonin-induced relaxation or contraction. (A): Effect of SB269970 (◊: 10−7 M, n = 6), ketanserin (∇: 10−6 M, n = 6), methiothepin (×: 10−8 M, n = 6), L-NNA (□: 10−4 M, n = 6), L-NNA (10−4 M) + SB269970 (10−7 M) + methiothepin (10−8 M)) (Δ, n = 4), L-NNA(10−4 M) + SB269970 (10−7 M) + ketanserin (10−6 M) (○, n = 4) on serotonin-induced relaxation (●: Control, n = 12). The relaxation induced by 10−4 M sodium nitroprusside was taken as 100%. Each point represents the mean ± SEM. *P < 0.05 compared with the value of Control. (B): Effects of methiothepin (10−8 M) and ketanserin (10−6 M) on serotonin-induced contraction (black: 10−5 M, Control). The contraction induced by 60 mM KCl was taken as 100%. Each bar represents the mean ± SEM for 6 snakes. **P < 0.01 compared with the value of Control.
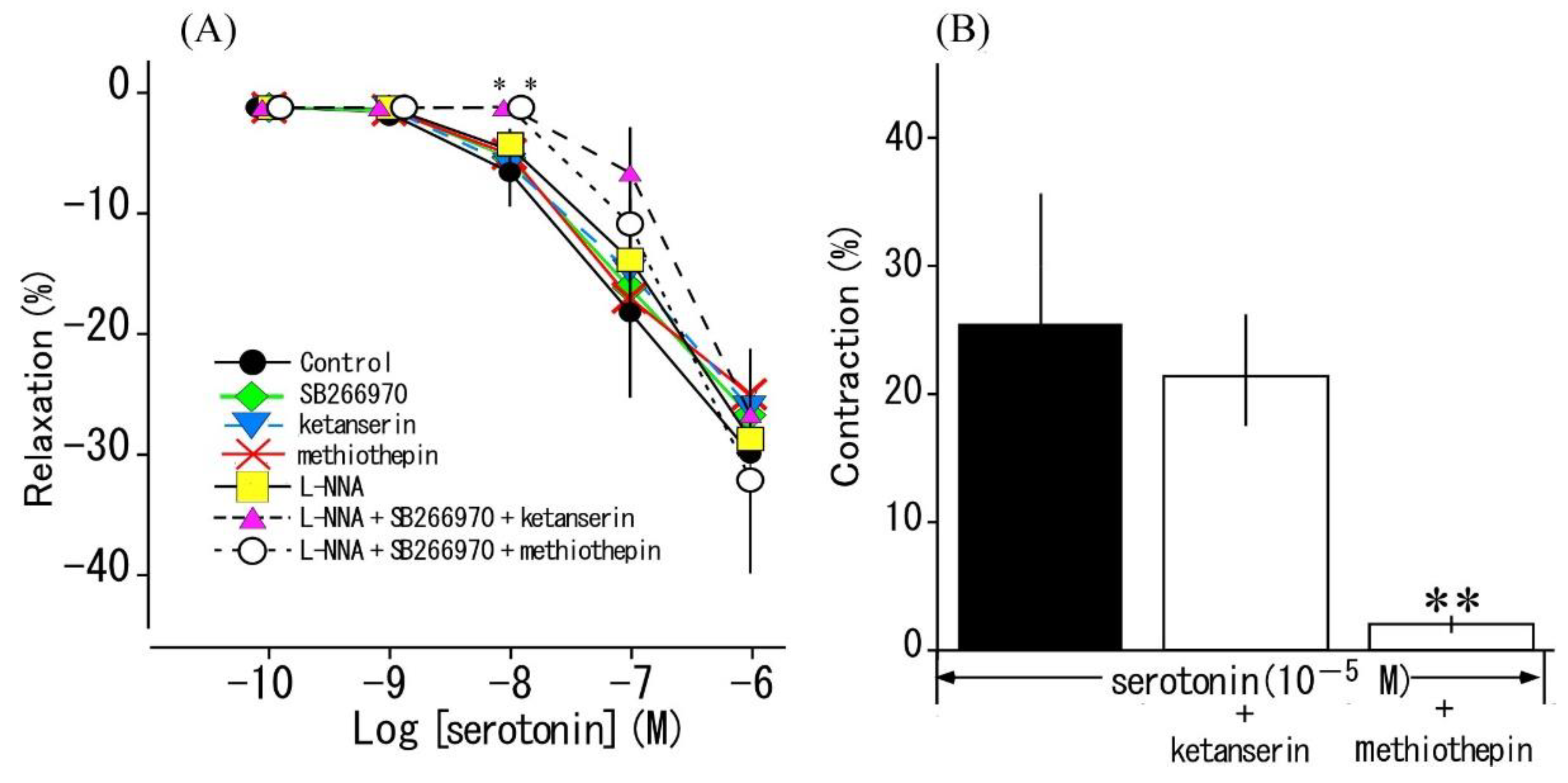
Figure 6.
Effects of atropine (10−6 M, a non-selective muscarine receptor antagonist), pirenzepine (10−6 M, an M1 receptor antagonist), methoctramine (10−6 M, an M2 receptor antagonist), pfHHSiD (10−6 M, an M3 receptor antagonist) and L-NNA (10−4 M) on acetylcholine-induced relaxation (black: 10−7 M) in thoracic aortas in resting tension. The relaxation induced by sodium nitroprusside (10−4 M) was taken as 100%. Each bar represents the mean ± SEM for 6 snakes.
Figure 6.
Effects of atropine (10−6 M, a non-selective muscarine receptor antagonist), pirenzepine (10−6 M, an M1 receptor antagonist), methoctramine (10−6 M, an M2 receptor antagonist), pfHHSiD (10−6 M, an M3 receptor antagonist) and L-NNA (10−4 M) on acetylcholine-induced relaxation (black: 10−7 M) in thoracic aortas in resting tension. The relaxation induced by sodium nitroprusside (10−4 M) was taken as 100%. Each bar represents the mean ± SEM for 6 snakes.
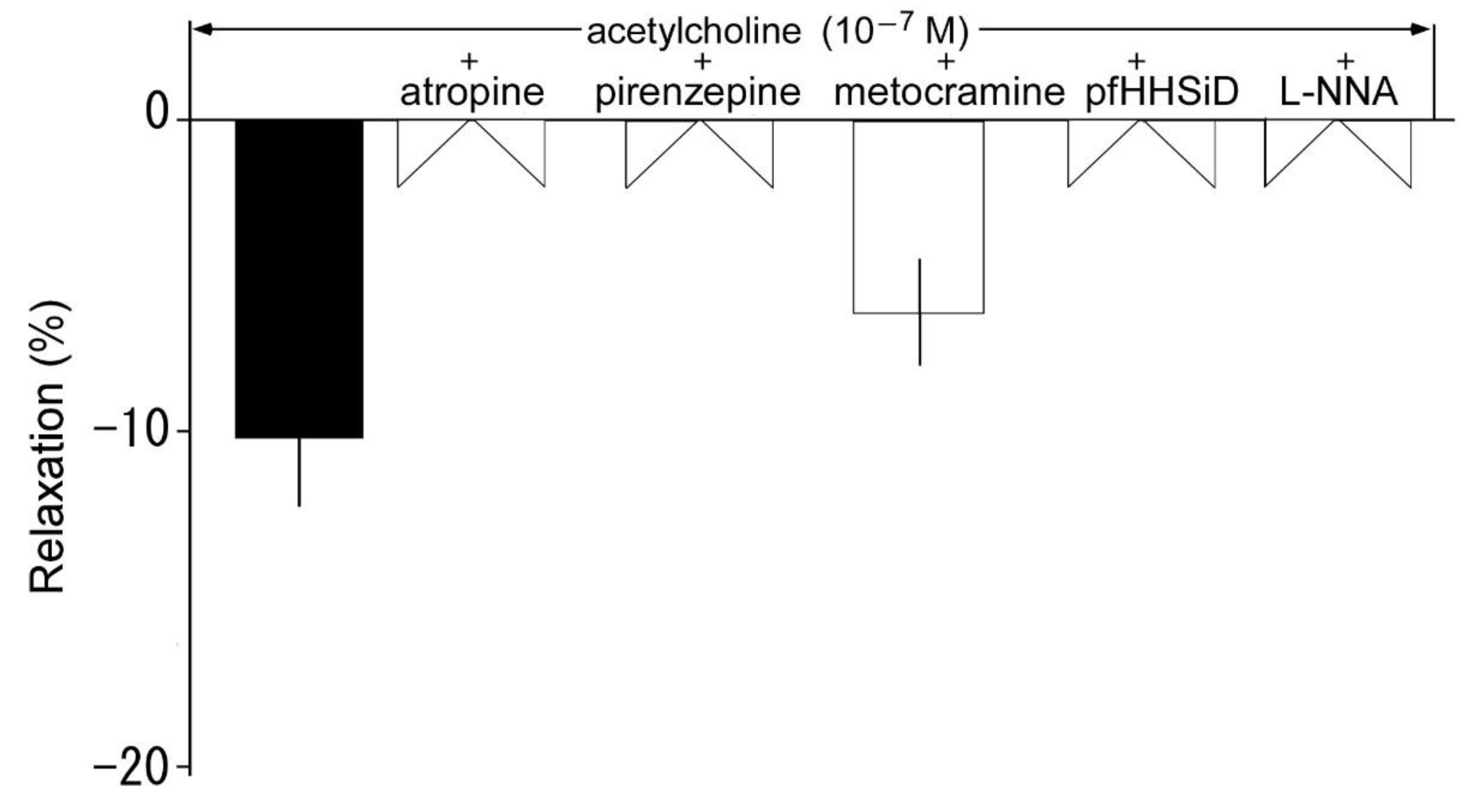
Figure 7.
Effects of atropine (10−6 M, a non-selective M receptor antagonist), pirenzepine (10−6 M, an M1 receptor antagonist), methoctramine (10−6 M, an M2 receptor antagonist), and pfHHSiD (10−6 M, an M3 receptor antagonist) on acetylcholine-induced contraction (black: 10−5 M, Control) in Tokara habu thoracic aortas in the presence of L-NNA (10−4 M). The contraction induced by 60 mM KCl was taken as 100%. Each bar represents the mean ± SEM for 5 snakes. **P < 0.01 compared with the value of Control.
Figure 7.
Effects of atropine (10−6 M, a non-selective M receptor antagonist), pirenzepine (10−6 M, an M1 receptor antagonist), methoctramine (10−6 M, an M2 receptor antagonist), and pfHHSiD (10−6 M, an M3 receptor antagonist) on acetylcholine-induced contraction (black: 10−5 M, Control) in Tokara habu thoracic aortas in the presence of L-NNA (10−4 M). The contraction induced by 60 mM KCl was taken as 100%. Each bar represents the mean ± SEM for 5 snakes. **P < 0.01 compared with the value of Control.

Figure 8.
Nitric oxide (NO) production ([NO2−] + [NO3−]) induced by serotonin (10−6 M), acetylcholine (10−7 M), and isoproterenol (10−6 M), and the effects of L-NNA (10−4 M) on the resultant NO production. Control: no treatment. Each column represents the mean ± SEM of arteries from 10 snakes. (⁎P < 0.05 vs. control).
Figure 8.
Nitric oxide (NO) production ([NO2−] + [NO3−]) induced by serotonin (10−6 M), acetylcholine (10−7 M), and isoproterenol (10−6 M), and the effects of L-NNA (10−4 M) on the resultant NO production. Control: no treatment. Each column represents the mean ± SEM of arteries from 10 snakes. (⁎P < 0.05 vs. control).
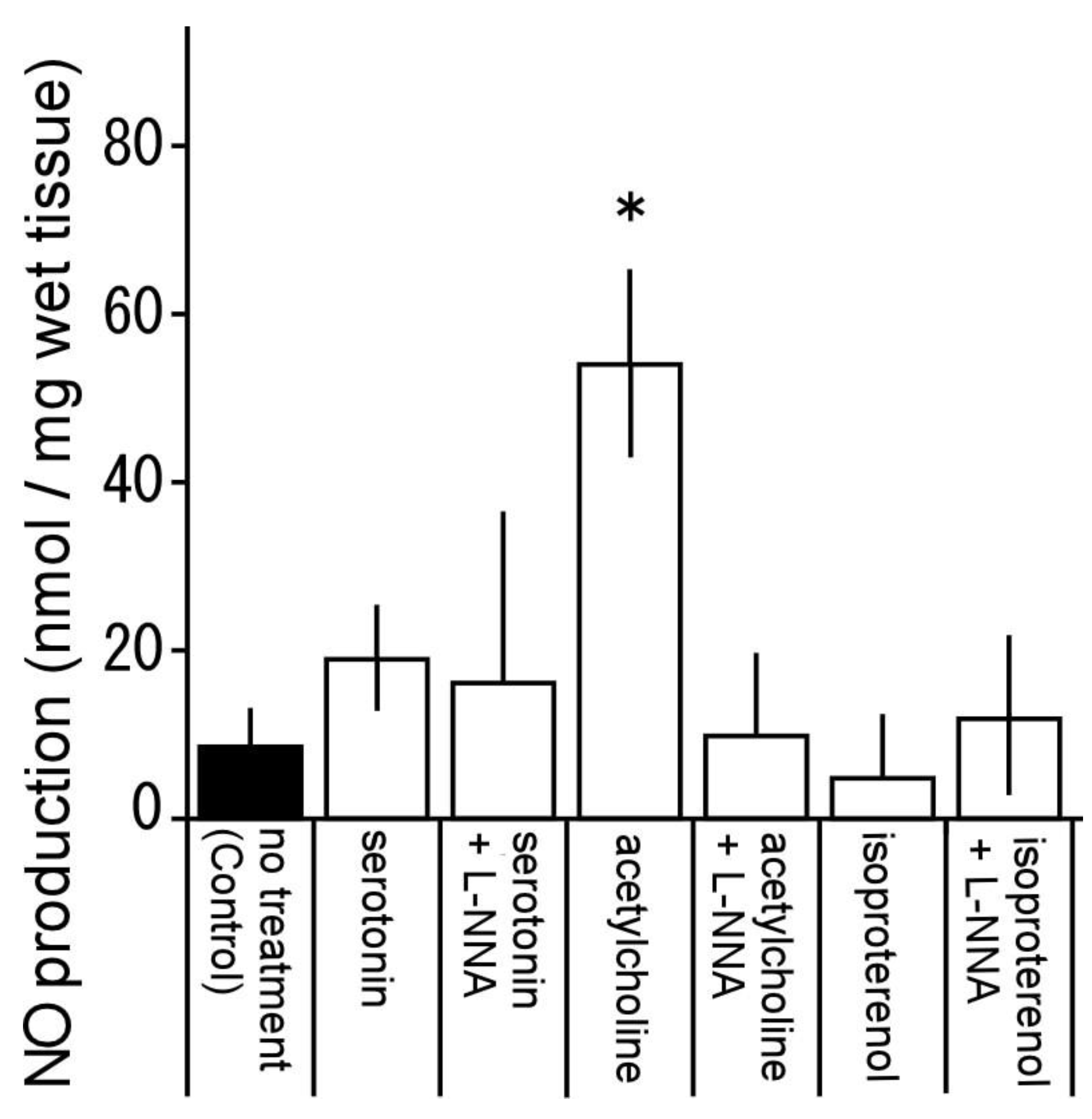

Table 1.
pEC50 values and maximal response (Emax) to agonists.
| Agonists | pEC50 | Emax (%) (reactivity) |
|---|---|---|
| Resting condition (2.4 mN) | ||
| Noradrenaline | 6.04 ± 0.14 | 130.2 ± 13.0 (contraction a) |
| Angiotensin II | 7.39 ± 0.06 | 46.4 ± 10.3 (contraction a) |
| Histamine | − | 0 (no response) |
| Rattlesnake bradykinin | − | 0 (no response) |
| Isoproterenol | 7.11 ± 0.07 | 22.9 ± 0.3 (relaxation b) |
| Serotonin | 7.41 ± 0.28 | 27.6 ± 4.9 (relaxation b) |
| Acetylcholine | 8.25 ± 0.19 | 10.2 ± 3.1 (relaxation b) |
a Contraction induced by 60 mM KCl was taken as 100%. b Relaxation induced by sodium nitroprusside (10−4 M) was taken as 100%.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



