
Submitted:
25 September 2023
Posted:
26 September 2023
You are already at the latest version
Abstract
This research explored the influence of spatiotemporal variations in phreatic levels and salinity on the patterns and interactions of soil mesofauna communities in a tropical urban coastal wetland. It delineated significant variations in phreatic levels and salinity, with distinct regimes characterizing different habitats and hydroperiods. A total of 120 mesofauna morphotypes across 42 taxa were identified, revealing habitat differences and similarities, and the ecosystem biodiversity. Moderate correlations were established between phreatic levels, salinity, and mesofauna richness and abundance, with habitat type and phreatic level emerging as significant predictors in the regression model. Our findings indicate that shallow phreatic levels in conjunction with oligohaline salinity conditions foster a more favorable abiotic environment, thereby promoting increased biodiversity. The study underscores the dynamic and adaptive nature of mesofauna communities in response to changing environmental conditions, offering valuable insights for effective costal urban wetland management, particularly in the context of multiple stressors such as global and regional climate change, sea level rise, and human activities. The bio-sensor capacity of soil mesofauna emerges as a vital tool for monitoring and facilitating adaptive ecosystem management, thereby ensuring their long-term health and sustainability.
Keywords:
urban wetland
; coastal wetlands
; Puerto Rico
; hydroperiods
; phreatic level
; salinity conditions
; soil mesofauna
; biodiversity
1. Introduction
Wetlands provide heterogeneous ecological niches for unique soil arthropods assemblages; being mesofauna morphotypes the most abundant and diverse (Batzer et al., 2020). Among them, Mites (Acari) and springtails (Collembola), are micro-arthropods that range in size from 0.1 to 2 mm. They are present in different substrates with most of their assemblages concentrated in hot spots zones or resource patches [1] within the litter system [2,3] comprised of loose litter layer and upper 1-5 cm of soil. Mites and springtails have a significant impact on the decomposition processes and nutrient mobilization as they are plant litter transformers via a) fragmentation/comminution, b) ingestion of plant debris, and c) feces deposition for further mineralization by soil microflora (fungi and bacteria). They also foster the growth and dispersal of microbial populations and interact at different trophic levels through the litter decomposition process [4,5,6,7].
Soil mites mainly occur in three taxa: Oribatida (fungivores and detritivores), Mesostigmata (free-living soil predators) and Prostigmata (predators and fungal feeders). Springtails (fungivores and detritivores) are grouped in two: Arthropleona and Symphypleona [8,9]. The distribution patterns and interactions of these organisms occur at different spatiotemporal dynamics [1,4,6,7], regulated by the combined effects of scale-dependent variables: climate (temperature, precipitation), edaphic properties (porosity, structure, nutrients, humidity, pH, salinity), vegetation (resource quality and quantity), microtopography and species inter and intra specific interactions [7]. Soil higher temperatures and lower humidity may lead to the reduction of Mesostigmata mites’ population densities [10]. Prostigmata mites dominate in soils with low nutrient content and low humidity, while Oribatid mites and Springtails inhabit in nutrient rich humid environments [4,11,12]. Direct and indirect plant effects determine soil food web structure and mesofauna assemblage, as is the case of the legume Lotus corniculatus and the non-leguminous forb Plantago lanceolata [13]. Springtails distribution is correlated with soil pore-size, relative humidity, and food availability [14]. Trophic cascade effect is seen as microtopography affects the soil environment and the diversity and composition of soil fungal communities [15], which in turn may influence the food resources and habitat quality for Springtails and Oribatids [10].
In coastal wetlands, the spatiotemporal variations in the abundance of soil mesofauna are also influenced by drying/wetting cycles or hydro-patterns effects on soil bio- physicochemical conditions [16]. Wetlands wetting cycles include waterlogging or flooding time, frequency, duration, rate of water rises, and depth [17]. It is modified by different water sources that enter via in-situ precipitation, freshwater inputs, and seawater flows [18]. The hydrological regime determines the degree of salinity in the wetland substrate and influences the spatiotemporal vegetation cover, litter quality and quantity, and mesofauna composition [18,19]. In The Netherlands floodplains groundwater levels influence collembolans distribution with a shift in community composition along elevation gradients from regularly inundated areas to dry, seldom flooded zones [20,21]. A laboratory study that investigated the effect of salinity on soil species of Acari (mites), and Collembola (springtails), found that their reproduction cycles were differentially affected by increases in salinity. Mites were the least sensitive maintaining its reproduction cycle, while springtails reproduction significantly decreased [22]. In peatlands from Canada, China, Minnesota, and northern England, hydro patterns variations induced spatial and temporal bio-physicochemical factors fluctuations that influenced Acari and Collembola diversity and abundance [23].
Although mesofauna communities play an essential role in wetlands biological processes, there is still a gap in knowledge about how wetlands spatio-temporal dynamics influence mesofauna diversity, especially in coastal wetlands. More research is needed, especially in the Caribbean, to understand wetland marine-terrestrial-marine connectivity, bio-physicochemical components, future climate change scenarios and their effects on mesofauna assemblages. Because Caribbean coastal wetlands were anthropogenically modified since colonial times, a mosaic of physicochemical conditions, habitats, and vegetation cover characterizes these ecosystems; with undergoing global and regional climate variability, sea-level rise, land use and land cover changes acting as additional stressors (Figure 1).
The combined effect between these anthropogenic stressors and the predominant wetland mosaic environment influences the hydro-pattern regime, bio-physicochemical components, and soil mesofauna diversity, abundance, and functional relationships among taxa [24,25]. For example, sea level rise impacts coastal soil processes and modifies ecosystems' net primary production due to variations in salinity, which have direct and indirect effects on soil mesofauna diversity and function [26]. In the Zhanjiang Plain wetland, a field experiment that emulated phreatic level changes from precipitation variations showed evidence that these communities were significantly influenced by substrate water level. Slight increases in total abundance were documented under natural water level dynamics, with significant reductions under constant water level conditions [27].
Land use/land cover change, along with concomitant changes in hydrology, has a strong influence on altering vegetation patterns and soil salinity concentrations in Caribbean coastal urban wetlands, where a mosaic of different habitat type, instead of gradients, are the norm rather than the exception [18]. Given that mesofauna play an essential role in wetland biological processes and have specific environmental requirements, determining how climate variability, and habitat bio-physicochemical constituents influence their composition, becomes an important tool in adaptive wetland management to global and regional climate change, sea level rise and increased anthropic use of the region. Our aim is to determine how spatiotemporal variations in phreatic level and salinity affect patterns and interactions of soil mesofauna communities in a tropical urban coastal wetland. We hypothesis that habitat differences significantly modulate mesofauna diversity and abundance in tropical urban coastal wetlands, with spatio-temporal variations in phreatic levels and salinity exerting critical influence on community structures and interactions of soil mesofauna.
2. Materials and Methods
2.1. Study Area
The study took take place in 2.2 ha (Research Area) within Ciénaga Las Cucharillas Natural Reserve, a palustrine-estuarine coastal urban wetland on the northern coast of the Caribbean Island of Puerto Rico. The reserve is in the municipality of Cataño (18 26’25.27” N, 66 08’08.39” W). The wetland comprises the western side of the San Juan Bay (Figure 2).
Average monthly temperature ranges from 31°C to 25°C from May to October, and 22°C to 28°C from December to March. The area has a humid climate with an average annual precipitation of 1920 mm and a bimodal distribution with lesser precipitation from December until April-May and two peaks of rainfall (May-June), and September to November [28]. The study was carried out from 2020 to 2021 (Figure 3). In 2020, the wettest month was July, with monthly mean precipitation of 9.64 millimeters and 24 rainy days. The driest month was May, with monthly mean precipitation of 0.51 millimeters and 6 rainy days. In 2021, the wettest month was September, with monthly mean precipitation of 8.63 millimeters and 18 rainy days. The driest month was May, with monthly mean precipitation of 1.78 millimeters and 13 rainy days.
Ciénaga Las Cucharillas Natural Reserve is representative of how coastal wetlands in the Tropics, especially in the Caribbean, have been hydrologically modified from colonial times to the present. The hydrological modifications include (a) drainage channels for agricultural use from the 17th century until the mid-20th century [29,30]; (b) the construction of a flood control channel (La Malaria channel) in the late 1940s, bringing a direct flow of fresh water to the wetland from the upper and middle parts of the basin [31]; and (c) restricted seawater exchange due to the dike effect of an outflow water pump structure at the mouth of the channel [32] (Figure 4). As a result, tidal interaction in this wetland occurs via deep subsurface flow [33] Historical and present hydrological modifications bring about a mosaic of physicochemical conditions, habitats, and vegetation cover.
2.2. Research Area
Four study plots (3, 5, 6 and 10) of 100 square meters (Figure 2 C), with different spatiotemporal physicochemical factors and habitat type, were established in a research area at the Natural Reserve. Plot locations were selected based on geospatial analysis, presence of predominant wetland plant functional types and presence in all plots, and onsite measurements of substrate abiotic factors [18]. Each habitat has a predominant plant assemblage and variations in vegetation cover, soil type, and substrate salinity (Table 1). Category-4 Hurricane Maria (September 19-20, 2017) caused significant damage and defoliation in the study area. Twenty-seven per cent (27%) of the area was flooded for nearly six months [33].
Plot 3 is a mature mangrove woodland habitat (MMW), mostly covered with Laguncularia racemosa C.F.Gaertn. Plot 5, a 25-year restoration area (RMW) with a dominant cover of L. racemosa and herbaceous species belonging to the Cyperaceae, Vitaceae and Polygonacae families. It also has patches of grasses species belonging to Poaceae family. Plot 6 was a transitioning mangrove forest (natural recolonization-MWR) with a dominant cover of young and seedlings L. racemosa followed by grasses of Poaceae family. Plot 10 includes a mature 50-year shrub (SM) of Dalbergia ecastaphyllum (L.) Taub forest and a 6-year succession area (GF) composed mostly of Echinochloa polystachya (Kunth) Hitchc.
The research area is characterized by two soil types: Saladar muck (Sm) series and Martin Peña (Mp) series [34]. Saladar muck (Sm) series consists of black, highly decomposed (peat) autochthonous vegetation materials, that reach down to bedrock depth in the substrate (Table 1). Martin Peña (Mp) series contains deposits of organic material (0- 20 cm) close to surface, over mineral sediments, which includes silty clay loam (20-45 cm) embedded in the peat down to bedrock depth. At the study site, the layers of mineral sediments found in the soil are the result of anthropogenic allochthonous infills from upper terrestrial sources placed to prepare the land for shanty town establishment. MWR, GF and S habitats are on Saladar muck” (Sm) soil series, whereas MMW and RMW habitats area on Martin Peña (Mp) soil series.

Table 1.
Physicochemical factors and plots/habitats characteristics at the study site. Source [18].
Table 1.
Physicochemical factors and plots/habitats characteristics at the study site. Source [18].
| Plot | 3 | 5 | 6 | 10 | ||||
|---|---|---|---|---|---|---|---|---|
| Habitat Type | Mangrove woodland (MMW) | Rehabilitated Mangrove woodland (RMW) | Mangrove woodland (MWR) | > 50 years Shrub (SM) |
6 years grass & ferns (GF) |
|||
| Stage | Mature |
Rehabilitated, damaged in Hurricane Maria | Natural Recolonization Damaged in Hurricane Maria | Mature | Early successional |
|||
|
% Cover Plants Species |
92.6 % L. racemosa 3.2 % Acrostichum sp. 4.2% grasses of Poaceae family |
59.9 % young and seedlings L. racemosa, 33.8 % Herbs and vines 4.2% grasses of Poaceae family 2.0 % Acrostichum sp |
46.0 % Young and seedlings L. racemosa, 7.9 % Acrostichum sp, 13.3 % D. ecastaphyllum, 32.8 % grasses of Poaceae family |
40.4 % D. ecastaphyllum | 2.2 % L. racemosa (young trees), 0.4 % Acrostichum sp., 56.9 % Echinochloa sp |
|||
| Plant Type | Woody, Fern and Grass | Woody, Fern, Herbs and Grass | Woody, Fern, Shrub and Grass | Shrub | Woody, Fern and Grass | |||
| Soil Type | Mineral allochthonous embedded in an organic matrix (Martín Peña) |
Organic (peat) Autochthonous (Saladar muck) |
||||||
| Mean Salinity * | Wet Period | 3.4±2 O |
2.8±1 O |
2.4±2 O |
2.1±2 O |
2.1±2 O |
||
| Dry Period | 4±0.4 O |
12± 2 M |
11± 5 M |
7±1 M |
7±1 M |
|||
* Salinity values: O - Oligohaline refers to salinity levels between > 0.5ppt to 5.0 ppt, M - Mesohaline refers to salinity levels between > 5.0ppt to 18.0 ppt.
2.3. Data Collection
Sampling was carried out based on hydroperiod conditions. Substrate samples were collected on five dates: during Moderate Dry (June 18-25, 2020), Flood (October 23, 2020), Moist/Between Floods (March 19, 2021) and Wet (June 9, 2021) conditions. These conditions were strongly influenced by monthly variations in mean precipitation and the number of rainy days occurring before and on the sampling date (see Figure 5) [16,29]. They were also influenced by factors such as the amount of water stored in the soil from previous flooding and local rainfall (reflecting soil antecedent patterns of drying/wetting cycles) [35] and subsurface tidal effects. For instance, during prolonged dry periods, bimodal high tide reaches the study site in 20 minutes, whereas it takes up to 2 hours during wet periods [36].
Sampling period conditions were classified as moderate dry/moist when at the sampling date the mean phreatic level values were between -0.30m and -0.60 m, respectively, Wet/Flood when at the sampling date mean phreatic level values were between -0.15 m and above ground level (0 m) (Figure 5) [16]. We also consider rainy days conditions 14 days before sampling day. Months after the Wet/Flood sampling date, the wetland was affected by two tropical storms, Isaias (July 29-30, 2020) and Laura (August 22-23, 2020) and a prolonged rainy period until the end of October 2020 that caused frequent intense rainfall and four (4) months of flooding at the study site [37,38]. The flood sampling period coincided with the receding of floodwaters.
The sampling occurred in the morning, between 7:00 am to 10:00 am to reduce high tides influence (table 2), temperature, and water content variations. Five (5) plant species were chosen based on their functional type and presence in all habitats [18]: D. ecastaphyllum (shrub), E.polystachya (grass), Poaceae family (grass), A.danaeifolium (woody herb) and L. racemosa (tree). D. ecastaphyllum was present in two plots (6 and 10), E. polystachya was present in plot 10, while the three other species were present in plots 3, 5 and 6. In each habitat type, three plants per functional type were chosen. Three substrate samples per plant were collected every sampling date. Each substrate sample measuring 7.62 cm diameter x 5 cm depth, was separated into loose litter (relatively undecomposed) and old litter (partly to fully decomposed). Phreatic level was measured on site. Samples were taken to the laboratory, fresh weight was determined, and placed in lighted Tullgren-Berlese extractors for one week [25,39]. The extracted arthropods were preserved in 70% ethanol solution placed under each extractor. Morphotypes were separated according to their morphology and identified up to the level of order or family. For each sample they were identified and counted using Amscope SF2TRA stereoscopic binocular microscope or a Nikon Eclipse 80i microscope. After extraction, samples were oven dried at 60°C for a period of seven days. A subsample was mixed with distilled water (1:1) and homogenized to determined salinity using EcoSense® conductivity meter [40].
2.4. Data analysis
Habitat differences were evaluated by examining variations in phreatic level, salinity, and mesofauna richness and abundance. Taxa were delineated into categories of "dominant," "common," and "rare," based on their relative abundance: dominant taxa comprised those with a relative abundance ≥10%, common taxa ranged between 1% and 10%, and rare taxa constituted those with a relative abundance of less than 1% [41,42]. Habitat and plant type similarities were assessed utilizing the Jacobsen-Sørensen similarity index [43,44].
Morphotype richness and abundance were appraised to analyze differences in arthropod distribution and composition according to hydroperiods, phreatic levels, and salinity, both within and between habitat types [43]. Abundance values were normalized to represent the number of individuals per square meter (m²) [43,45]. To facilitate the analysis and discern patterns of diversity and abundance variations, phreatic level and salinity values were categorized as follows:
Phreatic Level Categories
- High Levels: 0.09 to 0.11 m
- Shallow/Near Surface: -0.03 to -0.07 m
- Slightly Moderate: -0.12 to -0.17 m
- Moderate: -0.36 to -0.43 m
- Significantly Deep: -0.51 to -0.64 m
Salinity Categories [46]
- Freshwater: 0 to 0.5 ppt
- Oligohaline: >0.5 to 5.0 ppt
- Mesohaline: >5.0 to 18.0 ppt
- Polyhaline: >18.0 to 33.0 ppt
Non-parametric statistical methods including the Wilcoxon/Kruskal-Wallis test and post-hoc Tukey’s tests were utilized to identify habitat differences and analyze the spatio-temporal influences of phreatic level and salinity on mesofauna diversity and abundance. Spearman’s Rho correlation analysis delineated the significant correlations between these factors and mesofauna composition. Further, a general regression model was employed to elucidate the relationships and to determine the significance and the intensity of the effects of phreatic level and salinity variables. All statistical analyses were conducted utilizing SAS JMP® Pro 16 statistical software.
3. Results
3.1. Variations in habitat phreatic level and salinity
Overall, significant variations in phreatic levels were observed among different hydroperiods (p < 0.05). These hydroperiods can be ranked based on phreatic level measurements as follows: Flood > Wet > Moist > Moderate dry (Table 3). From a habitat perspective, substantial differences in phreatic levels were identified. During the flood period, the water level was above the soil surface for mature mangrove woodland (MMW) and natural recolonization mangrove woodland (MWR), whereas it remained near the soil surface for rehabilitated mangrove woodland (RMW), early successional grass and ferns (GF), and shrub vegetation older than 50 years (SM). In the moist period, RMW exhibited the lowest phreatic levels, while GF, MWR, and SM displayed higher values, which were significantly different from the phreatic levels in other habitats (p < 0.0001). However, no significant differences in phreatic levels were observed between MMW and MWR, as well as between SM and GF during the moderate dry period and among GF, MWR, and SM during the wet period.
Furthermore, significant negative correlations between hydroperiods, phreatic levels with salinity were identified, as illustrated in Table 4 and Figure 6. Specifically, when the phreatic level was near the surface (with mean values of -0.03 ± 0.10m and -0.11 ± 0.10m), freshwater and oligohaline conditions tended to prevail. Conversely, when the phreatic level was lower, at approximately -0.44 ± 0.10m and -0.57 ± 0.08m (mean values), mesohaline and polyhaline conditions became more dominant.
Salinity conditions varied across habitats throughout different hydroperiods. During the moist and moderately dry periods, mesohaline (salinity ranging from >5.0 to 18.0 ppt) and polyhaline (salinity ranging from >18.0 to 30.0 ppt) conditions were prevalent, with significantly higher salinity levels observed across all habitats (see Figure 7). MWR, MMW, and RMW exhibited the highest mean salinity levels during the moist period. In contrast, during the wet and flood periods, salinity levels predominantly ranged from oligohaline (salinity >0.5 to 5.0 ppt) to freshwater (salinity ranging from 0 to 0.5 ppt) for all habitats.
Salinity variations between habitats across different hydroperiods were also assessed and quantified (Figure 8). The salinity differences between MMW, MWR, and RMW during the moist period, ranging from high mesohaline to polyhaline levels, were found to be statistically insignificant. However, when considered collectively, they exhibited a notable contrast with GF and SM, which featured significantly lower mesohaline salinity levels. In contrast, during the moderately dry period, SM and GF exhibited salinity conditions ranging from oligohaline to low mesohaline. These conditions were notably distinct from the mesohaline and polyhaline conditions observed in MWR and RMW. During the flood period, the freshwater salinity in MMW and MWR was significantly different from the oligohaline conditions observed in the other habitats. During this period, the phreatic level in both habitats was above the soil surface. In the wet period, the salinity in MMW, primarily low mesohaline, was significantly different from the oligohaline conditions in the other habitats.
3.2. Mesofauna diversity and abundance between habitat types
A total of 120 mesofauna morphotypes belonging to 42 different taxa were identified. The analyses revealed significant variations in both mesofauna richness and abundance among the different habitats, as illustrated in Figure 9. Among the habitats, SM and RMW exhibited the highest mesofauna richness, while GF displayed the highest mesofauna abundance. Both findings were statistically distinct from the other habitats. Conversely, no statistically significant differences in mesofauna richness were observed between GF, MMW and MWR, as well as between MMW and MWR, when considering taxa abundance.
In total, these habitats shared 27 common taxa, as depicted in Figure 10, with an estimated similarity of approximately 64%. The remaining 36% of the taxa, which were not shared, contributed to the distinctions between the habitats. These non-shared taxa included Acaridae, Bdellidae, Cheyletidae, Cryptognathidae, Eniochthoniidae, Erythraeidae, Haplozetidae, Nothridae, Rhagidiidae, Schlerobatidae, Scutacaridae, Sejidae, Stigmaeidae, Trhypocthoniidae, and Veigaiidae.
The Jaccard Index of similarity between habitats indicated that SM and RMW were the most similar, sharing 88% of the total taxa, whereas MMW and MWR exhibited somewhat lower similarity, with a shared taxa percentage of 73% (details in Figure 10 and Table 5).
Among the studied habitats, mesofauna populations distributed between litter and soil organic layers exhibited significant variations in richness for GF and RMW, as well as in abundance for SM.
Biodiversity variations, classified into dominant, common, and rare taxa, were quantified across the different habitats (see Figure 11). In the GF habitat, a total of three dominant, ten common, and twenty-two rare taxa were identified. The dominant groups in this habitat were Collembola-Arthropleona (including Brachystomellidae and Isotomidae), which made up 42.8% of the mesofauna composition, along with Oribatids (Malaconothridae) at 14.4%. MMW habitat comprised three dominant, fourteen common, and twenty rare taxa. In this habitat, Oribatids (including Damaeidae and Tegoribatidae) held dominance at 22.7%, together with Collembola-Arthropleona (Isotomidae) that accounted for 12.7% of the mesofauna composition. MWR habitat contained three dominant taxa, mainly characterized by Oribatids (Ceratozetidae, Malaconothridae, and Tegoribatidae) at 43.2%, along with twelve common and twenty-one rare taxa. RMW habitat had one dominant taxon, Oribatid-Ceratozetidae at 23%, along with seventeen common and twenty rare taxa. In the SM habitat, there were three dominant taxa, with Oribatids (Malaconothridae and Damaeidae) making up 31.1% of the composition, while Collembola-Arthropleona (Isotomidae) contributed 10.5% to the dominant groups. There were also fourteen common and twenty rare taxa identified in this habitat. Summarizing, Oribatids families, Malaconothridae, Ceratozetidae, Tegoribatidae, and Damaeidae were dominant taxa among the habitats. In addition, the uniqueness of mesofauna taxa among the habitats can be ranked as follows: GF > MWR > MMW = SM = RMW.
3.3. Influence of hydroperiod phreatic level and salinity on mesofauna diversity and abundance
Moderate correlations were estimated between hydroperiod phreatic level and mesofauna richness and abundance (Table 6). The relationship assessed was positive with richness, indicating that as the phreatic level increased, mesofauna richness tended to increase as well. Conversely, the relationship with abundance was negative, suggesting that as the phreatic level increased, mesofauna abundance tended to decrease. Additionally, a moderate negative correlation was identified between salinity and mesofauna abundance, indicating that higher salinity levels were associated with lower mesofauna abundance.
The generalized regression model revealed that habitat type, phreatic level, and salinity collectively accounted for 45% (r-square = 0.45) of the variability in mesofauna richness and 40% (r-square = 0.40) in mesofauna abundance, as indicated in Table 7. The effect analysis suggests that the most influential predictors in explaining these variations were habitat type and phreatic level. These two factors had the strongest impact on mesofauna richness and abundance in the model.
3.3.1. Hydroperiods
Variations in habitat richness and abundance across hydroperiods were observed, as illustrated in Figure 12 and Figure 13, highlighting the dynamic nature of mesofauna richness and abundance at each and between the different habitats in response to changes in hydroperiods phreatic levels.
Habitat-specific variations in mesofauna richness across different hydroperiods were quantified (Figure 12). In the GF habitat, significant differences in richness were observed among all periods. The wet period had the highest richness value, while the moist period had the lowest in contrast, in the MMW habitat, richness during the flood and wet periods showed no significant differences, but both were significantly different from the moderate dry and moist periods. MWR exhibited significantly higher richness during the wet and moderate dry periods, with no significant differences between the flood and moist periods (between floods). For RMW, there were no significant differences between the moist, moderate dry, and flood periods, which were all different from the wet period. SM displayed significantly higher richness during the moist period, which was twice as high as that of the other periods. Regarding abundance, high values were quantified for all habitats during the flood and wet periods.
Among the various habitats, the highest mesofauna richness was observed during the wet period, as shown in Figure 13. Notably, SM consistently exhibited the highest richness across all periods, with significant differences from mean values in the other habitats. Specifically, SM doubled its richness compared to other habitats during moist periods, while GF, MMW, and MWR showed richness levels that were not significantly different from each other. Moderate dry period exhibited the lowest overall mesofauna richness. During this period, there were no significant differences in richness between SM and RMW, as well as between MMW and MWR, while GF had the lowest richness.
Mesofauna abundance was higher during the wet and flood period and lower during the moderate dry and moist periods, with significant differences observed between habitats. This suggests that these hydroperiods had a substantial impact on mesofauna abundance across the different habitats (Figure 14). However, no significant differences in mesofauna abundance were found between MMW and MWR during the flood period, between SM and RMW during the moist period, and between GF and RMW as well as between MMW and SM during the wet period. A notable observation from the study is that the GF habitat exhibits a twofold increase in mesofauna abundance compared to other habitats during the flood period.
Differentiation between mesofauna taxa among hydroperiods was achieved by quantifying family prevalence and absence, as depicted in Figure 14.
The mesofauna community's prevalence and response to hydroperiod phreatic level vary significantly among different families. Ninety eight percent (98%) of all families were prevalent during the wet period, while thirty six percent (36%) were absent during disturbances, including flood and post-flood (moist) periods. Notably, the oribatid family Trhypochthoniidae was absent under conditions of extreme phreatic level fluctuations, being absent both during moderate dry periods characterized by low phreatic levels, and during flood periods characterized by high phreatic levels. Concurrently, the Scheloribatidae family exhibited a doubling in their population during the flood period.
3.3.2. Phreatic level
The variations in mesofauna richness and abundance both within and across different habitats in response to shifts in phreatic levels underscore the critical role these levels play in shaping diversity dynamics. Within individual habitats, we observed distinct patterns of species richness at different phreatic levels, as illustrated in Figure 15. The habitats of GF, MWR, and RMW exhibited higher richness at slightly moderate phreatic levels. In contrast, the MMW habitat demonstrated greater richness at high and shallow levels, while the SM habitat was more diverse at moderate levels. Moreover, higher abundances were noted at shallow levels in the SM, GF, and MMW habitats, and at slightly moderate levels for MWR and RMW.
Comparatively, when examining between habitats, the SM habitat stood out with superior mean richness during shallow, slightly moderate, moderate, and significantly deep phreatic levels (depths of -0.51 to -0.64 meters). This richness was statistically distinct compared to other habitats, as depicted in Figure 16. The high, above-surface phreatic levels were only observed in MMW and MWR habitats, with a notable difference in richness between them; MMW harbored considerably higher richness than MWR. Furthermore, the SM habitat markedly surpassed others at the moderate phreatic level, establishing itself as a biodiversity hotspot at this level. Interestingly, the slightly moderate category emerged as the most biodiverse across all habitats. However, a decline in richness was evident across all habitats at the significantly deep category, albeit with continued significant disparities in richness levels. In terms of abundance, the slightly moderate and shallow categories showcased significantly higher values across all habitats, with the GF habitat doubling its abundance in the latter category. A notable discrepancy was observed at the significantly deep phreatic level, where the RMW habitat exhibited the highest abundance, significantly differing from the GF and SM habitats. Lower abundances were recorded for all habitats at moderate and significantly deep phreatic levels.
The distribution patterns and presence or absence of various mesofauna families across distinct phreatic gradients offer a rich source of data for understanding the adaptive responses and habitat preferences of mesofauna species, as delineated in Figure 17. The data reveals a nuanced picture of the ecological dynamics at play in different environmental contexts. In conditions characterized by high and slightly moderate phreatic levels, there was a noted absence of 4.8% of the taxa, specifically within the Oribatids-Acaridae and Prostigmata Rhagidiidae families. A more substantial proportion of taxa, amounting to 28.6%, were not found in areas with high and shallow conditions locales prone to flooding or waterlogging. This category encompassed a diverse range of families including Oribatids-Cryptognathidae, Haplozetidae, Eniochthoniidae, Lohmanniidae, Eupodidae, Schlerobatidae, Suctobelbidae, Glycyphagidae, and Nothridae, as well as Prostigmata-Bdellidae, Hermanniidae, and Mesostigmata encompassing Pachylaelapidae and Veigaiidae. Conversely, a segment constituting 4.8% of the taxa exhibited a preference for slightly moderate to moderate conditions, with representation from the Prostigmata Stigmaeidae and Oribatid Trhypocthoniidae families. In moderate to significantly deep zones, which are typically drier, there was an absence of 4.8% of the taxa, including families such as Prostigmata Cheyletidae and Scutacaridae. The Prostigmata Erythraeidae family was notably absent in environments with high to significantly deep phreatic levels, indicative of extreme conditions.
3.3.3. Salinity conditions
Mesofauna richness and abundance varied within and between habitats in response to changes in salinity conditions. Within habitats, a significantly higher richness was observed in GF, MMW, MWR, and RMW at oligohaline salinities, whereas SM exhibited this increase in richness at mesohaline conditions (Figure 18). Moreover, significantly higher abundance was recorded for all habitats under oligohaline conditions. Between habitats, significant differences in richness were observed between oligohaline and mesohaline salinities, in contrast to freshwater and polyhaline conditions. SM exhibited higher richness levels at low mesohaline conditions, which were significantly different from the other habitats (Figure 19). At freshwater salinities, MMW displayed the highest richness levels, which were significantly different from those of MWR, GF and RMW. RMW and MWR were the only habitats that experienced polyhaline conditions, and at these salinity levels, they exhibited significant differences in richness. In relation to mesofauna abundance, it doubled its values at oligohaline conditions compared to freshwater, mesohaline, and polyhaline conditions, which showed no significant differences. At oligohaline salinities, there were no differences in abundance between GF and RMW, both exhibiting the highest abundance, while MMW and MWR had lower abundance levels. SM, on the other hand, exhibited significantly different abundance levels compared to the other habitats. These results suggest that mesofauna richness and abundance are influenced by salinity conditions and vary across different habitats, with RMW and MWR standing out in terms of richness under specific conditions and SM displayed unique characteristics in terms of abundance.
Quantifying the prevalence and absence of mesofauna families across various salinity levels provided valuable insights into how mesofauna species distribute and acclimate in response to varying salinity conditions, as illustrated in Figure 20. Ninety-five percent (95%) of mesofauna taxa are predominant in oligohaline and mesohaline salinities. In contrast, Prostigmata Scutacaridae and Cheyletidae are exclusive to oligohaline salinities. Additionally, twenty-four percent (24%) of the taxa can acclimate to both freshwater and polyhaline salinities. Meanwhile, twenty-five percent (25%) of the families were found in freshwater conditions, and seventeen percent (17%) were observed in polyhaline salinity. In summary, the mesofauna community exhibits a spectrum of salinity preferences and adaptability among its taxa, with some taxa being highly specialized, while others more versatile in their habitat choices.
4. Discussion
Ciénaga Las Cucharillas Natural Reserve has undergone hydrological modifications since colonial times, altering its marine-terrestrial-marine connectivity and bio-physicochemical components. Consequently, a mosaic of physicochemical conditions and habitat types characterizes these ecosystems. The observed variations in phreatic levels across different hydroperiods and habitats underscore the complex interplay between hydrological processes and habitat characteristics. The significant negative correlations between phreatic levels and salinity further elucidate the pivotal role these factors play in shaping the habitat conditions. It's important to recognize that these environmental factors are influenced by various water sources that enter the wetland, including in-situ precipitation, freshwater inputs, and seawater flows [18].
Mesohaline and polyhaline conditions are favored when the phreatic level is moderate and significantly deep, as observed during the moist and moderate dry periods. During these periods, a bimodal high tide reaches the study site in just 20 minutes. Conversely, freshwater and oligohaline conditions occur at higher and near surface phreatic levels, particularly during the flood and wet periods, when it takes up to 2 hours for the bimodal high tide to reach the study site [36]. This tidal interaction in this wetland occurs via deep subsurface flow and causes notable fluctuations in substrate salinity concentrations within habitats. The moderated dry and moist periods are influenced by marine intrusion/tides and the flood and wet periods by freshwater input by precipitation and runoff. This periodic fluctuations in phreatic level and salinity exert a substantial influence on mesofauna dynamics and distribution patterns within and between habitats (Figure 21), underscoring their sensitivity to changes of these environmental factors.
During both the wet and flood periods, we observed the peak of mesofauna richness and abundance, respectively, across various habitats (Figure 22). Remarkably, a vast majority of mesofauna families flourished during these phases, accounting for 98% during the wet period and 74% during the flood period. This prevalence suggests that the habitat conditions of the wet period, characterized by a moderately low phreatic level (-0.12 ± 0.03 m) and oligohaline salinity (1.61 ± 1.07 ppt), foster an optimal microenvironment for a diverse array of mesofauna families. A plausible explanation for this trend is the series of environmental changes preceding the wet period. After enduring alternating flood and dry conditions for several months, factors such as substrate moisture, salinity, and food availability undergo significant shifts across habitats [16,48,49]. In wetland ecosystems, terrestrial plant litter accumulates during dry spells and undergoes partial in-situ decomposition, enriching the humic material reservoir. The onset of flooding leads to the death of some terrestrial plants, while aquatic plants sprout, grow, and eventually decay, further augmenting the mixture and quantity of detrital accumulation. As floodwaters recede, transitioning into the wet period, the mixture and exposure of these organic materials creates zones ripe for recolonization, enhancing the activity of microflora [50] and paving the way for a resurgence of mesofauna. This resurgence includes fungivorous entities such as oribatid mites and Collembola, as well as their predators, including Mesostigmata and Prostigmata mites [10]. An exemplification of this recolonization is observed in the Oribatid Scheloribatidae family, which exhibited a doubling in their population during the flood sampling period, coinciding with the receding of floodwaters.
The lowest richness and abundance observed during the moderate dry hydroperiod indicate that the prevailing conditions, which include deeper phreatic levels ( -0.46 ± 0.06) and moderate mesohaline to polyhaline salinities (10.53 ± 2.97 ppt, 18.94 ± 0.48 ppt) across the different habitats, had a substantial impact on mesofauna distribution and dynamics.
Between habitat-specific variations in richness and abundance, the mature 50-year shrubland of Dalbergia ecastaphyllum (L.) (SM) emerges as a distinctive environment, consistently showcasing the highest richness throughout all observed periods (Figure 22). This heightened richness is likely fostered by the unique characteristics of the SM habitat, including a moderate phreatic level (mean value= -0.22 ± 0.16 m), the presence of oligohaline to low mesohaline salinities, canopy closure providing shade to the understory and soil, and enhanced litter inputs. These factors collectively cultivate a microenvironment conducive to a diverse mesofauna community, instigating shifts in the population dynamics [50,51]. During the moist period, which occurs between flood events, the SM habitat exhibited a richness that was twice that of other habitats. This period was characterized by a moderate phreatic level and a prevailing low mesohaline salinity of 8.32 ± 2.09 ppt. The elevated richness during this period can be attributed to the higher micro-elevation of the SM habitat, which acts as a buffer against the high phreatic levels typically experienced during flood disturbances, a protection not afforded to other habitats especially MMW and MWR that had similar microenvironemnts. This distinctive microenvironment fosters conditions favorable for the colonization by a variety of mesofauna taxa that are well-adapted to predominantly mesohaline salinities [19,23]. Furthermore, it is noteworthy that during this moist period, the SM habitat maintained a relatively stable mesofauna community, as evidenced by the lack of significant fluctuations in the richness and abundance of key groups such as Oribatids, Mesostigmata, and Collembolas. This stability suggests a resilient community structure, capable of withstanding the environmental changes occurring during this period and underscores the critical role of the SM habitat in harboring a rich and stable mesofauna community. This finding highlights the importance of understanding the unique characteristics and dynamics of different habitats in fostering biodiversity in tropical urban coastal wetlands.
Significant variability exists in how different mesofauna taxa respond to phreatic levels and salinity, suggesting distinct tolerances to these environmental factors. Both Spearman's Rho correlation analysis and the generalized regression model offer empirical evidence of the significant impact of phreatic level and salinity on mesofauna richness and abundance. These diverse responses among taxa can be attributed to various factors, including life cycle variants, physiological speciation, and behavioral adaptations.
Significantly, around thirty-six percent (36%) of the mesofauna families were absent during periods of marked shifts in phreatic levels, including the flood and subsequent post-flood (moist) phases. This absence underscores the sensitivity of a considerable portion of mesofauna families to such hydrological disturbances. A case in point is the oribatid family Trhypocthoniidae, which exhibited a pronounced sensitivity to extreme fluctuations in phreatic levels. This family was conspicuously absent not only during the moderate dry periods, characterized by deep phreatic levels, but also during the flood periods where high phreatic levels prevail. This pattern of absence across a spectrum of extreme phreatic conditions highlights the vulnerability of Oribatid Trhypocthoniidae to substantial variations in hydroperiods, pointing to a narrow ecological amplitude with regard to phreatic level tolerances.
Regarding salinity preferences, Oribatids dominate across all salinity conditions (Figure 23). Collembola taxa show increased richness and abundance at oligohaline and freshwater salinities, while Mesostigmata and Prostigmata increase at oligohaline and mesohaline conditions. However, Prostigmata families such as Scutacaridae and Cheyletidae are exclusive to oligohaline salinities, indicating their specialization in this salinity range.The mesofauna community's prevalence and response to fluctuations in salinity conditions exhibit significant variations among different families. The majority of mesofauna families (ninety-five percent) thrive in oligohaline and mesohaline salinities. A portion of the taxa (twenty-four percent), which includes families like Oribatids Damaeidae, Mesostigmata Laelapidae, Prostigmata Cunaxidae, and Collembola Isotomidae, demonstrates adaptability to both freshwater and polyhaline salinities, showcasing their ability to acclimate to a broader salinity spectrum. Approximately twenty-five percent of mesofauna families were found in freshwater conditions (notably families such as Oribatids Ceratozetidae, Mesostigmata Digamasellidae, Prostigmata Cunaxidae, and Collembola Sminthuridae), and seventeen percent were observed in polyhaline salinity, with dominant families being Oribatids Galumnidae and Suctobelbidae, Mesostigmata Blattisociidae, Prostigmata Eupodidae, and Collembola Isotomidae.
The findings delineated above unveil a complex interplay of inter- and intra-specific responses to the fluctuating phreatic levels and salinity conditions prevalent during the hydroperiods of wetland habitats. This underscores the intricate adaptations and ecological strategies that various mesofauna families have evolved to acclimate to the periodic landscape dynamics inherent to tropical urban coastal wetlands. Consequently, this research offers profound insights into the community structures and operational dynamics of mesofauna in these ecosystems, elucidating their resilience and adaptability in the face of environmental variations.
5. Conclusions
In conclusion, the study underscores the significance of understanding mesofauna's intricate inter- and intra-specific responses to fluctuations in wetland hydroperiod phreatic levels and salinity conditions. The diverse range of responses exhibited by the mesofauna community in this wetland emphasizes their varying degrees of adaptability and resilience to changes in their microenvironment. This insight into mesofauna community dynamics is invaluable for effective wetland management, particularly in the context of multiple stressors such as global and regional climate change, sea level rise, and human activities. The bio-sensor capacity of soil mesofauna emerges as a crucial tool for monitoring and adaptive ecosystems management to ensure their long-term health and sustainability.
Author Contributions
GO-R and EC conceived and designed the study, analyzed the data, and wrote the manuscript. GO-R, EH, and SP-P field sampling and analysis. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the NSF CREST – Center for Innovation Research and Education in Environmental Nanotechnology (CIRE2N) HRD 1736093, NSF HRD 1806129, and the Center for Applied Tropical Ecology and Conservation (CATEC) of the University of Puerto Rico.
Data Availability Statement
The authors commit to making the raw data that supports the conclusions of this article available without undue reservation.
Acknowledgments
The authors would like to acknowledge the technical assistance provided by the Center for Applied Tropical Ecology and Conservation (CATEC). Appreciation is also extended to the coordinator of the Process and Functions Ecosystem Lab, Larry Díaz, and to the undergraduate students — Diego E. Otero-Santiago, Cecilia M. Fadhel-Alvarez, Jorge L. Gonzalez-Santana, Thalia M. Rosa-Agosto, Alexis Roque Sotomayor, and Alexander Cántres — for their substantial contributions to both field and laboratory work.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Lavelle, P.; Spain, V.A. Soil Ecology. Kluwer Academic Publishers: New York, Boston, Dordrecht, London, Moscow, 2003.
- Mulder, C.; Den Hollander, H. A.; Vonk, J. A.; Rossberg, A. G.; op Akkerhuis, G. A. J.; Yeates, G. W. Soil resource supply influences faunal size–specific distributions in natural food webs. Naturwissenschaften 2009, 96, 813–813. [Google Scholar] [CrossRef] [PubMed]
- Swift, M. J.; Heal, O. W.; Anderson, J. M.; Anderson, J. M. Decomposition in terrestrial ecosystems, Vol. 5. Univ of California Press: 1979.
- Coleman, D. C.; Callaham, M.; Crossley Jr, D. A. Fundamentals of soil ecology. Academic Press: 2017.
- Culliney, T.W. Role of Arthropods In Maintaining Soil Fertility. Plant Epidemiology and Risk Analysis Laboratory, Plant Protection, and Quarantine, Center for Plant Health Science and Technology, USDA-APHIS. Agriculture 2013, 3, 629–659. [CrossRef]
- Lavelle, P.; Blanchart, E.; Martin, A.; Martin, S.; Alister, S. A Hierarchical Model for Decomposition in Terrestrial Ecosystems: Application to Soils of the Humid Tropics. 2013, 25, 130–150.
- Wardle, D. A.; Bardgett, R. D.; Klironomos, J. N.; Setälä, H.; Van Der Putten, W. H.; Wall, D. H. Ecological linkages between aboveground and belowground biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef] [PubMed]
- Collins, W. W.; Qualset, C. O. (Eds.). Biodiversity in agroecosystems. CRC Press: 1998.
- Chaos of Delight. Soil animals-springtails, soil mites and mesofauna. Available online: https://www.chaosofdelight.org/.
- Kagainis, U.; Jucevica, E.; Salmane, I.; Ventins, J.; Melecis, V. Does Climate Warming Affect Soil Mesofauna? Available online: https://www.researchgate.net/publication/320559698_Does_Climate_Warming_Affect_Soil_Mesofauna.
- Ghiglieno, I.; Simonetto, A.; Orlando, F.; Donna, P.; Tonni, M.; Valenti, L.; Gilioli, G. Response of the Arthropod Community to Soil Characteristics and Management in the Franciacorta Viticultural Area (Lombardy, Italy). Agronomy 2020, 10, 740. [Google Scholar] [CrossRef]
- Socarrás, A. Mesofauna edáfica: indicador biológico de la calidad del suelo. Pastos y forrajes 2013, 36, 5–13. [Google Scholar]
- Bezemer, T. M.; Fountain, M. T.; Barea, J. M.; Christensen, S.; Dekker, S. C.; Duyts, H.; Van der Putten, W. H. Divergent composition but similar function of soil food webs of individual plants: plant species and community effects. Ecology 2010, 91, 3027–3036. [Google Scholar] [CrossRef] [PubMed]
- Haarlov, N. Vertical Distribution of Mites and Collembola in Relation to Soil Structure. In Soil Zoology; Mc Kevan, D.K.E., Ed.; Butter Worths: London, 1955; pp. 167–179. [Google Scholar]
- Liu, Y.; Du, J.; Xu, X.; Kardol, P.; Hu, D. Microtopography-induced ecohydrological effects alter plant community structure. Geoderma 2020, 362, 114119. [Google Scholar] [CrossRef]
- Batzer, P. D. ; Sharitz,R.R. Ecology of Freshwater and Estuarine Wetlands. The University of California: 2006.
- Kim, J.; Lee, J.; Cheong, T.; Kim, R.; Koh, D.; Ryu, J.; Chang, H. Use of time series analysis for the identification of tidal effect on groundwater in the coastal area of Kimje, Korea. Journal of Hydrology 2005, 300, 188–198. [Google Scholar] [CrossRef]
- Hernández, E. Ecophysiological responses of plant functional groups to environmental conditions in a coastal urban wetland, Ciénaga Las Cucharillas in Northeastern Puerto Rico. Ecolab. Department of Environmental Science. The University of Puerto Rico: 2022.
- Batzer, P. D.; Wu, H. Ecology of Terrestrial Arthropods in Freshwater Wetlands. Annual Reviews of Entomology 2020. Available online: www.annualreviews.org.
- Krediet, A.F.; Ellers, J.; Berg, M.P. Collembola community contains larger species in frequently flooded soil. Pedobiologia 2023. Available online. [CrossRef]
- van Dijk, J.; Didden, W. A.; Kuenen, F.; van Bodegom, P. M.; Verhoef, H. A.; Aerts, R. Can differences in soil community composition after peat meadow restoration lead to different decomposition and mineralization rates? Soil Biology and Biochemistry 2009, 41, 1717–1725. [Google Scholar] [CrossRef]
- Pereira, C.S.; Lopes, I.; Sousa, J.P.; Chelinho, S. Effects of NaCl and seawater induced salinity on survival and reproduction of three soil invertebrates species. Chemosphere 2015, 135, 116–122. [CrossRef]
- Batzer, D.; Wu, H.; Wheeler, T.; Eggert, S. Peatland invertebrates. In: Invertebrates in freshwater wetlands; 2016; pp. 219–250.
- Bardgett, R. D.; Yeates, G. W.; Anderson, J. M. Patterns and determinants of soil biological diversity. In: Biological diversity and function in soils; 2005; pp. 100–118.
- Barberena-Arias, M. F.; Cuevas, E. Physicochemical Foliar Traits Predict Assemblages of Litter / Humus Detritivore Arthropods. 2018, 1–20.
- Mazhar, S.; Pellegrini, E.; Contin, M.; Bravo, C.; De Nobili, M. Impacts of salinization caused by sea level rise on the biological processes of coastal soils - A review. Frontiers in Environmental Science 2022, 10. Available online: https://www.frontiersin.org/articles/10.3389/fenvs.2022.909415.
- Wu, P.; Zhang, H.; Wang, Y. The response of soil macroinvertebrates to alpine meadow degradation in the Qinghai–Tibetan Plateau, China. Applied Soil Ecology 2015, 90, 60–67. [Google Scholar] [CrossRef]
- National Weather Service. Climatological Data for TOA BAJA LEVITTOWN, PR – Year 2020 to 2021. Available online: https://www.weather.gov/wrh/climate?wfo=sju (accessed on 9 September 2023).
- Kennaway, T.; Helmer, E. H. The forest types and ages cleared for land development in Puerto Rico. GIScience Remote Sens. 2007, 44, 356–382. [Google Scholar] [CrossRef]
- Pumarada-O’Neill, L. Los Puentes Históricos de Puerto Rico. Centro de Investigación y Desarrollo, Recinto de Mayagüez, Universidad de Puerto Rico: Mayagüez, 1991.
- Webb, R. M.; Gómez-Gómez, F. Synoptic Survey of Water Quality and Bottom Sediments, San Juan Bay Estuary System, Puerto Rico, December 1994-July 1995. Water Resources Investigations Report. U.S. Geological Survey: Denver, CO, 1998. Available online, 19 December. [CrossRef]
- Cuevas, E. Personal communication. Ecolab. University of Puerto Rico: Rio Piedras, February 2023.
- Branoff, B.; Cuevas, E.; Hernández, E. Assessment of Urban Coastal Wetlands Vulnerability to Hurricanes in Puerto Rico. DRNA: San Juan, 2018. Available online: http://drna.pr.gov/wp-content/uploads/2018/09/FEMA-Wetlands-Report.pdf (accessed on 3 August 2023).
- United States Department of Agriculture (USDA). NRCS Web Soils Survey. Ciénaga las Cucharillas. Available online: https://websoilsurvey.usda.gov/ (accessed on 16 June 2023).
- Chen, Y.; Wang, B.; Pollino, C. A.; Cuddy, S. M.; Merrin, L. E.; Huang, C. Estimate of flood inundation and retention on wetlands using remote sensing and GIS. Ecohydrology 2014, 7, 1412–1420. [Google Scholar] [CrossRef]
- Hernández, E. Personal communication. Ecolab. University of Puerto Rico: Rio Piedras, June 2021.
- Weather.gov. Tropical Storm Isaias - July 29-31, 2020. US Dept of Commerce. National Oceanic and Atmospheric Administration. National Weather Service. Available online: https://www.weather.gov/sju/isaias2020 (accessed on 16 June 2023).
- Weather.gov. August 21-23, 2020. US Dept of Commerce. National Oceanic and Atmospheric Administration. National Weather Service. Available online: https://www.weather.gov/sju/laura2020 (accessed on 16 June 2023).
- Barberena-Arias, M. F. Single tree species effects on temperature, nutrients, and arthropod diversity in litter and humus in the Guánica dry forest. Ph.D. diss., The University of Puerto Rico: 2008.
- USDA Soil Quality Institute. Soil Quality Test Kit. Agricultural Research Service. Natural Resources Conservation Service: 1999.
- Zheng, X.; Wang, H.; Tao. Y.; Kou, X.; He, C.; Wang, Z. Community diversity of soil meso-fauna indicates the impacts of oil exploitation on wetlands. Ecological Indicators 2022, 144, 109451. Available online: https://www.sciencedirect.com/science/article/pii/S1470160X22009244.
- Li, K.; Bihan, M.; Yooseph, S.; Methé, B.A. Analyses of the microbial diversity across the human microbiome. PLoS One 2012, 7, e32118. [Google Scholar] [CrossRef] [PubMed]
- Daghighi, E.; Koehler, H.; Kesel, R.; Filser, J. Long-term succession of Collembola communities in relation to climate change and vegetation. Pedobiologia 2017, 64, 25–38. [Google Scholar] [CrossRef]
- Kiernan, D. Natural Resources Biometrics. Open SUNY. Milne Library (IITG PI). State University of New York at Geneseo: Geneseo, NY, 2014. Available online: https://open.umn.edu/opentextbooks/textbooks/205.
- Herrera, F. Artrópodos del suelo como bioindicadores de recuperación de sistemas perturbados. Venesuelos 2003, 11, 67–78. [Google Scholar]
- Environmental Protection Agency. Chapter 14 of the Volunteer Estuary Monitoring Manual, A Methods Manual, Second Edition, EPA-842-B-06-003. 2006.
- Lugo, A. E.; Medina, E.; Cuevas, E.; González, O. R. Ecological and physiological aspects of Caribbean shrublands. Caribbean Naturalist 2019, 58, 1–38. [Google Scholar]
- Cordes, P.; Maraun, M.; Schaefer, I. Dispersal patterns of oribatid mites across habitats and seasons. Experimental and Applied Acarology 2022, 86. [Google Scholar] [CrossRef]
- Walter, D.E.; Proctor, H.C. Life Cycles, Development and Size. In Mites: Ecology, Evolution & Behaviour; Springer: Dordrecht, 2013. [Google Scholar] [CrossRef]
- Heydari, M.; Eslaminejad, P.; Kakhki, F. V.; Mirab-balou, M.; Omidipour, R.; Prévosto, B.; Lucas-Borja, M. E. Soil quality and mesofauna diversity relationship are modulated by woody species and seasonality in semiarid oak forest. Forest Ecology and Management 2020, 473, 118332. [Google Scholar] [CrossRef]
- Barrios, E.; Sileshi, G.; Shepherd, K.; Sinclair, F. Agroforestry and Soil Health: Linking Trees, Soil Biota, and Ecosystem Services. 2012. [CrossRef]
Figure 1.
Historical changes in costal wetland land use cover since colonial times followed by abandonment, modified its hydrology and marine-terrestrial-marine connectivity.
Figure 1.
Historical changes in costal wetland land use cover since colonial times followed by abandonment, modified its hydrology and marine-terrestrial-marine connectivity.
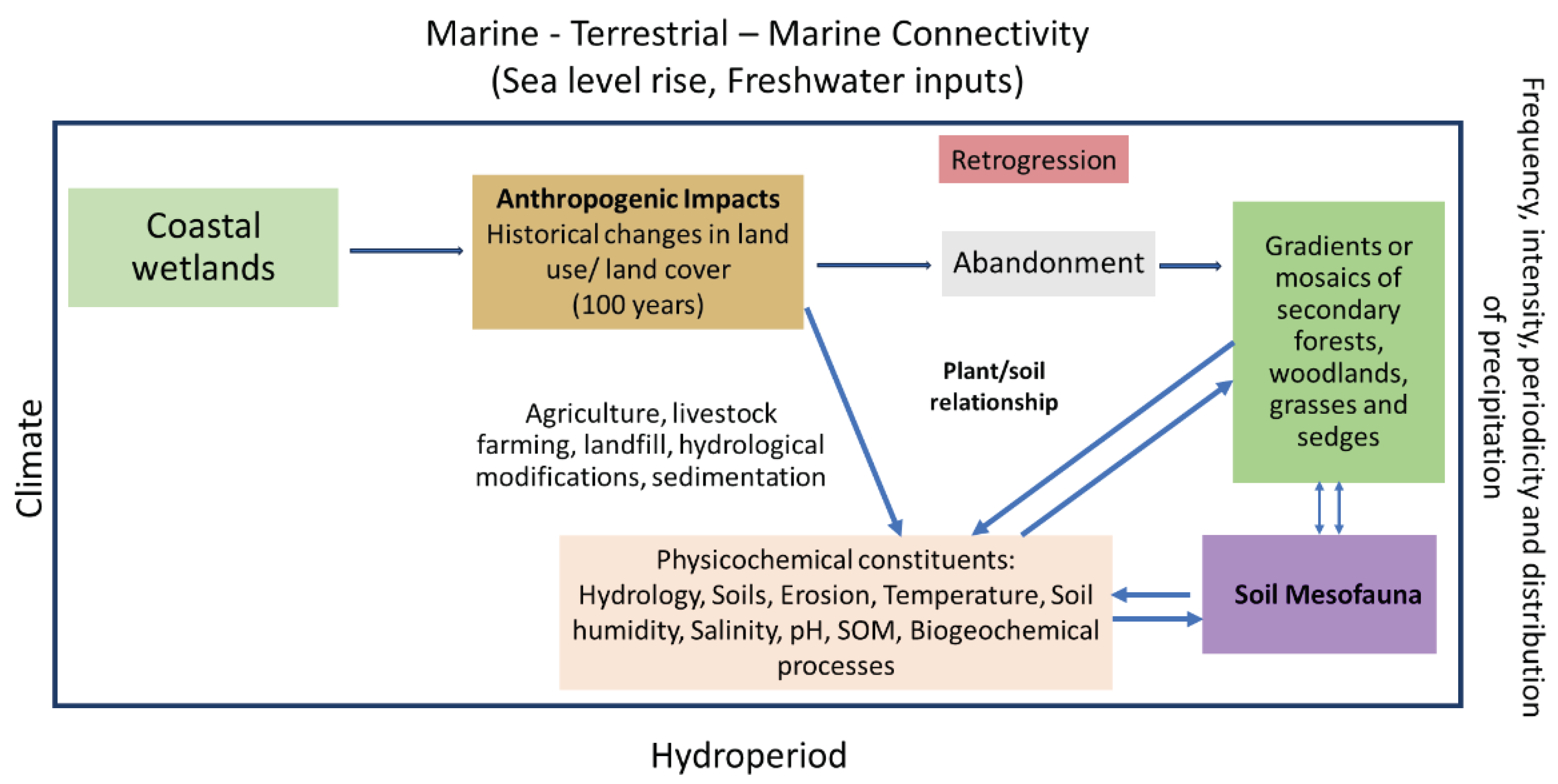
Figure 2.
A) Ciénaga las Cucharillas located in the northwestern side of the San Juan Bay, B) Study area (2.2ha), and C) Study Plots 3, 5, 10 and 6.
Figure 2.
A) Ciénaga las Cucharillas located in the northwestern side of the San Juan Bay, B) Study area (2.2ha), and C) Study Plots 3, 5, 10 and 6.
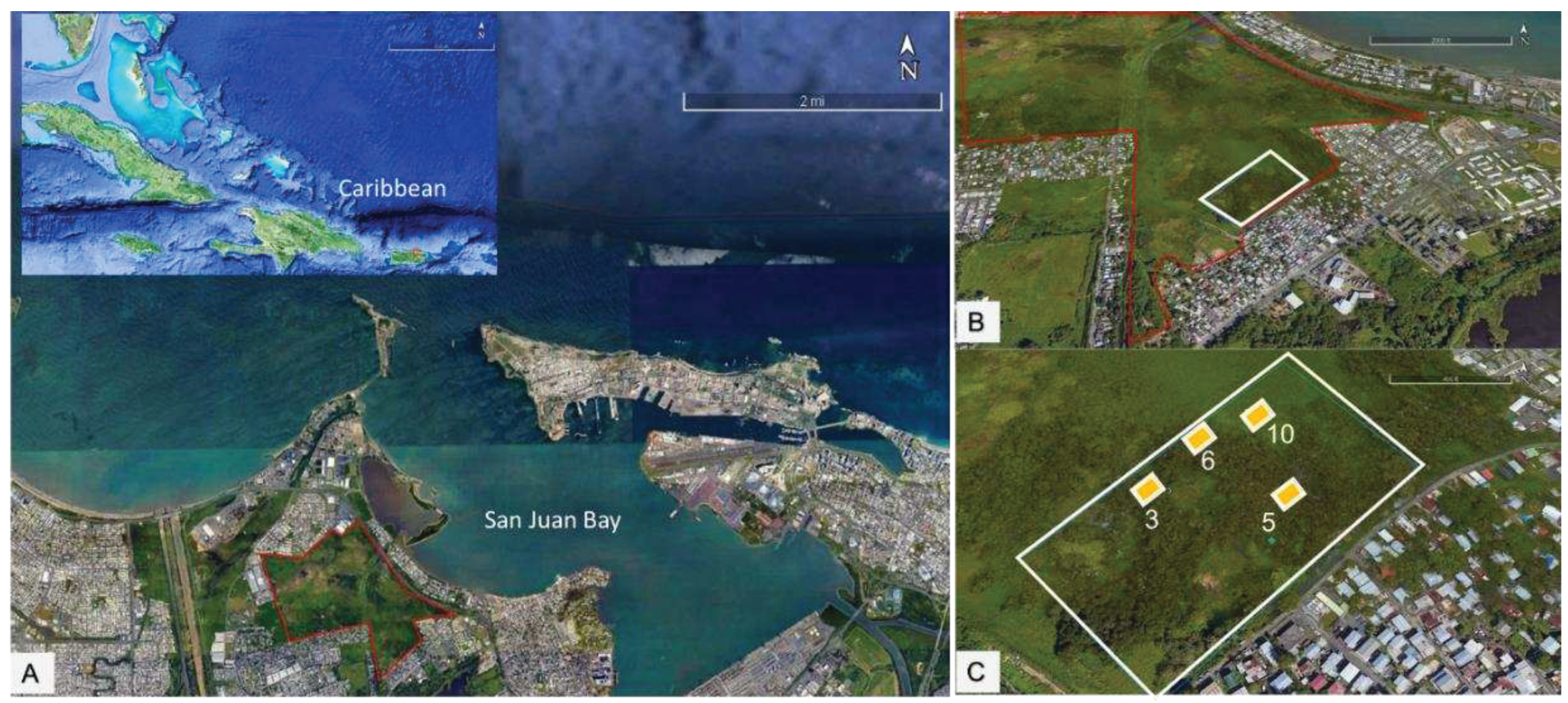
Figure 3.
a) Climate diagram of the study area; b) Mean monthly precipitation and total rainy days (January 2020 - November 2021) climatological data for Toa Baja Levittown, PR Meteorological Station (NOAA National Weather Service, 2021).
Figure 3.
a) Climate diagram of the study area; b) Mean monthly precipitation and total rainy days (January 2020 - November 2021) climatological data for Toa Baja Levittown, PR Meteorological Station (NOAA National Weather Service, 2021).
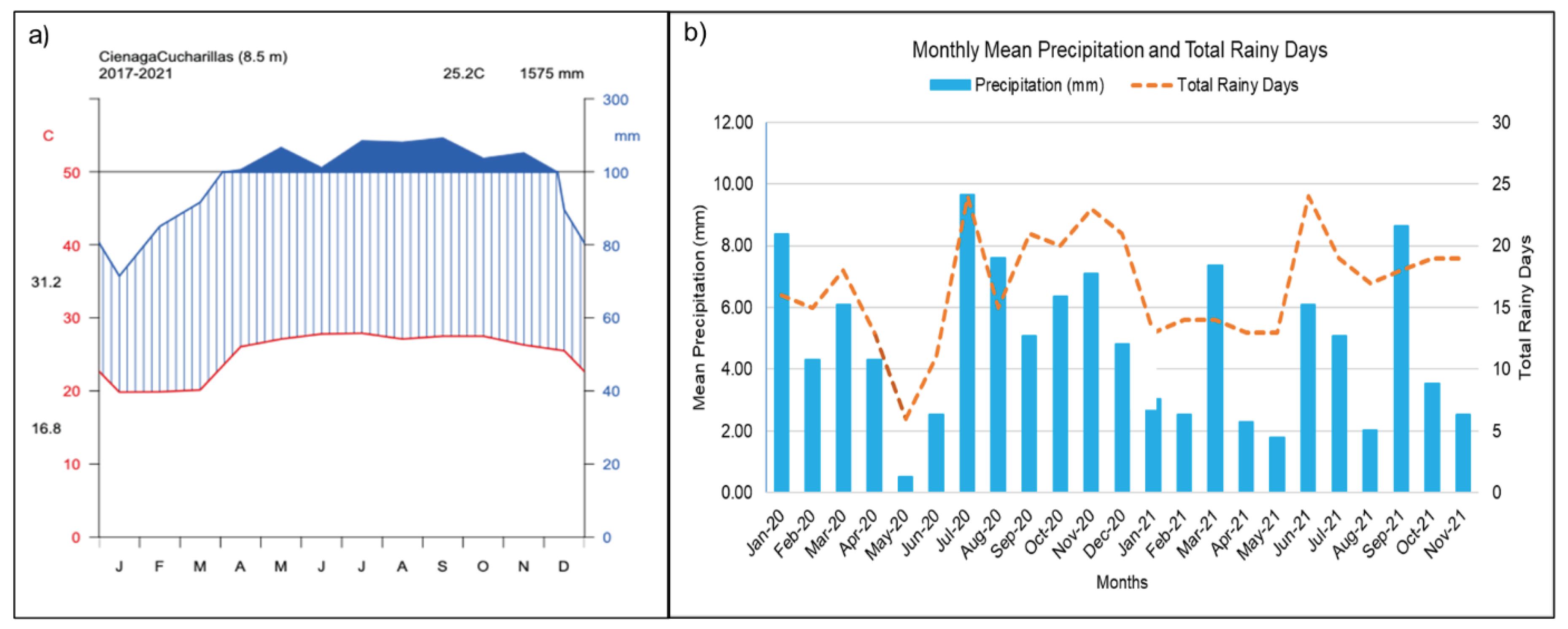
Figure 4.
La Malaria flood control channel, located northwest of the research area, and the outflow water pump structure at the mouth of the channel.
Figure 4.
La Malaria flood control channel, located northwest of the research area, and the outflow water pump structure at the mouth of the channel.
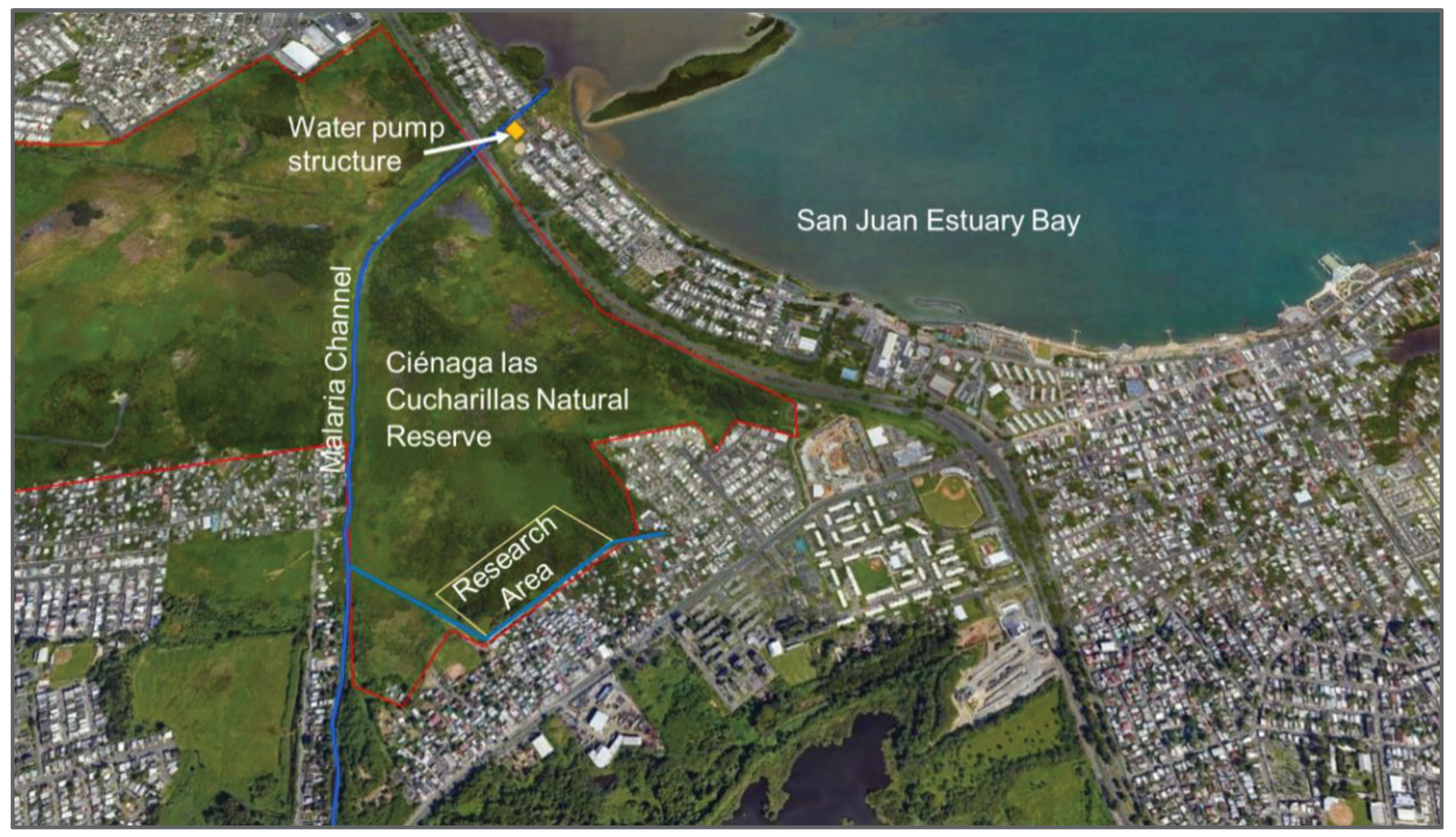
Figure 5.
Schematic diagram of hydroperiod condition on sampling date, total rainy days (cm) and mean phreatic level (m) 14 days before and at the sampling date. These variations strongly influenced litter system patterns at the sampling date.
Figure 5.
Schematic diagram of hydroperiod condition on sampling date, total rainy days (cm) and mean phreatic level (m) 14 days before and at the sampling date. These variations strongly influenced litter system patterns at the sampling date.
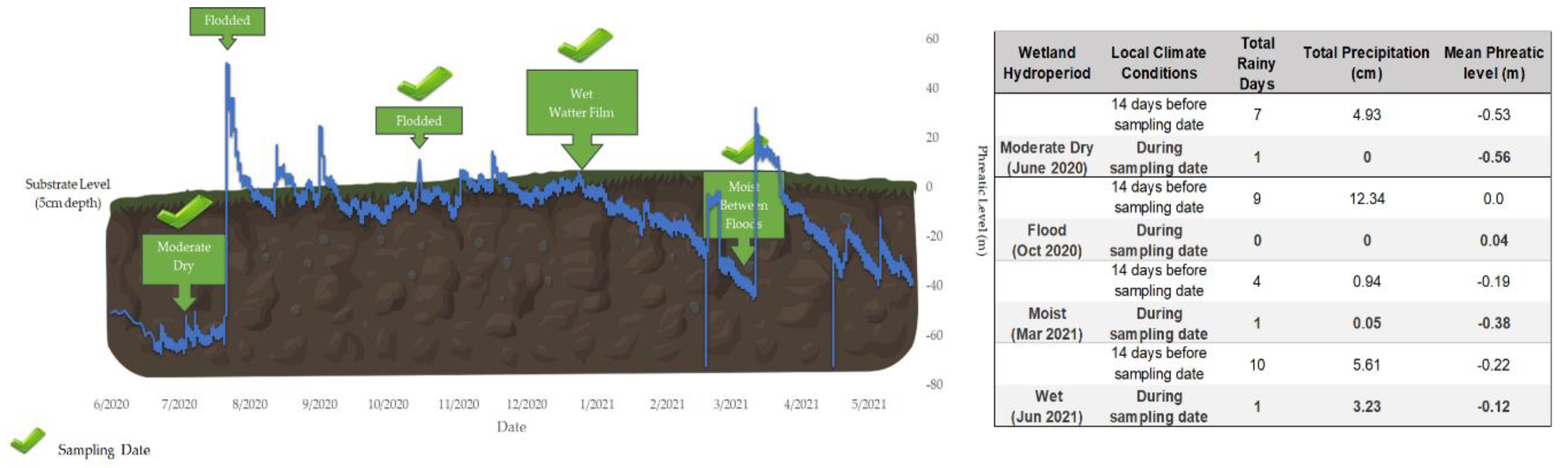
Figure 6.
Variations in salinity conditions correlated with hydroperiod phreatic level.
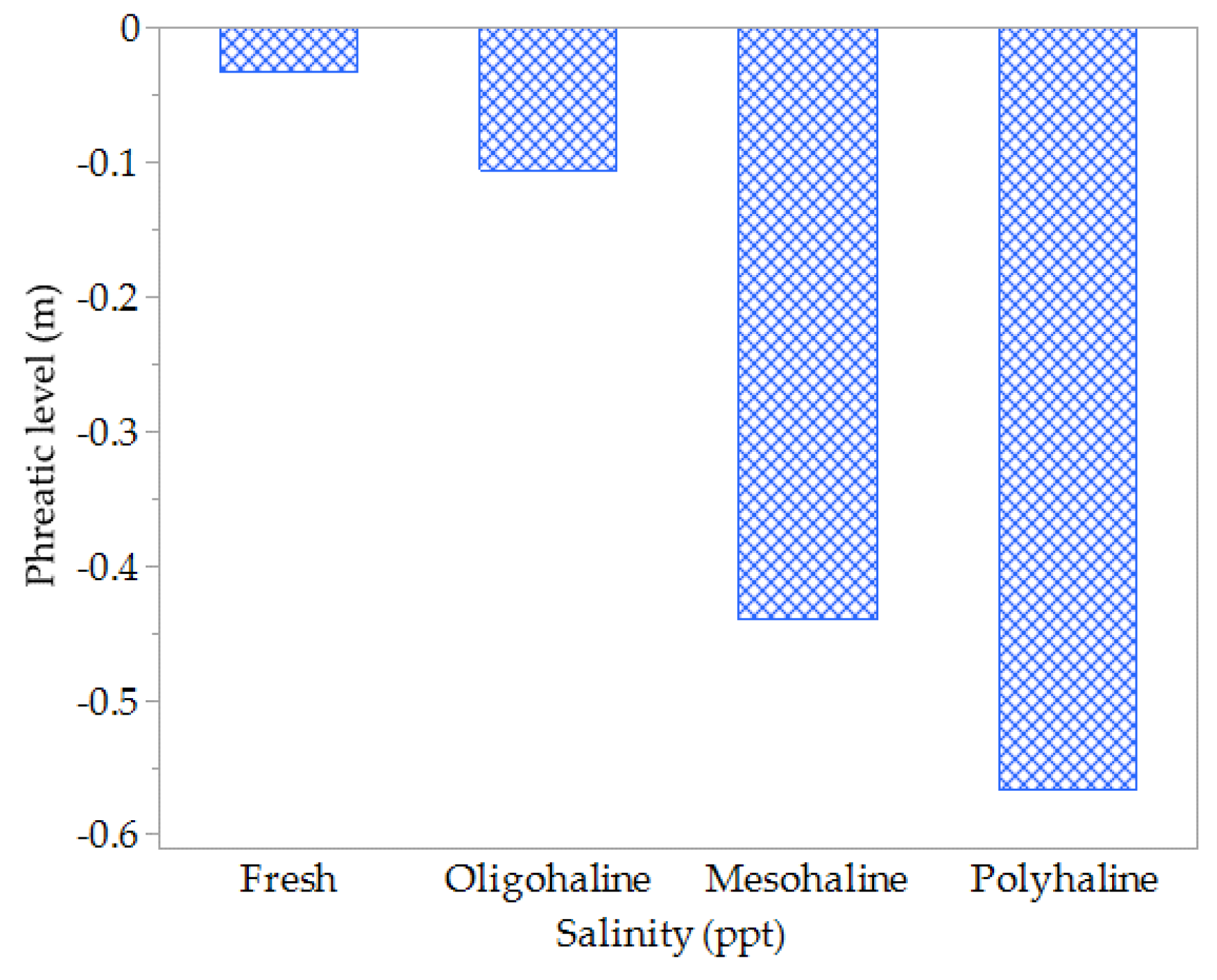
Figure 7.
Significant differences of habitats salinity among hydroperiods. Values with different letters indicates means with significant differences (p<.05).
Figure 7.
Significant differences of habitats salinity among hydroperiods. Values with different letters indicates means with significant differences (p<.05).
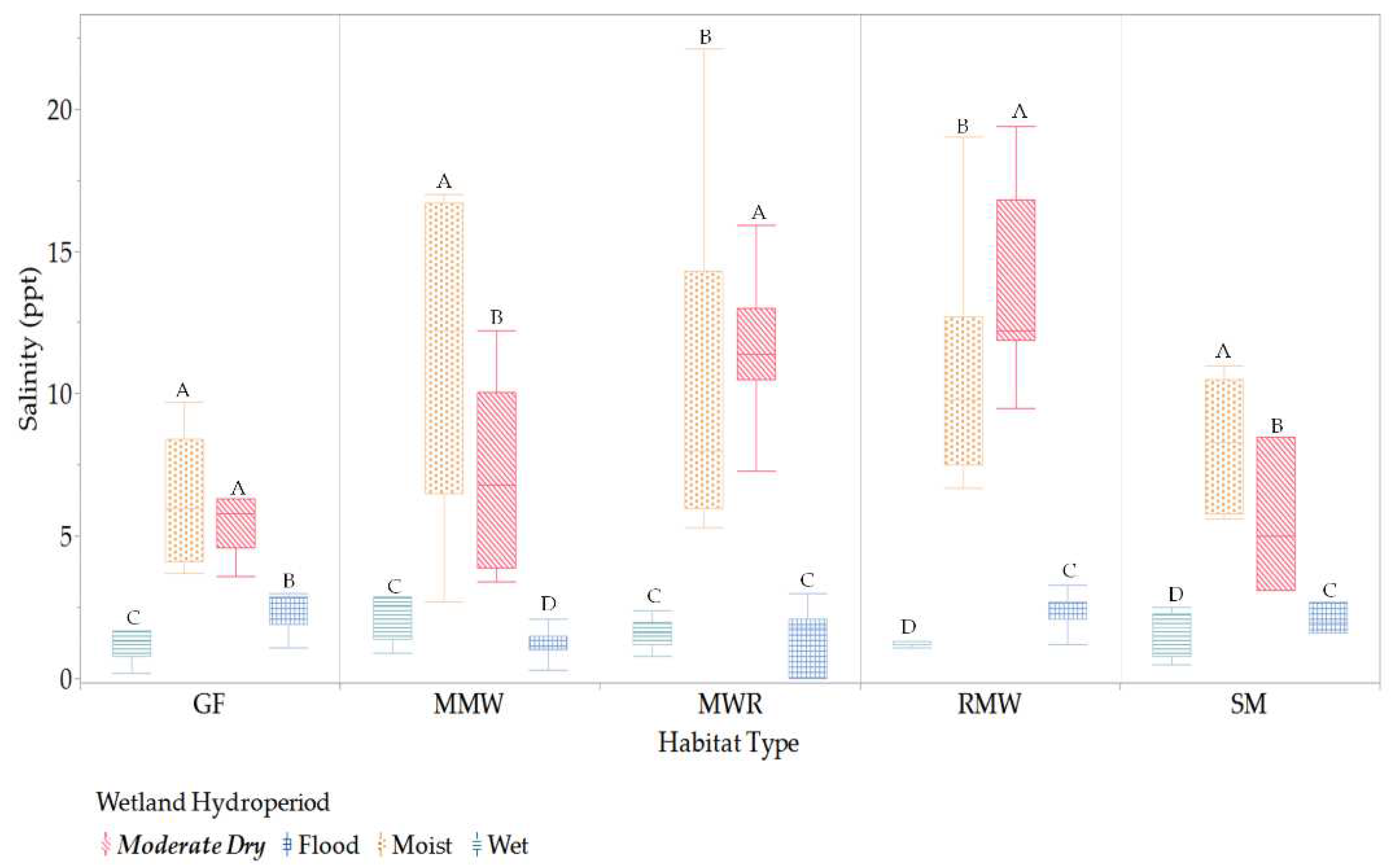
Figure 8.
Significant differences between habitats salinity per hydroperiods. Values with different letters indicates means with significant differences (p<.05).
Figure 8.
Significant differences between habitats salinity per hydroperiods. Values with different letters indicates means with significant differences (p<.05).
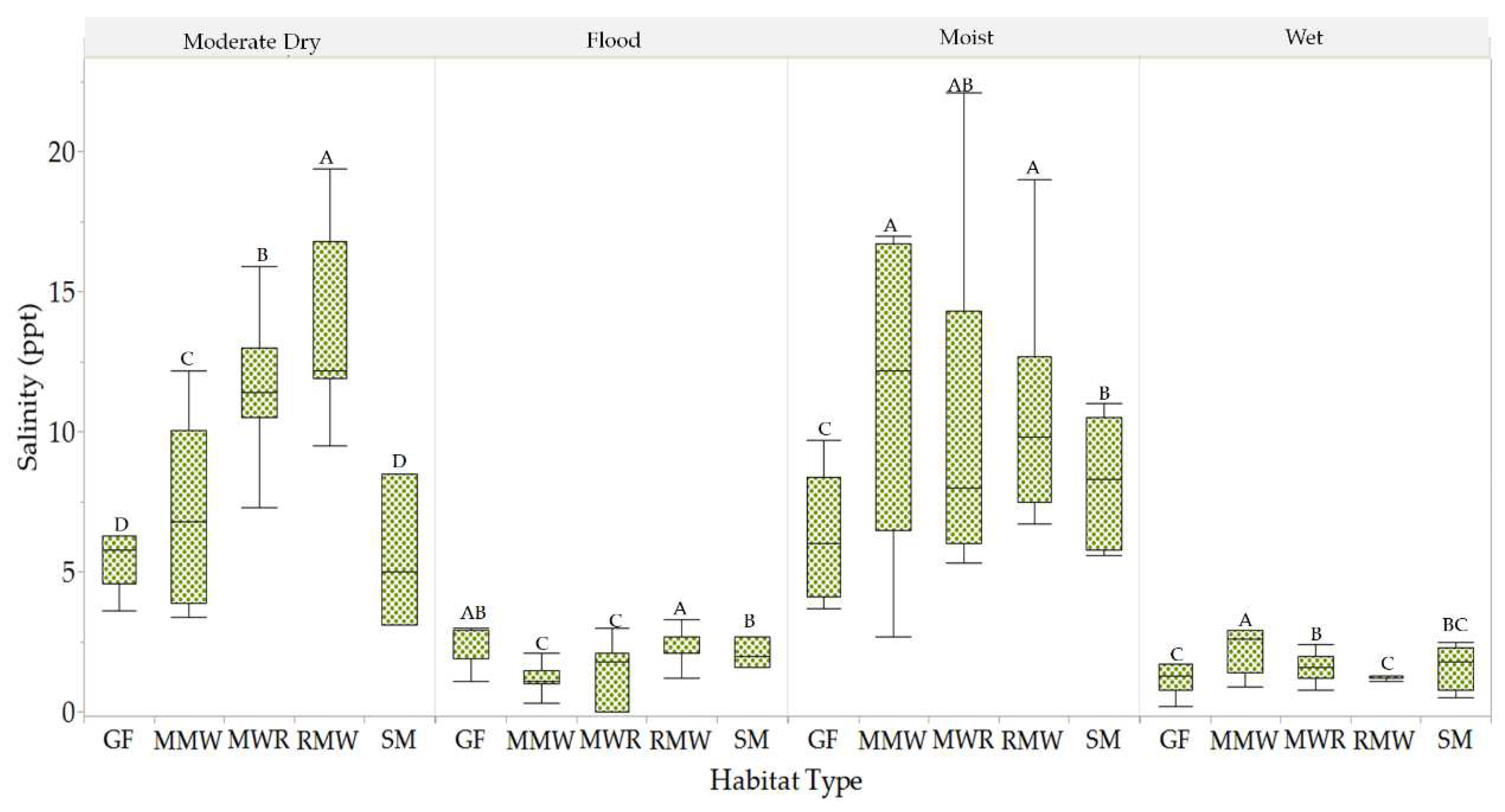
Figure 9.
Significant differences in mesofauna richness and abundance between habitats. Values with different letters indicates means with significant differences (p<.05).
Figure 9.
Significant differences in mesofauna richness and abundance between habitats. Values with different letters indicates means with significant differences (p<.05).
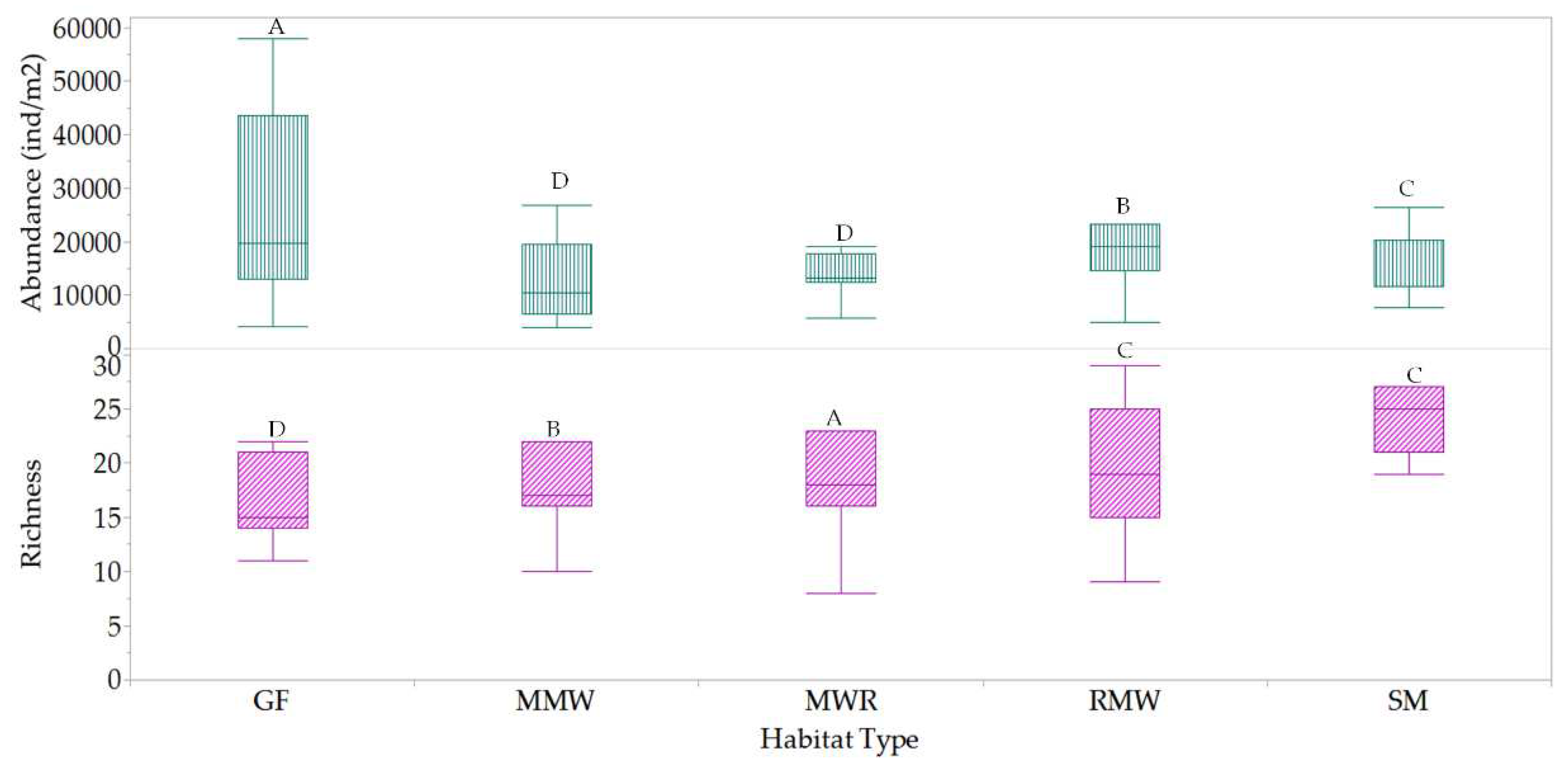
Figure 10.
Venn diagram showing the number of A) shared and B) not shared taxa among habitat types.
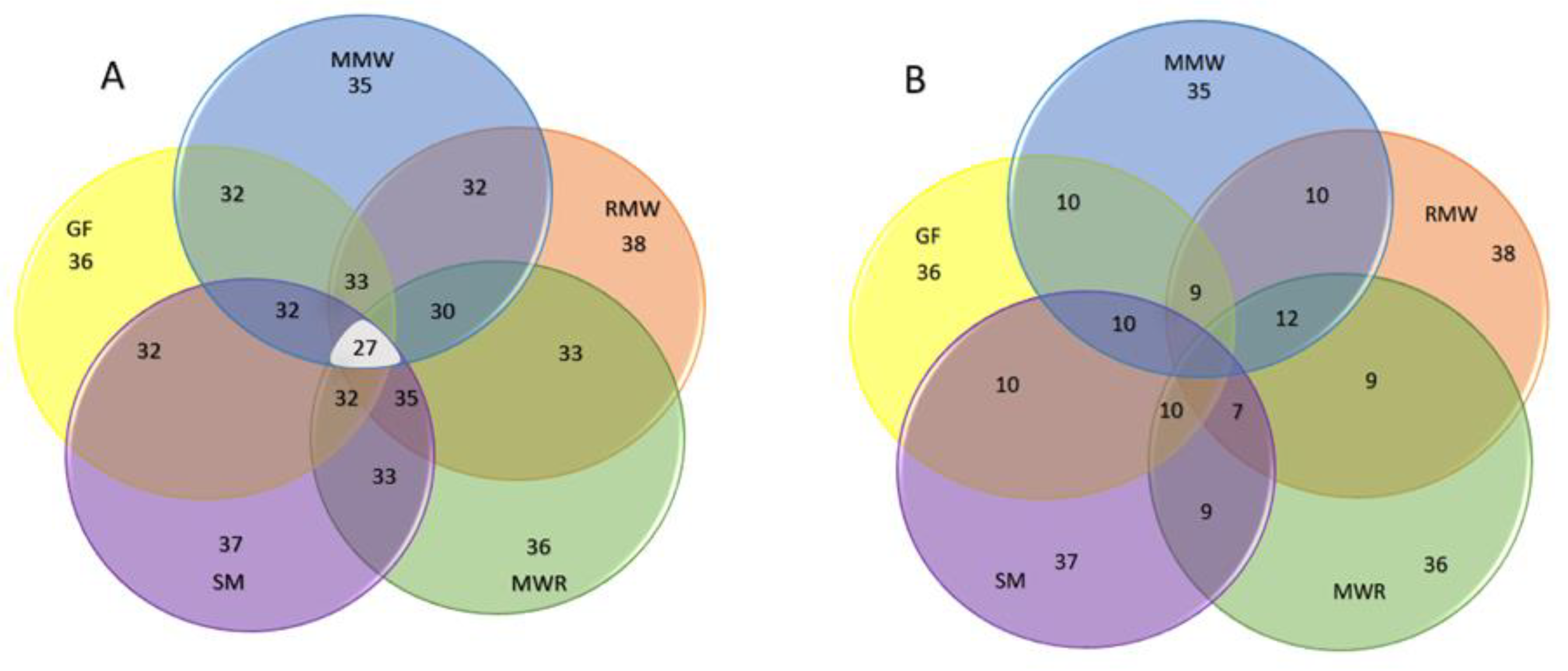
Figure 11.
Mesofauna dominant, common, and rare taxa composition by habitat [43].
Figure 11.
Mesofauna dominant, common, and rare taxa composition by habitat [43].
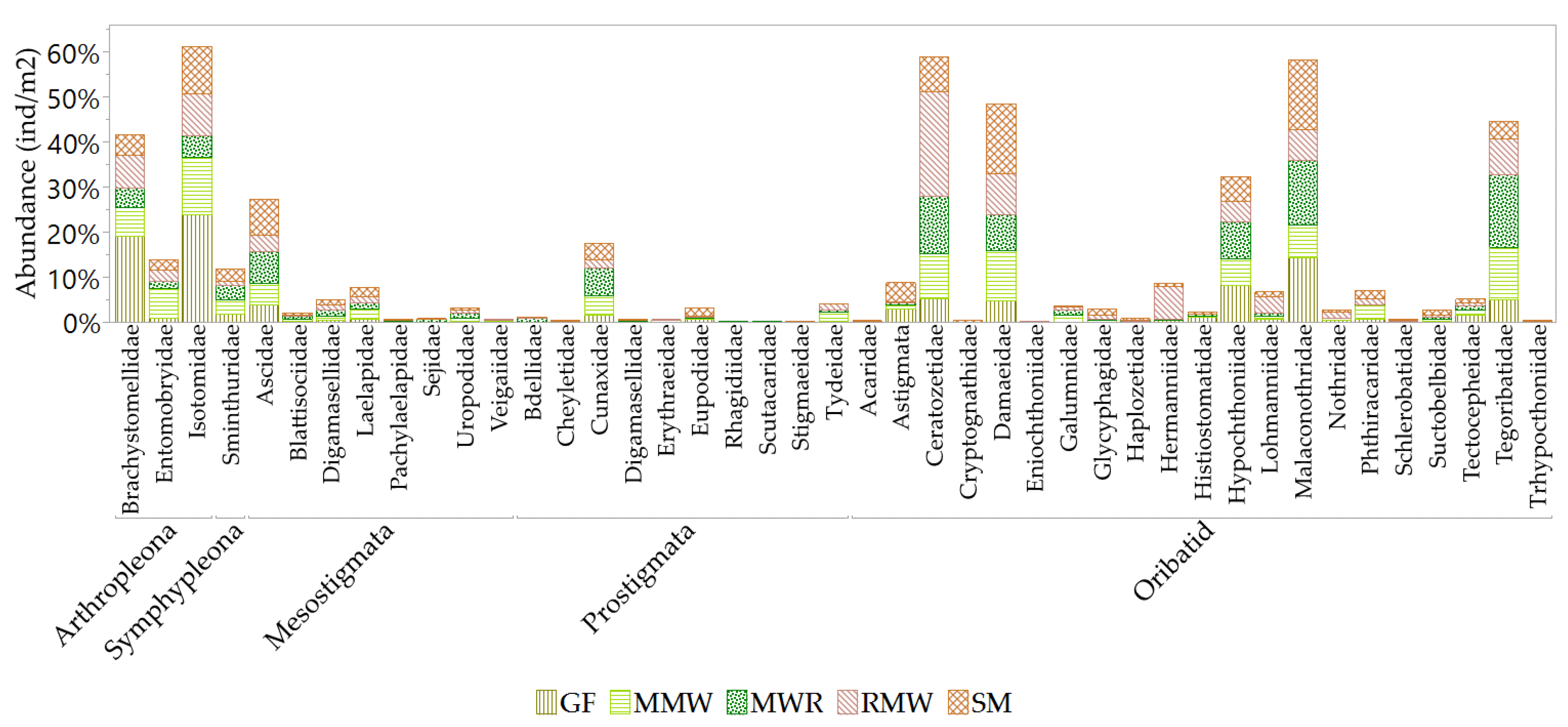
Figure 12.
Statistical differences of mesofauna richness and abundance within habitats among hydroperiods. Values with different letters indicates means with significant differences (p<.05)].
Figure 12.
Statistical differences of mesofauna richness and abundance within habitats among hydroperiods. Values with different letters indicates means with significant differences (p<.05)].
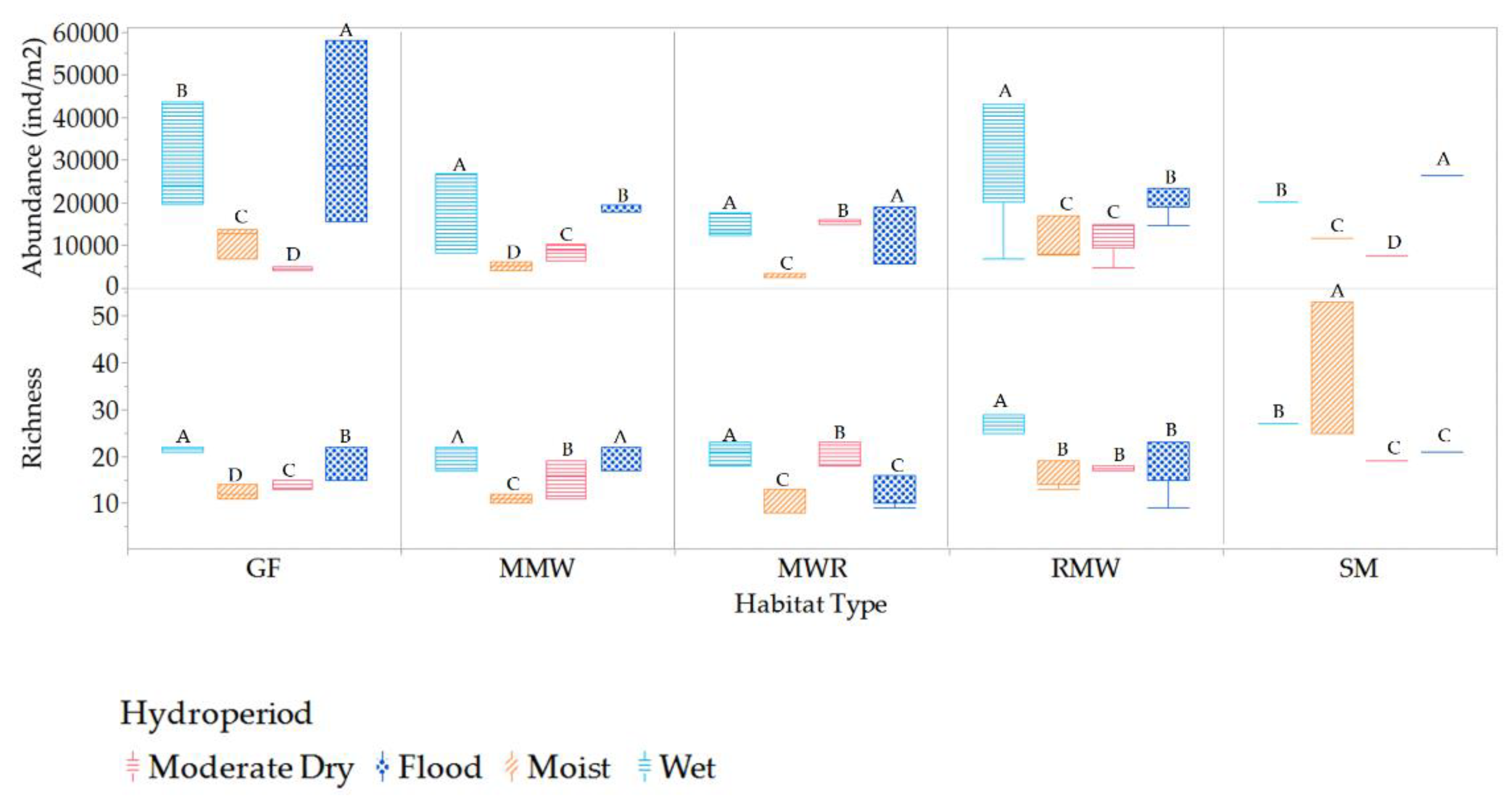
Figure 13.
Statistical differences of mesofauna richness and abundance between habitats among hydroperiods. Values with different letters indicates means with significant differences (p<.05).
Figure 13.
Statistical differences of mesofauna richness and abundance between habitats among hydroperiods. Values with different letters indicates means with significant differences (p<.05).
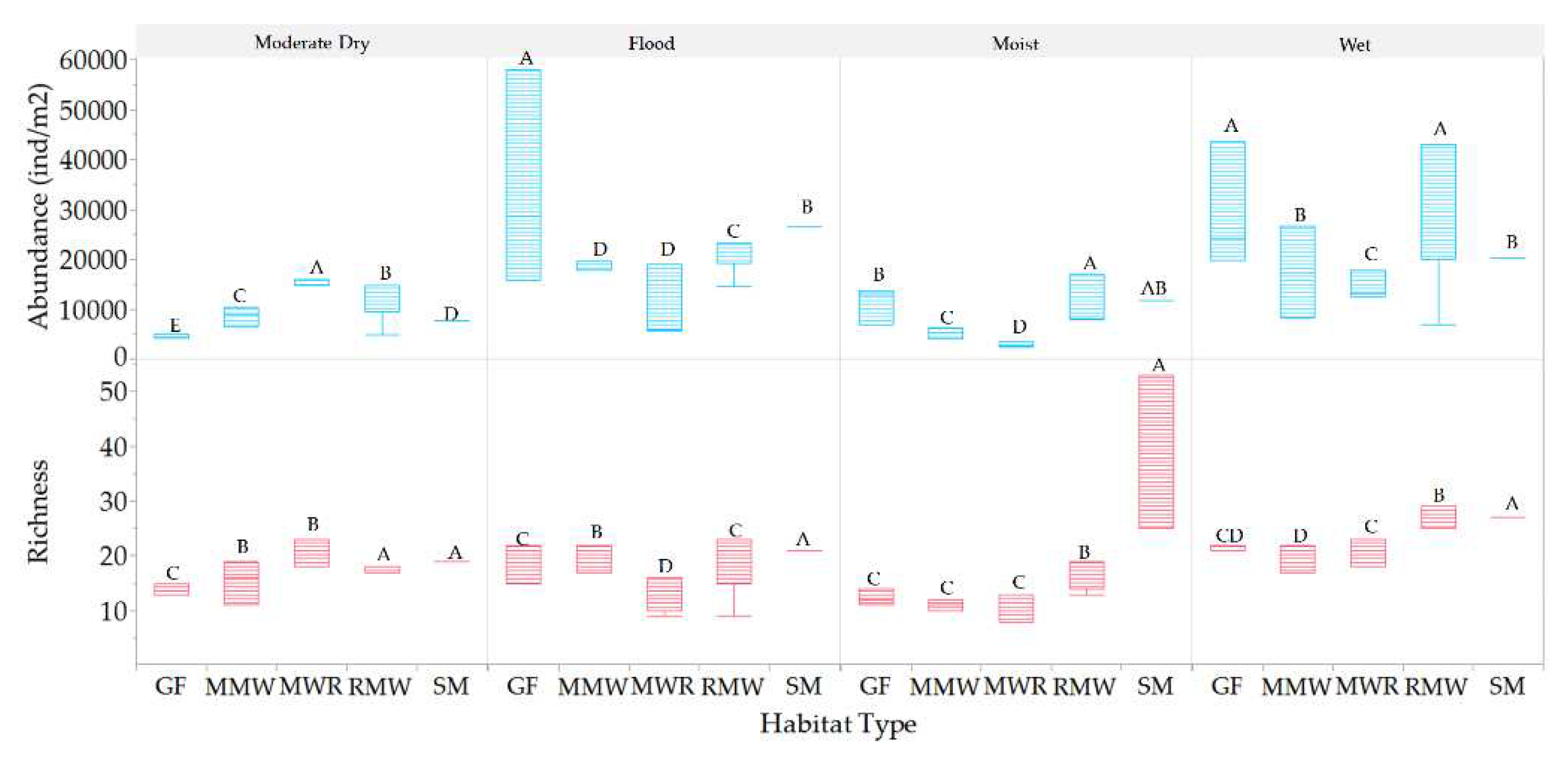
Figure 14.
Variations of mesofauna taxa among hydroperiods. Green circles indicate taxa absent during the flood period. Yellow circles represent taxa absent during the moist period. Blue circles denote taxa absent during the wet period. Red circles correspond to taxa absent during both flood and moist periods. Pink circles signify taxa absent during both flood and moderate dry periods.
Figure 14.
Variations of mesofauna taxa among hydroperiods. Green circles indicate taxa absent during the flood period. Yellow circles represent taxa absent during the moist period. Blue circles denote taxa absent during the wet period. Red circles correspond to taxa absent during both flood and moist periods. Pink circles signify taxa absent during both flood and moderate dry periods.
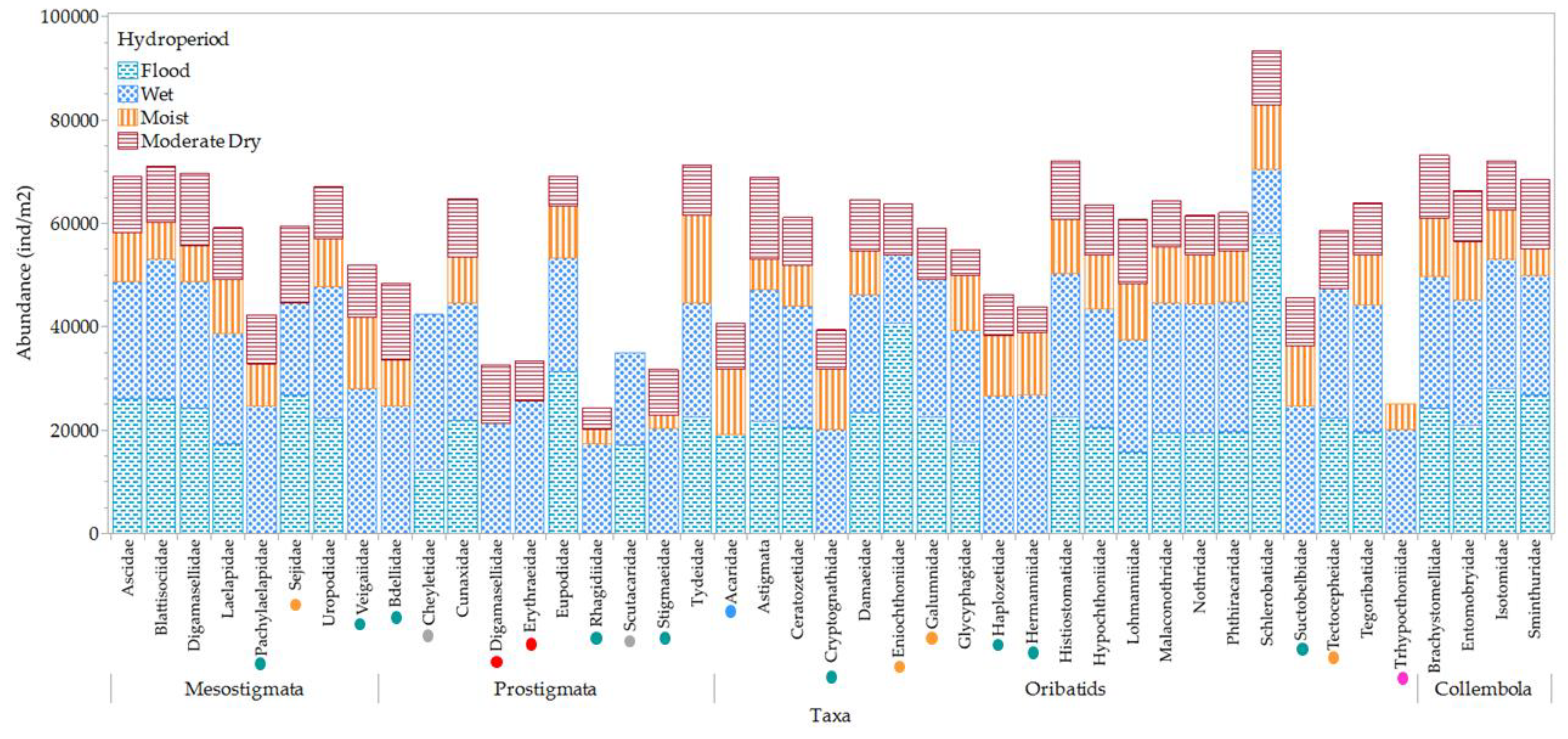
Figure 15.
Statistical differences of mesofauna richness and abundance within habitats among phreatic level categories. Values with different letters indicates means with significant differences (p<.05).
Figure 15.
Statistical differences of mesofauna richness and abundance within habitats among phreatic level categories. Values with different letters indicates means with significant differences (p<.05).
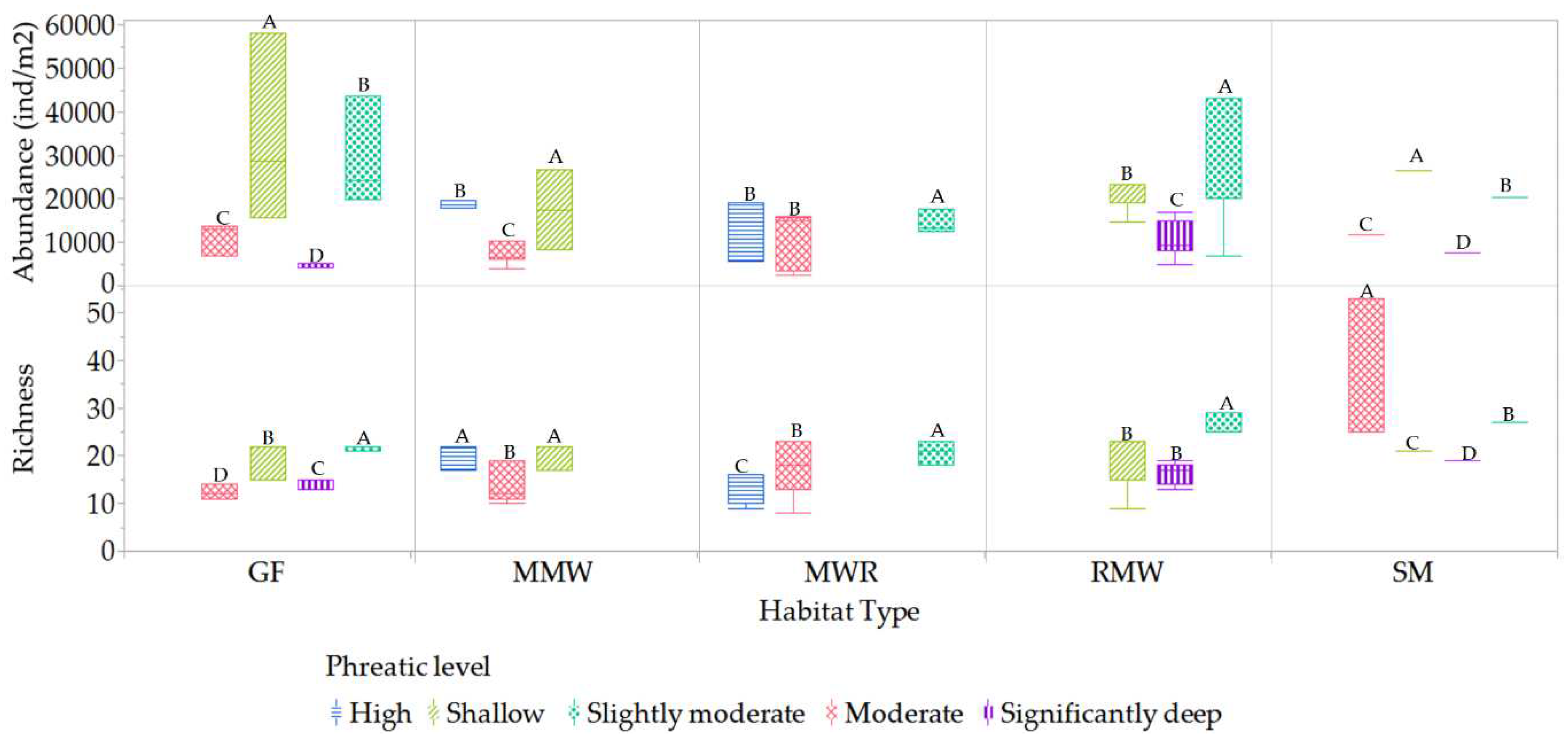
Figure 16.
Statistical differences of mesofauna richness and abundance between habitats among phreatic level categories. Values with different letters indicates means with significant differences (p<.05).
Figure 16.
Statistical differences of mesofauna richness and abundance between habitats among phreatic level categories. Values with different letters indicates means with significant differences (p<.05).
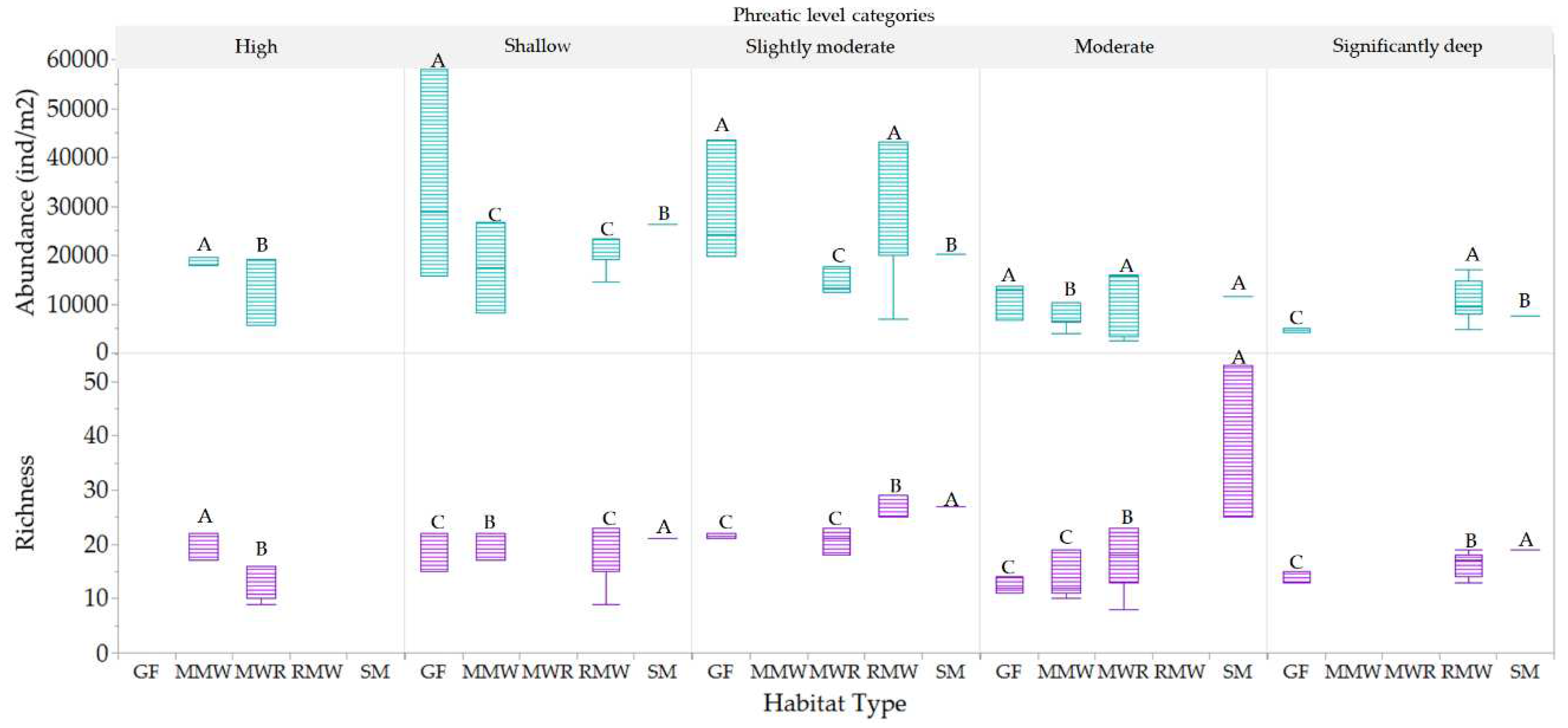
Figure 17.
Distribution and habitat preferences of different mesofauna taxa in various phreatic environments. In this figure, different colored circles represent the presence or absence of various taxa at different phreatic levels: Blue-green circles: Taxa absent in high and slightly moderate phreatic levels. Yellow circles: Taxa exclusively present in slightly moderate and moderate conditions. Dark blue circles: Taxa absent in high phreatic levels, characterized by flooded conditions. Light blue circles: Taxa absent in high and near-surface phreatic levels, typically associated with flood or waterlogged conditions. Orange circles: Taxa absent in both moderate and significantly deep phreatic levels, which are known for drier conditions. Purple circles: Taxa absent in environments with high to significantly deep phreatic levels, indicative of extreme conditions. Grey circles: Taxa absent in near-surface phreatic levels.
Figure 17.
Distribution and habitat preferences of different mesofauna taxa in various phreatic environments. In this figure, different colored circles represent the presence or absence of various taxa at different phreatic levels: Blue-green circles: Taxa absent in high and slightly moderate phreatic levels. Yellow circles: Taxa exclusively present in slightly moderate and moderate conditions. Dark blue circles: Taxa absent in high phreatic levels, characterized by flooded conditions. Light blue circles: Taxa absent in high and near-surface phreatic levels, typically associated with flood or waterlogged conditions. Orange circles: Taxa absent in both moderate and significantly deep phreatic levels, which are known for drier conditions. Purple circles: Taxa absent in environments with high to significantly deep phreatic levels, indicative of extreme conditions. Grey circles: Taxa absent in near-surface phreatic levels.
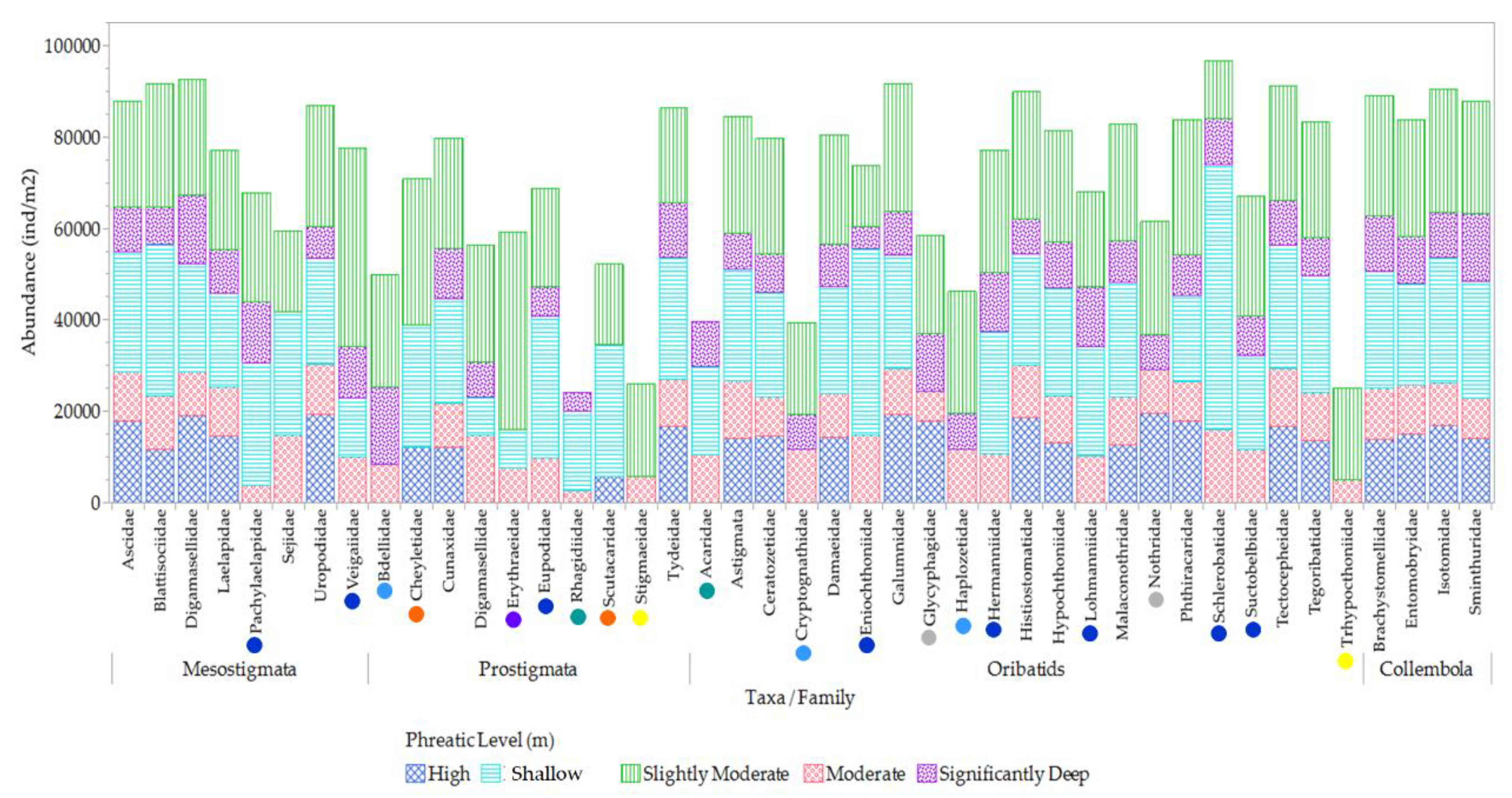
Figure 18.
Statistical differences of mesofauna richness and abundance within habitats among salinity conditions. Values with different letters indicates means with significant differences (p<.05).
Figure 18.
Statistical differences of mesofauna richness and abundance within habitats among salinity conditions. Values with different letters indicates means with significant differences (p<.05).
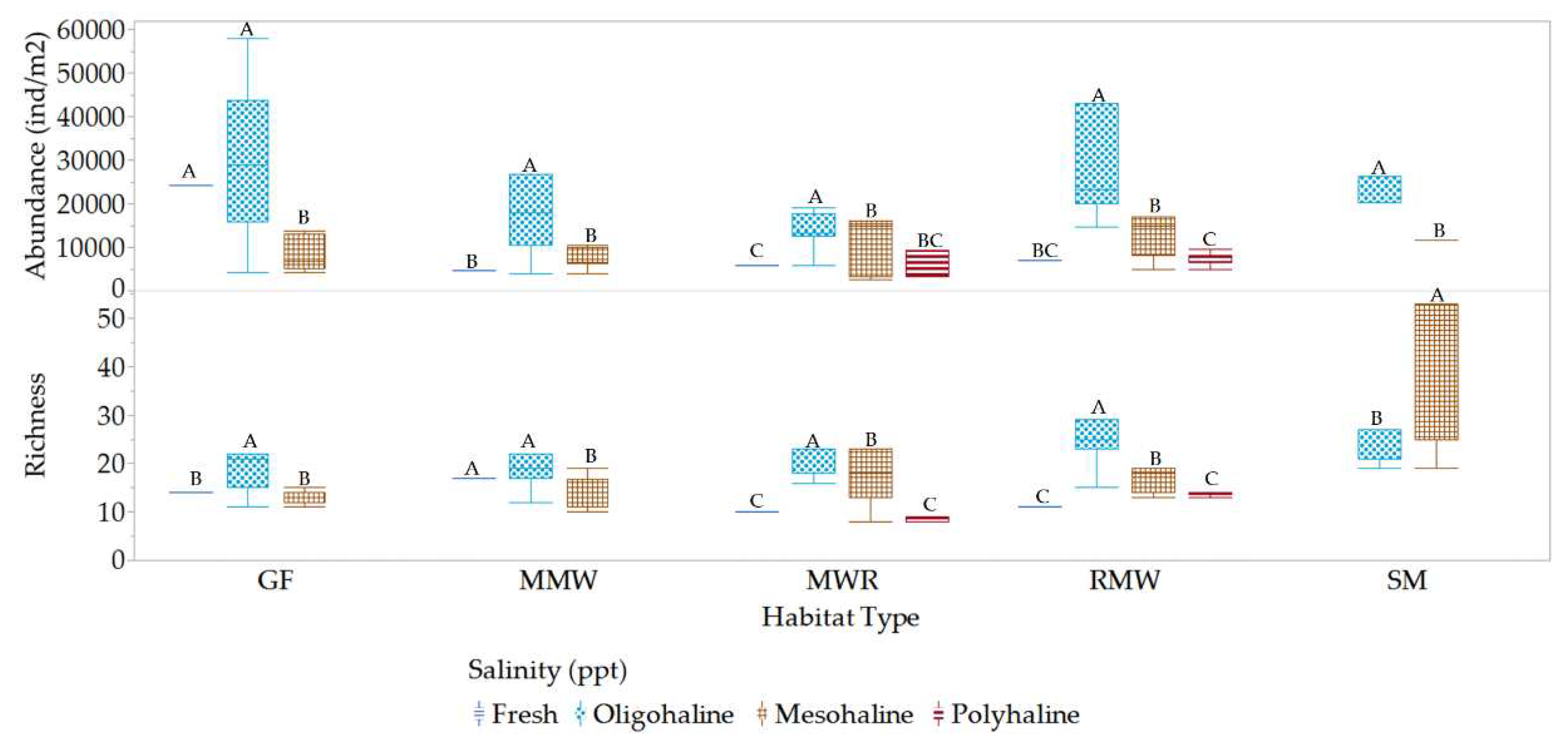
Figure 19.
Statistical differences of habitats mesofauna richness and abundance among salinity conditions. Values with different letters indicates means with significant differences (p<.05).
Figure 19.
Statistical differences of habitats mesofauna richness and abundance among salinity conditions. Values with different letters indicates means with significant differences (p<.05).
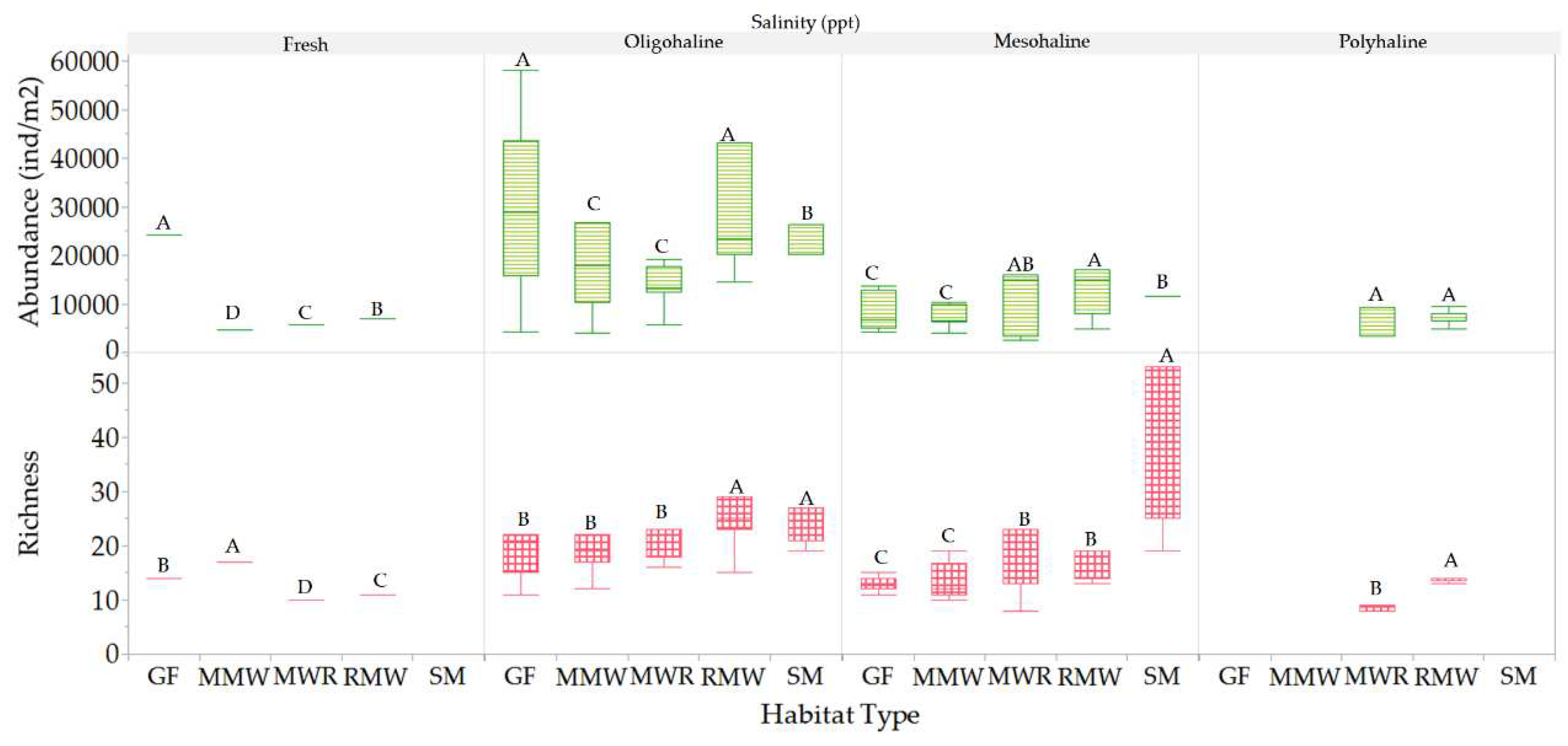
Figure 20.
Variations of mesofauna taxa among salinity conditions. Green circles represent taxa: exclusive to oligohaline salinities. Yellow circles indicate taxa tolerant of freshwater and polyhaline conditions. Blue circles correspond to taxa found at freshwater salinities. Red circles are taxa present in polyhaline conditions.
Figure 20.
Variations of mesofauna taxa among salinity conditions. Green circles represent taxa: exclusive to oligohaline salinities. Yellow circles indicate taxa tolerant of freshwater and polyhaline conditions. Blue circles correspond to taxa found at freshwater salinities. Red circles are taxa present in polyhaline conditions.
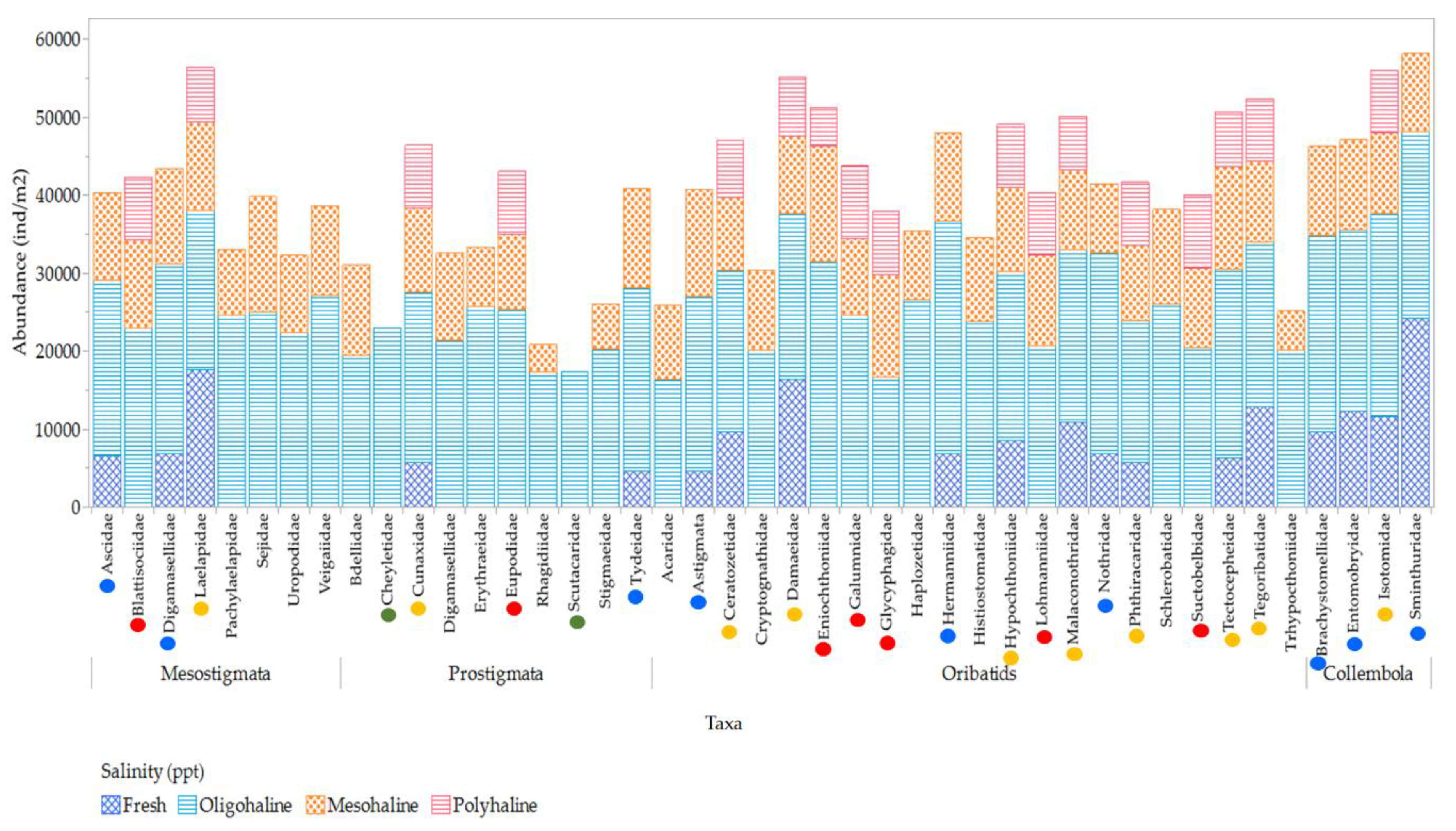
Figure 21.
Schematic diagram of the factors and gradients on wetlands that contribute to soil mesofauna diversity and abundance (Adapted from [47]).
Figure 21.
Schematic diagram of the factors and gradients on wetlands that contribute to soil mesofauna diversity and abundance (Adapted from [47]).
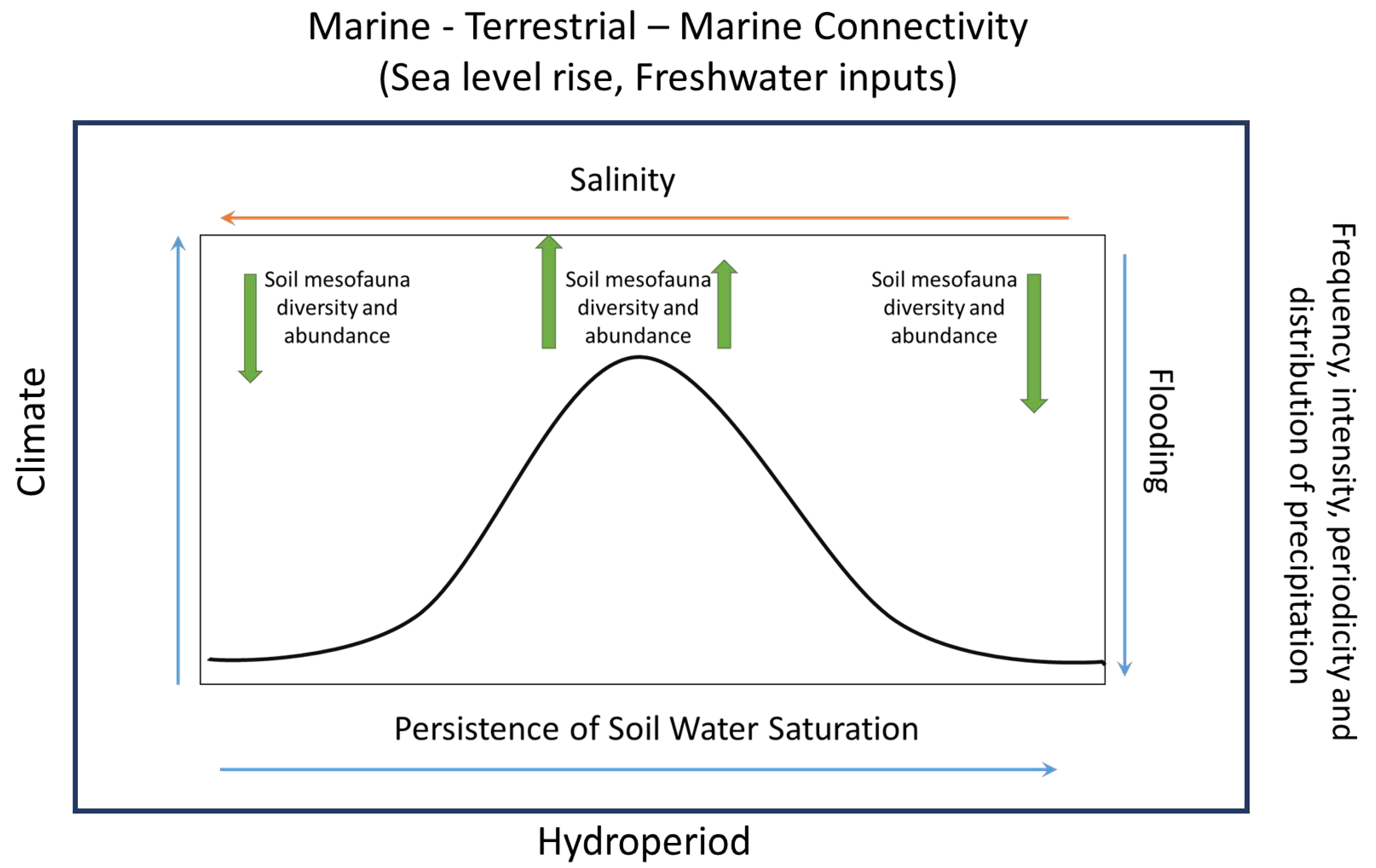
Figure 22.
Variations of mesofauna richness and abundance (ind/m2) between habitats among hydroperiod and salinity conditions.
Figure 22.
Variations of mesofauna richness and abundance (ind/m2) between habitats among hydroperiod and salinity conditions.
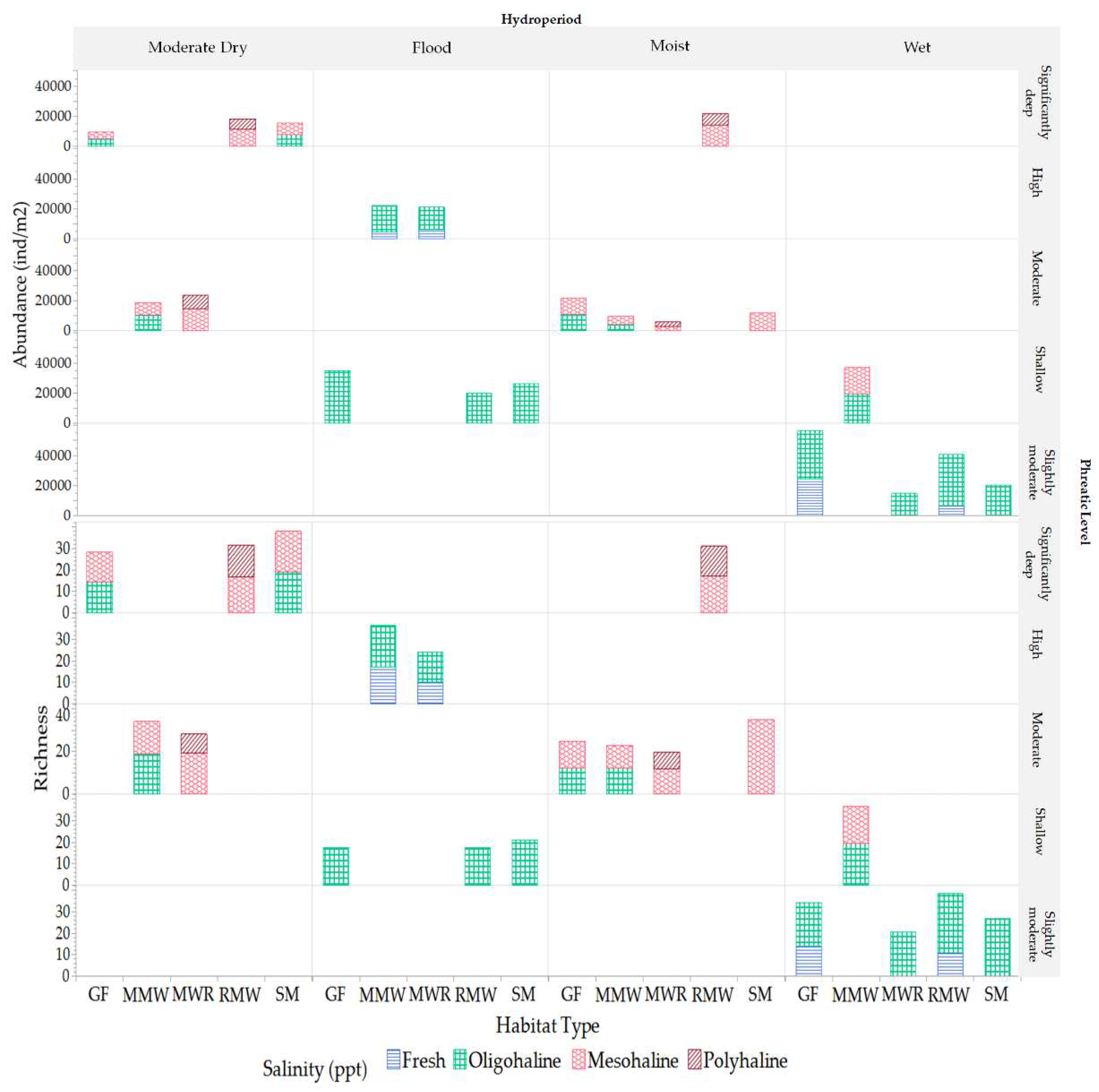
Figure 23.
Variations of mesofauna taxa among salinity conditions.
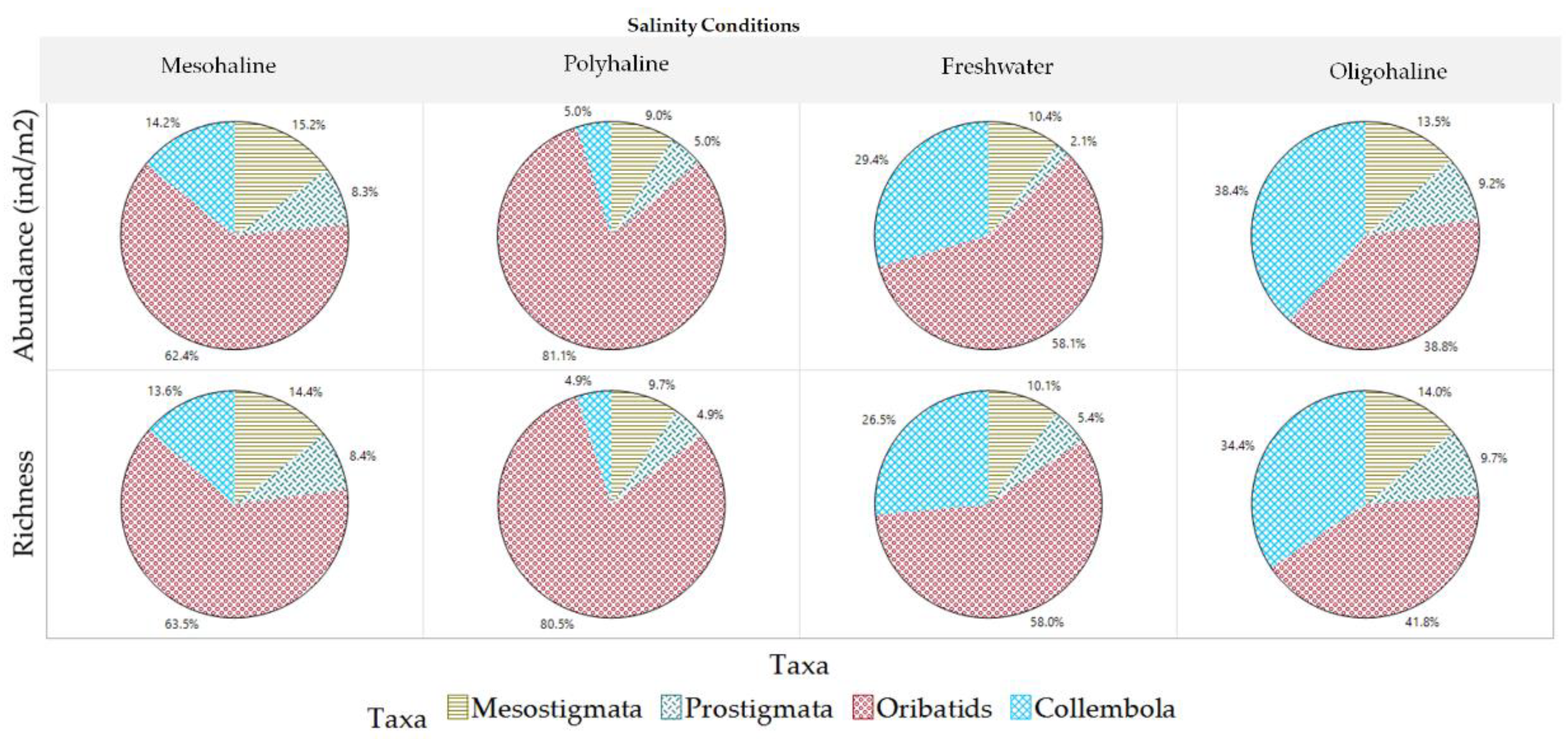
Table 2.
Tide conditions during sampling dates and hours. NOAA 9755371, La Puntilla Station, San Juan, Puerto Rico.
Table 2.
Tide conditions during sampling dates and hours. NOAA 9755371, La Puntilla Station, San Juan, Puerto Rico.
| Hydroperiod | Sampling Date | Sampling Time* | Tide (m) | Tide description |
|---|---|---|---|---|
| Moderate Dry | 6/18/2020 | 7:00 | 0.22 | Low |
| 10:00 | 0.32 | High | ||
| 6/25/2020 | 7:00 | 0.48 | High | |
| 10:00 | 0.23 | Low | ||
| Flood | 10/23/2020 | 7:00 | 0.31 | High |
| 10:00 | 0.14 | Low | ||
| Moist | 3/19/2021 | 7:00 | 0.29 | High |
| 10:00 | 0.15 | Low | ||
| Wet | 6/9/2021 | 7:00 | 0.17 | Low |
| 10:00 | 0.2 | High |
* Sampling occurred in the morning, between 7:00 am to 10:00 am.
Table 3.
Phreatic level (m) (mean ± std) between habitats and hydroperiods. Values with different capital letters indicates means with significant differences (p<.05) between habitats, lowercase letters indicate means with significant differences (p<.05) between hydroperiods.
Table 3.
Phreatic level (m) (mean ± std) between habitats and hydroperiods. Values with different capital letters indicates means with significant differences (p<.05) between habitats, lowercase letters indicate means with significant differences (p<.05) between hydroperiods.
| Habitat | Hydroperiod Phreatic Level (m) | |||
|---|---|---|---|---|
| Moderate Dry | Flood | Moist | Wet | |
| -0.49±0.06 d | -0.01±0.07 a | -0.44±0.12 c | -0.12±0.03 b | |
| GF | -0.51B | -0.05 D | -0.36 A | -0.12B |
| MMW | -0.41A | 0.11 A | -0.43 B | -0.07A |
| MWR | -0.41A | 0.09 B | -0.36 A | 0.12B |
| RMW | -0.54C | -0.03 C | -0.64 C | -0.17C |
| SM | -0.51B | -0.05 D | -0.36 A | -0.12B |
Table 4.
Spearman’s Rho correlation analysis showing significant correlations between hydroperiods phreatic level and habitat bio-physicochemical factors.
Table 4.
Spearman’s Rho correlation analysis showing significant correlations between hydroperiods phreatic level and habitat bio-physicochemical factors.
| Habitat type | Phreatic Level by Salinity | |
|---|---|---|
| Spearman ρ | Prob>|ρ| | |
| GFM | -0.5 | <.0001 |
| MMW | -0.8 | <.0001 |
| MWR | -0.8 | <.0001 |
| RMW | -0.6 | <.0001 |
| SM | -0.7 | <.0001 |
Table 5.
Jaccard index matrix showing similarities between habitat types.
| MMW | RMW | MWR | SM | GF | |
|---|---|---|---|---|---|
| - | 0.78 | 0.73 | 0.80 | 0.82 | MMW |
| 0.78 | - | 0.80 | 0.88 | 0.80 | RMW |
| 0.73 | 0.80 | - | 0.83 | 0.80 | MWR |
| 0.80 | 0.88 | 0.83 | - | 0.78 | SM |
| 0.82 | 0.80 | 0.80 | 0.78 | - | GF |
Table 6.
Spearman’s Rho correlation analysis showing correlations between mesofauna richness index (Menhinick's index) and total abundance with study site physicochemical factors.
Table 6.
Spearman’s Rho correlation analysis showing correlations between mesofauna richness index (Menhinick's index) and total abundance with study site physicochemical factors.
| Variable | By Variable | Spearman p | Prob>|p| |
|---|---|---|---|
| Richness |
Phreatic Level | -0.5 | <.0001* |
| Salinity (ppt) | 0.3 | <.0001* | |
| Abundance (ind/m2) | Salinity (ppt) | -0.5 | <.0001* |
| Phreatic Level | 0.5 | <.0001* |
Table 7.
Generalized regression model effect report providing information about the magnitude of the effect of each predictor on mesofauna richness index and total abundance (ind/m2).
Table 7.
Generalized regression model effect report providing information about the magnitude of the effect of each predictor on mesofauna richness index and total abundance (ind/m2).
| Mesofauna richness | Wald ChiSquare | Prob > ChiSquare | Mesofauna total abundance | Wald ChiSquare | Prob > ChiSquare |
| Habitat Type | 658 | <.0001* | Habitat Type | 568 | <.0001* |
| Phreatic level | 472 | <.0001* | Phreatic level | 260 | <.0001* |
| Salinity | 57 | <.0001* | Salinity | 12 | 0.0005* |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



