
Submitted:
26 September 2023
Posted:
27 September 2023
You are already at the latest version
Abstract
Abstract: The species composition, the structure of phytoplankton communities, hydrochemical parameters in the artificial cooling reservoir of one of the largest thermal power plants (TPP) in the north-east of Europe in the Pechora River basin, Komi Republic were studied in the ice-open period in June and August. Altogether 81 species of algae and cyanobacteria was revealed with the last one predominated. Among cyanobacteria and algae (Bacillariophyta, Chlorophyta), indicators of water quality were found. Assessment of water quality according to the species composition and abundance of indicator phytoplankton species showed that the waters of the Pechorskoe Reservoir can be at-tributed to the III class of water quality, beta-mesosaprobic self-purification zone, with water quality of satisfactory purity, retaining the ability to self-purify. An increase in the diversity of plankton communities and high values of their abundance and biomass under the influence of the warming effect of TPP water outlet were noted and revealed its spatial distribution over the reservoir surface. The plankton of the northern reservoir under the influence of warm waters indicates a slight in-crease in the trophic status of the studied water body, which is also confirmed by the data of hydrochemical analysis. Ecological mapping for the chemical and biological variables as well as pollution indices over the water area of the reservoir was carried out. The results obtained can be used to make forecasts of changes in the phototrophic biota of small northern water bodies under the conditions of climate change.
Keywords:
phytoplankton
; diversity
; bioindicators
; water quality indices
; thermal impact
; Pechorskoe Reservoir
; European North
1. Introduction
Artificial reservoirs - reservoirs, cooling ponds, fishponds, recreational ponds in city parks - play a significant role in economic activity and have been used by mankind since ancient times [1]. The control of the main characteristics of these objects, primarily the physicochemical parameters of water and production indicators, is used to assess the state of water bodies and their conservation and is also the basis for managing the functioning of these water bodies [2]. Along with physicochemical parameters, data on the state of the reservoir and the processes occurring in it reflect the dynamics of plankton communities. Plankton is a key link in the functioning of aquatic ecosystems, forming the main part of the biomass of water bodies available for consumption by other aquatic organisms [3]. The dynamics of the composition and production characteristics of plankton communities in artificial reservoirs change under the influence of both natural and anthropogenic factors [4].
A long-term and versatile study of the patterns of functioning of plankton in res-ervoirs, including in connection with anthropogenic impact, on the territory of Russia was mainly carried out for Volga River Basin Reservoirs [5,6], reservoirs of Siberia on the Ob River [7] and Yenisei River [8]. Conducted a study of plankton in reservoirs and reservoirs-coolers of the Kama River basin [9,10,11] and Ural River basin [12], and Primorsky Krai [13]. The phytoplankton of the Dnieper reservoirs of Ukraine was studied in detail [14,15,16]. There are works on the influence of the temperature factor on the phytoplankton of reservoirs in Belarus [17], China [4,18,19,20], and Israel [21].
At the same time, there are few data on biota and the state of the aquatic environment in small reservoirs and ponds (water bodies with a volume of less than one million m3 that do not have permanent spillways). These reservoirs, despite their small volumes, nevertheless, are important both for the economic activity of people, per-forming the functions assigned to them (fish breeding, cooling, accumulation of water mass, recreation, biological treatment, etc.), and for the natural environment, participating in the regulation of the water regime of the territory, self-purification of aquatic ecosystems, and conservation of biological diversity [22]. In this regard, the study of the features of functioning, monitoring and assessment of the state of these ecosystems is especially relevant.
On the territory of the studies (in the Komi Republic) in the north-east of European Russia, artificial reservoirs are predominantly small. The largest of them, reservoirs, are located in its southern regions (in the Vychegda River basin); the planktonic com-munities of such reservoirs have been studied in sufficient detail [23]. Thanks to hydrobiological studies carried out in their water area in recent years, we have enough information about the composition and structure of the aquatic invertebrates inhabiting them, including plankton, as well as information about the composition of the leading phytoplankton groups [23].
The object of our research, the Pechorskoe Reservoir, is used as a cooling reservoir, which distinguishes it from other reservoirs in the study area. As a result of the inflow of heat-exchange water into the reservoir, its chronic thermal pollution occurs. The effect of heat load, as a rule, leads to an increase in the content of biogenic substances, and as a result, an increase in the trophic content of the reservoir. In reservoirs affected by heated waters, there is an increase in secondary pollution and an increase in the toxicity of harmful substances [24].
We have not been able to find information on the effect of thermal pollution on planktonic communities of small reservoirs in high northern latitudes, although the study of such communities is of interest not only for bioindication but is important for understanding the processes of formation and change of planktonic communities of small water bodies of the North under conditions of climate warming [25,26,27]. The plankton communities of the Pechorskoe Reservoir have not yet been studied.
Our task was to characterize with the phytoplankton communities the influence of hot water from a thermal power plant on the northern reservoir ecosystem.
2. Materials and Methods
2.1. Description of study site
The studies were carried out in the artificial reservoir - Pechorskoe, which is located in the basin of the middle reaches of the Pechora River, 3 km from Pechora town (Figure 1). The reservoir was created in 1984 on the site of a swamp massif. In the process of its creation, two lakes and adjacent swampy forest areas were flooded [28]. This is a small shallow-water reservoir of a bulk type with an area of about 600 hectares; its volume is about 33 million m3, the surface area is 5.74 km2 [28]. The maximum depth of the reservoir is 7.6 m. The coastline of the reservoir is slightly indented, the southern shore is reinforced with concrete slabs. Until 2014, the reservoir was filled with waters from the Pechora River, now the reservoir is fed by three forest streams with swampy watersheds and atmospheric precipitation. The total annual water consumption in the circulating water supply system of the reservoir is 502,697.15 m3 per year. The temperature difference at the inlet and outlet is 7.3°C on average for the year.
The water temperature at the spillway and the water area adjacent to it reaches 12−29°С in summer and 6−9°С in winter; no more than 30% of the area is covered with ice.
The period with an average water temperature above 10°C lasts from mid-April to the end of October, which is 2 months longer than in natural water bodies of this region [29].
2.2. Methods of Plankton Sampling and Hydrological Investigation
Hydrobiological and hydrochemical sampling at the Pechorskoe Reservoir in 2022 was carried out twice per season: June 20–21 (beginning of summer) and August 16–17 (end of summer) at three stations, at 8 observation points in the reservoir, evenly spaced along water area of the reservoir (Figure 1 and Figure 2; Table 1).
At each station, samples were taken in the center of the reservoir, as well as near the northern and southern shores of the reservoir. Calculation of average indicators for each station was performed by the sum of indicators for three samples. Samples were taken from the boat. A total of 17 hydrobiological samples were examined.
2.2.1. Hydrochemical samples
The water samples were taken with standard methods [31]. They were taken the water body surface in a volume of 4.5 L (glass packaging) and at a depth of 30–50 сm with a 2-L Ruttner bathometer of 1.5 L (polyethylene packaging). The samples were placed into a cooling bag immediately after collection. A quantitative chemical analysis of the water samples based on 26 variables was performed with metrologically attested methods in an ecoanalytical laboratory at IB KSC. Ecoanalytical laboratory IB FRC Komi SC UB RAS is conformed to requirements of ISO/IEC 17025:2017 (GOST ISO/IEC 17025-2019) and recorded to the Register of Accredited CABs under number РОСС RU.0001.511257. A more detailed description of the hydrochemical analysis methods used was published by us earlier [32].
2.2.2. Phytoplankton samples
Water (50 L) was sampled from the surface horizons for the phytoplankton study. The sample was concentrated with a plankton net (mill gas no. 77 was used) [31]. We studied live samples and samples fixed with formaldehyde solution (4%). Nikon Eclipse80i microscopes (equipped with a system of differential interference contrast and video recording of images) (Nikon, Japan) and Bimam-I microscopes (Lomo, Russia) were used to identify algae. Diatoms were identified in temporary, calcined, and permanent preparations prepared with Elyashev’s medium [31,33]. The main international keys were used for identification [31,34,35,36,37,38,39,40], and others with upgrade the taxa name according [41]. The abundance and biomass were quantified via direct cell counting with a Goryaev camera [31]. The mean and standard deviation of five replicates were calculated for each point in Excell Program. The first three species (up to five in the case of identical values) in the ranked series of abundances/biomasses were attributed to the dominants, and the following three (or four) species were assigned to the subdominants.
2.3. Index Saprobity Determination
The index saprobity of the aquatic environment was calculated based on a semi-quantitative method for the assessment of the abundance of phytoplankton species-indicators on a 9-point scale, which, in turn, was based on the relative (percentage) abundance of individual taxa [42]. The points indicate the following values of the copy number: 1 point, less than 1% of the total number counted; 2 points, 2–3%; 3 points, 4–10%; 5 points, 11–20%; 7 points, 21–40%; 9 points, 41–100%. The water sap-robity index was calculated according to Pantle-Buck as modified by Sládeček [43]: S = Σhs/Σh, where S is the saprobic index of phytoplankton community, s is the spe-cies-specific saprobic valence [43], and Σh is the sum of points on a 9-point scale. The ecological characteristics of the species are given according to the literature [42,44]. Bioindicator analysis was carried out with the help of species-specific ecological preferences of revealed algae and cyanobacteria [42,44,45]. Water quality classes are established on the basis of the complex ecological classification of surface waters [42,44].
2.4. Statistical Analysis
Statistical maps were constructed in Statistica 13.0 program on the base of parameter value and GIS coordinates of sampling stations [45]. The statistical mapping method was applied to visualize the distribution of the variables. Statistical maps can be constructed for all available chemical, biological indicators and calculated indices for reservoir. Maps are drawn for the reservoir surface and outlined by its contour, al-lowing us to draw into the analysis landscape features and the location of both points and diffuse influencing factors of the catchment basin. To clarify the revealed, difference in the composition and abundance of phytoplankton communities, a correlation analysis was carried out in the Program JASP [46].
3. Results
3.1. Characteristics of the hydrochemical parameters in investigated water bodies
Table 2 presents data on hydrochemical parameters of the station at which samples were taken. It can be seen that the conditions in the three stations in different sampling periods were quite similar. In general, the surface waters at all of the stations are characterized by a comparatively low mineralization, which is suggested by interrelated values of conductivity. Hydrogen-carbonate ions and calcium ions are dominant in the ion balance with a relatively low content of chloride and sulfate ions which corresponds to the hydrogen-carbonate water class. The active reaction of the water medium is neutral. The concentration of nitrate nitrogen, phosphates, ammonium, and other indicators were low (Table 2).
The difference between the water temperature of the current outlet of circulating water used in the technical water supply system of the thermal power plant (according to the data of its environmental laboratory) at the time of intake and discharge in June is 6–9 °C, in August 5–6 °C. Indicators of water temperature at the surface and at depth for the studied stations, as well as air temperature at the time of sampling are presented in Figure 3.
The water masses of the reservoir are noticeably affected by the wind effect (Figure 1b), as a result, intensive mixing and movement of water masses occurs. On the day of sampling in June, a northerly, northwesterly wind with an average speed of 4 m/s, gusts up to 10 m/s prevailed, and in August - a northwesterly, westerly wind with an average speed of 3 m/s, gusts up to 11 m/s (Figure 1 b). The wind directions these days generally corresponded to the average data of the wind rose for June and August (http://pogodaiklimat.ru).
3.2. Study of the Phytoplankton of Reservoir and the Indicators
In the phytoplankton of the Pechorskoe Reservoir in the summer of 2022, 11 species of cyanobacteria (Cyanobacteria) and 70 algae taxa from 7 phyla (81 taxa in total) were identified, of which 9 belonged to the Euglenozoa, 32 to Bacillariophyta, 3 to Miozoa (Dinophyceae), 2 to Ochrophyta, 18 to Chlorophyta, 6 to Charophyta (Table 3, Table Appendix A-1). Below is the characteristic of phytoplankton for two periods of research at observed stations.
In June, phytoplankton in the reservoir was represented by 55 species and forms of algae, in August by 51 taxa (Table 3, Table Appendix A-1). In both observation periods, the highest species richness was noted for diatoms, green algae, and cyanobacteria are in third place (Table 3, Figure 9a). The diversity of other discovered taxonomic groups is much lower. In August, the difference between divisions somewhat decreases due to a decrease in the number of diatom species and an increase in other groups (Figure 9a).
The distribution of species abundance at the stations is shown in the figure (Figure 4 a-h). In June, a greater number of species were recorded in the northern part of the reservoir, at the point of discharge of warm waters. A greater number of species of cyanobacteria, diatoms and green algae were noted there than in the middle and southern parts of the reservoir. The waters of the reservoir after the winter period do not heat up well enough, and during this period the inflow of heated waters has a more noticeable effect on the phytoplankton communities. By the end of summer in August, the temperature regime in the reservoir levels off and the differences in the distribution of species along the surface of the reservoir are not as obvious as at the beginning of the growing season. The number of diatom species continues to be the highest in the upper part of the reservoir, and the diversity of cyanobacteria and green algae in the zone of warm water discharge remains approximately the same as in June, and increases at other stations, their increase is most noticeable in the coastal zone (Figure 4 e-h).
The distribution of dominant species by stations was similar in both June and August. The basis of the dominant complex was formed by cyanobacteria, diatoms and green algae (Table 3). The absolute dominant in abundance both in June and August at all stations was the cyanobacteria Woronichinia naegeliana, the development of which in the reservoir reaches the stage of water “blooming” (Figure 5 a,b). The dominant complex also includes diatoms - Aulacoseira italica, Fragilaria crotonensis and green algae - Hindakia tetrachotoma, Pediastrum duplex (Figure 5 d-f). In August, the complex of dominants is supplemented by cyanobacteria – Aphanizomenon flos–aquae (Figure 5 e) and Planktothrix agardhii.
Abundance indicators of phytoplankton at the stations in different observation periods varied widely (Table 2). In June, the range of values was from 4.2 to 6.5 million cells L-1. In August, the figures were in the range of 2.3 to 7.7 million cells L-1. The maximum values were recorded in summer at st. 4 in the central part of the reservoir. The basis of abundance both in June and August at all stations is mainly formed by cyanobacteria, for representatives of other divisions, quantitative indicators are much lower (Table 3, Figure 6). The distribution of phytoplankton abundance in the water area of the reservoir was heterogeneous (Figure 6 a,e). The highest concentration of algae in June was recorded at st. 5 in the southwestern coastal part. As can be seen from the figure (Figure 6c), cyanobacteria prevail in phytoplankton in terms of quantitative indicators. Statistical analysis showed that the concentration of cyanobacteria in the central part of the reservoir was significantly higher than in the upper (p=0.037) and lower (p=0.042) parts of the reservoir. Representatives of Bacillariophyta (Figure 6b) in June are distributed relatively evenly over the entire area of the reservoir, except for a narrow band in the area of st. 8, which can be attributed to the error in the for-mation of a graphic display. Green algae in June are mainly concentrated at st. 1–3 in the upper part of the reservoir, at the place of warm water discharge (Figure 6d).
In August, analysis of the distribution of phytoplankton abundance showed a clear concentration of algae in the central and lower parts of the reservoir at st. 4–8 (Figure 6 e). Analysis of variance also confirmed the concentration of algae in the low-er part of the reservoir, where their number was significantly higher relative to the upper part (p=0.016). As can be seen in Figure 6f,g, in August, representatives of Bacillariophyta and cyanobacteria are concentrated in the central (p=0.003) and lower (p=0.003) parts of the reservoir. These groups determine the basis of phytoplankton abundance. The concentration of green algae cells (Figure 6h) is diametrically opposed to the two sections discussed above, the maximum values for this group of algae are noted at st. 3, 6 in the reservoir coastal zone.
Phytoplankton biomass in June was lower than in August (Table 3). In June, the range of values was from 0.39 to 1.31 mg L-1, in August 0.40 to 2.36 mg L-1. In June, the graphs of the projections of the biomass and abundance of plankton algae differ (Figure 6a andFigure 7a). The maximum biomass indicators were noted in the upper part of the reservoir at st. 3, with the concentration of algae in the central part at st. 5. The discrepancy between quantitative indicators is due to the fact that the main biomass in the plankton community in the upper part of the reservoir in June is formed by large-celled forms of diatoms. Total phytoplankton biomass and diatom biomass have a similar distribution (Figure 7a,b). While small-celled cyanobacteria form the basis of abundance, but are inferior to diatoms in terms of biomass (Table 2, Figure 7c). For August, the biomass graph is in good agreement with the graph of the total abundance of phytoplankton (Table 2, Figure 6e andFigure 7e), as well as the abundance, biomass in the central (st. 4 and st. 5) and lower (st. 7) parts of the reservoir noticeably higher than in the upper part (st. 1–3) (p=0.0021). The biomass indicators of individual groups of algae are consistent with the graphs of the distribution of the total biomass.
Figure 8 shows a JASP network diagram of correlations and it can be seen that the phytoplankton communities of the Pechorskoe Reservoir have separated from 3 clusters. The first cluster united stations 1–3, these are communities of sites experiencing the greatest impact of warm water discharge. In the second cluster, stations 4 and 7 are phytoplankton communities of the central part of the reservoir (in the deepest parts of the reservoir) and in the third cluster, plankton communities of the coastal zone of the reservoir have united.
3.3. Indicators of Habitat Conditions for Phytoplankton
The general appearance of the composition of indicator species in the Pechorskoe Reservoir was highly similar for both months (Table Appendix A-1). Figure 9, Figure 10 and Figure 11 show comparative histograms of bioindication results by the number of taxa and their environmental variables in the Pechorskoe Reservoir in June and August. Among organisms-indicators of ecological conditions, the largest number of species was noted among diatoms and green algae (Figure 9 a), in spring the number of indicator species is slightly higher.
The confinement to the type of habitat in the communities of reservoir in June and August was dominated by planktonic-benthic forms (Figure 9b), with a slight in-crease in planktonic species at the end of the summer season.
Algae indicators of temperature are dominated by types of moderate temperature regime or indifferent by algae preferring moderate temperatures (temp), in June their share is higher than in August (Figure 9c). There are few indicators of warm waters: one heat-loving species (warm) was noted with a low abundance, the diatom Ulnaria acus. Also, four eurythermal species (eterm), predominantly from Euglena, mainly develop in August with a small abundance. Among the cold-loving forms cold-loving species (cool), were recorded the diatom Aulacoseira italica was noted in the complex of dominants in all observation periods, and Aulacoseira italica var. tenuissima was noted among the subdominants.
Indicators of the hydrodynamic conditions (oxygen regime and the mobility of water masses) are mainly represented by in different species or species that prefer waters of moderate fluidity (st-str). The remaining groups are small (Figure 9d), but species that prefer stagnant waters are predominantly represented by dominants – Planktothrix agardhii, Woronichinia naegeliana, Hindakia tetrachotoma. The ratio of this groups remains the same in spring and summer periods.
Among the indicators of salinity, the largest number of species is noted among diatoms (Table Appendix A-1). The most represented were indifferent species (i), further in the ranking were salt-loving species (hl) (Figure 10a). Indifferent species were recorded in the composition of phytoplankton at all studied stations in both periods of research.
Water pH indicators in the Pechorskoe Reservoir show slightly more alkaline conditions the waters were neutral to slightly alkaline. Indicators of oligosaprobic waters formed about 68% of the abundance and were included in the dominant complexes. In August, the proportion of oligo-alpha-mesosaprobionts increased to 17%, indicating an increase in the trophic status of water bodies.
Indicators of oligosaprobic waters already accounted for about 94% of the abundance in August and were included in the dominant complexes. The share of indicator species of β-saprobic conditions accounted for about 3%.
Ecological saprobity groups according to Watanabe [47] are known for a third of diatom species. Of these, saprophytes predominated, among which were species of the dominant complex from the genus Aulacoseira. Saproxenes are rare, eurysaprobes are not noted. Among the trophic groups, eutrophens (e) predominate in both periods, half as much as meso-eutraphentes (em). These two groups include dominant species. Mesotraphentes (m) are represented by 2–3 species. A slight increase in trophicity in Au-gust may be indicated by a decrease in the number of oligo-mesotraphentes (o-m) (three species in June and one in August) and the appearance of the highly eutrophic water species Stephanodiscus minutulus (he).
3.4. Statistical Mapping
For visualization of the identified trends the method of statistical mapping was applied. For analysis, several maps were in a complex system of mutual influence of indicators in all station in the Pechorskoe Reservoir. For the current analysis, several maps were selected showing the distribution of the most critical indicators (Figure 12).
As can be seen on the maps, the maximum phytoplankton biomass in the Pechorskoe Reservoir in June was noted in the upper part of the reservoir, in the zone of influence of incoming warm water from the thermal power plant (Figure 12c), when the natural water temperature of the reservoir differs to a greater extent from the TPP water outlet temperature. In summer, the impact of the warming effect on the phytoplankton biomass is reduced due to the equalization of the water temperature at all stations (Table 1). The concentration of algae in the water area of the reservoir (Figure 12d) to a greater extent during this period is affected by wind mixing of water masses (Figure 3). In addition, at stations st. 1–4, along with the effect of the warming effect of thermal power plant effluents, the mass development of planktonic organisms can be influenced by organic pollution from the catchment area by the stream that feeds the reservoir. This is also indirectly indicated by an increase in the values of the S index in this part of the reservoir, especially in spring (Figure 12a,b). However, its values for the entire observation period at all stations were in the range of 1.72-1.82, which corresponds to class III water quality, beta-mesosaprobic self-purification zone, with water quality of satisfactory purity [43].
4. Discussion
In the phytoplankton of the Pechorskoe Reservoir in the summer of 2022, 81 taxa from 7 phyla were identified (Table 3, Table Appendix A-1). The highest species richness was noted for diatoms and green algae, cyanobacteria are in third place. Abundance and biomass fluctuated in the range of 2.3–7.7 million cells L-1 and 0.39–2.36 mg L-1 (Table 3). The diversity and quantitative indicators of phytoplankton communities are typical for northern lentic ecosystems in the study area [23,48]. In total, 976 taxa of algae from 10 divisions were found for the Pechora River basin in different types of water bodies [48]. About 300 species of algae and cyanobacteria were previously identified in small artificial reservoirs plankton communities in the study region, abundance and biomass for these water bodies fluctuated in the range of 0.23–5.48 million cells L-1 and 0.45–0.98 mg L-1 respectively [23]. The revealed biodiversity in the reservoir corresponds in composition to phytoplankton communities of aquatic ecosystems of the temperate zone with a near-neutral pH of the water environment [49,50].
The distribution of algae in the water surface of the Pechorskoe Reservoir is influenced by a whole range of natural factors, including wind surge, the presence of shallow waters, bank collapse, the influence of the streams joins the reservoir, as well as anthropogenic impact associated with the chronic inflow of heated waters. All this is reflected in the composition and structure of phytoplankton communities. Therefore, it is quite difficult to separately assess the impact of elevated water temperature on phytoplankton communities. It is well known that water temperature affects the rate of growth and reproduction of most planktonic algae, with the maximum growth rate being achieved by most phytoplankton at a water temperature of about 20°C [51].
An increase in water temperature above 25°C leads to an increase in the maxi-mum growth rate of cyanobacteria, as a result, they can outnumber other phytoplankton groups [51]. This explains the fact that under conditions of constant inflow of heated water into the reservoir, already at the very beginning of the growing season, at all stations of the Pechorskoe Reservoir, a high diversity and biomass of cyanobacteria, as well as diatoms and green algae, was noted (Table 3). In August, when the water warms up naturally throughout the water area, all indicators of the development of phytoplankton communities increase (Table 3). The influence of warm waters is also indicated by the presence of the heat-loving species Ulnaria acus and four eurythermal species in the phytoplankton (Figure 9, Table Appendix A-1). The most heat-loving species found are represented by Euglenozoa. They were noted mainly in August in different parts of the reservoir with a small abundance.
The influence of thermal pollution on the species of phytoplankton was also shown by JASP correlation analysis (Figure 8), algae communities of st. 1–3 united into a separate cluster. These are the communities of sites that are most affected by the release of warm waters. We also associate the dominance of the cyanobacteria Woronichinia naegeliana at all observation Stations with the influence of warm waters. The mass development of this species for northern water bodies, reaching the stage of “blooming” of water, is quite rare [52,53,54], but cyanobacteria blooming is known even in the permafrost zone water bodies [55]. Usually, a complex of dominants, especially at the beginning of the growing season, is formed in northern water bodies by Bacillariophyta [48,49,50]. Diatoms are known to prefer colder water and are abundant in temperate lakes and reservoirs in autumn and spring when water temperatures are cooler [56,57]. It should also be noted that in all the observed periods, Bacillariophyta predominates in terms of the number of species in the Pechorskoe Reservoir; in August, their number decreases, but remains quite high. While for many other reservoirs and cooling ponds, the predominance of Chlorophyta and an increase in the species rich-ness of Bacillariophyta in the autumn period are usually noted [10,11,49,50]. In general, it can be concluded that the discharge of heated waters does not significantly affect the composition and quantitative indicators of algae, since both species characteristic of cold waters and species of moderately warm waters are found in a reservoir with a high abundance.
Another factor that caused the difference in the distribution of quantitative indicators of algae in the Pechorskoe Reservoir is the strength and direction of the wind. During the sampling period in June 2022, a strong northern, northwestern wind with gusts up to 10 m/s was observed, which drove water masses to the central and south-western parts of the reservoir (st. 4–5, st. 7–8) (Figure 1). The wind surge of water masses did not have a noticeable effect on the species diversity and structure of the dominant complexes (Table 3, Figure 4). This factor primarily affected the total number of algae, which increased in the direction of the wind (Figure 6). An increase in biomass at st. 4–6 was observed only for cyanobacteria, with total algae biomass reaching a maximum in the northeastern part of the reservoir at st. 3 (Figure 7). This discrepancy in biomass is due to the fact that, against the background of the dominance of cyanobacteria, a noticeable development of planktonic-benthic diatoms and green algae was observed in the northeastern part, forming large massive colonies. This phenomenon was also noted by other authors for northern lakes [57].
The northern and northeastern shores of the Pechorskoe Reservoir crumble, the coastal part here is relatively shallow, streams flow into the reservoir in these places. All this probably leads to water agitation, its heating, an increase in nutrients, and, as a result, to the active development of plankton-benthic species. At the same time, a typ-ical plankter with gas vacuoles Woronichinia naegeliana although remains the undisputed dominant in the water area of the entire reservoir, is partly carried along with water masses during strong winds to the southwestern part of the Pechorskoe Reservoir at st. 4–8 (Figure 6g, Figure 7g). Similar patterns are observed in large reservoirs. For example, for Lake Baikal, the influence of the nature of the flooded water masses on the concentration of chlorophyll a was revealed [58], which increased or decreased depending on which waters were brought by one or another wind direction. For sea and ocean waters, the influence of the impact of mesoscale eddy dynamics on the bioproductivity of marine systems in terms of changes in abundance, biomass, and chlorophyll concentration is also noted, while the issue of the impact of eddy dynamics on the species composition of planktonic organisms has been poorly studied, at the moment it is difficult to reliably explain the principles of observed changes in species composition. composition of algae under the influence of eddy dynamics [59].
In August 2022, during the sampling period, the wind was blowing less intensely. The species diversity of phytoplankton communities was not associated with wind surge (Figure 4). The distribution of quantitative indicators of the total abundance and biomass of phytoplankton and individual groups of algae in the water area of the Pechorskoe Reservoir, as well as in June, can also be associated with the direction of the wind (Figure 6 e-h, Figure 7 e-h).
Species of the dominant phytoplankton complex of the Pechorskoe Reservoir are predominantly indifferent in relation to salinity and pH, which is typical for northern water bodies [48,49,50,57,60] in contrary of the southern reservoirs where green algae prevail [16,19,20]. The distribution of these indicator groups at different stations of the reservoir also did not differ significantly, which indicates the relative homogeneity of hydrochemical conditions at different stations (Table 2). The trophic level of the Pechorskoe Reservoir according to algae indicators corresponds to the mesotrophic type [43]. The values of the Saprobity index (Figure 12a,b) indicate that the ecosystem is still successfully coping with both allochthonous and autochthonous organic pollution.
5. Conclusions
In first time algological studies carried out in the small northern reservoir, which is under the influence of chronic thermal effects, have shown that phytoplankton communities at the present stage of development of the reservoir do not undergo noticeable structural and qualitative changes. Assessment of water quality according to the species composition and abundance of indicator phytoplankton species showed that the waters of the Pechorskoe Reservoir can be attributed to the III class of water quality, beta-mesosaprobic self-purification zone, with water quality of satisfactory purity, retaining the ability to self-purify. Based on the data obtained, it can be concluded that the reservoir ecosystem at the present stage of its development is a stable self-healing and self-regulating system, despite the anthropogenic impact of a thermal power plant. The main groups of taxonomic and indicators distribution with the eco-logical mapping help us to reveal the factors of the wind direction and temperature as most important for forming stability and self-purification ability of the Pechorskoe Reservoir ecosystem.
The results obtained can be used to make forecasts of changes in the phototrophic biota of small northern water bodies under the conditions of climate change.
Author Contributions
Conceptualization, E.P., S.B.; methodology S.B., E.P. and M.S.; sample collection E.P. and M.S.; formal analysis all authors; investigation all authors; data curation all authors; writing – original draft preparation all authors; writing – review and editing E.P., S.B. and J.S.; visualization S.B., E.P. and M.S.; supervision and project administration E.P., S.B.; funding acquisition E.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the research was carried out within the framework of the state tasks IB FRC Komi SC UB RAS №122040600026-9.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Appendix A
Table Appendix A-1: Ecological preferences of phytoplankton species of the Pechorskoe Reservoir in June-August 2022.

Table A1.
Ecological preferences of phytoplankton species of the Pechorskoe Reservoir in June-August 2022.
Table A1.
Ecological preferences of phytoplankton species of the Pechorskoe Reservoir in June-August 2022.
| No. | Taxon | June | August | HAB | T | OXY | HAL | pH | D | SAP | Index S | AUT-HET | TRO |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bacillariophyta | |||||||||||||
| 1 | Acanthoceras zachariasii (Brun) Simonsen | 1 | 1 | P | - | st-str | i | ind | - | o-b | 1.4 | - | - |
| 2 | Achnanthidium minutissimum (Kützing) Czarn. | 1 | 1 | P-B | eterm | st-str | i | ind | es | b | 0.95 | ate | e |
| 3 | Asterionella formosa Hassall | 1 | 1 | P | temp | st-str | i | alf | sx | b | 1.35 | ate | me |
| 4 | Aulacoseira granulata (Ehrenberg) Simonsen | 1 | 1 | P-B | temp | st-str | i | alf | es | b | 2.0 | ate | e |
| 5 | Aulacoseira italica (Ehrenberg) Simonsen | 1 | 1 | P-B | cool | st-str | i | ind | es | b | 1.45 | ate | me |
| 6 | Aulacoseira italica var. tenuissima (Grunow) Simonsen | 1 | 1 | P-B | cool | st-str | i | ind | es | b | 1.3 | ate | me |
| 7 | Diatoma moniliformis (Kützing) D.M.Williams | 1 | 1 | P-B | temp | st-str | i | alf | - | x-o | 0.4 | - | - |
| 8 | Diatoma tenuis C.Agardh | 1 | 0 | P-B | temp | st-str | hl | alf | - | b-a | 2.4 | - | om |
| 9 | Encyonema cespitosum Kützing | 1 | 0 | B | temp | st | i | alf | es | o-b | 1.4 | - | - |
| 10 | Encyonema silesiacum (Bleisch) D.G.Mann | 1 | 1 | B | temp | st-str | i | ind | - | - | - | - | - |
| 11 | Entomoneis ornata (Bailey) Reimer | 0 | 1 | B | - | st-str | i | alf | - | b | 2.0 | hne | - |
| 12 | Epithemia sorex Kützing | 1 | 0 | B | temp | st-str | i | alf | - | - | - | - | - |
| 13 | Fragilaria crotonensis Kitton | 1 | 1 | P-B | temp | st-str | i | alf | - | - | - | - | - |
| 14 | Gomphonella olivacea (Hornemann) Rabenhorst | 1 | 0 | B | temp | st-str | i | alf | - | b | 2.3 | ate | om |
| 15 | Gomphonema acuminatum Ehrenberg | 1 | 0 | B | temp | st-str | i | ind | - | x-b | 0.8 | - | - |
| 16 | Gomphonema truncatum Ehrenberg | 1 | 0 | B | temp | st-str | i | ind | - | b | 2.0 | - | - |
| 17 | Gyrosigma acuminatum (Kützing) Rabenhorst | 1 | 1 | B | temp | st-str | i | alf | - | - | - | - | - |
| 18 | Iconella splendida (Ehrenberg) Ruck & Nakov | 0 | 1 | P-B | - | st-str | i | alf | - | - | - | - | - |
| 19 | Luticola mutica (Kützing) D.G.Mann | 1 | 1 | B,S | temp | st-str | hl | ind | - | o-a | 1.9 | ats | e |
| 20 | Navicula cryptotenella Lange-Bertalot | 1 | 0 | P-B | temp | st-str | i | ind | - | - | - | - | - |
| 21 | Navicula reinhardtii (Grunow) Grunow | 1 | 0 | - | - | - | - | - | - | - | - | - | - |
| 22 | Neidiomorpha binodis (Ehrenberg) M.Cantonati, Lange-Bertalot & N.Angeli | 0 | 1 | B | - | str | i | alf | - | o | 1.0 | - | - |
| 23 | Nitzschia frustulum (Kützing) Grunow | 1 | 0 | P-B | temp | st-str | hl | alf | es | a-o | 2.7 | - | - |
| 24 | Nitzschia gracilis Hantzsch | 1 | 0 | P-B | temp | st-str | i | ind | - | - | - | - | - |
| 25 | Nitzschia sigma (Kützing) W.Smith | 1 | 0 | B | temp | st-str | mh | alf | - | - | - | - | - |
| 26 | Rhopalodia constricta (Brébisson) Krammer | 1 | 0 | B | - | st-str | hl | alf | - | - | - | ats | e |
| 27 | Stephanodiscus hantzschii Grunow | 1 | 1 | P | temp | st-str | i | alf | sx | - | - | - | - |
| 28 | Stephanodiscus minutulus (Kützing) Cleve & Möller | 0 | 1 | P | temp | st-str | i | alb | es | a-o | 3.6 | hne | he |
| 29 | Surirella gracilis Grun. | 0 | 1 | B | - | st-str | i | ind | - | - | - | - | - |
| 30 | Tabellaria fenestrata (Lyngb.) Kütz. | 0 | 1 | P-B | - | st-str | i | ind | - | o-a | 1.9 | - | - |
| 32 | Ulnaria acus (Kützing) Aboal | 1 | 1 | P-B | warm | st-str | i | alf | es | o-a | 1.85 | ate | me |
| 31 | Ulnaria ulna (Nitzsch) Compère | 1 | 0 | P-B | temp | st-str | i | alf | es | b-a | 2.4 | ate | e |
| Charophyta | |||||||||||||
| 33 | Closterium sp. | 1 | 0 | - | - | - | - | - | - | - | - | - | - |
| 34 | Cosmarium bioculatum var. depressum (Schaarschmidt) Schmidle | 0 | 1 | P-B | - | st-str | hb | ind | - | x-o | 0.5 | - | m |
| 35 | Mougeotia sp. ster. | 0 | 1 | - | - | - | - | - | - | - | - | - | - |
| 36 | Spirogyra sp. ster. | 0 | 1 | P-B | - | - | - | alf | - | - | - | - | - |
| 37 | Spondylosium planum (Wolle) West & G.S.West | 0 | 1 | P-B | - | - | i | ind | - | - | - | - | - |
| 38 | Staurodesmus triangularis (Lagerheim) Teiling | 0 | 1 | P | - | - | i | acf | - | - | - | - | - |
| Chlorophyta | |||||||||||||
| 39 | Chlamydocapsa planctonica (West & G.S.West) Fott | 1 | 1 | P-B | - | - | - | alf | - | o | 1.2 | - | - |
| 40 | Coelastrum astroideum De Notaris | 0 | 1 | P | - | st-str | - | - | - | b | 2.2 | - | e |
| 41 | Coelastrum microporum Nägeli | 1 | 1 | P-B | - | st-str | i | ind | - | b | 2.3 | - | e |
| 42 | Desmodesmus communis (E.Hegewald) E.Hegewald | 1 | 1 | - | - | - | - | - | - | - | - | - | - |
| 43 | Eudorina elegans Ehrenberg | 0 | 1 | P | - | st-str | i | - | - | b | 2.3 | - | - |
| 44 | Eutetramorus globosus Walton | 1 | 1 | - | - | - | - | - | - | - | - | - | - |
| 45 | Hindakia tetrachotoma (Printz) C.Bock, Pröschold & Krienitz | 1 | 1 | P | - | st | i | - | - | b | 2.3 | - | - |
| 46 | Lemmermannia tetrapedia (Kirchner) Lemmermann | 0 | 1 | P-B | - | st-str | i | ind | - | b | 2.0 | - | e |
| 47 | Oedogonium sp. | 1 | 0 | - | - | - | - | - | - | - | - | - | - |
| 48 | Pandorina morum (O.F.Müller) Bory | 1 | 1 | P | - | st | i | - | - | - | - | - | m |
| 49 | Pediastrum duplex Meyen | 1 | 1 | P | - | st-str | i | ind | - | - | - | - | e |
| 50 | Pseudopediastrum boryanum (Turpin) E.Hegewald | 1 | 1 | P-B | - | st-str | i | ind | - | b | 2.1 | - | e |
| 51 | Pseudopediastrum boryanum var. longicorne (Reinsch) Tsarenko | 1 | 1 | P-B | - | st-str | - | - | - | b | 2.1 | - | e |
| 52 | Scenedesmus ellipticus Corda | 1 | 1 | P-B, S | - | st-str | - | - | - | b-o | 1.7 | - | - |
| 53 | Stauridium tetras (Ehrenberg) E.Hegewald | 1 | 1 | P-B | - | st-str | i | ind | - | - | - | - | om |
| 54 | Tetradesmus lagerheimii M.J.Wynne & Guiry | 1 | 0 | P-B | - | st-str | i | ind | - | b | 2.15 | - | e |
| 55 | Thelesphaera alpina Pascher | 1 | 1 | B | - | str | - | - | - | - | - | - | - |
| 56 | Ulothrix zonata (F.Weber & Mohr) Kützing | 1 | 0 | P-B | - | st-str | i | ind | - | o-a | 1.8 | - | - |
| Cyanobacteria | |||||||||||||
| 57 | Aphanizomenon flos-aquae Ralfs ex Bornet & Flahault | 1 | 1 | P-B | - | - | hl | alb | - | o-a | 1.95 | - | m |
| 58 | Aphanothece sp. | 0 | 1 | - | - | - | - | - | - | - | - | - | - |
| 59 | Calotrix sp. | 1 | 1 | - | - | - | - | - | - | - | - | - | - |
| 60 | Chroococcus minimus (Keissler) Lemmermann | 0 | 1 | P-B | - | - | hl | - | - | - | - | - | e |
| 61 | Dichothrix ramenskii Elenkin | 1 | 0 | - | - | - | - | - | - | - | - | - | - |
| 62 | Dolichospermum smithii (Komárek) Wacklin, L.Hoffmann & Komárek 2009 | 0 | 1 | P | - | - | - | - | - | - | - | - | - |
| 63 | Leptolyngbya sp. | 1 | 0 | - | - | - | - | - | - | - | - | - | - |
| 64 | Microcystis aeruginosa (Kützing) Kützing | 1 | 1 | P-B | - | - | hl | acf | - | b | 2.2 | - | me |
| 65 | Oscillatoria sp. | 1 | 1 | - | - | - | - | - | - | - | - | - | - |
| 66 | Planktothrix agardhii (Gomont) Anagnostidis & Komárek | 1 | 1 | P-B | - | st | hl | - | - | - | - | - | - |
| 67 | Woronichinia naegeliana (Unger) Elenkin | 1 | 1 | P | - | st | - | - | - | o-a | 1.8 | - | e |
| Euglenozoa | |||||||||||||
| 68 | Astasiidae cf. | 1 | 0 | - | - | - | - | - | - | - | - | - | - |
| 69 | Euglena sp. | 0 | 1 | - | - | - | - | - | - | - | - | - | - |
| 70 | Euglena texta (Dujardin) Hübner | 0 | 1 | P | eterm | st-str | - | ind | - | a-o | 2.9 | - | - |
| 71 | Phacus sp. | 1 | 1 | - | - | - | - | - | - | - | - | - | - |
| 72 | Trachelomonas hispida (Perty) F.Stein | 0 | 1 | P-B | eterm | st-str | i | acf | - | b | 2.2 | - | - |
| 73 | Trachelomonas ssp. | 1 | 0 | - | - | - | - | - | - | - | - | - | - |
| 74 | Trachelomonas volvocina (Ehrenberg) Ehrenberg | 0 | 1 | P-B | eterm | st-str | i | ind | - | b | 2.0 | - | - |
| Miozoa | |||||||||||||
| 75 | Ceratium hirundinella (O.F.Müller) Dujardin | 1 | 1 | P | - | st-str | i | - | - | o | 1.3 | - | e |
| 76 | Peridiniaceae sp. | 0 | 1 | - | - | - | - | - | - | - | - | - | - |
| 77 | Peridiniopsis quadridens (F.Stein) Bourrelly | 1 | 1 | P | - | - | - | - | - | o-b | 1.4 | - | - |
| Ochrophyta | |||||||||||||
| 78 | Dinobryon sertularia Ehrenberg | 1 | 1 | P-B | - | - | i | - | - | o | 1.3 | - | - |
| 79 | Tribonema vulgare Pascher | 0 | 1 | P-B | - | - | i | - | - | o-b | 1.4 | - | - |
Note: “0” – not recognized. Abbreviation of ecological groups. Habitat (HAB): P—planktonic; P–B—plankto-benthic; B—benthic; S—soil. Temperature (T): cool—cool water; temp—temperate temperature; eterm—eurythermic; warm— thermophilic. Oxygenation and water moving (OXY): st—standing water; st-str—low streaming water, str – fast mowing water. Salinity ecological groups according to Hustedt (1938–1939) [61] (HAL): hb – oligohalobes-halophobes, i – oligohalobes-indifferent, hl – halophiles; mh – mesohalobes. Water pH preferences groups according to Hustedt (1957) [62] (pH): alf—alkaliphiles; ind—indifferent; acf—acidophiles; alb—alkalibiontes. Organic pollution indicators according to Watanabe (D) [47]: sx—saproxenes; es—eurysaprobes, sp – saprophiles. Saprobity groups according to Sládeček [43] (SAP): o—oligosaprob; o–b—oligo–beta–msosaprob; b–o—beta–oligosaprob; o–a—oligo–alpha–mesosaprob; b—beta–mesosaprob; b–a—beta–alpha–mesosaprob; a-o—alpha-oligosaprob; x-b, xeno-beta-mesosaprobiont; x-o, xeno-oligosaprobiont. Index S – Species-specific index saprobity. Nutrition type as nitrogen uptake metabolism (Van Dam et al., 1994) [63] (AUT-HET): ate—nitrogen–autotrophic taxa; ats—tolerating elevated concentrations of organically bound nitrogen; hne—facultatively nitrogen–heterotrophic taxa, needing periodically elevated concentrations of organically bound nitrogen. Trophic state indicators (Van Dam et al., 1994) [63] (TRO): om—oligo–mesotraphentic; m—mesotraphentic; me—meso–eutraphentic; e—eutraphentic; he—hypereutraphentic.
References
- Protasov, A. A. Several aspects of application and optimization of EU Water Framework Directive approaches in view of assessment of ecological state of technoecosystem. Hydrobiol. J. 2018, 54, 55–68. [Google Scholar] [CrossRef]
- Uzunov, Y. I.; Protasov, A. A. Concept of ecosystem services as applied to water technoecosystems. Hydrobiol. J. 2019, 55, 3–17. [Google Scholar] [CrossRef]
- Field, C. B.; Behrenfeld, M. J.; Randerson, J. T.; Falkowski, P. Primary production of the biosphere: integrating terrestrial and oceanic components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Luo, J.; Xing, L.; Yu, X.; Zhang, J.; Chen, S. Effects of temperature–control curtain on algae biomass and dissolved oxygen in a large stratified reservoir: Sanbanxi Reservoir case study. J. of environmental management 2019, 248, 109250. [Google Scholar] [CrossRef]
- Korneva L., G. Phytoplankton of Volga River Basin Reservoirs; Kostromskoy pechatniy dom: Kostroma, Russia, 2015; p. 284. (In Russian) [Google Scholar]
- Kopylov, A. I.; Lazareva, V. I.; Mineeva, N. M.; Zabotkina, E. A. Planktonic community of a large eutrophic reservoir during a period of anomalously high-water temperature. Inland Water Biology 2020, 13, 339–348. [Google Scholar] [CrossRef]
- Kuksn, M. S.; Chaikovskaya T., S. Interannual fluctuations in the species composition and biomass of phytoplankton in the Novosibirsk Reservoir. In Comprehensive studies of the Novosibirsk reservoir. Gidrometeoizdat: Moscow, Russia, 1985; pp. 76-84. (In Russian).
- Gold, Z.G.; Anufrieva, T.N.; Muchkina, E.Y.; Kozhevnikova, N.A. et al. The dynamics of structural characteristics of the ecosystem in the Krasnoyarsk deep-water reservor (1977–1999). In Biodiversity and dynamics of ecosystems in North Eurasia: Int. Biodiversity Observ. Year, Novosibirsk, Russia, 21–26 Aug. 2000. IC&G: Novosibirsk, Russia, 2000, 5; pp. 125–127.
- Mukhutdinov, V. F.; Butakova, E. A. Production characteristics of phytoplankton in early years of the existence of Yumaguzin Reservoir. Inland Water Biology 2012, 5, 317–321. [Google Scholar] [CrossRef]
- Belyaeva, P. G. The dynamics of the composition and structure of the phytoplankton reservoir-cooler of the Karmanovsk State District Power Station (Bashkiria). Bot. Zhurn. 2018, 104, 1698–1711. (In Russian) [Google Scholar] [CrossRef]
- Belyaeva, P. G. Interannual dynamics of the composition and distribution of phytoplankton in the Votkinsk reservoir (Kama basin, Perm Territory). Bot. Zhurn. 2018, 103, 297–312. (In Russian) [Google Scholar] [CrossRef]
- Dzhaian, E. A. Interannual Сhanges in Phytoplankton of the Iriklinsky Reservoi. Inland Water Biology 2020, 13, 549–555. [Google Scholar] [CrossRef]
- Goncharov, A. A. Algoflora of the Primorsky Cooling Reservoir (Primorsky Krai). Bot. Zhurn. 1996, 81, 32–40. (In Russian) [Google Scholar]
- Oksiyuk, O. P.; Davydov, O. A.; Melenchuk, G. V. Formation of phytoplankton species diversity in the river sections of the Dnieper reservoirs. Algology 2005, 15, 78–85. (In Russian) [Google Scholar]
- Novoselova, T. N.; Protasov, A. A. Phytoplankton of cooling ponds of techno–ecosystems of nuclear and thermal power stations (a review). Hydrobiol. J. 2015, 51, 37–52. [Google Scholar] [CrossRef]
- Novoselova, T.; Barinova, S.; Protasov, A. Phytoplankton Indicators in the Assessment of the Ecological Status of Two Reservoirs with Different Purposes in Southern Ukraine. Ecologies 2022, 3, 96–119. [Google Scholar] [CrossRef]
- Genkal, S. I.; Kulikovskiy, M. S.; Mikheeva, T. M.; Kuznetsova, I. V.; Lukyanova, E.V. Plankton diatoms of the Svisloch River and its reservoirs; Nauchniy mir: Moscow, Russia, 2013; p. 236. ISBN 978-5-91522-357-7. (In Russian) [Google Scholar]
- Xu, Y.; Cai, Q.; Ye, L.; Shao, M. Asynchrony of spring phytoplankton response to temperature driver within a spatial heterogeneity bay of Three-Gorges Reservoir. China. Limnologica 2011, 41, 174–180. [Google Scholar] [CrossRef]
- Barinova, S.; Liu, N.; Ding, J. , An, Y.; Qin, X; Wu, C. Ecological assessment of water quality of the Songhua River upper reaches by algal communities. Acta Ecologica Sinica, 2016, 3, 126–132. [Google Scholar] [CrossRef]
- Barinova, S.; Liu, N.; Ding, J.; An, Y.; Qin, X.; Wu, C. Bioindication of water quality of the Xinlicheng Reservoir by algal communities. Transylvanian Review of Systematical and Ecological Research. The Wetlands Diversity 2020, 22.3, 1–16. [Google Scholar] [CrossRef]
- Barinova, S.; Alster, A. Algae and Cyanobacteria Diversity and Bioindication of Long–Term Changes in the Hula Nature Reserve, Israel. Diversity 2021, 13, 583. [Google Scholar] [CrossRef]
- Biggs, J.; Von Fumetti, S.; Kelly–Quinn, M. The importance of small waterbodies for biodiversity and ecosystem services: implications for policy makers. Hydrobiologia 2017, 793, 3–39. [Google Scholar] [CrossRef]
- Baturina, M. A.; Kononova, O. N.; Fefilova, E. B.; Teteryuk, B. Yu.; Patova, E. N.; Stenina, A. S.; Sterlyagova, I. N. Present State of Biota of Small Komi Republic Reservoirs. J. of Siberian Federal University. Biology 2017, 10, 422–445. [Google Scholar] [CrossRef]
- Antonova, L. N.; Kanyuk, G. I.; Pogonina, T. E.; Omelchenko, L. N.; Fokina, A. R. Purpose and features of the operating conditions of cooling ponds of thermal and nuclear power plants. East European Journal of Advanced Technologies 2012, 2, 56–63. [Google Scholar]
- Mason, C. F. Water pollution biology. Pollution: causes, effects and control, 3rd ed.; Mason, C. F., Ed.; Longman: Harlow, Essex, England, 1996; pp. 82–112. [Google Scholar]
- Peeters, F.; Straile, D.; Lorke, A.; Livingstone, D. M. Earlier onset of the spring phytoplankton bloom in lakes of the temperate zone in a warmer climate. Global Change Biology 2007, 13, 1898–1909. [Google Scholar] [CrossRef]
- Paerl, H. W.; Huisman, J. Blooms like it hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [PubMed]
- Zhilenkov, V.N. Experience in developing criteria for designing a filtration–selective water intake in the technical water supply system of the Pechorskaya State District Power Station. Izvestiya VNIIG im. B.E. Vedeneeva 2002, 240, 206–218. (In Russian) [Google Scholar]
- Boznak, E. I.; Zakharov, A. B. Fish population of an industrial reservoir under conditions of multifactorial anthropogenic impact. In Aquatic ecology: Modern problems of reservoirs and their watersheds, Proceed. of the Intern. Scient. Confer., Perm, Russia, 30 May – 2 June, 2009; Perm State Research University: Perm, Russia, 2009; Volume 2; pp. 220–224. (In Russian) [Google Scholar]
- Rafikov, R. R.; Boznak, E. I. Ichthyological characteristics of bleak Alburnus alburnus (Сyprinidae) from a cooling pond in the Pechora River basin. J. of Ichthyology 2021, 61, 646–649. [Google Scholar] [CrossRef]
- Methodology for studying biogeocenoses of inland water bodies. Mordukhai–Boltovskoi, F. D., Ed.; Nauka: Moscow, 1975; p. 240. (In Russian) [Google Scholar]
- Patova, E. N.; Stenina, A. S; Shabalina, Y. N.; Sterlyagova, I. N. Phytoplankton Communities of the Vychegda River in the Area of Treated-Wastewater Intake from the Pulp and Paper Industry. Contemporary Problems of Ecology 2021, 14, 633–641. [Google Scholar] [CrossRef]
- Diatoms of the USSR (fossil and modern). Gleser, S. I.; Jouse, A. P., Makarova, I. V., Proschkina-Lavrenko, A. I., Eds.; Sheshukova-Poretzkaja, V. S. Eds. Nauka: Leningrad, USSR, 1974; Volume 1; pp. 403. (In Russian) [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota 1. Teil: Chroococcales. In Süsswasserflora von Mitteleuropa 19/1; Unaltered, repr., Ettl, H., Gärtner, G., Heynig, G., Mollenhauer, D., Eds.; Spektrum Akademischer Verlag: Heidelberg, Germany, 2008; p. 548. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota 2. Teil: Oscillatoriales. In Süsswasserflora von Mitteleuropa 19/2; Büdel, B., Gärtner, G., Krienitz, L., Schlager, M., Eds.; Unaltered repr., 2. print; Spektrum Akademischer Verlag: Heidelberg, Germany, 2008; p. 759. [Google Scholar]
- Komárek, J. Cyanoprokaryota 3. Teil: Heterocytous genera. In Süsswasserflora von Mitteleuropa 19/3; Büdel, B., Gärtner, G., Krienitz, L., Schlager, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; p. 1133. [Google Scholar]
- Krammer, K.; Lange Bertalot, H. Bacillariophyceae 1. Naviculaceae. In Sǘßwasserflora von Mittelflora, 2/1. G. Fischer: Jena, Stuttgart, Lubeck, Ulm., 1986; pp. 876.
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae 2. Bacillariaceae, Epithemiaceae, Surirellaceae. In Süsswasserflora von Mitteleuropa, 2/2; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; G. Fischer: Jena, Germany, 1988; p. 596. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae 3. Centrales, Fragilariaceae, Eunotiaceae. Susswasserflora von Mitteleuropa, 2/3; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fisher Verlag: Stuttgart, 1991; p. 576. [Google Scholar]
- Škaloud, P.; Rindi, F.; Boedeker, C.; Leliaert, F. Freshwater Flora of Central Europe. Chlorophyta: Ulvophyceae. Bd. 13. In Süβwasserflora von Mitteleuropa. Springer Spektrum: Berlin/ Heidelberg, Germany, 2018; p. 289. [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. In World-Wide Electronic Publication; National University of Ireland: Galway, Ireland, 2023; Available online: http://www.algaebase.org (accessed on 20 September 2023).
- Barinova, S.; Bilous, O.; Tsarenko, P.M. Algal indication of water bodies in Ukraine: methods and perspectives. Haifa University Publishing House: Haifa, Kiev, 2019. pp. 367. (In Russian).
- Sládeček, V. System of water quality from the biological point of view. Arch. Hydrobiol. 1973, 7, 218. [Google Scholar]
- Barinova, S. S.; Medvedeva, L. A.; Anisimova, O. V. Diversity of Algal Indicators in Environmental Assessment. Pilies Studio: Tel-Aviv, Israel, 2006, pp. 498. (In Russian).
- Barinova, S. Essential and practical bioindication methods and systems for the water quality assessment. Int. J. Environ. Sci. Nat. Resour. 2017, 2, 79–89. [Google Scholar] [CrossRef]
- Love, J.; Selker, R.; Marsman, M.; Jamil, T.; Dropmann, D.; Verhagen, J.A.; Ly, A.; Gronau, F.Q.; Smira, M.; Epskamp, S.; et al. JASP: Graphical statistical software for common statistical designs. J. Stat. Softw. 2019, 88, 1–17. [Google Scholar] [CrossRef]
- Watanabe, T.; Asai, K.; Houki, A. Numerical estimation of organic pollution of flowing water by using the epilithic diatom assemblage—Diatom Assemblage Index (DAIpo). Sci. Total Environ. 1986, 55, 209–218. [Google Scholar] [CrossRef]
- Getsen, M. V. Algae of the Pechora River basin. Composition and distribution. Nauka: Leningrad, USSR, 1973, pp. 147. (In Russian).
- Habib, O. A.; Tippett, R.; & Murphy, K. J. J. Seasonal changes in phytoplankton community structure in relation to physico–chemical factors in Loch Lomond, Scotland. Hydrobiologia 1997, 350, 63–79. [Google Scholar] [CrossRef]
- Willén, E. Dominance patterns of planktonic algae in Swedish forest lakes. Hydrobiologia 2003, 502, 315–324. [Google Scholar] [CrossRef]
- Paerl, H. W.; Hall, N. S.; Calandrino, E. S. Controlling harmful cyanobacterial blooms in a world experiencing anthropogenic and climatic–induced change. Science of the total environment 2011, 409, 1739–1745. [Google Scholar] [CrossRef] [PubMed]
- Patova, E. N. Bloom-forming Cyanoprokaryotes in Kharbeyskie lakes of Bolshezemelskaya tundra. J. of the Siberian Federal University. Biology 2014, 7, 282–290. [Google Scholar] [CrossRef]
- Kurashov, E. A.; Barbashova, V. A.; Dudakova, D. S.; Kapustina, L. L.; Mitrukova, G. G.; Rusanov, A. G.; Aleshina, D. G.; Iofina, I. V.; Protopopova, Ye. V.; et al. Ladoga Lake ecosystem: present–day conditions and trends in the late XX to early XXI century. Biosphere 2018, 2, 65–121. (In Russian) [Google Scholar] [CrossRef]
- Dreher, T. W.; Matthews, R.; Davis II, E. W.; Mueller, R. S. Woronichinia naegeliana: A common nontoxigenic component of temperate freshwater cyanobacterial blooms with 30 % of its genome in transposons. Harmful Algae 2023, 125, 102433. [Google Scholar] [CrossRef]
- Gabyshev, V.; Davydov, D.; Vilnet, A.; Sidelev, S.; Chernova, E.; Barinova, S.; Gabysheva, O.; Zhakovskaya, Z. Gloeotrichia cf. natans (Cyanobacteria) in the Continuous Permafrost Zone of Buotama River, Lena Pillars Nature Park, in Yakutia (Russia). Water 2023, 15, 2370. [Google Scholar] [CrossRef]
- Winder, M.; Sommer, U. Phytoplankton response to a changing climate. Hydrobiologia 2012, 698, 5–16. [Google Scholar] [CrossRef]
- Barinova, S.; Gabyshev, V.; Genkal, S.; Gabysheva, O. Diatoms of Small Water Bodies as Bioindicators in the Assessment of Climatic and Anthropogenic Impacts on the Coast of Tiksi Bay, Russian Arctic. Water 2023, 15, 1533. [Google Scholar] [CrossRef]
- Pislegina, E. V.; Shchapov, K. S.; Izmestyeva, L. R. Influence of wind carrying over on an abundance of a plankton in direct thermal stratification of 2009 in Southern Baikal (settlement the Bolshie Coty). Bulletin of the Irkutsk State University. Biology. Ecology 2011, 4, 67–72. (In Russian) [Google Scholar]
- Mikaelyan, A. S.; Zatsepin, A. G.; Kubryakov, A. A. Effect of mesoscale eddy dynamics on bioproductivity of the marine ecosystems (review). Physical Oceanograph 2020, 27, 590–618. [Google Scholar] [CrossRef]
- Barinova, S.; Gabyshev, V.; Genkal, S. Diversity of Diatom Algae in the Lena Delta Nature Reserve and the Adjacent Territory in the Specific Ecological Factors of the Arctic. Diversity 2023, 15, 802. [Google Scholar] [CrossRef]
- Hustedt, F. Systematische und Ökologische Untersuchungen über die Diatomeenflora von Java, Bali und Sumatra. Archiv. Hydrobiol. Suppl. 1938–1939, 15–16, 131–177, 393–506, 638–790, 1–155, 274–394.
- Hustedt, F. Die Diatomeen flora des Flußsystems der Weser im Gebiet der Hansestadt Bremen. Abhandlungen Naturwissenschaftlichen Vereins Bremen 1957, 34, 181–440. [Google Scholar]
- Van Dam, H.; Mertens, A.; Sinkeldam, J. A coded checklist and ecological indicator values of freshwater diatoms from the Netherlands. Netherlands Journal of Aquatic Ecology 1994, 28(1), 117–133. [Google Scholar] [CrossRef]
Figure 1.
Location map of the Pechorskoe Reservoir. 1-8 – location of research stations in the water area. The red arrow is the place where warm waters are discharged; the blue arrow is the place where water is input to the cooling system; (a) – fragment of the satellite image (for 07 June 2022; (b) – wind rose in June and August 2022; (c) – location of the reservoir on the northern Asia map; (d) – statistical map of the reservoir surface with the sampling station position.
Figure 1.
Location map of the Pechorskoe Reservoir. 1-8 – location of research stations in the water area. The red arrow is the place where warm waters are discharged; the blue arrow is the place where water is input to the cooling system; (a) – fragment of the satellite image (for 07 June 2022; (b) – wind rose in June and August 2022; (c) – location of the reservoir on the northern Asia map; (d) – statistical map of the reservoir surface with the sampling station position.
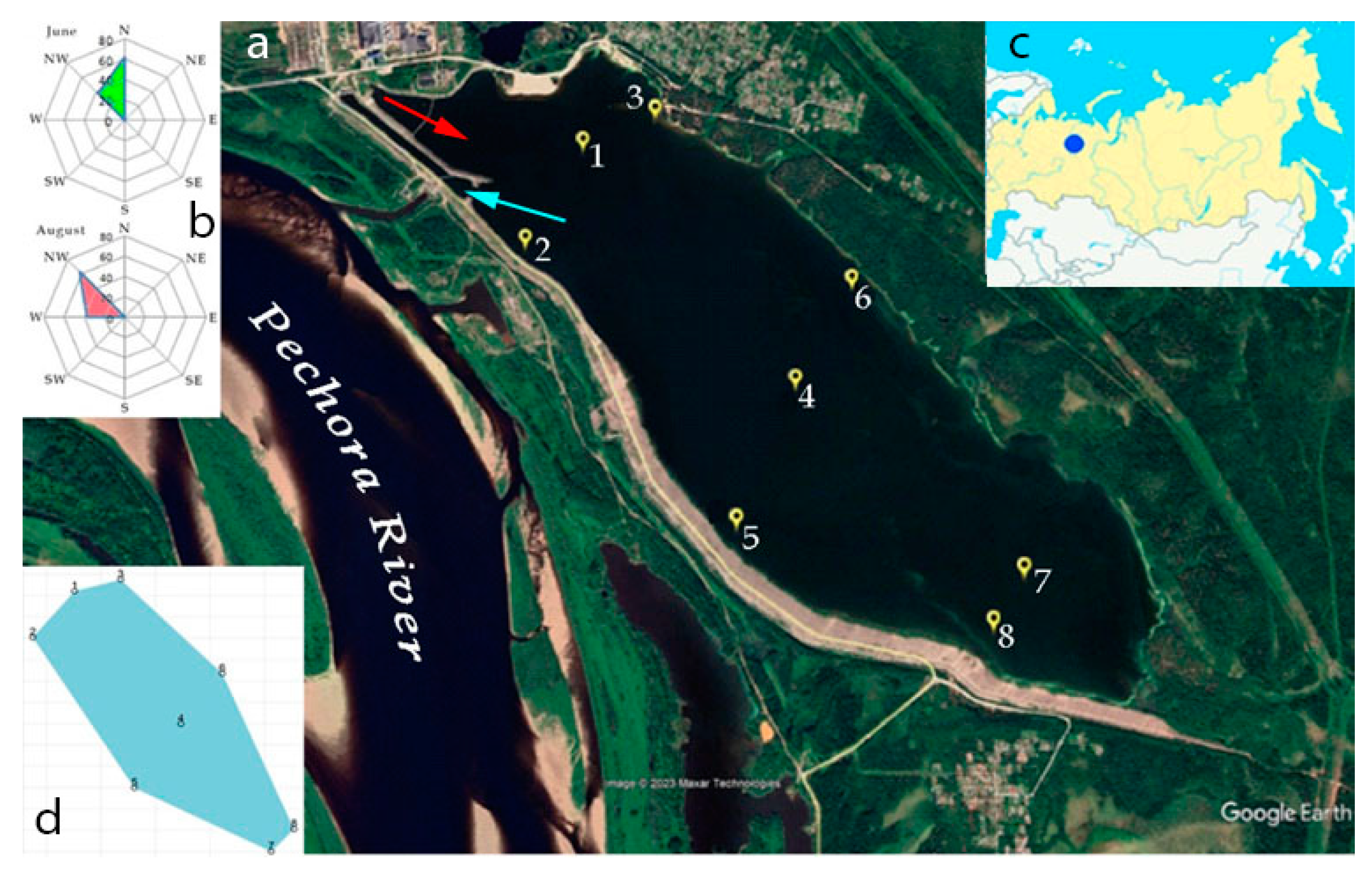
Figure 2.
General view of stations in the Pechorskoe Reservoir. View of the studied stations: (a) — st. 1 upper part of the reservoir, center; (b) — st. 2 upper part, near the southern coast (place of discharge of warm waters); (c) — st. 4 central part of the reservoir; (d) — st. 6 in the central part of the reservoir on the north coast; (e) — st. 7 lower part, center; (f) — st. 8 lower part, near the southern coast.
Figure 2.
General view of stations in the Pechorskoe Reservoir. View of the studied stations: (a) — st. 1 upper part of the reservoir, center; (b) — st. 2 upper part, near the southern coast (place of discharge of warm waters); (c) — st. 4 central part of the reservoir; (d) — st. 6 in the central part of the reservoir on the north coast; (e) — st. 7 lower part, center; (f) — st. 8 lower part, near the southern coast.

Figure 3.
Temperatures of water of the Pechorskoe Reservoir (at a depth of 0.2 and 5 m) and air at the studied stations on the day of sampling; (a) – in June, (b) – in August.
Figure 3.
Temperatures of water of the Pechorskoe Reservoir (at a depth of 0.2 and 5 m) and air at the studied stations on the day of sampling; (a) – in June, (b) – in August.

Figure 4.
Statistical maps of the distribution of the total species diversity of phytoplankton (a, e) and leading phyla: diatoms (b, f), cyanobacteria (c, g), and green algae (d, h) by stations of the Pechorskoe Reservoir in June-August 2022.
Figure 4.
Statistical maps of the distribution of the total species diversity of phytoplankton (a, e) and leading phyla: diatoms (b, f), cyanobacteria (c, g), and green algae (d, h) by stations of the Pechorskoe Reservoir in June-August 2022.
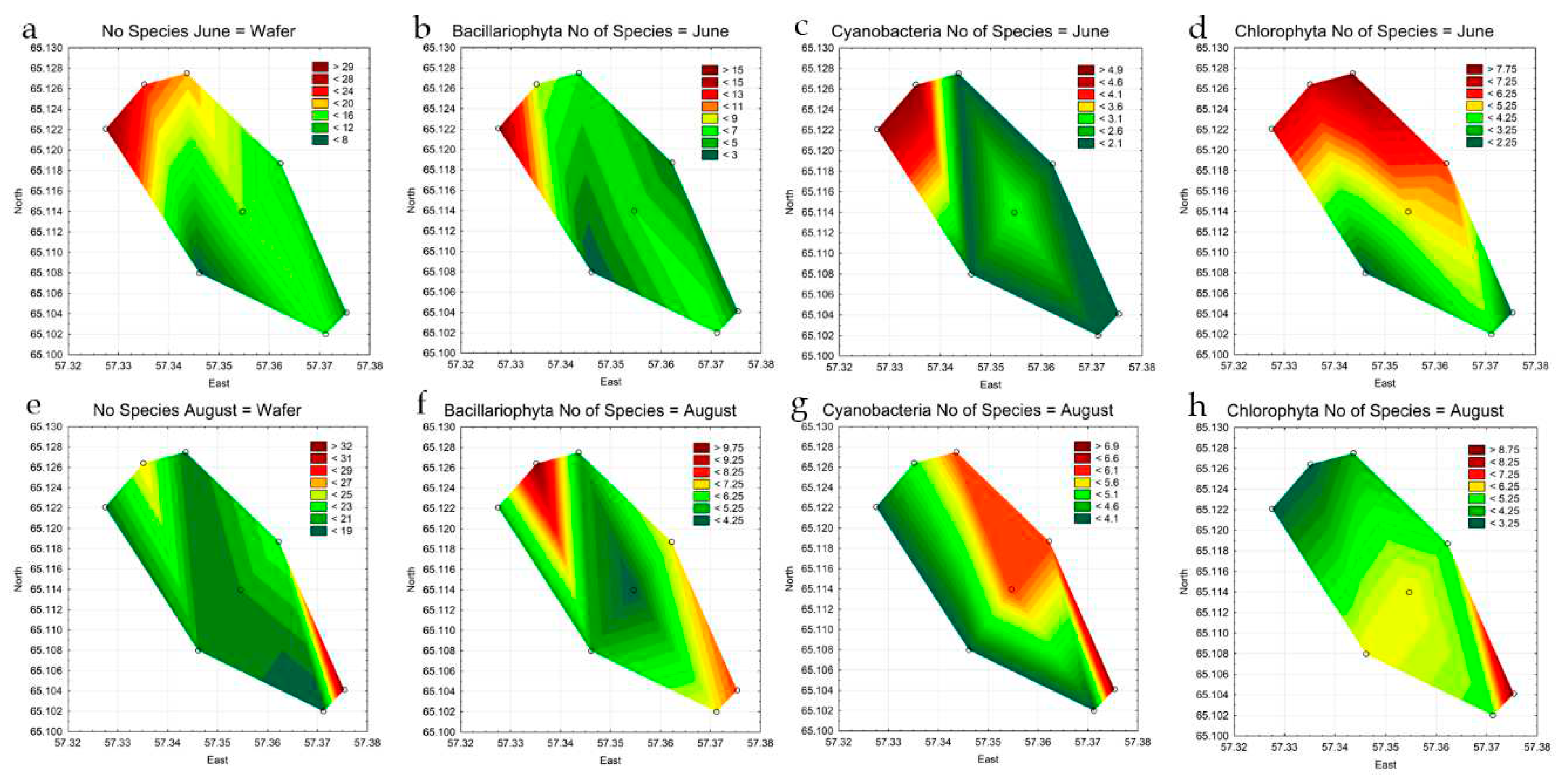
Figure 5.
Cyanobacteria and algae species dominant in phytoplankton of the Pechorskoe Reservoir (June, August 2022): a – cyanobacterial water bloom; a, b – Woronichinia naegeliana; a, c - Aphanizomenon flos-aquae; d - Pediastrum duplex; e - Aulacoseira italica; f - Hindakia tetrachotoma.
Figure 5.
Cyanobacteria and algae species dominant in phytoplankton of the Pechorskoe Reservoir (June, August 2022): a – cyanobacterial water bloom; a, b – Woronichinia naegeliana; a, c - Aphanizomenon flos-aquae; d - Pediastrum duplex; e - Aulacoseira italica; f - Hindakia tetrachotoma.
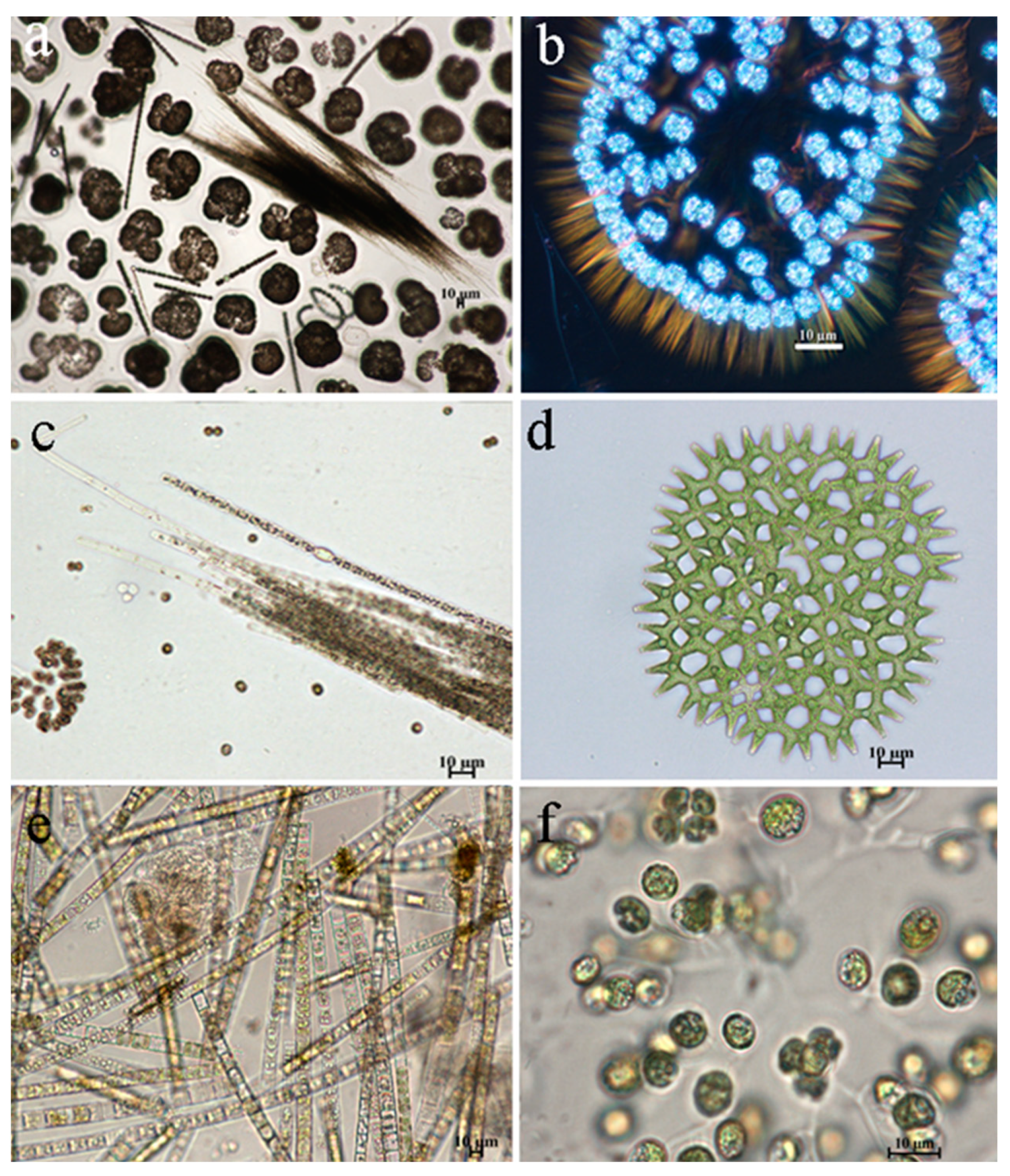
Figure 6.
Statistical maps of distribution of abundance (million cells L-1) of all phytoplankton groups (a, e) and leading phyla: diatoms (b, f), cyanobacteria (c, g), and green algae (d, h) in the Pechorskoe Reservoir (June-August 2022).
Figure 6.
Statistical maps of distribution of abundance (million cells L-1) of all phytoplankton groups (a, e) and leading phyla: diatoms (b, f), cyanobacteria (c, g), and green algae (d, h) in the Pechorskoe Reservoir (June-August 2022).
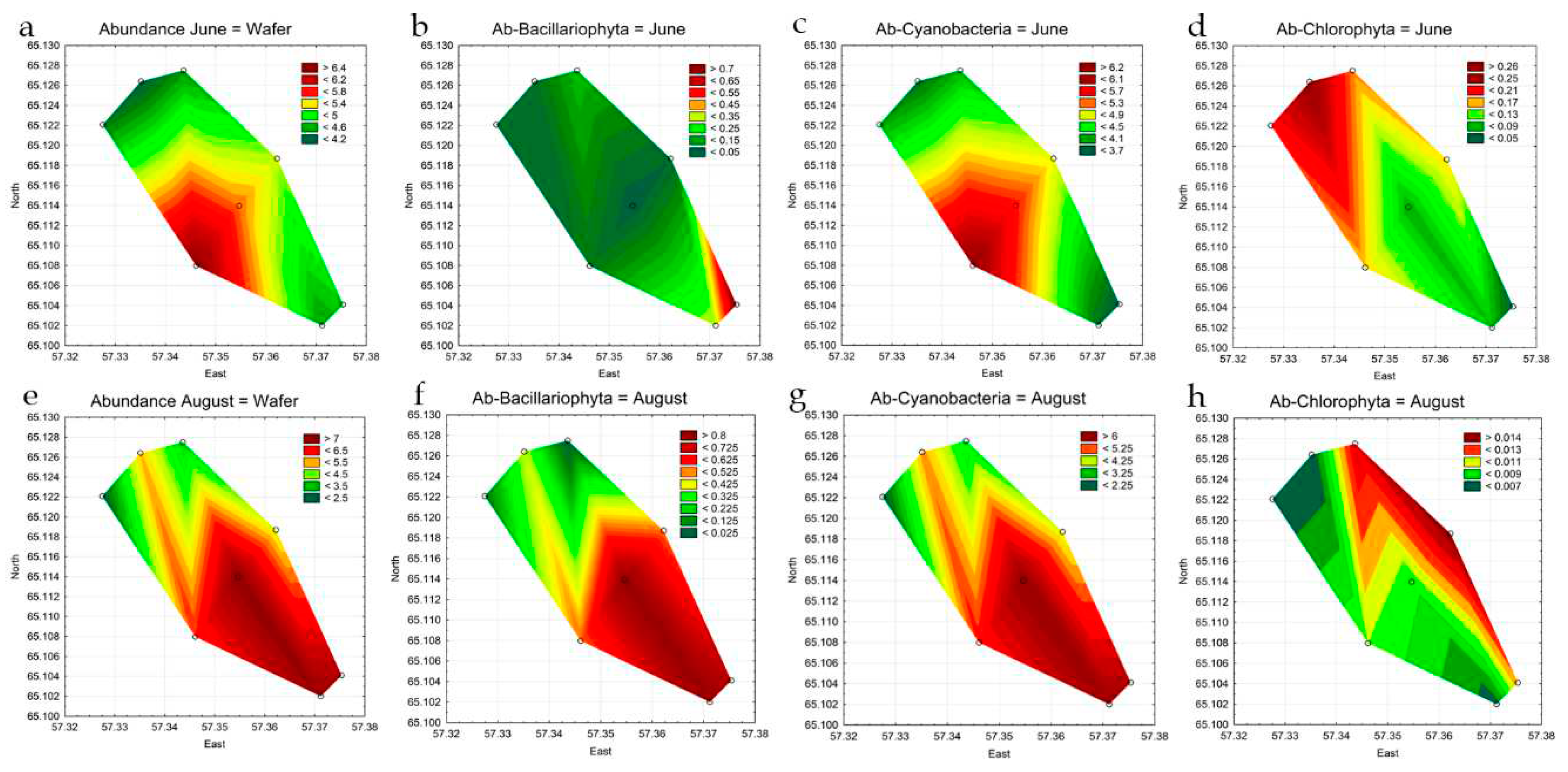
Figure 7.
Statistical maps of distribution of biomass (million cells L-1) of all phytoplankton groups (a, e) and leading phylum’s: diatoms (b, f), cyanobacteria (c, g), and green algae (d, h) in the Pechorskoe Reservoir (June-August 2022).
Figure 7.
Statistical maps of distribution of biomass (million cells L-1) of all phytoplankton groups (a, e) and leading phylum’s: diatoms (b, f), cyanobacteria (c, g), and green algae (d, h) in the Pechorskoe Reservoir (June-August 2022).
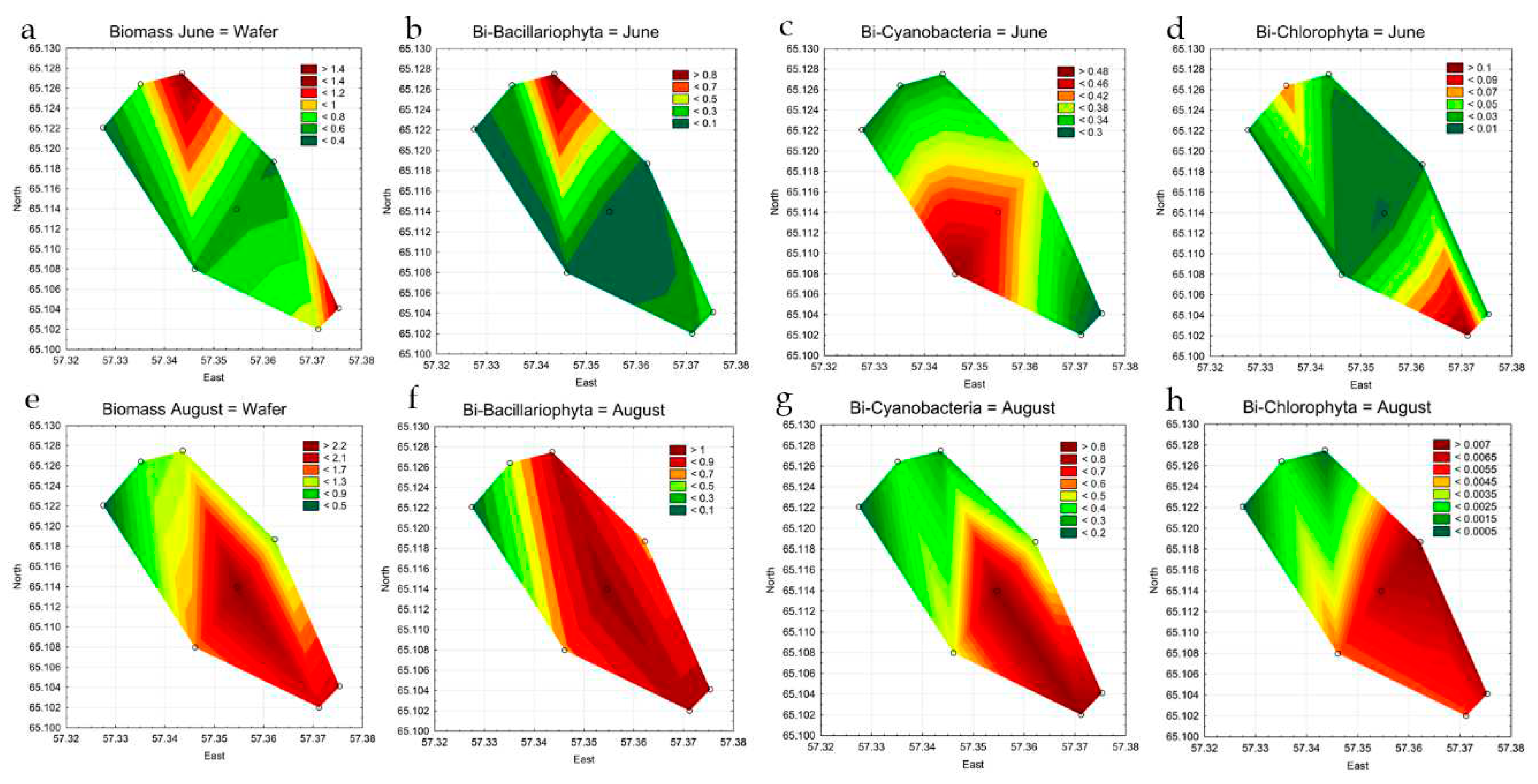
Figure 8.
JASP correlation network plot for the phytoplankton communities of the Pechorskoe Reservoir by sampling stations. The strongest links are shown by the thickest lines. Positive correlations are shown in blue lines, negative ones in red.
Figure 8.
JASP correlation network plot for the phytoplankton communities of the Pechorskoe Reservoir by sampling stations. The strongest links are shown by the thickest lines. Positive correlations are shown in blue lines, negative ones in red.
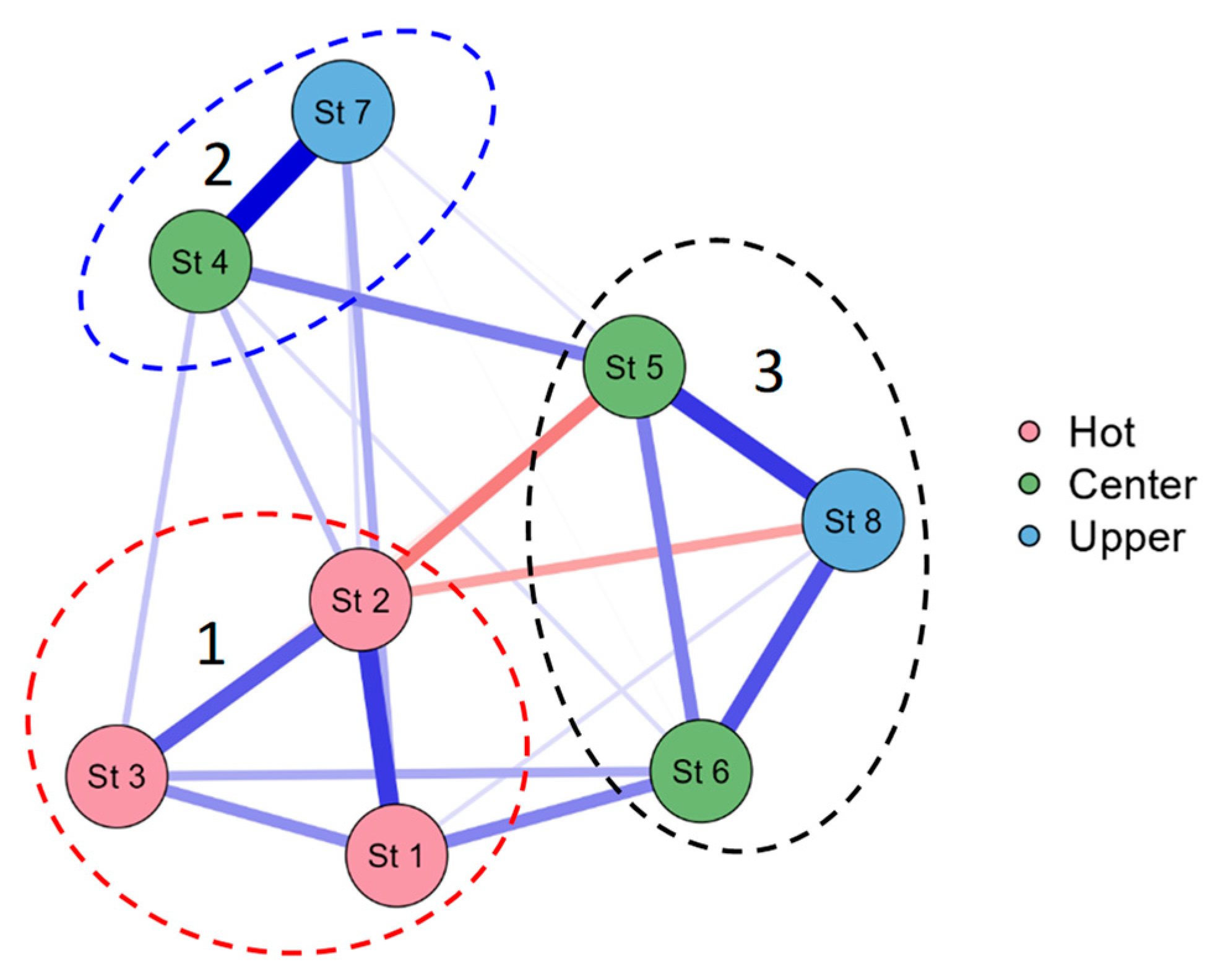
Figure 9.
Distribution of taxonomic content ecological indicators (a), indicators of habitat (b), water temperature (c) and oxygen (d) for phytoplankton communities in the Pechorskoe Reservoir. Habitat: P—planktonic; P–B—plankto-benthic; B—benthic. Temperature: cool—cool water; temp—temperate temperature; eterm—eurythermic; warm—thermophilic. Oxygenation and water moving: st—standing water; st-str—low streaming water; str – fast streaming water.
Figure 9.
Distribution of taxonomic content ecological indicators (a), indicators of habitat (b), water temperature (c) and oxygen (d) for phytoplankton communities in the Pechorskoe Reservoir. Habitat: P—planktonic; P–B—plankto-benthic; B—benthic. Temperature: cool—cool water; temp—temperate temperature; eterm—eurythermic; warm—thermophilic. Oxygenation and water moving: st—standing water; st-str—low streaming water; str – fast streaming water.
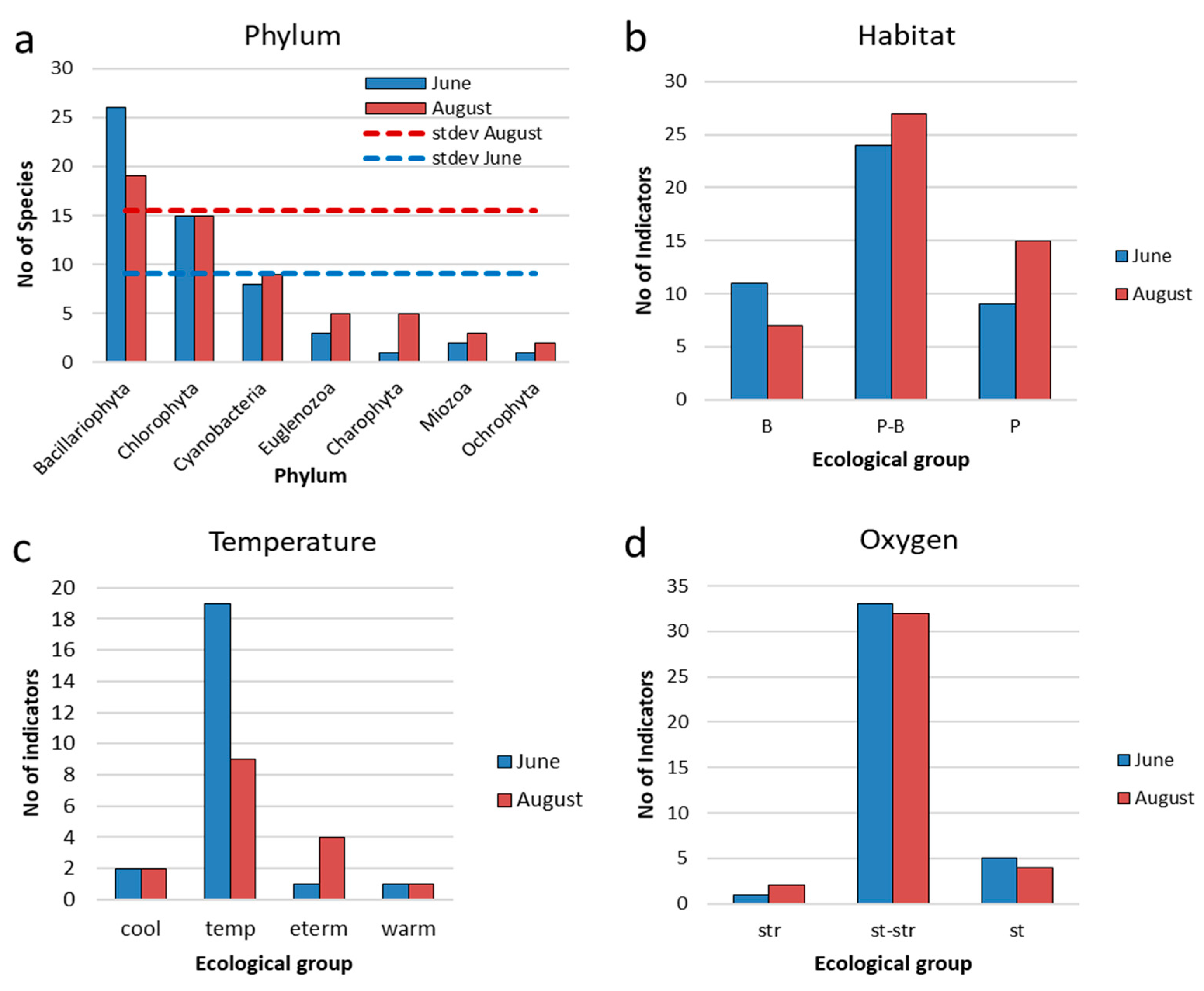
Figure 10.
Distribution of indicators of salinity (a), pH (b), saprobity (c) and nutrition type (d) for phytoplankton communities in the Pechorskoe Reservoir. Halobity degree (Salinity): hb – halophobes; i—oligohalobes–indifferent; hl—halophiles; mh—masohalobes. Acidity (pH): alf—alkaliphiles; ind—indifferent; acf—acidophiles; alb—alkalibiontes. Organic pollution indicators according to Watanabe: sx—saproxenes; es—eurysaprobes. Nitrogen uptake metabolism (autotrophy-heterotrophy): ate—nitrogen–autotrophic taxa; ats—tolerating elevated concentrations of organically bound nitrogen; hne—facultatively nitrogen–heterotrophic taxa, needing periodically elevated concentrations of organically bound nitrogen.
Figure 10.
Distribution of indicators of salinity (a), pH (b), saprobity (c) and nutrition type (d) for phytoplankton communities in the Pechorskoe Reservoir. Halobity degree (Salinity): hb – halophobes; i—oligohalobes–indifferent; hl—halophiles; mh—masohalobes. Acidity (pH): alf—alkaliphiles; ind—indifferent; acf—acidophiles; alb—alkalibiontes. Organic pollution indicators according to Watanabe: sx—saproxenes; es—eurysaprobes. Nitrogen uptake metabolism (autotrophy-heterotrophy): ate—nitrogen–autotrophic taxa; ats—tolerating elevated concentrations of organically bound nitrogen; hne—facultatively nitrogen–heterotrophic taxa, needing periodically elevated concentrations of organically bound nitrogen.
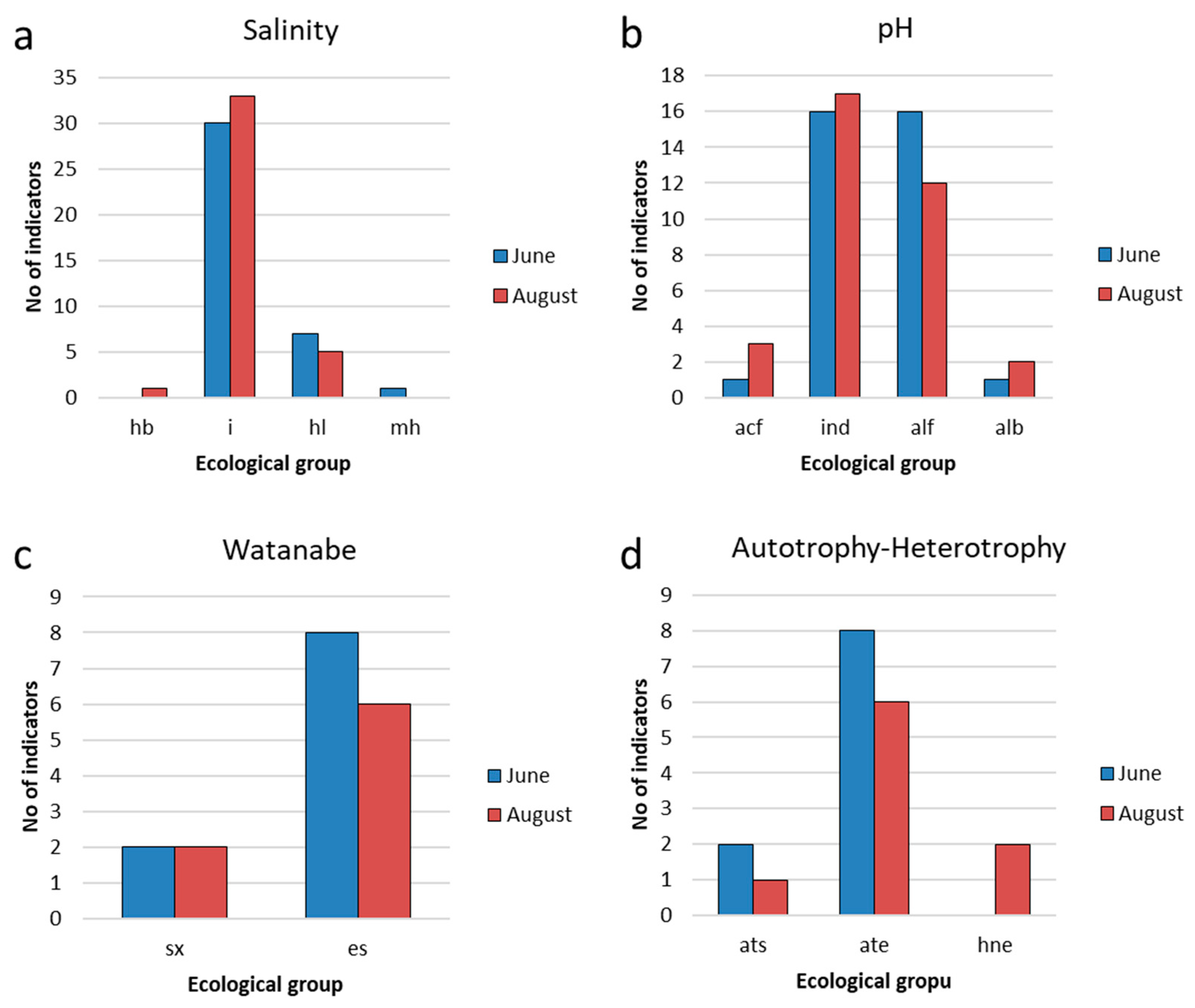
Figure 11.
Distribution of indicators of trophic state (a) and class of water quality (b) for phytoplankton communities in the Pechorskoe Reservoir. Trophic state: om—oligo–mesotraphentic; m—mesotraphentic; me—meso–eutraphentic; e—eutraphentic; he—hypereutraphentic. The water quality class is determined as the sum of indicators whose species-specific index saprobity S from Table Appendix A-1 is within the range of each class. Classes of water quality colored in EU color code.
Figure 11.
Distribution of indicators of trophic state (a) and class of water quality (b) for phytoplankton communities in the Pechorskoe Reservoir. Trophic state: om—oligo–mesotraphentic; m—mesotraphentic; me—meso–eutraphentic; e—eutraphentic; he—hypereutraphentic. The water quality class is determined as the sum of indicators whose species-specific index saprobity S from Table Appendix A-1 is within the range of each class. Classes of water quality colored in EU color code.
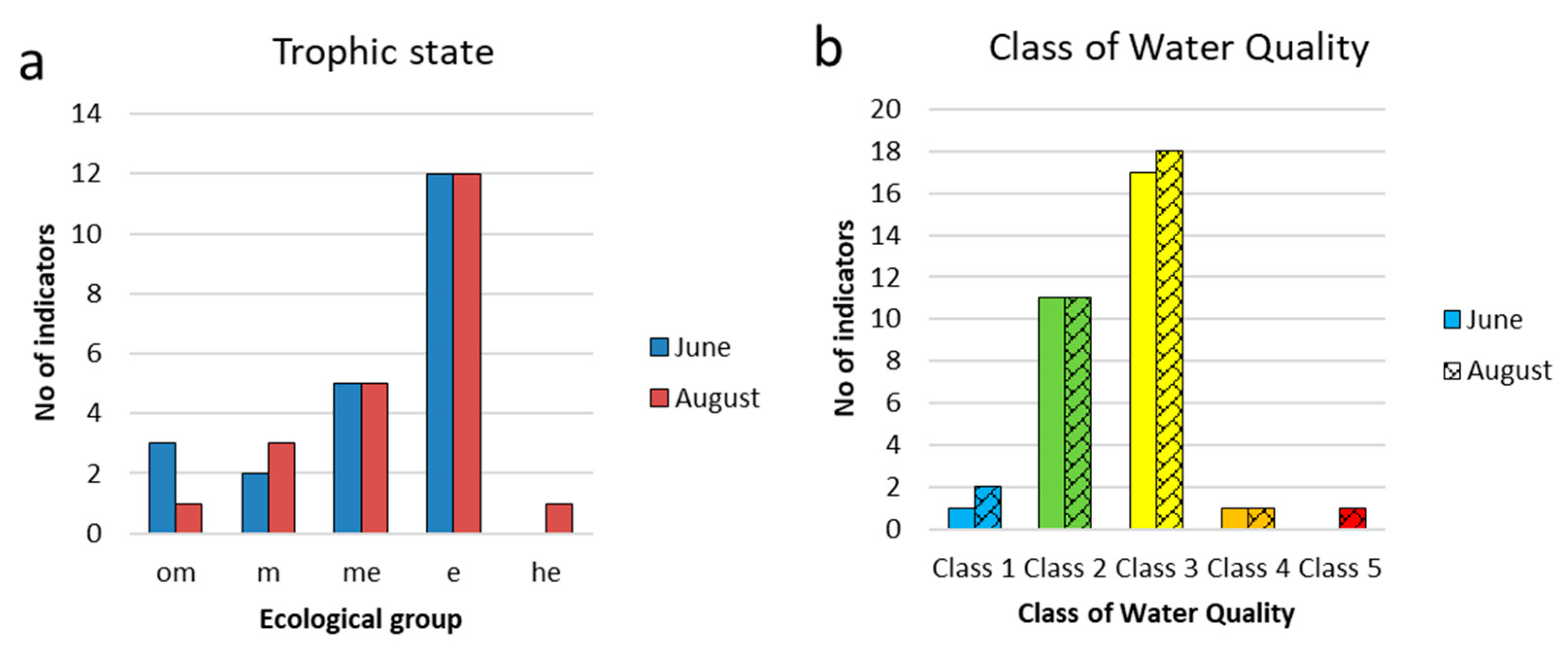
Figure 12.
Combined statistical maps for the Pechorskoe Reservoir. Index saprobity S in June (a) and August (b); Biomass in June (c) and August (d). Colors are given for the value amplitude of each mapped variable from green (lower boxes in the legend key) to red (upper boxes in the legend key).
Figure 12.
Combined statistical maps for the Pechorskoe Reservoir. Index saprobity S in June (a) and August (b); Biomass in June (c) and August (d). Colors are given for the value amplitude of each mapped variable from green (lower boxes in the legend key) to red (upper boxes in the legend key).
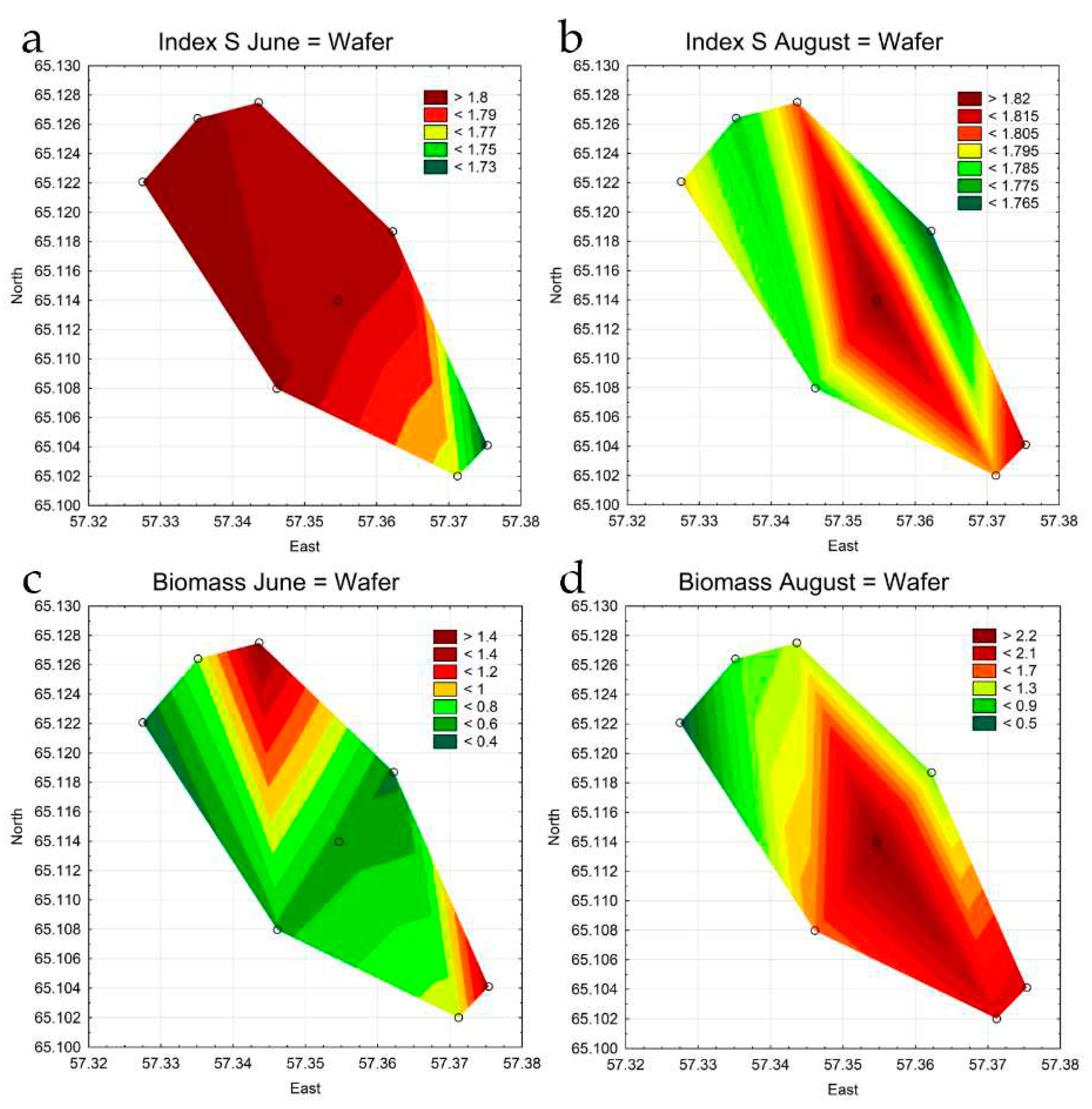
Table 1.
Characteristics sampling stations at the Pechorskoe Reservoir (June, August 2022) with coordinates and measured in 0.2 m surface horizon water pH and dissolved oxygen.
Table 1.
Characteristics sampling stations at the Pechorskoe Reservoir (June, August 2022) with coordinates and measured in 0.2 m surface horizon water pH and dissolved oxygen.
| Station | Location in reservoir |
Latitude | Longitude | Depth, m | Temperature, °С June |
Temperature, °С August |
pH June |
pH August |
Oxygen, mg O2 dm-3 June |
Oxygen, mg O2 dm-3 August |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Upper part, center | 65.126408° | 57.335149° | 5.8 | 21.1 | 18.6 | 7.2 | 7.6 | 8.51 | 8.25 |
| 2 | Upper part, near the southern coast (place of discharge of warm waters) | 65.122078° | 57.327509° | 4.3 | 24.5 | 19.9 | 7.2 | 7.6 | 8.43 | 8.77 |
| 3 | Upper part, on the north coast | 65.127499° | 57.343625° | 5.7 | 24.5 | 18.5 | 7.2 | 7.9 | 8.60 | 8.85 |
| 4 | Central part of the reservoir | 65.113962° | 57.354654° | 6.9 | 22.4 | 17.8 | 7.2 | 7.7 | 8.50 | 8.86 |
| 5 | Central part, near the southern coast | 65.107971° | 57.346128° | 4.0 | 21.1 | 18.2 | 7.1 | 7.9 | 8.74 | 9.18 |
| 6 | Central part. on the north coast | 65.118693° | 57.362238° | 5.0 | 21.3 | 18.3 | 7.1 | 7.9 | 8.71 | 8.96 |
| 7 | Lower part, center | 65.102001° | 57.371222° | 3.9 | 21.3 | 18.8 | 7.1 | 8.1 | 8.61 | 9.58 |
| 8 | Lower part, near the southern coast | 65.104101° | 57.375376° | 2.8 | 21.1 | 18.9 | 7.1 | 8.1 | 8.82 | 9.60 |
Table 2.
Averaged chemical variables of the Pechorskoe Reservoir in June and August 2022 with standard deviation.
Table 2.
Averaged chemical variables of the Pechorskoe Reservoir in June and August 2022 with standard deviation.
| Station | 1 | 4 | 7 | 1 | 4 | 7 | |
|---|---|---|---|---|---|---|---|
| Variable | Unit | June | June | June | August | August | August |
| рН | - | 7.2±0.2 | 7.2±0.2 | 7.1±0.2 | 7.6±0.2 | 7.7±0.2 | 8.1±0.2 |
| Electrical Conductivity | µS cm-1 | 79±4 | 78±4 | 78±4 | 87±4 | 87±4 | 89±4 |
| HCO3 | mg dm-3 | 43±5 | 43±5 | 69±8 | 47±6 | 45±5 | 48±6 |
| Total alkalinity | mmol dm-3 | 0.71±0.09 | 0.70±0.08 | 1.14±0.14 | 0.77±0.09 | 0.73±0.09 | 0.79±0.09 |
| PV (Permanganate Value) | mg dm-3 | 5.7±0.6 | 5.9±0.6 | 5.9±0.6 | 6.3±0.6 | 6.3±0.6 | 6.3±0.6 |
| COD | mgO dm-3 | 21±6 | 22±7 | 18±5 | 20±6 | 21±6 | 20±6 |
| CI- | mg dm-3 | 1.8±0.3 | 1.9±0.3 | 1.9±0.3 | 1.61±0.29 | 1.61±0.29 | 1.80±0.3 |
| SO42- | mg dm-3 | 4.5±0.8 | 3.8±0.8 | 3.8±0.8 | 3.9±0.8 | 4.1±0.8 | 3.7±0.8 |
| PO43- | mg dm-3 | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 | <0.05 |
| TSS | mg dm-3 | 0.0 | 1.45±0.26 | 0.63±0.14 | 1.17±0.21 | 1.06±0.19 | 2.00±0.40 |
| Fe | mg dm-3 | <0.050 | <0.050 | <0.050 | <0.050 | <0.050 | <0.050 |
| Cu | µg dm-3 | <1 | 6.1±1.4 | 4.2±1.0 | 5.3±2.2 | 5.1±2.1 | 4.4±1.9 |
| Phenol | µg dm-3 | <0.25 | 0.35±0.14 | 0.28±0.11 | <0.25 | <0.25 | <0.25 |
| Zn | µg dm-3 | <5 | <5 | <5 | <5 | <5 | <5 |
| Sr | µg dm-3 | 40±10 | 41±11 | 35±9 | 41±11 | 41±11 | 42±10 |
| Mn | µg dm-3 | 1.7±0.4 | <1.0 | 1.3±0.4 | <1.0 | <1.0 | <1.0 |
| Ni | µg dm-3 | <1.0 | <1.0 | <1.0 | 2.6±1.1 | 2.1±0.9 | 2.0±0.8 |
| Ca | mg dm-3 | 10.0±1.6 | 10.2±1.6 | 9.05±1.4 | 12.7±2.0 | 12.6±2.0 | 12.8±2.1 |
| Mg | mg dm-3 | 2.3±0.3 | 2.3±0.3 | 2.0±0.3 | 2.6±0.4 | 2.7±0.3 | 2.7±0.4 |
| K | mg dm-3 | 0.86±0.21 | 0.82±0.20 | 0.73±0.18 | 0.83±0.20 | 0.83±0.20 | 0.87±0.21 |
| Na | mg dm-3 | 3.2±0.5 | 3.3±0.5 | 3.0±0.4 | 3.2±0.5 | 3.2±0.5 | 3.3±0.5 |
| N-NO3 | mg dm-3 | <0.010 | <0.010 | <0.010 | 0.018±0.008 | <0.010 | 0.015±0.008 |
| N-NO2 | mg dm-3 | <0.010 | <0.010 | <0.010 | 0.016±0.006 | <0.010 | <0.010 |
| N-NН4 | mg dm-3 | 0.010±0.002 | 0.011±0.002 | 0.041±0.009 | 0.061±0.014 | 0.031±0.007 | 0.070±0.016 |
| Stotal | mg dm-3 | 1.7±0.4 | 1.8±0.4 | 1.6±0.4 | 1.8±0.4 | 1.8±0.4 | 1.8±0.4 |
| Ptotal | mg dm-3 | <0.020 | <0.020 | <0.020 | 0.027±0.011 | <0.020 | <0.020 |
Note: < - marks values below the detection range.
Table 3.
Indicators of phytoplankton development of the Pechorskoe Reservoir at the studied stations - number of species, abundance, biomass, Indices of saprobity.
Table 3.
Indicators of phytoplankton development of the Pechorskoe Reservoir at the studied stations - number of species, abundance, biomass, Indices of saprobity.
| Variable | St 1 | St 2 | St 3 | St 4 | St 5 | St 6 | St 7 | St 8 |
|---|---|---|---|---|---|---|---|---|
| No Species June | 23 | 30 | 20 | 16 | 7 | 13 | 15 | 10 |
| No Species August | 26 | 19 | 19 | 20 | 20 | 23 | 18 | 33 |
| Bacillariophyta, no. species June | 8 | 16 | 6 | 6 | 2 | 3 | 7 | 3 |
| Chlorophyta, no. species June | 7 | 7 | 8 | 5 | 2 | 6 | 4 | 2 |
| Cyanobacteria, no. species June | 5 | 5 | 2 | 3 | 2 | 2 | 2 | 2 |
| Euglenozoa, no. species June | 1 | 0 | 3 | 0 | 0 | 0 | 0 | 1 |
| Miozoa, no. species June | 2 | 1 | 1 | 1 | 0 | 1 | 1 | 1 |
| Ochrophyta, no. species June | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 1 |
| Charophyta, no. species June | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Bacillariophyta, no. species August |
10 | 5 | 5 | 4 | 5 | 7 | 7 | 8 |
| Charophyta, no. species June | 3 | 2 | 3 | 1 | 2 | 1 | 1 | 4 |
| Chlorophyta, no. species June | 3 | 3 | 4 | 6 | 6 | 5 | 5 | 9 |
| Cyanobacteria, no. species June | 5 | 4 | 6 | 6 | 4 | 6 | 4 | 7 |
| Euglenozoa, no. species June | 2 | 3 | 0 | 1 | 1 | 1 | 0 | 2 |
| Miozoa, no. species June | 2 | 2 | 1 | 2 | 2 | 2 | 1 | 2 |
| Ochrophyta, no. species June | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 1 |
| Total Abundance, million cells L-1 June |
4.1470 | 4.3348 | 4.4660 | 5.5973 | 6.4875 | 5.1283 | 4.4813 | 4.6775 |
| Total Abundance, million cells L-1 August | 5.2998 | 2.2891 | 3.5435 | 7.7418 | 6.0063 | 4.8970 | 7.6073 | 7.2395 |
| Bacillariophyta, million cells L-1 June | 0.0675 | 0.0490 | 0.1700 | 0.0313 | 0.0800 | 0.0525 | 0.3353 | 0.7400 |
| Chlorophyta, million cells L-1 June | 0.2603 | 0.1993 | 0.1745 | 0.0823 | 0.1613 | 0.1580 | 0.0885 | 0.0445 |
| Cyanobacteria, million cells L-1 June | 3.8105 | 4.0513 | 4.1163 | 5.4663 | 6.2450 | 4.8925 | 3.7825 | 3.6463 |
| Bacillariophyta, million cells L-1 August | 0.3698 | 0.0735 | 0.0088 | 0.8425 | 0.5325 | 0.5288 | 0.8513 | 0.7578 |
| Chlorophyta, million cells L-1 August | 0.0060 | 0.0060 | 0.0140 | 0.0093 | 0.0100 | 0.0158 | 0.0065 | 0.0118 |
| Cyanobacteria, million cells L-1 August | 4.8825 | 2.1960 | 3.4670 | 6.7825 | 5.3950 | 4.3113 | 6.6425 | 6.3263 |
| Total Biomass, mg L-1 June | 0.8096 | 0.3873 | 1.4320 | 0.5467 | 0.5757 | 0.4696 | 0.8639 | 1.3103 |
| Total Biomass, mg L-1 August | 1.0799 | 0.4032 | 1.2327 | 2.3626 | 1.5968 | 1.1466 | 1.9572 | 1.9903 |
| Bacillariophyta, mg L-1 June | 0.1859 | 0.0403 | 0.9847 | 0.0339 | 0.0647 | 0.0432 | 0.1309 | 0.2950 |
| Chlorophyta, mg L-1 June | 0.0688 | 0.0174 | 0.0211 | 0.0085 | 0.0134 | 0.0174 | 0.1061 | 0.0226 |
| Cyanobacteria, mg L-1 June | 0.3135 | 0.3018 | 0.3203 | 0.4351 | 0.4965 | 0.3851 | 0.3008 | 0.2901 |
| Bacillariophyta, mg L-1 August | 0.4954 | 0.0761 | 0.9239 | 1.0520 | 0.6741 | 0.6681 | 0.9559 | 0.9512 |
| Chlorophyta, mg L-1 August | 0.0025 | 0.0004 | 0.0006 | 0.0067 | 0.0049 | 0.0076 | 0.0051 | 0.0066 |
| Cyanobacteria, mg L-1 August | 0.3884 | 0.1752 | 0.2783 | 0.8494 | 0.4975 | 0.4281 | 0.8421 | 0.7784 |
| Index S June | 1.81 | 1.81 | 1.80 | 1.81 | 1.81 | 1.81 | 1.76 | 1.72 |
| Index S August | 1.78 | 1.80 | 1.80 | 1.82 | 1.78 | 1.76 | 1.80 | 1.82 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



