Submitted:
16 August 2023
Posted:
17 August 2023
You are already at the latest version
Abstract
Introduction: congenital ureteral stenosis is one of the leading causes of impaired urinary drainage and subsequent dilatation of urinary collecting system, known as hydronephrosis or ureterohy-dronephrosis. Mechanism that leads to obstruction is not clearly known. Multiple studies in rat models have shown increased angiotensin 11 and TGFβ levels in obstructed ureteral tissue. The aim of the study is to investigate expression of fibrosis related genes in obstructive and normal ureteral tissue.
Material and methods: it is a monocentric study in which nineteen patients were selected prospec-tively. 17 patients of which underwent Hynes-Anderson pyeloplasty due to the PUJO, 2 patients underwent ureteroneocystostomy due to ureterovesical junction obstruction (UVJO) and 6 patients were chosen for control group: 5 underwent nephrectomies due to the kidney tumor and 1- upper pole heminephrectomy due to the duplex kidney with normal pyeloureteric junctions in all. Tissue RNA was chemically extracted after freezing the biopsy samples in liquid nitrogen, with cDNA synthesis performed immediately after nucleic acid isolation. qPCR to evaluate the relative expres-sion of Tgfb1, Mmp1, Timp1, Pai1, Ctgf and Vegfa was performed. Expression levels of Gapdh and Gpi genes (geometric average) were used to calculate the relative expression of the investigated genes. Outliers were removed prior to calculating confidence intervals for the experimental groups, and Wilcoxon rank-sum test was performed to determine statistical significance of the differences.
Results: significant differences between healthy and stenotic tissue samples in Ctgf gene expression levels were observed, with the samples from afflicted tissue showing lower expression. No statistical difference in expression levels of Tgfb1, Timp1, Vegfa, Mmp1, Pai1 was found.
Discussion and conclusions: These findings suggest that tissue fibrosis similar to other tissues and organs is not the leading cause of stenosis, at least at the moment of surgery. Decreased CTGF ex-pression is indicative of developmental origin of obstruction.
Keywords:
Pyeloureteric junction
; Pyeloureteric junction obstruction
; Gene expression
; Fibrosis related genes
1. Introduction
Congenital ureteral stenosis is the leading cause of impaired urinary drainage and subsequent dilatation of urinary collecting system, condition known as hydronephrosis or ureterohydronephrosis, when ureteric dilatation is present. Ureteral obstruction is a very serious clinical condition, leading to damage of renal parenchyma because of obstruction induced increased pressure, reduced glomerular filtration, activation of profibrotic factors, cell apoptosis and subsequent loss of kidney function which has been investigated in animal models in a series of studies. [1], [2] Early recognition of obstruction, followed by timely treatment usually results in recovery of a kidney, mainly because of the better reproduction potential of renal stem cells and recovery of parenchyma at an early age. [3]
Several theories of pathogenesis leading to the development of hydronephrosis were learnt from animal models. The reasons of hydronephrosis can be generally regarded as obstructive and non-obstructive.
In systematic analysis which reviewed data from 10 articles was revealed that 15 genes had changed expression in ureteral stricture tissue. Genes which were up regulated were (ET1, 87 ACTA2, MCP-1, TGFB1, NFKB1, IL-6, HIF1A, S100A1, SYP) and the expression of six genes was down regulated (ADM, NOS2, EGF, PDGFRA, UCHL1, NGFR). The products of these genes are components of HIF-1 signaling pathway and participate in the development of vasculature. Some of these factors act as inductor of signaling pathways and particularly affect Ras signaling system. There may also be a connection of current factors with others that are related with tissue hypoxia, fibrosis and the inflammation. [4]
Histological studies of obstructed human ureters identified excessive amount of collagen in extracellular matrix and impaired distribution of its isoforms, increased proportion of elastin and apoptosis of myocytes. Altogether it leads to the abnormal development of tissue. [5], [6] In the intracellular level, immunofluorescence and PCR analysis revealed that cytoskeleton structural proteins were impaired in PUJO smooth muscle cells probably explaining the increased apoptosis of smooth muscle cells in PUJO specimens. [7] Aforementioned pathological findings suggest that the pathogenesis of the development of ureteral obstruction involves the processes of fibrosis, apoptosis and cell differentiation arrest.
The aim of our study was to analyze several fibrosis related factors that are known to participate in general mechanism of fibrosis in multiple sites of human body and to discuss their involvement in previously described chains of pathogenesis.
We have chosen to investigate fibrosis-associated genes expression in congenitally obstructed ureteral tissue. As obstruction involves abnormal cell differentiation as well as changes in extracellular matrix, expression profiles of Tgfb1, Ctgf, Vegfa, Timp1, Pai1 and Mmp1 genes were chosen to be analyzed according to previous studies where they were analyzed as the fibrosis inducing factors. [8,9,10,11,12,13] We could not identify any studies describing the expression analysis of CTGF, MMP1, PAI-1 in obstructed ureteral tissue over the last 10 years. Nevertheless, these genes were chosen as closely related to Tgfb pathway and implicated in the development of tissue fibrosis in other organs, such as the heart. [14]
2. Materials and Methods
Collection of samples
It is a monocentric study in which patients were included prospectively from 2019 to 2021. Samples of pyeloureteric junctions and 2 ureterovesical junctions were collected during surgery and stored in -80°C until RNA extraction.
The decision of whether or not to operate on the patient was based on our local protocol: the operation was performed when there was an increase in the anterior-posterior diameter of renal pelvis by 20% on subsequent ultrasound scans, when the differential renal function (DRF) was <40% of the affected kidney or a decrease in DRF by >5% was observed on subsequent renal scans, or when symptoms could be attributed to hydronephrosis.
RNA extraction and qPCR
Tissue RNA was extracted after freezing the samples in liquid nitrogen, mechanically dissociating them and applying Trizol reagent (Thermo Fisher) according to the manufacturer’s protocol. 1 mL of Trizol reagent was added per 100 mg of tissue. Isolated RNA was dissolved in nuclease-free water, and its quantity and quality were evaluated using the NanoDrop device (Thermo Fisher).
cDNA synthesis was performed using Maxima H Minus First Strand cDNA Synthesis Kit (Thermo Fisher). Each sample had reverse transcription negative (RT-) control. qPCR was performed using Luminaris Color HiGreen qPCR Master Mix (Thermo Fisher) with the Eppendorf realplex4 quantitative PCR device. Expression levels of Gapdh and Gpi genes (geometric average) were used to calculate relative expression of the investigated genes. The Common base method, as described in [15] was used to calculate relative expression of the genes, factoring the qPCR reaction efficiency. Primer sequences used for qPCR were derived from Primer Bank (https://pga.mgh.harvard.edu/primerbank/)
Statistical analysis
Results were analyzed using RStudio Version 1.2.5033. Outliers were removed prior to analysis. Wilcoxon rank-sum test was carried out to determine the significance of differences between healthy and obstructed tissue (significance level α = 0.05), and assessment of confidence intervals was carried out to further understand the differences between the groups.
3. Results
17 patients underwent Hynes-Anderson pyeloplasty due to the PUJO, 2 patients underwent ureteroneocystostomy due to ureterovesical junction obstruction (UVJO). Six patients were chosen for control group: 5 underwent nephrectomies due to the kidney tumor and 1- upper pole heminephrectomy due to the duplex kidney with normal pyeloureteric junctions in all. Median age was 15.2 months [9.07; 66.2] at the time of the surgery and median age of control group (4 boys and 2 girls) was 60 months [35; 80.3].
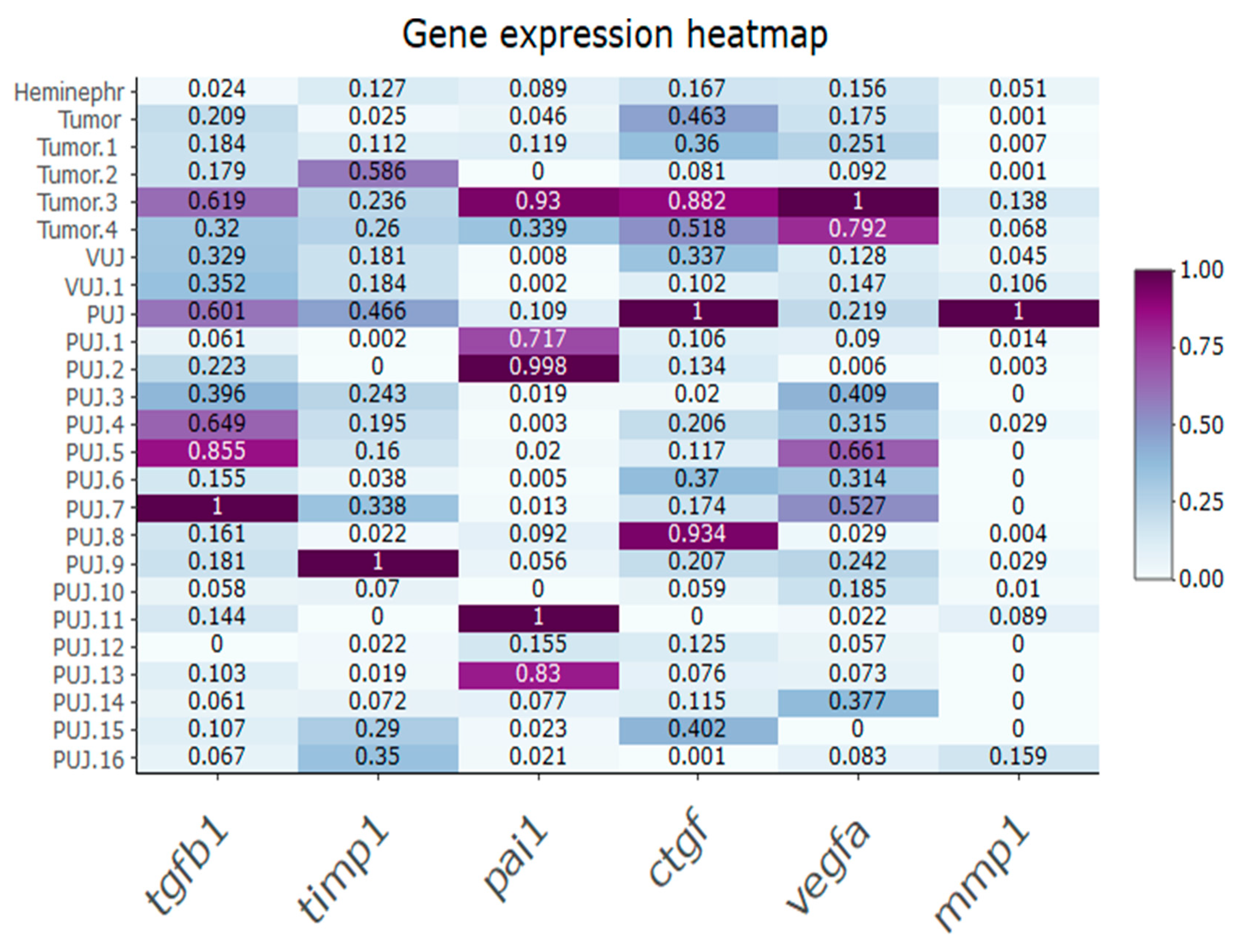
The expression of fibrosis-related genes in every specimen are depicted in the heatmap. (Figure 1). Variable expression of all studied genes was noted both in obstructed and in normal junctions.
Figure 2.
Relative expression of fibrosis-related genes in control and strictured ureter tissue samples. Medians (dot/triangle) and 95% confidence intervals (whiskers) shown. Star (*) indicates statistical significance between the groups (p < 0.05) as determined by Wilcoxon rank-sum test. Sample sizes are 6 for control and 19 for strictured group.
Figure 2.
Relative expression of fibrosis-related genes in control and strictured ureter tissue samples. Medians (dot/triangle) and 95% confidence intervals (whiskers) shown. Star (*) indicates statistical significance between the groups (p < 0.05) as determined by Wilcoxon rank-sum test. Sample sizes are 6 for control and 19 for strictured group.
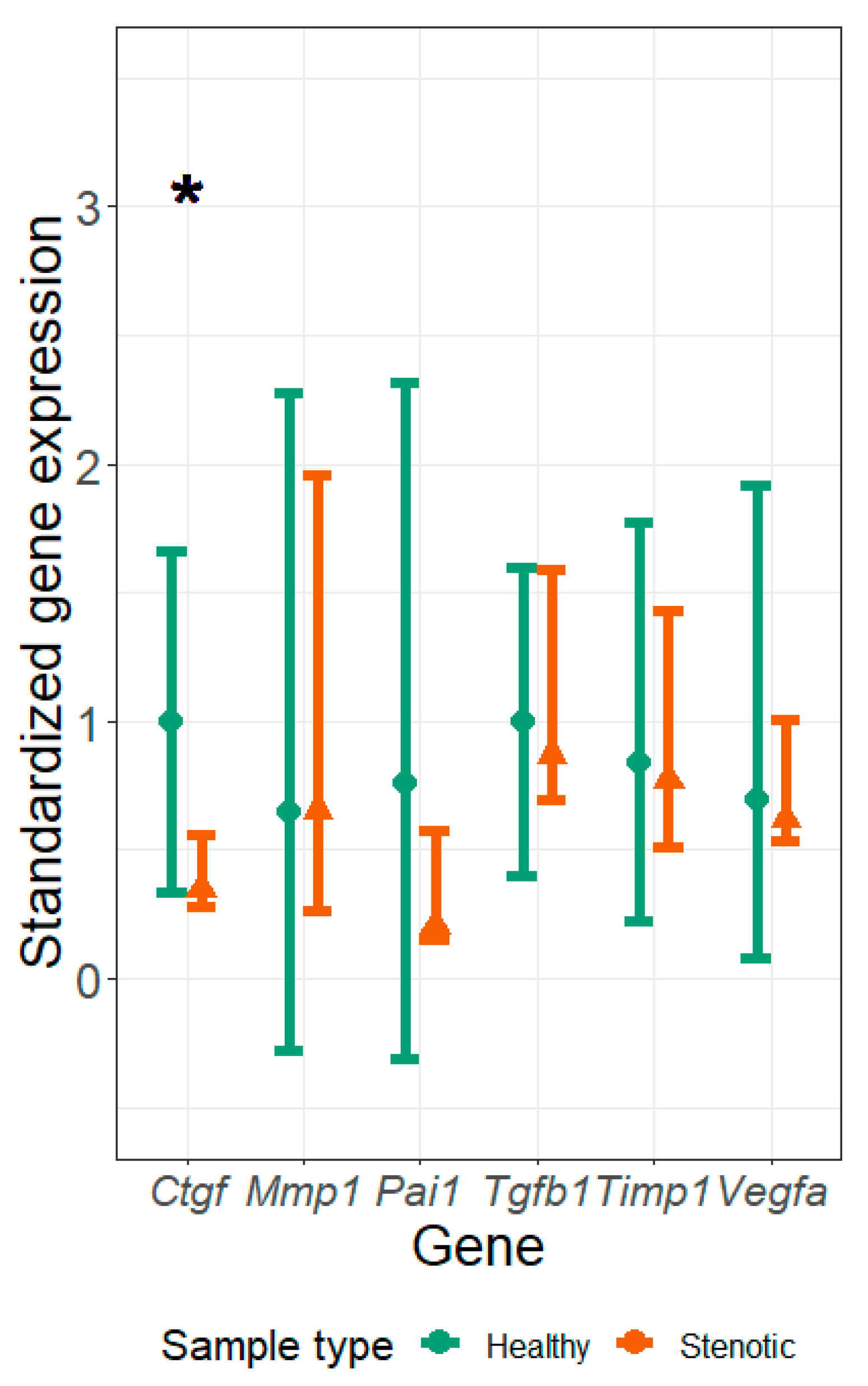
Wilcoxon rank-sum test showed significant differences between groups in Ctgf gene expression level. We further decided to compare the confidence intervals between groups, and the intervals of Ctgf gene showed little overlap, indicating differences between obstructed and control tissue samples. (Figure 2) No statistically significant differences between the groups were found in the expression of other studied genes.
Figure 1.
Heatmap of relative expression of fibrosis-related genes in control and strictured ureter tissue samples. Results standardized to highest gene expression levels for each gene. PUJ – pyeloureteric junction obstruction; Tumor – pyeloureteric junction from kidney tumor specimen; Heminephr. – pyeloureteric junction from heminephrectomy specimen; VUJ – vesicoureteral junction obstruction.
Figure 1.
Heatmap of relative expression of fibrosis-related genes in control and strictured ureter tissue samples. Results standardized to highest gene expression levels for each gene. PUJ – pyeloureteric junction obstruction; Tumor – pyeloureteric junction from kidney tumor specimen; Heminephr. – pyeloureteric junction from heminephrectomy specimen; VUJ – vesicoureteral junction obstruction.
4. Discussion
Out of our selected markers the best described group of fibrosis-related factors in the literature belong to transforming growth factor (TGF) family, particularly TGFs 1 to 3, sharing similar functions with minor structural differences. [16] Out of the three, the most well studied and ubiquitous is TGFβ1, a polypeptide that regulates cell differentiation, proliferation, adhesion and immune behavior. It is found in sites of injury, inflammation, cancer and elsewhere. The protein positively and negatively regulates other growth factors. The mature peptide may form heterodimers with other TGF beta family members. [17] It is stored in the extracellular matrix (ECM) as a complex with its pro-domain. Activation of TGFβ-1 requires the binding of alpha-V integrin, present on the membrane of residing cells, to an RGD sequence in the TGF prodomain and exertion of force on this domain, which is held in the extracellular matrix by latent TGFβ binding proteins (LTBP). As the synthesis and release of TGFβ involves many steps of proteolytic processing, binding and release, the expression and activity of TGFβ is both time and situation-dependent.
Among the disease-relevant ROS-dependent genomic targets (reactive oxygen species), TGF-beta1 stimulates expression of the potent profibrotic matricellular PAI-1 and CTGF genes as well as those that code ECM structural elements (fibronectin, collagen I). PAI-1 is one of the most highly regulated part of the TGFβ1/SMAD3-induced group, a prominent ROS responsive gene and involved in tissue fibrosis process. As the main inhibitor of plasmin generation, PAI-1 limits ECM degradation alleviating accumulation of matrix structural elements at the injury site. PAI-1 deficiency is renal-protective whereas transgenic PAI-1 overexpression promotes an increased fibrotic response with associated recruitment of inflammatory cells, macrophages and myofibroblasts. As proof-of-concept, unilateral ureter-obstructed PAI-1−/− mice develop a significantly attenuated inflammatory response suggesting that PAI-1 directly promotes infiltration of macrophages and T-cells. [17]
However, it is worth mentioning that out of these studies we could identify only one when polymerase chain reaction (PCR) analysis was applied in human ureters and showed that the amount of TGFβ1 mRNA in stenotic tissue was higher than in controls. [18] Another study found that immunohistochemical staining of TGFβ3 in obstructed tissue was higher than in the control tissue. [19] Even though the results of our study of TGFβ in patients with PUJO could not replicate these findings, this could be due to the observations that TGFβ1 is often expressed in the early stages of diseases, especially when macrophage activity is high, as well as when inflammation is present. Experimental studies have also advocated the increase of mRNA TGF-β in postoperative UPJO. Under these circumstances such finding can be explained by an acute onset of obstruction activating muscle fiber and collagen elaboration. [20]
We found only one study analyzing the role of PAI-1 in an obstructed kidney model and showing that TIMP-1 and PAI-1 in rat model are involved in the progression of tubulointerstitial scarring of renal tissue. [21]
With regard to one of the theories of UB branching, AngII is named as one of the enhancers for this process. PAI-1 has also been described as a downstream target of AngII in endothelial cells.[22] Is there a relation between potentially decreased PAI-1 and AngII, as its potential effector in congenitally obstructed ureter, remains to be answered. PAI-1 has been shown to play a role in ECM maintenance and remodeling, counteracting the effects of matrix metalloproteinases. [23]
We were able to identify only one study where VEGF was investigated in UPJO and a control group in human specimens: no color staining of VEGF was found in both obstructed and control groups and subsequently showed no difference in staining potential regardless of obstruction pattern. [24] Our results in VEGF expression have corresponded to the results of an aforementioned study, finding no difference in this gene expression in stenotic versus healthy tissue.
We could not identify any previous publication regarding Ctgf expression profile in congenital ureteral obstruction.
Our results have demonstrated that relative expression of Ctgf in stenotic ureteral tissue was significantly lower than in the control tissue supporting a potential role of this gene in the processes of stricture development. Ctgf has important roles in many biological processes, including cell adhesion, migration, proliferation, angiogenesis, skeletal development, and tissue wound repair, and is critically involved in fibrotic disease and several forms of cancers [9], [25]. It is thought that CTGF can cooperate with TGFβ to induce fibrosis and extracellular matrix production in association with other fibrosis-related conditions. [26] Overexpression of Ctgf in fibroblasts promotes fibrosis at the beginning of the development of stricture in the skin, kidney, and lung. We may only speculate that the reduction in our patients may be indicative on the stage of fibrosis – for example, in a nephropathy model in mice, it was shown that the expression level of Ctgf gradually increases over a 2-week period and is static afterwards; our results indicate there may be a significant reduction in the expression of this gene following the primary stage of disease development or gene can be only expressed for a certain period of time. [27]
Evident similarity in expression of other studied genes may suggest that fibrosis process is not the most important in the development of congenital PUJO and we should rather concentrate on other aspects of development.
Considering other aspects of cell distribution, the most prominent alteration comprises interstitial cells of Cajal and telocytes which are significantly reduced in comparison to normal ureteral tissue in both UPJO and UVJO. Differentiation of Cajal cells may be stimulated by overexpression of CTGF gene. [28] Also, SOX9 was found regulating CTGF/CCN2 transcription in chondrocytes. [29] Hypomotility of the ureter was induced by Sox9, a homeobox gene, the loss of which led to smooth muscle cell differentiation arrest. [30] Another study showed an essential role of Sox9 in regulation of differentiation of ureteric mesenchyme. Primary dilatation of renal collecting system without ureteral obstruction was induced by Keap1 knockout and expression of aquaporins with spontaneous mutations which caused urinary hyperproduction and secondary dilatation of renal collecting system. [31]
The drawback of our study is the relatively low and heterogeneous by age sample size. Although statistical analysis was possible, more specimens would be needed to elucidate the potential differences in the expression of our studied genes. Another drawback is a small and non-homogenous control group when 5 pyeloureteric junctions were obtained from renal tumor specimens and 1 pyeloureteric junctions from duplex kidney system. None of them had macroscopic signs of stricture, however the results can be influenced by preoperative chemotherapy and maldevelopment of the ureteral wall when compared to normal ureters. [32] Observed tendencies comparing PUJ, UVJ, dilated UVJ and normal PUJ after chemotherapy may be further investigated.
In summary, our findings suggest that decreased Ctgf expression in strictured ureteral specimen may be a sign of a progressive failure determined by a genetic trigger occurring at the embryonic period. Further studies have to be conducted to verify hypotheses.
5. Conclusions
In summary, our findings suggest that decreased Ctgf expression in strictured ureteral specimen may be a sign of a progressive failure determined by a genetic trigger occurring at the embryonic period. Further studies have to be conducted to verify hypotheses.
Author Contributions
All authors have accepted responsibility for the entire content of this manuscript and approved its submission. All authors prepared the manuscript.
Funding
This project has received funding from European Regional Development Fund No 01.2.2-LMT-K-718-03-0087 under grant agreement with the Research Council of Lithuania (LMTLT).
Institutional Review Board Statement
This study was approved by the Vilnius Regional Bioethics Committee (158200-18/6-1044-544, the date of approval 05/06/2018).
Conflicts of Interest
None.
Abbreviations
| UB | ureteric bud; |
| PUJO | obstructed pyeloureteric junction; |
| EGFR | epidermal growth factor receptor; |
| AT1R | angiotensin type 1 receptor; |
| AT2R | angiotensin type 2 receptor; |
| UVJO | ureterovesical junction obstruction; |
| ChKD | chronic kidney disease; |
| ESRD | end stage renal disease; |
| LTBP | latent TGFβ binding proteins |
References
- Zhao, S.; Li, W.; Yu, W.; Rao, T.; Li, H.; Ruan, Y.; Yuan, R.; Li, C.; Ning, J.; Li, S.; et al. Exosomal miR-21 from tubular cells contributes to renal fibrosis by activating fibroblasts via targeting PTEN in obstructed kidneys. Theranostics 2021, 11, 8660–8673. [Google Scholar] [CrossRef] [PubMed]
- Chang, I.Y.; Kim, J.N.; Jun, J.Y.; You, H.J.; Jeon, Y.J.; Park, K.-S.; Yoon, S.P. Repression of apurinic/apyrimidinic endonuclease by p53-dependent apoptosis in hydronephrosis-induced rat kidney. Free. Radic. Res. 2011, 45, 728–734. [Google Scholar] [CrossRef]
- Deng, Q.-F.; Chu, H.; Peng, B.; Liu, X.; Cao, Y.-S. Outcome analysis of early surgery and conservative treatment in neonates and infants with severe hydronephrosis. J. Int. Med Res. 2021, 49. [Google Scholar] [CrossRef]
- Isali, I.; McClellan, P.; Wong. , T.R.; Gupta, S.; Woo, L. A systematic review of underlying genetic factors associated with ureteropelvic junction obstruction in stenotic human tissue. J. Pediatr. Urol. 2022, 18, 629–641. [Google Scholar] [CrossRef]
- Senkul, T.; Kucukodaci, Z.; Iseri, C.; Karademir, K.; Erden, D.; Baloglu, H.; Narin, Y. The Smooth Muscle Ratio at the Renal Pelvis in Adults: Does It Predict Surgical Outcome? Urol. Int. 2004, 73, 248–251. [Google Scholar] [CrossRef]
- Kajbafzadeh, A.-M.; Payabvash, S.; Salmasi, A.H.; Monajemzadeh, M.; Tavangar, S.M. Smooth Muscle Cell Apoptosis and Defective Neural Development in Congenital Ureteropelvic Junction Obstruction. J. Urol. 2006, 176, 718–723. [Google Scholar] [CrossRef]
- Wishahi M. et al., “SMOOTH MUSCLE CELLS AND COLLAGEN FIBRES ARCHITECTURE IN EQUIVO-CAL URETEROPELVIC JUNCTION OBSTRUCTION: ELECTRON MICROSCOPY STUDY WITH CLINI-CAL CORRELATION IN ADULT EGYPTIANS,” 2020.
- Hinz, B. The extracellular matrix and transforming growth factor-β1: Tale of a strained relationship. Matrix Biol. 2015, 47, 54–65. [Google Scholar] [CrossRef]
- E Lipson, K.; Wong, C.; Teng, Y.; Spong, S. CTGF is a central mediator of tissue remodeling and fibrosis and its inhibition can reverse the process of fibrosis. Fibrogenesis Tissue Repair 2012, 5, 2–9. [Google Scholar] [CrossRef]
- Shang, J.; He, Q.; Chen, Y.; Yu, D.; Sun, L.; Cheng, G.; Liu, D.; Xiao, J.; Zhao, Z. miR-15a-5p suppresses inflammation and fibrosis of peritoneal mesothelial cells induced by peritoneal dialysis via targeting VEGFA. J. Cell. Physiol. 2019, 234, 9746–9755. [Google Scholar] [CrossRef]
- Nie, Q.-H.; Duan, G.-R.; Luo, X.-D.; Xie, Y.-M.; Luo, H.; Zhou, Y.-X.; Pan, B.-R. Expression of TIMP-1 and TIMP-2 in rats with hepatic fibrosis. World J. Gastroenterol. 2004, 10, 86–90. [Google Scholar] [CrossRef]
- Ghosh, A.K.; Vaughan, D.E. PAI-1 in tissue fibrosis. J. Cell. Physiol. 2012, 227, 493–507. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.; Bakir, A.S.; Shabana, S.S. Serum TGF-β, Serum MMP-1, and HOMA-IR as non-invasive predictors of fibrosis in Egyptian patients with NAFLD. Saudi J. Gastroenterol. 2012, 18, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Ladouceur, M.; Baron, S.; Nivet-Antoine, V.; Maruani, G.; Soulat, G.; Pereira, H.; Blanchard, A.; Boutouyrie, P.; Paul, J.L.; Mousseaux, E. Role of myocardial collagen degradation and fibrosis in right ventricle dysfunction in transposition of the great arteries after atrial switch. Int. J. Cardiol. 2018, 258, 76–82. [Google Scholar] [CrossRef]
- Ganger, M.T.; Dietz, G.D.; Ewing, S.J. A common base method for analysis of qPCR data and the application of simple blocking in qPCR experiments. BMC Bioinform. 2017, 18, 534. [Google Scholar] [CrossRef] [PubMed]
- Horbelt, D.; Denkis, A.; Knaus, P. A portrait of Transforming Growth Factor β superfamily signalling: Background matters. Int. J. Biochem. Cell Biol. 2012, 44, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Samarakoon, R.; Overstreet, J.M.; Higgins, P.J. TGF-β signaling in tissue fibrosis: Redox controls, target genes and therapeutic opportunities. Cell. Signal. 2013, 25, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhou, X.; Gao, H.; Ji, S.J.; Wang, C. The expression of epidermal growth factor and transforming growth factor-beta1 in the stenotic tissue of congenital pelvi-ureteric junction obstruction in children. J. Pediatr. Surg. 2003, 38, 1656–1660. [Google Scholar] [CrossRef]
- Koca, O.; Kaya, C.; Ozturk, M.I.; Gunes, M.; Gumrukcu, G.; Karaman, M.I. Analysis of expression of TNF-alpha and TGF-beta3 in intrinsic ureteropelvic junction obstruction. Bratisl. Med J. 2013, 114, 498–502. [Google Scholar] [CrossRef]
- Seremetis, G.M.; Maizels, M. TGF-beta mRNA expression in the renal pelvis after experimental and clinical ureteropelvic junction obstruction. J. Urol. 1996, 156, 261–266. [Google Scholar] [CrossRef]
- Duymelinck, C.; Dauwe, S.E.; De Greef, K.E.; Ysebaert, D.K.; Verpooten, G.A.; De Broe, M.E. TIMP-1 gene expression and PAI-1 antigen after unilateral ureteral obstruction in the adult male rat. Kidney Int. 2000, 58, 1186–1201. [Google Scholar] [CrossRef]
- Vaughan, D.E.; Lazos, S.A.; Tong, K. Angiotensin II regulates the expression of plasminogen activator inhibitor-1 in cultured endothelial cells. A potential link between the renin-angiotensin system and thrombosis. J. Clin. Investig. 1995, 95, 995–1001. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.K.; Vaughan, D.E. PAI-1 in tissue fibrosis. J. Cell. Physiol. 2012, 227, 493–507. [Google Scholar] [CrossRef] [PubMed]
- Görbe. ; Magyar, Z.; Schönleber, J.; Romics, M.; Hruby, E.; Nagy, B.; Sulya, B.; Beke, A.; Harmath,.; Jeager, J.; et al. Expression of VEGF in Neonatal Urinary Obstruction: Does Expression of VEGF Predict Hydronephrosis? Med. Sci. Monit. 2015, 21, 1319–1323. [Google Scholar] [CrossRef] [PubMed]
- Bickelhaupt, S.; Erbel, C.; Timke, C.; Wirkner, U.; Dadrich, M.; Flechsig, P.; Tietz, A.; Pföhler, J.; Gross, W.; Peschke, P.; et al. Effects of CTGF Blockade on Attenuation and Reversal of Radiation-Induced Pulmonary Fibrosis. J. Natl. Cancer Inst. 2017, 109. [Google Scholar] [CrossRef]
- Valle-Tenney, R.; Rebolledo, D.L.; Lipson, K.E.; Brandan, E. Role of hypoxia in skeletal muscle fibrosis: Synergism between hypoxia and TGF-β signaling upregulates CCN2/CTGF expression specifically in muscle fibers. Matrix Biol. 2020, 87, 48–65. [Google Scholar] [CrossRef] [PubMed]
- Roestenberg, P.; van Nieuwenhoven, F.A.; Joles, J.A.; Trischberger, C.; Martens, P.P.; Oliver, N.; Aten, J.; Höppener, J.W.; Goldschmeding, R. Temporal expression profile and distribution pattern indicate a role of connective tissue growth factor (CTGF/CCN-2) in diabetic nephropathy in mice. Am. J. Physiol. Physiol. 2006, 290, F1344–F1354. [Google Scholar] [CrossRef]
- Wang X. and Cretoiu D., “Advances in Experimental Medicine and Biology 913 Telocytes Connecting Cells.” [Online]. Available online: http://www.springer.com/series/5584.
- Huang, B.-L.; Brugger, S.M.; Lyons, K.M. Stage-specific Control of Connective Tissue Growth Factor (CTGF/CCN2) Expression in Chondrocytes by Sox9 and β-Catenin*. J. Biol. Chem. 2010, 285, 27702–27712. [Google Scholar] [CrossRef]
- Airik, R.; Trowe, M.-O.; Foik, A.; Farin, H.F.; Petry, M.; Schuster-Gossler, K.; Schweizer, M.; Scherer, G.; Kist, R.; Kispert, A. Hydroureternephrosis due to loss of Sox9-regulated smooth muscle cell differentiation of the ureteric mesenchyme. Hum. Mol. Genet. 2010, 19, 4918–4929. [Google Scholar] [CrossRef]
- Ampawong, S.; Klincomhum, A.; Likitsuntonwong, W.; Singha, O.; Ketjareon, T.; Panavechkijkul, Y.; Zaw, K.-M.; Kengkoom, K. Expression of Aquaporin-1, -2 and -4 in Mice with a Spontaneous Mutation Leading to Hydronephrosis. J. Comp. Pathol. 2012, 146, 332–337. [Google Scholar] [CrossRef]
- Paces-Fessy, M.; Fabre, M.; Lesaulnier, C.; Cereghini, S. Hnf1b and Pax2 cooperate to control different pathways in kidney and ureter morphogenesis. Hum. Mol. Genet. 2012, 21, 3143–3155. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



