
Submitted:
17 August 2023
Posted:
18 August 2023
You are already at the latest version
Abstract
Oral ferric citrate hydrate (FCH) is effective for iron deficiency in hemodialysis patients.; however, it remains unclear how iron balance in the body affects iron absorption from the intestinal tract. This prospective, observational study (Riona-Oral Iron Absorption Trial, R-OIAT, UMIN 000031406), conducted at 42 hemodialysis centers in Japan, wherein 268 hemodialysis patients without inflammation were enrolled and treated with a fixed amount of FCH for 6 months. We assessed the predictive value of hepcidin-25 for iron absorption and iron shift between ferritin (FTN) and red blood cells (RBCs) following FCH therapy. Serum iron changes at 2 h (ΔFe2h) after FCH ingestion were evaluated as iron absorption. The primary outcome was the quantitative delineation of iron variables on ΔFe2h and the secondary outcome was the description of the predictors of body iron balance. Generalized estimating equations (GEE) were used to identify the determinants of iron absorption during each phase of FCH treatment. ΔFe2h increased when hepcidin-25 and MCH decreased (-0.155, -0.242 to -0.068, p = 0.001; -2.574, -4.421 to -0.726, p = 0.006, respectively) in GEE. Hepcidin-25 increased when erythropoiesis-stimulating agent (ESA) decreased (-0.002, -0.002 to -0.001, p=0.000). FTN increased when RBC and ESA decreased (-0.466, -0.605 to -0.327, p=0.000; -0.002, -0.004 to -0.000, p=0.05, respectively. Limiting erythropoiesis to maintain hemoglobin levels induces RBC reduction in hemodialysis patients, resulting in increased hepcidin-25 and FTN levels. Hepcidin-25 production may prompt an iron shift from RBC-iron to FTN-iron, inhibiting iron absorption even with continued FCH intake.
Keywords:
oral iron absorption
; ferric citrate hydrate
; hemodialysis
; hepcidin-25
; iron shift
1. Introduction
Patients undergoing hemodialysis lose 1.5–3.0 g of iron annually owing to dialysis and periodic laboratory evaluations [1], leading to iron deficiency. Oral supplementation of ferrous iron is inconvenient for iron deficiency anemia in dialysis patients due to gastrointestinal side effects [2], while the use of highly soluble ferric citrate (FC) is increasing because it has fewer side effects [3]. The long-term administration of FC increases ferritin (FTN) and transferrin saturation (TSAT), reduces intravenous iron and erythropoiesis-stimulating agent (ESA) dose requirements, and maintains hemoglobin (Hb) levels in hemodialysis patients [4,5,6]. Nevertheless, it remains unclear what fraction of FC is absorbed, what type of iron state promotes iron absorption, and whether long-term FC administration causes iron overload.
Ferritin is an index of stored iron [7], but its appropriate value in the body remains unknown. There are large differences in FTN levels in hemodialysis patients across countries, with patients in Japan and the USA having the lowest and highest FTN levels, respectively [8]. It is speculated that if a large difference exists in the amount of iron stored, there will be a difference in iron absorption regulated by hepcidin-25; however, the previous FC studies have reported the same phenomena [4,5]. Yokoyama et al. [4]. found that FTN essentially plateaued at week 28, increasing from 57 ng/mL at baseline to 227 ng/mL after 52 weeks, whereas ESA doses gradually decreased during this period, and Lewis et al. [5] reported that FTN levels increased from 593 ng/mL at baseline to 899 ng/mL at 52 weeks, plateauing at 6 months, while the ESA dose decreased in FC studies.
Ferritin correlates strongly and positively with hepcidin-25, an iron-regulatory hormone, as measured using surface-enhanced laser desorption/ionization time-of-flight mass spectrometry, liquid chromatography-tandem mass spectroscopy (LC-MS/MS), and enzyme-linked immunosorbent assay [9,10,11]. Thus, iron states with high FTN levels were expected to be associated with high hepcidin expression. At higher serum hepcidin-25 levels, hepcidin-25 binds to the iron exporter ferroportin (FPN) on the basolateral surface of duodenal enterocytes, thereby inducing its internalization and degradation, and blocking intestinal iron absorption [12]. However, the details of this process are unclear because iron absorption and serum hepcidin-25 levels were not determined in these studies [4,5,6].
Moreover, whether long-term FC administration induces an iron overload remains unknown. The plateauing of FTN levels at 6 months despite continued FC administration may be related to the strict regulation and saturation of iron absorption [6]. However, in addition to the hepcidin-ferroportin axis and the erythroferrone produced during erythroblast production [13], or guideline-based arbitrary reduction of the ESA dose that regulates red blood cell (RBC) production, no iron-regulatory metabolic mechanism is currently known. Iron metabolism is closely linked to erythropoiesis [14], and inadequate erythropoiesis may disturb iron metabolism. However, the precise mechanisms that regulate iron flow in patients undergoing hemodialysis with ESA therapy are unknown. Hence, we conducted the Riona-Oral Iron Absorption Trial (R-OIAT) using ferric citrate hydrate (FCH; Riona, Torii Pharmaceutical Co. Ltd., Tokyo, Japan) as an analytical, prospective, observational study to intercept the crosstalk between erythropoiesis and iron metabolism using intestinal iron absorption and serum hepcidin-25 as the main indices. In this study, we did not aim to supplement iron but rather to observe iron states in patients who received a fixed amount of FCH as a phosphate binder for 6 months.
2. Results
2.1. Baseline characteristics of the study participants
A total of 268 patients were included in the R-OIAT (Figure 1). Between June 1, 2017, and June 1, 2019, 584 hemodialysis patients using FCH as a phosphate binder were enrolled in the R-OIAT. According to the exclusion criteria, 213 and 78 participants were excluded at M3 and M6, respectively. Number of patients with a serum CRP level > 0.5 mg/dL at M0, M3, and M6 was 103, 37, and 25, respectively. Finally, 268 patients who continued to receive a fixed amount of FCH for 6 months were analyzed. The trial concluded in June 2020. The baseline characteristics of the participants are shown in Table 1. The mean age of the participants was 63.0 years (SD, 11.7), 57% of patients were male, 55.6% received 750 mg of FCH, and 49.6% were injected with darbepoetin alfa.
2.2. Association of changes in clinical indices at different 3-month intervals
The clinical measurements on the day of R-OIAT testing at M0, M3, and M6 were summarized in Table 2. FTN levels increased significantly from 100.7 ng/mL at baseline to 116.7 ng/mL at 24 weeks, while the ESA dose decreased and Hb concentrations remained stable. Effects of the amount of FCH on the changes of iron absorption (ΔFe2h) and iron variables for 6 months were shown in Table 3. In patients receiving 750 mg (n = 149), 1,500 mg (n = 101), or 2,250 mg (n = 18) of FCH, no significant interaction between the amount of FCH and exposure time was observed for all iron variables; that means three independent groups change like each other by time (Table 3). Although ΔFe2h levels were significantly lower in the low-dose group than in the high-dose group, the differences remained mostly unchanged throughout the 6 months. The FTN and hepcidin-25 levels were significantly lower in the low-dose group than in the high-dose group at M0, and the difference was maintained at M3 and M6. In each group, there was significant increase in FTN, Hb, MCH and TSAT levels over 6 months. When RBC ≤ 350 × 104 /μL, cases with Hb exceeding 12 g/dl were only 2 (0.7 %).
2.3. Association of ΔFe2h with iron variables
GEE was used to identify the determinants of ΔFe2h during each phase of FCH treatment, M0, M3 and M6. ΔFe2h was affected by hepcidin-25 and MCH (Table 4). ΔFe2h increases by 0.155 μg/dL per one ng/mL decrease in hepcidin-25 [95%CI: -0.242 to -0.068, p=0.001], and increases by 2.574 μg/dL per one pg/cell decrease in MCH [95%CI: -4.421 to -0.726, p=0.006]. Hb level was excluded because of collinearity with RBC based on the formula; Hb = RBC x MCH. In the three data sets of iron markers per patient at M0, M3, and M6 (n = 804), there were significant negative correlations of ΔFe2h with hepcidin-25 (r = 0.161, p < 0.001, Figure 2A) and MCH (r = 0.196, p < 0.001, Figure 2B).
2.4. Association of hepcidin-25 with iron variables
Predictors for hepcidin-25 were analyzed by GEE using iron variables excluding ΔFe2h (Table 5). Hepcidin-25 was affected by MCH, TSAT, FTN and ESA. Hepcidin decreased by 2.908 ng/ml per one pg/cell decrease in MCH [95%CI: -0.196 to 6.011, p=0.042], by 0.511 ng/ml per one % decrease in TSAT [95%CI: 0.216 to 0.806, p=0.001], by 0.224 ng/ml per one ng/ml decrease in FTN [95%CI: 0.183 to 0.266, p=0.001] and by 0.002 ng/ml per one IU/week increase in ESA [95%CI: - 0.002 to 0.001, p=0.001]. In the three datasets of iron markers per patient at M0, M3, and M6 (n = 804), there were significant negative correlations of hepcidin-25 with ESA (r = 0.201, p =0.000, Figure 3A), FTN (r = 0.646, p =0.000, Figure 3B), TSAT (r = 0.278, p =0.000, Figure 3C) and MCH (r = 0.065, p = 0.063, Figure 3D).
2.5. Patterns of FTN fluctuations
Contrary to expectation, because FTN did not necessarily rise gradually with continuous administration of FCH, the fluctuation pattern of FTN was analyzed. Patients were classified into four groups based on 3-month changes in FTN from M0 to M3 or from M3 to M6 (denoted as ΔFTNM3-M0 and ΔFTNM6-M3, respectively); P-1 (ΔFTNM3-M0: positive, ΔFTNM6-M3: positive; n = 63), P-2 (ΔFTNM3-M0: positive, ΔFTNM6-M3: negative; n = 84), P-3 (ΔFTNM3-M0: negative, ΔFTNM6-M3: positive; n = 84), P-4 (ΔFTNM3-M0: negative, ΔFTNM6-M3: negative; n = 37). FTN gradually increased only in 63 cases (23.5%) during both 3-month intervals (Figure 4) and gradually decreased in 37 cases (13.8%). Interestingly, an increase in FTN corresponded to a significant decrease in RBC and Hb, and vice versa, with mirror-image in each group. Moreover there was a negative correlation between ΔFTNM3-M0 and ΔFTNM6-M3 (r= 0.57, Figure 5A). Eighty four of 147 patients (57.1%) whose ΔFTNM3-M0 were positive, were negative in ΔFTNM6-M3, and 84 of 121patients (69.4%) whose ΔFTNM3-M0 were negative, were positive in ΔFTNM6-M3. Similar negative correlations were seen in hepcidin-25, Hb and RBC between the changes in the first and second 3 months (r =0.53, 0.59, and 0.60, respectively, Figure 5B–D).
2.6. Background of changes in FTN
Predictors for FTN were also analyzed by GEE using iron variables excluding ΔFe2h (Table 5). FTN was affected by hepcidin-25, RBC and ESA. FTN increased by 0.775 ng/ml per one ng/mL increase in hepcidin-25 [95%CI: 0.579 to 0.971 p=0.000], by 0.466 ng/ml per one x104/μl decrease in RBC [95%CI: -0.605 to -0.327, p=0.000], by 0.002 ng/ml per one IU/week decrease in ESA [95%CI: - 0.004 to 0.000, p=0.050].
To analyze the effect of the change in RBC counts on FTN values over 3 months, levels at M0 and M3 were treated as starting-points for the first and second 3-month periods, and levels at M3 and M6 were as 3 month-points, respectively. Two data sets of iron variables were evaluated equally and the changes in iron variables over 3 months were represented as Δ3M (n=536). Each data was classified into 4 groups, G-1, G-2, G-3, and G-4, according to the RBC value at start point M0 0r M3; RBC ≤ 300, 300 < RBC ≤ 350, 350 < RBC ≤ 400, and RBC > 400 x 104/μl, respectively. Furthermore, each case was classified into G-a when each ΔRBC3M was negative and G-b when positive. The start and end points were vectorized as the mean values of MCH and RBC (Figure 6A). In any RBC region, changes in ferritin for 3 months in each group decreased in RBC increase group (a) and increased in RBC decrease group (b) (Figure 6B).
3. Discussion
The features of this study are that participants were taking a fixed amount of FCH without changing the dose for 6 months and had no inflammation that affected hepcidin-25 expression via IL6R [15]. Under these conditions, hepcidin-25 and MCH were the strongest inverse explanatory factors for intestinal iron absorption based on GEE. At the molecular level, hepcidin-25 has been shown to control the distribution density of FPN on cell membrane surface [16] and intestinal divalent metal-iron transporter 1expression [17], leading to the influx of iron from the intestinal epithelial cells into the blood. The results of this study reconfirmed that the clinical measurement of hepcidin-25 would be a great tool for estimating iron absorption in clinical practice as well. Similar phenomenon may be seen also in hepatocytes and macrophages which are bearing FPN on membranes [18].
MCH was also an explanatory factor for iron absorption. MCH is calculated by dividing Hb by RBC and indicates the averaged amount of Hb including in one RBC. MCH has an upper limit of 33 [19,20], up to which erythroblasts can take up iron for Hb synthesis, so MCH could be taken as an indicator of iron storage capacity in RBC. It is speculated that when the level of MCH is low, meaning large reserve-iron storage capacity in RBC, serum iron may be readily supplied to erythroblasts and iron absorption from the intestinal tract would increase to replenish it. Erythroblasts require large amounts of iron for hemoglobin synthesis; therefore, they express very high levels of TfR1 [21]. Although MCH has received less attention in the assessment of iron metabolism, it could be used to infer the reserve storage capacity of iron in the RBC.
FTN was not an explanatory factor of iron absorption, although Eshbach et al [22] reported that food iron absorption is increasing when the serum ferritin is below 30 ng/ml, decreasing when more than 100ng/ml by evaluating total body isotope activity at 2 weeks after ingestion of extrinsic tag of radioiron salt added to a meal. Since the amount of iron in blood is as low as 2-3 mg [23], it is speculated that a decrease in serum iron concentration and hepcidin-25 due to rapid iron consumption by erythropoiesis after ESA administration [24] may occur regardless of the value of stored iron, FTN. Given that FPN is the only known iron transporter currently, the rapid decrease in hepcidin-25 levels after ESA administration in hemodialysis patients, which means expansion of erythropoiesis, may be immensely advantageous for iron absorption in the intestine.
ESA and FTN were explanatory factors for hepcidin-25 in addition to TSAT, which is an index of serum iron concentration that stimulates TfR2 in hepatocytes to induce expression of hepcidin [25]. In our previous study [10], serum iron, hepcidin-25, and FTN levels rapidly decreased after ESA administration and then recover during the interval to next ESA administration. Chaston et al. [16] reported that the FPN response to hepcidin-25 differs among cells, e.g., reticuloendothelial macrophages, hepatocytes, and enterocytes, and that a rapid increase in hepcidin-25 reduces macrophage FPN expression after 2 hours, but enterocyte FPN reduction appears only when hepcidin-25 levels have remained continuously high for 24 hours. These phenomena may be responsible for the complicated relationship between iron absorption and hepcidin-25 and these reactions cannot be captured by one-time measurements. Time-course measurements should be included in future studies.
In the present study, predictors of FNT were analyzed by GEE because FTN has been often used to assess body iron stores [26]. Hepcidin-25 was a positive predictor, and RBC and ESA were negative predictors of FTN. These findings may be due to the arbitrary adjustment of ESA, such that Hb could be maintained at 10‒12 g/dL according to clinical guidelines [27,28]. Approximately 80% of the body’s iron is found in RBCs [29], with 90% of daily iron consumption used by erythroblasts to synthesize Hb [30]. The erythroid system maintains iron homeostasis as an iron reservoir [30] whereby iron is conserved and recycled in the body [31]. In a closed system of iron metabolism, limiting erythropoiesis by reducing ESA doses could induce an increase in serum iron, followed by hepcidin-25, inhibit delivery of iron to the blood by macrophages, and concomitantly suppress intestinal iron absorption. As a result, ferritin increases in macrophages that have phagocytized senescent erythrocytes. This phenomenon appears to be a shift from erythrocyte-iron to ferritin-iron or vice versa, which resembles the function of bellows in closed systems (Figure 7). Our findings provide an insight into the reasons why the rate of increase in FTN is reduced at 24 weeks and why it reaches a plateau despite continuous FC exposure for 52 weeks [4,5,6]. When Hb level is ≥ 12 g/dL, ESA is arbitrarily reduced and Hb levels is adjusted between 10 and 12 g/dL according to the guidelines. As shown in Figure 8, Hb level, which was 13 g/dL in RBC 450 × 104 /μL (MCH 28.9 pg, point A), was reduced to 11.6 g/dL when reducing ESA and resetting RBC to 400 × 104 /μL (point B). During this period, the FTN level increased because the RBC-iron capacity decreased and the corresponding amount of iron shifted to FTN iron. When FCH administration was continued, MCH increased with iron absorption, and Hb level easily exceeded 12 g/dL (RBC to 400 × 104 /μL, MCH 32.5 pg, point C). If ESA was reduced again and RBC reset to 350 × 104 /μL, Hb level improved to 11.4 g/dL (point D) with a re-elevation in the FTN value. When RBC is ≤ 350 × 104 /μL, there is little risk of Hb being ≥ 12 g/dL even if FCH therapy is continued, because there is an upper limit against MCH, which falls normally within the range 27–33 pg [19,20]. At this stage, there is no need to further reduce the ESA, and the FTN reaches a plateau without rising. Such a series of reactions may occur regardless of the FTN value at baseline, such as 227 ng/mL in Japan and 899 ng/mL in the United States [4,5,6]. Our concern with FCH therapy is whether the long-term use of FC may cause iron overload. These findings suggest that setting RBC to 300‒350 × 104/μL initially, followed by oral iron supplementation, in which Hb levels do not exceed 12 g/dL, would minimize fluctuations in FTN. Whether such a procedure will reduce the requirement for ESA and prevent iron overload should be clarified in future studies.
The upper limit of hepcidin-25 production in terms of the relationship with FTN has not been reported previously. We showed that physiological concentrations of hepcidin-25 have an upper limit at more than 100 ng/ml FTN. Similar trends have been observed in myelodysplastic syndromes [32], thalassemia [33], and dialysis patients [34], but there was no description of the upper limit of hepcidin in these papers. When cellular labile iron pool (LIP) increases, FTN expression is stimulated via iron responsive elements and iron regulatory proteins at the translational level, and the ferrous iron of LIP is mobilized to the FTN as ferric iron to reduce the risk of radical development in cytoplasm [7]. On the other hand, elevated LIP enhances the expression of bone morphogenetic protein 6 (BMP6) in hepatic non-parenchymal cells, which binds to hepatocyte BMP receptor in paracrine and stimulates hepcidin expression via the BMP6/SMAD pathway to prevent iron export via FPN to blood [35]. If it is hypothesized that there is an upper limit on the levels of LIP to prevent cytoplasm from radical exposures, BMP6 and hepcidin-25 in its downstream will also have an upper limit and their serum levels will fluctuate according to LIP. When FTN remains undegraded in cytoplasm, levels of ferritin might increase cumulatively without an upper limit. In myelodysplastic syndromes, patients with more than 10,000 ng/ml of ferritin were reports [36]. Hepcidin-25 may be a predictor of LIP and BMP6, and cumulative increase in FTN may indicate that the LIP has repeatedly reached the upper limit. This may be the reason why FC was effective regardless of the FTN levels in previous studies [4,5,6]. Although we targeted only cases without inflammation, the difference from the hepcidin synthesis via STAT3 stimulated by IL-6 during inflammation should be examined in the future studies [31].
In conclusion, our data suggested that a key molecule for facilitating the crosstalk between hematopoietic and iron metabolic systems to keep the balance between iron consumption from serum and iron supply to serum was hepcidin-25, and ESA was shown to be a trigger to cause an iron shift between body stored iron and RBC stored iron. The limiting erythropoiesis was one of the causes of FTN increase in hemodialysis patients undergoing FCH therapy, in addition to iron absorption from the intestinal tract. A major factor altering iron-related factors in dialysis patients may be the fluctuation of RBCs, which have large capacities for iron storage. If the RBC count is maintained below 350 × 104/µL, iron overload would not occur even with a sustained oral supply of iron. Although our results do not provide evidence for an adequate RBC number, our study provides a basis for clinical trials evaluating RBC-restricting strategies to minimize the ESA dose and improve iron metabolism in patients with renal anemia.
4. Materials and methods
4.1. Study design and data sources
The R-OIAT recruited patients from 42 hemodialysis centers in Japan. Eligible subjects (n=584) were adult patients with end stage renal disease on three times per week hemodialysis for at least one month before the R-OIAT trial who had been prescribed 750mg-2250mg doses of FCH as phosphate binder. Eligible hemodialysis patients were not pregnant and had no malignancies or infections. Patients were excluded if they received intravenous iron supplementation, were transferred to a different clinic, ceased using FCH, changed FCH dosage, or had inflammation (C-reactive protein [CRP] level > 0.5 mg/dL). Finally, 268 individuals who received a fixed amount of FCH as a phosphate binder for 6 months were enrolled;750mg (n = 149), 1500mg (n =101), 2250 mg (n = 18), respectively. This study was conducted in accordance with the Declaration of Helsinki and approved by the Kanazawa Medical University Ethics Committee (No. M401). All the participants provided written informed consent.
4.2. Procedures
The patients received FCH three times a day after meals. One tablet of FCH (250 mg) contains approximately 60 mg of ferric iron. The participants consumed their regular diets during the study period. Thus, on the day of the R-OIAT, blood was drawn early in dialysis, and patients received one-third of the total daily FCH dose shortly thereafter. After 2 h, another blood sample was obtained. The patients did not fast before the test, but they did fast until the second sampling after the FCH dose. The R-OIAT sampling was repeated thrice: at baseline (M0), 3 months (M3), and 6 months (M6). Blood samples were centrifuged at 1,500 × g for 10 min, and the serum was immediately frozen at -20 °C. Samples were shipped on dry ice to Kanazawa Medical University to determine serum iron, FTN, unsaturated iron-binding capacity (UIBC), and hepcidin-25 levels. Serum iron and UIBC levels were measured using colorimetry; TSAT was calculated using the following formula: serum iron/total iron-binding capacity × 100. The FTN levels were measured using a chemiluminescent enzyme immunoassay. Serum hepcidin-25 was measured using LC-MS/MS [37] at MCProt Biotechnology (Ishikawa, Japan), with a dynamic range of 0–1,000 ng/mL. ESA doses were calculated on each oral iron absorption trial day. Because patients were taking different ESAs (i.e., epoetin (IU), darbepoetin (μg) or epoetin beta pegol (μg), their units were converted to international unit (IU) based on previously reported data [38]. The conversion for darbepoetin and epoetin beta pegol was based on reports finding that 1 μg is equivalent to 200 IU [39]. Amounts of ESAs were assessed as average weekly doses (IU/week).
4.3. Estimating iron absorption
The amount of iron absorbed after FCH administration (ΔFe2h) was determined by subtracting the serum iron concentration before administration from the serum iron concentration measured 2 h later according to previous studies [40]. In preliminary experiments, ΔFe2h ranged from 0 to 23 mg/dL in 36 control hemodialysis patients without FCH administration, and the median (inter-quartile range, IQR) of ΔFe2h was 0.5 (IQR: 0–7) mg/dL.
4.4. Statistical analysis
Continuous variables were summarized as mean and standard deviation. Categorical variables were described as numbers and relative frequencies (%) of the subjects in each category. Comparisons between patient groups were carried out using one-way analysis of variance (ANOVA) or repeated-measures ANOVA with post hoc Bonferroni’s multiple comparison tests. Two-way repeated-measures ANOVA test (condition [750mg vs 1500mg vs 2250mg of FCH] x time [M0 vs M3 vs M6]) was used to analyze the effects of FCH on iron variables. Univariate correlations between variables were assessed using the Pearson’s correlation test. After adjusting for independent variables, generalized estimating equations (GEE) were used to identify the determinants of iron absorption during each phase of FCH treatment, M0, M3 and M6; the B coefficient with their 95% CI and the corresponding P values < 0.05 were used to declare the determinants of iron absorption. Independent predictors of hhepcidin-25 and FTN were also assessed using GEE. Differences in iron variables between ranges of RBC were compared using independent t-tests for parametric data. All p values were two-tailed, with an α level of < 0.05. Database construction and statistical analyses were performed using Statistical Package for the Social Sciences (SPSS) version 24 (IBM SPSS Inc., Armonk, NY).
Author Contributions
NT and YK conceived the studies and obtained funding. All authors contributed to the design of the trials. NT and YK conducted the studies. NT analysed the data and wrote the first draft of the manuscript. All authors have read and agreed to the final version of the manuscript.
Funding
This work was supported by Kanazawa Medical University grant, Torii Pharmaceutical Co., Ltd. (Tokyo, Japan) grant (primary funding source) and Japan Tobacco Inc. (Tokyo, Japan) grant. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Institute Review Board Statement
This study was conducted in accordance with the Declaration of Helsinki and approved by the Institutional Review Board of the Kanazawa Medical University Ethics Committee (No. M401, approval September 2016).
Informed Consent Statement
Informed consent was obtained from all individual participants included in the study. All the participants provided written informed consent.
Acknowledgments
We would like to thank the participants and the participating nursing staff. All authors contributed to the study design and were responsible for data collection. Dr. Tomosugi and Dr. Koshino conceived the study and obtained funding. Dr Tomosugi analyzed the data and drafted the manuscript. Dr. Koshino created all figures. All authors have contributed to the final version of this manuscript. The corresponding author had full access to all data in the study and had the final responsibility for the decision to submit it for publication.
Conflicts of Interest
Dr. Tomosugi is a professor at Kanazawa Medical University and the president of Medical Care Proteomics Biotechnology Co., Ltd. However, this association did not alter the authors’ adherence to all policies regarding the sharing of data. The authors have declared that no conflict of interest exists.
References
- Fishbane, S.; Spinowitz, B. Update on anemia in ESRD and earlier stages of CKD: core curriculum 2018. Am J Kidney Dis. 2018, 71, 423–35. [CrossRef]
- Besarab, A.; Coyne, D.W. Iron supplementation to treat anemia in patients with chronic kidney disease. Nat Rev Nephrol. 2010, 6, 699–710. [CrossRef]
- Pergola, P.E.; Fishbane, S.; Ganz, T. Novel oral iron therapies for iron deficiency anemia in chronic kidney disease. Adv Chronic Kidney Dis. 2019, 26, 272–91. [CrossRef]
- Yokoyama, K.; Akiba, T.; Fukagawa, M.; Nakayama, M.; Sawada, K.; Kumagai, Y.; Chertow, G.M.; Hirakata H. Long-term safety and efficacy of a novel iron-containing phosphate binder, JTT-751, in patients receiving hemodialysis. J Ren Nutr. 2014, 24, 261–267. [CrossRef]
- Lewis, J.B.; Sika, M.; Koury, M.J.; Chuang, P.; Schulman, G.; Smith, M.T.; Whittier, F.C.; Linfert, D.R.; Galphin, C.M.; Athreya, B.P. et al. Ferric citrate controls phosphorus and delivers iron in patients on dialysis. J Am Soc Nephrol. 2015, 26, 493–503. [CrossRef]
- Umanath, K.; Jalal, D.I.; Greco, B.A.; Umeukeje, E.M.; Reisin, E.; Manley, J.; Zeig, S.; Negoi, D.G.; Hiremath, A.N.; Blumenthal, S.S. et al. Ferric citrate reduces intravenous iron and erythropoiesis-stimulating agent use in ESRD. J Am Soc Nephrol. 2015, 26, 2578–2587. [CrossRef]
- Kühn, L.C. Iron regulatory proteins and their role in controlling iron metabolism. Metallomics. 2015, 7, 232–243. [CrossRef]
- Ramanathan, G.; Olynyk, J.K.; Ferrari, P. Diagnosing and preventing iron overload. Hemodial Int 21(Suppl 1): 2017, S58–S67. [CrossRef]
- Roe, M.A.; Collings, R.; Dainty, J.R.; Swinkels, D.W.; Fairweather-Tait, S.J. Plasma hepcidin concentrations significantly predict interindividual variation in iron absorption in healthy men. Am J Clin Nutr. 2009, 89, 1088–1091. [CrossRef]
- Honda, H.; Kobayashi, Y.; Onuma, S.; Shibagaki, K.; Yuza, T.; Hirao, K.; Yamamoto, T.; Tomosugi, N.; Shibata, T. Associations among erythroferrone and biomarkers of erythropoiesis and iron metabolism, and treatment with long-term erythropoiesis-stimulating agents in patients on hemodialysis. PLoS One 2016, 11, e0151601. [CrossRef]
- Pak, M.; Lopez, M.A.; Gabayan, V.; Ganz, T.; Rivera, S. Suppression of hepcidin during anemia requires erythropoietic activity. Blood 2006, 108, 3730–3735. [CrossRef]
- Gulec, S.; Anderson, G.J.; Collins, J.F. Mechanistic and regulatory aspects of intestinal iron absorption. Am J Physiol Gastrointest Liver Physiol. 2014,307, G397–G409. [CrossRef]
- Kautz, L.; Jung, G.; Valore, E.V.; Rivella, S.; Nemeth, E.; Ganz, T. Identification of erythroferrone as an erythroid regulator of iron metabolism. Nat Genet. 2014, 46, 678-684. [CrossRef]
- Li, H.; Ginzburg, Y.Z. Crosstalk between iron metabolism and erythropoiesis. Adv Hematol. 2010, 2010, 605435. [CrossRef]
- Verga Falzacappa, M.V.; Vujic Spasic, M.; Kessler, R.; Stolte, J.; Hentze, M.W.; Muckenthaler, M.U. STAT3 mediates hepatic hepcidin expression and its inflammatory stimulation. Blood. 2007, 109, 353–358. [CrossRef]
- Chaston, T.; Chung, B.; Mascarenhas, M.; Marks, J.; Patel, B.; Srai, S.K.; Sharp, P. Evidence for differential effects of hepcidin in macrophages and intestinal epithelial cells. Gut 2008, 57, 374–382. [CrossRef]
- Brasse-Lagnel, C.; Karim, Z.; Letteron, P.; Bekri, S.; Bado, A.; Beaumont, C. Intestinal DMT1 cotransporter is down-regulated by hepcidin via proteasome internalization and degradation. Gastroenterology 2011, 140, 1261–1271. [CrossRef]
- Nemeth, E.; Ganz, T. Hepcidin-Ferroportin Interaction Controls Systemic Iron Homeostasis. Int J Mol Sci. 2021, 22, 6493. [CrossRef]
- Wintrobe, M.M. The erythrocyte. In: Wintrobe MM, editor. Clinical hematology. 6 th ed. Philadelphia: Lea & Febiger, 1967, Chap 2: 63–103.
- Vajpayee, N.; Graham, S.S.; Bem, S. Basic examination of blood and bone marrow. In: McPherson RA, Pincus MR, editor. Henry’s clinical diagnosis and management by laboratory methods. 22nd ed. Philadelphia: WB Saunders, 2011, Chap 30: 509-35.
- Kawabata, H. Transferrin and transferrin receptors update. Free Radic Biol Med. 2019, 133, 46-54. [CrossRef]
- Eschbach, J.W.; Cook, J.D.; Scribner, B.H.; Finch, C.A. Iron balance in hemodialysis patients. Ann Intern Med. 1977, 87, 710-713. [CrossRef]
- Camaschella, C.; Nai, A.; Silvestri L. Iron metabolism and iron disorders revisited in the hepcidin era. Haematologica. 2020, 105, 260-272. [CrossRef]
- Egrie, J.C.; Strickland, T.W.; Lane, J.: Aoki, K.; Cohen, A.M.; Smalling, R.; Trail, G.; Lin, F.K.; Browne, J.K.; Hines, D.K. Characterization and biological effects of recombinant human erythropoietin. Immunobiology 1986, 172, 213–224. [CrossRef]
- Kawabata, H..; Fleming, R.E.; Gui, D.; Moon, S.Y.; Saitoh, T.; O'Kelly, J.; Umehara, Y.; Wano, Y.; Said, J.W.; Koeffler, H.P. Expression of hepcidin is down-regulated in TfR2 mutant mice manifesting a phenotype of hereditary hemochromatosis, Blood 2005,105, 376–381. [CrossRef]
- Daru, J.; Colman, K.; Stanworth, S.J; De La Salle, B.; Wood, E.M.; Pasricha, S.R. Serum ferritin as an indicator of iron status: what do we need to know?Am J Clin Nutr. 2017, 106, 1634S-1639S. [CrossRef]
- Babitt, J.L.; Eisenga, M.F.; Haase, V.H.; Kshirsagar, A.V.; Levin, A.; Locatelli, F.; Małyszko, J.; Swinkels, D.W.; Tarng, D.C.; Cheung, M. et al. Controversies in optimal anemia management: conclusions from a kidney disease: Improving Global Outcomes (KDIGO) Conference. Kidney Int. 2021, 99,1280–1295. [CrossRef]
- Tsubakihara, Y.; Nishi, S.; Akiba, T.; Hirakata, H.; Iseki, K.; Kubota, M.; Kuriyama, S.; Komatsu, Y.; Suzuki, M.; Nakai, S. et al. 2008 Japanese Society for Dialysis Therapy: guidelines for renal anemia in chronic kidney disease. Ther Apher Dial. 2010, 14,240–275. [CrossRef]
- Soe-Lin, S.; Apte, S.S.; Andriopoulos, B.Jr.; Andrews, M.C.; Schranzhofer, M.; Kahawita, T.; Garcia-Santos, D.; Ponka, P. et al. Nramp1 promotes efficient macrophage recycling of iron following erythrophagocytosis in vivo. Proc Natl Acad Sci USA. 2009, 106, 5960–5965. [CrossRef]
- Zhang, D.L.; Ghosh, M.C.; Ollivierre, H.; Li, Y.; Rouault, T.A. Ferroportin deficiency in erythroid cells causes serum iron deficiency and promotes hemolysis due to oxidative stress. Blood 2018, 132, 2078–2087. [CrossRef]
- Coffey, R.; Ganz, T. Iron homeostasis: an anthropocentric perspective. J Biol Chem. 2017, 292, 12727–12734. [CrossRef]
- Santini, V.; Girelli, D.; Sanna, A.; Martinelli, N.; Duca, L.; Campostrini, N.; Cortelezzi, A.; Corbella, M.; Bosi, A.; Reda, G. et al. Hepcidin levels and their determinants in different types of myelodysplastic syndromes. PLOS ONE 2011, 6, e23109. [CrossRef]
- Tantiworawit, A.; Khemakapasiddhi, S.; Rattanathammethee, T.; Hantrakool, S.; Chai-Adisaksopha, C.; Rattarittamrong, E.; Norasetthada, L.; Charoenkwan, P.; Srichairatanakool, S.; Fanhchaksai, K. Correlation of hepcidin and serum ferritin levels in thalassemia patients at Chiang Mai University Hospital. Biosci Rep 2021, 41, BSR20203352. [CrossRef]
- Daimon, S. Efficacy for anaemia and changes in serum ferritin levels by long-term oral iron administration in haemodialysis patients. Ther Apher Dial 2019, 23, 444–450. [CrossRef]
- Rausa, M.; Pagani, A.; Nai, A.; Campanella, A.; Gilberti, M.E.; Apostoli, P.; Camaschella, C.; Silvestri, L. Bmp6 expression in murine liver non parenchymal cells: a mechanism to control their high iron exporter activity and protect hepatocytes from iron overload? PLoS One 2015, 10, e0122696. [CrossRef]
- Kawabata, H.; Usuki, K.; Shindo-Ueda, M.; Kanda, J.; Tohyama, K.; Matsuda, A.; Araseki, K.; Hata, T.; Suzuki, T.; Kayano, H. et al. Serum ferritin levels at diagnosis predict prognosis in patients with low blast count myelodysplastic syndromes. Int J Hematol 2019, 110, 533-42. [CrossRef]
- Murao, N.; Ishigai, M.; Yasuno, H.; Shimonaka, Y.; Aso Y. Simple and sensitive quantification of bioactive peptides in biological matrices using liquid chromatography/selected reaction monitoring mass spectrometry coupled with trichloroacetic acid clean-up. Rapid Commun Mass Spectrom. 2007, 21, 4033–4038. [CrossRef]
- Aljama, P.; Bommer, J.; Canaud, B.; Carrera, F.; Eckardt, K.U.; Hörl, W.H.; Krediet, R.T.; Locatelli, F.; Macdougall, I.C.; Wikström, B. Practical guidelines for the use of NESP in treating renal anaemia. Nephrol Dial Transpl. 2001,16 (Suppl 3), 22-28. [CrossRef]
- Carrera, F.; Lok, C.E.; de Francisco, A.; Locatelli, F.; Mann, J.F.; Canaud, B.; Kerr, P.G.; Macdougall, I.C.; Besarab, A.; Villa, G. et al. Maintenance treatment of renal anaemia in haemodialysis patients with methoxy polyethylene glycolepoetin beta versus darbepoetin alfa administered monthly: a randomized comparative trial. Nephrol Dial Transpl. 2010, 25, 4009-4017. [CrossRef]
- Conway, R.E.; Geissler, C.A.; Hider, R.C.; Thompson, R.P.; Powell, J.J. Serum iron curves can be used to estimate dietary iron bioavailability in humans. J Nutr. 2006, 136, 1910–1914. [CrossRef]
Figure 1.
Flow chart depicting the study population selection.
Figure 2.
Correlation between ΔFe2h and hepcidin-25, and MCH. Correlation between ΔFe2h and hepcidin-25 (A), and MCH (B). The data included all samples from 268 patients at M0, M3, and M6 during FCH therapy for 6 months (n = 804). ΔFe2h, iron absorption; MCH, mean corpuscular hemoglobin.
Figure 2.
Correlation between ΔFe2h and hepcidin-25, and MCH. Correlation between ΔFe2h and hepcidin-25 (A), and MCH (B). The data included all samples from 268 patients at M0, M3, and M6 during FCH therapy for 6 months (n = 804). ΔFe2h, iron absorption; MCH, mean corpuscular hemoglobin.
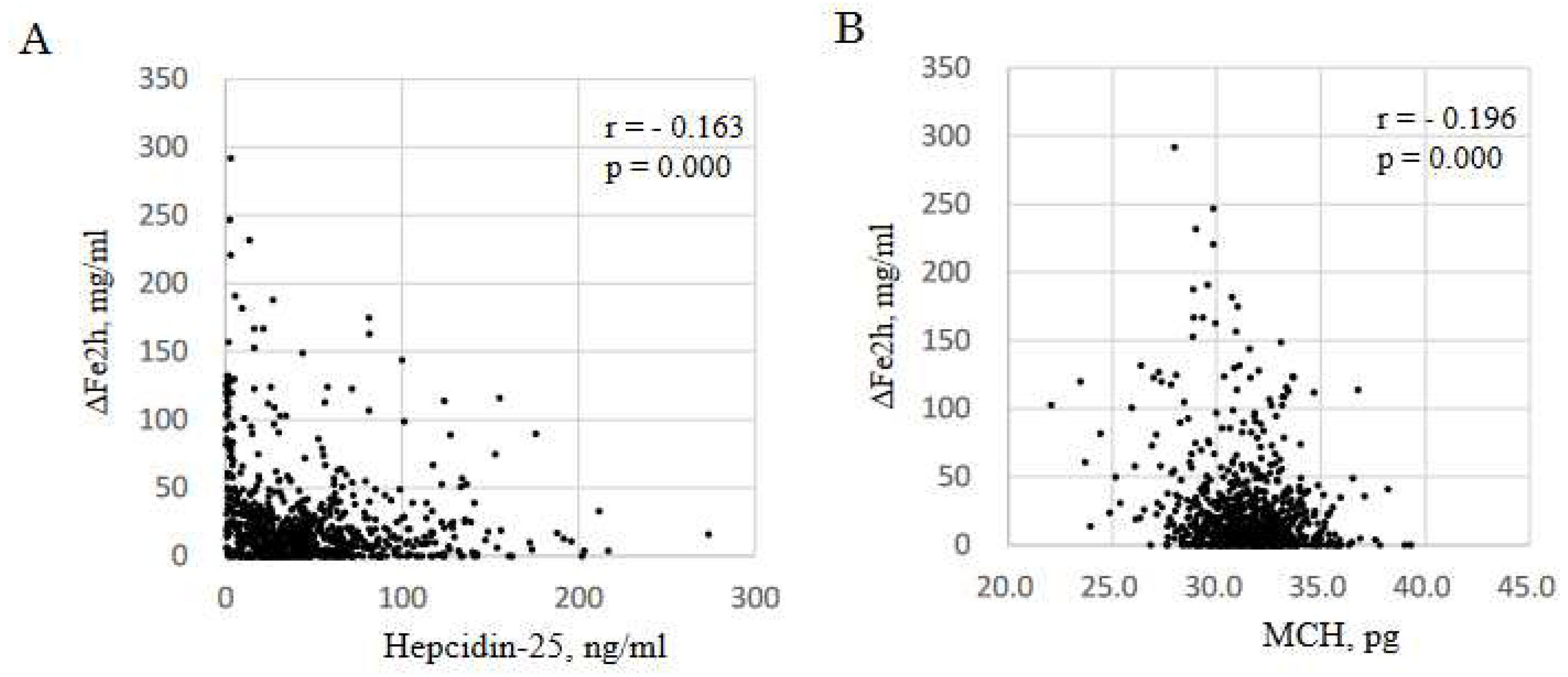
Figure 3.
Correlation between hepcidin-25 and iron markers. Correlation between hepcidin-25 and ESA (A), ferritin (B), MCH (C) and TSAT (D). Data included all samples from 268 patients at M0, M3 and M6 during FCH therapy for 6 months (n = 804). ESA, erythropoiesis-stimulating agent; MCH, mean corpuscular hemoglobin; TSAT, transferrin saturation.
Figure 3.
Correlation between hepcidin-25 and iron markers. Correlation between hepcidin-25 and ESA (A), ferritin (B), MCH (C) and TSAT (D). Data included all samples from 268 patients at M0, M3 and M6 during FCH therapy for 6 months (n = 804). ESA, erythropoiesis-stimulating agent; MCH, mean corpuscular hemoglobin; TSAT, transferrin saturation.
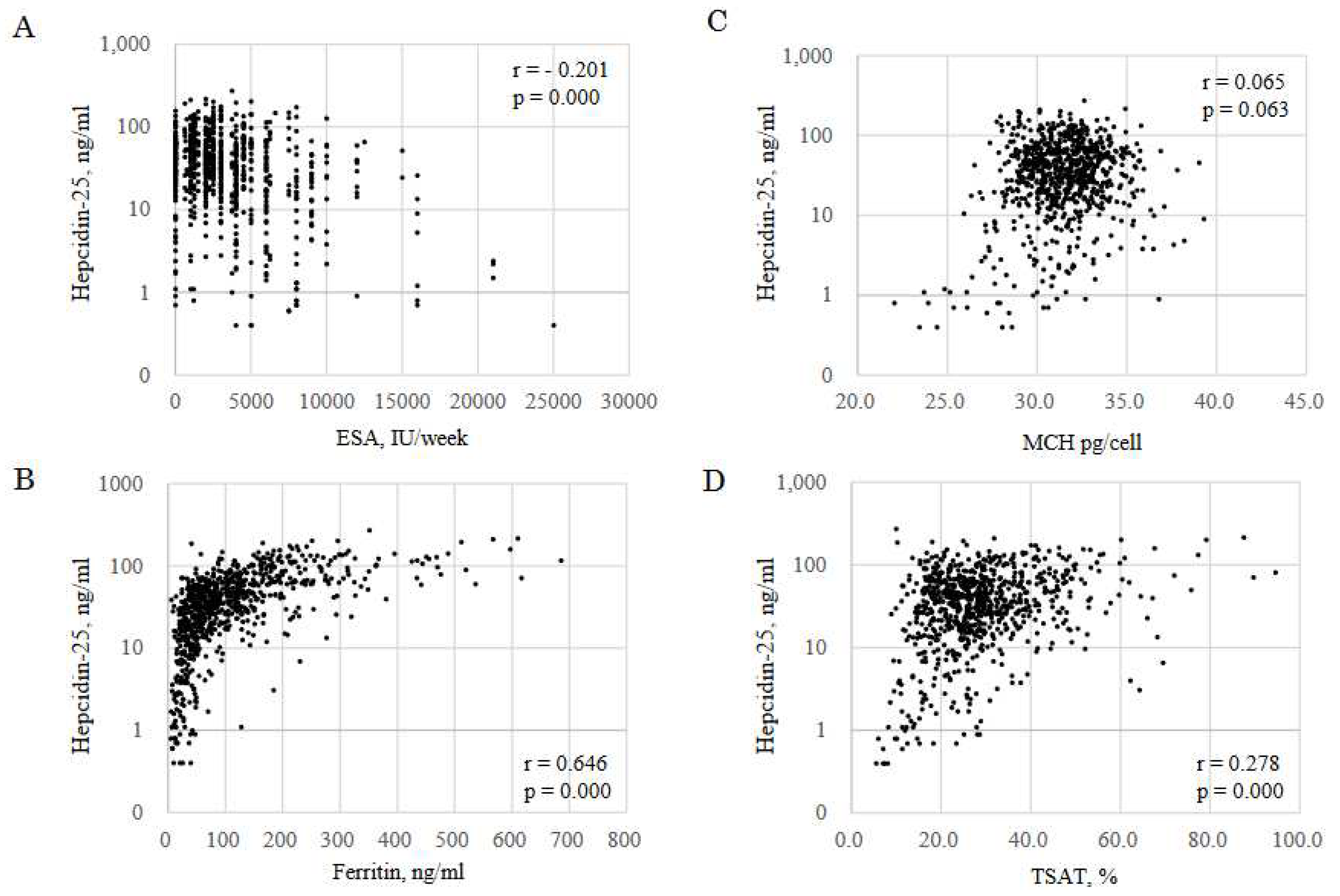
Figure 4.
Changes of FTN and iron variables. Patients were classified into 4 groups based on positive or negative of ΔFTNM3-M0 and ΔFTNM6-M3; P-1(positive, positive), P-2(positive, negative), P-3(negative, positive), P-1(negative, negative), (n=268). FTN, ferritin; RBC, red blood cell; Hb, hemoglobin; MCH, mean corpuscular hemoglobin. The levels of iron variables at M0 were compared with those at M3 and M6. *P<0.05. **P<0.001.
Figure 4.
Changes of FTN and iron variables. Patients were classified into 4 groups based on positive or negative of ΔFTNM3-M0 and ΔFTNM6-M3; P-1(positive, positive), P-2(positive, negative), P-3(negative, positive), P-1(negative, negative), (n=268). FTN, ferritin; RBC, red blood cell; Hb, hemoglobin; MCH, mean corpuscular hemoglobin. The levels of iron variables at M0 were compared with those at M3 and M6. *P<0.05. **P<0.001.
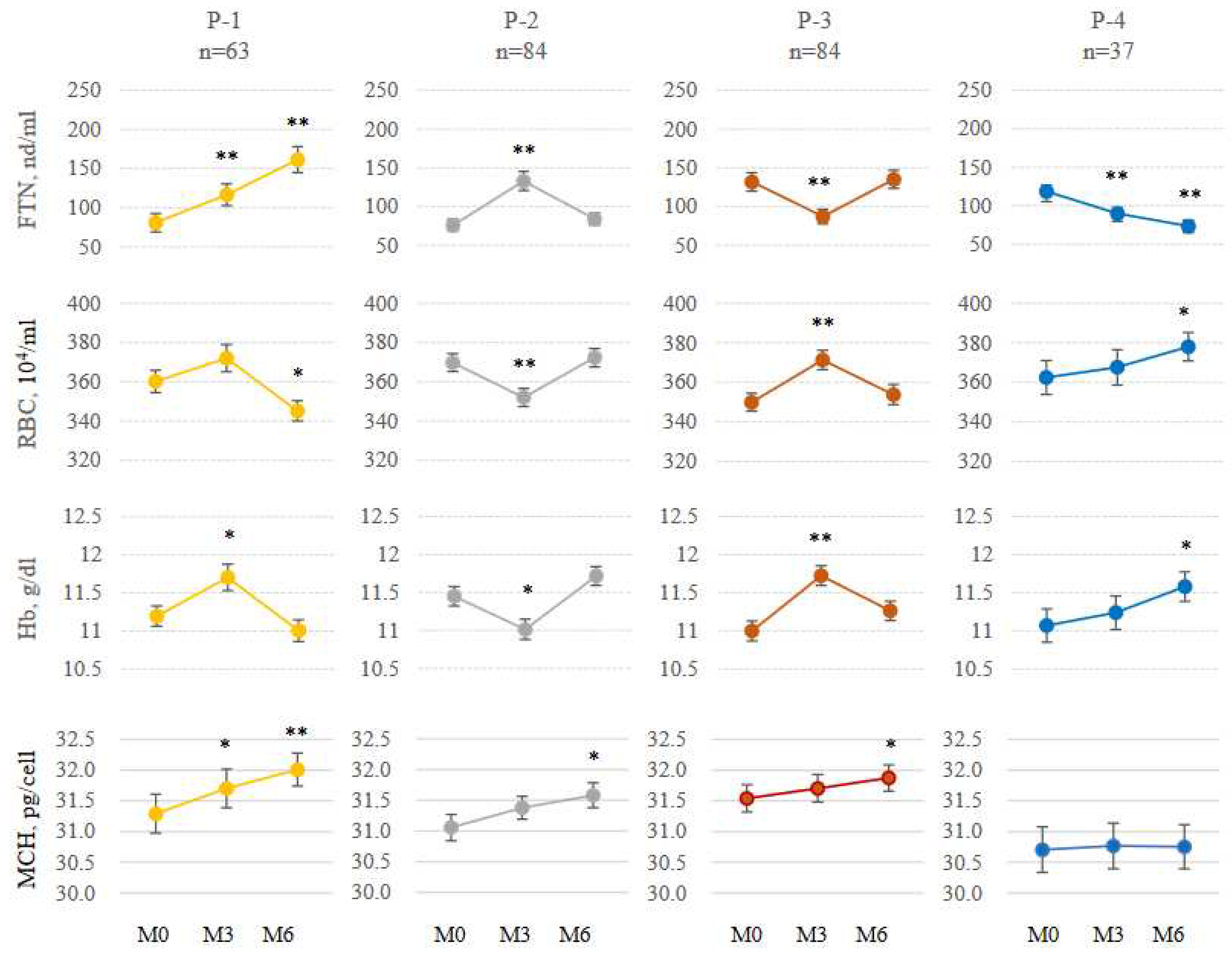
Figure 5.
Inverse correlations between the changes of iron variables from M0 to M3, and those from M3 to M6 (n=268). A: ΔFTNM3-M0 vs ΔFTNM6-M3, B: ΔRBC M3-M0 vs ΔRBC M6-M3, C: ΔHb M3-M0vs ΔHb M6-M3, D: ΔHEP M3-M0 vs Δhepcidin-25 M6-M3. Only 63 cases (23.5%) were positive for both ΔFTN M3-M0 and Δ FTN M6-M3. ΔM3-M0: changes from M0 to M3, ΔM6-M3: changes from M3 to M6
Figure 5.
Inverse correlations between the changes of iron variables from M0 to M3, and those from M3 to M6 (n=268). A: ΔFTNM3-M0 vs ΔFTNM6-M3, B: ΔRBC M3-M0 vs ΔRBC M6-M3, C: ΔHb M3-M0vs ΔHb M6-M3, D: ΔHEP M3-M0 vs Δhepcidin-25 M6-M3. Only 63 cases (23.5%) were positive for both ΔFTN M3-M0 and Δ FTN M6-M3. ΔM3-M0: changes from M0 to M3, ΔM6-M3: changes from M3 to M6
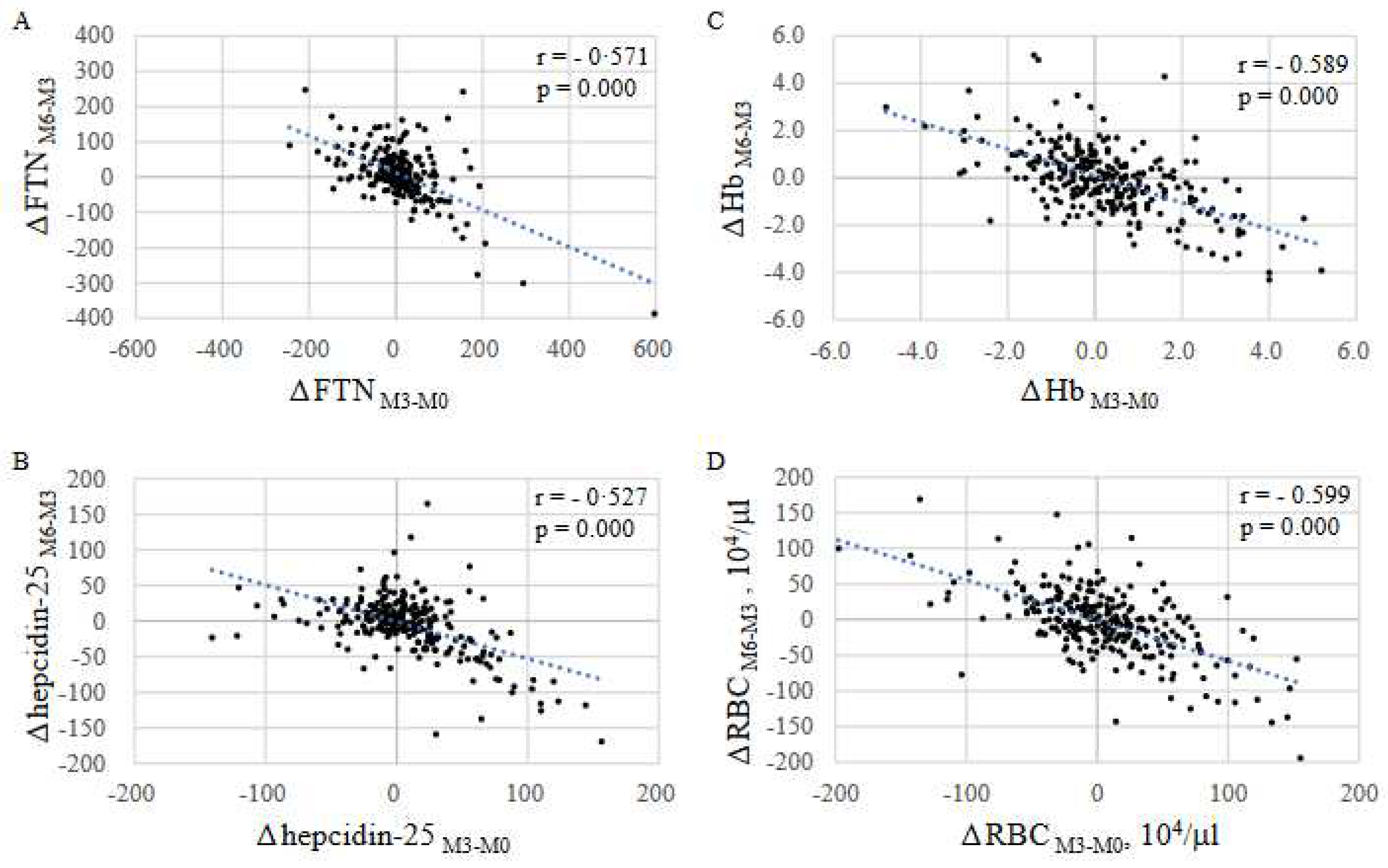
Figure 6.
Effects of RBC on changes of FTN. Two data sets of iron variables were obtained from each patient as independent 3-month periods; ΔM3-M0 and ΔM6-M3. Both data were evaluated equally and the changes in iron variables over 3 months were represented as Δ3M (n=536). To analyze the effect of the change in RBC counts on FTN values over 3 months, each case was classified into 4 groups, G-1, G-2, G-3, and G-4, according to the RBC value at start points (M0 and M3) ; RBC ≤ 300, 300 < RBC ≤ 350, 350 < RBC ≤ 400, and RBC > 400 x104/ml, respectively. Furthermore, each case was classified into G-a when each ΔRBC3M was negative and G-b when positive. Panel A: The start and end points were vectorized as the mean values of MCH and RBC. Hb10g/dl-line (blue) and Hb12g/dl-line (red) are shown based on the formula Hb = RBC × MCH. Panel B presents ΔFTN3M in each group. Data are shown as means ± SEM of samples.
Figure 6.
Effects of RBC on changes of FTN. Two data sets of iron variables were obtained from each patient as independent 3-month periods; ΔM3-M0 and ΔM6-M3. Both data were evaluated equally and the changes in iron variables over 3 months were represented as Δ3M (n=536). To analyze the effect of the change in RBC counts on FTN values over 3 months, each case was classified into 4 groups, G-1, G-2, G-3, and G-4, according to the RBC value at start points (M0 and M3) ; RBC ≤ 300, 300 < RBC ≤ 350, 350 < RBC ≤ 400, and RBC > 400 x104/ml, respectively. Furthermore, each case was classified into G-a when each ΔRBC3M was negative and G-b when positive. Panel A: The start and end points were vectorized as the mean values of MCH and RBC. Hb10g/dl-line (blue) and Hb12g/dl-line (red) are shown based on the formula Hb = RBC × MCH. Panel B presents ΔFTN3M in each group. Data are shown as means ± SEM of samples.
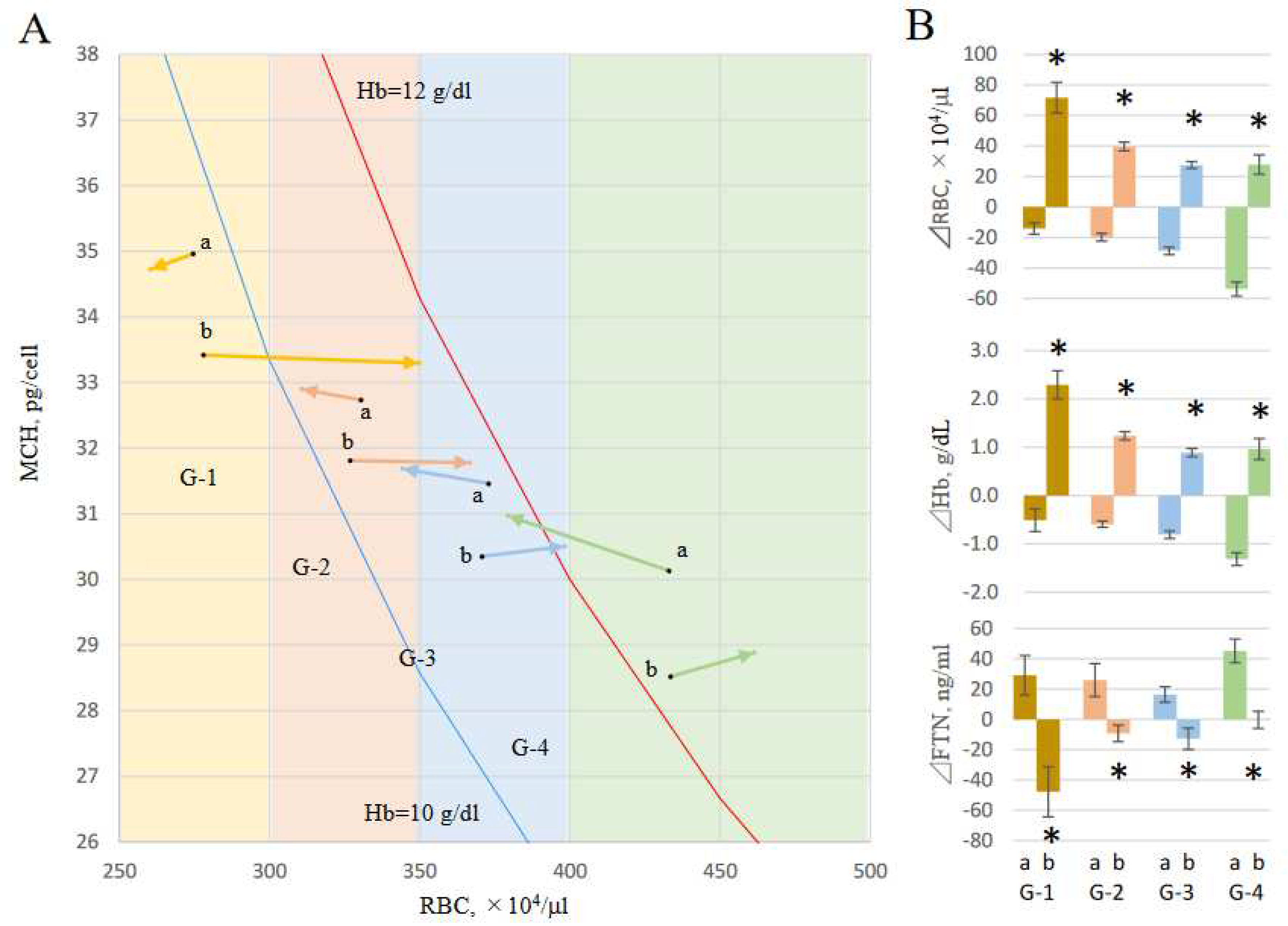
Figure 7.
Model of crosstalk between erythropoietic system and iron metabolic system. Stimulation of erythropoiesis by ESA is a trigger for crosstalk. When ESA stimulates hematopoiesis (A), serum iron is rapidly consumed and decreased. This triggers a decrease in expression of hepcidin-25. Iron is then supplied from iron-stored cells via FPN. If the supply of iron recovered from senescent red blood cells alone is inadequate, stored iron are consumed. This phenomenon appears to be a shift of ferritin-iron to RBC-iron. In this situation iron is readily absorbed from the intestinal tract. When erythropoiesis decreases by limitation of ESA (B), serum iron increases because of reduced iron consumption in bone marrow, and hepcidin-25 also increases. As a result, iron supply to blood is suppressed and unsupplied iron is stored in cells. Iron recovered from senescent red blood cells is also stored. This appears to be a shift of RBC-iron to ferritin-iron. In this situation, iron absorption from the intestinal tract is suppressed.
Figure 7.
Model of crosstalk between erythropoietic system and iron metabolic system. Stimulation of erythropoiesis by ESA is a trigger for crosstalk. When ESA stimulates hematopoiesis (A), serum iron is rapidly consumed and decreased. This triggers a decrease in expression of hepcidin-25. Iron is then supplied from iron-stored cells via FPN. If the supply of iron recovered from senescent red blood cells alone is inadequate, stored iron are consumed. This phenomenon appears to be a shift of ferritin-iron to RBC-iron. In this situation iron is readily absorbed from the intestinal tract. When erythropoiesis decreases by limitation of ESA (B), serum iron increases because of reduced iron consumption in bone marrow, and hepcidin-25 also increases. As a result, iron supply to blood is suppressed and unsupplied iron is stored in cells. Iron recovered from senescent red blood cells is also stored. This appears to be a shift of RBC-iron to ferritin-iron. In this situation, iron absorption from the intestinal tract is suppressed.
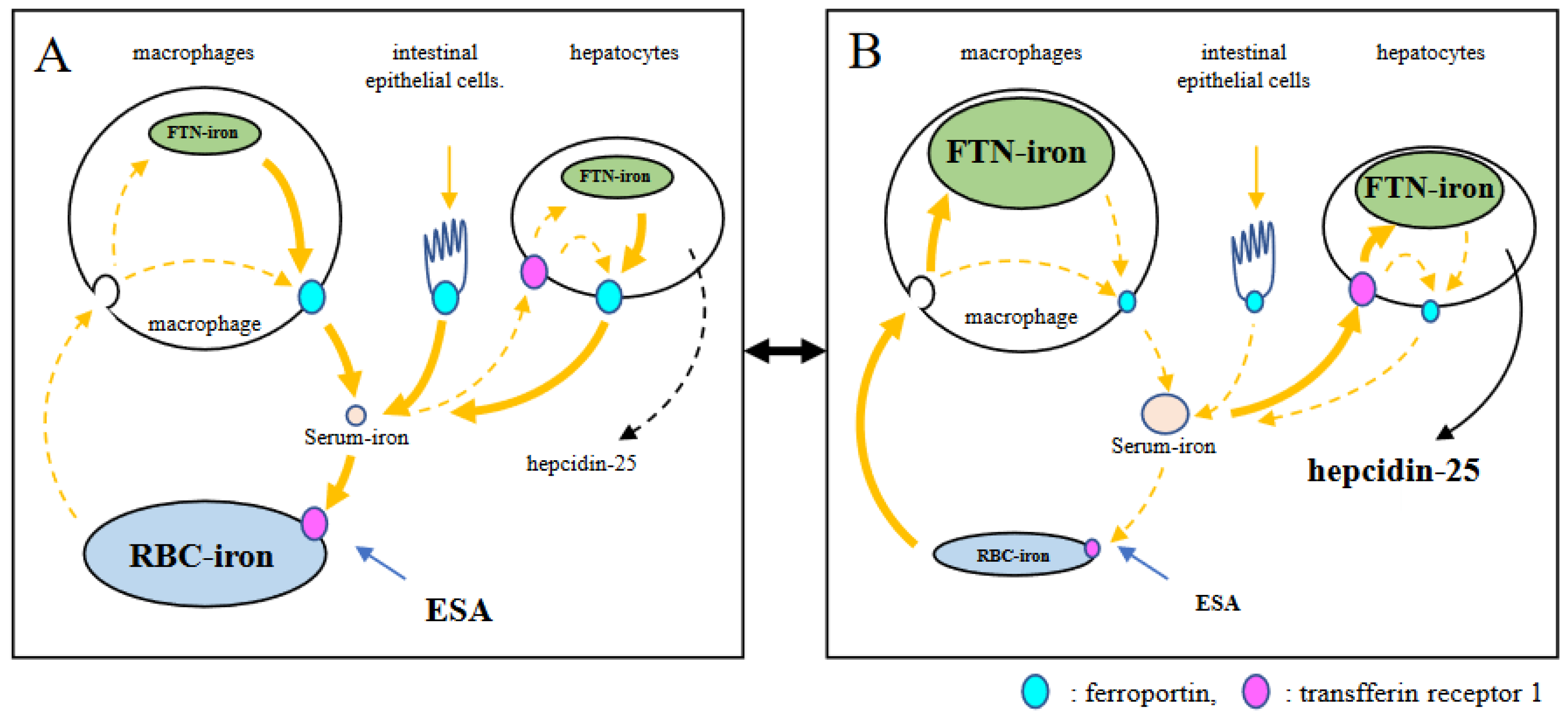
Figure 8.
Hypothesis of Hb adjustment shown in RBC/MCH diagram. When Hb level is ≥ 12 g/dL, ESA is arbitrarily reduced and Hb levels is adjusted between 10 and 12 g/dL according to the guidelines. Hb level, which was 13 g/dL in RBC 450 × 104 /μL (MCH 28.9 pg, point A), was reduced to 11.6 g/dL when reducing ESA and resetting RBC to 400 × 104 /μL (point B). During this period, the FTN level increased because the RBC-iron capacity decreased and the corresponding amount of iron shifted to FTN iron. When FCH administration was continued, MCH increased with iron absorption, and Hb level easily exceeded 12 g/dL (RBC to 400 × 104 /μL , MCH 32.5 pg, point C). If ESA was reduced again and RBC reset to 350 × 104 /μL, Hb level improved to 11.4 g/dL (point D) with a re-elevation in the FTN value. When RBC is ≤ 350 × 104 /μL, there is little risk of Hb being ≥ 12 g/dL even if FCH therapy is continued, because there is an upper limit against MCH, which falls normally within the range 27–33 pg. At this stage, there is no need to further reduce the ESA, and the FTN reaches a plateau without rising.
Figure 8.
Hypothesis of Hb adjustment shown in RBC/MCH diagram. When Hb level is ≥ 12 g/dL, ESA is arbitrarily reduced and Hb levels is adjusted between 10 and 12 g/dL according to the guidelines. Hb level, which was 13 g/dL in RBC 450 × 104 /μL (MCH 28.9 pg, point A), was reduced to 11.6 g/dL when reducing ESA and resetting RBC to 400 × 104 /μL (point B). During this period, the FTN level increased because the RBC-iron capacity decreased and the corresponding amount of iron shifted to FTN iron. When FCH administration was continued, MCH increased with iron absorption, and Hb level easily exceeded 12 g/dL (RBC to 400 × 104 /μL , MCH 32.5 pg, point C). If ESA was reduced again and RBC reset to 350 × 104 /μL, Hb level improved to 11.4 g/dL (point D) with a re-elevation in the FTN value. When RBC is ≤ 350 × 104 /μL, there is little risk of Hb being ≥ 12 g/dL even if FCH therapy is continued, because there is an upper limit against MCH, which falls normally within the range 27–33 pg. At this stage, there is no need to further reduce the ESA, and the FTN reaches a plateau without rising.
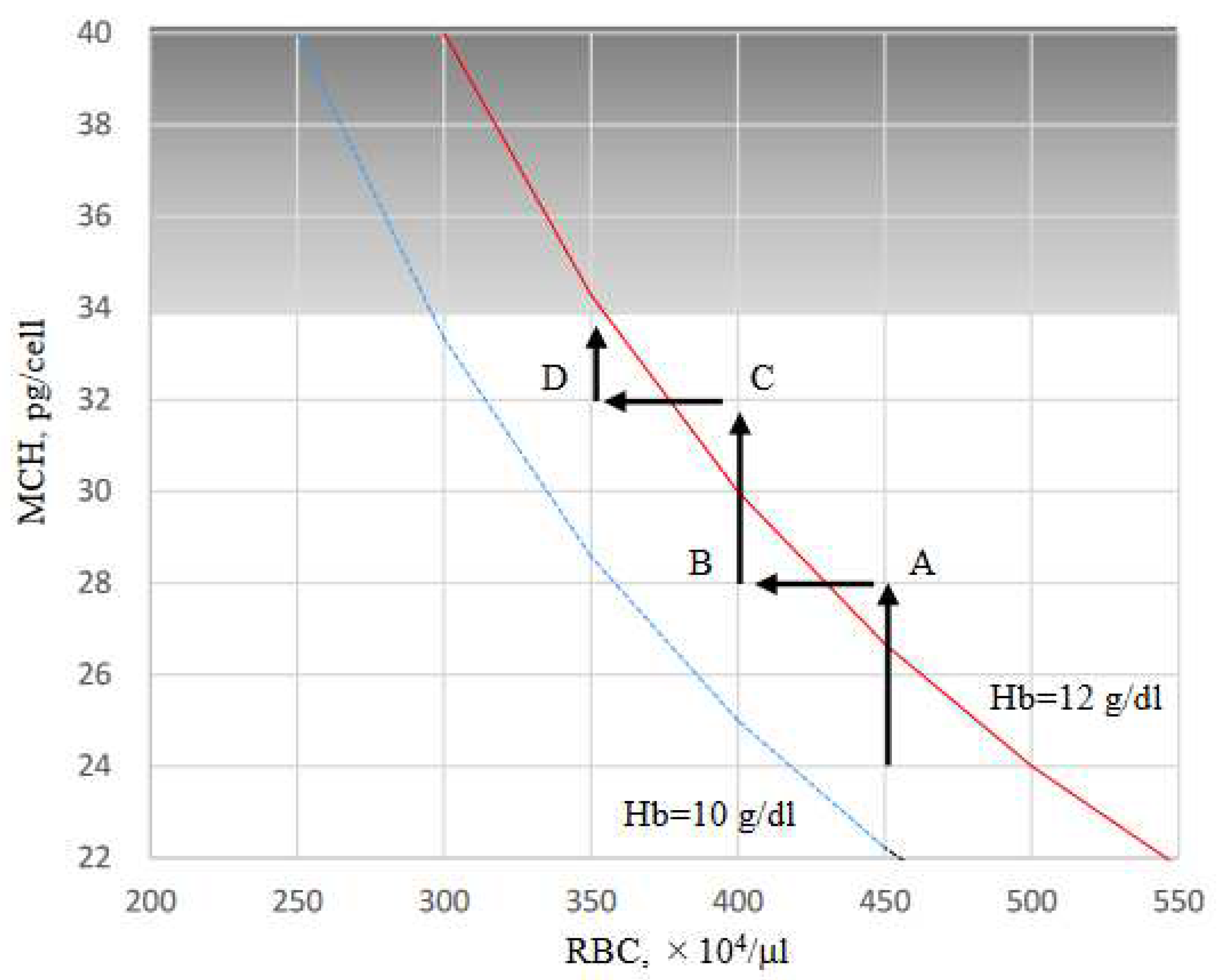

Table 1.
Baseline characteristics of R-OIAT participants.
| Characteristics | Baseline (n = 268) |
|---|---|
| Age (years) | 63.0 (11.6) |
| Males, age, n (%) | 63.8 (11.3), 153 (57%) |
| Females, age, n (%) | 62.0 (12.0), 115 (43%) |
| Riona | |
| 3 tablets (750 mg of FCH) | 149 (55.6%) |
| 6 tablets (1500 mg of FCH) | 101 (37.7%) |
| 9 tablets (2250 mg of FCH) | 18 (6.7 %) |
| ESA | |
| EPO, n (%) | 40 (14.9%) |
| DP, n (%) | 133 (49.6%) |
| MC, n (%) | 65 (24.3%) |
| No ESA, n (%) | 30 (11.2%) |
Data are mean (sd) or n (%). FCH, ferric citrate hydrate; ESA, erythropoiesis-stimulating agent; EPO, erythropoietin; DP, darbepoetin alfa; MC, mircera.
Table 2.
Characteristics of participants and iron status at the time of iron absorption test.
| Baseline | 3 months | 6 months | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M0 (n = 268) | M3 (n = 268) | M6 (n = 268) | |||||||||||
| mean | (sd) | mean | (sd) | mean | (sd) | p | |||||||
| ESA, IU/week | 3679.3 | (3406.8) | 3256.1 | (3205.9) | 3147.7 | (3026.6) | 0.001 | ||||||
| RBC, 104/ml | 360.3 | (44.5) | 364.9 | (48.4) | 360.9 | (45.5) | 0.216 | ||||||
| Hb, g/dl | 11.2 | (1.2) | 11.4 | (1.3) | 11.4 | (1.2) | 0.015 | ||||||
| Ht, % | 34.6 | (3.8) | 35.1 | (4.0) | 34.9 | (3.6) | 0.135 | ||||||
| MCV | 95.8 | (8.8) | 96.6 | (5.5) | 96.6 | (7.5) | 0.117 | ||||||
| MCH, pg/cell | 31.2 | (2.2) | 31.5 | (2.1) | 31.7 | (2.0) | 0.000 | ||||||
| Plat, 104/ml | 19.2 | (5.9) | 20.1 | (14.8) | 18.8 | (6.1) | 0.169 | ||||||
| S-Fe, mg/dl | 65.7 | (26.2) | 70.3 | (28.4) | 69.8 | (27.8) | 0.053 | ||||||
| Ferritin, ng/ml | 100.7 | (93.2) | 108.9 | (99.6) | 116.7 | (102.7) | 0.001 | ||||||
| TSAT, % | 27.4 | (11.6) | 28.9 | (12.0) | 29.5 | (12.5) | 0.047 | ||||||
| Hepcidin-25, ng/ml | 42.9 | (38.7) | 50.5 | (41.6) | 45.6 | (35.6) | 0.006 | ||||||
| DFe2h, mg/dl | 26.6 | (37.2) | 24.7 | (35.8) | 22.9 | (32.3) | 0.270 | ||||||
| P, mg/dl | 5·5 | (1.3) | 5.5 | (1.3) | 5.5 | (1.3) | 0.994 | ||||||
| Albumin, g/dl | 3.9 | (3.4) | 4.1 | (4.1) | 3.9 | (3·5) | 0.282 | ||||||
| AST, IU/L | 13.5 | (6.7) | 13.8 | (6.3) | 13.9 | (7.8) | 0.553 | ||||||
| ALT, IU/L | 11.4 | (5.4) | 11.9 | (7.6) | 12.2 | (9.4) | 0.152 | ||||||
| Al-P, IU/ml | 239·5 | (110.5) | 241.7 | (110.2) | 238 | (111.6) | 0.654 | ||||||
| γ-GTP, IU/L | 21·3 | (21.0) | 20.7 | (17.0) | 22.2 | (25.0) | 0.483 | ||||||
The obtained data are expressed as the mean ± standard deviations (SD). A one-way repeated measures ANOVA was used for the comparison of the data at M0, M3 and M6. The statistical significance was set at p < 0.05.
Table 3.
Effects of the amount of FCH on the changes of iron absorption (ΔFe2h) and iron variables for 6 months.
Table 3.
Effects of the amount of FCH on the changes of iron absorption (ΔFe2h) and iron variables for 6 months.
| time course | F | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| iron variable | FCH (mg) | M0 | M3 | M6 | FCH | time | interaction | ||||||||||
| amounts | course | effects | |||||||||||||||
| DFe2h, mg/dl | 750 | 23.0 | (37.5) | 18.5 | (36.6) | 17.2 | (28.0) | 3.83* | 0.96 | 0.44 | |||||||
| 1500 | 29.9 | (37.1) | 29.6 | (35.3) | 29.1 | (36.2) | |||||||||||
| 2250 | 25.6 | (44.9) | 27.8 | (50.2) | 21.2 | (46.5) | |||||||||||
| Ferritin, ng/ml | 750 | 72.8 | (72.0) | 81.3 | (70.4) | 84.5 | (78.0) | 20.64** | 6.58* | 1.08 | |||||||
| 1500 | 136.7 | (103.8) | 147.9 | (120.8) | 160.3 | (114.4) | |||||||||||
| 2250 | 129.4 | (112.3) | 118.6 | (105.2) | 139.2 | (121.8) | |||||||||||
| Hepcidin-25, ng/ml | 750 | 33.9 | (32.2) | 41.1 | (32.8) | 36.6 | (24.5) | 15.97** | 2.40 | 0.67 | |||||||
| 1500 | 53.8 | (43.1) | 63.0 | (49.6) | 54.7 | (41.9) | |||||||||||
| 2250 | 56.4 | (44.7) | 58.9 | (39.5) | 68.8 | (51.2) | |||||||||||
| TSAT, % | 750 | 26.5 | (11.5) | 29.1 | (12.5) | 28.7 | (11.2) | 0.67 | 4.52* | 0.35 | |||||||
| 1500 | 28.4 | (11.7) | 28.4 | (11.2) | 29.9 | (13.6) | |||||||||||
| 2250 | 28.3 | (12.1) | 30.1 | (12.2) | 32.8 | (15.4) | |||||||||||
| RBC, 104/ml | 750 | 360.3 | (42.5) | 361.2 | (47.6) | 358.2 | (44.9) | 4.62* | 0.56 | 0.72 | |||||||
| 1500 | 365.3 | (44.9) | 372.5 | (48.7) | 368.7 | (42.5) | |||||||||||
| 2250 | 332.3 | (50.3) | 353.8 | (50.9) | 339.8 | (58.8) | |||||||||||
| Hb, g/dl | 750 | 11.2 | (1.1) | 11.3 | (1.2) | 11.3 | (1.2) | 4.46* | 5.77* | 0.70 | |||||||
| 1500 | 11.4 | (1.2) | 11.6 | (1.4) | 11.6 | (1.1) | |||||||||||
| 2250 | 10.6 | (1.5) | 11.3 | (1.4) | 11.1 | (1.5) | |||||||||||
| MCH, pg/cell | 750 | 31.1 | (2.3) | 31.5 | (2.2) | 31.6 | (2.2) | 1.37 | 10.31* | 0.92 | |||||||
| 1500 | 31.2 | (2.0) | 31.4 | (1.8) | 31.6 | (1.8) | |||||||||||
| 2250 | 32.1 | (2.0) | 32.2 | (2.5) | 32.3 | (2.0) | |||||||||||
| ESA, IU/week | 750 | 3701 | (3318) | 3231 | (3367) | 3015 | (3031) | 0.97 | 0.68 | 2.08 | |||||||
| 1500 | 3606 | (3477) | 3092 | (2738) | 3096 | (2773) | |||||||||||
| 2250 | 3902 | (3901) | 4375 | (4134) | 4527 | (4052) | |||||||||||
The obtained data are expressed as the mean ± standard deviations (SD). A two-way (amount× time) repeated measures ANOVA was used for the comparison of the data at M0, M3 and M6. When a significant difference was detected, a multiple comparison test was performed using the post hoc Bonferroni correction. The statistical significance was set at p < 0.05. FCH=750 mg (n=149), FCH=1500 mg (n=101), FCH 2250 mg (n=18). FCH, ferric citrate hydrate; ΔFe2h, iron absorption ; FTN, ferritin; HEP, hepcidin-25; TSAT, transferrin saturation; RBC, red blood cell; Hb, hemoglobin; MCH, mean corpuscular hemoglobin. *p<0.05. **p<0.001
Table 4.
Predictors for ΔFe2h.
| Parameter estimates | ||||||||||
| DFe2h, mg/dl | 95%CI | p value | ||||||||
| Variables | B | SE | lower | upper | ||||||
| hepcidn-25, ng/ml | -0.155 | 0.045 | -0.242 | -0.068 | 0.001 | |||||
| RBC, 104/ml | 0.030 | 0.038 | -0.044 | 0.104 | 0.423 | |||||
| MCH, pg/cell | -2.574 | 0.943 | -4.421 | -0.726 | 0.006 | |||||
| TSAT, % | -0.115 | 0.106 | -0.322 | 0.092 | 0.275 | |||||
| Ferritin, ng/ml | 0.010 | 0.021 | -0.030 | 0.050 | 0.621 | |||||
| ESA, IU/week | 0.001 | 0.001 | 0.000 | 0.002 | 0.113 | |||||
| FCH, 750mg | -10.87 | 9.973 | -30.416 | 8.676 | 0.276 | |||||
| FCH, 1500mg | 0.103 | 9.759 | -19.025 | 19.231 | 0.992 | |||||
| FCH, 2250mg | 0a | |||||||||
| age | 0.018 | 0.123 | -0.224 | 0.259 | 0.884 | |||||
| sex, female | -1.888 | 3.096 | -7.956 | 4.179 | 0.542 | |||||
| sex, male | 0a | |||||||||
a; This parameter is redundant and is set to 0.
Table 5.
Predictors for hepcidin-25 and ferritin.
| Parameter estimates | |||||||||||||||||||||
| hepcidin-25, ng/dl | 95%CI | p value | ferritin, ng/dl | 95%CI | p value | ||||||||||||||||
| Variables | B | SE | lower | upper | B | SE | lower | upper | |||||||||||||
| hepcidn-25, ng/ml | 0.775 | 0.099 | 0.579 | 0.971 | 0.000 | ||||||||||||||||
| RBC, 104/ml | -0.468 | 0.077 | -0.607 | -0.328 | 0.637 | -0.466 | 0.071 | -0.605 | -0.327 | 0.000 | |||||||||||
| MCH, pg/cell | 2.908 | 1.583 | -0.196 | 6.011 | 0.042 | 2.809 | 1.589 | -0.305 | 5.923 | 0.077 | |||||||||||
| TSAT, % | 0.511 | 0.151 | 0.216 | 0.806 | 0.001 | 0.379 | 0.221 | -0.054 | 0.812 | 0.086 | |||||||||||
| Ferritin, ng/ml | 0.224 | 0.0211 | 0.183 | 0.266 | 0.000 | ||||||||||||||||
| ESA, IU/week | -0.002 | 0.000 | -0.002 | -0.001 | 0.000 | -0.002 | 0.001 | -0.004 | -0.000 | 0.050 | |||||||||||
| FCH, 750mg | -13.863 | 6.137 | -25.891 | -1.835 | 0.024 | -21.650 | 20.939 | -62.689 | 19.389 | 0.148 | |||||||||||
| FCH, 1500mg | -8.856 | 6.437 | -21.472 | 3.761 | 0.169 | 31.967 | 22.078 | -11.306 | 75.239 | 0.148 | |||||||||||
| FCH, 2250mg | 0a | 0a | |||||||||||||||||||
| age | 0.017 | 0.092 | -0.164 | 0.197 | 0.851 | -0.028 | 0.354 | -0.723 | 0.666 | 0.936 | |||||||||||
| sex, female | 10.051 | 2.491 | 5.170 | 14.933 | 0.000 | -6.049 | 8.075 | -21.876 | 9.778 | 0.454 | |||||||||||
| sex, male | 0a | 0a | |||||||||||||||||||
a; This parameter is redundant and is set to 0.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



