Submitted:
07 September 2023
Posted:
11 September 2023
Read the latest preprint version here
Abstract
Integrated cultivation strategies can significantly improve rice yield. However, it is unclear how integrated cultivation practices improve soil microbial diversity and its related mechanism in rice field. Therefore, four integrated cultivation practices i.e., no N application (N0), local farmers’ practice (FP), high-yield and high-efficiency practice (HYEY), and super-high-yield cultivation practice, impacts on soil properties and structure composition of microbial community were explored. Compared with N0, SHY treatment, significantly increased soil alkaline nitrogen and organic matter contents. HYHE and SHY treatments significantly altered soil fungal community and Alpha diversity. Based on Venn diagram analysis, the composition of bacterial communities under all treatments was similar, but with different compositions of fungal communities. Cluster analysis indicates that the Proteobacteria was the absolute dominant group of bacterial communities, and Chloroflexi, Bacteroidetes, and Acidobacteria were the dominant group. Different cultivation methods also led to changes in the main factors affecting soil bacteria and fungi composition. Available potassium and organic matter were the main environmental factors that affected bacteria, with the strong of available potassium on the soil. To sum up, SHY and HYEY were beneficial management options in terms of improved soil fertility Alpha diversity of the soil bacterial community respectively.
Keywords:
rice
; integrated cultivation practices
; microbial diversity
; environmental factors
1. Introduction
Microorganisms are ubiquitous in various environments, while the soil is the habitat with the most abundant microbial diversity on the earth [1]. Soil microorganisms can play a key role in soil carbon mineralization and accumulation, which determines soil nutrient circulation and soil fertility [2]. Nevertheless, soil microbes are very sensitive to subtle environmental differences caused by environmental pressures or natural disturbances [3], while changes in the soil can promote biogeochemical cycles that contribute directly or indirectly to crop growth and quality, nutrient cycling, and sustainability of soil productivity [4]. There is a need to assess how cultivation, fertilization, and irrigation can affect soil microbial diversity and community structure in rice paddy fields. There identifying efficient management practices in terms of improved soil microbial and nutrient status are urgently needed.
Fertilization changed soil nutrient status, such as increased soil organic carbon and total nitrogen, which provided sufficient nutrients for microorganisms and thus induces changes in soil microbial community composition [5,6]. Long-term application of nitrogen fertilizer affected the composition of the rhizosphere bacterial community, and the diversity of rhizosphere bacteria increased with the increase of nitrogen application [7]. Nitrogen fertilizer decreased the diversity of nitrification gene amoA in the paddy soil and significantly changed the community composition of amoA and hao [8]. Phosphate fertilizer promoted soil phosphorus activation and release, significantly increased soil available phosphorus content, and then changed soil microbial community composition [9]. The relative abundance of beneficial microorganisms under the action of phosphorus fertilizer was increased and became the dominant flora. Balanced nitrogen, phosphorus, and potassium fertilization in the rice ecosystem significantly increased bacterial diversity [10]. The irrigation mode directly affected the change in soil water content, while the soil microbial community structure and function were directly affected by the degree of soil water fluctuation [11]. Even the drastic fluctuation of soil moisture affected the microbial-mediated turnover of soil elements such as carbon and nitrogen, and then continuously affected subsequent soil processes and ecosystems [12]. Dangi et al. [13] showed that furrow irrigation significantly improved the number of gram-positive bacteria and actinobacteria, while drip irrigation significantly increased the number of fungal gram-negative bacteria. Changes in irrigation mode can significantly affect the functional diversity of soil microbial community, and the carbon sources used by soil microorganisms in flooded irrigation for a long time were mainly lipids, the carbon sources used in intermittent irrigation and moistening irrigation were mainly amino acids and sugars, respectively [14]. Changes in cultivation mode strongly altered the soil microenvironment mainly through soil disturbance, and then affected the structure and functional diversity of the soil microbial community [15]. The combination of conventional tillage and rotary tillage to retain crop residues could help to establish a stable and functional soil environment for soil bacterial communities in double-cropping paddy fields in southern China, long-term conventional tillage and rotation contributed to the improvement of microbial richness, Shannon index, and Macintosh index [16]. The microbial community structure of light and simplified direct seeding rice field and mechanized transplanting rice field was significantly different from transplanting treatment [17]. Rice seedling broadcasting partially overlapped with transplanting, but the impact of four cultivation methods on the Shannon index and Simpson index of soil microorganisms did not reach a significant level.
Heilongjiang Province is the main distribution area of rice in the cold region of China, and is the largest and the most important japonica rice production area. The rice planting area has reached 3.72 million hectare and the total rice yield reached 26.114 million tons in 2019 [18]. In recent years, reducing the yield gap and increasing total crop yield via., integrated cultivation techniques is a hot topic in rice production. However, at present, the effect of multiple cultivation techniques on soil microbes is not clear. Therefore, it is particularly important to construct several cultivation techniques and explore how these practices change soil microorganisms and soil nutrient status. In our previous studies, we found that increased N, P, and K accumulation from the heading stage to the mature stage under HYEY and SHY treatment may contribute to increased rice yield [19]. Comparing the differences in soil microbial diversity and community structure in rice fields under different cultivation methods elucidated the mechanism of rice yield difference in the cold regions.
2. Materials and Methods
2.1. Plant materials and experimental field
The experiment was carried out in the paddy field of farmers in Wangjiaweizi (46°40′ N, 125°07′ E), Daqing City, Heilongjiang Province in 2021 and 2022. The soil of the experimental field was sodic saline-alkali soil. The rice variety Kenjing 8 (13 leaves on the main stem) was used as the experimental material. The seeding, transplanting, and harvest were performed on 19 April, 16 May, and 23 September, respectively.
2.2. Experimental design
Experiments were performed using a randomized block design with a plot size of 150m2 three replicates per treatment. The experiment consisted of four treatments: no N application (N0), local farmers’ practice (FP), high-yield and high-efficiency practice (HYEY), which applies alternate wetting and drying irrigation and increasing manure to improve yield and resource-use efficiency, and super-high-yield practice (to obtain a high yield regardless of labor or fertilizer input; SHY). Treatment details are given in Table 1. Seedlings were raised using dry bed method under greenhouse conditions (with a sowing rate of 250 g/m2 of sprouts) and pot method (with a sowing rate of 3 sprouts per pot hole). Seedlings were transplanted after 35 days of sowing with a parallel row (transplanting density 30 cm × 10 cm) and alternate wide and narrow rows planting patters [transplanting density (20-40) cm × 13.3cm]. Irrigation including shallow water flood irrigation i.e., maintained a 2-3 cm shallow water layer during whole growth period and alternate wetting and drying irrigation for example, first irrigation was 2-3 cm shallow water layer until the soil water potential reached -20 kPa (monitored the soil water potential with a negative pressure gauge, placed the bottom of the clay head at the 15 cm soil layer). After this, irrigated a 2-3 cm shallow water layer frequently during entire growth period and stopped irrigation one week before harvest (light sun drying at the end of tillering stage). Chemical fertilizers included urea (N46%), diammonium phosphate (N18%, P2O5 46%), potassium sulfate (K2O 50%), nano silicon fertilizer (effective silicon ≥ 55%, manufacturer's name). Organic fertilizer was classified as compound fertilizer (N+P+K ≥ 5%, organic matter ≥ 40%). Nano-silicon fertilizer was applied to seedling bed before transplanting. Guifuji was applied as basal fertilizer. Nitrogen fertilizer was applied at 4:3:1:2 ratio during pre-transplanting, re-greening, primary tillering, panicle initiation, and spikelet differentiation stages, respectively. Phosphate fertilizer (P2O5) was applied as basal fertilizer. Potassium fertilizer (K2O) was applied at the ratio of 6:4 at the pre-transplanting and panicle initiation stage, respectively. The pre-transplanting, re-greening, primary tillering, panicle initiation, and spikelet differentiation fertilizer were applied at 12d before transplanting (May 1, 2021 and May 4, 2022), vegetative (May 19, 2021and May 28, 2022),8.5 leaf age (June 2, 2021 and June 25,2022) and 10.5 leaf age (July 6, 2021 and July 12,2022), respectively. Each treatment was irrigated and drained separately to prevent mutual influence. Weeds, pest and diseases were controlled.
2.3. Soil sample collection
At the heading stage, rhizosphere soil was collected at depth of 20 cm in 2022. Then tried to take the soil close to the root surface (mixed 3 rice plants to 1 sample for each plot). A soil sample was naturally dried, grinded and sifted by 1 mm sifter for soil chemical determination. A fresh soil sample was transported to the laboratory in the ice bags and was stored at −80 ℃ refrigerator for the determination of soil microbial diversity.
2.4. Measurement of soil physical and chemical properties
Soil organic matter content was determined by potassium dichromate volumetric method and external heating method; soil alkali-hydrolyzed nitrogen content was determined by diffusion absorption method. Soil available phosphorus content was determined by extraction of sodium bicarbonate-molybdenum antimony anti-colorization method. Soil available potassium content was determined by ammonium acetate extract-AAS method [20]. Soil microbial diversity was investigated by Biomarker Technologies Co,LTD (Shanghai). The Illumina HiSeq technique was used for samples clustering and diversity analysis.
2.5. Data analysis
Microsoft Excel 2010 was used for data sorting, and the physical and chemical properties of soil and microbial diversity index were statistical analyzed by SPSS 170.0 software. The differences were identified statistically significant at P<0.05 level. The bioinformatic analysis in this study was completed using the Biomarker Biocloud Platform (www.biocloud.org, accessed on 20 November 2020).
3. Results
3.1. Effects of integrated cultivation practices on soil nutrients in paddy field
Changes in available nutrients, organic matter, and pH of paddy soil under integrated cultivation practices are shown in Table 2. The soil alkaline nitrogen content among the four treatments is in the order SHYP>HYP>FP>N0. The alkali hydrolyzed nitrogen content of SHYP and HYP was significantly higher than other treatments. Compared with N0 and FP, SHYP treatment increased alkali-hydrolyzed nitrogen by 43.4% and 28.1%, while HYP treatment increased alkali-hydrolyzed nitrogen by 28.2% and 14.5%, respectively. The content of available phosphorus and available potassium under N0 soil was the highest, while the content of FP was the lowest. Available potassium under HYP and SHYP was increased by 38.4% and 15.9%, respectively. SHYP significantly increased the content of soil organic matter by 25.6%, 14.9%, and 11.5% compared with N0, FP, and HYP, respectively; Compared with N0, FP, HYP, and SHYP showed a decreasing trend in soil pH.
3.2. Soil microbial diversity in paddy fields (Alpha diversity)
Compared with FP treatment, the bacterial ACE, Chao1, and Shannon index under HYP treatment was increased, while the Simpon index significantly decreased (Table 3). Under SHYP treatment, the bacterial ACE, Chao1, and Shannon index were decreased, while the Simpon index increased. The fungal ACE index and Chao1 index showed N0>FP>SHYP>HYP order. The fungal ACE index under FP, HYP, and SHYP treatments was significantly reduced by 30.09%, 36.12%, and 28.93% compared with N0, respectively. The Chao1 index of fungi under FP, HYP, and SHYP treatment was reduced by 30.06%, 36.50%, and 28.76% compared with N0, respectively. Fungal Simpson was in the following order HYP>SHYP>N0>FP. N0, FP, and SHYP were reduced by 89.44%, 93.22%, and 87.08% compared to HYP, respectively. The FP decreased by 35.76% compared to N0; SHYP increased by 22.42% compared to N0. Integrated cultivation practices mainly affect the fungal community Alpha diversity index, bacterial community Alpha diversity index has little impact.
3.3. Correlation analysis of soil nutrients and microbial diversity index
Soil organic matter significantly affected bacterial ACE and Chao1, with correlation coefficients of -0.583 (Table 4). Therefore, higher organic matter content can result in lower soil bacterial richness. However, the bacterial diversity index of Simpson and Shannon were not significantly affected by soil nutrients. Compared with bacteria, there is a certain difference in the impact of soil nutrients on fungal diversity. Fungal richness index ACE and Chao1 are significantly or extremely affected by alkaline nitrogen, available potassium, and organic matter. Soil-available potassium was the main influencing factor. However, the fungi diversity index Simpson and Shannon were not significantly affected by soil nutrients.
3.4. Wayne map of the OUT distribution of soil bacteria
A total number of bacterial communities OTUs was 1898, and the bacterial OTUs number common by N0, FP, HYEY, and SHY was 1894, 99.79% of the total, while specific OUT’s numbers of all four treatment were 0. FP, HYEY, and SHY could not increase the OTUs number of the bacterial community specificity, and the composition of bacterial community was similar under integrated cultivation practices (Figure 1). The total number of fungal communities OTUs was 1413, and the fungal OTUs number common by N0, FP, HYEY, and SHY was 395, 27.95% of the total, while specific OTUs numbers of N0, FP, HYEY, and SHY were 86, 123, 53 and 107, 26.11% of the total.
3.5. Principal component analysis of microbiology
The contribution of the first principal component (PC1) of abscissa to the OTU difference of bacteria was 34.19%, the contribution of the second principal component (PC2) of ordinate to the difference of bacteria was 18.80%, and the cumulative contribution rate of the first principal component (PC1) and the second principal component (PC2) was 52.99% (Figure 2a). Soil bacterial community composition under the four cultivation methods was similar. The results of Figure 2b showed that the contribution of the first principal component (PC1) of abscissa to the OTU difference of fungal was 22.04%, the contribution of the second principal component (PC2) of ordinate to the difference of fungal was 17.67%, and the cumulative contribution rate of the first principal component (PC1) and the second principal component (PC2) was 39.71%. HYEY and SHY had obvious aggregation, while N0 and FP behaved as separate separations.
3.6. Composition structure of bacterial and fungal community at the phylum level
To explore the changes in species composition of bacteria and fungi in rice fields under different cultivation methods, the dominant phyla of the top 10 with relative abundance > 1% were analyzed by cluster analysis (Figure 3a, b). At the phylum level of bacterial (Figure 3a), Proteobacteria was an absolute dominant group, 34.03% - 35.50% of the total, Chloroflexi, Bacteroidetes, and Acidobacteria were preponderant groups of each treatment, 17.47% - 20.61%, 16.03% - 20.17% and 10.75% - 11.61%, respectively. The relative abundance of Proteobacteria of FP, HYEY, and SHY increased by 2.59%, 4.32%, and 3.73% respectively compared with that of N0. The relative abundance of Chloroflexi showing FP>HYEY>N0>SHY, and the trend of Bacteroidetes was opposite of Chloroflexi, showed SHY >N0>HYEY> FP. The relative abundance of Chloroflexi of FP and HYEY increased by 10.19% and 5.35%, respectively compared with N0; the relative abundance of Chloroflexi of SHY decreased by 6.53% compared with N0. The relative abundance of Bacteroidetes of SHY was increased by 7.06% compared with N0; the relative abundance of Bacteroidetes of FP and HYEY was decreased by 14.92% and 12.31%, respectively compared with N0. This indicated that there was some competition between Chloroflexi and Bacteroidetes. The Patescibacteria, Actinobacteria and Verrucomicrobia of FP, HYEY, and SHY was increased compared with N0. A similar trend of Patescibacteria and Verrucomicrobia was HYEY>FP>SHY>N0, and the Actinobacteria of FP was the largest. The relative abundance of Acidobacteria, Nitrospirae, Gemmatimonadetes, and Firmicutes of HYEY and SHY all showed a trend of less than N0 and FP.
At the phylum level of funguses (Figure 3b), the relative abundance of phyla of others (the relative abundance ≤1 or unknown phyla) of four treatments was larger, 34.03% - 35.50% of the total. Ascomycota was an absolute dominant group, 27.71% - 46.64% of the total, while the remaining nine phyla had a low relative abundance. The relative abundance of Ascomycota of FP, HYEY and SHY was increased by 42.80%, 68.31%, and 5.27%, respectively compared with N0. This explained that FP, HYEY, and SHY had changed the dominant phyla of fungi. The Mortierellomycota, Glomeromycota, Kickxellomycota, and Blastocladiomycota of FP, HYEY, and SHY were increased over N0. Notable, Kickxellomycota and Blastocladiomycota were not present in N0, but Kickxellomycota and Blastocladiomycota appeared in FP, HYEY, and SHY. While Aphelidiomycota only existed in N0 and HYEY, the relative abundance was 0.029% and 0.075%, respectively. Moreover, the Basidiomycota of FP and SHY all increased compared with N0. Rozellomycota, Chytridiomycota, and Monoblepharomycota of FP, HYEY, and SHY were decreased compared with N0.
Based on RDA linear model, the relationship between soil bacteria, fungal community and environmental factors in rice paddy fields was explored (Figure 4a, b). The principal elements 1 and 2 of environmental factors respectively explained 18.72% and 15.79% of the bacterial community, and the total explained 34.51% (Figure 4a). The environmental factors that affected the composition of species at the phylum level of bacterial communities were available potassium and organic matter. Proteobacteria was positively correlated with soil alkali-hydrolyzed nitrogen and organic matter and, negatively correlated with available phosphorus, pH, and available potassium. Chloroflexi was negatively correlated with alkali-hydrolyzed nitrogen, available phosphorus, available potassium, organic matter, and pH. This also explained the low relative abundance of Chloroflexi in N0. Bacteroidetes were the opposite, which all positively correlated with alkali-hydrolyzed nitrogen, available phosphorus, available potassium, organic matter, and pH. Acidobacteria was positively correlated with soil alkali-hydrolyzed nitrogen and organic matter, and negatively correlated with available phosphorus, pH, and available potassium.
According to Figure 4b, the principal elements 1 and 2 of the environmental factors respectively explained 29.79% and 14.23% of the fungal community, and a total explained 44.02%. Available potassium was the main factor that affected the species composition of the fungal community at the phylum level, followed by organic matter. Ascomycota, Basidiomycota, Mortierellomycota, and Blastocladiomycota were positively correlated with soil alkali-hydrolyzed nitrogen and organic matter, and negatively correlated with available phosphorus, available potassium, and pH, but the degree of correlation varied by fungal species. Rozellomycota was negatively correlated with soil alkali-hydrolyzed nitrogen and organic matter, and positively correlated with available phosphorus, available potassium, and pH.
4. Discussion
4.1. Effect of integrated cultivation practices on the Alpha diversity of soil microorganisms
Alpha diversity occurs within a particular region or ecosystem, and is a comprehensive indicator reflecting the species 'richness and diversity of a single sample [21]. In the current study, integrated cultivation practices affected the richness index (ACE, Chao1 index) and diversity index (Shannon, Simpson index) of soil bacteria. HYEY had the highest richness indexes (ACE and Chao1 index) and the highest Shannon index, and had the lowest Simpson index. This indicated that high-yield cultivation methods can not only improve bacterial community richness, but also improve bacterial community diversity. The soil microbial diversity of FP was second only to that of HYEY. The changing trend of soil microbial diversity of SHY was contrary to HYEY. SHY had the lowest richness indexes (ACE and Chao1 index) and Shannon index, and the highest Simpson index due to excessive nitrogen application. Microbial diversity is very sensitive to environmental changes and management practices [22], like fertilization or long-term nutrient inputs can easily affect the richness and diversity of the soil bacterial communities [23]. Overuse of nitrogen application in dryland soil led to reduced diversity of bacterial communities [24]. It has been shown that bacterial diversity was negatively associated with nitrogen application [25]. Therefore, excessive nitrogen application may be a factor in reduced bacterial diversity under SHY soil. The effect of nitrogen application on the bacterial diversity in the soil was also significantly related to the water management system [26]. This may be because bacteria were able to move and acquire nutrients in the soil environment using the flow of water membranes [27]. Hamer et al. [28] believed that with soil moisture content reduced, part of the active water-sensitive microbial cells disintegrated and died. Alternating dry and wet conditions can lead to reduced soil bacteria diversity in the rice ecosystem [29]. Nitrogen application level of HYEY may be suitable for saline-alkali soil microbial growth, and on the other hand, Guifuji biological organic fertilizer may be conducive to the formation of microbial diversity. The combined use of two factors eliminated the negative effects of alternate drying and wetting. High levels of nitrogen application and alternate drying and wetting may be the main reasons for reduced microbial diversity for SHY. Soil pH has been shown the main factor affecting the structure and abundance of soil microbial communities [30]. In particularly, bacteria were most significantly affected by pH [31]. The relevant analysis of this test found that organic matter was the main factor affecting the bacterial diversity of soil bacteria, and the higher the soil organic matter content, the lower the richness and degree of soil bacterial community diversity. After the application of nitrogen and phosphorus and potassium fertilizer, the soil pH was weaker acidic (i.e. neutral or alkaline) which determine soil organic carbon as a key factor affecting the soil bacterial diversity [26]. In this test, due to the obstacles of saline-alkali soil itself, changes in soil pH from N0 to SHY were small, and the impact of soil pH on bacterial diversity may also be reduced, and organic matter, as the matrix of heterotrophic bacteria, was more important in affecting bacterial diversity than soil pH. In addition, rice transplanting method, fertilization, soil available nutrients, and other biological and non-biological factors all affected the bacterial diversity of the soil.
In this study, compared to the Alpha diversity of soil bacterial communities, the cultivation methods had a more significant influence on the richness and diversity of the soil fungal phyla. The ACE and Chao1 indexes of the fungal community of 4 treatments all showed N0>FP>SHY>HYEY, and those of N0 were significantly higher than those of FP, HYEY, and SHY; the Shannon index of the fungal community of 4 treatments showed FP>N0> SHY>HYEY, and that of FP was significantly higher than N0, HYEY, and SHY; This suggested that HYEY and SHY were detrimental to fungal community growth. Comparison with Alpha diversity of soil bacterial communities showed that HYEY was beneficial to bacterial growth and inhibited fungal growth. This was likely because the HYEY soil environment was suitable for bacterial growth, increasing the number of some beneficial microbial bacteria in the bacterial community, and the coexistence of multiple soil microorganisms inhibited each other [32]. Through soil microbial competitive exclusion [33], certain beneficial microbial bacteria in the bacteria community may inhibit harmful fungi in the fungus community, thus reducing the richness and diversity of fungal phyla in HYEY soil. Inorganic fertilizers not only significantly reduced fungal species, but also led to a decrease in fungal richness and diversity, and altered dominant fungal populations [34]. The application of nitrogen reduced the richness and diversity of fungal classification and changed the community structure [35]. This may be because fertilization stimulated the mass growth of certain specific microorganisms, but suppressed other microorganism growth, causing a decline in diversity [32]. Studies also believed that root secretions affected fungal diversity [36]. Peanut root secretions had more influence on the fungal community than on the bacterial community, and the growth of Mortierella and terrestrial bi-basidiomycetes was inhibited by the peanut root secretion of phenolic acid, thus reducing the relative abundance [37]. Fertilization promoted crop root metabolism and increased root secretion [38]. Therefore, rice root secretions may be one of the factors affecting the diversity of fungal communities. In addition, the mycelium of soil fungus can both redistribute water [39], or obtain nutrients from the soil pores to relieve the pressure of water stress [40], so the fungus is more resistant to water stress than bacteria [41]. Bacterial growth was reduced by alternating dry and wet, and the fungus remained unchanged [42]. In this test, dry-wet alternation may have less or no impact on the diversity of fungal communities in HYEY, SHY soil. Alkali-hydrolyzed nitrogen and organic matter were significantly negatively related with ACE, Chao1 index, available potassium was significantly positively related with ACE, Chao1 index, alkali-hydrolyzed nitrogen, the relationship between available potassium, organic matter, and Shannon、Simpson index was insignificant. The high content of alkali-hydrolyzed nitrogen and organic matter in HYEY, SHY soil resulted in a reduction of soil fungal richness. Available potassium content in N0 soil was the highest which resulted in increased soil fungal richness.
4.2. Effect of integrated cultivation practices on the composition of soil microbial community
In this study, the Proteobacteria in rhizosphere soil were all absolute dominant groups under different cultivation methods, Chloroflexi, Bacteroidetes, and Acidobacteria were dominant groups. These bacteria may play an important role in rhizosphere soil. Proteobacteria is a symbiotic bacterium that grows rapidly in a C-rich environment [43] and is involved in the biocirculation of essential mineral nutrients in the soil [44].The high proportion of Proteobacteria in rhizosphere soil was conducive to the maintenance of soil fertility and plant growth [45].The Proteobacteria was highly responsive to N input and the relative abundance increased with nitrogen application [26]. The test results showed that FP, HYEY, SHY increased the relative abundance of Proteobacteria compared to N0, and Proteobacteria abundance of HYEY, SHY was higher than FP. RDA analysis showed that Proteobacteria was positively associated with alkali-hydrolyzed nitrogen and organic matter. This indicated that the increase of Proteobacteria abundance in rhizosphere soil favored the accumulation of alkali-hydrolyzed nitrogen and organic matter, while high organic matter content had high organic carbon content. This in turn stimulated Proteobacteria growth. Proteobacteria was negatively associated with available phosphorus, available potassium, and pH. This further explained why the Proteobacteria relative abundance of SHY was slightly lower than HYEY. In this study, FP, HYEY, and SHY reduced the relative abundance of the Firmicutes compared with N0, and showed the opposite trend to Proteobacteria. There was a nutritional competition between Proteobacteria and Firmicute [16]. Rodrigues et al. [46] believed that Firmicutes was also a trophic organism, and high carbon stimulated its growth. But Firmicutes was also an oligotrophic bacterium [47], and high nitrogen application reduced its abundance [48]. According to the above views, the presence of Proteobacteria and the relationship between nitrogen application amount and Firmicutes explained the decrease of the Firmicutes relative abundance of FP, HYEY, and SHY. RDA analysis found that Firmicutes had a positive correlation with alkali-hydrolyzed nitrogen, available phosphorus, available potassium, organic matter, and pH. This explained the reason that the Firmicute’s relative abundance of HYEY, and SHY was higher than that of FP. Chloroflexi was mainly inclined to anaerobic conditions [49], was the main degradation of sugar and polysaccharide [50], enhanced the crop absorption of soil nutrients [51]. This study showed that FP, and HYEY increased the relative abundance of Chloroflexi compared to N0, while SHY decreases the relative abundance of Chloroflexi. RDA analysis showed that Chloroflexi had a negative correlation with alkali-hydrolyzed nitrogen, available phosphorus, available potassium organic matter, and pH, which suggested that the reduced relative abundance of Asperglum favors the preservation of SHY soil fertility. Bacteroidetes was an anaerobic or facultative anaerobic eutrophic bacteria, a relative abundance of Bacteroidetes increased at a high organic level [52]. This study showed that Bacteroidete’s relative abundance of FP, HYEY decreased compared to N0, and the soil organic matter content of SHY was significantly higher than N0, FP, HYEY, thus the relative abundance of Bacteroidetes was the highest. Bacteroidetes had a Phosphate-solubilizing function and was positively associated with available phosphorus content [53]. Acidobacteria was recognized as oligotrophic bacteria [54]. The high nutritional environment of HYEY, SHY reduced the relative abundance of the Acidobacteria compared to FP. Generally, acid bacteria can adapt to acidic soil environments with low pH [55]. This test also showed that Acidobacteria had a negative association with pH.
In agricultural soils, Ascomycota was the largest group of fungi, which degraded nonpeer fishable organic matter in the soil, and increased as nutrients increased [56]. The experimental study showed that Ascomycota in rhizosphere soil of rice fields was the absolute dominant group under different cultivation methods, compared with N0, Ascomycota of FP, HYEY, and SHY was increased by 42.80%, 68.31%, and 5.27%, respectively. This suggested that high nutrient input (SHY) did not increase Ascomycota abundance, but inhibited. This was because the promoting effect of alkali-hydrolyzed nitrogen and organic matter on Ascomycota did not compensate for the inhibition of available phosphorus, available potassium, and pH on Ascomycota. In addition, Ascomycota had strong drought resistance and could survive in soil moisture-limited environments [57]. Most Basidiomycota fungi were rotten or parasitic fungi, which were also important decompositions in the soil [58], and liked the soil conditions with better ventilation [59]. This study showed that higher abundance of Basidiomycota in SHY, which may be due to the high level of organic matter in SHY soils. HYEY, SHY changed the growth environment of the fungal community, increased the soil available nutrients and organic matter content more suitable for the growth of rotten fungi, but the HYEY soil environment was more suited to the Ascomycota fungus, while SHY was more inclined to the Basidiomycota. This also suggested that a different fungal community adaptation to different environments led to fungal community changes under N0, FP, HYEY, and SHY.
5. Conclusions
Different cultivation practices altered soil bacterial diversity and fungal communities. Compared with bacterial community diversity, the cultivation method significantly affected fungal community diversity. The practice HYEY increased the richness and diversity of bacterial communities, but significantly reduced the richness and diversity of fungal communities. The richness and diversity of bacterial communities under SHY were decreased but fungal community abundance was increased for N0. Both, HYEY and SHY practices improved soil nutrient content, especially alkali-hydrolyzed nitrogen and organic matter. Under different cultivation methods, bacteria took Proteobacteria as the absolute dominant group, Chloroflexi, Bacteroidetes, and Acidobacteria as the dominant group; fungi took Ascomycota as the absolute dominant group. Available potassium and organic matter were the main environmental factors that affected the species composition at the phylum level of the bacterial community. Soil-available potassium was the major environmental factor that affected species composition at the phylum level of the fungal community.
Author Contributions
Conceptualization, C.Z.; Data curation, Y.H.; Formal analysis, Y.G. and Y.Z.; Funding acquisition, X.G.; Investigation, L.X.; Project administration, H.L.; Supervision, X.G. and Y.L.; Writing—original draft, X.G.; Writing—review and editing, Y.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Opening Foundation of Collaborative Innovation Center for Genetic Improvement and High Quality and Efficiency Production of Northeast Japonica Rice, grant number KF2022-02 and Heilongjiang Provincial Higher Education Teaching Reform Project, grant number SJGY20220466.
Data Availability Statement
Data available from the author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vogel, T.M.; Simonet, P.; Jansson, J.K.; Hirsch, P.R.; Tiedje, J.M.; van Elsas, J.D.; Bailey, M.J.; Nalin, R.; Philippot, L. TerraGenome: a consortium for the sequencing of a soil metagenome. Nature Reviews Microbiology. 2009, 7, 252. [Google Scholar] [CrossRef]
- Zhang, W.W.; Wang, C.; Xue, R.; Wang, L.J. Effects of salinity on the soil microbial community and soil fertility. Journal of Integrative Agriculture. 2019, 18, 1360–1368. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; Gormley, N.; Gilbert, J.A.; Smith, G.; Knight, R. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. The ISME Journal. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.M.; Liu, S.R.; Wang, H.; Hu, Z.D.; Li, Z.G.; You, Y.M. Changes of soil microbial biomass carbon and community composition through mixing nitrogen-fixing species with Eucalyptus urophylla in subtropical China. Soil Biology and Biochemistry. 2014, 73, 42–48. [Google Scholar] [CrossRef]
- Chen, X.F.; Li, Z.P.; Liu, M.; Jiang, C.Y.; Che, Y.P. Microbial community and functional diversity associated with different aggregate fractions of a paddy soil fertilized with organic manure and/or NPK fertilizer for 20years. Journal of Soils and Sediments. 2015, 15, 292–301. [Google Scholar] [CrossRef]
- Xia, X.; Shi, K.; Huang, Q.R.; Li, D.M.; Liu, M.Q.; Li, H.X.; Hu, F.; Jiao, J.G. The changes of microbial community structure in red paddy soil under long-term fertilization. Acta Pedologica Sinica. 2015, 52, 697–705. (in Chinese). [Google Scholar]
- Jorquera, M.A.; Martínez, O.A.; Marileo, L.G.; Acuña, J.J.; Saggar, S.; Mora, M.L. Effect of nitrogen and phosphorus fertilization on the composition of rhizobacterial communities of two Chilean Andisol pastures. World Journal of Microbiology and Biotechnology. 2014, 30, 99–107. [Google Scholar] [CrossRef]
- Chen, C.L.; Wu, M.N.; Wei, W.X. Effect of long-term application of nitrogen fertilizer on the diversity of nitrifying genes (amoA and hao) in paddy soil. Environment Science. 2011, 32, 1489–1496. (in Chinese). [Google Scholar]
- Grover, J.P. Predation, competition, and nutrient recycling: a stoichiometric approach with multiple nutrients. Journal of Theoretical Biology, 2004, 229, 31–43. [Google Scholar] [CrossRef]
- Su, J.Q.; Ding, L.J.; Xue, K.; Yao, H.Y.; Quensen, J.; Bai, S.J.; Wei, W.X.; Wu, J.S.; Zhou, J.Z.; Tiedje, J.M.; Zhu, Y.G. Long-term balanced fertilization increases the soil microbial functional diversity in a phosphorus-limited paddy soil. Molecular Ecology. 2015, 24, 136–150. [Google Scholar] [CrossRef]
- Meisner, A.; Jacquiod, S.; Snoek, B.L.; Ten, H.F.C.; van der Putten, W.H. Drought legacy effects on the composition of soil fungal and prokaryote communities. Frontiers in Microbiology. 2018, 9, 294. [Google Scholar] [CrossRef] [PubMed]
- Manzoni, S.; Schimel, J.P.; Porporato, A. Responses of soil microbial communities to water stress: results from a meta-analysis. Ecology. 2012, 93, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Dangi, S.R.; Zhang, H.H.; Wang, D.; Gerik, J.; Hanson, B.D. Soil microbial community composition in a peach orchard under different irrigation methods and postharvest deficit irrigation. Soil Science. 2016, 181, 208–215. [Google Scholar] [CrossRef]
- Li, Q.; Li, C.; Fang, B.H.; Liu, Y.; Kuang, W.; Chen, K.L.; Zhou, X.Q.; Wang, W.X.; Zhang, Y.Z. Effect of different irrigation modes in paddy soil on microbial community functional diversity. Genomics and Applied Biology. 2020, 39, 1632–1641. (in Chinese). [Google Scholar]
- Gil, S.V.; Meriles, J.; Conforto, C.; Basanta, M.; Radl, V.; Hagn, A.; Schloter, M.; March, G.J. Response of soil microbial communities to different management practices in surface soils of a soybean agroecosystem in Argentina. European Journal of Soil Biology. 2010, 47, 55–60. [Google Scholar]
- Tang, H.M.; Xiao, X.P.; Li, C.; Cheng, K.K.; Shi, L.H.; Pan, X.C.; Li, W.Y.; Wen, L.; Wang, K. Tillage and crop residue incorporation effects on soil bacterial diversity in the double-cropping paddy field of southern China. Archives of Agronomy and Soil Science. 2021, 67, 435–446. [Google Scholar] [CrossRef]
- Lin, X.B.; Shi, H.; Wu, L.; Cheng, Y.H.; Cai, S.; Huang, S.S.; He, S.L.; Huang, Q.R.; Zhang, K. Effects of cultivation methods on soil microbial community structure and diversity in red paddy. Ecology and Environmental Sciences. 2020, 29, 2206–2214. (in Chinese). [Google Scholar]
- Chen, Z.G.; Dai, Y.Y. Analysis report of heilongjiang rice market in 2019. Heilongjiang Liangshi. 2020, 5, 23–27. (in Chinese). [Google Scholar]
- Lv, Y.D.; Yue Hu, Y.; Jiang, H.F.; Lan, Y.C.; Wang, H.Y.; Xu, L.Q.; Yin, D.W.; Zheng, G.P.; Guo, X. H.Agronomic practices affect rice yield and nitrogen, phosphorus, and potassium accumulation, allocation and translocation. Agronomy journal. 2020, 112, 1238–1249. [Google Scholar] [CrossRef]
- Bao, S.D. Soil Agrochemical Analysis (Third Edition); (in Chinese). China Agricultural Press: Beijing, 2005. [Google Scholar]
- Grice, E.A.; Kong, H.H.; Conlan, S.; Deming, C.B.; Davis, J.; Young, A.C.; Bouffard, G.G.; Blakesley, R.W.; Murray, P.R.; Green, E.D.; Turner, M.L.; Segre, J.A. Topographical and temporal diversity of the human skin microbiome. Science. 2009, 324, 1190–1192. [Google Scholar] [CrossRef]
- Amadou, A.; Song, A.; Tang, Z.X.; Li, Y.L.; Wang, E.Z.; Lu, Y.Q.; Liu, X.D.; Yi, K.K.; Zhang, B.; .Fan, F.L. The effects of organic and mineral fertilization on soil enzyme activities and bacterial community in the below-and above-ground parts of wheat. Agronomy. 2020, 10, 1452. [Google Scholar] [CrossRef]
- Zhong, W.H.; Gu, T.; Wang, W.; Zhang, B.; Lin, X.G.; Huang, Q.R.; Shen, W.S. The effects of mineral fertilizer and organic manure on soil microbial community and diversity. Plant and Soil. 2010, 326, 523. [Google Scholar] [CrossRef]
- Zeng, J.; Liu, X.J.; Song, L.; Lin, X.G.; Zhang, H.Y.; Shen, C.C.; Chu, H.Y. Nitrogen fertilization directly affects soil bacterial diversity and indirectly affects bacterial community composition. Soil Biology and Biochemistry. 2016, 92, 41–49. [Google Scholar] [CrossRef]
- Ling, N.; Chen, D.M.; Guo, H.; Wei, J.X.; Bai, Y.F.; Shen, Q.R.; Hu, S.J. Differential responses of soil bacterial communities to long-term N and P inputs in a semi-arid steppe. Geoderma. 2017, 292, 25–33. [Google Scholar] [CrossRef]
- Dai, Z.M.; Su, W.Q.; Chen, H.H.; Barberán, A.; Zhao, H.C.; Yu, M.J.; Yu, L.; Brookes, P.C.; Schadt, C.W.; Chang, S.X.; Xu, J.M. Long-term nitrogen fertilization decreases bacterial diversity and favors the growth of Actinobacteria and Proteobacteria in agro-ecosystems across the globe. Global Change Biology. 2018, 24, 3452–3461. [Google Scholar] [CrossRef]
- Evans, S.E.; Wallenstein, M.D. Soil microbial community response to drying and rewetting stress: does historical precipitation regime matter? Biogeochemistry. 2012, 109, 101–116. [Google Scholar] [CrossRef]
- Hamer, U.; Unger, M.; Makeschin, F. Impact of air-drying and rewetting on PLFA profiles of soil microbial communities. Journal of Plant Nutrition and Soil Science. 2007, 170, 259–264. [Google Scholar] [CrossRef]
- Zhuang, L.J. Effects of wetting and drying conditions on the abundance of anaerobic ammonium oxidation bacteria and ammonia oxidizing archaea in soil. Shanghai: Donghua University, 2017. (in Chinese).
- Zhao, J.; Ni, T.; Li, Y.; Xiong, W.; Ran, W.; Shen, B.; Shen, Q.R.; Zhang, R.F. Responses of bacterial communities in arable soils in a rice-wheat cropping system to different fertilizer regimes and sampling times. PloS ONE. 2014, 9, e85301. [Google Scholar] [CrossRef]
- Ahn, J.H.; Song, J.; Kim, B.Y.; Kim, M.S.; Joa, J.H.; Weon, H.Y. Characterization of the bacterial and archaeal communities in rice field soils subjected to long-term fertilization practices. Journal of Microbiology. 2012, 50, 754–765. [Google Scholar] [CrossRef]
- Chen, D.M.; Chen, X.M.; Liang, Y.J.; Huo, X.J.; Zhang, C.H.; Duan, Y.Q.; Yang, Y.H.; Yuan, L. Influence of crop rotation on soil nutrients,microbial activities and bacterial community structures. Acta Prataculturae Sinica 2015, 24, 56–65. (in Chinese). [Google Scholar]
- Eldridge, D.J.; Delgado-Baquerizo, M.; Travers, S.K.; Val, J. , Oliver, L.; Hamonts, K.; Singh, B.K. Competition drives the response of soil microbial diversity to increased grazing by vertebrate herbivores. Ecology. 2017, 98, 1922–1931. [Google Scholar] [CrossRef]
- Chen, D.M.; Yuan, L.; Huang, J.G.; Ji, J.H.; Hou, H.Q.; Liu, Y.R. Influence of long-term fertilizations on nutrients and fungal communities in typical paddy soil of south China. Acta Agronomica Sinica. 2017, 43, 286–295 (in Chinese). (in Chinese). [Google Scholar] [CrossRef]
- Ji, L.F.; Ni, K.; Ma, L.F.; Chen, Z.J.; Zhao, Y.Y.; Ruan, J.Y.; Guo, S.W. Effect of different fertilizer regimes on the fungal community of acidic tea-garden soil. Acta Ecologica Sinica. 2018, 38, 8158–8166. (in Chinese). [Google Scholar]
- Liu, H.; Pan, F.J.; Han, X.Z.; Song, F.B.; Zhang, Z.M.; Yan, J.; Xu, Y.L. A comprehensive analysis of the response of the fungal community structure to long-term continuous cropping in three typical upland crops. Journal of Integrative Agriculture. 2020, 19, 866–880. [Google Scholar] [CrossRef]
- Li, X.G.; Ding, C.F.; Hua, K.; Zhang, T.L.; Zhang, Y.N.; Zhao, L.; Yang, Y.R.; Liu, J.G.; Wang, X.X. Soil sickness of peanuts is attributable to modifications in soil microbes induced by peanut root exudates rather than to direct allelopathy. Soil Biology and Biochemistry. 2014, 78, 149–159. [Google Scholar] [CrossRef]
- Gao, R.; Lv, J.L. Study on the enzyme activities and fertility change of soils by a long-term located utilization of different fertilizers. Chinese Journal of Eco-Agriculture. 2005, 13, 143–145. (in Chinese). [Google Scholar]
- de Vries, F.T.; Liiri, M.E.; Bjørnlund, L.; Bowker, M.A.; Christensen, S.; Setälä, H.M.; Bardgett, R.D. Land use alters the resistance and resilience of soil food webs to drought. Nature Climate Change. 2012, 2, 276–280. [Google Scholar] [CrossRef]
- Barnard, R.L.; Osborne, C.A.; Firestone, M.K. Responses of soil bacterial and fungal communities to extreme desiccation and rewetting. The ISME Journal. 2013, 7, 2229–2241. [Google Scholar] [CrossRef]
- Zhu, Y.Z.; Li, Y.Y.; Han, J.G.; Yao, H.Y. Effects of changes in water status on soil microbes and their response mechanism: A review. Chinese Journal of Applied Ecology. 2019, 30, 4323–4332. (in Chinese). [Google Scholar]
- Bapiri, A.; Bååth, E.; Rousk, J. Drying-rewetting cycles affect fungal and bacterial growth differently in an arable soil. Microbial Ecology. 2010, 60, 419–428. [Google Scholar] [CrossRef]
- Jenkins, S.N. , Rushton, S.P., Lanyon, C.V., Whiteley, A.S., Waite, L.S., Brookes, P.C., Kemmitt, S.; Evershed, R.P.; O’Donnell, A.G. Taxon-specific responses of soil bacteria to the addition of low level C inputs. Soil Biology and Biochemistry. 2010, 42, 1624–1631. [Google Scholar] [CrossRef]
- Celine, L.; Dimitris, P.; Sean, M.; Bernard, O.; Steven, S.; Safiyh, T.; Donald, Z.; Daniel, V.D.L. Elevated atmospheric CO2 affects soil microbial diversity associated with trembling aspen. Environmental Microbiology. 2008, 10, 926–941. [Google Scholar]
- Chaudhry, V.; Rehman, A.; Mishra, A.; Chauhan, S.P.; Nautiyal, C.S. Changes in bacterial community structure of agricultural lnd due to long-term organic and chemical amendments. Microbial Ecology. 2012, 64, 450–460. [Google Scholar] [CrossRef]
- Rodrigues, J.L.M.; Pellizari, V.H.; Mueller, R.; Baek, K.; Jesus, E.D.C.; Paula, F.S.; Mirza, B.; Hamaoui, G.S.; Tsai, S.M.; Feigl, B.; Tiedje, J.M.; Bohannan, B.J.M.; Nüsslein, K. Conversion of the Amazon rainforest to agriculture results in biotic homogenization of soil bacterial communities. Proceedings of the National Academy of Sciences of the United States of America. 2013, 110, 988–993. [Google Scholar] [CrossRef]
- Freedman, Z.; Eisenlord, S.D.; Zak, D.R.; Xue, K.; He, Z.L.; Zhou, J.Z. Towards a molecular understanding of N cycling in northern hardwood forests under future rates of N deposition. Soil Biology and Biochemistry. 2013, 66, 130–138. [Google Scholar] [CrossRef]
- Li, Y.Z.; Song, D.P.; Liang, S.H.; Dang, P.F.; Qin, X.L.; Liao, Y.C.; Siddique, K.H.M. Effect of no-tillage on soil bacterial and fungal community diversity: A meta-analysis. Soil and Tillage Research. 2020, 204, 104721. [Google Scholar] [CrossRef]
- Li, X.; Zhang, H.H.; Sun, M.L.; Xu, N.; Sun, G.Y.; Zhao, M.C. Land use change from upland to paddy field in mollisols drives soil aggregation and associated microbial communities. Applied Soil Ecology. 2020, 146, 103351. [Google Scholar] [CrossRef]
- Podosokorskaya, O.A.; Bonch-Osmolovskaya, E.A.; Novikov, A.A.; Kolganova, T.V.; Kublanov, I.V. Ornatilinea apprima gen. nov. sp. nov. a cellulolytic representative of the class Anaerolineae. International Journal of Systematic and Evolutionary Microbiology. 2013, 63, 86–92. [Google Scholar] [CrossRef]
- Kragelund, C.; Levantesi, C.; Borger, A.; Thelen, K.; Eikelboom, D.; Tandoi, V.; Kong, Y.; van der Waarde, J.; Krooneman, J.; Rossetti, S.; Thomsen, T.R.; Nielsen, P.H. Identity, abundance and ecophysiology of filamentous Chloroflexi species present in activated sludge treatment plants. FEMS Microbiology Ecology. 2007, 59, 671–682. [Google Scholar] [CrossRef]
- Li, X.; Sun, M.L.; Zhang, H.H.; Xu, N.; Sun, G.Y. Use of mulberry-soybean intercropping in salt-alkali soil impacts the diversity of the soil bacterial community. Microbial Biotechnology. 2016, 9, 293–304. [Google Scholar] [CrossRef]
- Wang, D.; Zhao, Y.G.; Ma, R.; Yang, P.; Zhang, C.; Zhou, D.J.; Sun, F.J.; Zhang, F.H. Effects of microbial fertilizers on soil improvement and bacterial communities in saline-alkali soils of lycium barbarum. Journal of Agricultural Biotechnology. 2020, 28, 1499–1510. (in Chinese). [Google Scholar]
- Pascault, N.; Ranjard, L.; Kaisermann, A.; Bachar, D.; Christen, R.; Terrat, S.; Mathieu, O.; Lévêque, J.; Mougel, C.; Henault, C.; Lemanceau, P.; Péan, M.; Boiry, S.; Fontaine, S.; Maron, P.A. Stimulation of different functional groups of bacteria by various plant residues as a driver of soil priming effect. Ecosystems. 2013, 16, 810–822. [Google Scholar] [CrossRef]
- Lauber, C.L.; Strickland, M.S.; Bradford, M.A.; Fierer, N. The influence of soil properties on the structure of bacterial and fungal communities across land-use types. Soil Biology and Biochemistry. 2008, 40, 2407–2415. [Google Scholar] [CrossRef]
- Leff, J.W.; Jones, S.E.; Prober, S.M.; Barberán, A.; Borer, E.T.; Firn, J.L.; Harpole, W.S.; Hobbie, S.E.; Hofmockel, K.S.; Knops, J.M.H.; McCulley, R.L.; La, P.K.; Risch, A.C.; Seabloom, E.W.; Schütz, M.; Steenbock, C.; Stevens, C.J.; Fierer, N. Consistent responses of soil microbial communities to elevated nutrient inputs in grasslands across the globe. Proceedings of the National Academy of Sciences of the United States of America. 2015, 112, 10967–10972. [Google Scholar] [CrossRef]
- Yan, R.; Feng, W.; Wang, X.J. Composition and diversity of soil fungi community under different sand-fixing plants in the Hulun Buir Desert. Science of Soil and Water Conservation. 2021, 19, 60–68. (in Chinese). [Google Scholar]
- Yelle, D.J.; Ralph, J.; Lu, F.C.; Hammel, K.H. Evidence for cleavage of lignin by a brown rot basidiomycete. Environmental Microbiology. 2008, 10, 1844–1849. [Google Scholar] [CrossRef]
- Sui, X.; Zhang, R.T.; Xu, N.; Liu, Y.N.; Chai, C.R.; Wang, J.F.; Fu, X.L.; Zhong, H.X.; Ni, H.W. Fungal community structure of different degeneration deyeuxia angustifolia wetlands in Sanjiang plain. Environmental Science. 2016, 37, 3598–3605. (in Chinese). [Google Scholar]
Figure 1.
Venn graph showed the number of shared and unique bacterial (Figure 1a), fungal (Figure 1b) OTUs among different treatments. N0: no nitrogen application; FP: local farmer practice; HYEY: high-yield and high-efficiency practice; SHY: super-high-yield practice.
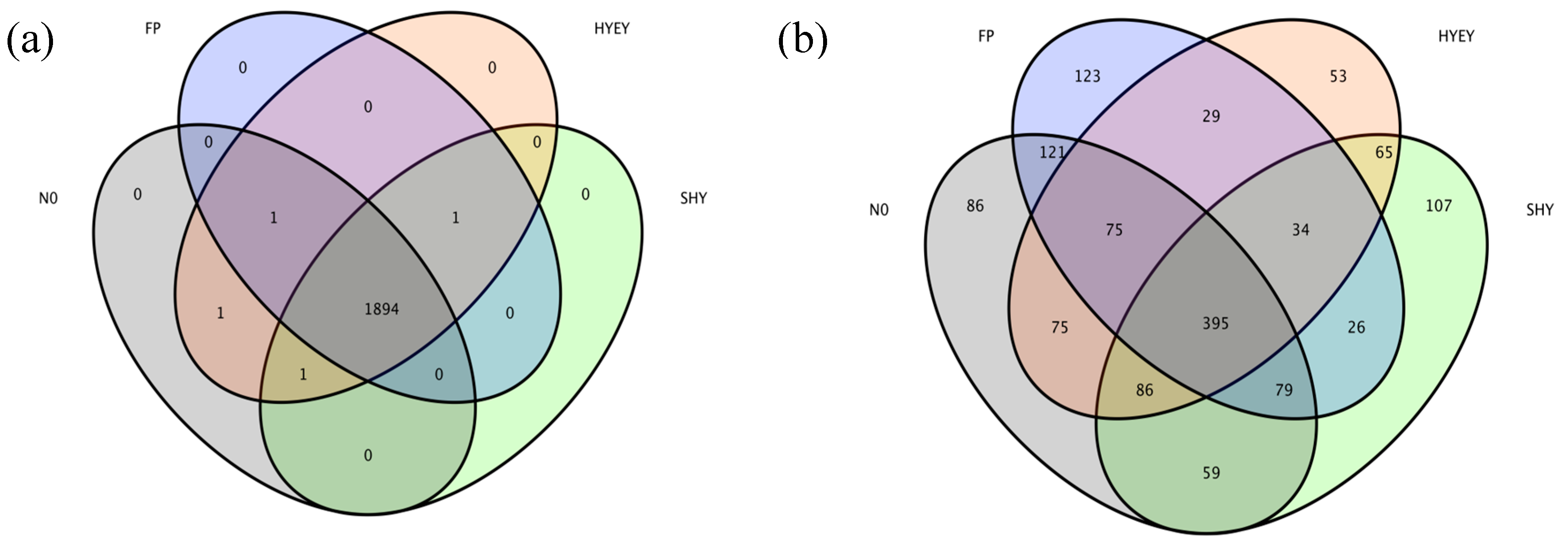
Figure 2.
Principal component analysis of the soil bacterial community (Figure 2a) and fungal community (Figure 2b) in the soil under different treatments. N0: no nitrogen application; FP: local farmer practice; HYEY: high-yield and high-efficiency practice; SHY: super-high-yield practice.
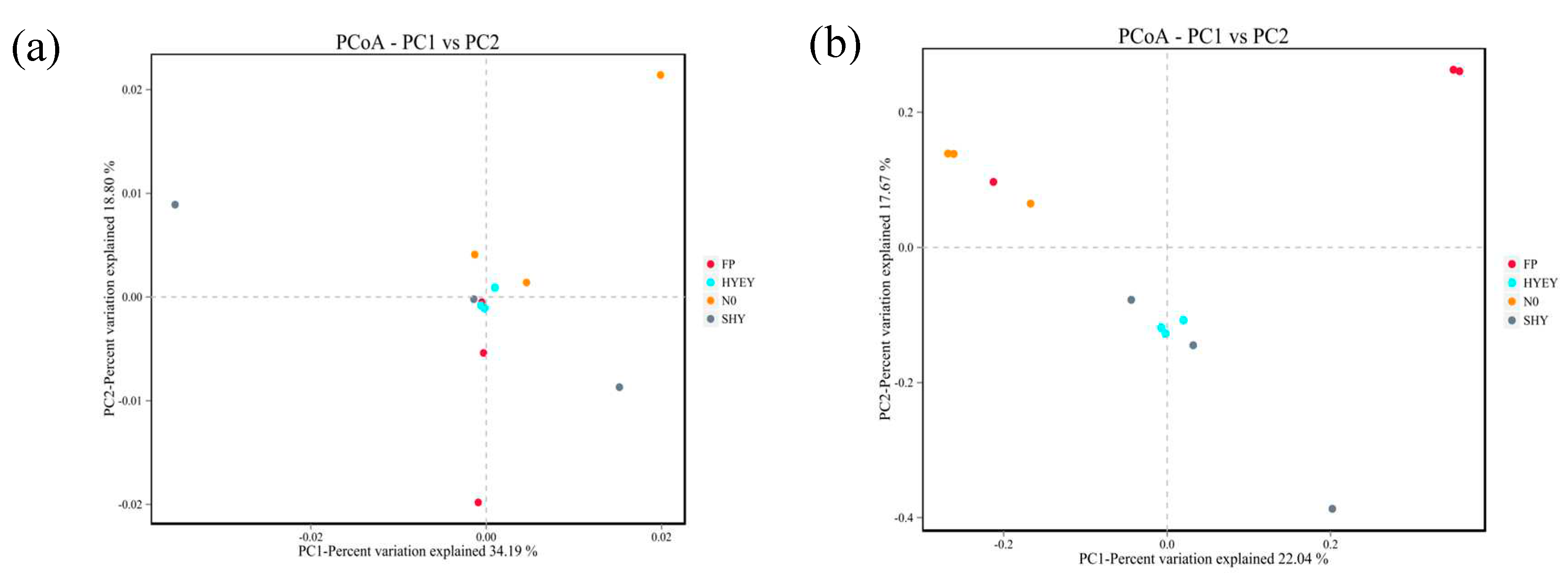
Figure 3.
Structure and composition of the bacterial (Figure 3a) and fungal (Figure 3b) community at the phylum level under different treatments. N0: no nitrogen application; FP: local farmer practice; HYEY: high-yield and high-efficiency practice; SHY: super-high-yield practice.
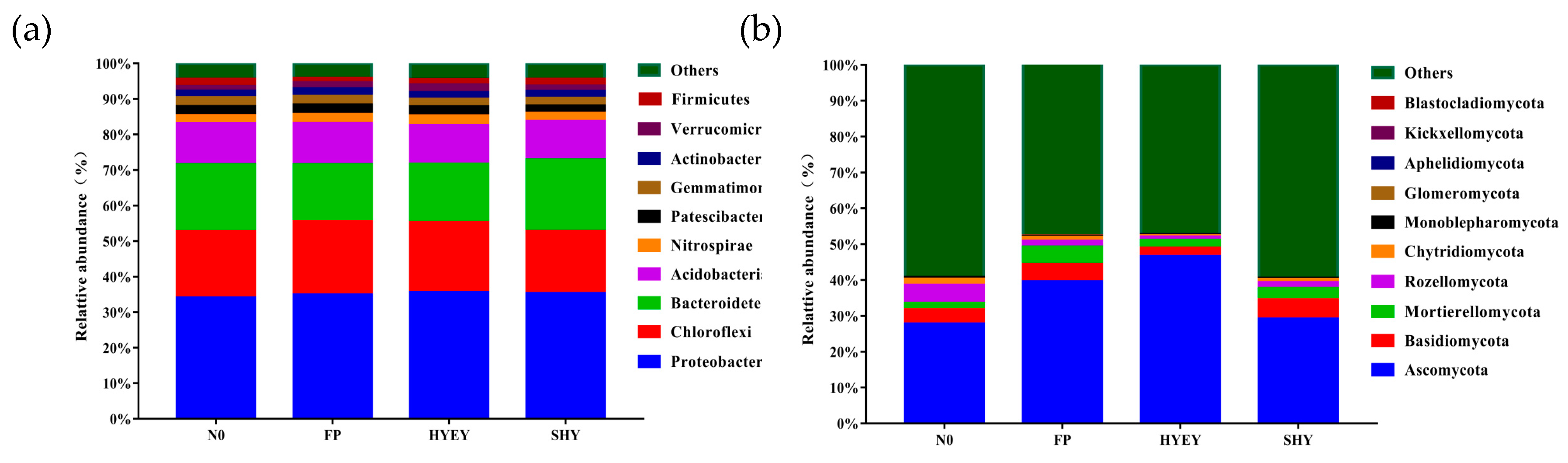
Figure 4.
Redundancy analysis of the relationship between soil microbial community (Figure 4a bacterial, Figure 4b fungal) distribution and soil properties under different treatments. N0: no nitrogen application; FP: local farmer practice; HYEY: high-yield and high-efficiency practice; SHY: super-high-yield practice. AHN, alkaline hydrolyzable nitrogen; AK, available potassium; AP, available phosphorus; SOM, soil organic matter.
Figure 4.
Redundancy analysis of the relationship between soil microbial community (Figure 4a bacterial, Figure 4b fungal) distribution and soil properties under different treatments. N0: no nitrogen application; FP: local farmer practice; HYEY: high-yield and high-efficiency practice; SHY: super-high-yield practice. AHN, alkaline hydrolyzable nitrogen; AK, available potassium; AP, available phosphorus; SOM, soil organic matter.
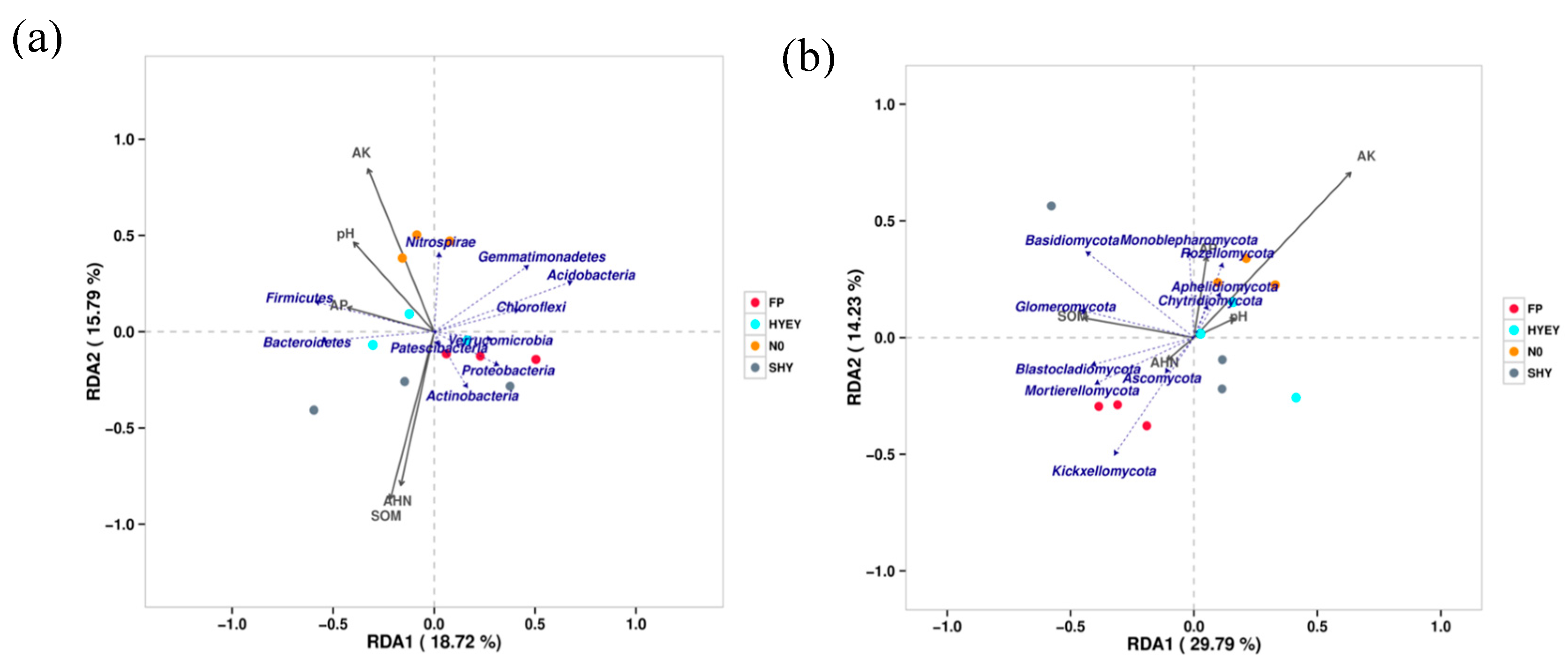

Table 1.
Fertilization application regime and cultivation practices under different cultivation modes.
Table 1.
Fertilization application regime and cultivation practices under different cultivation modes.
| Treatment | SRP | PP | IM | TNFA (ha−1) |
CF (ha−1) | |
|---|---|---|---|---|---|---|
| Nano-Silicon | Guifuji | |||||
| N0 | DBM | PW | SWF | 0 | - | - |
| FP | DBM | PW | SWF | 150 | - | - |
| HYEY | DBM | PW | AWD | 160 | 450 | - |
| SHY | PRM | AWNW | AWD | 180 | 450 | 15 |
SRP: seedling-raising patterns; PP: planting patters; IM: irrigation methods; AI: amount of irrigation; TNFA: total N fertilizer application; CF: compound fertilizer; N0: no nitrogen application; FP: local farmer practice; HYP: high-yield and high-efficiency practice; SHY: super-high-yield practice; DBM: dry bed method; PRM: potted raising method; PW: parallel row; AWNW: alternating wide and narrow rows; SWF: shallow water flood irrigation; AWD: alternate wetting and drying irrigation.
Table 2.
Effects of integrated cultivation practices on soil nutrients in paddy field.
| Treatments | AHN mg·kg−1 |
AP mg·kg−1 |
AK mg·kg−1 |
SOM g·kg−1 |
pH |
|---|---|---|---|---|---|
| N0 | 144.06d | 39.00a | 192.48a | 33.03c | 8.44a |
| FP | 161.29c | 36.23a | 112.83d | 36.12b | 8.40a |
| HYEY | 184.70b | 39.16a | 156.10b | 37.21b | 8.36a |
| SHY | 206.55a | 38.90a | 130.82c | 41.50a | 8.33a |
N0: no nitrogen application; FP: local farmer practice; HYEY: high-yield and high-efficiency practice; SHY: super-high-yield practice. AHN, alkali-hydrolyzed nitrogen; AP, available phosphorus; AK, available potassium; SOM, soil organic matter. Different letters indicate significant differences (p < 0.05).
Table 3.
Effects of integrated cultivation practices on the soil microbial diversity.
| Microbial species | Samples | ACE | Chao1 | Shannon | Simpson |
|---|---|---|---|---|---|
| Bacteria | N0 | 1885.72ab | 1890.88a | 6.72ab | 0.0026bc |
| FP | 1886.69ab | 1891.02a | 6.70ab | 0.0029ab | |
| HYEY | 1895.37a | 1897.22a | 6.78a | 0.0024c | |
| SHY | 1867.09b | 1871.71a | 6.63b | 0.0031a | |
| Fungus | N0 | 731.04a | 744.75a | 5.11ab | 0.0165b |
| FP | 511.07b | 520.88b | 5.59a | 0.0106b | |
| HYEY | 466.97c | 472.93c | 3.53c | 0.1563a | |
| SHY | 519.52b | 530.58b | 4.89b | 0.0202b |
N0: no nitrogen application; FP: local farmer practice; HYEY: high-yield and high-efficiency practice; SHY: super-high-yield practice. Different letters indicate significant differences (p < 0.05).
Table 4.
Correlation analysis of soil nutrients and microbial diversity.
| Microbial species | Parameters | AHN | AP | AK | SOM | pH |
|---|---|---|---|---|---|---|
| Bacteria | ACE | -0.221 | 0.002 | 0.145 | -0.583* | -0.080 |
| Chao1 | -0.226 | -0.074 | 0.153 | -0.583* | -0.113 | |
| Simpson | 0.36 | -0.169 | -0.538 | 0.545 | -0.16 | |
| Shannon | -0.256 | 0 | 0.317 | -0.423 | -0.082 | |
| Fungus | ACE | -0.661* | 0.066 | 0.743** | -0.633* | 0.39 |
| Chao1 | -0.651* | 0.068 | 0.732** | -0.623* | 0.369 | |
| Simpson | 0.258 | 0.114 | 0.153 | 0.032 | -0.122 | |
| Shannon | -0.353 | -0.15 | -0.166 | -0.122 | 0.154 |
AHN, alkali-hydrolyzed nitrogen; AP, available phosphorus; AK, available potassium; SOM, soil organic matter. *, ** Significant at the p = 0.05 and p = 0.01 probability level, respectively.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



