
Submitted:
21 August 2023
Posted:
22 August 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
We report two cases of pancytopenia in patients after recovering from a mild COVID-19, now presenting as paroxysmal nocturnal hemoglobinuria (PNH) and aplastic anemia. These cases il-lustrate a common pathway whereby a viral trigger causes clonal expansion of a hematological disorder. Although the association in both cases are temporal, and that COVID-19 may be an in-cidental diagnosis, the growing evidence related to hematological effects of SARS-CoV-2 infec-tion highlights the need for further investigation into the hematological consequences of COVID-19, particularly in the post-pandemic era.
Keywords:
Paroxysmal nocturnal hemoglobinuria
; aplastic anemia
; SARS-CoV-2
1. Introduction
COVID-19, caused by the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), has caused more than 6.8 million deaths globally as of March 22, 2023 [1]. There are known associations of hematological disorders with other viruses from the Coronaviridae family, such as SARS-CoV-1 and Middle East Respiratory Syndrome coronavirus (MERS-CoV) [2]. Similarly, SARS-CoV-2 has been linked to a broad spectrum of hematological abnormalities ranging from mild leukopenia to fulminant bone marrow failure. More recently, acquired bone marrow failure syndromes, including aplastic anemia and paroxysmal nocturnal hemoglobinuria, have been reported in the literature as a sequela of COVID-19 infection [3,4,5,6].
The pathophysiology of marrow aplasia can be subdivided into three main categories: Physical or chemical damage, immune-mediated destruction, or a constitutional genetic defect [7]. The most common etiology of aplastic anemia is immune-mediated, and 80% of these cases respond remarkably well to immunosuppressive therapy [7]. The mechanism behind immune-mediated aplastic anemia is not fully understood, however, culminating evidence points to the role of viral infections leading to dysregulated immune responses and destruction of hematopoietic stem cells (HSCs) [7].
PNH is an acquired disorder of HSCs characterized by somatic mutations in the glycosylphosphatidylinositol (GPI) anchor protein synthesis, which leads to a deficiency of complement regulatory proteins and unregulated complement-mediated hemolysis [7,8,9]. More often, AA and PNH can present in the same patient as a moving target in the same spectrum.
Though etiologically challenging to prove the cause for AA/PNH, it is imperative to report this rare condition in association with the recent SARS-COV2 pandemic. Here we describe two cases of aplastic anemia and PNH presenting as pancytopenia associated with SARS-CoV-2.
Objective
To present two cases of acquired pancytopenia associated with COVID-19 manifesting as PNH and AA.
2. Case Series
2.1. Case 1
A previously healthy 21-year-old male presents to the hematology clinic for pancytopenia that was discovered during pre-operative evaluation for an ankle open reduction and internal fixation (ORIF) in April 2022. His only medical history is that of mild COVID-19 infection confirmed by PCR and no family history of any hematological disorders. Initial labs revealed a hemoglobin of 9.3 g/dL, white blood cell count at 4.1 x 10^3 cells/uL, absolute neutrophil count at 2.74 x 10^3 cells/uL, and platelet count of 34 x 10^3 cells/uL. His coagulation profile (PT/INR, APTT), D-dimer and fibrinogen levels were within reference range. COVID-19 by nasopharyngeal polymerase chain reaction (PCR) was negative. Hemolytic labs revealed an LDH of 595 U/L and an elevated total bilirubin of 1.4 mg/dL. Bone marrow biopsy showed decreased trilineage hematopoiesis, but no blasts. Further cytogenetic studies showed an abnormal 13q deletion in 2 out of 20 cells. PNH flow cytometry identified PNH clones in granulocytes (19.53%), monocytes (19.77%), and RBCs (3.61%; 0.22% type II cells and 3.39% type III cells). A diagnosis of hemolytic paroxysmal nocturnal hemoglobinuria/moderate aplastic anemia combination syndrome was made. He was given the appropriate immunization for encapsulated organisms prior to starting on ravulizumab. Five months after diagnosis, and after completing 3 doses of ravulizumab, he has improvement in counts, and remains transfusion independent.
2.2. Case 2
A 52-year-old female presents to hematology clinic with increased bruising. She has a medical history of anterior uveitis and fibromyalgia, however, no family history of hematological disorders. She contracted a mild COVID-19 infection in May 2022 confirmed by PCR and finished a course of nirmatrelvir-ritonavir (Paxlovid) without experiencing any respiratory compromise. A month following her recovery, she was noted to have easy bruising which prompted a visit to her primary care physician where basic labs were drawn and she was found to be pancytopenic. On exam, she did not have any active bleeding, petechiae, abdominal pain or melena. Of note, she completed her initial vaccination series for SARS-CoV-2 with Johnson & Johnson/Janssen vaccines in 2021 and seven months later, she followed up with a Pfizer-BioNtech booster. Initial laboratory studies revealed hemoglobin of 11.6 mg/dL, white blood cell count of 3.0 x 10^3 cells/uL, absolute neutrophil count of 0.9 x 10^3 cells/uL and platelet count of 18 x 10^3 cells/uL. Bone marrow biopsy revealed decreased trilineage hematopoiesis and no increase in blasts, however, rare, small irregular lymphoid aggregates composed of small mature T and B lymphocytes were present. Flow cytometry revealed normal immunophenotypic results. Next generation sequencing and cytogenetic studies did not reveal any abnormalities, but PNH flow cytometry identified minute PNH clones in FLAER and CD157 negative neutrophils and monocytes (0.5%). The patient was diagnosed with moderate aplastic anemia with a non-hemolytic subclinical PNH clone and was managed with observation and close monitoring of blood counts.
The clinical characteristics from both cases are summarized below in Table 1.
3. Discussion
Epidemiological studies estimate the annual incidence of AA in 2019 to be around 2.0 per million in Western countries and higher in Asia (3.0-5.0 per million) [8]. Comparatively, the annual incidence for PNH in the US is 5.7 per million [9]. PNH can affect any age-group; however, in the US, the most affected age group is between third and fifth decades. At the time of writing, incidence rates of AA and PNH in the post-COVID-19 era remain unknown. Several studies have reviewed the hematologic manifestations of COVID-19, and the most frequently observed laboratory findings include lymphopenia, neutrophilia, anemia, and thrombocytopenia [2,10,11]. However, as we reported here, some patients have a predilection towards acquiring bone marrow failure syndromes after COVID-19 infection. A brief compilation of such cases is presented here in Table 2. Although the association with SARS-CoV-2 infection seems temporal and, in some instances, incidental given the wide prevalence of the virus, an increasing number of new AA and PNH cases related to COVID-19 warrant further exploration of a viral trigger leading to marrow failure as a sequela.
This emerging relationship between COVID-19, AA, and PNH is being investigated globally. A survey from the UK examined the emergence of AA in patients recovering from COVID-19 infection and discovered 3 cases of AA (diagnosed as severe or very severe AA) developed a few weeks after positive SARS-CoV-2 result, 2 cases of AA relapse (as confirmed with marrow hypocellularity), and 15 cases of hematologic decline in known AA which required treatment, transfusion support, and monitoring [15]. Additionally, there has been an increase in the number of cases reporting exacerbation of PNH with COVID-19 infection since the start of the pandemic: Iannuzzi et al. reported 14 cases of known PNH patients and 7 cases of AA/PNH patients who presented with worsening hemolysis symptoms following infection with SARS-CoV-2 [14]. This underscores the need to include the worsening clinical spectrum in estimating the true incidence of AA/PNH during the COVID-19 pandemic.
A few mechanisms have been proposed regarding the effect of SARS-CoV-2 on the hematopoietic system include direct effects of the virus on the bone marrow, cytokine storm causing immune-mediated damage and direct effect of virus on erythroid precursors [2]. A summary of these mechanisms is illustrated below in Figure 1. Viral infections, such as SARS-CoV-1, MERS-CoV, and SARS-CoV-2, have been shown to induce a pro- inflammatory state wherein activated white blood cells and cytokines are released in a positive feedback loop [8]. Because of this background inflammatory state, immune system hyperactivation results in multiorgan dysfunction [16]. Ratajczak and Kucia (2020) evaluated the role of the Nlrp3-inflammasome complex in generating an inflammatory micro-environment, how the innate immune system interacts with the inflammasome and the secretion of pro-inflammatory cytokines in COVID-19 and HSCs (Figure 1) [17]. Their work demonstrated the overexpression of inflammasome complex in HSCs during an active infectious period and that SARS-CoV-2 virus entry receptor angiotensin-converting enzyme 2 (ACE2) is also expressed on the surface of HSCs [17]. They hypothesize that SARS-CoV-2 may direct transcription of pro-inflammatory mediators in HSCs by spike protein interaction with the ACE2 receptor, and may also induce uncontrolled Nlrp3 inflammasome expression leading to hematopoietic stem cell death via pyroptosis [17,18].
The pro-inflammatory microenvironment generated by COVID-19 also has an role in the immune-mediated acquired aplastic anemia mechanism. The development of abnormal autoimmune responses, including cytotoxic T-cells that activate, expand, and circulate as oligoclones, causing the release of myelosuppressive cytokines and inducing cell death of HSCs and progenitor cells [19]. Nevertheless, the inciting antigens for such T-cell response remain undetermined, and HLA polymorphisms and aberrant expression of T-cell receptor signaling genes may also play a role in T-cell dysfunction [20]. Whether the SARS-CoV-2 virus can act as an inciting antigen remains to be clarified.
PNH clonal expansion is found in almost 50% of patients with immune-mediated acquired aplastic anemia [20]. Clonal expansion can be from intrinsic (such as PNH cells being conferred an intrinsic growth advantage via acquired mutations) or extrinsic (such as an environment targeting the destruction of normal HSCs giving rise to PNH HSC selection) in nature. It must be noted that both mechanisms can also co-occur simultaneously [7]. It has also been proposed that PNH cells have acquired the capacity to evade apoptotic stimuli and inflammatory cytokines and escape the HSC-directed immune attack as found in aplastic anemia [20].
Moreover, if SARS-CoV-2 catalyzes an immunologic attack against bone marrow progenitors, these cases illustrate the variability in the virus’ ability to cause PNH clonal expansion and foster an autoimmune marrow environment (Figure 2). Here, case 1 demonstrated a larger PNH clone population leading to the hemolytic presentation, and case 2 was found to have a small amount of PNH clones. Although the reason for this variability can be multifactorial, a reduced immunologic attack should correspond to lower levels of PNH clone expansion [20]. Whether the temporal relationship between somatic PIGA mutation and immunologic attack (i.e., COVID-19 infection) determines the degree of clone expansion, as well as the effects of COVID vaccination and/or antiviral treatment remains an open question for future studies.
4. Conclusions
In summary, we report two cases of new-onset PNH/aplastic anemia syndrome and aplastic anemia associated with COVID-19. It is possible that SARS-CoV-2 does plays a role in the development of PNH clonal expansion and bone marrow failure, but the exact mechanism is still unknown. Regardless, aplastic anemia is a common ground, perhaps stimulating the formation of clonal populations. Although the association in both cases are temporal, and that COVID-19 may be an incidental diagnosis, the growing evidence related to hematological effects of SARS-CoV-2 infection highlights the need for further investigation of this phenomenon.
Funding
This research received no external funding.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Acknowledgments
Figures 1 and 2 were created with BioRender.com.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jha, P.; Brown, P. E.; Ansumana, R. Counting the global COVID-19 dead. Lancet 2022, 399(10339), 1937–1938. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Niloofa, R.; Jayarajah, U.; De Mel, S.; Abeysuriya, V.; Seneviratne, S. L. Hematological Abnormalities in COVID-19: A Narrative Review. Am J Trop Med Hyg 2021, 104. [Google Scholar] [CrossRef] [PubMed]
- Hines, A.; Hakim, N.; Barrientos, J. COVID-19 infection presenting as paroxysmal nocturnal hemoglobinuria. Clin Case Rep 2021, 9 (8), e04636. [Google Scholar] [CrossRef]
- Lee, N. C. J.; Patel, B.; Etra, A.; Bat, T.; Ibrahim, I. F.; Vusirikala, M.; Chen, M.; Rosado, F.; Jaso, J. M.; Young, N. S.; et al. SARS-CoV-2 infection associated with aplastic anemia and pure red cell aplasia. Blood Adv 2022, 6, 3840–3843. [Google Scholar] [CrossRef]
- Otieno, S. B.; Altahan, A.; Kaweeta, F.; Karri, S.; Alnoor, F.; Johnson, R. Severe Hemolysis in a COVID-19 Patient with Paroxysmal Nocturnal Hemoglobinuria. Case Rep Hematol 2021, 2021, 6619177. [Google Scholar] [CrossRef]
- Sumbly, V.; Siddiqui, R.; Alshamam, M.; Kurbanova, T.; Rizzo, V. New Onset Aplastic Anemia after a COVID-19 Infection: A Case Report. American Journal of Medical Case Reports 2021, 9, 451–455. [Google Scholar]
- Hill, A.; DeZern, A. E.; Kinoshita, T.; Brodsky, R. A. Paroxysmal nocturnal haemoglobinuria. Nat Rev Dis Primers 2017, 3, 17028. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X. F.; He, H. L.; Wang, S. Q.; Tang, J. Y.; Han, B.; Zhang, D. H.; Wu, L. Q.; Wu, D. P.; Li, W.; Xia, L. H.; et al. Current Treatment Patterns of Aplastic Anemia in China: A Prospective Cohort Registry Study. Acta Haematol 2019, 142, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Jalbert, J. J.; Chaudhari, U.; Zhang, H.; Weyne, J.; Shammo, J. M. Epidemiology of PNH and Real-World Treatment Patterns Following an Incident PNH Diagnosis in the US. Blood 2019, 134 (Supplement_1), 3407–3407. [Google Scholar] [CrossRef]
- Guan, W. J.; Ni, Z. Y.; Hu, Y.; Liang, W. H.; Ou, C. Q.; He, J. X.; Liu, L.; Shan, H.; Lei, C. L.; Hui, D. S. C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N Engl J Med 2020, 382, 1708–1720. [Google Scholar] [CrossRef]
- Taneri, P. E.; Gómez-Ochoa, S. A.; Llanaj, E.; Raguindin, P. F.; Rojas, L. Z.; Roa-Díaz, Z. M.; Salvador, D.; Groothof, D.; Minder, B.; Kopp-Heim, D.; et al. Anemia and iron metabolism in COVID-19: a systematic review and meta-analysis. Eur J Epidemiol 2020, 35, 763–773. [Google Scholar] [CrossRef]
- Hock, H.; Kelly, H. R.; Meyerowitz, E. A.; Frigault, M. J.; Massoth, L. R. Case 31-2021: A 21-Year-Old Man with Sore Throat, Epistaxis, and Oropharyngeal Petechiae. N Engl J Med 2021, 385, 1511–1520. [Google Scholar] [CrossRef]
- Chakravarthy, R.; Murphy, M. L.; Ann Thompson, M.; McDaniel, H. L.; Zarnegar-Lumley, S.; Borinstein, S. C. SARS-CoV-2 infection coincident with newly diagnosed severe aplastic anemia: A report of two cases. Pediatr Blood Cancer 2022, 69 (4), e29433. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, A.; Parrella, A.; De Ritis, F.; Cammarota, A.; Berloco, L.; Paudice, F.; D'Angelo, G.; Aliberti, E.; Iannuzzo, G. Pancytopenia in a Case of Aplastic Anaemia/Paroxysmal Nocturnal Haemoglobinuria Unmasked by SARS-CoV-2 Infection: A Case Report. Medicina (Kaunas) 2022, 58. [Google Scholar] [CrossRef] [PubMed]
- Avenoso, D.; Marsh, J. C. W.; Potter, V.; Pagliuca, A.; Slade, S.; Dignan, F.; Tholouli, E.; Mittal, S.; Davis, B.; Tauro, S.; et al. SARS-CoV-2 infection in aplastic anemia. Haematologica 2022, 107, 541–543. [Google Scholar] [CrossRef]
- Fara, A.; Mitrev, Z.; Rosalia, R. A.; Assas, B. M. Cytokine storm and COVID-19: a chronicle of pro-inflammatory cytokines. Open Biol 2020, 10 (4), e29433. [Google Scholar] [CrossRef]
- Ratajczak, M. Z.; Kucia, M. SARS-CoV-2 infection and overactivation of Nlrp3 inflammasome as a trigger of cytokine "storm" and risk factor for damage of hematopoietic stem cells. Leukemia 2020, 34, 1726–1729. [Google Scholar] [CrossRef] [PubMed]
- Kucia, M.; Ratajczak, J.; Bujko, K.; Adamiak, M.; Ciechanowicz, A.; Chumak, V.; Brzezniakiewicz-Janus, K.; Ratajczak, M. Z. An evidence that SARS-Cov-2/COVID-19 spike protein (SP) damages hematopoietic stem/progenitor cells in the mechanism of pyroptosis in Nlrp3 inflammasome-dependent manner. Leukemia 2021, 35, 3026–3029. [Google Scholar] [CrossRef]
- Zeng, Y.; Katsanis, E. The complex pathophysiology of acquired aplastic anaemia. Clin Exp Immunol 2015, 180, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Fattizzo, B.; Ireland, R.; Dunlop, A.; Yallop, D.; Kassam, S.; Large, J.; Gandhi, S.; Muus, P.; Manogaran, C.; Sanchez, K.; et al. Clinical and prognostic significance of small paroxysmal nocturnal hemoglobinuria clones in myelodysplastic syndrome and aplastic anemia. Leukemia 2021, 35, 3223–3231. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Mechanisms of SARS-CoV-2 on hematopoietic cells. The SARS-CoV-2 virus has two primary mechanisms: direct viral entry (above, inset) and through inciting an immune response (below). Direct viral entry, via ACE2 receptors, may upregulate Nlrp3 inflammasome expression leading to pyroptosis, viral replication within HSCs, and apoptosis. Immune response causes activation of T cells which respond against HSCs and subsequent immune attack, leading to proliferation of inflammatory signals and cytokine storm. HSC: hematopoietic stem cell, ACE2: angiotensin converting enzyme 2, Nlrp3: NOD-like receptor family pyrin domain containing 3.
Figure 1.
Mechanisms of SARS-CoV-2 on hematopoietic cells. The SARS-CoV-2 virus has two primary mechanisms: direct viral entry (above, inset) and through inciting an immune response (below). Direct viral entry, via ACE2 receptors, may upregulate Nlrp3 inflammasome expression leading to pyroptosis, viral replication within HSCs, and apoptosis. Immune response causes activation of T cells which respond against HSCs and subsequent immune attack, leading to proliferation of inflammatory signals and cytokine storm. HSC: hematopoietic stem cell, ACE2: angiotensin converting enzyme 2, Nlrp3: NOD-like receptor family pyrin domain containing 3.
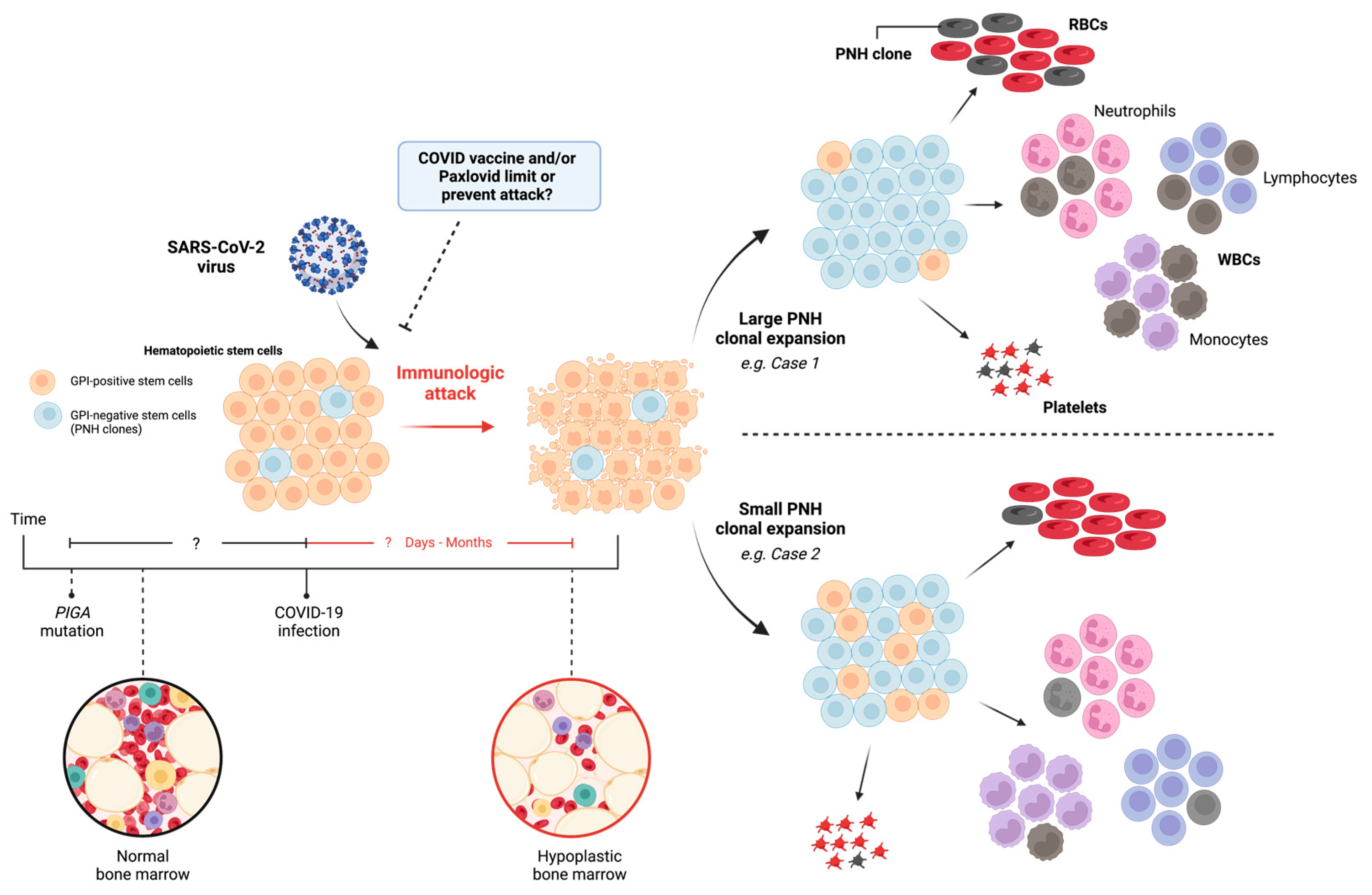
Figure 2.
Implications of SARS-CoV-2 virus: immunologic attack and PNH clonal expansion. A normal bone marrow, with or without PIGA mutation, encounters SARS-CoV-2 virus leading to immunologic attack. Direct or indirect mechanisms may cause HSC death, resulting in variable PNH clonal expansion and bone marrow hypoplasia. RBC: Red blood cell, WBC: White blood cell, PNH: paroxysmal nocturnal hemoglobinuria, GPI: glucose phosphate isomerase, PIGA: Phosphatidylinositol N-acetylglucosaminyltransferase subunit A.
Figure 2.
Implications of SARS-CoV-2 virus: immunologic attack and PNH clonal expansion. A normal bone marrow, with or without PIGA mutation, encounters SARS-CoV-2 virus leading to immunologic attack. Direct or indirect mechanisms may cause HSC death, resulting in variable PNH clonal expansion and bone marrow hypoplasia. RBC: Red blood cell, WBC: White blood cell, PNH: paroxysmal nocturnal hemoglobinuria, GPI: glucose phosphate isomerase, PIGA: Phosphatidylinositol N-acetylglucosaminyltransferase subunit A.
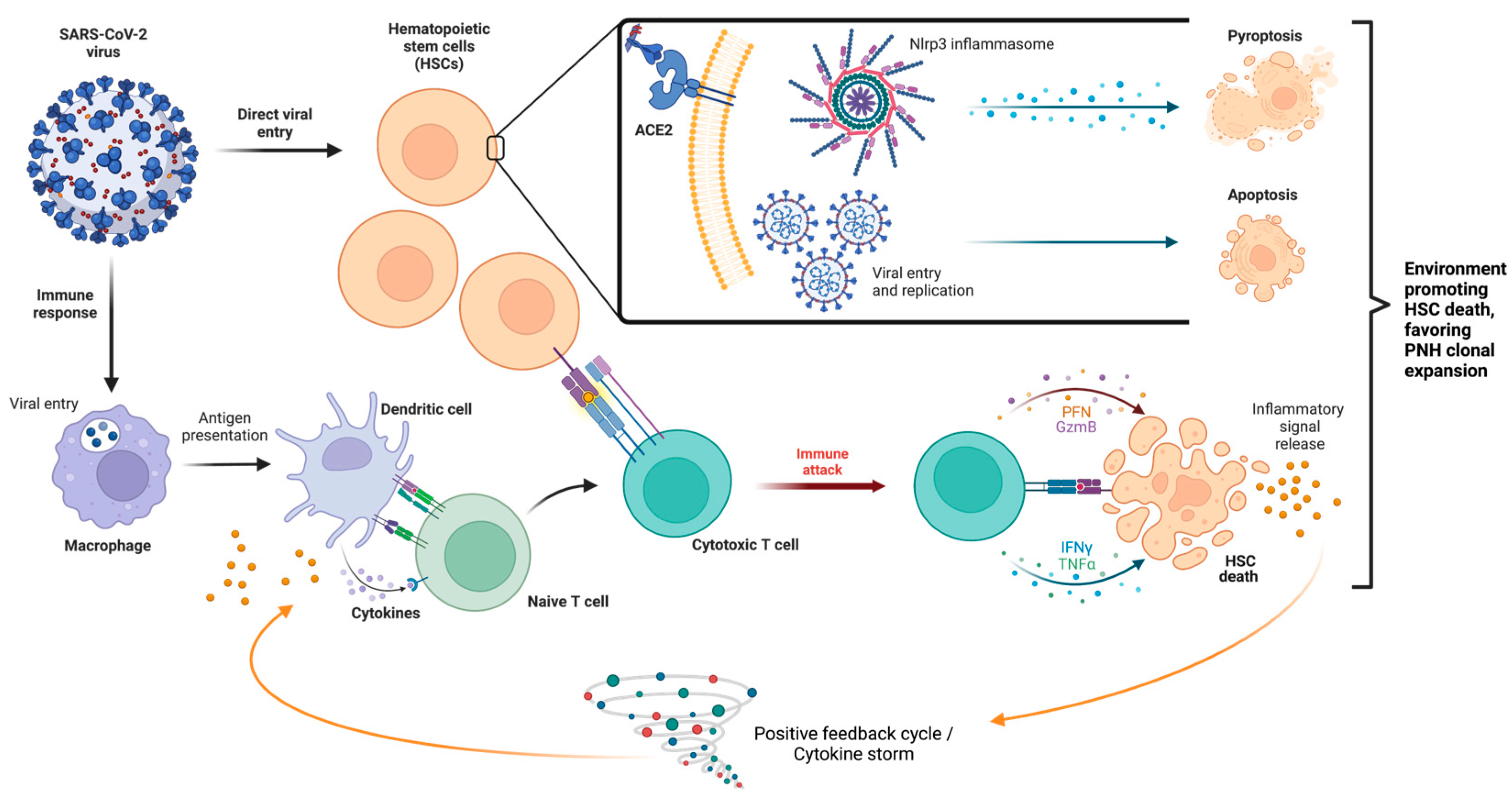

Table 1.
Clinical information for patients with SARS-CoV-2-related aplastic anemia.
| Variable | Patient Case | ||
| 1 | 2 | ||
| Age | 21 | 52 | |
| Sex | Male | Female | |
|
Interval between COVID infection and pancytopenia |
4 months | 1 month | |
| CBC | WBC ANC Platelets HGB MCV |
4.1 1.2 34 9.3 106.2 |
3.0 0.9 18 11.6 94 |
| Bone Marrow Biopsy Cellularity | 10-20% | 10-15% | |
| PNH clones | Granulocytes Monocytes RBCs |
19.53% 19.77% 3.61% |
N/A 0.5% N/A |
| History of autoimmune disease | None | Anterior uveitis | |
| SARS-CoV-2 Vaccination | None | Johnson and Johnson/Janssen Seven months later, Pfizer booster |
|
| Treatment | Ravulizumab | N/A | |
Table 2.
Cases of SARS-CoV-2-related aplastic anemia / PNH.
| Clinical Data | Diagnosis |
Bone Marrow Biopsy Cellularity |
Onset of cytopenia after COVID infection | Therapy | Reference(s) |
| 21, M* | Hemolytic PNH / Aplastic anemia | 10-20% | 4 months | Ravulizumab | This report |
| 52, F* | Aplastic anemia (with subclinical PNH clones) | 10-15% | 1 month | Observation | This report |
| 35, M | Hemolytic PNH | Normal | 0 days ‡ |
Eculizumab, transfusion support |
[3] |
| 22, F | Severe aplastic anemia | 5% | 10 days | Sibling HSCT | [4] |
| 69, F | Severe aplastic anemia | 5-10% | 2 days | Cyclosporine, antithymocyte globulin, eltrombopag | [4] |
| 76, M | Pure red cell aplasia |
20-30% | 4 months | Cyclosporine | [4] |
| 21, M | Severe aplastic anemia (with subclinical PNH clones) |
<5% | < 1 month | Cyclosporine, antithymocyte globulin, eltrombopag; eculizumab† |
[4] |
| 69, F | Severe aplastic anemia (with subclinical PNH clones) |
5% | 5 months | Cyclosporine, antithymocyte globulin, eltrombopag | [4] |
| 19, F | Hemolytic PNH | 40-50% § |
0 days ‡ |
Eculizumab, Ravulizumab | [5] |
| 28, F | Severe aplastic anemia | 20-30% | 3 months | Cyclosporine, antithymocyte globulin, eltrombopag | [4,6] |
| 21, M | Severe aplastic anemia | <5% | 2 months | Sibling HSCT | [12] |
| 12, F | Severe aplastic anemia | 10% | 0 days ‡ |
Antithymocyte globulin, cyclosporine |
[13] |
| 18, M | Severe aplastic anemia | 10% | 0 days ‡ |
Antithymocyte globulin, cyclosporine |
[13] |
| 78, F | Hemolytic PNH / Aplastic anemia | Poor | 0 days ‡ |
-- || |
[14] |
* Our case report. † Received eculizumab due to expansion of PNH clone. ‡ SARS-CoV-2 PCR was positive on presentation; timing between infection and cytopenia onset unclear. § Biopsy also showed large areas less than 10% cellular. || Patient was voluntarily discharged prior to start of any treatment; follow-up unknown.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



