Submitted:
15 September 2023
Posted:
19 September 2023
You are already at the latest version
Abstract
The body of knowledge on ocean acidification gives a better understanding of biological sensitivity to low pH. Key parameters such as life-history strategies or local adaptation were identified as keys to predict species sensitivity and resolve previously some of the unexplained species- and population-specific differences. Encapsulation has been suggested as one of these keys as it exposed the embryo to low pH conditions, or ontogenetic hypercapnia, leading to physiological adaptation. We tested this hypothesis on the nudibranch Aeolidiella glauca by exposing their egg-strings containing large number of eggs to two different pH (8.1 and 7.3). The fertilized eggs developed 1 egg-cell, over early cleavage up to morula, blastula, gastrula, rhomboid-shaped rotating gastrula, early rotating veliger larvae with developed shell, to free-swimming well developed veliger larvae. Despite a corrosive environment, the exposure to low pH had no significant effect on the developmental rate. The only significant effects were a slightly smaller and narrower shell in larvae raised at low pH as compared to the high pH. Our results showed a remarkable resilient to low pH in a calcifying mollusc and support the idea that ontogenic hypercapnia is leading to low sensitivity to ocean acidification.
Keywords:
development
; nudibranch
; acidification
1. Introduction
Human induced rise in atmospheric CO2 concentration leads to changes in sea water carbonate chemistry including a decrease in seawater pH, a process known as ocean acidification (OA). A large body of evidence is documenting various effects on marine species and ecosystem [1]. The early life stages of marine invertebrate are especially sensitive to a decrease in seawater pH. These include an increase in larval mortality rate and the percentage of abnormally developing larvae as well as delay in development [2,3].
The nudibranch Aeolidiella glauca (Figure 1a,b) is one of the key organisms living in Zostera marina shallow eelgrass meadows. Along the Swedish West coast, it lays large egg-strings on the eelgrass during summer (Figure 1c,d). Like all nudibranchs, A. glauca is hermaphrodite with internal fertilization and have functional female and male genitalia simultaneously [4]. A. glauca does not perform self-fertilization. It cross-fertilizes and transfers sperms via external spermatophores on the partner’s back, from which the sperms migrate to the gonopore of the receiver [4]. Thereafter, sperms enter the reproductive organs and fertilize the eggs.
From mid-June to early August, A. glauca preforms multiple matings and can lay several egg-strings. Several matings stimulate the string and egg laying. The egg-strings contained a huge number of eggs, up to 1000 eggs per cm [4]. The string size and number of fertilized eggs depends on the number of sperms reaching the female reproductive organs [4]. For example, one mating may not always be sufficient to transfer enough sperms to fertilize all eggs in the egg-string. The receiver animal can also regulate the possibility for the sperms to fertilize the eggs by closing the gonopore. After reproduction A. glauca dies.
The development of A. glauca is still poorly characterized. Preliminary observations performed in 2019 showed that egg development undergo spiral cleavage including morula, blastula and gastrula, followed by larval development up to free-swimming veliger larvae (Figure 2, Figure 3, Figure 4 and Figure 5). The whole developing process in aquarium with eelgrass meadow and natural environmental conditions takes around 7 to 8 days.
While the changes in carbonate chemistry associated with ocean acidification are well defined for the open ocean, these are more complex for the coastal environment. Large fluctuations can be observed in regions with high biological activity because of the balance between respiration and calcification (producing CO2) and photosynthesis (consuming CO2). For example, in the Gullmars Fjord in Sweden, pH can vary between 8.6 to 7.6 and this variability will shift down to a decrease of 0.4 pH unit by the end of the century [2]. Recent work demonstrated that species sensitivity to pH is strongly modulated by the extreme of the present range of variability as a consequence of local adaptation [5,6]. This hypothesis was tested in the Gullmars Fjord on several species confirming this hypothesis with a threshold of around 7.5 for the green sea urchin Strongylocentrotus droebachiensis [2,7] and around 7.7 for the blue mussel Mytilus edulis [3]. Based on this hypothesis, we could hypothesize that A. glauca development may be tolerant to pH down to 7.6 and that negative effect on the embryo and larval development will be observed under more extreme low pH.
Life-history traits are also modulating species sensitivity to low pH and explain contrasting responses observed between species living in the same environment. Benthic invertebrates have developed a variety of life-history strategies that can be divided into two main strategies: (i) planktotrophy, where a parent invests energy in producing millions of small eggs that will develop into planktotrophic larvae feeding on exogenous sources; and (ii) lecithotrophy, where parents produce comparatively small numbers of large, yolky eggs and lecithotrophic larvae that derive their nutrition from energy stored in the egg itself. The comparison of the two strategies in echinoderms inhabiting the Gullmars Fjord, it was shown that while species with planktotrophic strategy were much more sensitive to low pH, no negative effects were observed in lecithotrophic ones [8]. Another life-history trait that was proposed to modulate species sensitivity to ocean acidification is the encapsulation of embryos. Based on studies performed on the cephalopod Sepia officinalis, Melzner et al. [9] proposed that the idea that encapsulation can lead to physiological adaptation to low pH. While partly protecting from the external environment, encapsulation is exposing the embryos to low pH and low O2, what can be referred to ontogenetic hypercapnia [10]. This is a strategy used by many species of molluscs where they lay their eggs in different type of egg mass such as strings, ribbons, capsules, or globose. These species can then be expected to be resilient to low pH well beyond the extremes of the present range of natural variability.
To test this hypothesis, we designed an experiment comparing the impact of two different pH: (i) 8.1, the present average in seawater pH at the sampling site; and, (ii) 7.3, the extreme pH within the natural variability as expected by 2100 and well below the extreme of the present range of variability (tipping point of pH 7.6; [2]). We followed the developmental rates from 1-cell stage to free swimming A. glauca veliger larvae. A photographic documentation and description of this development was also performed.
2. Materials and Methods
2.1. Animals and egg-strings
The nudibranch Aeolidiella glauca (Figure 1a) was collected by scuba diving or snorkeling in the eelgrass (Zostera marina) communities in the Gullmars Fjord, Skagerrak, off the Swedish west coast (58°21′N, 11°06′E; 58°21′N, 11°07′E), during late June to early August, 2019, 2022, 2023. A. glauca was found on Z. marina leaves; depth 0.5-4 m. Collected nudibranchs were kept in small aquaria with running, fresh seawater from the Gullmars Fjord at pHT 8.0 and 18 °C at the Kristineberg Marine Research Station, Fiskebäckskil, University of Gothenburg. Z. marina and the sea anemone Sagartiogeton viduatus, the favorite food prey of A. glauca (Figure 1b), were also placed in the aquaria. In nature, A. glauca attaches the egg-string on the leaves of Z. marina (Figure 1d). When captured, A. glauca also attached strings on the glass sides (Figure 1c) and on the bottom of the aquaria. Some strings were free-floating on water surface of the aquaria. The starting region of the laid egg-string was broad with closely packed eggs compared to the narrow egg-string end, with less packed eggs (Figure 1c,d). The develop analyses started at the narrow string end with the latest laid eggs. Egg-strings from 2022 were used to document the impact of pH 8.1 and 7.3 on the development from one egg cell to free-swimming veliger. Pictures taken over the 3 years illustrate in detail the larval development.
2.2. Experimental designs and carbonate chemistry
The experiment was performed between 9 July and 18 August 2022. At total of 28 egg-strings laid by A. glauca were analysed, from which 6 free-floating strings were selected for the experiment. Free-floating egg-strings were easier to analyse consistently than strings attached on eelgrass. The egg-string replicate were placed, one by one in Petri-dishes with seawater at pH 8.1 or 7.3 (n=3 per pH), covered with a lid to limit the contact with the atmosphere and avoid fluctuations in pH. The Petri-dishes were kept in a thermoconstant room at 18°C.
Aerated deep seawater was used for the high pH while low pH was achieved by bubbling with pure CO2. Seawater physico-chemistry (temperature, salinity, pH and alkalinity) was measured 16 times over the course of the experiment. On each seawater batch, pH was measured in on the total scale (pHT) using a glass electrode calibrated with TRIS buffer and alkalinity was measured by titration. Other parameters of the seawater chemistry (pCO2, Wcalcite, Waragonite) were calculated from temperature, salinity, pHT and alkalinity using CO2sys with the dissociation constants from Mehrbach et al. [11] as refitted by Dickson & Millero [12].
2.3. Endpoints
Along the narrow string-end, each replicate string was divided into ten string-parts in a row. The percentage of the different developmental stages were here quantified at every sampling time, keeping track of the egg-string replicates and tested pH. Each string parts were photographed under a stereomicroscope with 16x magnification, and the number of eggs at each specific development stages were counted. In total around 200 different eggs were counted each time from the selected areas. The documentation of the developmental stages was done in the same string areas each time. Pictures were taken to document the different cell-cleavage stages up to larval development and to well-developed veliger larvae (Figure 2).
The cleavages from one cell to 2, 4, 8, 16, 32 cells (early morula) were documented every hour (Figure 2a–d). After the egg-strings had reached the morula stage (early morula Figure 2d), and start developing into late morula (Figure 2e), documentations were performed 4 times daily from now on to eggs developed to blastula (Figure 2f); gastrula (Figure 2f); ciliated moving or rotating rhomboid-shaped larvae with markings for velar apparatus (Figure 2g); rotating larvae with velar apparatus and small shell (Figure 2h); and rotating, well-developed veliger larvae with large shells (Figure 2i). On free-swimming veliger larvae, floating on the surface-water in the Petri-dishes (Figure 5p), the shell size (length x width) was measured in dorsal view.
Analyses with light microscope (40x) were performed to illustrate in details early cleavage stages (Figure 3); early morula (Figure 4a,b); late morula (Figure 4c,d); blastula (Figure 4e,f); gastrula (Figure 4g,h); ciliary movements and rotation of rhomboid-shaped gastrula (Figure 4i,j); development of the velar apparatus (Figure 4k,l); up to well-developed veliger larvae with shell (Figure 5). Early rotating, successively growing veliger larvae in different positions inside the egg-shell show different structures as liver, stomach, eyes, statocysts, growing shell and foot (Figure 5a–d). In the free-swimming veliger larvae, these structures were more prominent (Figure 5e–j).
2.4. Microscope
Examinations of the egg-strings from early egg cleavage to free swimming veliger larvae were made using a Leica M205C (Leica Microsystems, Wetzlar Germany) stereomicroscope and a Leitz DMRBE (Leica Microsystems, Wetzlar Germany) light microscope (LM) equipped with interference-contrast optics, 100x/1.30 PL, fluotar objective. Both microscope were connected to a Leica DFCA20C camera with an application suite, version 3.8 (LAS V3.8The stereomicroscope Leica M205C was connected to the CAMERA AXICAM 305 color, and the Leitz DMRBE light microscope was connected to the CAMERA AXICAM 705 color. Both cameras were equipped with the software ZEN, 31 blue edition, Zeiss. Accuracy measurements in the LM was generally + 0.5 mm, and + 0.2 mm for the smallest details.
2.5. Statistical analyses
For 3 strings per pH treatment, the duration of each developmental stage (early cleavage, morula, blastula, gastrula, gastrula moving, rhomboid-shaped; veliger larvae without shell; veliger larvae with shell) was estimated as the mean development time (MDT in hour) between 2 successive stages. MDT was calculated as the time when 50% of the organisms in a culture had passed that stage, which in turn was estimated from stage–frequency data converted to stage proportion. The cumulative proportion of each stage over time was plotted against time and a linear regression was fitted (adapted from [13]). The effect of pH on the MDT for six developmental stages was tested using a one-way analysis of variance (ANOVA). Effects of pH and strings (replicates) were tested using a two-ways ANOVA. Difference between strings were evaluated using a post-hoc Scheffe’s test. The Shapiro-Wilk statistic W [14] was used to check the data for normality of distribution. Analyses were performed using SAS/STAT. All results as presented as mean with the standard error of mean (sem).
3. Results
3.1. Description of A. glauca early development (2019, 2022, 2023)
During egg laying, A. glauca slowly moved in ovals or circles around the starting point of the egg-string (Figure 1c). Laying the egg-string took up three to four hours. Eggs in the string-end thus started their cleavage some hours later than eggs at the string-starting region. The documentation of the development was easier in the narrow string-end with not so densely packed eggs compared to at the starting region (Figure 1c,d). In free-floating egg-strings the small sections of eggs in the narrow string-end (Figure 1c) were easier to select and document compared to on strings attached at the seagrass (Figure 1d). At the end of A. glauca’s reproduction season, egg-strings were often narrower, shorter and contained less number eggs as compared to strings laid in the beginning of, or during the point of A. glauca’s reproduction season.
Right after the individual finished laying the eggs, 100% of the eggs were in the 1-cell stage in the narrow end (compare Figure 2a and Figure 3a), whereas in the thicker starting string end 72% were already in the 2-cells stage (Figure 2b). One hour later, 100% were still in the 1-cell stage in the narrow string end, and in the 2-cells stage in the thick end. Future observations focused on the narrow string end. After 3 hours all cells were in the 2-4-cells stages (Figure 3b), and in the 4-8-cells stages after 6 hours (Figure 2c and Figure 3c–h). At 8 hours, some embryos were still at the 4-cell stage, but the majority reached the 12-16-cells (Figure 3i,l). Starting with the third cleavage, the 8-cells embryo undergoes spiral cleavage, a cleavage pattern more or less clearly recognizable up to morula (Figure 3h–l). Early morula (~32-cell stage) appeared after 11 hours (compare Figure 4a,b) and late morula (64 cell stage) (Figure 4c,d) after 12 hours. At 18 hours, late morula was the dominant stage in the string. Early morula stage has larger cells than late morula, as seen in Figure 2d,e and Figure 4a–d. Faint cilia movements were visible after 21 hours.
The three stages: late morula, blastula gastrula could appear at the same time in the selected string-parts (Figure 6). To separate late morula (Figure 4c,d) from blastula (Figure 2e,f and Figure 4e,f) is not always easy. Blastula with hollow center, the blastocoel appeared at 27 hours. The gastrula invagination appeared after 30 hours. The invagination is clearly visible in side view (Figure 2f and Figure 4g) and the blastopore can be focused in ventral/dorsal view (Figure 4h).
Between 3 and 4 days, the rounded gastrula with marginal incision or central ventral/dorsal blastopore (Figure 4g,h) transformed into an irregular rhomboid-shaped larvae (Figure 4i,j). Faint ciliary movements causing the gastrula to weakly sway back and forth (Figure 4i), which later turned to greater wobbling’s and rotations of the larvae (Figure 4j). Faint velar markings (Figure 2f) turned to butterfly-shaped wings on early veliger larvae (Figure 4j,k,l).
The small shell on top of the larva (Figure 4l) continued to grow to a fully developed large shell on the growing, rotating veliger larvae (Figure 5a–g). On this larval stage, still inside the egg-shell, stomach, liver, eyes, statocyst and retractor tendon/muscles had developed. After 7-8 days, a fully developed, free swimming veliger larvae was visible (Figure 5e–h) with long beating ciliated velar apparatus outside the partly transparent shell. The cilia created a current transporting small particle towards the mouth, located between the two velar wings (Figure 5e,g). The mouth was surrounded by small, cilia. Particles were transported to the stomach through a canal system (Figure 5e–h). On the lower side of the stomach wall, an u-shaped, a small dense structure was visible, towards which particles were circulating (Figure 5h). Large cuboidal, epithelial cells surrounded the large stomach lumen (Figure 5i). The large liver, placed inside the bulging shell, consisted of large, dense cuboidal epithelial cells (Figure 5e–h). The larva was attached to the shell by a broad retractor muscle/tendon (Figure 5c,f). A thin tendon/muscle connected the stomach to the shell (Figure 5h). A pair of prominent eyes could be seen on larvae when inside or partly outside the shell (Figure 5a,e,i). In partly contracted larvae seen from the ventral shell side, eyes and foot could be seen inside the opercular shell part (Figure 5i,j).
3.2. Carbonate chemistry
The carbonate chemistry of the water used during the experiment is summarized in the Table 1. The target pH 8.0 and 7.3 were reached with a corresponding pCO2 of 411 and 2739 matm, respectively. At pH 8.0, the seawater was super saturated for both calcite and aragonite while it was corrosive for both calcium carbonate forms at pH 7.3 (Wca and War<1).
3.3. Effect of pH on the dynamic of the early development
An example of developmental succession is presenting in Figure 6. From the morula stage, the duration of each stage was estimated as the mean development time (MDT) between 2 successive stages (Figure 7). Different stages had different durations: (i) Late morula, 16±3 h (Figure 4c,d); (ii) blastula, 25±3 h (Figure 4e,f); (iii) gastrula, 25±3 h (Figure 4g,h); (iv) moving, rhomboid-shaped gastrula, 18±2 h (Figure 4i,j); (v) rotating veliger with no shell, 39±5 h (Figure 4k,l); and, (vi) well developed veliger larvae with shell, 40±4 h (Figure 5a–d). pH had no significant effect on these mean developmental time (Figure 6, Table 2).
3.4. Effect of pH on the morphology
After hatching, shell length was significantly impacted by pH (ANOVA 2; model: F5,941=51.40, p<0.0001; pH: F1=134.44, p<0.0001) with significant differences between replicates (F4=21.30, p<0.0001). Shells from strings maintained at pH 7.3 were 5% smaller than the ones raised at pH 8.1 (Figure 8; pH 8.1: 146.04±0.31 mm; pH 7.3: 138.53±0.54 mm).
A Scheffe’s post-hoc test (Figure 8) showed that shell length from all strings were significantly different from each other with the exceptions of R1-R2 at pH 8.1, R1-R2 at pH 7.3 and R3-pH 8.1 and R2-pH 7.3.
There was a significant linear relationship between shell length and width for pH 8.1 and pH 7.3 (Figure 9). The ratio between shell length and width was significant difference between the two tested pH (ANCOVA; model: F2,941=670.28, p<0.0001; pH: F1=133.50, p<0.0001). The ratio was higher at pH 7.3 (0.63±0.03) as compared to pH 8.1 (0.59±0.03). Overall, larvae raised at low pH have a 10% narrower shell for a given length, but this difference decreased with increasing size (Figure 9).
4. Discussion
As the development of A. glauca was poorly characterized, we documented and described the evolution and dynamic of its development from fertilization to the free-swimming larvae. Like all mollusks, the egg cells of the nudibranch Aeolidiella glauca undergo holoblastic spiral cleavage, a form of embryonic development, which occurs widely within invertebrates such as mollusks, annelids, nemerteans and flatworms [15]. The first three cleavage stages up to early morula (32 cell stages) went rather fast and were reached after 11-12 hours. The development continued following a classic pattern through late morula, blastula and early gastrula. The growing gastrula stages became rhomboid-shaped. The tiny wobbling movement caused by cilia in the velar region increased with time and cilia continue to grow on the velar region. Rhomboid-shaped gastrulas with velar markings developed into rotating gastrulas. The different gastrula stages including the early none-moving gastrula, the rhomboid-shaped gastrula with increasing ciliary movement, the rotating growing rhomboid–shaped gastrula with no or a tiny piece of shell and early developing stomach and liver regions spanned over a period of approximately 75-80 hours. Approximately 120 hours were needed to grow a rotating veliger larva with a growing large shell and developing internal functional organs and about one more day before hatching and the release of the free-swimming larvae.
Exposure to pH 7.3, well below the extreme of the present range of variability at the sampling site, had little effect on the early development of A. glauca. The development progression was not affected, and the corrosive water did not prevent the formation and maintenance of the aragonite shell. The only significant effect was a subtle change in shell allometries. Under low pH, the shells were more triangular-shaped and slightly narrower than shells developed under pH 8.0.
The impact of ocean acidification on nudibranch is poorly documented with only 2 articles published so far and only focusing on adult stages. The nudibranch Corambe stenbergae is a predator of bryozoan. Short exposure to pH as low as 7.1 only had no effect on their consumption rate [16]. In another study, adults of 5 different species of nudibranch (Aeolidia papillosa, Cuthono gymnota, Dendronotus frondosus, Onchidoris bilamellata and Flabellina verrucosa) were collected from intertidal and subtidal zones and exposed to pH 7.8 and 7.0. Exposure to low pH did not trigger any stress behavior in any of the tested species and a mild but significant effect on the heart rates was only observed in 2 of the 5 tested species [17]. Overall, these results also suggest a high tolerance for low pH in adult nudibranch.
Maybe more surprisingly, we show high tolerance to low pH for the planktotrophic veliger larvae of A. glauca and its aragonite shell, despite growing in corrosive water. Marine calcifiers are generally sensitive to low pH as the extra energy costs associated with acid-base regulation can lead to negative effects on fitness (e.g. [18]). However, calcifiers are not pieces of calcium carbonate and they can inhabit chemically challenging environments [19]. For example, dense clusters of the vent mussel Bathymodiolus brevior were observed at pH values as low as 5.36 [20]. This is possible in species developing physiological adaptation to buffer or compensate for the environmental challenges associated with low pH (acid-base regulation, e.g. [18]; compensatory calcification, e.g. [3]). In A. glauca, this may be a consequence of ontogenic hypercapnia. Encapsulation is a method to protect and buffer against some chemical changes [21] but it also exposed the embryos to more extreme environments than their surrounding seawater, including undersaturated low pH waters. These can be amplified under ocean acidification conditions and can drive negative response when the limit of the pH tolerance is reached. The encapsulated embryo of Littorina obtusata exposed to pH 7.6 has a small decrease in viability, an increased developmental time, decreased heart rate, and changes in the shell morphology [22].
We hypothesized that as a consequence of ontogenic hypercapnia, the early life-stages of A. glauca would be tolerant to pH far below the extreme of the present range of the natural variability. The tested low pH (7.3) was 0.3 unit lower than the present extreme of the natural variability (7.6) but did not elicit a strong response beyond some subtle changes in shell morphology. These changes are difficult to interpret as it could be an expression of plasticity as previously observed for other invertebrate larvae exposed to low pH (e.g. [23]). These results are then in agreement with our hypothesis. While this is the first article documenting the impact of low pH on the early life stage of a nudibranch, some additional support for this hypothesis can be found in the grey literature. Carr & Podolsky [24] compared 6 different species of nudibranch (A. papillosa, Diaulula sandigensis, Doris montereyensis, Hermissenda crassicornis, Melibe leonine and, Triopha catalinae) laying egg mass using different strategies (Strings or ribbons). Similarly to our results, exposure to pH as low as 7.3 only triggered moderate responses such as small differences in shell length, time of hatching or weight of organic or inorganic masses.
In conclusion, available evidence supports the idea that encapsulation of eggs leads to pre-adaptation to low pH environment making nudibranch resilient to near-future ocean acidification.
Author Contributions
Conceptualization, C.Ö. and S.D.; methodology, C.Ö. and S.D.; experimentation, all; data analysis, All; writing, C.Ö. and S.D. All authors have read and agreed to the published version of the manuscript.
References
- Cooley, S.; Schoeman, D.; Bopp, L.; Boyd, P.; Donner, S.; Ito, S.; Kiessling, W.; Martinetto, P.; Ojea, E.; Racault, M.F.; et al. Oceans and Coastal Ecosystems and their Services. In IPCC AR6 WGII; Cambridge University Press: United Kingdom, 2022; pp. 379–550. [Google Scholar]
- Dorey, N.; Lancon, P.; Thorndyke, M.; Dupont, S. Assessing physiological tipping point for sea urchin larvae exposed to a broad range of pH. Glob. Chang. Biol. 2013, 19, 3355–3367. [Google Scholar] [CrossRef] [PubMed]
- Ventura, A.; Schulz, A.; Dupont, S. Maintained larval growth in mussel larvae exposed to acidified undersaturated seawater. Sci. Rep. 2016, 6, 23728. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, A. Reproduction of the Hermaphrodite Aeolidiella Glauca. Doctoral thesis, Acta Universitatis Upsaliensis, Uppsala, 2001.
- Vargas, C.; Lagos, N.; Lardies, M.; Duarte, C.; Manríquez, P.; Aguilera, V.; Broiman, B.; Widdicombe, S.; Dupont, S. Species-specific responses to ocean acidification should account for local adaptation and adaptive plasticity. Nature Ecol. Evol. 2017, 1, 84. [Google Scholar] [CrossRef]
- Vargas, C.A.; Cuevas, L.A.; Broitman, B.R. : San Martin, V.A.; Lagos, N.A.; Gaitán-Espitia, J.D.; Dupont, S. Upper environmental pCO2 drives sensitivity to ocean acidification in marine invertebrates. Nat. Clim. Change 2022, 12, 200–207. [Google Scholar] [CrossRef]
- Jager, T.; Ravagnan, E.; Dupont, S. Near-future ocean acidification impacts maintenance costs in sea-urchin larvae: identification of stress factors and tipping points using a DEB modelling approach. J. Exp. Mar. Biol. Ecol. 2016, 474, 11–17. [Google Scholar] [CrossRef]
- Dupont, S.; Lundve, B.; Thorndyke, M. Near future ocean acidification increases growth rate of the lecithotrophic larvae and juvenile of the seastar Crossaster papposus. J. Exp. Zool. 2010, 314B, 382–389. [Google Scholar] [CrossRef]
- Melzner, F.; Gutowska, M.A.; Langenbuch, M.; Dupont, S.; Lucassen, M.; Thorndyke, M.; Bleich, M.; Pörtner, H.O. Physiological basis for high CO2 tolerance in marine ectothermic animals: pre-adaptation through lifestyle and ontogeny? Biogeosciences 2009, 6, 2313–2331. [Google Scholar] [CrossRef]
- Gutowska, M.A.; Melzner, F. Abiotic conditions in cephalopod (Sepia officinalis) eggs: embryonic development at low pH and high pCO2. Mar. Biol. 2009, 156, 515–519. [Google Scholar] [CrossRef]
- Mehrbach, C.; Culberson, C.H.; Hawley, J.E.; Pytkowicx, R.M. Measurement of the apparent dissociation constants of carbonic acid in seawater at atmospheric pressure. Limnol. Oceanogr. 1973, 18, 897–907. [Google Scholar] [CrossRef]
- Dickson, A.; Millero, F.J. A comparison of the equilibrium constants for the dissociation of carbonic acid in seawater media. Deep-Sea Res. I: Oceanogr. Res. 1987, 34, 1733–1743. [Google Scholar] [CrossRef]
- Leandro, S.M.; Queiroga, H.; Rodríguez-Graña, L.; Tiselius, P. Temperature-dependent development and somatic growth in two allopatric populations of Acartia clausi (Copepoda: Calanoida). Mar. Ecol. Prog. Ser. 2006, 322, 189–197. [Google Scholar] [CrossRef]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Hall, B.K.; Wake, M.H. The origin and evolution of larval forms; Gulf Professional Publishing: United Kingdom, 1999. [Google Scholar]
- Seroy, S.K.; Grünbaum, D. Individual and population level effects of ocean acidification on a predator-prey system with inducible defenses: bryozoan-nudibranch interactions in the Salish Sea. Mar. Ecol. Prog. Ser. 2018, 607, 1–8. [Google Scholar] [CrossRef]
- Gibson, J.L; Harris, L.G. Cardiovascular responses to increased temperature and lower pH for six cold water Opisthobranch species. J. Exp. Mar. Biol. Ecol. 2023, 565, 151914. [Google Scholar] [CrossRef]
- Stumpp, M.; Hu, M.; Melzner, F.; Gutowska, M.A.; Dorey, N.; Himmerkus, N.; Holtmann, W.; Dupont, S.; Thorndyke, M.C.; Bleich, M. Acidified seawater impacts sea urchin larvae pH regulatory systems relevant for calcification. Proc. Natl. Acad. Sci. U.S.A. 2012, 109, 18192–18197. [Google Scholar] [CrossRef] [PubMed]
- Roleda, M.Y.; Boyd, P.W.; Hurd, C.L. Before ocean acidification: calcifier chemistry lessons. J. Phycol. 2012, 48, 840–843. [Google Scholar] [CrossRef] [PubMed]
- Tunnicliffe, V.; Davies, K.T.; Butterfield, D.A.; Embley, R.W.; Rose, J.M.; Chadwick, W.W., Jr. Survival of mussels in extremely acidic waters on a submarine volcano. Nat. Geosci. 2009, 2, 344–348. [Google Scholar] [CrossRef]
- Pechenik, J.A. Role of encapsulation in invertebrate life histories. Am. Nat. 1979, 114, 859–870. [Google Scholar] [CrossRef]
- Ellis, R.P.; Bersey, J.; Rundle, S.D.; Hall-Spencer, J.M.; Spicer, J.I. Subtle but significant effects of CO2 acidified seawater on embryos of the intertidal snail, Littorina obtusata. Aquat. Biol. 2009, 5, 41–48. [Google Scholar] [CrossRef]
- Chan, K.Y.K.; Garcia, E.; Dupont, S. Acidification reduced growth rate but not swimming speeds of larval sea urchins. Sci. Rep. 2015, 5, 9764. [Google Scholar] [CrossRef] [PubMed]
- Carr, G.M.; Podolsky, R.D. Effects of ocean acidification on embryo development: Does encapsulation matter? Friday Harbor Laboratories Student Research Papers, Friday Harbor, 2016.
Figure 1.
Nudibranchs and egg strings. a, d) Stereomicroscopic pictures. b, c) Digital camera picture (no scales). a) A. glauca on eelgrass showing dorsal cerata, antenna, rhinophores, eyes and an egg-string inside dorsal body (arrow). b) A. glauca eating the sea anemone Sagartiogeton viduatus. c) A. glauca laying an egg-string on aquarium wall. Note gonopore (go), from where the egg-string emerges. The first emerged string-part is broader than the latest emerging part. Arrows show invaginations of the egg-string and fractions of the egg-cells. d) An egg-string attached to eelgrass. Note: starting point (sp) of the egg-string, narrow part of the egg-string (en) and attachment sites of the string (arrows). Abbreviations: an, antenna; en, egg-string narrow part; ey, eyes; go, gonopore; rh, rhinophore; sp, starting point of egg-string. Figure 1b © Eduardo Infantes.
Figure 1.
Nudibranchs and egg strings. a, d) Stereomicroscopic pictures. b, c) Digital camera picture (no scales). a) A. glauca on eelgrass showing dorsal cerata, antenna, rhinophores, eyes and an egg-string inside dorsal body (arrow). b) A. glauca eating the sea anemone Sagartiogeton viduatus. c) A. glauca laying an egg-string on aquarium wall. Note gonopore (go), from where the egg-string emerges. The first emerged string-part is broader than the latest emerging part. Arrows show invaginations of the egg-string and fractions of the egg-cells. d) An egg-string attached to eelgrass. Note: starting point (sp) of the egg-string, narrow part of the egg-string (en) and attachment sites of the string (arrows). Abbreviations: an, antenna; en, egg-string narrow part; ey, eyes; go, gonopore; rh, rhinophore; sp, starting point of egg-string. Figure 1b © Eduardo Infantes.
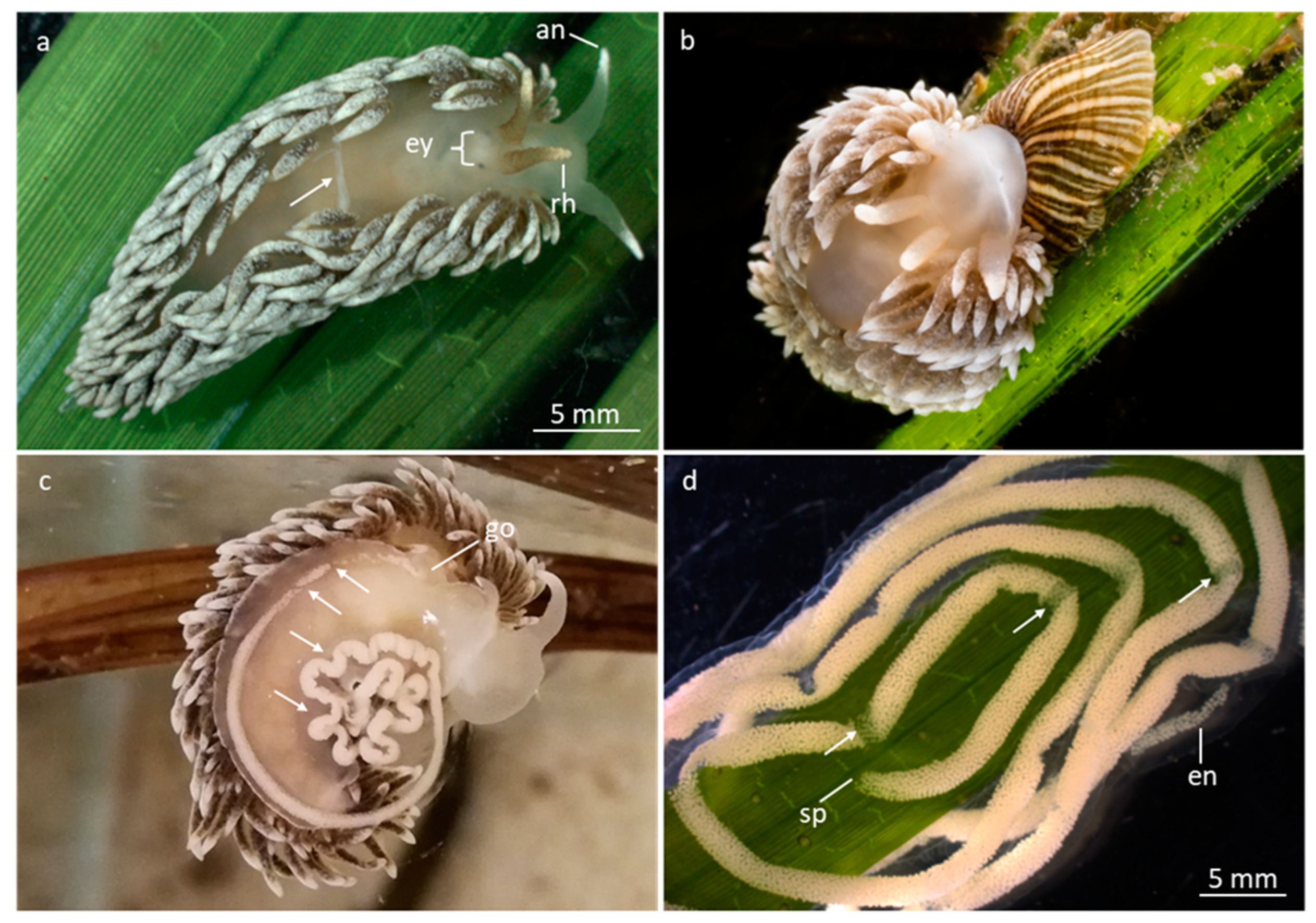
Figure 2.
Stereomicroscopic pictures. Development of egg cells from 1-cell stage to rotating veliger larvae in the egg-string in surface sea water, pH 8.0. a) 1-cell stage. b) 2 cells. c) 4 to 8 cells. Note spiral cleavage in third division. d) Early morula. e) Late morula. f) Blastula to gastrula with markings for velar apparatus. One gastrula in lateral view showing invagination, one rhomboid-shaped larva in bottom left corner. g) Gastrula in ventral/dorsal view with invagination and rhomboid-shaped larvae. h) Early rotating veliger larvae with velar apparatus, small shell, stomach, and eyes. i) Late veliger larvae with velar apparatus, stomach and liver, ready to leave the eggshell and the string. Note the larger size of larvae and shells compared to picture h. Abbreviations: ey, eye; gi, gastrula invagination; sh, shell; st, stomach; ve, velar apparatus; vm, velar marking.
Figure 2.
Stereomicroscopic pictures. Development of egg cells from 1-cell stage to rotating veliger larvae in the egg-string in surface sea water, pH 8.0. a) 1-cell stage. b) 2 cells. c) 4 to 8 cells. Note spiral cleavage in third division. d) Early morula. e) Late morula. f) Blastula to gastrula with markings for velar apparatus. One gastrula in lateral view showing invagination, one rhomboid-shaped larva in bottom left corner. g) Gastrula in ventral/dorsal view with invagination and rhomboid-shaped larvae. h) Early rotating veliger larvae with velar apparatus, small shell, stomach, and eyes. i) Late veliger larvae with velar apparatus, stomach and liver, ready to leave the eggshell and the string. Note the larger size of larvae and shells compared to picture h. Abbreviations: ey, eye; gi, gastrula invagination; sh, shell; st, stomach; ve, velar apparatus; vm, velar marking.

Figure 3.
Development from 1 to 16 cells in egg-strings (LM). Note light nucleus in cells. a) 1 cell. b) 2 to 4 cells. c, d) 4 cells in different developmental stages. e-g) Developing 4 to 8 cells. h) 8 cells (note the spiral cleavage; the 4 smaller cell present in grooves between the 4 larger cells). i, j) Developing 8 to 16 cells. k) 12 cells, approximately. l) 16 cells, approximately.
Figure 3.
Development from 1 to 16 cells in egg-strings (LM). Note light nucleus in cells. a) 1 cell. b) 2 to 4 cells. c, d) 4 cells in different developmental stages. e-g) Developing 4 to 8 cells. h) 8 cells (note the spiral cleavage; the 4 smaller cell present in grooves between the 4 larger cells). i, j) Developing 8 to 16 cells. k) 12 cells, approximately. l) 16 cells, approximately.
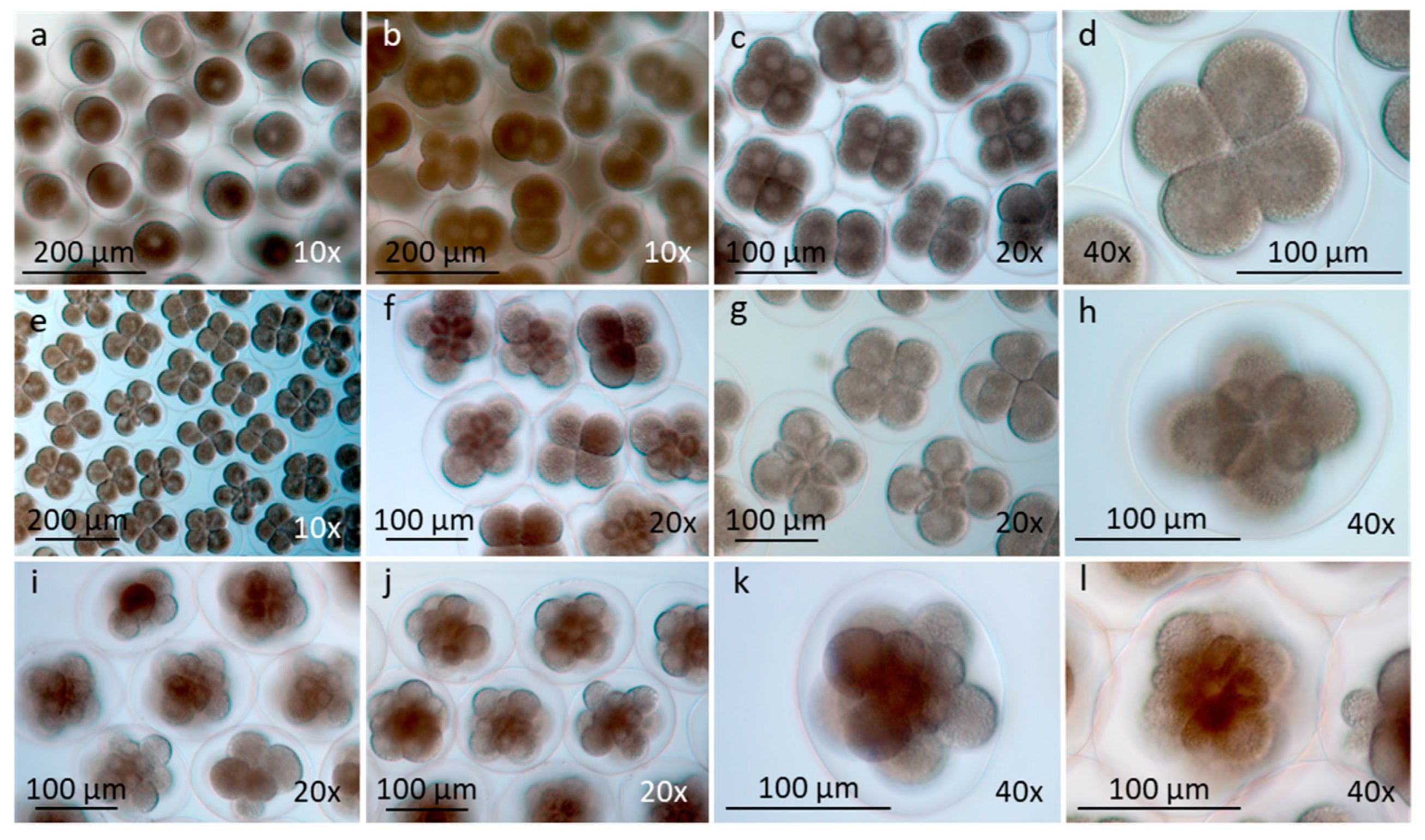
Figure 4.
Development in egg-strings from early morula to veliger larvae with developed small shell and velar apparatus (LM). a, b) Early morula up to 32 cells. c, d) Late morula up to 64 cells. e, f) Blastula with three polar bodies (f, arrow). g, h) Gastrula with invagination, lateral view (g, arrow) and dorsal view (h, arrow). i, j) Rhomboid shaped embryo with tiny cilia on velar region (i) and rotating (j). Note two polar bodies (arrow). Early developing rotating veliger larvae with ciliated velar apparatus, no shells (k) and small shell (l, arrows).
Figure 4.
Development in egg-strings from early morula to veliger larvae with developed small shell and velar apparatus (LM). a, b) Early morula up to 32 cells. c, d) Late morula up to 64 cells. e, f) Blastula with three polar bodies (f, arrow). g, h) Gastrula with invagination, lateral view (g, arrow) and dorsal view (h, arrow). i, j) Rhomboid shaped embryo with tiny cilia on velar region (i) and rotating (j). Note two polar bodies (arrow). Early developing rotating veliger larvae with ciliated velar apparatus, no shells (k) and small shell (l, arrows).
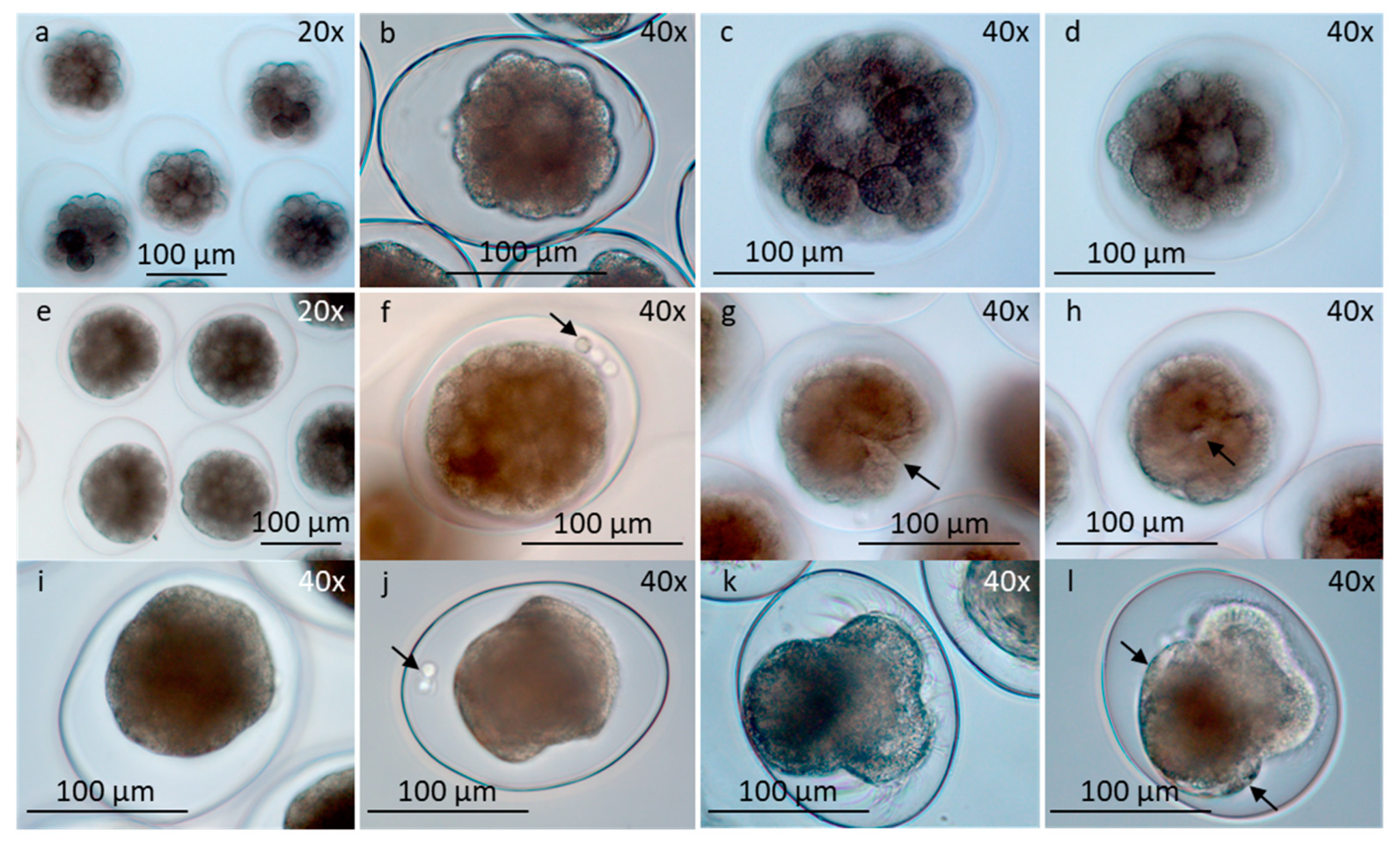
Figure 5.
Early developed larvae in egg-strings and free-swimming veliger larvae, seen in different views (LM). a-j) Larval development pH 8. a-d) Early developed larvae within eggshell showing velar apparatus, shell, eyes, statocysts, stomach liver and tendon. a) Rotating larvae organized in egg-string. b, c) Front view in different focuses. d) Side view. e-h) Free swimming veliger larvae showing eyes, mouth with tiny cilia and internal organs (e) tendon (f) mouth, stomach and liver (g), liver and canal system to stomach with structure inside (h). i) Larva with focus on stomach with cuboidal epithelia cells and a particle inside, eyes and foot. j) Larvae partly inside shell. Focus on eyes, foot and opercular part of shell. k, l) Development in pH 7.3. Defaulted larvae with velar apparatus, foot and unidentified internal organs as stomach and liver (* k), without shell (k), or small shell (l). m) Larvae completely inside shell. Note opercular part of shell. n) Empty rounded shell seen from below. o) Empty triangular shell seen from below. p) Living larvae in dorsal view inside shell, floating on surface water, measured length and width of shells (white lines). m,n,p) development in pH 8.0; o) development in pH 7.3. Abbreviations: ci, cilia; eg , eggshell; ey, eye; fo, foot; li, liver; lic, liver canal; mo, mouth; op, opercular part of shell; sh, shell; st, stomach; sta, statocyst; sts, structure in stomach; teb, tendon broad; ten, tendon narrow; ve, velar apparatus. Scale bars in all pictures are 100 µm.
Figure 5.
Early developed larvae in egg-strings and free-swimming veliger larvae, seen in different views (LM). a-j) Larval development pH 8. a-d) Early developed larvae within eggshell showing velar apparatus, shell, eyes, statocysts, stomach liver and tendon. a) Rotating larvae organized in egg-string. b, c) Front view in different focuses. d) Side view. e-h) Free swimming veliger larvae showing eyes, mouth with tiny cilia and internal organs (e) tendon (f) mouth, stomach and liver (g), liver and canal system to stomach with structure inside (h). i) Larva with focus on stomach with cuboidal epithelia cells and a particle inside, eyes and foot. j) Larvae partly inside shell. Focus on eyes, foot and opercular part of shell. k, l) Development in pH 7.3. Defaulted larvae with velar apparatus, foot and unidentified internal organs as stomach and liver (* k), without shell (k), or small shell (l). m) Larvae completely inside shell. Note opercular part of shell. n) Empty rounded shell seen from below. o) Empty triangular shell seen from below. p) Living larvae in dorsal view inside shell, floating on surface water, measured length and width of shells (white lines). m,n,p) development in pH 8.0; o) development in pH 7.3. Abbreviations: ci, cilia; eg , eggshell; ey, eye; fo, foot; li, liver; lic, liver canal; mo, mouth; op, opercular part of shell; sh, shell; st, stomach; sta, statocyst; sts, structure in stomach; teb, tendon broad; ten, tendon narrow; ve, velar apparatus. Scale bars in all pictures are 100 µm.
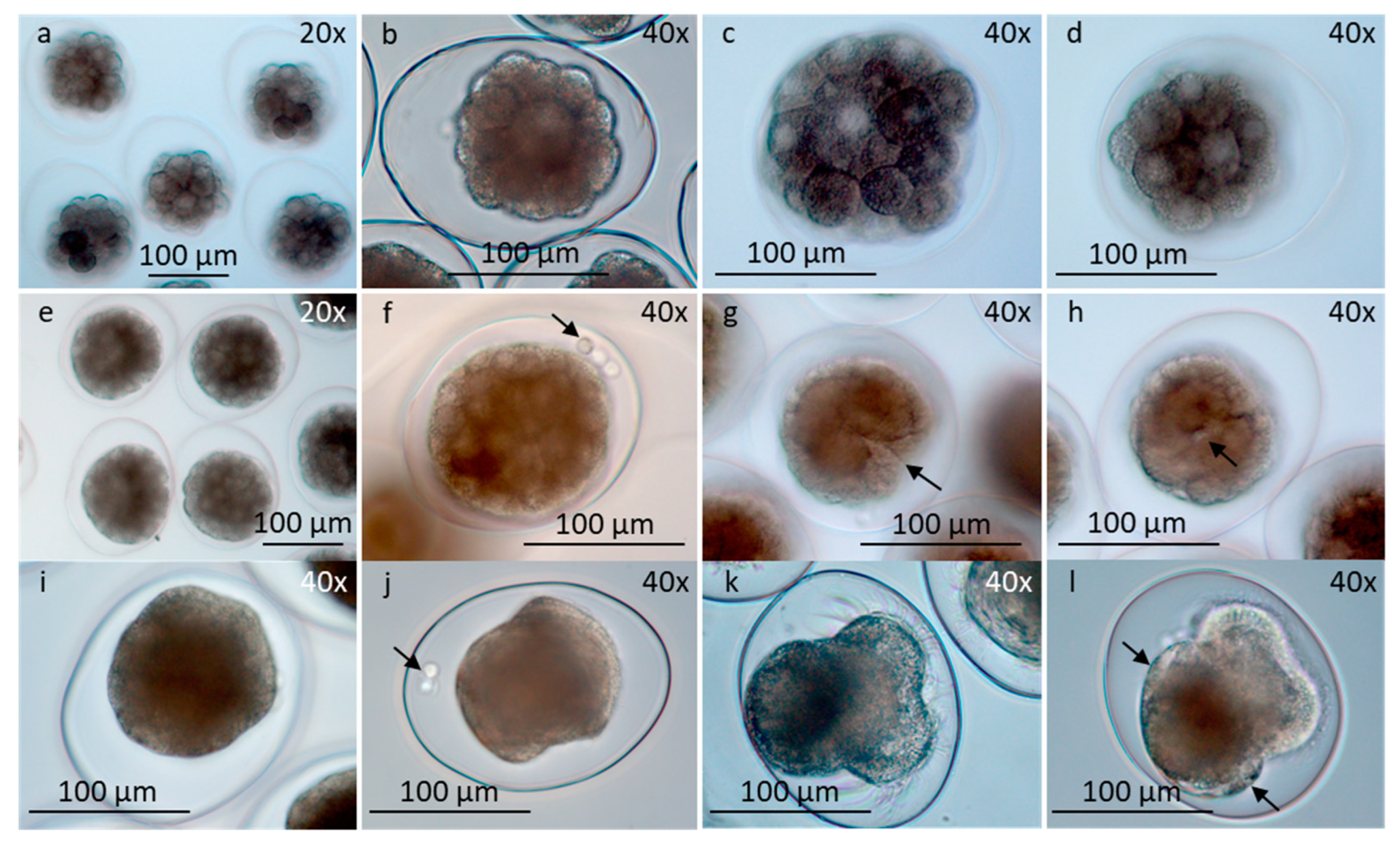
Figure 6.
Example of the development progression of Aeolidiella glauca eggs within a string (pH 8.0) over time (presented on the logarithmic scale), from 1 cell (0h) to free swimming veliger larvae (232h).
Figure 6.
Example of the development progression of Aeolidiella glauca eggs within a string (pH 8.0) over time (presented on the logarithmic scale), from 1 cell (0h) to free swimming veliger larvae (232h).
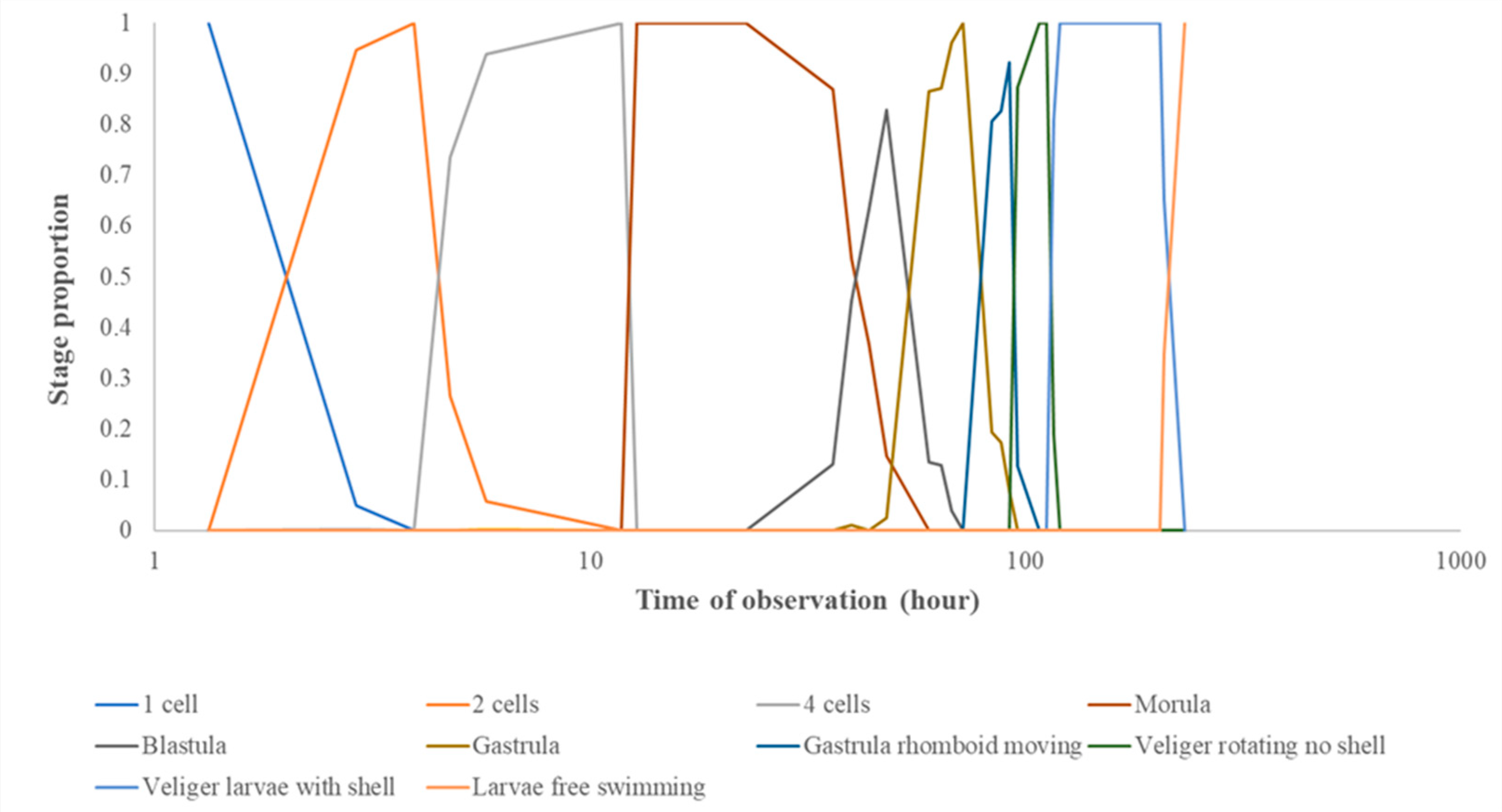
Figure 7.
Mean developmental time (MDT in h) for 6 developmental stages from late morula to veliger larvae with shell within egg-string.
Figure 7.
Mean developmental time (MDT in h) for 6 developmental stages from late morula to veliger larvae with shell within egg-string.
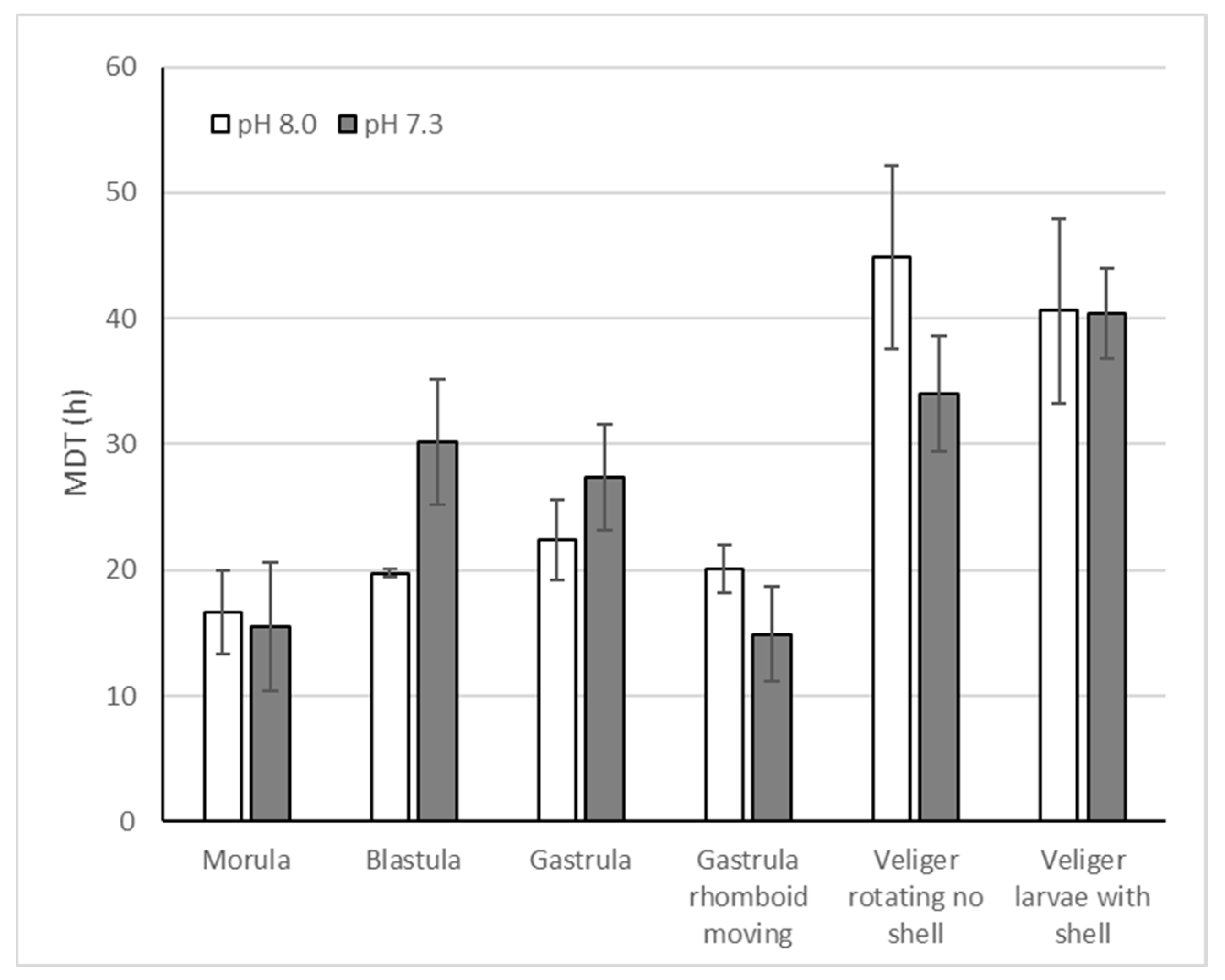
Figure 8.
Shell length (in mm) measured within 3 days post-hatching in 6 strings: 3 replicates (R1, R2, R3) and 2 pH (8.1, 7.3).
Figure 8.
Shell length (in mm) measured within 3 days post-hatching in 6 strings: 3 replicates (R1, R2, R3) and 2 pH (8.1, 7.3).
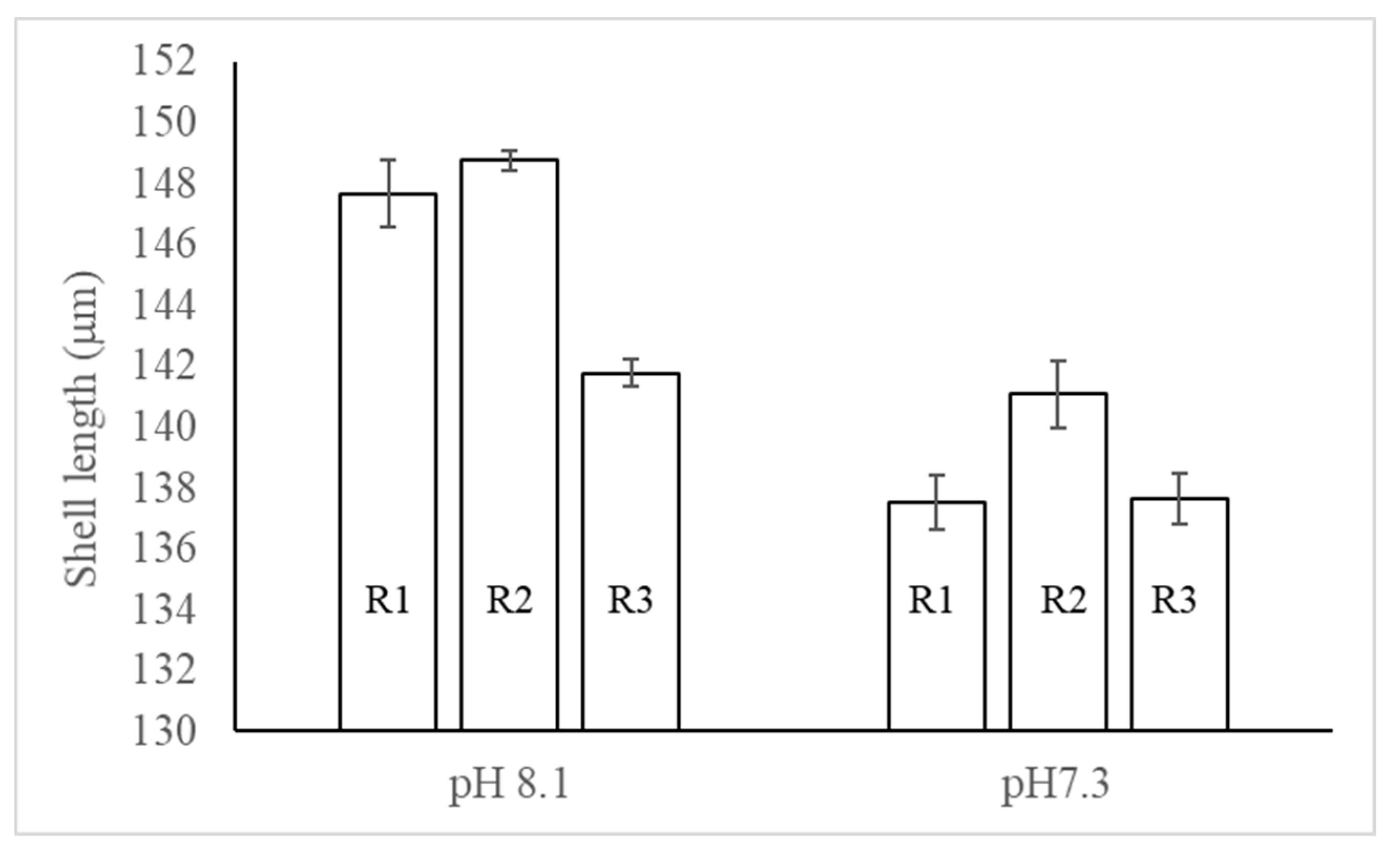
Figure 9.
Significant linear relationship between the larval shell length (in mm) and the larval shell width (in mm) measured within 3 days post-hatching for the 2 tested pH (8.1, 7.3). pH 8.1: F1,510=293.31, p<0.0001; pH 7.3: F1,430=417,35, p<0.0001.
Figure 9.
Significant linear relationship between the larval shell length (in mm) and the larval shell width (in mm) measured within 3 days post-hatching for the 2 tested pH (8.1, 7.3). pH 8.1: F1,510=293.31, p<0.0001; pH 7.3: F1,430=417,35, p<0.0001.
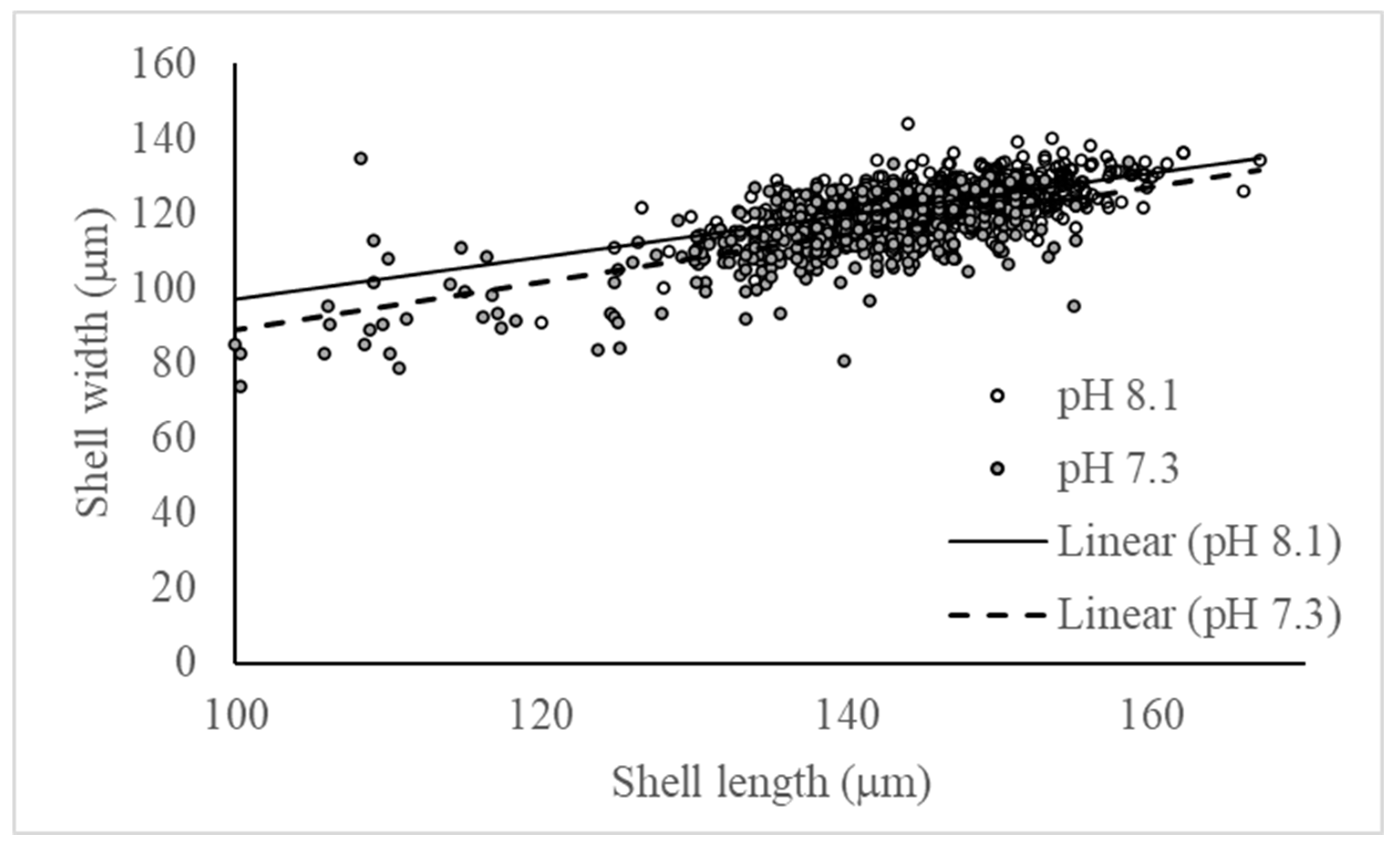

Table 1.
Seawater carbonate chemistry parameters presented as mean ± sem. Seawater pH on the total scale (pHT), temperature (18.1±0.6 °C), salinity (31±1) and total alkalinity (2402±11 mmol kg−1) were used to calculate CO2 partial pressure (pCO2; matm) as well as aragonite (Ωca) and calcite (Ωar) saturation states.
Table 1.
Seawater carbonate chemistry parameters presented as mean ± sem. Seawater pH on the total scale (pHT), temperature (18.1±0.6 °C), salinity (31±1) and total alkalinity (2402±11 mmol kg−1) were used to calculate CO2 partial pressure (pCO2; matm) as well as aragonite (Ωca) and calcite (Ωar) saturation states.
| Target pH | Measured | Calculated | ||
|---|---|---|---|---|
| pHT | pCO2 (matm) | Wca | War | |
| pH 8.0 | 8.07±0.02 | 411±20 | 4.38±0.16 | 2.81±0.10 |
| pH 7.3 | 7.31±0.01 | 2739±21 | 0.88±0.01 | 0.57±0.01 |
Table 2.
ANOVA of Mean Development Time as a function of pH for 6 developmental times.
| Stage | F1,5 | p |
|---|---|---|
| Morula | 0.04 | 0.86 |
| Blastula | 4.38 | 0.10 |
| Gastrula | 0.86 | 0.41 |
| Gastrula rhomboid moving | 1.56 | 0.28 |
| Veliger rotating no shell | 1.59 | 0.28 |
| Veliger larvae with shell | 0.01 | 0.98 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



