
Submitted:
25 August 2023
Posted:
29 August 2023
You are already at the latest version
Abstract
Abstract: Autophagy has a dual role in gliomagenesis in a microRNA-modulated environment. We investigated the potential relevance of autophagy in glioma development and survival by exploring the association of autophagy-associated genes and microRNAs in low- and high-grade gliomas. Real-time PCR (qPCR) was used to determine the expression of genes and microRNAs in 50 fresh glioma tissues while Formalin-fixed paraf-fin-embedded tissues of the same patients were used for immunohistochemistry. The Mann-Whitney U-test test, Spearman correlation test, and Kaplan-Meier survival analysis were performed to evaluate the expression, association, and overall survival in patients respectively. The expression of LC3, AKT, and miR-21 was increased in high-grade glioma compared to low-grade glioma while ULK2 expression was decreased in high-grade glioma. A strong positive correlation was observed for ULK2 with UVRAG, PTEN, miR-7, and miR-100, while the moderate correlation with mTOR, Beclin1, miR-30, miR-204, miR-374, miR-21 and miR-126 in low-grade glioma, while a moderate positive correlation between ULK2 and PI3K, PTEN, ULK1, VPS34, mTOR, Beclin1, UVRAG, AKT and miR-374, and between AKT and ULK1, VPS34, UVRAG, and miR-7 in high-grade gliomas. The low ULK2 and LC3 expression group was significantly associated with better overall survival in gliomas while miR-21 overexpression showed a poor prognosis in glioma patients. Therefore, miR-21, ULK2, and LC3 may serve as prognostic biomarkers for survival outcomes in glioblastoma.
Keywords:
Glioma
; Prognostic markers
; Autophagy
; Micro-RNA
; Overall Survival
1. Introduction
Gliomas are primary CNS tumors with significant molecular heterogeneity and a highly mutated genome [1]. Malignant glioma comprises 50% of primary brain tumors, with overall survival of 12-15 months despite surgical resection, chemotherapy, and radiation therapy. Despite substantial advances in glioma research and therapeutic modalities, the survival rates for glioma patients have not shown significant improvement, with recent highlighting the dysregulation (downregulation) of critical pathways, such as mutations in isocitrate dehydrogenase (IDH), p53, as key factors in glioma development and prognosis [2]. Dysfunctional cell death is a primary hallmark of malignant growth and underlies chemo-resistance gained by cancer cells [3]. Loss of apoptotic mechanisms has been implicated previously in gliomagenesis [4]. Autophagy, a caspase-independent process, mediates the degradation of cytoplasmic components under stress conditions, such nutrient deprivation, hypoxia, endoplasmic reticulum (ER stress), cancer therapy, and pathogenic infection via the formation of autophagosome [1,2]. Autophagy exerts a tumor-suppressive function during the initiation phase of cancer, whereas the evasion of autophagy during cancer progression and therapeutic interventions contributes to a tumor-promoting effect [3]. Furthermore, the PI3K/Akt/mTOR signaling cascade, which is an important component of the upstream autophagy pathway, is often found to be dysregulated in glioblastoma (GBM). The investigation of autophagy mechanisms represents significant focal points in the study of gliomagenesis and malignant transformation.
Non-coding RNAs, including microRNAs (miRNAs), have been known to regulate autophagy [4]. miRNAs are reportedly dysregulated in human cancers such as prostate, renal, colon, and glioblastoma, where the dysregulation is associated with initiation and progression of cancer, as well as resistance to chemo-radiotherapy [5,6]. miRNAs can function as both tumor suppressors and oncogenes, with single miRNAs regulating multiple targets and networks involved in tumor progression. Several studies have emphasized the importance of microRNAs in autophagy regulation and the significance of autophagy progression in various cancer stages and processing, while miRNAs can regulate autophagy-related genes [5,7]. However, the precise mechanism by which autophagy and miRNAs influence each other in the context of glioma progression and patient survival are still not completely understood. Elucidating these interactions will contribute to a better understanding of the underlying tumor biology and potentially help in identifying novel therapeutic strategies.
This study aims to determine and correlate the expression of autophagy markers, autophagy-associated signalling genes (PI3K/Akt/mTOR) and autophagy-associated microRNAs in low and high-grade gliomas.
2. Results
2.1. Clinical Characteristics of the study population
A total of 50 glioma patients were recruited in our study, out of which, 16 (32%) were females and 34 (68%) were males, with a median age of 38 years (range from 4-70). The most prevalent histopathology was as follows: glioblastoma 29 (58%), oligodendroglioma 15 (30%), and astrocytoma 6 (12%). Twenty-six (52%) patients had postoperative Karnofsky Performance Scale KPS scores of less than 80, while 22 (44%) of the patients had KPS scores above 80. Only 21 (42%) patients received adjuvant chemotherapy and radiotherapy, while 23 (46%) patients received either chemotherapy or radiotherapy unaided. The average life expectancy was 22.6 months, ranging from 4 to 35 months (Table 1). Moreover, IDH-1 mutation was found in 20 (40%) patients, while 80(60%) patients were labeled as IDH-1 wild type. The tumor suppressor gene p53 over-expression was found in 40 (80%) patients, while all tumor samples 50 exhibited ATRX retention.
Pearson chi-square correlation analysis showed that the prognostic marker Ki-67 was significantly correlated with age, histological type, radiotherapy, chemotherapy, tumor type, and tumor origin types (p<0.005). However, IDH-1, Ki-67, and p53 did not exhibit statistically significant association with age, histological type, radiotherapy, chemotherapy, tumor type, and tumor origin types (p > 0.05) (Table 2).
2.2. Expression of autophagy marker LC3-II in LGG and HGG samples
Out of 50 cases, 49 cases showed cytoplasmic LC3-II punctaes in various parts of the cells. The median Q score of LC3-II were 20 and 24 for LGG and HGG, respectively. the difference in cytoplasmic expression of LC3-II in LGG and HGG was not statistically significant (p=0.32) (Figure 1a and b). In the next step, the differential mRNA expression of LC3-II was examined in the LGG and HGG samples. The qPCR analysis revealed a statistically significant difference in LC3-II mRNA expression, with HGG samples exhibiting a 0.53 fold higher LC3-II expression as compared to LGG (p = 0.009) (Figure 1c).
2.3. Analysis of autophagy-associated genes in LGG and HGG samples
To further explore the differential expression of autophagy-associated genes in LGG and HGG groups, expression of seven genes related to the core autophagy activation pathway, namely ULK1 complexes (ULK1, ULK2), Beclin1/PI3K complex interacting proteins (Beclin1, UVRAG, Vps34), and ATG-8 ubiquitin-like conjugation system (LC3B) was investigated. No significant difference in expression of the ULK1 gene was observed between LGG and HGG samples (p=0.33), while analysis of ULK2 showed a significant 8.86 fold higher expression in LGG as compared to HGG (p=0.04). Other genes (VPS34, Beclin1, and UVRAG and mTOR) also did not exhibit significant difference (p>0.05) in expression between the two groups (Figure 2).
2.4. Analysis of PTEN/PI3K/AKT genes in LGG and HGG samples
Next, the differential expression of PTEN/PI3K/AKT genes was examined, which are component of the upstream autophagy pathway [9]. The mRNA expression of the AKT gene was found to be 0.5 fold higher (p=0.04) in HGG as compared to LGG samples. The expression of PI3K and PTEN, however, were comparable between the two groups (p=0.19) (Figure 3).
2.5. Analysis of differential expression of autophagy-associated miRNAs in LGG and HGG samples
Analysis of expression of autophagy-associated miRNAs showed the expression of miR-126 and miR-374 to be 0.35-fold (p=0.01) and 1.56-fold (p=0.01) higher in LGG as compared to HGG samples, while miR-21 expression was found to be 1.18 fold less in HGG as compared to LGG samples (p=0.000075). Expressions of other tested miRNA were comparable between the two groups (Figure 4) between the two groups.
2.6. Correlation of expression of microRNAs and other autophagy-associated genes in LGG and HGG samples
In LGG group, a significant strong positive correlation was observed between UKL2 and miR-7 (rs=0.85), ULK2 and miR-100 (rs=0.81). Similarly, a moderate positive correlation was observed between ULK2 and miR-30 (rs=0.56), ULK2 and miR-204 (rs=0.58), ULK2 and miR-374 (rs=0.68). Likewise, miR-21 exhibited a moderate correlation between miR-126 (rs=0.56, p<0.05). Similarly, a strong positive correlation was observed between ULK2 and PTEN (rs=0.92), ULK2 and UVRAG (rs=0.80) while a moderate correlation between ULK2 and mTOR (rs= 0.59) (Table 3).
In HGG group, a moderate positive correlation was observed between ULK2 and miR-7 (rs=0.40), ULK2 and miR-374 (rs=0.50). Likewise, AKT was moderately correlated with miR-7 (rs=0.66, p<0.01), AKT with miR-30 (rs=0.41) and AKT with miR-204 (rs=0.40). Also, miR-30 has a moderate correlation with miR-21 (rs=0.40). Similarly, AKT correlated positively with UVRAG (rs:0.69), VPS34 (rs:0.56), ULK1 (rs:0.59), ULK2 (rs:0.50), PTEN (rs:0.48), and PI3K (rs:0.48). ULK2 correlated with ULK1 (rs:0.53), PTEN (rs:0.63) PI3K (rs=0.49), VPS34 (rs=0.63), mTOR (rs=0.59), Beclin (rs=0.55), UVRAG (rs=0.66) and AKT (rs=0.50). A weak positive correlation of miR-21 was observed with AKT (rs:0.38) in high-grade glioma (Table 4).
2.7. Association of expression of autophagy-associated genes and microRNAs with patient’s survival and cancer prognosis
Considering that AKT, ULK2, LC3 and miR-21 expressions were significantly correlated with glioma grade, we hypothesized that these genes might affect the survival/cancer prognosis of the patients. To confirm this possibility, the patients were divided into high AKT, ULK2, LC3 and miR-21expression and low expression groups. Cox regression was performed on the two groups and high expression group was used as reference. The hazard ratios (HR) >1 and <1 was used as indicators of low and high hazard risk, respectively. We found AKT (HR: 2.829, p=0.03) and miR-21(HR: 2.637 p=0.03) to be independent prognostic factors for overall survival, while ULK2 (HR: 0.355, p=0.02) and LC3 (HR: 0.41, p:0.05) AKT (HR: -0.97, p=0.04) and miR-21(HR: -0.92, p=0.05) were found to be marginally associated with overall survival (Table 5).
In the next step, the above predictors (AKT, ULK2, LC3, miR-21) along with significantly expressed genes AKT, ULK2, LC3) and microRNAs miR-21, miR-216 and miR-374) were included for survival analysis. The results indicated that high expression of AKT was significantly associated with poor overall survival (log-rank: 0.025). Additionally, low PI3K, ULK2, and LC3 were found to be statistically correlated with poor overall survival (log-rank: 0.098, 0.024, 0.05, respectively) (Figure 5a). Furthermore, high miR-21 was significantly correlated with poor overall survival OS in glioma patients (log-rank: 0.033) (Figure 5b).
Finally, Kaplan-Meier plots showed that patients with high expression group of AKT and high Ki67 showed significant poor OS (log-rank: 0.012). Interestingly, patients with IDH-wild type showed a trend with poor OS with both low and high expression group with LC3 (log-rank: 0.066). Similarly, wild-type IDH1 and High Ki67 were significantly associated with poor OS in low ULK2 type tumors (log-rank: 0.022), (log-rank: 0.006) respectively. Similarly, high miR-21 expression also correlated with poor overall survival in IDH1 wildtype, High Ki67, and p53 mutant tumors. Close to significant correlations of low miR-126 were seen with poor OS in IDH1 wildtype and High Ki67 tumor patients (log-rank: 0.064) (log-rank: 0.002) respectively (Figure 6a & b).
3. Discussion
The current study was conducted to evaluate the association of autophagy-associated genes and microRNAs along with the clinical parameters and their significance in glioma patient’s outcome. The data showed that glioblastoma was the most common tumor followed by oligodendroglioma, and astrocytoma and the onset of glioma was more profound in ages below 40 years indicating the early risk for glioma development coinciding with a study conducted by Singh et al. [10]. Generally, both age group are susceptible to glioma but, data from our study showed that women are less likely to present glioma compared to men. This trend in data could be associated to gender discrepancy in developing countries where females are twice more likely to postpone their surgery due to lack of financial assistance, travelling and decision-making process [11,12].
Next, we investigated genetic alteration in glioma tumor using a panel of immunohistological markers including IDH1, ATRX, p53 and Ki-67 which are used routinely for initial diagnosis of glioma. The association of these common prognostic and diagnostic molecular markers were correlated with clinical parameters such as age, gender, tumor type and tumor recurrence and the results indicated a strong association with Ki-67 as reported by Stark et al. in recurrent glioma patients [13]. Subsequently, autophagy genes and microRNAs processed from fresh tissues were analyzed through RT-qPCR to determine the role of autophagy in increasing glioma grades.
Autophagy is an essential cellular process that helps maintain cellular homeostasis by degrading and recycling damaged or unnecessary cellular components [14]. While autophagy initially serves as a protective mechanism during metabolic stress, emerging research suggests that it can also play a role in promoting cancer and resistance to anticancer therapies. This dual role of autophagy highlights its complex and context dependent involvement in cancer biology, emphasizing the need for further investigation to better understand its precise mechanism [15]. As reported previously, LC3 is associated with the membrane of autophagosome involved in autophagy, and LC3 is widely used as an autophagy marker and can be tested using LC3 antibody instead of other methods in clinical tissue specimens recently. LC3 consists of three isoforms LC3-I, LC3-II and LC3-III and in immunohistochemistry LC3 shows three patterns, diffuse cytoplasmic, cytoplasmic and dot-like patterns, thus the functional LC3 in autophagy remains confusing [16]. So, LC3-II was selected in this study to measure the flux of autophagy and the result showed a high punctate expression of LC3-II in both grades of glioma with no statistical difference between glioma grades. The puncta assay can provide valuable information about autophagosome formation, it may not directly measure autophagic flux. Combining the puncta assay with other complementary methods, such as the LC3 turnover assay or substrate degradation assays, can provide a more comprehensive understanding of autophagic flux [17] . Thus we turn to RT-qPCR technique for LC3 quantification due to quantitative nature of assay and we found a high and statistically significant expression of LC3 transcript in high-grade glioma compared to low grade of glioma, which is in line with a study from China indicating the association of LC3-II with the advanced tumor stage [18]. Aoki et al. confirmed the detection of a strong expression of LC3 in the cytoplasm of 31 glioblastoma tissue in comparison to no LC3 staining in normal brain samples [19]. In the present study, a statistically significant downregulation was found in the expression of ULK2 in high glioma group as compared to low glioma group. The current study was supported by Shulka et al. [20] where the autophagy inducer gene ULK2 was hypermethylated and downregulated in GBM as compared to normal brain. These results are supported by studies which reported that that ULK1 and ULK2 are transcriptional targets of p53 necessary for sustained autophagy activity induced by DNA damage thus coinciding with our findings [20]. Thus, it is safe to assume that sustained autophagic potential in malignant high-grade tumors further promotes the surviving tumor cells towards disease progression.
Given the slow growth rate of low-grade gliomas compared to high grade, it is a reasonable assumption that nutrient stress is more in HHG than LGG [21] . Therefore, it is expected that mTOR activity is moderate and autophagic rate is higher in HHG than LGG. Our result for the increased autophagic activity is in line with the previous studies [22]. However, the contrast in mTOR expression could be due to several possibilities. First, from a technical standpoint, qPCR shows the gene expression at the transcriptional level while the actual protein may get altered at translation and post-translational level as well. It is possible that mTOR is modulated at the translational level in high-grade gliomas. Crabtree et al, also reported an inconclusive result of mTOR in their study [23].
Further, we evaluated the cell proliferative, autophagy upstream pathway, PI3K/AKT/mTOR. The AKT gene was significantly up regulated in HGG than in LGG samples indicating proliferation of tumor in the malignant stage. PTEN and PI3K remained unchanged when compared in both grades, and PTEN had a low expression in both grades of gliomas. This observation is supported by a study indicating essential role of PI3K/AKT signaling pathway in development and progression of glioma [24].
microRNAs (miRNA or miR) which modulate gene expression by targeting the 3’ UTR of the mRNA exert regulatory control over gene expression by modulating key pathways involved in various biological processes. In the present study, we also studied autophagy-associated miRs (miR-7, miR-21, miR-30, miR-100, miR-204, miR-126 and miR-374) and found a significant upregulation of autophagy-associated microRNAs miR-21, miR-126 and miR-374 in high-grade glioma tissues compared to low-grade glioma. This trend of up-regulation for most of the studied miRNAs is in line with the observation reported by Xu et al. [25]. miR-21 was found to be significant in this study and from the literature it is known to be one of the first microRNAs found in the human genome and the most important oncomir known to be up-regulated in all types of human cancer including (glioblastoma, breast, lung, esophageal, hepatocellular, prostate and lung) [26]. Oncomir miR-21 is consistently over-expressed in glioma and research has demonstrated that it engages in a wide variety of biological pathways, promoting tumor cell survival and invasiveness [27]. Furthermore, it has been implicated in resistance to treatment, both against chemotherapy and radiotherapy [28]. Yang et al. also reported a significant difference in miR-21 levels, measured with qRT-PCR, between LGG and GBM samples [29].
This study found up-regulation of miR-374 along with its target gene AKT in low grade glioma. The findings of the current study were also corroborated by Dong et al. demonstrated who showed that miR-374 level was significantly repressed in high-grade vs. low-grade gliomas which is consistent with the results of the present study [30]. Similarly, Zhang, et al. [31] , reported that miR-374 activates the PI3K/Akt pathway in myocardial damage thus autophagy reveals a novel function for miRNA in a critical cellular event with significant impacts on cancer development, progression, and treatment, and in other diseases [32].
On the contrary, the tumor suppressor miR-126 was found to be down-regulated within our malignant glioma samples. A previous study has shown an anti-tumor effect of miR-126 IN glioma thus supporting our result. The same study findings also suggested that a potential inverse relationship between miR-126, PI3K and AKT occurs in in glioma cell lines and that miR-126 acts as a tumor suppressor by targeting, among others, AKT and PI3K [33,34].
In the low-grade glioma group, we found a significant strong positive correlation between the expression of ULK2 and miR-7 and a moderate positive correlation with other autophagy gene panels except LC3. The correlation between expressions of miR-7, PI3K/AKT, mTOR and ULK2 were reported by numerous researchers in multiple cancers [35,36,37]. However, to the best of our knowledge, none of the researchers have reported the correlation based on grades of glioma. A study reported that miR-7 impairs autophagy to suppress pancreatic cancer progression by indirectly limiting LC3 dots. miR-7 could repress autophagy through up-regulation of LKB1-AMPK-mTOR signaling, and directly targeting the stages of autophagy induction and vesicle elongation to reduce the supply of intracellular glucose to glycolysis metabolism whereas, in our study we found a negative correlation between miR-7/LC3 but it was not significant [36]. One of the first studies on autophagy induction via mir-21 targeting LC3-II was reported in leukemia patients [38]. Another study reported that miR-204 was regulated by the tumor suppressor von Hippel–Lindau (VHL) Protein in renal carcinoma, directly controlled the level of LC3 and determined the level of autophagy.
The intercorrelation among autophagy genes was more profound with increasing glioma grade in the present study and could show evidence for possible pathways of gliomagenesis through the nexus of microRNA and autophagy markers. ULK2 and AKT continues to correlate significantly with most autophagy markers in HGG, showcasing its key role as a downstream target of the PI3k-Akt signaling pathway [39]. There is significant evidence for the PI3K/AKT/mTOR pathway as a key component in GBM progression, with evidence supporting the development of inhibitors for mTOR [40]. So far PTEN deficiency has been linked to promoting tumors indirectly through dysregulation of PI3K/AKT [41] and low PTEN expression correlates significantly with poor survival in our cohort.
We also investigated the association of autophagy associated mRNA and miRs with OS of patients and prognostic significance. The result demonstrated that the expression level of AKT and miR-21 were significantly associated with OS, with patients exhibiting higher expression levels with poor prognostic outcome. It has been reported that PI3K/AKT pathway is involved in the glioblastoma progression [42]. High LC3 expression has been correlated with better overall survival in our current study as well as in previous study reported by Aoki et al [43].
Multiple studies have provided evidence linking the expression level of miR-21 to prognostic outcomes, suggesting its potential as a prognostic marker in various cancers, including prostate cancer [44] and head and neck squamous cell carcinoma [45]. Based on these early discoveries, a prognostic predictive high expression for miR-21 in glioma has been confirmed in this study and many previous studies have proven miR-21 as an independent prognostic marker [46]. Therefore, autophagy-related genes and microRNA may serve as a potential prognostic marker for glioma.
We also investigated their impact on survival when subgrouping tumors according to well-established molecular markers of glioma i.e., IDH, p53, Ki-67. ULK2 expression appears to be a significant predictor across all subgroups, suggesting its importance as a potential biomarker. A study indicated a promotor hypermethylation of ULK2 in all grades of glioma suggesting that inhibition of autophagy by ULK1/ULK2 is essential for glioma development [20]. In our study, we found that the high expression level of miR-21 was significantly associated with poor prognosis when correlated with IDH wildtype, High Ki67 and with mutant. This suggests patients with higher miR-21 expression have shorter OS in line with a study reported by Nieland et al. [47] Similarly low expression of tumor suppressor miR-126 with high Ki67 predicted a worst prognostic effect in the current glioma patients. miR-126 induced autophagy has been well-established as an inhibitory mechanism against glioma survival, and its promotor methylation may play a role in tumor development [20,48,49].
High expression of autophagy genes such as LC3 in high-grade glioma compared to low-grade glioma shows the association of autophagy with tumor progression, with prognostic significance. MicroRNAs such as miR-126, miR-374 and miR-21 and their correlating targets, particularly the tumor promoting pathway gene AKT, may be ideal targets for inhibiting glioma formation. This will be an initial step in understanding the oncogenic role of autophagy in glioma tissues from the population. Further, cell culture models overexpressing or suppressing critical microRNAs will lead to better functional understanding of the effect on targeted genes and downstream pathways.
4. Methods
4.1. Patient selection
For this study, 50 fresh glioma tumor samples, divided as low grade glioma (LGG; Grade II; N=15) and high grade glioma (HGG; Grade III, Grade IV; N=35) and 50 matching formalin-fixed paraffin-embedded (FFPE) blocks from patients, who had a confirmed diagnosis of glioma on histology and CT (Computerized Tomography)/MRI (Magnetic Resonance Imaging), and underwent surgery at the Aga Khan University Hospital, Karachi between 2019-2022 were used. Patient details, such as therapeutic data (chemotherapy, radiotherapy), histopathology, tumor grade, and molecular marker (Ki-67) were retrieved from hospital medical records. Furthermore, survival data was obtained from their follow-up records. Patient data and samples were treated according to ethical and legal standards adopted by the Declaration of Helsinki 2013. This study was approved by the ethical review committee of Aga Khan University (AKU-ERC # 2019 1704-4925). All eligible participants involved in the study provided written informed consent for the utilization of their tissue samples.
4.2. Histopathology and immunohistochemistry
Histological sections of tumor FFPE tissues were examined and graded by a histopathologist at Aga Khan University. In each FFPE sample, IHC-based expression of ATRX, p53, IDH1-R132H, LC3-II was examined. Briefly, after deparaffinization in xylene and alcohol, antigen retrieval was conducted using Tris-ethylenediaminetetraacetic acid (EDTA; pH 9.0) in a microwave oven for a duration of 20 minutes, followed by cooling at room temperature. Following a 10-minute rinsing and blocking step with peroxidase, mouse monoclonal antibodies targeting IDH-1 R132H (clone: H09, Dianova), ATRX (clone: BSB-108), and p53 (DO7, Cell Marque), as well as a rabbit polyclonal antibody against LC3-II (ab51520), were administered. The incubations were conducted overnight in a humidity chamber at a temperature of 4ºC. Subsequently, the samples were washed three time using phosphate buffer saline (PBS) buffer with a pH of 7.6, with each wash lasting for a duration of 2 minutes. After that, biotinylated secondary antibodies were applied, followed by a washing step using a PBS solution. Subsequently, the samples were incubated with horseradish peroxidase (HRP) for a duration of 30 minutes, followed by another round of washing as previously described. The chromogen 3,3-diaminobenzidine (DAB) was applied for 1 minute as per the manufacturer’s protocol. The slides underwent counterstaining with hematoxylin, followed by dehydration in ethanol and subsequent clearing with xylene. Negative controls were prepared by substituting the primary antibody with PBS), while Brain tumor samples previously characterized for overexpression of IHC markers were used as positive controls. For the IHC assessment, all prepared slides were examined using Olympus BX43 light microscope at 20X objective magnification. Photographs were obtained from PC-driven digital camera (Olympus) and analyzed using CAPTURE infinity imaging solution tool. IDH-1, ATRX and p53 were scored as positive or negative. LC3-II punctae were quantified using four-tiered scale: score 1, punctae ≤ 10 per cells; score 2, punctae 11-20 per cells; score 3, punctae >20 per cells [6].
4.3. RNA extraction, DNase Treatment of RNA, and CDNA synthesis
Total RNA was extracted from glioma tissue using TRIzol®-chloroform (Invitrogen; Thermo Fisher Scientific, Inc; Cat # 15596018) method, following manufacturer’s instructions. RNA concentration and purity was assessed using Nanodrop® 2000c Spectrophotometer (Thermo Scientific, Waltham, MA, USA). The RNA samples were stored at -80 °C for further use. Prior to cDNA conversion, each RNA sample was treated with DNase-I (RNase-free) 1U/1μl (Thermo Fisher Scientific, Cat. No. EN0521). For this, 1μg of the total RNA template was pooled in a 200 μl tube with 1ul of DNase-I, a 10X reaction buffer comprising MgCl2, and a nuclease-free water for a final volume of up to 10 μl. The reaction was incubated at 37 °C for 30 minutes followed by addition of 1μl of 50 Mm EDTA, and incubation at 65 °C for 10 minutes. The treated RNA was stored at -80 °C for further use in the cDNA reaction. The RNA samples were converted to cDNA using RevertAid first strand cDNA synthesis kit (Thermo Fisher Scientific, Cat. No. K1612). In the first step, 1 μl of treated RNA was mixed with 1 μl of 10 μM Random primers, and 10 ul of nuclease-free water in 200 μl tube. The tube was incubated for 5 minutes at 65 °C, briefly centrifuged and incubated on ice for 1 minute. The reaction mixture was further supplied with 4 μl of 5X reaction buffer, 1μl of Ribonuclease inhibitor, 2 μl of 10 μM dNTPs mix each and 1 μl of RevertAid reverse transcriptase enzyme and incubated at following conditions: 50 °C for 5 minutes, then 64 °C for 60 minutes, and finally 70 °C, for 5 minutes. Additionally, the prepared cDNA was diluted by the addition of 180 µl nuclease-free water to make up the reaction volume 200 μl. The prepared cDNA samples were stored at -20°C until further use.
4.4. Analysis of gene expression using quantitative real-time PCR (qPCR)
Expression of autophagy genes, LC3, ULK1/2, UVRAG, Beclin1, UVRAG, and autophagy-associated signalling pathway genes, PI3K, AKT, mTOR, PTEN and autophagy-related microRNAs (miR-7, miR-21, miR-30, miR-100, miR-126 and miR-374) were determined through the quantitative real-time PCR (qPCR) using QIAGEN’S Rotor-Gene Q machine. A 10 μl reaction mixture was set by combining 2 μl of cDNA template with 5 μl of PowerUp™ SYBR™ Green Master Mix (Thermo Fisher Scientific, Cat. No. A25742), 1 μl forward and reverse primers (10 μM) each (Eurofins, USA), (Appendix A) and 2 μl of nuclease free water. The above reactions were subjected to the following thermal cycling conditions; initial hold for 2 mins at 50°C, another hold for 2 minutes at 95°C followed by 40 cycles for denaturation for 15 sec at 95°C, annealing for 60 sec at 60°C. A melt curve analysis was set up between 50°C to 90°C with an increment of 1°C at each step to plot the specificity of the products. Each reaction was run in duplicate with a non-template and non-primer as an experimental control. A β-Actin gene and small nuclear RNA-U6 were used as an endogenous control. A relative fold change in gene and microRNA expression in each sample was calculated using Livak 2-∆∆Ct method [8].
4.5. Statistical analysis
Shapiro-walk with Maan-Whitney tests were applied to determine the difference in gene expression between in the LGG and HGG groups. In the next step, we applied Spearman’s rank correlation coefficient test to correlate expression of different genes in LGG and HGG groups. Similarly, Pearson Chi-square was applied for the demographical analysis with clinical parameters. R statistical platform base software version 3.3.2 and R-studio version. Cherry Blossom (3c53477a, 2023-03-09) was used for satistical data analysis and graphical representation. In all analyses, a p-value less than 0.05 was considered statistically significant.
For survival analysis, patients were divided into low and high expression groups, using median expression of genes and microRNAs as the cut-off value, and a log-rank test was applied for survival analysis and Kaplan-Meier survival plots were generated. In addition, Cox regression analysis was performed on the same group of patients to quantify the comparative hazard.
Author Contributions
WA: Manuscript writing, experiment and data analysis. NM: Conceptualization of the study. Provided guidance in manuscript writing, analysis and interpretation of the results and finalization of paper. SAE: Conceptualization of study. Sample collection. Final draft review SHA: Supervision. Helped in manuscript writing and analysis and reviewed manuscript. SS: Data analysis and interpretation and in the final drafting of paper. SN, AAL, WA: Helped in the retrieval of data and gathering. KG: Supervision. Manuscript review and editing. MHB: Data analysis. Manuscript review.
Funding
This study was funded by Higher Education Commission (No: Ref No. 20-16919/NRPU/R&D/HEC/2021 2021).
Institutional Review Board Statement
All methods were carried out following relevant guidelines and regulations. This study is carried out after the approval of the Aga Khan University (AKU) Ethics Review Committee (AKU-ERC # 2019-1945-5110). Informed consent was obtained from all individual participants included in the study.
Informed Consent Statement
Written informed consent has been obtained from all participants to publish this paper.
Data Availability Statement
Original data are available on request.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
List of primers for autophagy associated genes and microRNAs used in the qPCR array
| Forward primer | Reverse primer | ||
| Genes & microRNAs | Beclin1 | 5’-AATGACTTTTTTCCTTAGGGGG-3’ | 5’ -GTGGCTTTTGTGGATTTTTTCT-3’ |
| Mtor | 5’-TGGGACAGCATGGAAGAATA-3’ | 5’- TGTTGTGCCAAGGAGAAGAG-3’ | |
| UVRAG | 5′- CTGTTGCCCTTGGTTATACTGC -3′ | 5′- GATGATTTCTTCTGCTTGCTCC -3′ | |
| VPS34 | 5′-GCTGTCCTGGAAGACCCAAT-3′ | 5′-TTCTCACTGGCAAGGCCAAA-3′ | |
| PTEN | 5’CCAAGCTTATGACAGCCATCATC-3’ | 5’-CGCGGATCCTCAGACTTTTGTAA-3’ | |
| ULK1 | 5’-GGACACCATCAGGCTCTTCC-3’ | 5’-GAAGCCGAAGTCAGCGATCT-3’ | |
| ULK2 | 5’-TTCCTGCTCTAAGGGTTTGCTT-3’ | 5’-CCAGCGAGGGAGAACAACTG-3’ | |
| PI3K | 5’ - ATGCAAATTCAGTGCAAAGG-3’ | 5’ - CGTGTAAACAGGTCAATGGC-3’ | |
| AKT | 5’ -GCAGCACGTGTACGAGAAGA-3’ | 5’ -GGTGTCAGTCTCCGACGTG-3’ | |
| miR-7 | 5’ -AAAACTGCTGCCAAAACCAC-3’ | 5’ -GCTGCATTTTACAGCGACCAA-3’ | |
| miR-30 | 5’ -GGGGTGTAAACATCCTCGACTG-3’ | 5’ -ATTGCGTGTCGTGGAGTCG-3’ | |
| miR-100 | 5’ -GAACCCGTAGATCCGAACT-3’ | 5’ -CAGTGCGTGTCGTGGAGT-3’ | |
| miR-126 | 5’ TATGGTTGTTCTCGACTCCTTCAC-3’ | 5’ TCGTCTGTCGTACCGTGAGTAAT-3’ | |
| miR-21 | 5’-GTCGTATCCAGTGATACGACTCAACA-3’ | 5’ -GTCGTATCCAGTGCAGGGTCC-3’ | |
| miR-374 | 5’ -CCCGGGTTATAATACAACCTG-3’ | 5’ -CTCAACTGGTGTCGTGGAGTC-3’ | |
| miR-204 | 5’ -GCTACAGTCTTTCTTCATGTG-3’ | 5’ -CCAGTGATGACAATTGAACG-3’ |
References
- Hersh, A.M.; Gaitsch, H.; Alomari, S.; Lubelski, D.; Tyler, B.M. Molecular Pathways and Genomic Landscape of Glioblastoma Stem Cells: Opportunities for Targeted Therapy. Cancers 2022, 14, 3743. [Google Scholar] [CrossRef] [PubMed]
- Chaurasia, A.; Park, S.-H.; Seo, J.-W.; Park, C.-K. Immunohistochemical Analysis of ATRX, IDH1 and p53 in Glioblastoma and Their Correlations with Patient Survival. J. Korean Med Sci. 2016, 31, 1208–1214. [Google Scholar] [CrossRef] [PubMed]
- Pistritto, G.; Trisciuoglio, D.; Ceci, C.; Garufi, A.; D’Orazi, G. Apoptosis as anticancer mechanism: function and dysfunction of its modulators and targeted therapeutic strategies. Aging 2016, 8, 603–619. [Google Scholar] [CrossRef]
- Wang, G.; Wang, J.J.; Tang, H.M.; To, S.S.T. Targeting strategies on miRNA-21 and PDCD4 for glioblastoma. Arch. Biochem. Biophys. 2015, 580, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Zarzynska, J.M. The Importance of Autophagy Regulation in Breast Cancer Development and Treatment. BioMed Res. Int. 2014, 2014, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Schlã¤Fli, A.; Berezowska, S.; Adams, O.; Langer, R.; Tschan, M. Reliable LC3 and p62 autophagy marker detection in formalin fixed paraffin embedded human tissue by immunohistochemistry. Eur. J. Histochem. 2015, 59, 2481. [Google Scholar] [CrossRef]
- Ju, J.; Fesler, A.; Liu, H.; Wu, N.; Liu, F.; Ling, P. Autophagy regulated by miRNAs in colorectal cancer progression and resistance. Cancer Transl. Med. 2017, 3, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2ˆ(–delta delta CT) method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinform. Biomath. 2013, 3, 71–85. [Google Scholar]
- Zhu, L.; Mou, Q.; Wang, Y.; Zhu, Z.; Cheng, M. Resveratrol contributes to the inhibition of liver fibrosis by inducing autophagy via the microRNA-20a-mediated activation of the PTEN/PI3K/AKT signaling pathway. Int. J. Mol. Med. 2020, 46, 2035–2046. [Google Scholar] [CrossRef] [PubMed]
- Thambi, R. Histopathological Analysis of Brain Tumours- A Seven Year Study from a Tertiary Care Centre in South India. J. Clin. Diagn. Res. 2017, 11, EC05–EC08. [Google Scholar] [CrossRef]
- Samad, L.; Jawed, F.; Sajun, S.Z.; Arshad, M.H.; Baig-Ansari, N. Barriers to Accessing Surgical Care: A Cross-Sectional Survey Conducted at a Tertiary Care Hospital in Karachi, Pakistan. World J. Surg. 2013, 37, 2313–2321. [Google Scholar] [CrossRef] [PubMed]
- Enam, S.A.; Abdullah, U.E.H.; Laghari, A.A.; Khalid, M.U.; Rashid, H.; Jabbar, A.A.; Mubarak, F.; Hafiz, A.; Shamim, S. Current management of glioma in Pakistan. Glioma 2019, 2, 139. [Google Scholar] [CrossRef]
- Stark, A.M.; Doukas, A.; Hugo, H.-H.; Mehdorn, H.M. The expression of mismatch repair proteins MLH1, MSH2 and MSH6 correlates with the Ki67 proliferation index and survival in patients with recurrent glioblastoma. Neurol. Res. 2010, 32, 816–820. [Google Scholar] [CrossRef] [PubMed]
- Parzych, K.R.; Klionsky, D.J. An Overview of Autophagy: Morphology, Mechanism, and Regulation. Antioxidants Redox Signal. 2014, 20, 460–473. [Google Scholar] [CrossRef] [PubMed]
- Casares-Crespo, L.; Calatayud-Baselga, I.; García-Corzo, L.; Mira, H. On the Role of Basal Autophagy in Adult Neural Stem Cells and Neurogenesis. Front. Cell. Neurosci. 2018, 12, 339. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; et al. Prognostic significance of p62/SQSTM1 subcellular localization and LC3B in oral squamous cell carcinoma. British journal of cancer 2014, 111, 944–954. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Abeliovich, H.; Agostinis, P.; Agrawal, D.K.; Aliev, G.; Askew, D.S.; Baba, M.; Baehrecke, E.H.; Bahr, B.A.; Ballabio, A.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy in higher eukaryotes. Autophagy 2008, 4, 151–175. [Google Scholar] [CrossRef]
- Jiang, T.; Wu, Z. Immunohistochemical assessment of autophagic protein LC3B and p62 levels in glioma patients. Int. J. Clin. Exp. Pathol. 2018, 11, 862–868. [Google Scholar]
- Aoki, H.; Kondo, Y.; Aldape, K.; Yamamoto, A.; Iwado, E.; Yokoyama, T.; Hollingsworth, E.F.; Kobayashi, R.; Hess, K.; Shinojima, N.; et al. Monitoring autophagy in glioblastoma with antibody against isoform B of human microtubule-associated protein 1 light chain 3. Autophagy 2008, 4, 467–475. [Google Scholar] [CrossRef]
- Shukla, S.; Patric, I.R.P.; Patil, V.; Shwetha, S.D.; Hegde, A.S.; Chandramouli, B.A.; Arivazhagan, A.; Santosh, V.; Somasundaram, K. Methylation Silencing of ULK2, an Autophagy Gene, Is Essential for Astrocyte Transformation and Tumor Growth. J. Biol. Chem. 2014, 289, 22306–22318. [Google Scholar] [CrossRef]
- Boyd, N.H.; Tran, A.N.; Bernstock, J.D.; Etminan, T.; Jones, A.B.; Gillespie, G.Y.; Friedman, G.K.; Hjelmeland, A.B. Glioma stem cells and their roles within the hypoxic tumor microenvironment. Theranostics 2021, 11, 665–683. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; et al. Knockdown lncRNA CRNDE enhances temozolomide chemosensitivity by regulating autophagy in glioblastoma. Cancer Cell International 2021, 21, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Crabtree, J.S.; Jelinsky, S.A.; Harris, H.A.; Choe, S.E.; Cotreau, M.M.; Kimberland, M.L.; Wilson, E.; Saraf, K.A.; Liu, W.; McCampbell, A.S.; et al. Comparison of Human and Rat Uterine Leiomyomata: Identification of a Dysregulated Mammalian Target of Rapamycin Pathway. Cancer Res 2009, 69, 6171–6178. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Wang, C.; Lei, F.; Zhang, L.; Zhang, X.; Liu, A.; Wu, G.; Zhu, J.; Song, L. miR-93 Promotes Cell Proliferation in Gliomas through Activation of PI3K/Akt Signaling Pathway. Oncotarget 2015, 6, 8286–8299. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Wang, W.; Su, N.; Zhu, X.; Yao, J.; Gao, W.; Hu, Z.; Sun, Y. miR-374a promotes cell proliferation, migration and invasion by targeting SRCIN1 in gastric cancer. FEBS Lett. 2014, 589, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Peralta-Zaragoza, O.; Deas, J.; Meneses-Acosta, A.; De la O-Gómez, F.; Fernández-Tilapa, G.; Gómez-Cerón, C.; Benítez-Boijseauneau, O.; Burguete-García, A.; Torres-Poveda, K.; Bermúdez-Morales, V.H.; et al. Relevance of miR-21 in regulation of tumor suppressor gene PTEN in human cervical cancer cells. BMC Cancer 2016, 16, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.-H.; Tian, D.; Yang, Z.-C.; Li, J.-L. Exosomal miR-21 promotes proliferation, invasion and therapy resistance of colon adenocarcinoma cells through its target PDCD4. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.-E.; Suh, H.-W.; Bahal, R.; Josowitz, A.; Zhang, J.; Song, E.; Cui, J.; Noorbakhsh, S.; Jackson, C.; Bu, T.; et al. Nanoparticle-mediated intratumoral inhibition of miR-21 for improved survival in glioblastoma. Biomaterials 2019, 201, 87–98. [Google Scholar] [CrossRef]
- Yang, C.H.; Yue, J.; Pfeffer, S.R.; Fan, M.; Paulus, E.; Hosni-Ahmed, A.; Sims, M.; Qayyum, S.; Davidoff, A.M.; Handorf, C.R.; et al. MicroRNA-21 Promotes Glioblastoma Tumorigenesis by Down-regulating Insulin-like Growth Factor-binding Protein-3 (IGFBP3). J. Biol. Chem. 2014, 289, 25079–25087. [Google Scholar] [CrossRef]
- Dong, Q.; Yuan, G.; Liu, M.; Xie, Q.; Hu, J.; Wang, M.; Liu, S.; Ma, X.; Pan, Y. Downregulation of microRNA-374a predicts poor prognosis in human glioma. Exp. Ther. Med. 2019, 17, 2077–2084. [Google Scholar] [CrossRef]
- Zhang, S.-B.; Liu, T.-J.; Pu, G.-H.; Li, B.-Y.; Gao, X.-Z.; Han, X.-L. MicroRNA-374 Exerts Protective Effects by Inhibiting SP1 Through Activating the PI3K/Akt Pathway in Rat Models of Myocardial Ischemia-Reperfusion After Sevoflurane Preconditioning. Cell. Physiol. Biochem. 2018, 46, 1455–1470. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Wu, H.; Liu, X.; Li, B.; Chen, Y.; Ren, X.; Liu, C.-G.; Yang, J.-M. Regulation of autophagy by a beclin 1-targeted microRNA, miR-30a, in cancer cells. Autophagy 2009, 5, 816–823. [Google Scholar] [CrossRef] [PubMed]
- Sibilano, M.; Tullio, V.; Adorno, G.; Savini, I.; Gasperi, V.; Catani, M.V. Platelet-Derived miR-126-3p Directly Targets AKT2 and Exerts Anti-Tumor Effects in Breast Cancer Cells: Further Insights in Platelet-Cancer Interplay. Int. J. Mol. Sci. 2022, 23, 5484. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-R.; Cai, W.-P.; Dai, X.-J.; Guo, A.-S.; Chen, H.-P.; Lin, G.-S.; Lin, R.-S. Research on miR-126 in glioma targeted regulation of PTEN/PI3K/Akt and MDM2-p53 pathways. Eur. Rev. Med Pharmacol. Sci. 2019, 23, 3461–3470. [Google Scholar] [PubMed]
- Akkoc, Y.; Gozuacik, D. MicroRNAs as major regulators of the autophagy pathway. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research 2020, 1867, 118662. [Google Scholar] [CrossRef] [PubMed]
- Gu, D.-N.; Jiang, M.-J.; Mei, Z.; Dai, J.-J.; Dai, C.-Y.; Fang, C.; Huang, Q.; Tian, L. microRNA-7 impairs autophagy-derived pools of glucose to suppress pancreatic cancer progression. Cancer Lett. 2017, 400, 69–78. [Google Scholar] [CrossRef]
- Korać, P.; Antica, M.; Matulić, M. MiR-7 in Cancer Development. Biomedicines 2021, 9, 325. [Google Scholar] [CrossRef]
- Seca, H.; Lima, R.; Lopes-Rodrigues, V.; Guimaraes, J.; Gabriela, G.; Vasconcelos, M. Targeting miR-21 Induces Autophagy and Chemosensitivity of Leukemia Cells. Curr. Drug Targets 2013, 14, 1135–1143. [Google Scholar] [CrossRef]
- Duzgun, Z.; Eroglu, Z.; Avci, C.B. Role of mTOR in glioblastoma. Gene 2016, 575, 187–190. [Google Scholar] [CrossRef]
- Mecca, C.; Giambanco, I.; Donato, R.; Arcuri, C. Targeting mTOR in Glioblastoma: Rationale and Preclinical/Clinical Evidence. Dis. Markers 2018, 2018, 1–10. [Google Scholar] [CrossRef]
- Hashemi, M.; Etemad, S.; Rezaei, S.; Ziaolhagh, S.; Rajabi, R.; Rahmanian, P.; Abdi, S.; Koohpar, Z.K.; Rafiei, R.; Raei, B.; et al. Progress in targeting PTEN/PI3K/Akt axis in glioblastoma therapy: Revisiting molecular interactions. BioMedicine 2023, 158, 114204. [Google Scholar] [CrossRef] [PubMed]
- Morelli, M.B.; Nabissi, M.; Amantini, C.; Maggi, F.; Ricci-Vitiani, L.; Pallini, R.; Santoni, G. TRPML2 Mucolipin Channels Drive the Response of Glioma Stem Cells to Temozolomide and Affect the Overall Survival in Glioblastoma Patients. Int. J. Mol. Sci. 2022, 23, 15356. [Google Scholar] [CrossRef] [PubMed]
- Cj, P.L.; et al. High LC3/Beclin expression correlates with poor survival in glioma: a definitive role for autophagy as evidenced by in vitro autophagic flux. Pathology & Oncology Research 2019, 25, 137–148. [Google Scholar]
- Stafford, M.C.; Willoughby, C.E.; Walsh, C.P.; McKenna, D.J. Prognostic value of miR-21 for prostate cancer: a systematic review and meta-analysis. Biosci. Rep. 2022, 42, BSR20211972. [Google Scholar] [CrossRef] [PubMed]
- Irimie-Aghiorghiesei, A.I.; et al. Prognostic value of MiR-21: an updated meta-analysis in Head and Neck Squamous Cell Carcinoma (HNSCC). Journal of clinical medicine 2019, 8, 2041. [Google Scholar] [CrossRef]
- Sufianov, A.; Begliarzade, S.; Ilyasova, T.; Liang, Y.; Beylerli, O. MicroRNAs as prognostic markers and therapeutic targets in gliomas. Non-coding RNA Res. 2022, 7, 171–177. [Google Scholar] [CrossRef]
- Nieland, L.; van Solinge, T.S.; Cheah, P.S.; Morsett, L.M.; El Khoury, J.; Rissman, J.I.; Kleinstiver, B.P.; Broekman, M.L.; Breakefield, X.O.; Abels, E.R. CRISPR-Cas knockout of miR21 reduces glioma growth. Mol. Ther. - Oncolytics 2022, 25, 121–136. [Google Scholar] [CrossRef]
- Cui, H.; Mu, Y.; Yu, L.; Xi, Y.-G.; Matthiesen, R.; Su, X.; Sun, W. Methylation of the miR-126 gene associated with glioma progression. Fam. Cancer 2015, 15, 317–324. [Google Scholar] [CrossRef]
- Shukla, A.; Sehgal, M.; Singh, T.R. Hydroxymethylation and its potential implication in DNA repair system: A review and future perspectives. Gene 2015, 564, 109–118. [Google Scholar] [CrossRef]
Figure 1.
LC3-II in low- and high-grade glioma. (a) Immunohistochemistry images. (b) Quantification for LC3-II punctea was based on the intensity score from 1-3, where score 1, punctae ≤ 10 per cells; score 2, punctae 11-20 per cells; score 3, punctae >20 per cells c) RT-qPCR shows a significant difference in median expression of LC3-II in low and high-grade glioma tissues using Mann-Whitney test.
Figure 1.
LC3-II in low- and high-grade glioma. (a) Immunohistochemistry images. (b) Quantification for LC3-II punctea was based on the intensity score from 1-3, where score 1, punctae ≤ 10 per cells; score 2, punctae 11-20 per cells; score 3, punctae >20 per cells c) RT-qPCR shows a significant difference in median expression of LC3-II in low and high-grade glioma tissues using Mann-Whitney test.
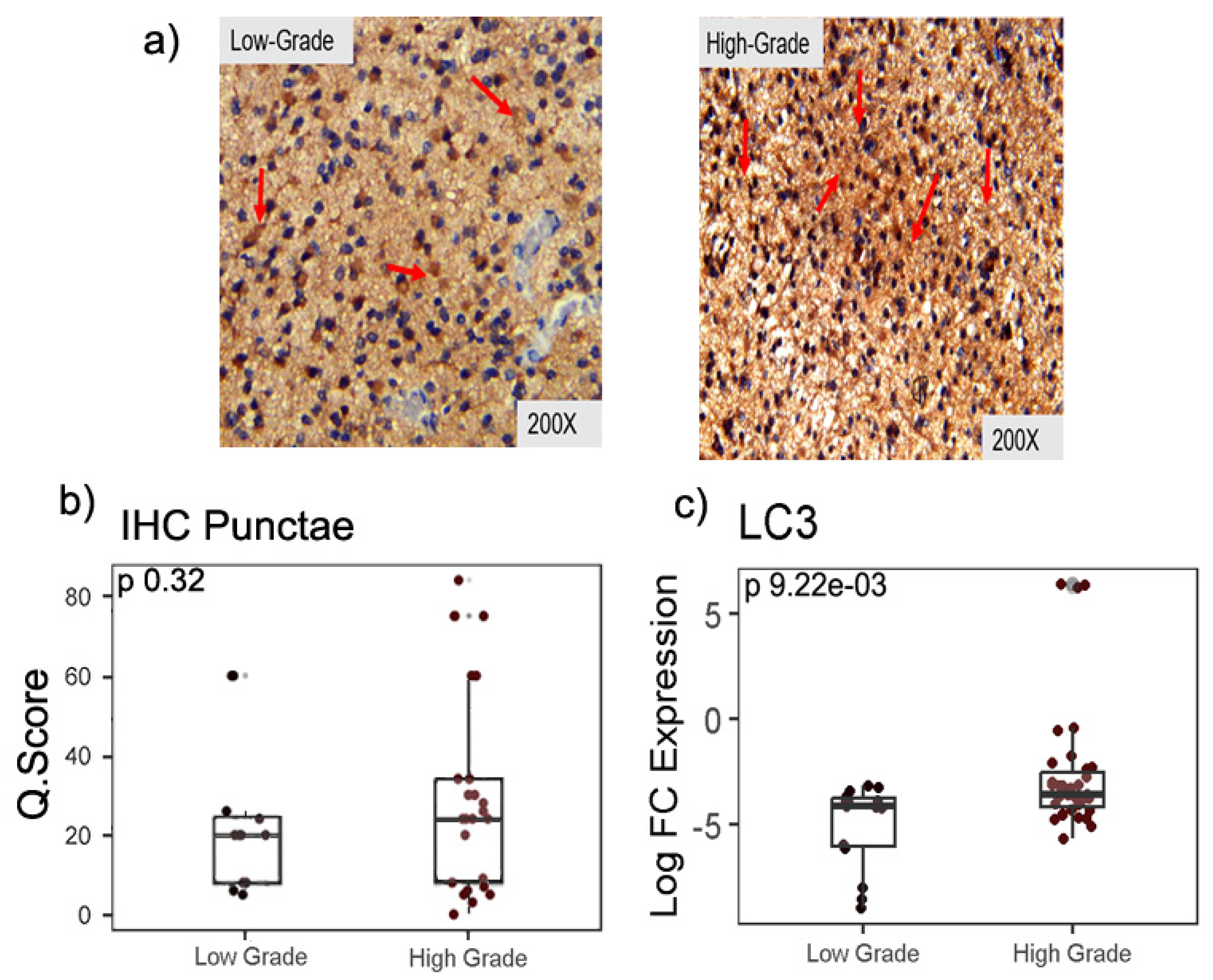
Figure 2.
RT-qPCR analysis for Beclin1, mTOR, ULK1, ULK2, UVRAG, Vps34 gene expression in low and high-grade glioma tissues. Significant difference in median expression of genes between LGG and HGG was determined using Mann-Whitney test.
Figure 2.
RT-qPCR analysis for Beclin1, mTOR, ULK1, ULK2, UVRAG, Vps34 gene expression in low and high-grade glioma tissues. Significant difference in median expression of genes between LGG and HGG was determined using Mann-Whitney test.
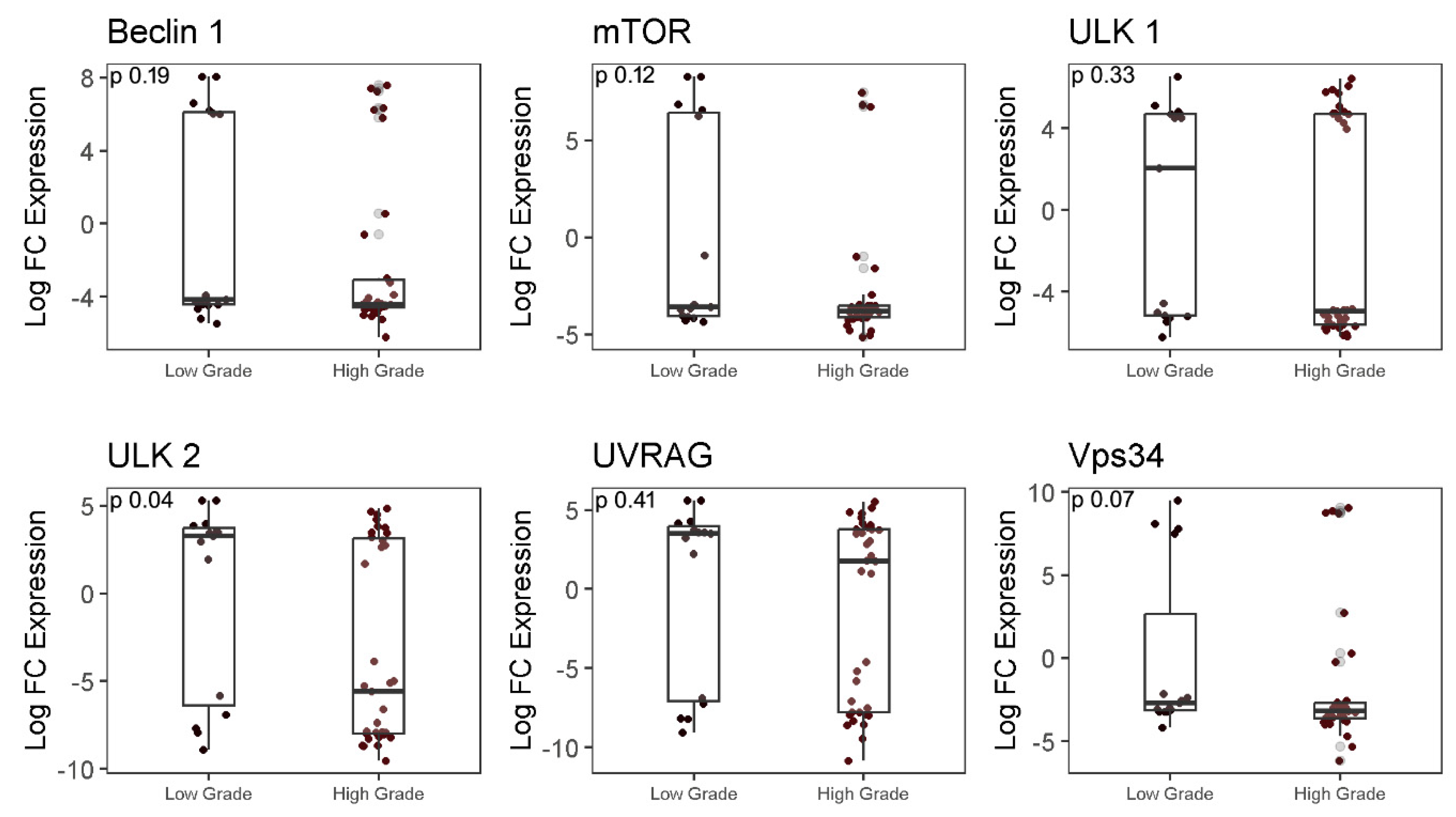
Figure 3.
RT-qPCR analysis for PI3K/PTEN/AKT gene expression in low- and high-grade glioma. A significant difference in median expression of genes between LGG and HGG was determined using Mann-Whitney test.
Figure 3.
RT-qPCR analysis for PI3K/PTEN/AKT gene expression in low- and high-grade glioma. A significant difference in median expression of genes between LGG and HGG was determined using Mann-Whitney test.
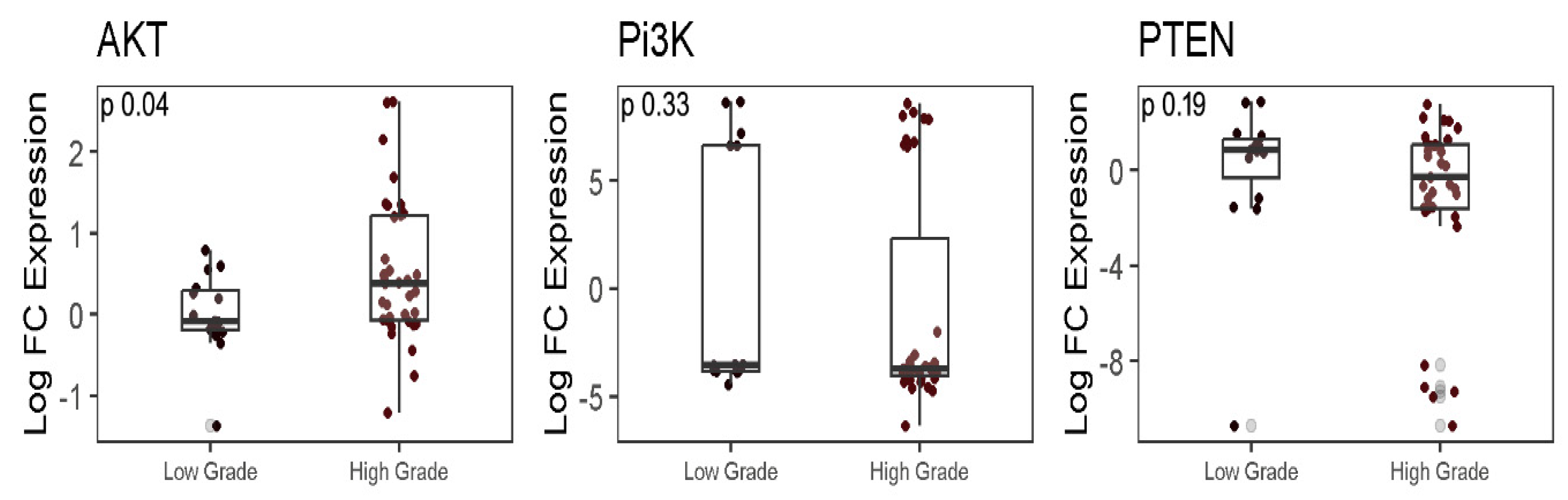
Figure 4.
RT-qPCR analysis for micro-RNA expression in low and high-grade glioma tissues. A significant difference in median expression of genes between LGG and HGG was determined using Mann-Whitney test.
Figure 4.
RT-qPCR analysis for micro-RNA expression in low and high-grade glioma tissues. A significant difference in median expression of genes between LGG and HGG was determined using Mann-Whitney test.
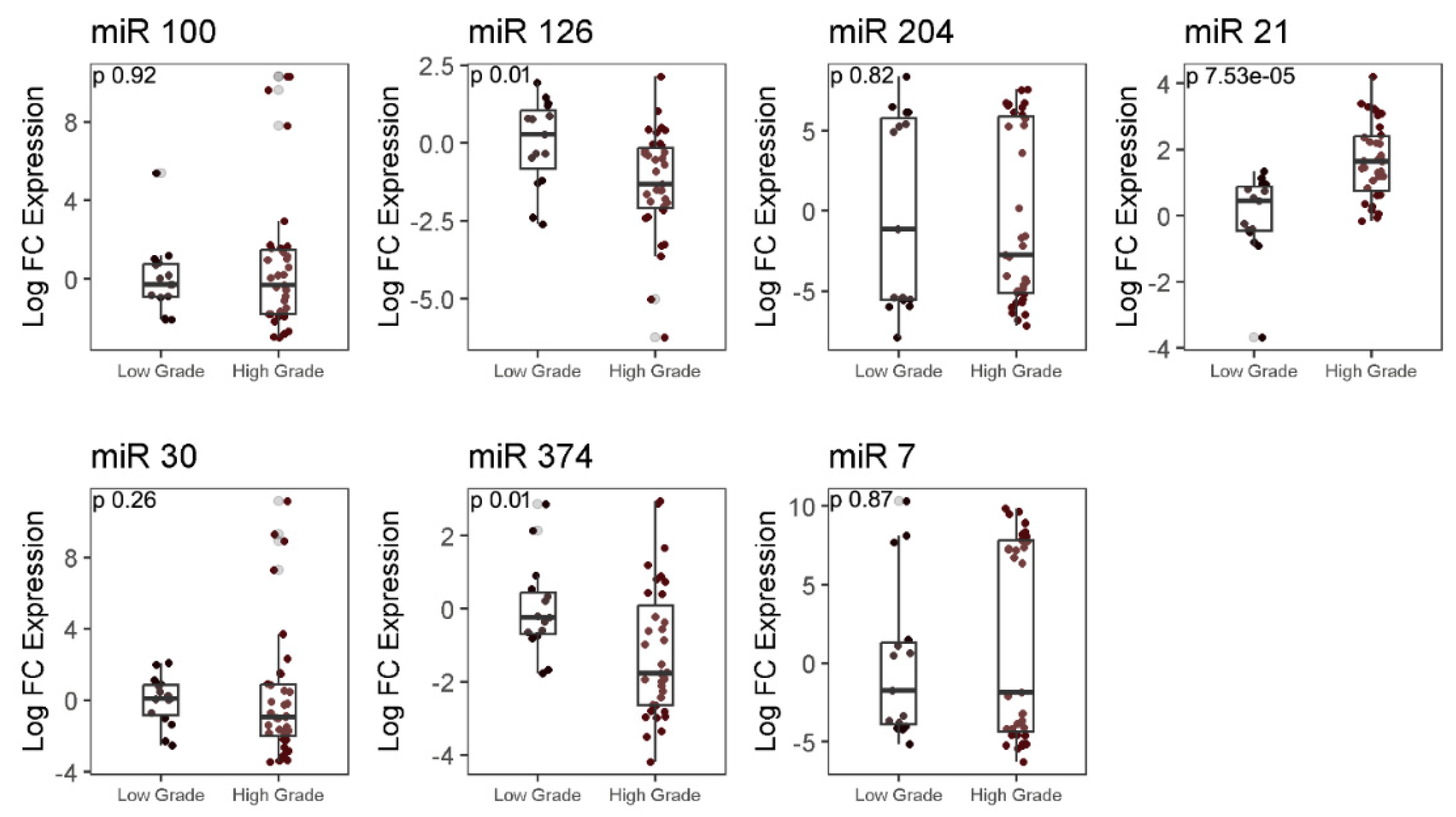
Figure 5.
a & b). The prognosis of autophagy genes and microRNAs in glioma tissue samples (n=50). Kaplan-Meier survival analysis was performed after glioma patients were clinically followed up for 40 months post-surgery and segregating patients into two groups based on median expression: patients with high expression of genes (red) and patients with low expression of genes (blue).
Figure 5.
a & b). The prognosis of autophagy genes and microRNAs in glioma tissue samples (n=50). Kaplan-Meier survival analysis was performed after glioma patients were clinically followed up for 40 months post-surgery and segregating patients into two groups based on median expression: patients with high expression of genes (red) and patients with low expression of genes (blue).
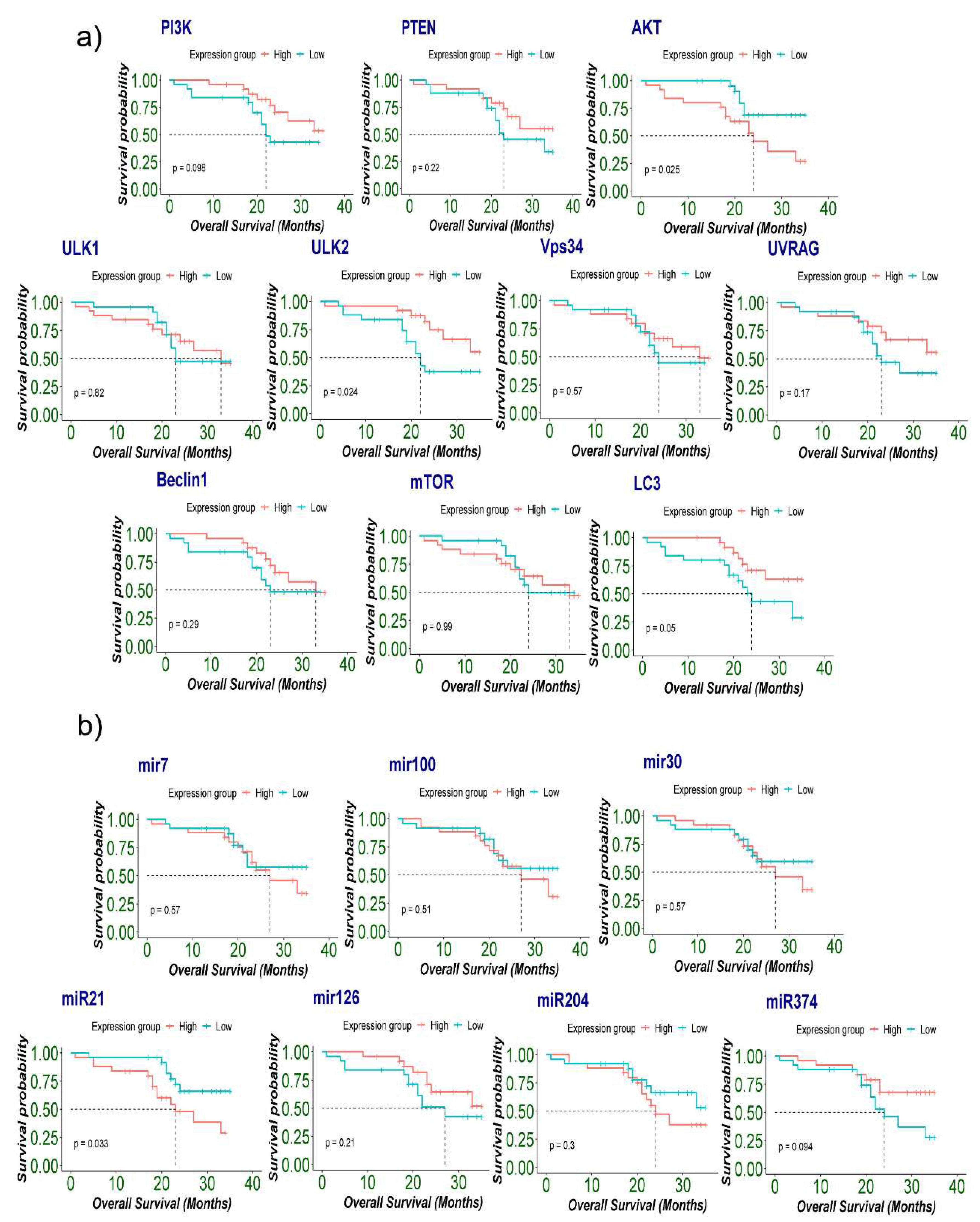
Figure 6.
a & b) The prognosis of autophagy genes (a) and miRNA (b) in background of IDH1, Ki67 and p53 genes in glioma tissue samples (n=50). Kaplan-Meier survival analysis was performed after glioma patients were clinically followed up for 40 months post-surgery. Molecular analysis of genes and miRs (AKT, ULK2, LC3, miR-21, miR-126 and miR-374) transcription levels in glioma tissue was measured by RT-qPCR, while IDH1, Ki67 and p53 protein expression in paraffin-embedded glioma tissue sections was detected using immunohistochemistry. .
Figure 6.
a & b) The prognosis of autophagy genes (a) and miRNA (b) in background of IDH1, Ki67 and p53 genes in glioma tissue samples (n=50). Kaplan-Meier survival analysis was performed after glioma patients were clinically followed up for 40 months post-surgery. Molecular analysis of genes and miRs (AKT, ULK2, LC3, miR-21, miR-126 and miR-374) transcription levels in glioma tissue was measured by RT-qPCR, while IDH1, Ki67 and p53 protein expression in paraffin-embedded glioma tissue sections was detected using immunohistochemistry. .
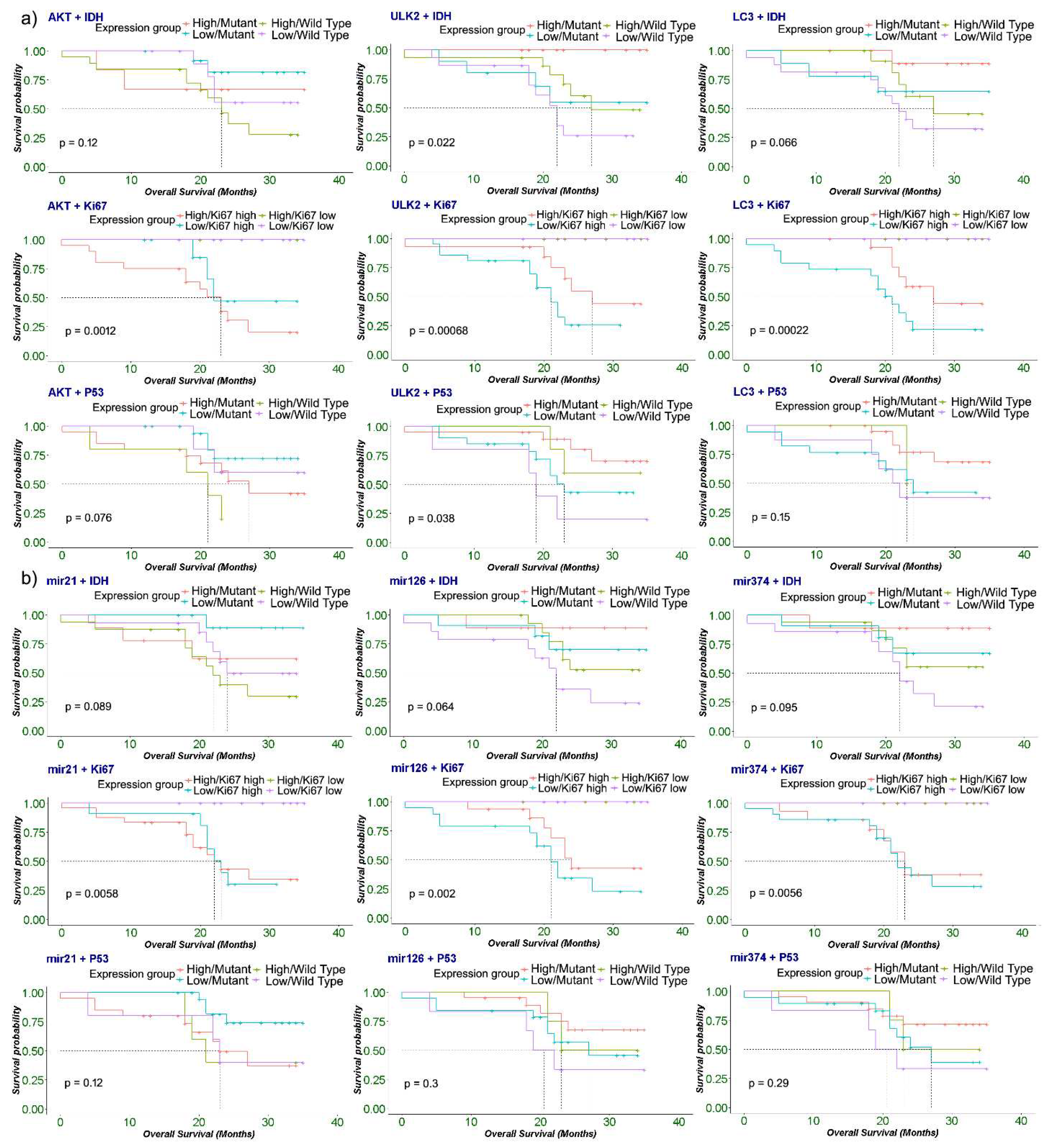

Table 1.
Summary of clinicopathological characteristics of glioma cases.
| Clinicopathological features | Values (%) |
|---|---|
|
Gender Male Female |
34 (68%) 16 (32%) |
|
Age (years) Medians (range) |
38 (4-70) |
|
Histological type (WHO grade) II III IV |
15 (30%) 5 (10%) 30 (60%) |
|
Histological group LGG HGG |
15 (30%) 35 (70%) |
|
Status at 4 years Dead Alive LTFU |
18 (36%) 31 (62%) 1 (20%) |
|
Recurrence No recurrence Recurrence |
30 (62%) 9 (38%) |
|
Radiotherapy Yes No |
23 (46%) 23 (46%) |
|
Chemotherapy Yes No |
23 (46%) 23 (46%) |
| Adjuvant chemoradiotherapy |
21 (42%) |
|
Postoperative KPS score >=80 <80 |
22 (44%) 26 (52%) |
|
Overall survival months Median (Range) |
22.6 (4-35) |
|
Molecular Profile IDH-1 Mutation ATRX Retained p53 Overexpression Ki-67 Overexpression |
20 (40%) 50 (100%) 40 (80%) 35 (35%) |
Table 2.
Correlation of molecular protein expression ATRX, IDH1, p53 and Ki67 with clinicopathological factors. Rows with variables exhibiting significant correlation (p<0.05) are shaded grey.
Table 2.
Correlation of molecular protein expression ATRX, IDH1, p53 and Ki67 with clinicopathological factors. Rows with variables exhibiting significant correlation (p<0.05) are shaded grey.
| Table 2 | Total No. of samples | ATRX | IDH1 | p53 | Ki67 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Factors | n=50 | Retained n=50 |
Loss n=0 |
Wild Type n=30 |
Mutant n=20 |
P-value | Positive n=40 |
Negative n=10 |
P-value | High n=35 |
Low n=15 |
P-value |
|
Age <40 |
32 | 32 (100%) | 0 (0%) | 19 (59.3%) | 13 (40.6%) | 1 | 27 (84.3%) | 5 (15.6%) | 0.5 | 18 (56.2%) | 14 (43.7%) |
0.005 |
| >40 | 18 | 18 (100%) | 0 (0%) | 11 (61.1%) | 7 (38.8%) | 13 (72.2%) | 5 (27.7%) | 17 (94.4%) | 1 (5.5%) | |||
|
Gender Female |
16 | 16 (100%) | 0 (0%) | 9 (56.2%) | 7 (43.7%) | 0.76 | 12 (75%) | 4 (25%) | 0.7 | 13 (81.2%) | 3 (18.7%) | 0.32 |
| Male | 34 | 34 (100%) | 0 (0%) | 21 (61.7%) | 13 (38.2%) | 28 (82.3%) | 6 (17.6%) | 22 (64.7%) | 12 (35.2%) | |||
|
Histological Type LGG (grade II) |
15 | 15 (100%) | 0 (0%) | 8 (53.3%) | 7 (46.66%) | 0.23 | 13 (86.66%) | 2 (13.33%) | 0.7 | 1(6.66) | 14 (93.33%) | <0.001 |
| HGG (III, IV) | 35 | 35(100%) | 0 (0%) | 22 (62.85%) | 13 (37.14%) | 27 (77.14%) | 8 (22.85%) | 34 (97.14%) | 1 (2.85%) | |||
|
Chemotherapy Yes |
23 | 23 (100%) | 0 (0%) | 11 (47.8%) | 12 (52.1%) | 0.23 | 19 (82.6%) | 4 (17.3%) | 0.7 | 21 (91.3%) | 2 (8.6%) | 0.001 |
| No | 23 | 23 (100%) | 0 (0%) | 16 (69.5%) | 7 (30.4%) | 17 (73.9%) | 6 (26%) | 10 (43.4%) | 13 (56.5%) | |||
|
Radiotherapy Yes |
23 | 23 (100%) | 0 (0%) | 11 (47.8%) | 12 (52.1%) | 0.23 | 19 (82.6%) | 4 (17.3%) | 0.7 | 20 (86.9%) | 3 (13%) | 0.01 |
| No | 23 | 23 (100%) | 0 (0%) | 16 (69.5%) | 7 (30.4%) | 17 (73.9%) | 6 (26%) | 11 (47.8%) | 12 (52.1%) | |||
|
Recurrence Yes |
19 | 19 (100%) | 0 (0%) | 12 (63.1%) | 7 (36.8%) | 0.62 | 16 (84.2%) | 3 (15.7%) | 0.8 | 15 (78.9%) | 4 (21%) | 0.54 |
| No | 30 | 30 (100%) | 0 (0%) | 18 (60%) | 12 (40%) | 23 (76.6%) | 7 (23.3%) | 19 (63.3%) | 11 (36.6%) | |||
|
Current Status Dead |
18 | 18 (100%) | 0 (0%) | 14 (77.7%) | 4 (22.2%) | 0.13 | 13 (72.2%) | 5(27.7%) | 0.2 | 16 (51.6%) | 15(48.3%) | 0.004 |
| Alive | 31 | 31 (100%) | 0 (0%) | 15 (48.3%) | 16 (51.6%) | 27 (87%) | 4(12.9%) | 6 (40%) | 9 (60%) | |||
|
Tumor Type Oligodendroglioma |
15 | 15 (100%) | 0 (0%) | 6 (40%) | 9 (60%) | 0.19 | 13 (86.6%) | 2 (13%) | 0.5 | 7 (46.6%) | 8 (53.3%) | <0.001 |
| Glioblastoma | 29 | 29 (100%) | 0 (0%) | 20 (68.9%) | 9 (31%) | 22 (75.8%) | 7 (24.1%) |
29 (100%) |
0 (0%) |
|||
| Astrocytoma | 6 | 6 (100%) | 0 (0%) | 4 (66.6%) | 2 (33.3%) | 5 (83.3%) | 1 (16.6%) | 1 (16.66%) | 5 (83.33%) | |||
Table 3.
Analysis of correlation between the expression of different genes in LGG group. Correlation was determined using Spearman’s correlation test. Each column shows the coefficient of correlation (rs). Correlations with significant p-values (p<0.05) are shown in bold.
Table 3.
Analysis of correlation between the expression of different genes in LGG group. Correlation was determined using Spearman’s correlation test. Each column shows the coefficient of correlation (rs). Correlations with significant p-values (p<0.05) are shown in bold.
| Correlation between Gene to miRs in Low Grade Glioma | |||||||||||||||
| Genes/miRs | miR-7 | miR-30 | miR-100 | miR-126 | miR-204 | miR-374 | miR-21 | ||||||||
| ULK2 | 0.85 | 0.56 | 0.81 | 0.15 | 0.58 | 0.68 | 0.21 | ||||||||
| AKT | 0.45 | 0.19 | 0.04 | 0.11 | 0.19 | 0.16 | 0.03 | ||||||||
| LC3 | -0.31 | -0.02 | 0.02 | 0.24 | -0.18 | -0.09 | 0.43 | ||||||||
| miR-21 | 0.12 | 0.05 | 0.01 | 0.56 | 0.23 | 0.08 | 1.00 | ||||||||
| Correlation between Gene to Genes in Low Grade Glioma | |||||||||||||||
|
Genes /miRs |
ULK2 | AKT | LC3 | miR-21 | PI3K | PTEN | ULK1 | Vps34 | mTOR | Beclin | UVRAG | ||||
| ULK2 | 1.00 | 0.39 | -0.08 | 0.21 | 0.46 | 0.92 | 0.41 | 0.49 | 0.59 | 0.51 | 0.80 | ||||
| AKT | 0.39 | 1.00 | -0.38 | 0.03 | -0.02 | 0.24 | -0.31 | -0.21 | -0.05 | 0.11 | 0.30 | ||||
| LC3 | -0.08 | -0.38 | 1.00 | 0.43 | -0.01 | -0.02 | -0.12 | 0.00 | -0.14 | 0.00 | 0.01 | ||||
| miR-21 | 0.21 | 0.03 | 0.43 | 1.00 | 0.09 | 0.31 | 0.04 | 0.37 | 0.30 | 0.13 | 0.23 | ||||
Table 4.
Analysis of correlation between the expression of different genes in HGG group. Correlation was determined using Spearman’s correlation test. Each column shows the coefficient of correlation (rs). Correlations with significant p-values (p<0.05) are shown in bold.
Table 4.
Analysis of correlation between the expression of different genes in HGG group. Correlation was determined using Spearman’s correlation test. Each column shows the coefficient of correlation (rs). Correlations with significant p-values (p<0.05) are shown in bold.
| Correlation between Gene to miRs in Low Grade Glioma | ||||||||||||||||
| Genes/miRs | miR-7 | miR-30 | miR-100 | miR-126 | miR-204 | miR-374 | miR-21 | |||||||||
| ULK2 | 0.40 | 0.27 | 0.16 | 0.32 | 0.12 | 0.50 | 0.21 | |||||||||
| AKT | 0.66 | 0.41 | 0.30 | 0.38 | 0.40 | 0.35 | 0.38 | |||||||||
| LC3 | -0.03 | 0.08 | 0.20 | 0.07 | 0.22 | 0.01 | -0.19 | |||||||||
| miR-21 | 0.26 | 0.40 | 0.12 | 0.08 | -0.14 | 0.11 | 1.00 | |||||||||
| Correlation between Gene to Genes in High Grade Glioma | ||||||||||||||||
| Genes/miRs | ULK2 | AKT | LC3 | miR-21 | PI3K | PTEN | ULK1 | Vps34 | mTOR | Beclin | UVRAG | |||||
| ULK2 | 1.00 | 0.50 | -0.02 | 0.21 | 0.49 | 0.63 | 0.53 | 0.63 | 0.59 | 0.55 | 0.66 | |||||
| AKT | 0.50 | 1.00 | 0.13 | 0.38 | 0.48 | 0.48 | 0.59 | 0.56 | 0.43 | 0.34 | 0.69 | |||||
| LC3 | -0.02 | 0.13 | 1.00 | -0.19 | -0.02 | 0.18 | 0.06 | 0.19 | 0.09 | -0.15 | 0.13 | |||||
| miR-21 | 0.21 | 0.38 | -0.19 | 1.00 | 0.12 | 0.03 | 0.24 | 0.09 | -0.05 | -0.14 | 0.29 | |||||
Table 5.
Univariate COX proportional hazard analysis of autophagy-associated genes and microRNAs for overall survival OS.
Table 5.
Univariate COX proportional hazard analysis of autophagy-associated genes and microRNAs for overall survival OS.
| Univariable Cox regression | |||
|---|---|---|---|
| Genes | HR Ratio | CI (Lower 0.95 - Upper 0.95) | p-value (Log Rank) |
| AKT | 2.829 | 0.135 : 0.921 | 0.03* |
| ULK2 | 0.355 | 1.111 : 7.113 | 0.02* |
| LC3 | 0.41 | 0.968: 6.127 | 0.05* |
| miR-21 | 2.637 | 0.150 : 0.955 | 0.03* |
| miR-126 | 0.56 | 0.722: 4.35 | 0.2 |
| miR-374 | 0.463 | 0.859: 5.414 | 0.09 |
Abbreviation: HR= Hazard ratio *p<0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



