
Submitted:
16 August 2023
Posted:
17 August 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
The COVID-19 pandemic has been met with an unprecedented response from the scientific community leading to the development, study, and authorization of vaccines and antivirals, ultimately reducing the impact of SARS-CoV-2 on global public health. However, SARS-CoV-2 is far from being eradicated, continues to evolve, and causes substantial health and economic burdens. In this paper, we posit essential points on SARS-CoV-2 and its management during the transition from the acute phase of the COVID-19 pandemic. As discussed, despite Omicron (sub)variant(s) causing clinically milder infections, SARS-CoV-2 is far from being a negligible pathogen. It requires continued genomic surveillance, particularly if one considers that its future (sub)lineages do not necessarily have to be milder. Antivirals and vaccines remain the essential elements in COVID-19 management. However, the former could benefit from further development and improvements in dosing, while the seasonal administration of the latter requires simplification to increase interest and tackle vaccine hesitancy. It is also essential to ensure accessibility of COVID-19 pharmaceuticals and vaccines in low-income countries and improve the understanding of their use in the context of long-term goals of SARS-CoV-2 management. Regardless of location, the primary role of COVID-19 awareness and education must be played by healthcare workers who directly communicate with patients and serve as role models for healthy behaviors.
Keywords:
SARS-CoV-2
; vaccination
; antivirals
; viral evolution
1. Introduction
The coronavirus disease (COVID-19), caused by SARS-CoV-2 and first reported by the Chinese authorities in late 2019, rapidly became an emerging, evolving situation, spreading inevitably to other Asian countries and continents. The World Health Organization (WHO) first declared a Public Health Emergency of International Concern (PHEIC) on January 30, 2020, and considered COVID-19 as a pandemic since March 11, 2020 1–3. On May 5, 2023, it was announced that COVID-19 no longer has PHEIC status 4. Within three years, three months, and five days that passed in between over 765 million SARS-CoV-2 infections were confirmed, with nearly 7 million deaths to COVID-19 5. However, the true toll of the pandemic is likely a few-fold higher due to underdiagnosis, underreporting, and excess mortality 6. In addition, a range of symptoms can persist or onset following acute SARS-CoV-2 infection, a condition known as a post-COVID-19 syndrome, post-acute sequelae of SARS-CoV-2, or long-COVID 7, which also cause significant burden if one considers its global prevalence has been estimated at 43% in the general population 8 and 25% in children and adolescents 9.
The COVID-19 crisis has led to implementing a hygiene regime, face masking, pursuing diagnostic testing daily, and imposing temporary school closures and national lockdown measures. Therefore, it also had a broad societal impact, exacerbated pre-existing deep-rooted structural inequalities, caused numerous changes in different strata of life, and resulted in economic loss 10–15.
The emergence of SARS-CoV-2 has also led to unprecedented scientific response encompassing essential research on diagnostic methods, studies of COVID-19 immunology, viral pathogenicity, and potential therapeutic targets 16,17. Various pharmaceuticals (e.g., arbidol hydroxychloroquine, darunavir, lopinavir, favipiravir, remdesivir, ribavirin, ritonavir, interferons, dexamethasone, and tocilizumab) has been repurposed for COVID-19 treatment with mixed effectiveness results 18–21. The use of convalescent plasma has been eventually abandoned due to the lack of clinical benefits observed in severely ill patients 22,23, while the effectiveness of different monoclonal antibodies has been dramatically impacted by the SARS-CoV-2 evolution 24,25. The development and authorization of the first-generation anti-SARS-CoV-2 oral drugs (nirmatrelvir/ritonavir and molnupiravir) have brought hopes in 2022 that they may represent a game-changer due to the possibility of their use outside the clinical settings 26. However, the relatively high price of these pharmaceuticals and interactions with other drugs have been limiting factors in their use 26.
In 2020, great efforts also focused on developing vaccines to circumvent the need for social distancing and personal protective equipment 27. This eventually led to their authorization in late 2020/early 2021, and the massive global vaccination campaigns pursued in the storm of misinformation, fake news, and anti-vaccine propaganda 28. As estimated, COVID-19 vaccines have averted 19.8 million deaths in 2021 alone 29. When WHO announced COVID-19 is no longer PHEIC in May 2023, over 5.5 billion individuals received at least one vaccine dose. Despite the high effectiveness of vaccines against severe disease and death 30,31, it soon became evident that due to a gradual decrease of serum antibodies, the vaccination does not offer long-term protection from SARS-CoV-2 infection, leading to the recommendation of subsequent booster doses. In addition, viral evolution has led to the emergence of lineages, such as Omicron and its descendants, characterized by an increased ability to escape humoral responses. Although the primary goal of COVID-19 vaccination is to decrease the rates of hospitalizations, admission to intensive care units, and deaths 32 and is often achievable due to the extended duration of vaccine-induced cellular immunity and its lower susceptibility to viral mutations 33–35, some individuals may still experience severe COVID-19 due to a worse response to immunization because of age-related immunosenescence, primary or secondary immune deficiencies, and various lifestyle factors 36–39.
All in all, SARS-CoV-2 is far from being eradicated in the near future. It remains, as also emphasized by the WHO, a global health threat 36. According to the official data, nearly 11 thousand COVID-19 deaths, with 64% in high-income countries, were reported in May 2023 since the WHO called off PHEIC. It cannot be excluded that COVID-19 will become endemic, meaning it will remain consistently present at predictable spread and occurrence rates. In this context, endemic does not necessarily imply that infection rates are low or that the disease is mild. For example, malaria is regarded as endemic in selected world regions, with its incidence rate and mortality remaining relatively stable since 2015, with approximately 600,000 deaths annually 40. The endemic phase of COVID-19 will also require appropriate management strategy and preparedness to decrease the disease burden systematically and by no means should be used as a synonym for safe infections 41.
Therefore, in this article, we highlight the essential issues regarding the benefits and future of COVID-19 vaccination, SARS-CoV-2 evolution and its impact on clinical significance, and the continuous need to pursue various control measures when exiting an acute phase of the COVID-19 pandemic, but still existing with the virus that can have profound effects on public health.
2. SARS-CoV-2 Is Here to Stay and Will Continue to Evolve
The priority of vaccinology has always been to decrease the clinical severity of infection. Preventing infection (whether symptomatic or asymptomatic) has been a secondary goal. The eradication of the pathogen is the most challenging task. Until today, smallpox remains the only human disease successfully eradicated due to vaccination campaigns 42. SARS-CoV-2 will continue circulating in the human population primarily because of the short-lived immune response following natural infection and vaccination, and due to viral evolution. SARS-CoV-2 belongs to RNA viruses that exhibit higher rates of spontaneous mutations than DNA viruses 43. The primary mechanism behind this phenomenon lies in the lower replication fidelity of polymerase enzyme, ultimately leading to point mutations. Frequently they do not affect virus biology or are deleterious to its further replication. A small minority of such mutations will provide fitness advantages and impact different aspects of virus biology, e.g., pathogenicity, infectivity, transmissibility, and antigenicity. SARS-CoV-2 mutation rate has been estimated at 1 × 10–6 – 2 × 10–6 mutations per nucleotide per replication cycle 41,44, which is lower than the rate of various other RNA viruses such as influenza viruses (3 x 10-5), human immunodeficiency virus (10−4 to 10−5), and hepatitis C virus (3.5 × 10–5 to 1.2 × 10–4) 45–47. This is because SARS-CoV-2's polymerase, similar to that of other coronaviruses, utilizes a proofreading 3′-to-5′ exoribonuclease of the nonstructural protein 14, a mechanism ensuring higher fidelity of replication, which is not present in the majority of RNA viruses. Nevertheless, the accumulation of point mutations in SARS-CoV-2 gives rise to novel lineages and sublineages that are competitive regarding transmissibility. An example of such mutation is D614G in spike protein that emerged in late January/early February 2020 and increased SARS-CoV-2 infectivity and soon became widespread 48.
The other process that can drive SARS-CoV-2 adaptation is recombination. It results from co-infection of the host cell with two genetically distinct viruses that, when recombined, produce viable hybrid progeny 49. The odds of this process playing a more significant role in SARS-CoV-2 evolution increased over time due to the emergence and subsequent co-circulation of genetically divergent viral (sub)lineages, a phenomenon particularly evident in the Omicron era 41. An example of such SARS-CoV-2 recombinant that gained global relevance is the XBB that emerged from the recombination of BA.2.10.1 and BA.2.75 sublineages 50. Further accumulation of point mutations within this recombinant lineage gave rise to XBB.1.5, which became dominant in various world regions in 2023 51.
SARS-CoV-2 will continue to evolve by accumulating advantageous point mutations and recombination events (Figure 1). It is pivotal to monitor its evolution and understand the key biological and clinical features of the emerging (sub)variants. One should note that the virus can become more adapted through various processes that depend on ecological and epidemiological factors. For example, mutations leading to enhanced evasion of humoral immunity are more likely to be subject to higher selective pressure when the population immunity levels gradually increase. Such vaccine-breakthrough and antibody-resistant mutations are observed in Omicron, and it can be expected that they might become a dominating pathway of SARS-CoV-2 evolution when most of the world is either infected or vaccinated 52. Importantly though, it does not imply the entire loss of the COVID-19 vaccines effectiveness since a vital role in antiviral response is played by the vaccine-induced adaptive cellular immunity 53, which is less prone to evasion through viral mutations as also evidenced in the case of various Omicron sublineages 54,55. Nevertheless, it indicates that managing the SARS-CoV-2 burden will require the systematic administration of booster doses and consideration of updated variant-adapted doses, particularly if one aims to increase protection from symptomatic infection. In parallel, the implementation of novel vaccination strategies is needed to utilize other antigens than spike protein due to a high number of mutations in its gene (second next to the gene encoding nonstructural protein 3) 56. In response to this need, approaches based on multiple antigen-targeted cell-mediated immunity were suggested to overcome waning antibody responses and attenuate infectious breakthrough events and disease severity of future SARS-CoV-2 variants 57.
3. Omicron Lineage Is Milder But Not Negligible
The SARS-CoV-2 variant belonging to the Omicron lineage was identified for the first time in November 2021 in Africa. It has been characterized by a large number of sense mutations, exceeding 30 in the gene encoding spike protein, including 10 in the receptor-binding domain 59. Its high transmissibility soon led to its global distribution, a rise of novel sublineages, and the replacement and ultimate extinction of previous viral variants (Figure 1). Numerous studies have consistently shown that the enhanced transmissibility of Omicron is due to its ability to better evade the humoral immunity of vaccinated and individuals with a history of SARS-CoV-2 infection 60,61 At the same time, there is mounting evidence that Omicron lineage is causing milder infections in humans. Firstly, experimental studies demonstrated its less efficient membrane fusion kinetics than previous SARS-CoV-2 lineages, preferential endocytic cell entry, and faster replication in the human bronchus, while less efficient in lung cells 62–64. All of these features translated into decreased severity of infection. This has been clearly reflected in vivo studies employing naïve animals, including rodents and non-human primates 65–69. Epidemiological analyses of various human populations confirmed that Omicron infections are characterized by decreased lower respiratory tract involvement, reduced odds of hospitalization due to severe COVID-19, and less mortality 70–74. Despite the emergence of subsequent Omicron sublineages, such as BA.4/BA.5 and XBB, the hospitalization and death risk remained lower compared to previous SARS-CoV-2 variants, such as Delta 75–77.
Although Omicron causes statistically-wise milder infections with better clinical prognosis, it should not be regarded as a negligible pathogen (Figure 2). Since the beginning of 2022, when Omicron became dominant in most world regions (Figure 1), until the first half of 2023, approximately 1,455 million deaths of COVID-19 patients were confirmed 5. The mean monthly death rate in the pre-Omicron period amounted to 228 thousand, while during the Omicron era, it fell over 2.5-fold to 86 thousand, which still is substantial and results in larger mortality than in the case of seasonal influenza, which annual toll is estimated globally at 290 000-650 000 deaths 76). This was also reflected by the results of a comparative study in patients requiring hospitalization which demonstrated that Omicron infection is associated with an approximately 1.5-fold higher risk of in-hospital all-cause mortality than seasonal influenza viruses 79. Moreover, due to high transmissibility and the ability to infect many individuals in a given period, the rising rates of hospitalizations due to this variant in some countries were even higher than during the Delta wave 80. Further, within the first 1.5 years of Omicron dominance, more than 478 million cases of infection were officially reported, representing 165% of cases in the first two years since early 2020 5. This indicates that Omicron, even if its infections are more frequently mild, is to substantially affect absenteeism from work and schools due to illness. Last but not least, individuals infected with the Omicron can also report, similarly to other SARS-CoV-2 variants, a range of persisting symptoms, collectively known as post-COVID-19 syndrome, post-acute sequelae of SARS-CoV-2, or long-COVID 7. Although the odds of this condition with Omicron were reported to be significantly reduced compared to Delta variant 81, its estimated rate in the post-acute cohort (6-15 months from infection) was 17%, most often characterized by chronic fatigue, but a range of symptoms encompassed systemic, cardiac, dermatologic, ocular, otologic, gastrointestinal, metabolic, musculoskeletal, neurologic, psychiatric and respiratory and urinary 82.
Omicron is not a "natural solution" to the COVID-19 problem, as some have suggested 83, and should not be, along with various sublineages, regarded as a negligible pathogen. Despite a milder course of the disease, it continues to cause substantial health and economic burdens, management of which requires appropriate awareness, preparedness, and resources.
4. Future Viral Variants May Not Necessarily Be Always Milder
It is challenging to predict the future clinical relevance of SARS-CoV-2. However, it is not certain that its further evolution will lead to a decrease in infection severity. As demonstrated by the recent study, the directions of change in intrinsic case severity across successive SARS-CoV-2 variant waves have been inconsistent. It increased continuously from the early lineages, through the Alpha variant to Delta lineage, to decrease substantially in the case of BA.1 Omicron and even further when BA.2 emerged 84. This contradicts the notion that SARS-CoV-2 transmissibility can only be enhanced at the expense of its pathogenicity since the Delta variant, infection with which was characterized by increased severity, was significantly more transmissible than preceding lineages 85. This advantage in the spread was gained predominantly by higher viral loads 85,86. In turn, the Omicron variant does not cause elevated viral loads in the respiratory tract compared to those observed for the Delta variant, while some studies reported that these loads might even be lower 87–89. In addition, it does not reveal a higher affinity to the angiotensin-converting enzyme-2 receptor and demonstrated attenuated fusogenicity due to decreased use of the cellular protease TMPRSS2 resulting in greater utilization of the endocytic pathway during cell entry 63,90,91. Its enhanced transmissibility is due to the efficient escape from humoral immunity of individuals with a history of SARS-CoV-2 infection and those who were vaccinated. SARS-CoV-2 may continue to evolve into a greater escape from infection- and vaccination-acquired immunity. This could lead to its high transmissibility without a significant increase in severity, particularly if immune escape will mostly concern humoral and not cellular responses. However, considering that SARS-CoV-2 is most transmissible prior to symptoms onset and at the beginning of the symptomatic phase 92, the mutation-enhancing viral loads could also lead to superior transmission yet be potentially accompanied by more severe infections due to a higher risk of hyperinflammation and disease severity under such a scenario 93.
Moreover, the viral evolution may lead to a gradual increase in fusogenicity, which is known to impact the disease severity 94. This process has already been reported for more newly emerging Omicron subvariants such as BA.4/BA.5 and XBB that demonstrated higher fusogenicity of spike protein compared to early BA.1 and BA.2 SARS-CoV-2 95–97. Experimental studies have shown a close relationship between enhanced viral fusogenicity and pathogenicity 95,98. Although epidemiological studies reveal some differences in clinical severity between original and later Omicron subvariants, they consistently indicate that it remains reduced compared to the Delta lineage 99–101. It is plausible that a history of immunization, be it SARS-CoV-2 infection, COVID-19 vaccination, or both, plays a protective role in attenuating an increased severity in the human population that would otherwise be expected.
When considering the future of SARS-CoV-2 evolution, one should note that it can also infect non-human hosts, including wild animals and livestock 102–106, and potentially return to the human population through contact with these species. In addition, Omicron can likely utilize a broader range of host species than other SARS-CoV-2 variants, while the risk of cross-species infection is higher to increased human mobility than in the case of the pre-Omicron era when various sanitary restrictions were imposed 107. The clinical consequences of such retransmission to the human population are challenging to predict since mutation-driven adaptations to a new host may lead to decreased adaptation to the human environment but also to better evasion of acquired immunity, including cellular response, and thus higher susceptibility to severe disease 108–111.
In conclusion, predictions of the exact clinical trajectories of future SARS-CoV-2 (sub)variants should be made cautiously to avoid communication disregarding the relevance of this pathogen but also fear-promoting messages. SARS-CoV-2 requires continuous genomic surveillance conducted globally with data sharing in the open domain and accompanied by in vitro and in vivo studies on viral biology, pathogenicity, and evasion of acquired immunity. This approach is essential for the timely implementation or modification of safety measures, including vaccines.
5. Vaccines Remain a Key Component of Primary COVID-19 Prevention
The benefits of COVID-19 vaccination are well documented. According to a mathematical modeling study, their administration has averted 19.8 million deaths in 2021 alone 29. Numerous analyses encompassing a period preceding the dominance of the Omicron demonstrate the public health impact of COVID-19 vaccines in different world regions regarding averted deaths, hospitalizations, and infection 112–120. According to a meta-analysis that included real-world studies conducted before Omicron's emergence, the overall COVID-19 vaccine effectiveness against SARS-CoV-2 infection, COVID-19-related hospitalization, admission to the intensive care unit, and death was 89.1, 97.2, 97.4, and 99.0%, respectively, with better effectiveness against infection observed for mRNA vaccines 113. Further, the majority of conducted studies have shown that vaccination reduced the risk of long-COVID 121–128.
However, vaccine effectiveness against infection decreased when the Omicron lineage emerged and became widespread due to its enhanced ability to escape humoral responses 31,129–131. According to meta-analysis, booster dose administration improved to some extend protection against symptomatic Omicron infection, reaching 57% within three months from administration but decreasing to 33% after six months 132. However, COVID-19 vaccines remained highly effective in protecting against severe COVID-19 and death in the era of Omicron, and this effect was further demonstrated to be improved/restored by the booster vaccinations 133,134. As indicated in the meta-analysis, the real-world effectiveness of booster doses against severe disease caused by Omicron infection was 86% 132. Another meta-analysis estimated effectiveness of booster doses against Omicron infection and hospitalization at 70% and 89%, respectively, decreasing to 43% and 71% at 112 days or later 135. In children and adolescent populations, pooled effectiveness of two COVID-19 vaccine doses against symptomatic Omicron infection was 51 and 61%, respectively, with the pooled effectiveness against hospitalization of 70% 136. As calculated in the UK, a booster dose program in autumn-winter 2021 averted 12.8 million cases, 1.1 million hospitalizations, and 290,000 deaths during the first three months of Omicron dominance in 2022 137. This clearly shows that even though the authorized COVID-19 vaccines are still not optimal, they save lives, protect health and decrease economic loss caused by SARS-CoV-2.
In response to the emergence of the Omicron lineage, novel bivalent booster mRNA vaccines were developed and authorized in the second half of 2022. Their administration provided additional protection against symptomatic SARS-CoV-2 in immunocompetent persons who previously received monovalent vaccines only 138. Early estimates show that in adults aged 18-49 years, the effectiveness of a bivalent mRNA booster dose (with mRNA encoding primary spike protein antigen and BA.4/BA.5 spike protein) given 2-3 months earlier compared to no bivalent booster was 52% against symptomatic infection with BA.5 Omicron subvariant and 48% against infection with XBB/XBB.1.5 139. A retrospective cohort study conducted in Israel confirmed that bivalent mRNA vaccines significantly reduced hospitalization and mortality in individuals aged ≥65 years 140. However, one should note that the effectiveness of these bivalent booster doses against infection with the Omicron variant was not as high as could be expected. This phenomenon may be due to immunological imprinting, according to which the immune system of those already vaccinated with monovalent vaccines was primed to respond to the ancestral strain of SARS-CoV-2. As a result, the administration of bivalent vaccines revoked response to epitopes shared by Omicron (BA.4/BA.5) and the ancestral strain rather than to unique epitopes of Omicron, as also directly demonstrated by the lack of BA.5-specific antibodies in the serum of individuals boosted with bivalent COVID-19 vaccines 141–144. Therefore, future booster doses are likely to be monovalent and lack the index-virus antigen, also because they have been adapted to lineages currently considered extinct 145.
Importantly, studies conducted during the Omicron dominance also show that vaccines continue to decrease the risk of long-COVID 146. As estimated, the booster dose administration in autumn-winter 2021 resulted in a 68% reduction of newly diagnosed long-COVID cases in the first quarter of 2022 when Omicron was the dominant SARS-CoV-2 lineage 137.
In summary, the available evidence consistently demonstrates that all individuals should stay up to date with recommended COVID-19 vaccines, including receiving updated doses. COVID-19 vaccination reduces the overall burden of SARS-CoV-2 regardless of dominating lineage, including clinically milder Omicron. Nevertheless, it requires booster doses, including those based on updated antigens. Importantly, the mRNA platform enables the rapid manufacturing of novel versions of COVID-19 vaccines if such a need arises 147.
At the same time, there is a need to pursue efforts to develop vaccine candidates that could confer more durable protection against SARS-CoV-2 infection and be less prone to mutations in gene encoding spike protein. One approach in this regard is focusing on the use of self-amplified mRNA vaccine candidates that aim to induce multiple antigen-targeted cell-mediated immunities in addition to neutralizing humoral responses in order to bypass waning antibody concentrations and attenuate infectious breakthrough and disease severity of future SARS-CoV-2 variants 57. Preclinical data shows that using dual antigen mRNA vaccines, encoding viral nucleocapsid and spike proteins, is superior in controlling SARS-CoV-2 (including the Omicron variant) in the lower and upper respiratory tract than immunization with mRNA encoding exclusively spike protein 148.
In addition, further efforts to develop effective intranasal COVID-19 vaccines are necessary as this route of administration may offer several advantages. Contrary to intramuscular vaccines, it can induce a significant mucosal immunity that plays a role in host defense in the upper respiratory airway, a primary entry site of viruses such as SARS-CoV-2, ultimately preventing virus infection, replication, shedding, transmission, and disease development and progression 149,150. Secondly, the components of these vaccines can be absorbed through the mucosa, leading to systemic immunity 151,152. Last but not least, the intranasal route of administration is less invasive and painless and may translate into lower vaccine fears and improved acceptance 153,154. So far, developing safe and efficacious intranasal COVID-19 vaccines remains a challenge 155. The intranasal COVID-19 adenoviral vaccine candidate has failed to induce robust mucosal and systemic immunities in phase 1 clinical trial 156, despite encouraging preclinical data 157. In turn, intranasal administration of lipid nanoparticles employed to encapsulate mRNA has led to inflammatory responses in the lungs and resulted in a high mortality rate 158. These findings indicate the need for better preclinical models for mucosal immunity in humans and to develop strategies for the safe delivery of some vaccines, i.e., based on the mRNA platform.
6. Simplifying COVID-19 Booster Vaccination Will Improve Vaccine Acceptance and Intake
Given that COVID-19 vaccines will remain a primary strategy to decrease the health burden caused by SARS-CoV-2, it is essential to simplify the vaccination protocols, particularly regarding booster administration. Various observations demonstrate a significant decline of vaccination-induced antibodies within six months from the previous dose and indicate that the biannual boosting with mRNA vaccines (most frequently used for this purpose) will induce the highest level of protection against infection 159–162. Under such an approach, the risk of breakthrough infection over six years was estimated at 7-11%. In comparison, annual boosting would also substantially reduce the 6-year risk to 25-31%. In turn, delaying boosting beyond two years yielded cumulative risks of future infection nearly as high as foregoing boosting entirely 162.
However, one should bear in mind that interest in COVID-19 vaccines decreases with subsequent boosting doses. For example, by June 2023, 76% of the population had received at least one dose of the COVID-19 vaccine in the European Economic Area, the primary course of vaccination was completed by 73%, the first booster was received by 55%, the second one by 14%, while a third one only by 2% 164. This trend has a multifactorial basis, including low perceived benefits of receiving a booster vaccine, a low subjective risk of severe COVID-19, disappointment in vaccines due to experience of breakthrough infection or adverse effects after the previous vaccine dose, and loss of trust in health authorities during the pandemic 164–167. All of these factors are more or less rooted in inappropriate communication on the role of COVID-19 vaccination in decreasing the overall SARS-CoV-2 burden on public and individual health as well as the economy. They may also arise from various unknowns on COVID-19 vaccines that existed when they were introduced (e.g., regarding the durability of immunity) and confusion about shifting public health guidelines regarding vaccine safety, changing the interval between doses, mixing particular vaccines brands, and introducing subsequent booster doses without knowing whether and when additional ones will be required 168. These issues are currently clarified, allowing for the simplification of COVID-19 booster strategies, translating into lower vaccine hesitancy and better acceptance.
SARS-CoV-2 reveals a seasonal behavior, which is generally in line with this seen for other respiratory viruses, such as influenza viruses and respiratory syncytial virus 169. For example, in Europe and the United States, the highest SARS-CoV-2 burden, i.e., infections, emergency visits, hospitalizations, and deaths, can be expected between autumn and early spring 170–172. This strongly advocates that the administration of seasonal booster vaccines should be performed in a similar timeframe as in the case of influenza. This timeframe also coincides with when RSV vaccines (currently gaining authorization for use in particular groups) will be recommended 173. This creates an opportunity, comfortable and less time-consuming from the perspective of those interested in vaccination, to offer seasonal booster COVID-19 vaccine simultaneously during the same visit as those against influenza and RSV. As recently shown, concurrently administering COVID-19 and influenza vaccines was not associated with additional safety risks and remained immunogenic, although marginally lower anti-S antibodies levels were observed compared to booster COVID-19 vaccination alone 174. Ultimately, the future may lie in combined vaccines. Some multicomponent mRNA vaccine candidates against COVID-19, influenza, and RSV have already entered clinical phases of testing (e.g., mRNA-1230, NCT05585632). mRNA platform enables the development of updated COVID-19 booster and seasonal influenza vaccine based on antigen selection approximately three months before the increased number of infections is expected 147. This approach should increase the protection levels, not only against severe disease but also symptomatic infection, during a period when respiratory diseases are the most overwhelming for healthcare systems. A decision to select a novel version of the SARS-CoV-2 antigen should be made based on genomic surveillance data and genetic divergence between currently and previously dominating viral sublineages.
Seasonal COVID-19 booster campaigns should preferentially target those at the highest risk of severe disease, including the elderly, individuals with comorbidities and immunodeficiencies, but also pregnant women, and healthcare workers. This approach has also been recommended recently in a joint statement by the European Centre for Disease Prevention and Control and the European Medicines Agency 170,171. We posit that these groups should be prioritized for reimbursed vaccines by local authorities. However, the seasonal COVID-19 vaccines should also be made available for other eligible groups, e.g., through commercial distribution in a similar fashion to how influenza vaccines are offered in different world parts. Receiving them also by those at lower risk of severe disease would decrease the risk of experiencing mild symptomatic infection and its consequences, such as long-COVID and being forced to abstain from work temporarily. It seems reasonable to recommend seasonal booster COVID-19 vaccinations, preferentially pursued at the same time as immunization against other respiratory viruses such as influenza and RSV.
7. Antivirals Represent a Strategy to Adapt to Long-Term Co-existence with SARS-CoV-2
COVID-19 treatment depends on the severity of the infection and the presence of risk factors in infected patients. Pharmaceuticals targeting SARS-CoV-2 aim to inhibit viral replication and prevent disease progression to a more severe form 171,172. For this purpose, they need to be applied in the early symptomatic phase 174,175. However, ensuring the targeted site is not subject to frequent mutations is critical. Otherwise, the effectiveness of such pharmaceuticals may soon decrease due to viral evolution. Such effect has been observed in the case of various monoclonal antibodies either used to treat infection or as preexposure prophylaxis 176.
In 2022, two oral antivirals specifically targeting the SARS-CoV-2 replication cycle were recommended for use in different world parts: nirmatrelvir/ritonavir and molnupiravir 176,177 (Figure 3). Both require 5-day treatment initiated no later than five days from symptoms onset. The former inhibits the main SARS-CoV-2 protease, pivotal to processing polyprotein precursors, ultimately leading to the inability of the virus to replicate. This mechanism is ensured by nirmatrelvir, which is extended in the presence of a low dose of ritonavir, acting as an inhibitor of CYP3A-mediated metabolism. The pivotal phase 2–3 double-blind, randomized, controlled trial on nirmatrelvir/ritonavir conducted in symptomatic, unvaccinated, nonhospitalized adults at high risk for progression to severe COVID-19 reported a reduction of hospital admission or death by 97% relative to placebo. Experimental studies demonstrated that it remains effective against various Omicron subvariants 176, the observations further confirmed by clinical trials and real-world studies conducted in different populations and reporting reduced hospitalizations and mortality in treated patients 177–179. In addition, a large cohort study found that treatment with nirmatrelvir/ritonavir during the period dominated by the Omicron variant is associated with reduced risk of long-COVID regardless of vaccination status and prior infection 180.
Molnupiravir is a small-molecule ribonucleoside pro-drug of N-hydroxycytidine 181, which was tested prior to the COVID-19 pandemic for potential use against SARS-CoV-1 and MERS-CoV 182,183. Its mechanism of action is based on so-called lethal mutagenesis, the process in which viral RNA-dependent RNA polymerase is misdirected to induce transition mutations throughout the genome during viral replication, ultimately leading to errors deleterious for the virus. A double-blind, randomized, placebo-controlled phase 3 clinical trial in symptomatic, unvaccinated, nonhospitalized adults has shown that the risk of death was 89% lower in the group receiving molnupiravir for five days 184. However, no clinical benefit was found in the clinical trial involving hospitalized patients 185. These studies were conducted during the dominance of viral variants other than Omicon. Nevertheless, experimental in vitro studies demonstrated that molnupiravir remains efficacious against this variant 176, which was further confirmed in clinical trials and real-world studies, also involving hospitalized COVID-19 patients 177,186,187. Moreover, molnupiravir use was also associated with reduced risk of long-COVID regardless of vaccination status and history of previous SARS-CoV-2 infection 188.
Apart from oral antivirals, which can be used in outpatient and inpatient settings, an important treatment option in patients hospitalized with COVID-19 includes remdesivir, an intravenously administrated non-canonical nucleotide developed prior to the COVID-19 pandemic (Figure 3). It acts as an inhibitor of the RNA-dependent RNA polymerase of RNA viruses of several families, including Paramyxoviridae, Filoviridae, and Coronaviridae. Based on evidence from clinical studies, remdesivir was authorized in 2020 by various health authorities to treat COVID-19 in adults and adolescents (>12 years with weight ≥ 40 kg) who require oxygen therapy. It can also be used in adults who do not require oxygen supplementation but represent a high-risk group for severe COVID-19 189. Experimental in vitro studies have shown that it remains efficacious against Omicron's sublineages, including BQ.1.1 and XBB 176. This was also confirmed in the real-world analysis in which remdesivir use in patients hospitalized during Omicron dominance was an independent predictor of lower mortality, similar to the period dominated by the Delta lineage, 190.
In summary, antivirals such as nirmatrelvir/ritonavir, molnupiravir, and remdesivir retain their effectiveness against the novel SARS-CoV-2 sublineages and continue to be important elements of COVID-19 therapy. It is pivotal to ensure their availability, particularly when an increased number of SARS-CoV-2 infections can be expected (e.g., during the autumn-winter season in the temperate zone). In this regard, oral antivirals are the pharmaceuticals of choice as they reduce healthcare costs through decreased hospitalization rates 191–193. At the same time, it is important to pursue research on potential benefits arising from therapies based on the combination of antivirals, assessed mostly as case reports 194 or in vivo rodent studies 195. Such combinations may slow the emergence of resistance mutations, as already evidenced for other pathogens 26,196,197. This may be particularly of interest in the case of immunocompromised patients since they are often characterized by an extended time of viral elimination even when treated with available antivirals 194,198–200. High dosing regimen recommended for nirmatrelvir/ritonavir (3 tablets administrated twice a day for five days) and molnupiravir (four capsules every 12h for five days) also represents a significant challenge due to a risk of missing a dose or inappropriate adherence, not possible to directly control outside the clinical setting. Moreover, nirmatrelvir/ritonavir tablets and molnupiravir capsules are relatively large (8.5 x 17.5 mm and 7.6 x 21.8 mm, respectively 201,202) and cannot be chewed or crushed and may be difficult to swallow by selected patients, including elderly for whom this issue has been particularly recognized 203,204. Improved formulations, requiring reduced dosing and based on the smaller size of swallowed tablets/capsules, would be ultimately desired. Simultaneously, the continued efforts to increase the portfolio of anti-SARS-CoV-2 pharmaceuticals, preferentially administrated orally and acting on different viral targets, is highly encouraged 203.
8. Leaving No Country Behind: Low-Income Regions Require Better Access to COVID-19 Vaccines and Antivirals
Considering that COVID-19 vaccines and SARS-CoV-2 antivirals remain essential tools during the transition from the acute phase of the pandemic, it is pivotal to pursue efforts to increase their availability and willingness to use in low-income countries. Although the case-fatality ratio for various low-income areas, e.g., the African continent, remains below that observed globally 205, it does not implicate that SARS-CoV-2 is a negligible pathogen. This is also because the reasons behind such epidemiological phenomenon are unclear, with various hypotheses put forward, including cross-protection from other infections or younger populations than in other worlds parts 206–208. However, low-income countries likely have the highest rates of underreported COVID-19 cases (including severely diseased) and COVID-19 mortality 209,210. At the same time, they are represented by low vaccination rates, with approximately 25% of the population of low-income countries having completed an initial vaccination protocol 5. This is due to several factors, including insufficient supply, limited local vaccine production, inequitable distribution, weak healthcare systems, low perceived risk, and high vaccine hesitancy 211.
The efforts should be continued to improve vaccine equity in low-income countries through better support of humanitarian initiatives from high-income countries, such as the COVID-19 Vaccines Global Access (COVAX) initiative. As of early 2023, it delivered 1.88 billion doses 212, despite its initial target to deliver 2 billion doses in 2021 213. This results in great discrepancies in vaccination rates in the world, i.e., after 2.5 years from COVID-19 vaccines authorization (June 2023), the percentage of the population with completed initial vaccination protocol in low-income countries is similar to that already reached by high-income regions after six months of the campaign in 2021 (Figure 4). This failure resulted mainly from the subdued efforts from wealthy regions, vaccine nationalism, and trade between high-income countries 32,214,215, summarized by the WHO's Director-General as a "handful of rich countries gobbling up the anticipated supply as manufacturers sell to the highest bidder, while the rest of the world scrambles for the scraps" 216. Notably, the most considerable benefits of COVID-19 vaccination, in terms of averted deaths, have been demonstrated for high-income and upper-middle-income regions, likely due to better logistics, swift rollout, and improved access to highly efficient mRNA vaccines 29. These findings also underline the need for vaccine aid and support in regions of lower income (Figure 4). As estimated, universal vaccination in low-income and lower-middle-income countries with three doses of an mRNA vaccine would avert as much as 1.5 million COVID-19 deaths in the period already dominated by the Omicron lineage 217
One should also note that prior to Omicron's emergence, researchers have continuously warned that vaccine inequity during the COVID-19 pandemic not only reflects a moral crisis but also increases the odds of the emergence of novel, problematic SARS-CoV-2 variants 32. Although the exact origins of the Omicron lineage remain unknown, it is suggested that it may arise during infection of the immunocompromised individual (e.g., HIV/AIDS patient) or even cross-infection between a group of them due to extended viral replication and selection of neutralization resistance mutations in such subjects 218,219. In addition, the study conducted before the emergence of Omicron has shown that the mutation frequency positively correlates with the percentage of unvaccinated individuals in a population, with the highest frequency found for regions with vaccination rates below 10–20%. In turn, the rate of individuals who completed a primary vaccination course in Africa, which has the highest population of people living with HIV (predominantly in the Sub-Saharan area), was approx. 5% at the time of Omicron identification (compared to nearly 55% in the USA, 65% in the European Union, and 50% in Oceania). Although infections with Omicron are milder compared to SARS-CoV-2 lineages preceding it, a lesson must be learned, particularly if one considers that viral genomic surveillance in low-income countries is limited 220. When various health authorities issue novel recommendations regarding COVID-19 vaccinations 220, there is no rationale to shape them differently for low and high-income regions since COVID-19 remains a global issue and should be treated equally regardless of one's origin or ethnicity 215. Importantly, it is crucial to ensure that low-income regions continue to move away from aid dependence through various mechanisms enabling local production of vaccines, including those based on innovative technologies such as mRNA platforms. This could be done by building on the existing capacity, developing sustainable financing mechanisms and quality control systems, prioritizing research funding and regional integration, and collaboration conceptions based on technology co-creation and co-ownership, as discussed elsewhere 147. Improvement of manufacturing and supply of vaccines based on technologies such as mRNA would be important also outside the COVID-19 realm if one considers their potential to deliver preventive tools against other infectious diseases 147,221, some of which are particularly burdensome in low-income countries and have zoonotic origins 222,223.
Simultaneously to vaccine equity, it is essential to improve accessibility to SARS-CoV-2 antivirals in low-income countries, particularly those available in the oral form 224. The first-generation oral antivirals are relatively expensive 225, highlighting the need for aid in delivering these pharmaceuticals to low-income regions, developing generic versions of these drugs, and pursuing efforts to produce them locally 26. This is particularly important since these antivirals can substantially reduce the risk that infected patients will require specialized healthcare, access to which is limited under low-income resources.
All of these efforts require integration with improved education and awareness campaigns to fight vaccine hesitancy, educate on infectious diseases, including COVID-19, and build trust in local authorities and vaccine manufacturers. These goals will also likely require external support, bringing together experience from vaccinations in developed regions and local specificity. Improvement of accessibility to pharmaceuticals and their acceptance in low-income areas should be an integral part of a strategy of pathogen management in high-income countries if one considers that in an increasingly connected modern world, the risks arising from infectious disease can be globally shared 226.
9. Healthcare Workers play a Crucial Role in Maintaining Public COVID-19 Awareness
Healthcare workers are a pivotal part of health communication as they interact directly with patients, including those at high risk of various diseases such as severe COVID-19. Moreover, they serve as role models of healthy behaviors, including vaccination decisions 227. In fact, their role in general COVID-19 awareness may even be more influential during the transition from the acute phase of the pandemic. This is because earlier, the regular communication with patients in this regard was likely curbed due to the significant allocation of healthcare resources to fight COVID-19, social distancing, closures of primary care units, and increased stress experienced by healthcare workers 228–231. At the same time, COVID-19 received high media coverage, with various information on preventive measures often reported daily 232. Vaccination campaigns, lifting sanitary restrictions, the spread of clinically milder Omicron lineage, and the emergence of other issues of public importance (e.g., a war in Ukraine) translated into a decreased interest in COVID-19 in traditional and social media. This may lead to the false assumption that COVID-19 is no longer a threat requiring any preventive measures, e.g., seasonal vaccinations.
Therefore, this is pivotal to ensure that healthcare workers continue their efforts to communicate the risks for particular groups of patients, follow the recommendations on vaccinations, and communicate them further in an understandable manner. As recently stated in the joint statement by the European Centre for Disease Prevention and Control and the European Medicines Agency, seasonal COVID-19 vaccination of healthcare workers should be considered because they have a higher risk of exposure to SARS-CoV-2 while playing a key role in the functioning of healthcare system 175. However, their decision to vaccinate is also likely to be influential for their patients 233. Therefore, ensuring an appropriate education level on COVID-19 vaccines among healthcare workers, including primary physicians, is crucial. As shown in a study led by WHO/Europe, healthcare workers are more confident in recommending COVID-19 vaccines to their patients if they undergo dedicated online training on how to communicate with patients regarding the vaccinations 234. They will also likely be more confident in discussing COVID-19 risks with particular groups of patients after completing the training course, updating them on current SARS-CoV-2 sublineages in circulation and their clinical relevance.
One should note that apart from physicians, the increasingly important role in vaccinations is played by pharmacists 235–237. A study conducted in the US demonstrated that one in four people who refused to receive influenza and pneumococcal vaccines could eventually decide to receive a vaccination after consultation with the pharmacist in pharmacy 238. As calculated in other analyses, including pharmacists in consultation services among seniors for influenza vaccination is cost-effective and improves vaccination rates in this group 239. Therefore, political and organizational barriers should not limit pharmacists' participation in COVID-19 vaccinations. Therefore, the local authorities shall consider increasing the rights of pharmacists in qualifying, prescribing, and vaccinating patients against COVID-19, ultimately simplifying the seasonal vaccination campaigns and likely translating into higher vaccination rates.
10. Conclusions
The acute phase of the COVID-19 pandemic may be over, as reflected by the WHO decision to call off PHEIC in May 2023, but SARS-CoV-2 continues to spread, evolve and cause economic and health burdens. Therefore, as highlighted in the present paper, it requires sustained genomic surveillance and promotion of prevention strategies that are simplified as possible, continuously supported by the healthcare community, and accessible also in low-income regions. All of these elements should be a part of the strategy to adapt to long-term co-existence with SARS-CoV-2 in a manner that prevents healthcare from being overwhelmed.
Author Contributions
Piotr Rzymski: Conceptualization, Project administration, Resources, Writing – original draft. Maria Pokorska-Śpiewak: Writing - review & editing. Teresa Jackowska: Writing - review & editing. Ernest Kuchar: Writing - review & editing. Aneta Nitsch-Osuch: Writing - review & editing. Małgorzata Pawłowska: Writing - review & editing. Mateusz Babicki: Writing - review & editing. Jerzy Jaroszewicz: Writing - review & editing. Leszek Szenborn: Writing - review & editing. Jacek Wysocki: Writing - review & editing. Robert Flisiak: Conceptualization, Supervision.
Funding
Pfizer organized the meeting of the Expert Committee on Prevention and Treatment of Infectious Diseases and funded editorial support to prepare this manuscript, but it had no effect on its content, which remains the sole intellectual property of the authors.
Conflicts of Interest
All authors were members of the Expert Committee on Prevention and Treatment of Infectious Diseases. P.R., T.J., R.F. report consultation and lecture fees from Moderna and Pfizer. E.K. reports consultation and lecture fees from AstraZeneca, GSK, Moderna, Novartis, Novavax, GSK, Pfizer and Sanofi. A.N-S. and J.W. report consultation and lecture fees from Astra Zeneca, GSK, Moderna and Pfizer. J.J. reports consultation and lecture fees from Gilead, Moderna and Pfizer. L.S. reports grants and lecture fees from GSK and Pfizer.
References
- Jiang, S.; Xia, S.; Ying, T.; Lu, L. A novel coronavirus (2019-nCoV) causing pneumonia-associated respiratory syndrome. Cell Mol Immunol. 2020, 17, 554–554. [Google Scholar] [CrossRef] [PubMed]
- Sohrabi, C.; Alsafi, Z.; O’Neill, N. World Health Organization declares global emergency: A review of the 2019 novel coronavirus (COVID-19). Int J Surg. 2020, 76, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Cucinotta, D.; Vanelli, M. WHO declares COVID-19 a pandemic. Acta Biomed. 2020, 91, 157–160. [Google Scholar]
- WHO Statement on the fifteenth meeting of the IHR (2005) Emergency Committee on the COVID-19 pandemic. Published 2023. Accessed , 2023. https://www.who. 7 June 2023.
- Mathieu, E.; Ritchie, H.; Rodés-Guirao, L. Coronavirus Pandemic (COVID-19). Our World in Data, /: Published online , 2020. Accessed December 14, 2022. https, 5 March 2020; 14. [Google Scholar]
- COVID-19 Excess Mortality Collaborators. Estimating excess mortality due to the COVID-19 pandemic: a systematic analysis of COVID-19-related mortality, 2020-21. Lancet. 2022, 399, 1513–1536. [Google Scholar] [CrossRef]
- Nalbandian, A.; Sehgal, K.; Gupta, A. Post-acute COVID-19 syndrome. Nat Med. 2021, 27, 601–615. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Haupert, S.R.; Zimmermann, L.; Shi, X.; Fritsche, L.G.; Mukherjee, B. Global prevalence of post-Coronavirus disease 2019 (COVID-19) condition or long COVID: A meta-analysis and systematic review. J Infect Dis. 2022, 226, 1593–1607. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Leon, S.; Wegman-Ostrosky, T.; Ayuzo Del Valle, N.C. Long-COVID in children and adolescents: a systematic review and meta-analyses. Sci Rep. 2022, 12, 9950. [Google Scholar] [CrossRef]
- Schnitzler, L.; Janssen, L.M.M.; Evers, S.M.A.A. The broader societal impacts of COVID-19 and the growing importance of capturing these in health economic analyses. Int J Technol Assess Health Care. 2021, 37, e43. [Google Scholar] [CrossRef]
- Shang, Y.; Li, H.; Zhang, R. Effects of pandemic outbreak on economies: Evidence from business history context. Front Public Health. 2021, 9, 632043. [Google Scholar] [CrossRef]
- Miller, I.F.; Becker, A.D.; Grenfell, B.T.; Metcalf, C.J.E. Disease and healthcare burden of COVID-19 in the United States. Nat Med. 2020, 26, 1212–1217. [Google Scholar] [CrossRef]
- Lenzen M, Li M, Malik A, et al. Global socio-economic losses and environmental gains from the Coronavirus pandemic. PLoS One. 2020, 15, e0235654. [Google Scholar]
- Sidor, A.; Rzymski, P. Dietary Choices and Habits during COVID-19 Lockdown: Experience from Poland. Nutrients. 2020, 12, 1657. [Google Scholar] [CrossRef] [PubMed]
- Onyeaka H, Anumudu CK, Al-Sharify ZT, Egele-Godswill E, Mbaegbu P. COVID-19 pandemic: A review of the global lockdown and its far-reaching effects. Sci Prog. 2021, 104, 368504211019854. [Google Scholar]
- Nowakowska J, Sobocińska J, Lewicki M, Lemańska Ż, Rzymski P. When science goes viral: The research response during three months of the COVID-19 outbreak. Biomed Pharmacother. 2020, 129, 110451. [Google Scholar]
- Ghebreyesus, T.A.; Swaminathan, S. Scientists are sprinting to outpace the novel coronavirus. Lancet. 2020, 395, 762–764. [Google Scholar] [CrossRef]
- Cusinato, J.; Cau, Y.; Calvani, A.M.; Mori, M. Repurposing drugs for the management of COVID-19. Expert Opin Ther Pat. 2021, 31, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Flisiak R, Zarębska-Michaluk D, Berkan-Kawińska A, et al. Remdesivir-based therapy improved the recovery of patients with COVID-19 in the multicenter, real-world SARSTer study. Pol Arch Intern Med. 2021, 131, 103–110. [Google Scholar]
- Zarębska-Michaluk D, Jaroszewicz J, Rogalska M, et al. Effectiveness of tocilizumab with and without dexamethasone in patients with severe COVID-19: A retrospective study. J Inflamm Res. 2021, 14, 3359–3366. [Google Scholar] [CrossRef]
- Flisiak, R.; Flisiak-Jackiewicz, M.; Rzymski, P.; Zarębska-Michaluk, D. Tocilizumab for the treatment of COVID-19. Expert Rev Anti Infect Ther, 16 June 2023. [Google Scholar] [CrossRef]
- Moniuszko-Malinowska A, Czupryna P, Zarębska-Michaluk D, et al. Convalescent plasma transfusion for the treatment of COVID-19-experience from Poland: A multicenter study. J Clin Med. 2020, 10, 28. [Google Scholar] [CrossRef]
- Simonovich VA, Burgos Pratx LD, Scibona P, et al. A randomized trial of convalescent plasma in Covid-19 severe pneumonia. N Engl J Med. 2021, 384, 619–629. [Google Scholar] [CrossRef]
- Jiang, S.; Hillyer, C.; Du, L. Neutralizing antibodies against SARS-CoV-2 and other human coronaviruses. Trends Immunol. 2020, 41, 355–359. [Google Scholar] [CrossRef]
- 25. Brobst B, Borger J. Benefits and Risks of Administering Monoclonal Antibody Therapy for Coronavirus (COVID-19), 2023.
- 26. Rahmah L, Abarikwu SO, Arero AG, et al. Oral antiviral treatments for COVID-19: opportunities and challenges. Pharmacological Reports, 2022. [CrossRef]
- Thanh Le T, Andreadakis Z, Kumar A, et al. The COVID-19 vaccine development landscape. Nat Rev Drug Discov. 2020, 19, 305–306. [Google Scholar] [CrossRef]
- Rzymski P, Borkowski L, Drąg M, et al. The strategies to support the COVID-19 vaccination with evidence-based communication and tackling misinformation. Vaccines (Basel). 2021, 9, 109. [Google Scholar] [CrossRef]
- Watson, O.J.; Barnsley, G.; Toor, J.; Hogan, A.B.; Winskill, P.; Ghani, A.C. Global impact of the first year of COVID-19 vaccination: a mathematical modelling study. Lancet Infect Dis. 2022, 22, 1293–1302. [Google Scholar] [CrossRef]
- Zheng, C.; Shao, W.; Chen, X.; Zhang, B.; Wang, G.; Zhang, W. Real-world effectiveness of COVID-19 vaccines: a literature review and meta-analysis. Int J Infect Dis. 2022, 114, 252–260. [Google Scholar] [CrossRef] [PubMed]
- 31. Rzymski P, Kasianchuk N, Sikora D, Poniedziałek B. COVID-19 vaccinations and rates of infections, hospitalizations, ICU admissions, and deaths in Europe during SARS-CoV-2 Omicron wave in the first quarter of 2022. J Med Virol, 6 September 2022. [CrossRef]
- Rzymski P, Camargo CA, Fal A, et al. COVID-19 Vaccine Boosters: The Good, the Bad, and the Ugly. Vaccines. 2021, 9, 1299. [Google Scholar] [CrossRef] [PubMed]
- 33. Woldemeskel BA, Garliss CC, Blankson JN. mRNA Vaccine-Elicited SARS-CoV-2-Specific T cells Persist at 6 Months and Recognize the Delta Variant. Clin Infect Dis, 25 October 2021. [CrossRef]
- Jordan SC, Shin BH, Gadsden TAM, et al. T cell immune responses to SARS-CoV-2 and variants of concern (Alpha and Delta) in infected and vaccinated individuals. Cell Mol Immunol. 2021, 18, 2554–2556. [Google Scholar] [CrossRef]
- 35. Jergovic M, Coplen CP, Uhrlaub JL, et al. Resilient T cell responses to B.1.1.529 (Omicron) SARS-CoV-2 variant. bioRxiv, 2: , 2022, 16 January 2022. [CrossRef]
- Collier DA, Ferreira IATM, Kotagiri P, et al. Age-related immune response heterogeneity to SARS-CoV-2 vaccine BNT162b2. Nature. 2021, 596, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Brosh-Nissimov T, Orenbuch-Harroch E, Chowers M, et al. BNT162b2 vaccine breakthrough: clinical characteristics of 152 fully vaccinated hospitalized COVID-19 patients in Israel. Clin Microbiol Infect. 2021, 27, 1652–1657. [Google Scholar] [CrossRef] [PubMed]
- Hallam, J.; Jones, T.; Alley, J.; Kohut, M.L. Exercise after influenza or COVID-19 vaccination increases serum antibody without an increase in side effects. Brain Behav Immun. 2022, 102, 1–10. [Google Scholar] [CrossRef]
- Rzymski P, Pazgan-Simon M, Kamerys J, et al. Severe breakthrough COVID-19 cases during six months of delta variant (B.1.617.2) domination in Poland. Vaccines (Basel). 2022, 10, 557. [Google Scholar] [CrossRef] [PubMed]
- WHO World malaria report 2022. Published 2023. https://apps.who. 1484.
- Markov PV, Ghafari M, Beer M, et al. The evolution of SARS-CoV-2. Nat Rev Microbiol. 2023, 21, 361–379. [Google Scholar] [CrossRef]
- Strassburg, M.A. The global eradication of smallpox. Am J Infect Control. 1982, 10, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Combe, M.; Sanjuán, R. Variation in RNA virus mutation rates across host cells. PLoS Pathog. 2014, 10, e1003855. [Google Scholar] [CrossRef] [PubMed]
- Amicone M, Borges V, Alves MJ, et al. Mutation rate of SARS-CoV-2 and emergence of mutators during experimental evolution. Evol Med Public Health. 2022, 10, 142–155. [Google Scholar] [CrossRef]
- Manzanares-Meza LD, Medina-Contreras O. SARS-CoV-2 and influenza: a comparative overview and treatment implications. Bol Med Hosp Infant Mex. 2020, 77, 262–273. [Google Scholar]
- Rawson JMO, Landman SR, Reilly CS, Mansky LM. HIV-1 and HIV-2 exhibit similar mutation frequencies and spectra in the absence of G-to-A hypermutation. Retrovirology. 2015, 12, 60. [Google Scholar] [CrossRef]
- Nakamura F, Takeda H, Ueda Y, et al. Mutational spectrum of hepatitis C virus in patients with chronic hepatitis C determined by single molecule real-time sequencing. Sci Rep. 2022, 12, 7083. [Google Scholar] [CrossRef]
- Korber B, Fischer WM, Gnanakaran S, et al. Tracking changes in SARS-CoV-2 Spike: Evidence that D614G increases infectivity of the COVID-19 virus. Cell. 2020, 182, 812–827. [Google Scholar] [CrossRef]
- Jackson B, Boni MF, Bull MJ, et al. Generation and transmission of interlineage recombinants in the SARS-CoV-2 pandemic. Cell. 2021, 184, 5179–5188. [Google Scholar] [CrossRef]
- Chakraborty, C.; Bhattacharya, M.; Chopra, H.; Islam, M.A.; Saikumar, G.; Dhama, K. The SARS-CoV-2 Omicron recombinant subvariants XBB, XBB.1, and XBB.1.5 are expanding rapidly with unique mutations, antibody evasion, and immune escape properties - an alarming global threat of a surge in COVID-19 cases again? Int J Surg. 2023, 109, 1041–1043. [Google Scholar] [CrossRef] [PubMed]
- Parums, D.V. Editorial: The XBB.1.5 ('Kraken’) subvariant of Omicron SARS-CoV-2 and its rapid global spread. Med Sci Monit. 2023, 29, e939580. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Chen, J.; Wei, G.W. Mechanisms of SARS-CoV-2 evolution revealing vaccine-resistant mutations in Europe and America. J Phys Chem Lett. 2021, 12, 11850–11857. [Google Scholar] [CrossRef]
- Sadarangani, M.; Marchant, A.; Kollmann, T.R. Immunological mechanisms of vaccine-induced protection against COVID-19 in humans. Nat Rev Immunol. 2021, 21, 475–484. [Google Scholar] [CrossRef]
- 54. Lasrado N, Collier ARY, Miller J, et al. Waning immunity against XBB.1.5 following bivalent mRNA boosters. bioRxivorg, 2: , 2023, 23 January 2023.
- 55. Muik A, Lui BG, Diao H, et al. Progressive loss of conserved spike protein neutralizing antibody sites in Omicron sublineages is balanced by preserved T-cell recognition epitopes. bioRxiv, 2: , 2022, 15 December 2022. [CrossRef]
- 56. Abbasian MH, Mahmanzar M, Rahimian K, et al. Global landscape of SARS-CoV-2 mutations and conserved regions. J Transl Med. [CrossRef]
- McCafferty S, Haque AKMA, Vandierendonck A, et al. A dual-antigen self-amplifying RNA SARS-CoV-2 vaccine induces potent humoral and cellular immune responses and protects against SARS-CoV-2 variants through T cell-mediated immunity. Mol Ther. 2022, 30, 2968–2983. [Google Scholar] [CrossRef] [PubMed]
- Nextstrain. Genomic epidemiology of SARS-CoV-2 with subsampling focused globally since pandemic start. Published 2023. Accessed , 2023. https://nextstrain. 14 June.
- Viana R, Moyo S, Amoako DG, et al. Rapid epidemic expansion of the SARS-CoV-2 Omicron variant in southern Africa. Nature. 2022, 603, 679–686. [Google Scholar] [CrossRef]
- Arora P, Zhang L, Rocha C, et al. Comparable neutralisation evasion of SARS-CoV-2 omicron subvariants BA.1, BA.2, and BA.3. Lancet Infect Dis. 2022, 22, 766–767. [Google Scholar] [CrossRef]
- 61. Liu L, Iketani S, Guo Y, et al. Striking antibody evasion manifested by the Omicron variant of SARS-CoV-2. Nature, 23 December 2021. [CrossRef]
- Hui KPY, Ho JCW, Cheung MC, et al. SARS-CoV-2 Omicron variant replication in human bronchus and lung ex vivo. Nature. 2022, 603, 715–720. [Google Scholar] [CrossRef]
- Suzuki R, Yamasoba D, Kimura I, et al. Attenuated fusogenicity and pathogenicity of SARS-CoV-2 Omicron variant. Nature. 2022, 603, 700–705. [Google Scholar] [CrossRef]
- Wang XJ, Yao L, Zhang HY, et al. Neutralization sensitivity, fusogenicity, and infectivity of Omicron subvariants. Genome Med. 2022, 14, 146. [Google Scholar] [CrossRef]
- Halfmann PJ, Iida S, Iwatsuki-Horimoto K, et al. SARS-CoV-2 Omicron virus causes attenuated disease in mice and hamsters. Nature. 2022, 603, 687–692. [Google Scholar] [CrossRef] [PubMed]
- Abdelnabi R, Foo CS, Zhang X, et al. The omicron (B.1.1.529) SARS-CoV-2 variant of concern does not readily infect Syrian hamsters. Antiviral Res. 2022, 198, 105253. [Google Scholar] [CrossRef] [PubMed]
- McMahan K, Giffin V, Tostanoski LH, et al. Reduced pathogenicity of the SARS-CoV-2 omicron variant in hamsters. Med (N Y). 2022, 3, 262–268. [Google Scholar]
- Chan, J.F.W.; Chu, H. Pathogenicity of SARS-CoV-2 Omicron BA.1.1 in hamsters. EBioMedicine. 2022, 80, 104035. [Google Scholar] [CrossRef] [PubMed]
- Yu W, Wang J, Yang Y, et al. SARS-CoV-2 Omicron (B.1.1.529) infection in rhesus macaques, hamsters, and BALB/c mice with severe lung histopathological damage. J Med Virol. 2023, 95, e28846. [Google Scholar] [CrossRef]
- Menni C, Valdes AM, Polidori L, et al. Symptom prevalence, duration, and risk of hospital admission in individuals infected with SARS-CoV-2 during periods of omicron and delta variant dominance: a prospective observational study from the ZOE COVID Study. Lancet. 2022, 399, 1618–1624. [Google Scholar] [CrossRef]
- Flisiak R, Rzymski P, Zarębska-Michaluk D, et al. Variability in the clinical course of COVID-19 in a retrospective analysis of a large real-world database. Viruses. 2023, 15, 149. [Google Scholar] [CrossRef]
- 72. Consolazio D, Murtas R, Tunesi S, et al. A comparison between Omicron and earlier COVID-19 variants’ disease severity in the Milan area, Italy. Front Epidemiol. [CrossRef]
- Nyberg T, Ferguson NM, Nash SG, et al. Comparative analysis of the risks of hospitalisation and death associated with SARS-CoV-2 omicron (B.1.1.529) and delta (B.1.617.2) variants in England: a cohort study. Lancet. 2022, 399, 1303–1312. [Google Scholar] [CrossRef]
- Bager P, Wohlfahrt J, Bhatt S, et al. Risk of hospitalisation associated with infection with SARS-CoV-2 omicron variant versus delta variant in Denmark: an observational cohort study. Lancet Infect Dis. 2022, 22, 967–976. [Google Scholar] [CrossRef]
- Jassat W, Abdool Karim SS, Ozougwu L, et al. Trends in cases, hospitalizations, and mortality related to the Omicron BA.4/BA.5 subvariants in South Africa. Clin Infect Dis. 2023, 76, 1468–1475. [Google Scholar] [CrossRef]
- 76. Pung R, Kong XP, Cui L, et al. Severity of SARS-CoV-2 Omicron XBB subvariants in Singapore. medRxiv, 2: , 2023, 10 May 2023. [CrossRef]
- 77. Karyakarte RP, Das R, Rajmane MV, et al. Chasing SARS-CoV-2 XBB.1.16 recombinant lineage in India and the clinical profile of XBB.1.16 cases in Maharashtra, India. medRxiv, 2: , 2023, 26 April 2023. [CrossRef]
- World Health Organization. Influenza. Published 2023. Accessed , 2023. https://www.who. 14 June.
- Portmann L, de Kraker MEA, Fröhlich G, et al. Hospital outcomes of community-acquired SARS-CoV-2 Omicron variant infection compared with influenza infection in Switzerland. JAMA Netw Open. 2023, 6, e2255599. [Google Scholar] [CrossRef] [PubMed]
- Taylor CA, Whitaker M, Anglin O, et al. COVID-19-associated hospitalizations among adults during SARS-CoV-2 Delta and Omicron variant predominance, by race/ethnicity and vaccination status - COVID-NET, 14 states, July 2021-January 2022. MMWR Morb Mortal Wkly Rep. 2022, 71, 466–473. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, M.; Pujol, J.C.; Spector, T.D.; Ourselin, S.; Steves, C.J. Risk of long COVID associated with delta versus omicron variants of SARS-CoV-2. Lancet. 2022, 399, 2263–2264. [Google Scholar] [CrossRef]
- Thaweethai T, Jolley SE, Karlson EW, et al. Development of a definition of postacute sequelae of SARS-CoV-2 infection. JAMA. 2023, 329, 1934–1946. [Google Scholar] [CrossRef]
- Li Wan Po, A. Omicron variant as nature’s solution to the COVID-19 pandemic. J Clin Pharm Ther. 2022, 47, 3–5. [Google Scholar] [CrossRef]
- 84. Pascall DJ, Vink E, Blacow R, et al. Directions of change in intrinsic case severity across successive SARS-CoV-2 variant waves have been inconsistent. J Infect, 1 June 2023. [CrossRef]
- Earnest R, Uddin R, Matluk N, et al. Comparative transmissibility of SARS-CoV-2 variants Delta and Alpha in New England, USA. Cell Rep Med. 2022, 3, 100583. [Google Scholar] [CrossRef] [PubMed]
- King KL, Wilson S, Napolitano JM, et al. SARS-CoV-2 variants of concern Alpha and Delta show increased viral load in saliva. PLoS One. 2022, 17, e0267750. [Google Scholar]
- Yuasa S, Nakajima J, Takatsuki Y, et al. Viral load of SARS-CoV-2 Omicron is not high despite its high infectivity. J Med Virol. 2022, 94, 5543–5546. [Google Scholar] [CrossRef]
- Laitman, A.M.; Lieberman, J.A.; Hoffman, N.G.; Roychoudhury, P.; Mathias, P.C.; Greninger, A.L. The SARS-CoV-2 omicron variant does not have higher nasal viral loads compared to the delta variant in symptomatic and asymptomatic individuals. J Clin Microbiol. 2022, 60, e0013922. [Google Scholar] [CrossRef]
- Puhach O, Adea K, Hulo N, et al. Infectious viral load in unvaccinated and vaccinated individuals infected with ancestral, Delta or Omicron SARS-CoV-2. Nat Med. 2022, 28, 1491–1500. [Google Scholar] [CrossRef]
- Wu L, Zhou L, Mo M, et al. SARS-CoV-2 Omicron RBD shows weaker binding affinity than the currently dominant Delta variant to human ACE2. Signal Transduct Target Ther. 2022, 7, 8. [Google Scholar] [CrossRef]
- Meng B, Abdullahi A, Ferreira IATM, et al. Altered TMPRSS2 usage by SARS-CoV-2 Omicron impacts infectivity and fusogenicity. Nature. 2022, 603, 706–714. [Google Scholar] [CrossRef]
- He X, Lau EHY, Wu P, et al. Temporal dynamics in viral shedding and transmissibility of COVID-19. Nat Med. 2020, 26, 672–675. [Google Scholar] [CrossRef] [PubMed]
- Fajnzylber J, Regan J, Coxen K, et al. SARS-CoV-2 viral load is associated with increased disease severity and mortality. Nat Commun. 2020, 11, 5493. [Google Scholar] [CrossRef] [PubMed]
- Saito A, Irie T, Suzuki R, et al. Enhanced fusogenicity and pathogenicity of SARS-CoV-2 Delta P681R mutation. Nature. 2022, 602, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Kimura I, Yamasoba D, Tamura T, et al. Virological characteristics of the SARS-CoV-2 Omicron BA.2 subvariants, including BA.4 and BA.5. Cell. 2022, 185, 3992–4007. [Google Scholar] [CrossRef]
- Xia S, Wang L, Jiao F, et al. SARS-CoV-2 Omicron subvariants exhibit distinct fusogenicity, but similar sensitivity, to pan-CoV fusion inhibitors. Emerg Microbes Infect. 2023, 12, 2178241. [Google Scholar] [CrossRef]
- Xia S, Jiao F, Wang L, et al. SARS-CoV-2 Omicron XBB subvariants exhibit enhanced fusogenicity and substantial immune evasion in elderly population, but high sensitivity to pan-coronavirus fusion inhibitors. J Med Virol. 2023, 95, e28641. [Google Scholar] [CrossRef]
- Yuan S, Ye ZW, Liang R, et al. Pathogenicity, transmissibility, and fitness of SARS-CoV-2 Omicron in Syrian hamsters. Science. 2022, 377, 428–433. [Google Scholar] [CrossRef]
- Wolter N, Jassat W, Walaza S, et al. Clinical severity of SARS-CoV-2 Omicron BA.4 and BA.5 lineages compared to BA.1 and Delta in South Africa. Nat Commun. 2022, 13, 5860. [Google Scholar] [CrossRef]
- Robertson, C.; Kerr, S.; Sheikh, A. Severity of Omicron BA.5 variant and protective effect of vaccination: national cohort and matched analyses in Scotland. Lancet Reg Health Eur. 2023, 28, 100638. [Google Scholar] [CrossRef]
- Ciuffreda L, Lorenzo-Salazar JM, García-Martínez de Artola D, et al. Reinfection rate and disease severity of the BA.5 Omicron SARS-CoV-2 lineage compared to previously circulating variants of concern in the Canary Islands (Spain). Emerg Microbes Infect. 2023, 12, 2202281. [Google Scholar] [CrossRef] [PubMed]
- Oude Munnink BB, Sikkema RS, Nieuwenhuijse DF, et al. Transmission of SARS-CoV-2 on mink farms between humans and mink and back to humans. Science. 2021, 371, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann M, Zhang L, Krüger N, et al. SARS-CoV-2 mutations acquired in mink reduce antibody-mediated neutralization. Cell Rep. 2021, 35, 109017. [Google Scholar] [CrossRef] [PubMed]
- Domańska-Blicharz K, Orłowska A, Smreczak M, et al. Mink SARS-CoV-2 infection in Poland - short communication. J Vet Res. 2021, 65, 1–5. [Google Scholar] [CrossRef]
- 105. Palmer MV, Martins M, Falkenberg S, et al. Susceptibility of white-tailed deer (Odocoileus virginianus) to SARS-CoV-2. J Virol. [CrossRef]
- Chandler JC, Bevins SN, Ellis JW, et al. SARS-CoV-2 exposure in wild white-tailed deer (Odocoileus virginianus). Proc Natl Acad Sci U S A. 2021, 118, e2114828118. [Google Scholar] [CrossRef]
- Li L, Han P, Huang B, et al. Broader-species receptor binding and structural bases of Omicron SARS-CoV-2 to both mouse and palm-civet ACE2s. Cell Discov. 2022, 8, 65. [Google Scholar] [CrossRef]
- Konishi, T. SARS-CoV-2 mutations among minks show reduced lethality and infectivity to humans. PLoS One. 2021, 16, e0247626. [Google Scholar] [CrossRef]
- Devaux, C.A.; Pinault, L.; Delerce, J.; Raoult, D.; Levasseur, A.; Frutos, R. Spread of mink SARS-CoV-2 variants in humans: A model of sarbecovirus interspecies evolution. Front Microbiol, 6: 12, 6755. [Google Scholar]
- Willgert K, Didelot X, Surendran-Nair M, et al. Transmission history of SARS-CoV-2 in humans and white-tailed deer. Sci Rep. 2022, 12, 12094. [Google Scholar] [CrossRef]
- Kuchipudi SV, Surendran-Nair M, Ruden RM, et al. Multiple spillovers from humans and onward transmission of SARS-CoV-2 in white-tailed deer. Proc Natl Acad Sci U S A. 2022, 119, e2121644119. [Google Scholar] [CrossRef]
- Gupta, S.; Cantor, J.; Simon, K.I.; Bento, A.I.; Wing, C.; Whaley, C.M. Vaccinations against COVID-19 May have averted up to 140,000 deaths in the United States. Health Aff (Millwood). 2021, 40, 1465–1472. [Google Scholar] [CrossRef] [PubMed]
- Kayano T, Sasanami M, Kobayashi T, et al. Number of averted COVID-19 cases and deaths attributable to reduced risk in vaccinated individuals in Japan. Lancet Reg Health West Pac. 2022, 28, 100571. [Google Scholar] [CrossRef] [PubMed]
- Mattiuzzi C, Henry BM, Lippi G. COVID-19 vaccination uptake strongly predicts averted deaths of older people across Europe. Biomed J. 2022, 45, 961–962. [Google Scholar] [CrossRef] [PubMed]
- Sikora D, Rzymski P. COVID-19 vaccination and rates of infections, hospitalizations, ICU admissions, and deaths in the European Economic Area during autumn 2021 wave of SARS-CoV-2. Vaccines (Basel). 2022, 10, 437. [Google Scholar] [CrossRef]
- Yi S, Choe YJ, Lim DS, et al. Impact of national Covid-19 vaccination Campaign, South Korea. Vaccine. 2022, 40, 3670–3675. [Google Scholar] [CrossRef] [PubMed]
- McCarthy CV, O’Mara O, van Leeuwen E, CMMID COVID-19 Working Group, Jit M, Sandmann F. The impact of COVID-19 vaccination in prisons in England and Wales: a metapopulation model. BMC Public Health. 2022, 22, 1003. [Google Scholar]
- Santos CVBD, Noronha TG de, Werneck GL, Struchiner CJ, Villela DAM. Estimated COVID-19 severe cases and deaths averted in the first year of the vaccination campaign in Brazil: A retrospective observational study. Lancet Reg Health Am. 2023, 17, 100418. [Google Scholar]
- Haas EJ, McLaughlin JM, Khan F, et al. Infections, hospitalisations, and deaths averted via a nationwide vaccination campaign using the Pfizer-BioNTech BNT162b2 mRNA COVID-19 vaccine in Israel: a retrospective surveillance study. Lancet Infect Dis. 2022, 22, 357–366. [Google Scholar] [CrossRef]
- 120. Sacco C, Mateo-Urdiales A, Petrone D, et al. Estimating averted COVID-19 cases, hospitalisations, intensive care unit admissions and deaths by COVID-19 vaccination, Italy, January-September 2021. Euro Surveill, 20 September. [CrossRef]
- Brannock MD, Chew RF, Preiss AJ, et al. Long COVID risk and pre-COVID vaccination in an EHR-based cohort study from the RECOVER program. Nat Commun. 2023, 14, 2914. [Google Scholar] [CrossRef]
- Simon, M.A.; Luginbuhl, R.D.; Parker, R. Reduced incidence of long-COVID symptoms related to administration of COVID-19 vaccines both before COVID-19 diagnosis and up to 12 weeks after. bioRxiv, 2: online , 2021, 18 November 2021. [Google Scholar] [CrossRef]
- Antonelli M, Penfold RS, Merino J, et al. Risk factors and disease profile of post-vaccination SARS-CoV-2 infection in UK users of the COVID Symptom Study app: a prospective, community-based, nested, case-control study. Lancet Infect Dis. 2022, 22, 43–55. [Google Scholar] [CrossRef]
- 124. Senjam SS, Balhara YPS, Kumar P, et al. Assessment of Post COVID-19 Health Problems and its Determinants in North India: A descriptive cross section study. bioRxiv, 2: 2021, 7 October 2021. [CrossRef]
- Ayoubkhani D, Bosworth ML, King S, et al. Risk of long COVID in people infected with severe acute respiratory syndrome Coronavirus 2 after 2 doses of a Coronavirus disease 2019 vaccine: Community-based, matched cohort study. Open Forum Infect Dis. 2022, 9, ofac464. [Google Scholar] [CrossRef] [PubMed]
- Al-Aly, Z.; Bowe, B.; Xie, Y. Long COVID after breakthrough SARS-CoV-2 infection. Nat Med. 2022, 28, 1461–1467. [Google Scholar] [CrossRef] [PubMed]
- Taquet, M.; Dercon, Q.; Harrison, P.J. Six-month sequelae of post-vaccination SARS-CoV-2 infection: A retrospective cohort study of 10,024 breakthrough infections. Brain Behav Immun. 2022, 103, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Notarte KI, Catahay JA, Velasco JV, et al. Impact of COVID-19 vaccination on the risk of developing long-COVID and on existing long-COVID symptoms: A systematic review. EClinicalMedicine. 2022, 53, 101624. [Google Scholar] [CrossRef]
- Yu X, Wei D, Xu W, et al. Reduced sensitivity of SARS-CoV-2 Omicron variant to antibody neutralization elicited by booster vaccination. Cell Discov. 2022, 8, 4. [Google Scholar] [CrossRef]
- Yu J, Collier ARY, Rowe M, et al. Neutralization of the SARS-CoV-2 omicron BA.1 and BA.2 variants. N Engl J Med. 2022, 386, 1579–1580. [Google Scholar] [CrossRef]
- Lau JJ, Cheng SMS, Leung K, et al. Real-world COVID-19 vaccine effectiveness against the Omicron BA.2 variant in a SARS-CoV-2 infection-naive population. Nat Med. 2023, 29, 348–357. [Google Scholar] [CrossRef]
- Mohammed H, Pham-Tran DD, Yeoh ZYM, et al. A systematic review and meta-analysis on the real-world effectiveness of COVID-19 vaccines against infection, symptomatic and severe COVID-19 disease caused by the Omicron variant (B.1.1.529). Vaccines (Basel). 2023, 11, 224. [Google Scholar] [CrossRef]
- Zarębska-Michaluk D, Hu C, Brzdęk M, Flisiak R, Rzymski P. COVID-19 vaccine booster strategies for Omicron SARS-CoV-2 variant: Effectiveness and future prospects. Vaccines (Basel). 2022, 10, 1223. [Google Scholar] [CrossRef]
- Solante R, Alvarez-Moreno C, Burhan E, et al. Expert review of global real-world data on COVID-19 vaccine booster effectiveness and safety during the omicron-dominant phase of the pandemic. Expert Rev Vaccines. 2023, 22, 1–16. [Google Scholar] [CrossRef]
- Wu N, Joyal-Desmarais K, Ribeiro PAB, et al. Long-term effectiveness of COVID-19 vaccines against infections, hospitalisations, and mortality in adults: findings from a rapid living systematic evidence synthesis and meta-analysis up to December, 2022. Lancet Respir Med. 2023, 11, 439–452. [Google Scholar] [CrossRef]
- Li, Y.; Liang, H.; Ding, X.; Cao, Y.; Yang, D.; Duan, Y. Effectiveness of COVID-19 vaccine in children and adolescents with the Omicron variant: A systematic review and meta-analysis. J Infect. 2023, 86, e64–e66. [Google Scholar] [CrossRef]
- 137. Mendes D, Chapman R, Aruffo E, et al. Public health impact of UK COVID-19 booster vaccination programs during Omicron predominance. Expert Rev Vaccines, 1: 2023, 3 January 2023.
- Link-Gelles R, Ciesla AA, Fleming-Dutra KE, et al. Effectiveness of bivalent mRNA vaccines in preventing symptomatic SARS-CoV-2 infection - Increasing Community Access to testing program, United States, September-November 2022. MMWR Morb Mortal Wkly Rep. 2022, 71, 1526–1530. [Google Scholar] [CrossRef]
- Link-Gelles R, Ciesla AA, Roper LE, et al. Early estimates of bivalent mRNA booster dose vaccine effectiveness in preventing symptomatic SARS-CoV-2 infection attributable to Omicron BA.5- and XBB/XBB.1.5-related sublineages among immunocompetent adults - Increasing Community Access to testing program, United States, December 2022-January 2023. MMWR Morb Mortal Wkly Rep. 2023, 72, 119–124. [Google Scholar]
- 140. Arbel R, Peretz A, Sergienko R, et al. Effectiveness of a bivalent mRNA vaccine booster dose to prevent severe COVID-19 outcomes: a retrospective cohort study. Lancet Infect Dis, 13 April 2023. [CrossRef]
- Collier ARY, Miller J, Hachmann NP, et al. Immunogenicity of BA.5 bivalent mRNA vaccine boosters. N Engl J Med. 2023, 388, 565–567. [Google Scholar] [CrossRef] [PubMed]
- Offit, P.A. Bivalent covid-19 vaccines - A cautionary tale. N Engl J Med. 2023, 388, 481–483. [Google Scholar] [CrossRef] [PubMed]
- Wang Q, Bowen A, Valdez R, et al. Antibody response to omicron BA.4-BA.5 bivalent booster. N Engl J Med. 2023, 388, 567–569. [Google Scholar] [CrossRef] [PubMed]
- 144. Carreño JM, Singh G, Simon V, Krammer F, PVI study group. Bivalent COVID-19 booster vaccines and the absence of BA.5-specific antibodies. Lancet Microbe, 1 May 2023. [CrossRef]
- WHO Statement on the antigen composition of COVID-19 vaccines. Published 2023. Accessed , 2023. https://www.who. 18 June 2023.
- Ballouz T, Menges D, Kaufmann M, et al. Post COVID-19 condition after Wildtype, Delta, and Omicron SARS-CoV-2 infection and prior vaccination: Pooled analysis of two population-based cohorts. PLoS One. 2023, 18, e0281429. [Google Scholar]
- 147. Rzymski P, Szuster-Ciesielska A, Dzieciątkowski T, Gwenzi W, Fal A. mRNA vaccines: The future of prevention of viral infections? J Med Virol, 10 February 2023. [CrossRef]
- Hajnik RL, Plante JA, Liang Y, et al. Dual spike and nucleocapsid mRNA vaccination confer protection against SARS-CoV-2 Omicron and Delta variants in preclinical models. Sci Transl Med. 2022, 14, eabq1945. [Google Scholar] [CrossRef]
- Alu, A.; Chen, L.; Lei, H.; Wei, Y.; Tian, X.; Wei, X. Intranasal COVID-19 vaccines: From bench to bed. EBioMedicine. 2022, 76, 103841. [Google Scholar] [CrossRef]
- Ramvikas, M.; Arumugam, M.; Chakrabarti, S.R.; Jaganathan, K.S. Nasal Vaccine Delivery. In: Micro and Nanotechnology in Vaccine Development. Elsevier; 2017:279-301.
- Sengupta A, Azharuddin M, Cardona ME, et al. Intranasal Coronavirus SARS-CoV-2 immunization with lipid adjuvants provides systemic and mucosal immune response against SARS-CoV-2 S1 spike and nucleocapsid protein. Vaccines (Basel). 2022, 10, 504. [Google Scholar] [CrossRef] [PubMed]
- Shim S, Soh SH, Im YB, et al. Induction of systemic immunity through nasal-associated lymphoid tissue (NALT) of mice intranasally immunized with Brucella abortus malate dehydrogenase-loaded chitosan nanoparticles. PLoS One. 2020, 15, e0228463. [Google Scholar]
- McLenon, J.; Rogers, M.A.M. The fear of needles: A systematic review and meta-analysis. J Adv Nurs. 2019, 75, 30–42. [Google Scholar] [CrossRef]
- Oladoye, M.J. Intranasal vaccines: a panacea to vaccine hesitancy? Med Res J. 2022, 7, 274–275. [Google Scholar] [CrossRef]
- Dhama K, Dhawan M, Tiwari R, et al. COVID-19 intranasal vaccines: current progress, advantages, prospects, and challenges. Hum Vaccin Immunother. 2022, 18, 2045853. [Google Scholar] [CrossRef] [PubMed]
- Madhavan M, Ritchie AJ, Aboagye J, et al. Tolerability and immunogenicity of an intranasally-administered adenovirus-vectored COVID-19 vaccine: An open-label partially-randomised ascending dose phase I trial. EBioMedicine. 2022, 85, 104298. [Google Scholar]
- van Doremalen N, Purushotham JN, Schulz JE, et al. Intranasal ChAdOx1 nCoV-19/AZD1222 vaccination reduces viral shedding after SARS-CoV-2 D614G challenge in preclinical models. Sci Transl Med. 2021, 13, eabh0755. [Google Scholar] [CrossRef]
- Ndeupen, S.; Qin, Z.; Jacobsen, S.; Bouteau, A.; Estanbouli, H.; Igyártó, B.Z. The mRNA-LNP platform’s lipid nanoparticle component used in preclinical vaccine studies is highly inflammatory. iScience. 2021, 24, 103479. [Google Scholar] [CrossRef]
- Đaković Rode O, Bodulić K, Zember S, et al. Decline of anti-SARS-CoV-2 IgG antibody levels 6 months after complete BNT162b2 vaccination in healthcare workers to levels observed following the first vaccine dose. Vaccines (Basel). 2022, 10, 153. [Google Scholar] [CrossRef]
- Hosseinian S, de Assis R, Khalil G, et al. Analysis and comparison of SARS-CoV-2 variant antibodies and neutralizing activity for 6 months after a booster mRNA vaccine in a healthcare worker population. Front Immunol. 2023, 14, 1166261. [Google Scholar] [CrossRef]
- Silva, M.F.S.; Pinto, A.C.M.D.; de Oliveira F de, C.E.; Caetano, L.F.; Araújo FM de, C.; Fonseca, M.H.G. Antibody response 6 months after the booster dose of Pfizer in previous recipients of CoronaVac. J Med Virol. 2023, 95, e28169. [Google Scholar] [CrossRef] [PubMed]
- Townsend, J.P.; Hassler, H.B.; Dornburg, A. Infection by SARS-CoV-2 with alternate frequencies of mRNA vaccine boosting. J Med Virol. 2023, 95, e28461. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. COVID-19 Vaccine Tracker. Published 2023. Accessed , 2022. https://vaccinetracker.ecdc.europa.eu/public/extensions/COVID-19/vaccine-tracker. 31 January.
- Fieselmann, J.; Annac, K.; Erdsiek, F.; Yilmaz-Aslan, Y.; Brzoska, P. What are the reasons for refusing a COVID-19 vaccine? A qualitative analysis of social media in Germany. BMC Public Health. 2022, 22, 846. [Google Scholar] [CrossRef]
- Rzymski, P.; Poniedziałek, B.; Fal, A. Willingness to receive the booster COVID-19 vaccine dose in Poland. Vaccines (Basel). 2021, 9, 1286. [Google Scholar] [CrossRef] [PubMed]
- Rzymski P, Sikora D, Zeyland J, et al. Frequency and nuisance level of adverse events in individuals receiving homologous and heterologous COVID-19 booster vaccine. Vaccines (Basel). 2022, 10, 754. [Google Scholar] [CrossRef]
- Sobierajski, T.; Rzymski, P.; Wanke-Rytt, M. Impact of the COVID-19 pandemic on attitudes toward vaccination: Representative study of polish society. Vaccines (Basel). 2023, 11, 1069. [Google Scholar] [CrossRef]
- Capurro, G.; Tustin, J.; Jardine, C.G.; Driedger, S.M. When good messages go wrong: Perspectives on COVID-19 vaccines and vaccine communication from generally vaccine accepting individuals in Canada. Hum Vaccin Immunother. 2022, 18, 2145822. [Google Scholar] [CrossRef]
- Hernandez, N.; Caetano-Anollés, G. Worldwide correlations support COVID-19 seasonal behavior and impact of global change. Evol Bioinform Online. 2023, 19, 11769343231169376. [Google Scholar] [CrossRef]
- Gavenčiak T, Monrad JT, Leech G, et al. Seasonal variation in SARS-CoV-2 transmission in temperate climates: A Bayesian modelling study in 143 European regions. PLoS Comput Biol. 2022, 18, e1010435. [Google Scholar]
- Wiemken TL, Khan F, Puzniak L, et al. Seasonal trends in COVID-19 cases, hospitalizations, and mortality in the United States and Europe. Sci Rep. 2023, 13, 3886. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. National emergency department visits for COVID-19, influenza, and respiratory syncytial virus. Published , 2023. Accessed June 14, 2023. https://www.cdc.gov/ncird/surveillance/respiratory-illnesses/index. 17 January.
- Harris, E. FDA clears RSV vaccine for adults aged 60 years or older. JAMA. 2023, 329, 1817. [Google Scholar] [CrossRef] [PubMed]
- Dulfer EA, Geckin B, Taks EJM, et al. Timing and sequence of vaccination against COVID-19 and influenza (TACTIC): a single-blind, placebo-controlled randomized clinical trial. Lancet Reg Health Eur. 2023, 29, 100628. [Google Scholar] [CrossRef] [PubMed]
- ECDC-EMA statement on updating COVID-19 vaccines composition for new SARS-CoV-2 virus variants. European Centre for Disease Prevention and Control. Published , 2023. Accessed June 14, 2023. https://www.ecdc.europa. 7 June.
- Imai M, Ito M, Kiso M, et al. Efficacy of antiviral agents against omicron subvariants BQ.1.1 and XBB. N Engl J Med. 2023, 388, 89–91. [Google Scholar] [CrossRef] [PubMed]
- Wan EYF, Yan VKC, Mok AHY, et al. Effectiveness of molnupiravir and nirmatrelvir-ritonavir in hospitalized patients with COVID-19 : A target trial emulation study. Ann Intern Med. 2023, 176, 505–514. [Google Scholar] [CrossRef]
- Weng C, Xie R, Han G, et al. Safety and efficacy of Paxlovid against omicron variants of Coronavirus disease 2019 in elderly patients. Infect Dis Ther. 2023, 12, 649–662. [Google Scholar] [CrossRef]
- Schwartz KL, Wang J, Tadrous M, et al. Population-based evaluation of the effectiveness of nirmatrelvir-ritonavir for reducing hospital admissions and mortality from COVID-19. CMAJ. 2023, 195, E220–E226. [Google Scholar] [CrossRef]
- Xie, Y.; Choi, T.; Al-Aly, Z. Association of treatment with nirmatrelvir and the risk of post-COVID-19 condition. JAMA Intern Med. 2023, 183, 554–564. [Google Scholar] [CrossRef]
- Imran M, Kumar Arora M, Asdaq SMB, et al. Discovery, development, and patent trends on molnupiravir: A prospective oral treatment for COVID-19. Molecules. 2021, 26, 5795. [Google Scholar] [CrossRef]
- 182. Agostini ML, Pruijssers AJ, Chappell JD, et al. Small-molecule antiviral β-d-N 4-hydroxycytidine inhibits a proofreading-intact Coronavirus with a high genetic barrier to resistance. J Virol. [CrossRef]
- Barnard DL, Hubbard VD, Burton J, et al. Inhibition of severe acute respiratory syndrome-associated coronavirus (SARSCoV) by calpain inhibitors and beta-D-N4-hydroxycytidine. Antivir Chem Chemother. 2004, 15, 15–22. [Google Scholar] [CrossRef]
- Jayk Bernal A, Gomes da Silva MM, Musungaie DB, et al. Molnupiravir for oral treatment of Covid-19 in nonhospitalized patients. N Engl J Med. 2022, 386, 509–520. [Google Scholar] [CrossRef]
- 185. Arribas JR, Bhagani S, Lobo SM, et al. Randomized trial of molnupiravir or placebo in patients hospitalized with covid-19. NEJM Evid. [CrossRef]
- Wong, C.K.H.; Au, I.C.H.; Lau, K.T.K.; Lau, E.H.Y.; Cowling, B.J.; Leung, G.M. Real-world effectiveness of early molnupiravir or nirmatrelvir-ritonavir in hospitalised patients with COVID-19 without supplemental oxygen requirement on admission during Hong Kong’s omicron BA.2 wave: a retrospective cohort study. Lancet Infect Dis. 2022, 22, 1681–1693. [Google Scholar] [CrossRef] [PubMed]
- Flisiak R, Zarębska-Michaluk D, Rogalska M, et al. Real-world experience with molnupiravir during the period of SARS-CoV-2 Omicron variant dominance. Pharmacol Rep. 2022, 74, 1279–1285. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Choi, T.; Al-Aly, Z. Molnupiravir and risk of post-acute sequelae of covid-19: cohort study. BMJ, e: 381, 0745. [Google Scholar]
- EMA Veklury. European Medicines Agency. Published , 2020. Accessed January 22, 2023. https://www.ema.europa. 23 June.
- Dobrowolska K, Zarębska-Michaluk D, Brzdęk M, et al. Retrospective analysis of the effectiveness of remdesivir in COVID-19 treatment during periods dominated by Delta and Omicron SARS-CoV-2 variants in clinical settings. J Clin Med. 2023, 12, 2371. [Google Scholar] [CrossRef] [PubMed]
- 191. Sandin R, Harrison C, Draica F, et al. Estimated impact of oral nirmatrelvir;Ritonavir on reductions in hospitalizations and associated costs within high-risk COVID-19 patients in the US. Research Square, 30 November 2022. [CrossRef]
- Savinkina, A.; Paltiel, A.D.; Ross, J.S.; Gonsalves, G. Population-level strategies for nirmatrelvir/ritonavir prescribing-A cost-effectiveness analysis. Open Forum Infect Dis. 2022, 9, ofac637. [Google Scholar] [CrossRef]
- Wai AKC, Chan CY, Cheung AWL, et al. Association of Molnupiravir and Nirmatrelvir-Ritonavir with preventable mortality, hospital admissions and related avoidable healthcare system cost among high-risk patients with mild to moderate COVID-19. Lancet Reg Health West Pac. 2023, 30, 100602. [Google Scholar] [CrossRef]
- 194. Marangoni D, Antonello RM, Coppi M, et al. Combination regimen of nirmatrelvir/ritonavir and molnupiravir for the treatment of persistent SARS-CoV-2 infection: A case report and a scoping review of the literature. Int J Infect Dis.
- Jeong JH, Chokkakula S, Min SC, et al. Combination therapy with nirmatrelvir and molnupiravir improves the survival of SARS-CoV-2 infected mice. Antiviral Res. 2022, 208, 105430. [Google Scholar] [CrossRef]
- Hill, J.A.; Cowen, L.E. Using combination therapy to thwart drug resistance. Future Microbiol. 2015, 10, 1719–1726. [Google Scholar] [CrossRef]
- 197. Moreno S, Perno CF, Mallon PW, et al. Two-drug vs. three-drug combinations for HIV-1: Do we have enough data to make the switch? HIV Med.
- Sun, F.; Lin, Y.; Wang, X.; Gao, Y.; Ye, S. Paxlovid in patients who are immunocompromised and hospitalised with SARS-CoV-2 infection. Lancet Infect Dis. 2022, 22, 1279. [Google Scholar] [CrossRef]
- Helleberg M, Niemann CU, Moestrup KS, et al. Persistent COVID-19 in an immunocompromised patient temporarily responsive to two courses of remdesivir therapy. J Infect Dis. 2020, 222, 1103–1107. [Google Scholar] [CrossRef]
- 200. Camprubí D, Gaya A, Marcos MA, et al. Persistent replication of SARS-CoV-2 in a severely immunocompromised patient treated with several courses of remdesivir. Int J Infect Dis.
- European Medicines Agency. Use of molnupiravir for the treatment of COVID-19. Published 2022. Accessed , 2023. https://www.ema.europa.eu/en/documents/referral/lagevrio-also-known-molnupiravir-mk-4482-covid-19-article-53-procedure-assessment-report_en. 14 June.
- European Medicines Agency. Paxlovid. Assessment report. Published 2022. Accessed , 2023. https://www.ema.europa.eu/en/documents/assessment-report/paxlovid-epar-public-assessment-report_en. 14 June.
- Stegemann, S.; Gosch, M.; Breitkreutz, J. Swallowing dysfunction and dysphagia is an unrecognized challenge for oral drug therapy. Int J Pharm. 2012, 430, 197–206. [Google Scholar] [CrossRef]
- Hummler H, Stillhart C, Meilicke L, et al. Impact of tablet size and shape on the swallowability in older adults. Pharmaceutics. 2023, 15, 1042. [Google Scholar] [CrossRef] [PubMed]
- Lawal, Y. Africa’s low COVID-19 mortality rate: A paradox? Int J Infect Dis. 2021, 102, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Osei, S.A.; Biney, R.P.; Anning, A.S.; Nortey, L.N.; Ghartey-Kwansah, G. Low incidence of COVID-19 case severity and mortality in Africa; Could malaria co-infection provide the missing link? BMC Infect Dis. 2022, 22, 78. [Google Scholar] [CrossRef]
- Ashworth J, Mathie D, Scott F, et al. Peptide microarray IgM and IgG screening of pre-SARS-CoV-2 human serum samples from Zimbabwe for reactivity with peptides from all seven human coronaviruses: a cross-sectional study. Lancet Microbe. 2023, 4, e215–e227. [Google Scholar] [CrossRef]
- Diop, B.Z.; Ngom, M.; Pougué Biyong, C.; Pougué Biyong, J.N. The relatively young and rural population may limit the spread and severity of COVID-19 in Africa: a modelling study. BMJ Glob Health. 2020, 5, e002699. [Google Scholar] [CrossRef] [PubMed]
- Gill CJ, Mwananyanda L, MacLeod WB, et al. What is the prevalence of COVID-19 detection by PCR among deceased individuals in Lusaka, Zambia? A postmortem surveillance study. BMJ Open. 2022, 12, e066763. [Google Scholar] [CrossRef] [PubMed]
- Levin AT, Owusu-Boaitey N, Pugh S, et al. Assessing the burden of COVID-19 in developing countries: systematic review, meta-analysis and public policy implications. BMJ Glob Health. 2022, 7, e008477. [Google Scholar] [CrossRef]
- Kunyenje CA, Chirwa GC, Mboma SM, et al. COVID-19 vaccine inequity in African low-income countries. Front Public Health. 2023, 11, 1087662. [Google Scholar] [CrossRef]
- Member State Briefing. Update on Global COVID-19 vaccination. Accessed , 2023. https://apps.who.int/gb/COVID-19/pdf_files/2023/05_01/Item1. 18 June.
- US international COVID-19 vaccine donations tracker, K.F.F. Published , 2023. Accessed June 11, 2023. https://www.kff. 9 June.
- Hassan, F.; Yamey, G.; Abbasi, K. Profiteering from vaccine inequity: a crime against humanity? BMJ, n: 374, 2027. [Google Scholar]
- Rzymski, P.; Szuster-Ciesielska, A. The COVID-19 vaccination still matters: Omicron variant is a final wake-up call for the rich to help the poor. Vaccines (Basel). 2022, 10, 1070. [Google Scholar] [CrossRef]
- WHO. COVAX Allocation. Published 2022. Accessed , 2022. https://www.who. 15 May.
- Savinkina A, Bilinski A, Fitzpatrick M, et al. Estimating deaths averted and cost per life saved by scaling up mRNA COVID-19 vaccination in low-income and lower-middle-income countries in the COVID-19 Omicron variant era: a modelling study. BMJ Open. 2022, 12, e061752. [Google Scholar] [CrossRef]
- Hoffman SA, Costales C, Sahoo MK, et al. SARS-CoV-2 neutralization resistance mutations in patient with HIV/AIDS, California, USA. Emerg Infect Dis. 2021, 27, 2720–2723. [Google Scholar] [CrossRef] [PubMed]
- Cele S, Karim F, Lustig G, et al. SARS-CoV-2 prolonged infection during advanced HIV disease evolves extensive immune escape. Cell Host Microbe. 2022, 30, 154–162. [Google Scholar] [CrossRef]
- Gonzalez-Reiche, AS. SARS-CoV-2 in low-income countries: the need for sustained genomic surveillance. Lancet Glob Health. 2023, 11, e815–e816. [Google Scholar] [CrossRef] [PubMed]
- 221. Le T, Sun C, Chang J, Zhang G, Yin X. MRNA vaccine development for emerging animal and zoonotic diseases. Viruses. [CrossRef]
- Chauhan, R.P.; Dessie, Z.G.; Noreddin, A.; El Zowalaty, M.E. Systematic review of important viral diseases in Africa in light of the “One Health” concept. Pathogens. 2020, 9, 301. [Google Scholar] [CrossRef]
- Gwenzi, W.; Skirmuntt, E.C.; Musvuugwa, T.; Teta, C.; Halabowski, D.; Rzymski, P. Grappling with (re)-emerging infectious zoonoses: Risk assessment, mitigation framework, and future directions. Int J Disaster Risk Reduct. 2022, 82, 103350. [Google Scholar] [CrossRef]
- Iacobucci, G. Covid-19: “Grotesque inequity” that only a quarter of paxlovid courses go to poorer countries. BMJ, o: 379, 2795. [Google Scholar]
- 225. Price of COVID treatments from Pfizer, Merck, GSK align with patient benefits -report. Reuters, 3 February 2022; 26.
- Baker RE, Mahmud AS, Miller IF, et al. Infectious disease in an era of global change. Nat Rev Microbiol. 2022, 20, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Watts, M.S. Physicians as role models in society. West J Med. 1990, 152, 292. [Google Scholar]
- Betancourt, J.A.; Rosenberg, M.A.; Zevallos, A.; Brown, J.R.; Mileski, M. The Impact of COVID-19 on Telemedicine Utilization Across Multiple Service Lines in the United States. Healthcare (Basel). [CrossRef]
- Bazan, D.; Nowicki, M.; Rzymski, P. Medical students as the volunteer workforce during the COVID-19 pandemic: Polish experience. Int J Disaster Risk Reduct. 2021, 55, 102109. [Google Scholar] [CrossRef]
- Benfante, A.; Di Tella, M.; Romeo, A.; Castelli, L. Traumatic stress in healthcare workers during COVID-19 pandemic: A review of the immediate impact. Front Psychol, 5: 11, 5699. [Google Scholar]
- Wittenberg, E.; Goldsmith, J.V.; Chen, C.; Prince-Paul, M.; Johnson, R.R. Opportunities to improve COVID-19 provider communication resources: A systematic review. Patient Educ Couns. 2021, 104, 438–451. [Google Scholar] [CrossRef]
- Mach KJ, Salas Reyes R, Pentz B, et al. News media coverage of COVID-19 public health and policy information. Humanit Soc Sci Commun. 2021, 8, 1–11. [Google Scholar]
- Fotiadis K, Dadouli K, Avakian I, et al. Factors associated with healthcare workers’ (HCWs) acceptance of COVID-19 vaccinations and indications of a role model towards population vaccinations from a cross-sectional survey in Greece, may 2021. Int J Environ Res Public Health. 2021, 18, 10558. [Google Scholar] [CrossRef] [PubMed]
- 234. World Health Organization. Regional Office for Europe. Evaluation Report for the Training Module “Communicating with Patients about COVID-19 Vaccination”: Greece, /: Regional Office for Europe; 2023. Accessed , 2023. https, 18 June 2023.
- Burson, R.C.; Buttenheim, A.M.; Armstrong, A.; Feemster, K.A. Community pharmacies as sites of adult vaccination: A systematic review. Hum Vaccin Immunother. 2016, 12, 3146–3159. [Google Scholar] [CrossRef] [PubMed]
- Sobierajski, T.; Rzymski, P.; Wanke-Rytt, M. The influence of recommendation of medical and non-medical authorities on the decision to vaccinate against influenza from a social vaccinology perspective: Cross-sectional, representative study of polish society. Vaccines (Basel). 2023, 11, 994. [Google Scholar] [CrossRef] [PubMed]
- Paudyal V, Fialová D, Henman MC, et al. Pharmacists’ involvement in COVID-19 vaccination across Europe: a situational analysis of current practice and policy. Int J Clin Pharm. 2021, 43, 1139–1148. [Google Scholar] [CrossRef]
- Queeno, B.V. Evaluation of inpatient influenza and pneumococcal vaccination acceptance rates with pharmacist education. J Pharm Pract. 2017, 30, 202–208. [Google Scholar] [CrossRef]
- Pullagura, G.R.; Waite, N.M.; Houle, S.K.D.; Violette, R.; Wong, W.W.L. Cost-utility analysis of offering a novel remunerated community pharmacist consultation service on influenza vaccination for seniors in Ontario, Canada. J Am Pharm Assoc (2003). 2019, 59, 489–497. [Google Scholar] [CrossRef]
Figure 1.
The emergence of SARS-CoV-2 variants over the course of the COVID-19 pandemic (A) and their phylogeny (B). The data and graphs retrieved from Nextstrain.org 58.
Figure 1.
The emergence of SARS-CoV-2 variants over the course of the COVID-19 pandemic (A) and their phylogeny (B). The data and graphs retrieved from Nextstrain.org 58.
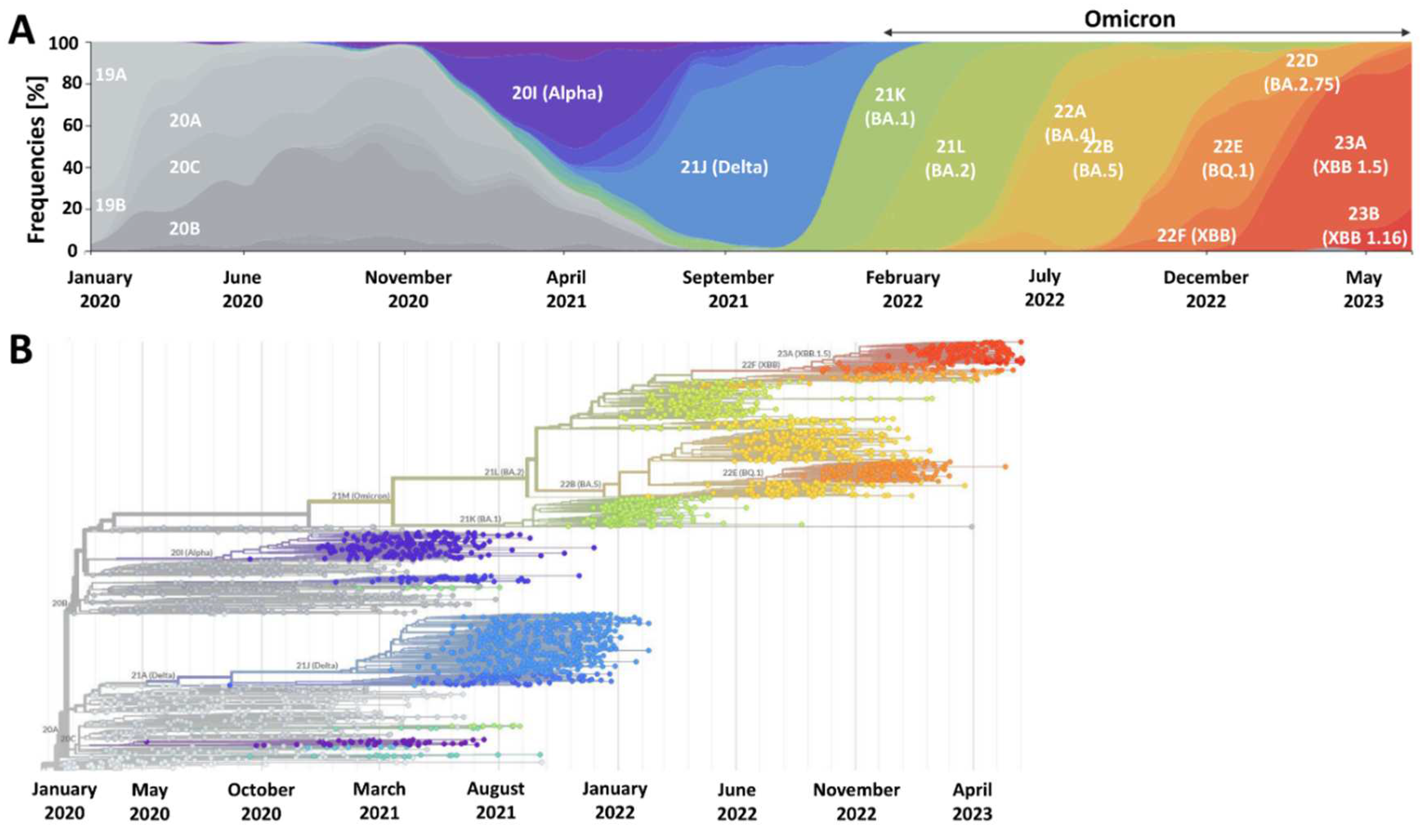
Figure 2.
A global death toll of SARS-CoV-2 Omicron during the first 1.5 years of its dominance compared to the pre-Omicron period. Graphs prepared based on data collected by Our World in Data 5.
Figure 2.
A global death toll of SARS-CoV-2 Omicron during the first 1.5 years of its dominance compared to the pre-Omicron period. Graphs prepared based on data collected by Our World in Data 5.
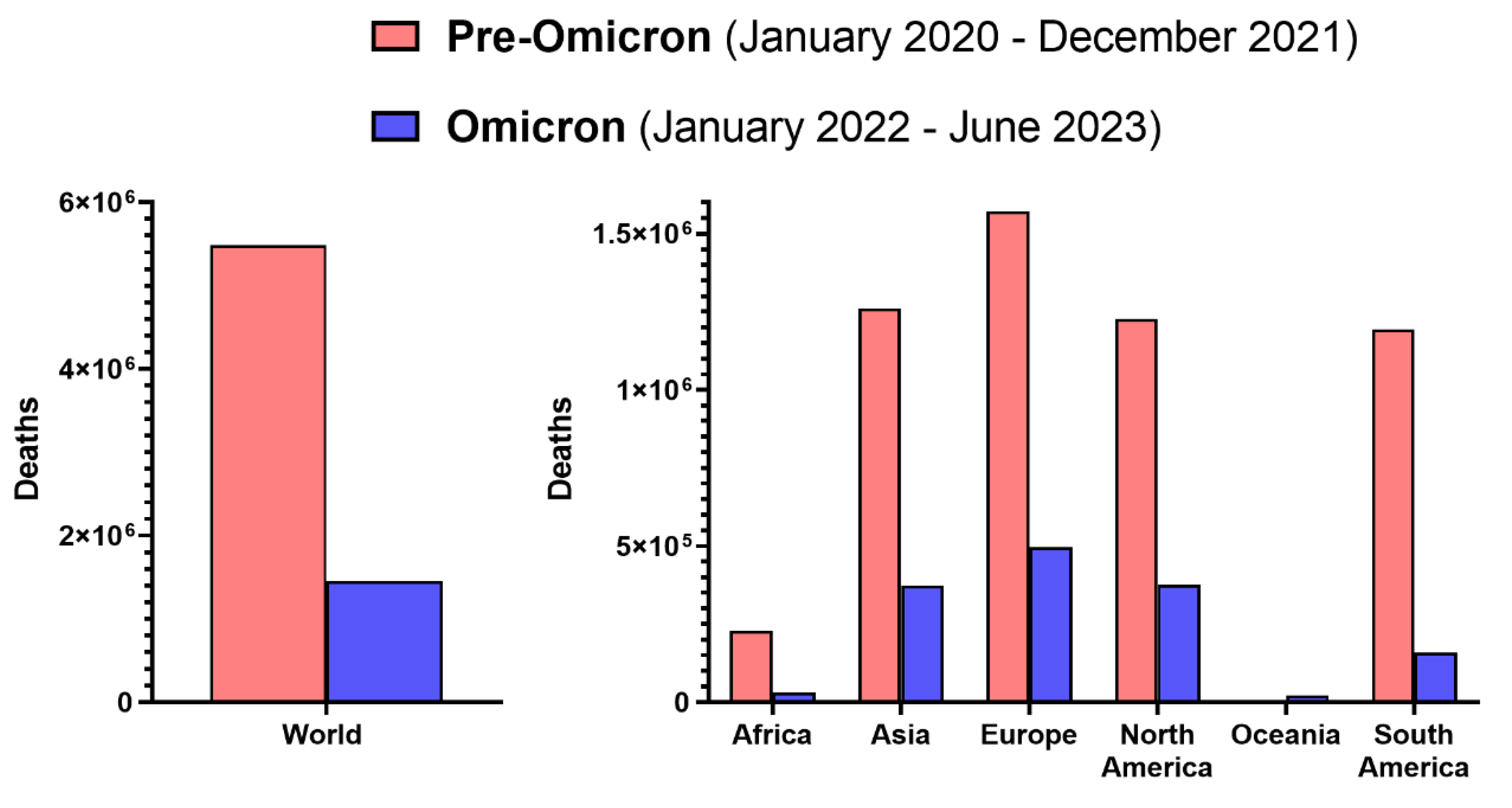
Figure 3.
Steps of the SARS-CoV-2 replication cycle in the human cell disrupted by oral antivirals nirmatrelvir/ritonavir and molnupiravir and intravenously administrated remdesivir. The scheme was used and modified by permission 191.
Figure 3.
Steps of the SARS-CoV-2 replication cycle in the human cell disrupted by oral antivirals nirmatrelvir/ritonavir and molnupiravir and intravenously administrated remdesivir. The scheme was used and modified by permission 191.
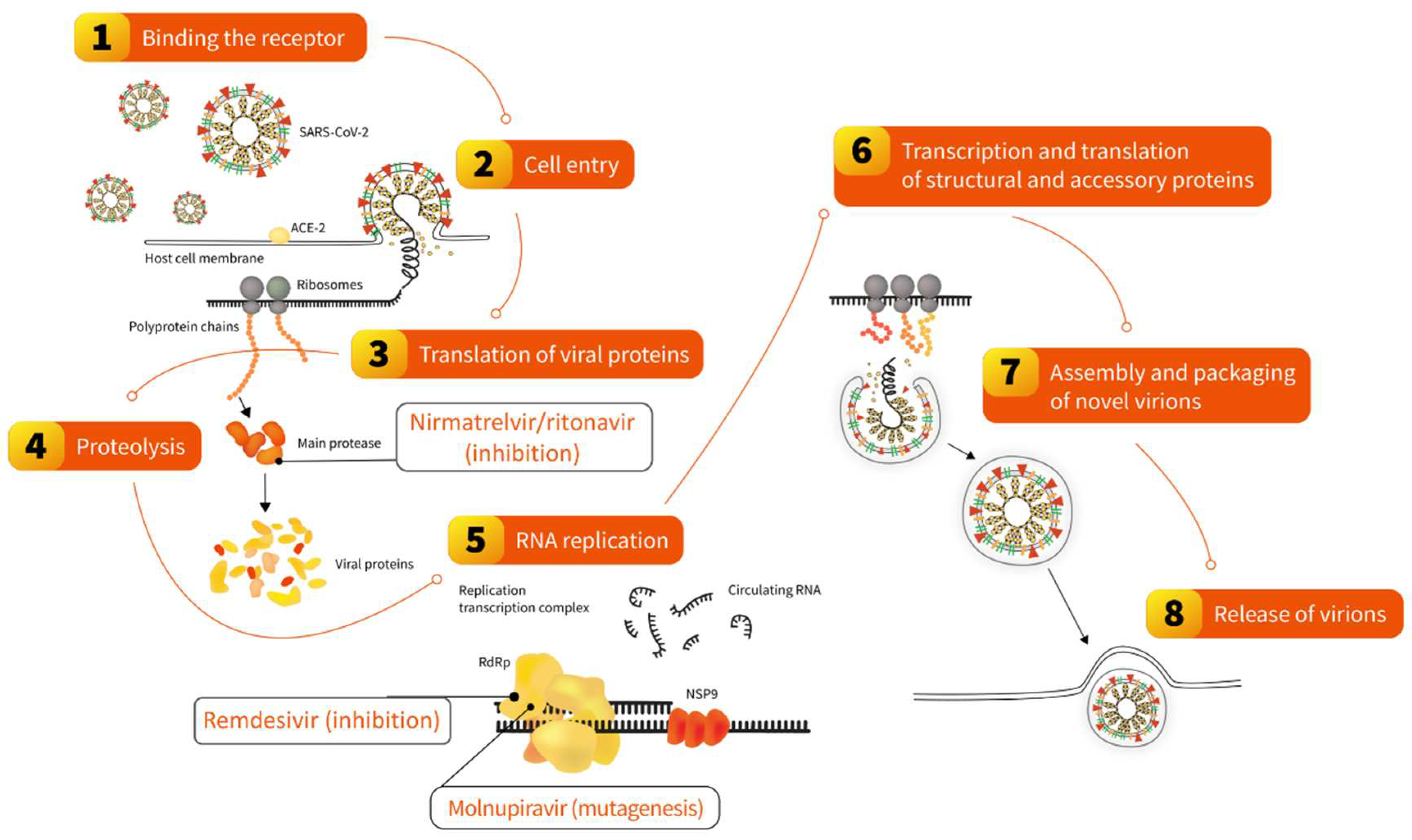
Figure 4.
The percentage of population with completed initial COVID-19 vaccination protocol by economic group. Prepared based on data from Our World in Data 5.
Figure 4.
The percentage of population with completed initial COVID-19 vaccination protocol by economic group. Prepared based on data from Our World in Data 5.
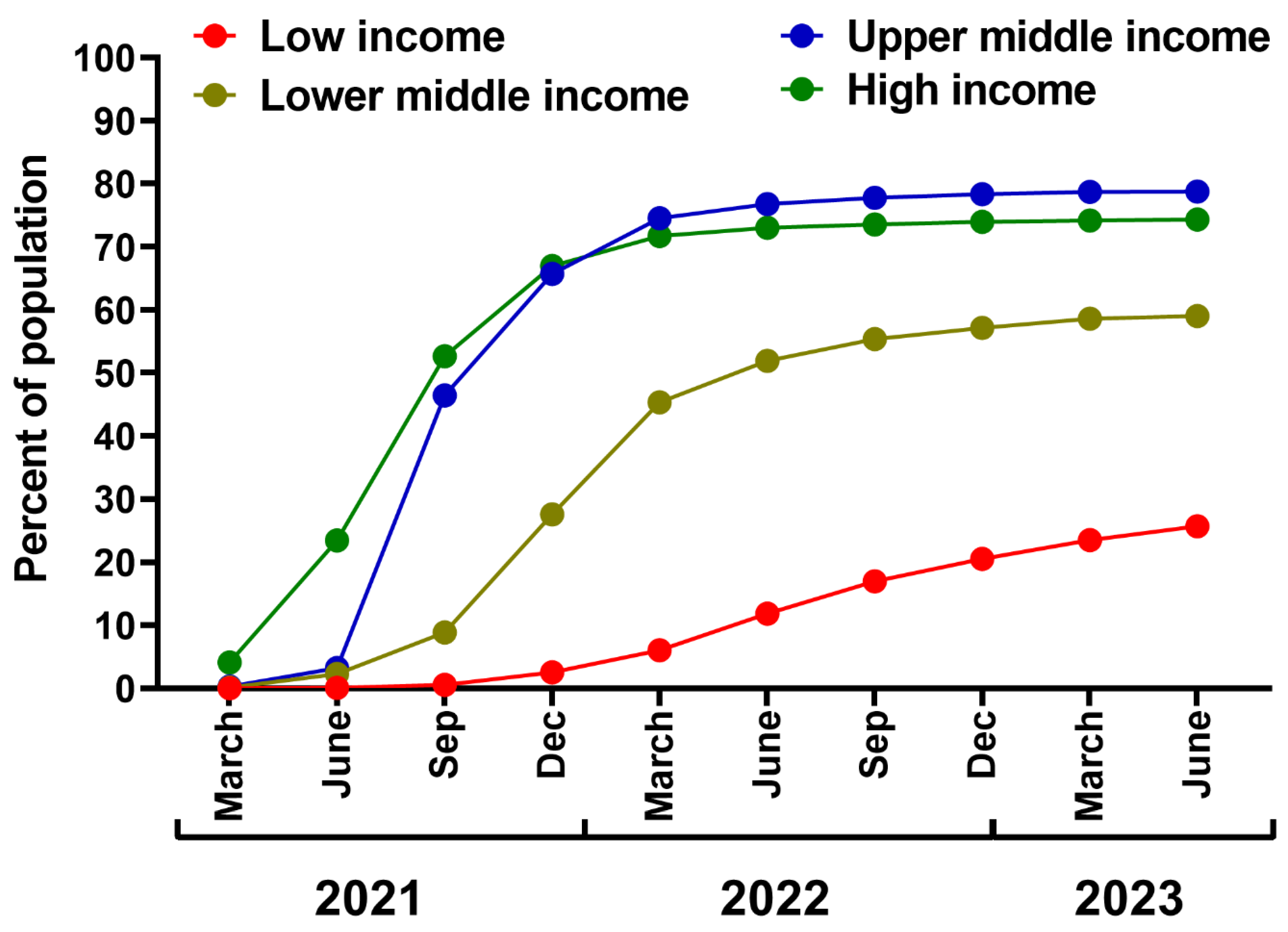
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



