
Submitted:
11 August 2023
Posted:
13 August 2023
You are already at the latest version
Abstract
Plant-based extracts have been recently employed as sustainable tools to improve biotic and abiotic stress tolerance and increase grape quality. However, knowledge about the effect of these extracts on secondary metabolism compounds, that are fundamental for grape and wine quality, is still scarce. In this study, a trial was installed in an experimental vineyard with the variety Touriga Franca located at University of Trás-os-Montes e Alto Douro, Baixo Corgo sub-region of the Douro Demarcated Region, in two growing seasons: 2019 and 2020. Foliar sprayings with the natural products, nettle extract (NE) and Japanese knotweed extract (JKE), were tested to evaluate their effect on downy mildew incidence and severity, on grapevines bioactive compounds contents and antioxidant activity, at veraison and harvest. NE and JKE have stimulated an improvement in the concentration of total carotenoids in leaves and the concentration of total phenolics in berries along with an improved concentration of total phenolics and flavonoids in leaves and berries. Thus, it was verified that these extracts enhance grapevines bioactive compounds and antioxidant capacity and, consequently, the physiological performance of the plant and the quality of the berries. Considering the challenges posed by climate change in the Mediterranean basin viticulture sector, the use of plant extracts might represent a sustainable tool to mitigate the increasing severity of drought, often associated to heatwaves and high irradiation. Moreover, downy mildew incidence results suggest that an integrated disease management approach combining conventional control with alternative fungicides can be used in the future for a more sustainable viticulture.
Keywords:
Antioxidant activity
; climate change
; Douro Demarcated Region
; phenolic compounds
; plant extracts
; sustainable viticulture
; Vitis vinifera L.
1. Introduction
In Mediterranean viticulture, such as in Douro Demarcated Region (DDR), climate is the main factor affecting grape and wine quality [1]. Extreme weather conditions, such as heatwaves, continuous drought and spring frost will have an impact on flowering and fruit set, ripening and berries size, which will decrease grapevine yield, and berries and wine quality[1,2]. On the other hand, climate change will increase the pressure of pests and diseases on the vines, due to milder winters [2,3]. Vitis vinifera L. is susceptible to many fungi and fungi-like pathogens, whose infections will also reduce fruit quality and decrease plant vigor and yield [4,5,6]. These diseases lead to excessive use of fungicides in viticulture, and thus there is a growing concern regarding its residues in wine and effects on human and environmental health [7,8,9,10]. Grapevine productivity and berries quality are the most important concerns for winegrowers because they directly determine their profits [1]. In recent years, 30% of the authorized products for downy mildew prevention have been removed from the market, mainly due to their toxic effects. According to the Kumming-Montereal Agreement, and the European Directive for Sustainable Use of Pesticides (2009/128/EC), a sustainable food system is the main objective, with a 30% reduction in pesticide use by 2030. Alternative fungicides to prevent grape fungal diseases are emerging, namely natural products derived from plant extracts. Some of these contain several active compounds that have a positive role in the physiological performance of the grapevines and, consequently, improve yield and fruit quality [11]. The search for alternative solutions to fungicides to be widely applied has been a goal in agriculture. Some plants, like Urtica species have antimicrobial effects on a variety of plant pathogens, such as Botrytis cinerea (in tomato) [12], Alternaria sp. (in tomato) [13] and Phytophthora infestans (in potato) [14]. Products based on nettle (Urtica spp.) extract are rich in nitrogen, promoting the growth and quality of some species, such as, celeriac (Apium graveolens var. rapaceum) [15], white head cabbage (Brassica oleracea var. capitata) [16] and bean (Phaseolus vulgaris) [17]. These extracts contain many oligoelements, which are used as acaricides and also have a repelling effect on many insects [18]. Japanese knotweed (Reynoutria spp.) extracts have been used as antifungal against Plasmopara viticola (in grapevine) [19] and Blumeria graminis (in barley) [20]. Plants of the Reynoutria genus contain a large complex of biologically active substances, including phenolic compounds, such as, resveratrol, piceid, furans, phenol carboxylic acids and their derivatives, lignans, coumarins, catechins, naphthoquinones, and anthraquinones [21,22,23].
In this context, the main objective of this work was to evaluate the effect of foliar applications of plant extracts (nettle and Japanese knotweed) on grapevine downy mildew incidence and on grapevine bioactive compounds and antioxidant capacity. Thus, berries quality was analyzed, aiming to contribute to the increase of the profitability and sustainability of vines production while reducing the use of chemicals to prevent and control fungal diseases.
2. Materials and Methods
2.1. Plant material and sampling
Samples were obtained from the black skinned Vitis vinifera cv. Touriga Franca, in two growing seasons: 2019 and 2020. The trial was installed in an experimental vineyard located at University of Trás-os-Montes e Alto Douro (41°17’14.8”N 7°44’14.8”W, 500m above sea level), Baixo Corgo sub-region of the Douro Demarcated Region, Vila Real, Northern Portugal. Vines were managed in rainfed conditions and grown using standard cultural practices commonly employed by commercial farmers. This area has peculiar climatic characteristics such as a warm-temperate climate with dry and hot summers, and higher precipitation during the winter months and very low during the summer. Monthly temperature and precipitation values were recorded by a weather station located near to the experimental site and are shown in Figure 1.
Four vineyard rows, with 35 plants each were sprayed between leaves unfolded (BBCH 11) and veraison (BBCH 81) [24]. Foliar sprayings were carried out during the morning, covering the whole canopy and according to disease incidence and weather conditions, in a total of ten and fourteen applications in 2019 and 2020, respectively. Four different foliar treatments were tested, namely: i) nettle extract - NE (Urtica spp.) (3%); ii) Japanese knotweed extract - JKE (Reynoutria japonica) (4.5%); iii) conventional treatment - CT (4.8% (p/p) cymoxanil + 40% (p/p) folpet + 8% (p/p) metalaxyl (SAPEC) or 10.1% (p/p) penconazole (SAPEC) or 50% (p/p) tebuconazole + 25% (p/p) trifloxystrobin (BAYER) (prepared according to the manufacturer guidelines), and iv) control – C (water) (4 treatments x 6 plants x 3 replicates). All the products were dissolved in water. Leaves and grapes were sampled directly to liquid nitrogen. Sampling occurred in two development stages: veraison (BBCH 81) and harvest (BBCH 89) [24], in three randomized blocks of six plants in each of the four treatment conditions. The samples were kept at -80°C until analysis.
2.2. Downy mildew incidence
Grapevine downy mildew (Plasmopara viticola) was evaluated according to the guidelines EPPO/OEPP PP 1/31 (3) [25]. To assess disease incidence (0 no disease or 1 disease) and severity (percentage of leaves and bunches with downy mildew symptoms), six vines randomly selected from each of the three replicates at each evaluation date were examined. Disease incidence was assessed in leaves and bunches, in three growth stages: pea-size (BBCH 75), veraison (BBCH 81) and harvest (BBCH 89) [24]. The artificial inoculation of downy mildew under controlled conditions has been attempted, however without success.
2.3. Determination of bioactive compounds
For sample extraction, in the case of leaves, 10mg of fresh material of each sample were weighted and then 4 mL of 80% (v/v) acetone were added. Samples were vortexed 3 minutes, sonicated 10 minutes and finally centrifuged at 4000rpm, for 5 minutes, at 4°C. For berries, 40mg of dry material (lyophilized and powdered with liquid nitrogen) of each sample were weighted, then added 950µL of 70% (v/v) methanol, mixed thoroughly the extract in a vortex and the mixture submitted to 70°C for 30 minutes, and finally centrifuged at 13000rpm, at 1°C, for 15 minutes. These extracts were stored at -20°C and used for the determination of the total carotenoids, total phenolics, and flavonoids contents and in antioxidant activity assays. All results are expressed as the average of six replicates with standard error (SE) shown.
2.3.1. Total carotenoids
The concentration of total carotenoids was quantified according to Lichtenthaler (1987) [26], using the leaves and berries sample extracts. The absorbance was measured at 663, 645 and 470nm. The results were expressed as µg.g-1 fresh weight (FW) in leaves or dry weight (DW) in berries and obtained according to the following equation: Total carotenoids = (1000*Abs470 – 1.82*Cla – 85.02*Clb)/198)*0.001, where Cla (Chlorophyll a = (12.7*Abs663 – 2.69*Abs645)/1000) and Clb (Chlorophyll b = (22.9*Abs645 – 4.68*Abs663)/1000).
2.3.2. Total phenolics
The concentration of total phenolics was determined according to the Folin–Ciocalteu colorimetric method at 765nm [27], using a gallic acid calibration curve and the results were expressed as mg of gallic acid equivalents (GAE).g-1 of FW in leaves or DW in berries.
2.3.3. Flavonoids
Flavonoid concentration in the same extracts was determined at 510nm according to the colorimetric method [28]. A calibration curve was prepared with catechin, and the results were expressed as mg of catechin equivalents (CE).g-1 of FW in leaves or DW in berries.
2.3.4. Total anthocyanins
The total monomeric anthocyanin (TMA) content was determined according to Lee et al., Meng et al., and Ali Shelat et al. [29,30,31]. To obtain the extracts, 5mL of methanol acidified with 1% HCl were added to 50mg of berries or 20mg of leaves. The mixture was shaken and placed in the dark at 4°C for 1 hour. It was then centrifuged at 4000rpm for 15 minutes at 4°C. Subsequently, the supernatant was collected. In a microplate, the mixture of 50μL of extract and 250μL of 0.025M KCl (pH= 1.0) or 50μL of extract and 250μL of 0.4M sodium acetate buffer (pH= 4.5) was pipetted into different wells. Finally, absorbances were read at 510 and 700nm. The concentration of total monomeric anthocyanins was expressed as mg of cyanidin-3-O-glucoside equivalents (CGE).g-1 of FW in leaves or DW in berries, according to the following formula: TMA = , where MW is the molecular weight of cyanidin-3-O-glucoside (449 g.mol-1); DF is the dilution factor; ε is the molar extinction coefficient of cyanidin-3-O-glucoside (29.600); C is the concentration of extracted volume and A = (A510 – A700)pH1.0 – (A510 – A700)pH4.5.
2.4. Antioxidant activity assays
2.4.1. ABTS•+ radical-scavenging activity
To determine the radical-scavenging activity of leaf and fruit extracts the discoloration assay ABTS•+ (2,2′-azino-bis (3-ethylbenzothiazoline-6-sulphonic acid)) was used, according to Re et al. and Stratil et al. [32,33]. To prepare the ABTS•+ work solution, 7mM ABTS were mixed with 140mM K2S2O8 in double distilled water. The mixture was incubated for 12–16 hours in the dark at room temperature and then its absorbance was adjusted to 0.7-0.8 with absolute ethanol in a wavelength of 734nm. After that, 15µL of leaf or berry extracts and 285µL of the ABTS•+ work solution was mixed and left for 10 minutes in the dark, and the absorbance was read at 734nm. The blank was made with 80% acetone in leaves or 70% methanol in berries plus ABTS•+ solution. The results were expressed as µmol Trolox.µg-1 of FW or DW in leaves or berries, respectively, using a Trolox calibration curve.
2.4.2. DPPH radical-scavenging activity
The DPPH (2,2-diphenyl-1-picrylhydrazyl) radical-scavenging activity assay was carried out according to Brand-Williams et al., Sánchez-Moreno et al., and Siddhraju and Becker [34,35,36], mixing 15µL of leaf or berry extracts with 285µL methanolic solution containing DPPH radicals (10-5 mol.L-1). The blank was made with 15µL of 80% acetone in leaves or 70% methanol in berries and 285µL methanolic solution containing DPPH radicals. The mixture was vigorously shaken and left to stand for 30 minutes in the dark. The reduction of the DPPH radical was detected by measuring samples absorbance at 517nm. Using a Trolox calibration curve the results were expressed as µmol Trolox.µg-1 of FW or DW in leaves or berries, respectively.
2.4.3. FRAP assay
The FRAP (Ferric Reducing Antioxidant Power) assay used in this study was a modification of the previous methods of Stratil et al. and Benzie and Strain [33,37]. To prepare the FRAP reagent, 1 volume of an aqueous 10mM solution of TPTZ (2,4,6-Tri(2-pyridyl)-s-triazine) in 40mM HCl was mixed with the 1 volume of 20mM FeCl3.6H2O and 10 volumes of 300mM acetate buffer, pH 3.6. Then, 25µL of leaf or berry extracts were mixed with 275µL of FRAP reagent. The blank was made with 25µL of 80% acetone in leaves or 70% methanol in berries and 275µL of FRAP reagent. The mixture was vigorously shaken and left to stand for 5 minutes in the dark and the absorbance at 593nm was recorded. Using a Trolox calibration curve the results were expressed as µmol Trolox.µg-1 of FW or DW in leaves or berries, respectively.
2.5. Statistical analysis
Data were analyzed by using SPSS Statistics for Windows (IBM SPSS Statistics for Windows, Version 23.0. Armonk, NY: IBM Corp). Statistical differences between treatments in each phenological stage and in each year, were evaluated by one-, two-, and three-way ANOVA, followed by Duncan’s multiple range test at P < 0.05. Pearson’s correlation analysis was used to determine the relationship between bioactive compounds and antioxidant activity in leaves and berries.
3. Results
3.1. Downy mildew incidence
In 2019, downy mildew was not detected in the trial under study. Although precipitation occurred in April, the temperatures were not favorable for Plasmopara viticola development, and in the following months the precipitation values were very low and temperatures high (Figure 1).
In 2020, downy mildew manifested itself with some intensity, mainly in bunches (Figure 2). In May of 2020 meteorological conditions were favorable for downy mildew development, given the rainfall and mild temperatures (Figure 1). It was also verified in leaves and berries that the incidence of the disease was high, however the severity was low. The spraying with plant extracts had a positive effect in reducing the incidence of downy mildew in 2020 on leaves, at pea-size (JKE and NE) and at harvest (NE), when compared to control (C), although with no significant differences (Figure 2A). It was also found a positive effect on disease severity, mainly at pea-size and harvest (Figure 2C). In berries, at veraison, there was a decrease in disease incidence in plants treated with JKE compared to untreated plants (Figure 2B). At harvest, mainly in berries (Figure 2B), the tested treatments did not show effectiveness to reduce downy mildew incidence, but JKE seems to decrease the severity in comparation to C (Figure 2D). The rainfall during the months of August and September (Figure 1), may have led to the progression of the disease that was already installed in these grapevines in the previous stages (pea-size and veraison).
3.2. Effect on leaf and berry bioactive compounds
The total carotenoids concentration in leaves and berries from cv. Touriga Franca is shown in Figure 3. In leaves the concentration of total carotenoids was affected by year (P < 0.001), by phenological stage (P < 0.001), by the interaction between treatment and year (P < 0.001), by the interaction between year and phenological stage (P < 0.05) and by the interaction of treatment, year and phenological stage (P < 0.001) (Table S1). In case of berries was affected by treatment (P < 0.01), by the phenological stage (P < 0.001) and by the interaction between year and phenological stage (P < 0.001) (Table S1). The total carotenoids concentration was higher in average, for both leaves (328 µg.g-1) and berries (55 µg.g-1), at harvest of 2019. In this year and in all treatments, the carotenoids concentration in leaves and berries increased in average 54% and 80%, respectively, from veraison to harvest. In 2020, the carotenoids concentration at veraison and harvest stages was very similar, for both leaves and berries.
At veraison of 2019, significant differences were detected in leaves, specifically between nettle extract (NE) and the remaining treatments, with higher total carotenoids concentration (253 µg.g-1) being observed for this treatment (Figure 3A). In terms of total carotenoids in the berries, no major differences were found between treatments, however CT seemed to slightly increase the total carotenoids in berries (about 10 µg.g-1), at harvest stage, when compared to control (C) (Figure 3).
Table 1 shows the data of total phenolic, total anthocyanin, and flavonoid concentrations, in leaves and berries, sampled in veraison and harvest of 2019 and 2020 in grapevines treated with NE, JKE, CT (conventional fungicide) and C (water).
For total phenolics, total anthocyanin and flavonoid concentration, data recorded in berries were higher than in leaves. Furthermore, comparing the two analyzed years these parameters were higher in 2020 than in 2019 (not considering the total anthocyanins content in leaves, which was residual in both years). The differences were more pronounced in the flavonoids content in berries at veraison, with values in 2020 being 32% higher in average than in 2019. For the remaining parameters, the values obtained in 2020 were also higher than those of 2019, ranging from 71% (flavonoids in leaves at harvest) to 48% (total anthocyanins in berries at harvest) higher in average.
The concentration of total phenolics in leaves were influenced by the treatment (P < 0.01) and year (P < 0.001) (Table S1). At veraison of 2020, significant differences were detected among treatments, with an increase of 19% in NE (27.33 mg.g-1) and 21% in JKE (28.09 mg.g-1) compared to control (22.19 mg.g-1) grapevines. In berries was verified an influence of year (P < 0.001), phenological stage (P < 0.001), the interaction between treatment and year (P < 0.05), the interaction between year and phenological stage (P < 0.001) (Table S1). At veraison 2019, the total phenolics content was significantly higher in plants treated with NE with an increase of 30% when compared to C. Although with no significant differences, in 2020, both in berries at veraison and at harvest, the highest content of total phenolics was detected in grapevines treated with JKE (increasing 21% at veraison and 18% at harvest, in comparison to C).
In this study, the total anthocyanins content was affected by the treatment (P < 0.001), the year (P < 0.001), the phenological stage (P < 0.001) the interaction between treatment and year (P < 0.05) and the interaction between year and phenological stage (P < 0.001) (Table S1). The concentration of total anthocyanins in leaves was residual (data not shown). In berries, the concentration of total anthocyanins increased 1.04 times in 2019 and 3.02 times in 2020 between veraison and harvest, of both years. At veraison and harvest of 2019, sprayings with plant extracts lead to berries with higher total anthocyanins content than control, namely in NE (more 35% at veraison and 28% at harvest) and JKE (more 38% at veraison and 11% at harvest). In fact, in both phenological stages and years, there was always at least one natural treatment with anthocyanin contents higher than those detected in the control berries (veraison 2019 – NE and JKE; veraison 2020 - NE; harvest 2019 – NE and JKE; harvest 2020 –JKE) (Table 1).
Flavonoids content in leaves were affected by treatment (P < 0.001), year (P < 0.001) and phenological stage (P < 0.01) (Table S1) and in berries by year (P < 0.001), phenological stage (P < 0.001) and by the interaction between year and phenological stage (P < 0.001) (Table S1). It was verified that flavonoids content in leaves increased in average 20% in 2019 and 11% in 2020 from veraison to harvest, in both years and in all treatment conditions, except for CT in 2020. In the opposite, in berries, in all treatments, the flavonoids content decreased in average 2% and 46% from veraison to harvest of 2019 and 2020, respectively. The highest flavonoids content (16.80 mg.g-1) was found in berries at veraison 2020. The flavonoids content was higher at veraison (2020) and harvest (2019) in leaves sprayed with NE (11% and 19% higher, respectively) and JKE (16% and 11% higher, respectively) than in control leaves. At harvest of 2020, flavonoids content in leaves was also higher in treated plants compared to untreated ones (24% higher in JKE-treated grapevines). In berries, at the veraison of 2019 and harvest of 2020, the flavonoids contents were higher in both plant extract treatments compared to CT or C grapevines: NE, 5.67 and 9.03 mg.g-1; JKE, 7.19 and 9.61 mg.g-1; conventional treatment, 4.33 and 9.33 mg.g-1 and control, 4.60 and 8.11 mg.g-1, respectively (Table 1).
3.3. Effect on leaf and berry antioxidant activity
In this study three antioxidant assays, ABTS•+, DPPH, and FRAP, were used to access the antioxidant activity in leaves and berries at veraison and harvest of 2019 and 2020 (Table 2). In case of ABTS•+ significant differences were found for leaves in treatment (P < 0.001) and year (P < 0.001) for berries in treatment (P < 0.05), year (P < 0.001) and phenological stage (P < 0.01) (Table S1). DPPH was affected by year (P < 0.001) and phenological stage (0.05 < P < 0.01) in leaves and berries. It was also verified an influence of year (P < 0.001) and the interaction between treatment and year (P < 0.01) in FRAP for both leaves and berries (Table S1).
Antioxidant activity was much higher (60 times in average) in fruits than in leaves, as expected. Unlike the antioxidant compound contents, which were higher in 2020 than 2019, the ABTS•+ assay revealed higher antioxidant activity in 2019 than in 2020 in leaves and berries at veraison and harvest, with more pronounced differences in leaves at harvest. DPPH in berries and FRAP in leaves were also higher in 2019 than in 2020 both at veraison and harvest. Conversely, DPPH in leaves and FRAP in berries were higher in 2020 than 2019 both at veraison and harvest.
There was a significant effect of the treatments in the antioxidant activity in leaves and berries at veraison and harvest for most treatments (for ABTS•+ at veraison and harvest of both years and berries at veraison of both years; for DPPH in leaves at both stages of 2019 and in berries at veraison of 2019; in FRAP in leaves at both stages of 2019 and in veraison of 2020 in berries in both stages and years). At least one of the tested products (NE, JKE) revealed a positive effect on the antioxidant activity of the leaves in both phenological stages and years in comparison to the conventional treatment, although lower in comparison to control. Nevertheless, for FRAP, at veraison of 2019, JKE-treated leaves (0.74 µmol Trolox.µg-1) showed a positive effect in comparison with control (0.70 µmol Trolox.µg-1). Similar situation was verified in harvest of 2019 for the natural products (NE 0.62 µmol Trolox.µg-1 and JKE 0.66 µmol Trolox.µg-1) in comparison with control (0.58 µmol Trolox.µg-1). The antioxidant capacity evaluated by the different methodologies revealed a significative increment in leaves from plants treated with the natural products (NE and JKE) in comparison to the conventional treatment at veraison 2019 and 2020 and at harvest 2019 (ABTS•+ and DPPH).
In berries, both plant extracts revealed a positive effect (1.1 times in average) on the antioxidant capacity at veraison 2019 and harvest 2019 in relation to CT, measured by FRAP and DPPH, respectively. The sprayings with NE (34.58 µmol Trolox.µg-1 in 2019 and 69.77 µmol Trolox.µg-1 in 2020) and Japanese knotweed extracts (24. 87 µmol Trolox µg-1 in 2019 and 66.21 µmol Trolox µg-1 in 2020) led to berries with higher antioxidant capacity (determined by FRAP) at veraison and with NE (70.52 µmol Trolox µg-1 in 2019 and 61.74 µmol Trolox µg-1 in 2020) (determined by DPPH) at harvest, in both years compared to CT. In 6 of the 12 combinations berry/year/phenological stage/antioxidant activity assay, NE-treated berries revealed a higher antioxidant capacity than the CT.
Analyzing the correlations between non-enzymatic antioxidant compounds and antioxidant activity, in leaves FRAP showed a statistically significant (P < 0.01) positive correlation with total phenolics and flavonoids in 2019 at veraison (R2 = 0.584 and R2 = 0.530) and harvest (R2 = 0.654 and R2 = 0.404). In 2020, in leaves, a statistically significant (P < 0.01) positive correlation was found between FRAP and total phenolics at veraison (R2 = 0.476) and harvest (R2 = 0.438).
For berries, in 2019 FRAP assay showed a significant (P < 0.01) and positive correlation with total phenolics in veraison and harvest (R2 = 0.841 and R2 = 0.607, respectively) and in 2020, a positive correlation with total phenolics was verified at veraison (R2 = 0.560, P < 0.01) and at harvest (R2 = 0.495, P < 0.05).
4. Discussion
4.1. Downy mildew incidence
During the year of 2019, downy mildew was not detected in the trial, and according to ADVID [38] this year was considered of low pressure of diseases. The year 2020 was considered hot and dry [39], and differences in disease incidence and severity were observed in the trial. During the season of 2020, the leaves at pea-size and harvest sprayed with NE and JKE revealed a reduction of downy mildew severity, when compared to control plants. Berries at pea-size sprayed with NE showed a reduction of downy mildew incidence, and berries sprayed with JKE at harvest a reduction on severity. Extracts obtained from these plants are a mixture of active ingredients, that contain many secondary metabolites which display several mechanisms of actions, such as direct toxicity against the pathogen or a capacity to stimulate the defense system of the vine [40,41,42,43]. New products with formulations involving nettle and Japanese knotweed extracts could be tested in the future as eco-friendly strategies to control downy mildew.
4.2. Leaf and berry bioactive compounds
Carotenoids protect the photosynthetic apparatus from photooxidative damage, preventing the formation of harmful reactive oxygen species (ROS), and by being directly involved in the extinction of these activated oxygen species. If this protective mechanism fails or becomes overloaded, ROS can accumulate, leading to lipid peroxidation [44], irreversible DNA damage and cell death [45]. NE and JKE increased the total carotenoids concentration in leaves, indicating potential in the protection of the photosynthetic apparatus, as well as helping prevent lipid peroxidation in plants under summer stress. In fact, NE and JKE tended to increase primary metabolites in the leaves, such as chlorophyll a, chlorophyll b, and total soluble sugars, among others (data not shown). Moreover, it is known that the application of natural products may contribute to fruit nutritional value due to the increment of secondary metabolites, such as carotenoids [46,47].
In 2020 the concentration of total phenolics, total anthocyanins and flavonoids in berries was higher than in 2019. These differences can be explained due to differences in the climatic conditions of both years (Figure 1). Bunch exposure to high temperature and radiation may increase anthocyanins, flavonols, and flavanols synthesis in grapes due to increased activity of the phenylalanine ammonium lyase (PAL) enzyme [48]. In addition, the year of 2020 also recorded higher precipitation values and warmer temperatures in March and May (Figure 1), leading to a high pressure of diseases, with emphasis on downy mildew due to its precocity. These factors may have led to the accumulation of polyphenols, and specifically of anthocyanins in grapevine, in response to the stress conditions in the year of 2020.
The total phenolics content in leaves was increased in the treatments under study; with similar results being obtained by Singh et al. and Irani et al. [49,50] with the application of chitosan and a seaweed extract, respectively. In berries at veraison 2019 and at both phenological stages in 2020, the total phenolics content was significantly higher in grapevines sprayed with the plant extracts under study, compared to control grapevines. Since berry quality, particularly of the grapevine cv. Touriga Franca, is largely dependent on the accumulation of secondary metabolites such as polyphenols and volatiles [51], these extracts could be used to improve berry quality in the future. Similar results were obtained by Kok [52], who described an increase in phenolic content in the table grapevine Cardinal treated with gibberellic acid. Other studies using chitosan [47] and different biostimulants [53], in species such as strawberry and rocket plant (Eruca vesicaria subsp. sativa) [46] also reported an increase in total phenolics content. There are many functions attributed to phenolic compounds, highlighting their action in defense mechanisms, fruit color and their antioxidant effect. Given the Mediterranean climate conditions to which the plants under study were subjected, which is characterized by water deficit, high temperature and excessive sunlight radiation in the summer, the synthesis of phenolics is very important, as these compounds are capable of reducing the effects of the stress conditions, offering resistance to photooxidation by absorption of UV radiation [54]. Furthermore, given the antioxidant activity of some phenolic compounds, they may be able to neutralize ROS [55].
In this study, the total anthocyanins content was affected by the foliar application of plant extracts. In berries, NE and/or JKE treatment increased anthocyanins content when compared to control: at veraison and harvest 2019 in plants treated with NE and JKE, at veraison 2020 in plants treated with NE; and at harvest 2020 in plants treated with JKE. Increases of anthocyanins content in treated grapevines have also been verified by other authors, which studied the effect of chitosan in the cv. Touriga Franca [49], and with the application of Ascophyllum nodosum extracts in the cv. Sangiovese [56].
It is known that flavonoids represent a huge fraction of the soluble phenolics present in grapevine and play different physiological roles, being often involved in protection against biotic and abiotic stress [57]. The concentration of flavonoids in berries varied during the different development stages: the accumulation starts on fruit set until 1–2 weeks after veraison (the onset of ripening) and then their level decrease [57], as we verified in this study. The application of NE and JKE extracts increased the flavonoids content in leaves and grapes, which can improve the plant resistance to summer stress, a major problem in DDR [58,59].
4.3. Leaf and berry antioxidant activity
Antioxidant activity determination in the leaves of grapevines treated with NE and JKE extracts revealed a significant increase compared to the CT but was lower than in control (C). In general, JKE-treated leaves presented the highest antioxidant activity, suggesting that switching from conventional fungicides to natural alternatives does not compromise, or even improve the antioxidant capacity of the grapevines. Singh et al. [49] observed an increase in antioxidant activity of leaves from Touriga Franca plants sprayed with chitosan.
In berries, treatments revealed a significative effect on the antioxidant capacity in FRAP in both years and stages; in ABTS•+ at veraison of both years; and in DPPH at harvest 2019. NE-treated grapevines had higher antioxidant capacity than the CT plants. Similar results were obtained by Godlewska et al. [16], in Brassica oleracea var. capitata sprayed with nettle extract. It is known that the application of alternative compounds can lead to changes in enzyme activity and influence the antioxidant properties of compounds, such as, lycopene, ascorbic acid, and phenolic compounds [60]. In plants, several antioxidants can help to handle adverse growth conditions [61]. In this case, NE appears to influence antioxidant activity and could be used in the future to mitigate the effects of climate change in plants under summer stress. The compounds naturally present in plants are responsible for the antioxidant activity [62], and different bioactive compounds are found in both Japanese knotweed and nettle. In the case of Japanese knotweed, flavonoids, stilbenes, anthraquinones, coumarins, and lignans, can be found in roots, leaves, stems and flowers [40,42]. In the case of nettle, bioactive compounds can also be found in different parts of the plant, and include flavonoids, phenolic acids, amino acids, carotenoids, organic acids, and fatty acids [43]. It was verified that the antioxidant activity, namely FRAP, was correlated whit some non-enzymatic antioxidant compounds in leaves and berries, showing a statistically significant (P < 0.01) positive correlation with total phenolics in both leaves and berries, in both years and phenological stages. Other studies in grapevine, such as those of Rockenbach et al. and Kupe et al. [63,64], have also found a significant correlation between total phenol content and antioxidant activity. A statistically significant correlation between FRAP and free phenolic acids has also been reported by Speranza et al. [65] in sorghum kernels.
5. Conclusion
This study, comparing downy mildew protection using conventional fungicide and two alternative products (nettle and Japanese knotweed extracts) under field conditions, highlights their stimulatory effect on the leaves and berries non-enzymatic antioxidant compounds, as well as a slight increase in the berry antioxidant activity.
In 2020 was possible to evaluate the effect of the tested plant extracts on downy mildew, being verified that these putative alternative fungicides may decrease the incidence and severity of the disease, mainly in the leaves. In the future, with the need to reduce pesticides, these extracts could be an alternative, or could even be used in new formulations reducing the synthetic fungicides. They can be applied from the beginning of the growing season in order to reduce the incidence and severity of downy mildew in the leaves, as well as decreasing the probability of its incidence in the berries, reducing the use of chemicals and making viticulture more sustainable.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization: E.M., I.C. (Isabel Cortez), I.C. (Isaura Castro), and B.G. Investigation: E.M., M.B., S.P., S.C., and H.F. Writing—original draft: E.M. Writing—review and editing: E.M., M.B., S.P., S.C., H.F., I.C. (Isabel Cortez), I.C. (Isaura Castro), and B.G. Supervision: I.C. (Isabel Cortez), I.C. (Isaura Castro), and B.G. All authors approved the version to be published.
Funding
This research was funded by the European Regional Development Fund through the INTERREG Atlantic Area Program, under the project NASPA—Natural fungicides against air & soil borne pathogens in the Atlantic Area (EAPA-451/2016).
Acknowledgments
The author Eliana Monteiro acknowledges the financial support provided by the Portuguese Foundation for Science and Technology (FCT) (UI/BD/150730/2020), under the Doctoral Program “Agricultural Production Chains—from fork to farm” (PD/00122/2012). The authors acknowledge the support of National Funds by FCT, under the project UIDB/04033/2020.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gutiérrez-Gamboa G, Zheng W, Martínez de Toda F. Current viticultural techniques to mitigate the effects of global warming on grape and wine quality: A comprehensive review. Food Res Int 2021, 139. [CrossRef]
- Jones, G. Uma Avaliação do Clima para a Região Demarcada do Douro : Uma análise das condições climáticas do passado, presente e futuro para a produção de vinho. AVID: Portugal, 2013; pp. 5–80. [Google Scholar]
- Fraga H, De Cortázar Atauri IG, Malheiro AC, Moutinho-Pereira J, Santos JA. Viticulture in Portugal: A review of recent trends and climate change projections. OENOOne 2017, 51, 61–9. [Google Scholar] [CrossRef]
- Wong FP, Wilcox WF. Distribution of baseline sensitivities to azoxystrobin among isolates of Plasmopara viticola. Plant Dis 2000, 84, 275–81. [Google Scholar] [CrossRef]
- Aziz A, Poinssot B, Daire X, Adrian M, Bézier A, Lambert B, et al. Laminarin elicits defense responses in grapevine and induces protection against Botrytis cinerea and Plasmopara viticola. Mol Plant-Microbe Interact 2003, 16, 1118–28. [Google Scholar] [CrossRef]
- Belhadj A, Saigne C, Telef N, Cluzet S, Bouscaut J, Corio-Costet MF, et al. Methyl jasmonate induces defense responses in grapevine and triggers protection against Erysiphe necator. J Agric Food Chem 2006, 54, 9119–25. [CrossRef]
- Petit A, Wojnarowiez G, Panon M, Baillieul F, Clément C, Fontaine F, et al. Botryticides affect grapevine leaf photosynthesis without inducing defense mechanisms. Planta 2009, 229, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Jermini M, Blaise P, Gessler C. Quantification of the influence of the downy mildew (Plasmopara viticola) epidemics on the compensatory capacities of Vitis vinifera “Merlot” to limit the qualitative yield damage. Vitis - J Grapevine Res 2010, 49, 153–60. [Google Scholar]
- Delaunois B, Farace G, Jeandet P, Clément C, Baillieul F, Dorey S, et al. Elicitors as alternative strategy to pesticides in grapevine? Current knowledge on their mode of action from controlled conditions to vineyard. Environ Sci Pollut Res 2014, 21, 4837–46. [Google Scholar] [CrossRef]
- Garde-Cerdán T, Mancini V, Carrasco-Quiroz M, Servili A, Gutiérrez-Gamboa G, Foglia R, et al. Chitosan and Laminarin as alternatives to copper for Plasmopara viticola control: Effect on grape amino acid. J Agric Food Chem 2017, 65, 7379–86. [Google Scholar] [CrossRef]
- Monteiro E, Gonçalves B, Cortez I, Castro I. The role of biostimulants as alleviators of biotic and abiotic stresses in grapevine: A review. Plants 2022, 11, 396. [Google Scholar] [CrossRef]
- Ghazal HN, Al-Shahwany AW, Al-Dulaimy FT. Control of gray mold on tomato plants by spraying Piper nigrum and Urtica dioica extracts under greenhouse condition. Iraqi J Sci 2019, 60, 961–71. [Google Scholar] [CrossRef]
- Rodino S, Butu M, Butu A. Alternative antimicrobial formula for plant protection. Bull USAMV Ser Agric 2018, 75, 32–7. [Google Scholar] [CrossRef]
- Njogu M, Nyankanga R, Muthomi J, Muindi E. Studies on the effects of stinging nettle extract, phosphoric acid and conventional fungicide combinations on the management of potato late blight and tuber yield in the highlands of Kenya. J Agric Food Sci 2014, 2, 119–27. [Google Scholar]
- Godlewska K, Pacyga P, Michalak I, Biesiada A, Szumny A, Pachura N, et al. Field-scale evaluation of botanical extracts effect on the yield, chemical composition and antioxidant activity of celeriac (Apium graveolens L. var. rapaceum). Molecules 2020, 25, 4212. [Google Scholar] [CrossRef]
- Godlewska K, Pacyga P, Michalak I, Biesiada A, Szumny A, Pachura N, et al. Effect of botanical extracts on the growth and nutritional quality of field-grown white head cabbage (Brassica oleracea var. capitata). Molecules 2021, 26, 1992. [Google Scholar] [CrossRef]
- Maričić B, Radman S, Romić M, Perković J, Major N, Urlić B, et al. Stinging nettle (Urtica dioica L.) as an aqueous plant-based extract fertilizer in green bean (Phaseolus vulgaris L.) sustainable agriculture. Sustain 2021, 13. [CrossRef]
- Đurić M, Mladenović J, Bošković-Rakočević L, Šekularac G, Brković D, Pavlović N. Use of different types of extracts as biostimulators in organic agriculture. Acta Agric Serbica 2019, 24, 27–39. [Google Scholar] [CrossRef]
- Godard S, Slacanin I, Viret O, Gindro K. Induction of defence mechanisms in grapevine leaves by emodin- and anthraquinone-rich plant extracts and their conferred resistance to downy mildew. Plant Physiol Biochem 2009, 47, 827–37. [Google Scholar] [CrossRef]
- Hildebrandt U, Marsell A, Riederer M. Direct effects of physcion, chrysophanol, emodin, and pachybasin on germination and appressorium formation of the barley (Hordeum vulgare L.) powdery mildew fungus Blumeria graminis f. sp. hordei (DC.) Speer. J Agric Food Chem 2018, 66, 3393–401. [Google Scholar] [CrossRef]
- Zaki El-Readi M, Yehia Eid S, Al-Amodi HS, Wink M. Fallopia japonica: Bioactive secondary metabolites and molecular mode of anticancer. J Tradit Med Clin Naturop 2016, 05. [CrossRef]
- Oleszek M, Kowalska I, Oleszek W. Phytochemicals in bioenergy crops. Phytochem Rev 2019, 18, 893–927. [Google Scholar] [CrossRef]
- Borovaya S, Lukyanchuk L, Manyakhin A, Zorikova O. Effect of Reynoutria japonica extract upon germination and upon resistance of its seeds against phytopathogenic fungi Triticum aestivum L., Hordeum vulgare L., and Glycine max (L.) Merr. Org Agric 2020, 10, 89–95. [Google Scholar] [CrossRef]
- Lorenz DH, Eichhorn KW, Bleiholder H, Klose R, Meier U, Weber E. Growth stages of the grapevine: phenological growth stages of the grapevine (Vitis vinifera L. ssp. vinifera)—Codes and descriptions according to the extended BBCH scale. Aust J Grape Wine Res 1995, 1, 100–3. [Google Scholar] [CrossRef]
- EPPO. Plasmopara viticola. EPPO Bull 2001, 31, 313–7. [CrossRef]
- Lichtenthaler, HK. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzym 1987, 350–82. [Google Scholar] [CrossRef]
- Singleton V, Rossi J. Colorometry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am J Enol Vitic 1965, 144–58.
- Dewanto V, Wu X, Adom KK, Liu RH. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J Agric Food Chem 2002, 3010–4.
- Lee J, Durst RW, Wrolstad RE, Eisele T, Giusti MM, Haché J, et al. Determination of total monomeric anthocyanin pigment content of fruit juices, beverages, natural colorants, and wines by the ph differential method: Collaborative study. J AOAC Int 2005, 88, 1269–78. [Google Scholar] [CrossRef]
- Meng J-F, Fang Y-L, Qin M-Y, Zhuang X-F, Zhang Z-W. Varietal differences among the phenolic profiles and antioxidant properties of four cultivars of spine grape (Vitis davidii Foex) in Chongyi County (China). Food Chem 2012, 134, 2049–56. [Google Scholar] [CrossRef]
- Ali Shehat W, Sohail Akh M, Alam T. Extraction and estimation of anthocyanin content and antioxidant activity of some common fruits. Trends Appl Sci Res 2020, 15, 179–86. [Google Scholar] [CrossRef]
- Re R, Pellegrini N, Proteggente A, Pannala A, Yang M, Rice-Evans C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic Biol Med 1999, 26, 1231–7. [Google Scholar] [CrossRef] [PubMed]
- Stratil P, Klejdus B, Kubáň V. Determination of total content of phenolic compounds and their antioxidant activity in vegetablesevaluation of spectrophotometric methods. J Agric Food Chem 2006, 54, 607–16. [Google Scholar] [CrossRef] [PubMed]
- Brand-Williams W, Cuvelier ME, Berset C. Use of a free radical method to evaluate antioxidant activity. Lebnsm Wiss Technol 1995, 25–30. [Google Scholar]
- Sánchez-Moreno C, Larrauri JA, Saura-Calixto F. A procedure to measure the antiradical efficiency of polyphenols. J Sci Food Agric 1998, 76, 270–6. [Google Scholar] [CrossRef]
- Siddhraju P, Becker K. Antioxidant properties of various solvents extracts of total phenolic constituents from three different agroclimatic origins of drumstick tree (Moringa oleifera Lam) leaves. J Agric Food Chem 2003, 2144–55.
- Benzie IFF, Strain JJ. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal Biochem 1996, 239, 70–6. [Google Scholar] [CrossRef]
- ADVID. Boletim ano vitícola 2019; ADVID. Vila Real, 2019. [Google Scholar]
- ADVID. Boletim Ano Vitícola 2020; ADVID. Vila Real, 2020. [Google Scholar]
- Patočka J, Navrátilová Z, Ovando M. Biologically active compounds of Knotweed (Reynoutria spp.). Mil Med Sci Lett 2017, 86, 17–31. [Google Scholar] [CrossRef]
- Suteu D, Rusu L, Zaharia C, Badeanu M, Daraban G. Challenge of utilization vegetal extracts as natural plant protection products. Appl Sci 2020, 10, 8913. [Google Scholar] [CrossRef]
- Cucu A-A, Baci G-M, Dezsi Ş, Nap M-E, Beteg FI, Bonta V, et al. New approaches on Japanese knotweed (Fallopia japonica) bioactive compounds and their potential of pharmacological and beekeeping activities: challenges and future Directions. Plants 2021, 10, 2621. [Google Scholar] [CrossRef]
- Devkota HP, Paudel KR, Khanal S, Baral A, Panth N, Adhikari-Devkota A, et al. Stinging Nettle (Urtica dioica L.): nutritional composition, bioactive compounds, and food functional properties. Molecules 2022, 27, 5219. [Google Scholar] [CrossRef]
- Young AJ, Britton G. Carotenoids and oxidative stress; Curr. Res. Photosynth., Dordrecht: Springer Netherlands, 1990; pp. 3381–4. [Google Scholar] [CrossRef]
- Huang H, Ullah F, Zhou D-X, Yi M, Zhao Y. Mechanisms of ROS regulation of plant development and stress responses. Front Plant Sci 2019, 10, 1–10. [Google Scholar] [CrossRef]
- Mona, MA. The potential of Moringa oleifera extract as a biostimulant in enhancing the growth, biochemical and hormonal contents in rocket (Eruca vesicaria subsp. sativa) plants. Int J Plant Physiol Biochem 2013, 5, 42–9. [Google Scholar] [CrossRef]
- Rahman M, Mukta JA, Sabir AA, Gupta DR, Mohi-Ud-Din M, Hasanuzzaman M, et al. Chitosan biopolymer promotes yield and stimulates accumulation of antioxidants in strawberry fruit. PLoS One 2018, 13, 1–14. [Google Scholar] [CrossRef]
- Flamini R, Mattivi F, De Rosso M, Arapitsas P, Bavaresco L. Advanced knowledge of three important classes of grape phenolics: Anthocyanins, stilbenes and flavonols. Int J Mol Sci 2013, 14, 19651–69. [Google Scholar] [CrossRef]
- Singh RK, Soares B, Goufo P, Castro I, Cosme F. , Pinto-Sintra AL, et al. Chitosan upregulates the genes of the ROS pathway and enhances the antioxidant potential of grape (Vitis vinifera L. ‘Touriga Franca’ and ’Tinto Cão’) tissues. Antioxidants 2019, 8, 525. [Google Scholar] [CrossRef]
- Irani H, ValizadehKaji B, Naeini MR. Biostimulant-induced drought tolerance in grapevine is associated with physiological and biochemical changes. Chem Biol Technol Agric 2021, 8, 1–13. [Google Scholar] [CrossRef]
- Ferrandino A, Lovisolo C. Abiotic stress effects on grapevine (Vitis vinifera L.): Focus on abscisic acid-mediated consequences on secondary metabolism and berry quality. Environ Exp Bot 2014, 103, 138–47. [Google Scholar] [CrossRef]
- Kok, D. Grape growth, anthocyanin and phenolic compounds content of early ripening cv. Cardinal table grape (V. vinifera L.) as affected by various doses of foliar biostimulant applications with gibberellic acid. Erwerbs-Obstbau 2018, 60, 253–9. [Google Scholar] [CrossRef]
- Soppelsa S, Kelderer M, Casera C, Bassi M, Robatscher P, Matteazzi A, et al. Foliar applications of biostimulants promote growth, yield and fruit quality of strawberry plants grown under nutrient limitation. Agronomy 2019, 9, 1–22. [Google Scholar] [CrossRef]
- Macheix JJ, Fleuriet A, Billot J. Fruit phenolic; CRC: Press, Boca Raton. USA., 1990. [Google Scholar]
- Ryan D, Antolovich M, Prenzler P, Robards K, Lavee S. Biotransformations of phenolic compounds in Olea europaea L. Sci Hortic (Amsterdam) 2002, 92, 147–76. [Google Scholar] [CrossRef]
- Frioni T, Tombesi S, Quaglia M, Calderini O, Moretti C, Poni S, et al. Metabolic and transcriptional changes associated with the use of Ascophyllum nodosum extracts as tools to improve the quality of wine grapes (Vitis vinifera cv. Sangiovese) and their tolerance to biotic stress. J Sci Food Agric 2019, 99, 6350–63. [CrossRef]
- Braidot E, Zancani M, Petrussa E, Peresson C, Bertolini A, Patui S, et al. Transport and accumulation of flavonoids in grapevine (Vitis vinifera L.). Plant Signal Behav 2008, 3, 626–32. [CrossRef]
- Fraga H, Malheiro AC, Moutinho-Pereira J, Santos JA. An overview of climate change impacts on European viticulture. Food Energy Secur 2012, 1, 94–110. [Google Scholar] [CrossRef]
- Blanco-Ward D, Ribeiro A, Barreales D, Castro J, Verdial J, Feliciano M, et al. Climate change potential effects on grapevine bioclimatic indices: A case study for the Portuguese demarcated Douro Region (Portugal). BIO Web Conf 41st World Congr Vine Wine 2019, 12, 01013. [Google Scholar] [CrossRef]
- Drobek M, Frąc M, Cybulska J. Plant biostimulants: Importance of the quality and yield of horticultural crops and the improvement of plant tolerance to abiotic stress - A review. Agronomy 2019, 9. [Google Scholar] [CrossRef]
- Parađiković N, Teklić T, Zeljković S, Lisjak M, Špoljarević M. Biostimulants research in some horticultural plant species—A review. Food Energy Secur 2018, 8, 1–17. [Google Scholar] [CrossRef]
- Xu DP, Li Y, Meng X, Zhou T, Zhou Y, Zheng J, et al. Natural antioxidants in foods and medicinal plants: Extraction, assessment and resources. Int J Mol Sci 2017, 18, 20–31. [Google Scholar] [CrossRef]
- Rockenbach II, Rodrigues E, Gonzaga LV, Caliari V, Genovese MI, Gonalves AEDSS, et al. Phenolic compounds content and antioxidant activity in pomace from selected red grapes (Vitis vinifera L. and Vitis labrusca L.) widely produced in Brazil. Food Chem 2011, 127, 174–9. [Google Scholar] [CrossRef]
- Kupe M, Karatas N, Unal MS, Ercisli S, Baron M, Sochor J. Phenolic composition and antioxidant activity of peel, pulp and seed extracts of different clones of the Turkish grape cultivar ‘Karaerik.’. Plants 2021, 10, 2154. [CrossRef]
- Speranza S, Knechtl R, Witlaczil R, Schönlechner R. Reversed-phase HPLC characterization and quantification and antioxidant capacity of the phenolic acids and flavonoids extracted from eight varieties of sorghum grown in Austria. Front Plant Sci 2021, 12, 1–14. [Google Scholar] [CrossRef]
Figure 1.
Monthly average values of temperature (A) and precipitation (B) in the experimental vineyard during 2019 and 2020.
Figure 1.
Monthly average values of temperature (A) and precipitation (B) in the experimental vineyard during 2019 and 2020.
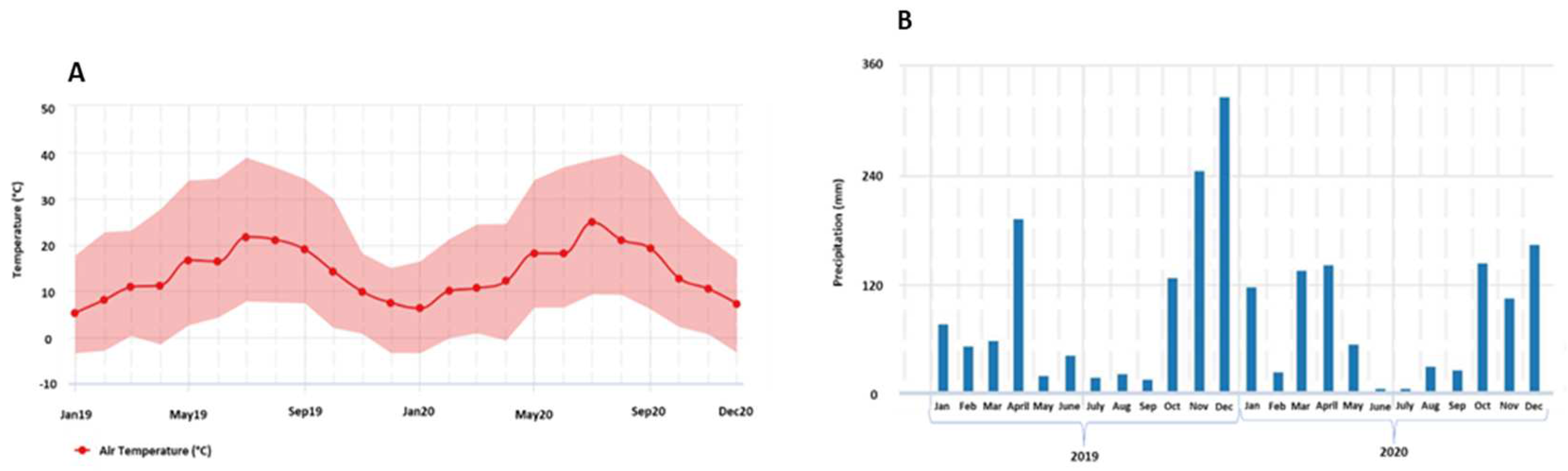
Figure 2.
Downy mildew incidence on leaves (A) and berries (B) and severity on leaves (C) and berries (D) of the cv. Touriga Franca during the season 2020. Values are presented as mean ± SE; different letters mean significant differences (P < 0.05, Duncan’s test) between treatments, within each phenological stage (numbers in letters refer to each phenological stage). CT – Conventional Treatment; JKE – Japanese Knotweed Extract; NE – Nettle Extract; C – Control.
Figure 2.
Downy mildew incidence on leaves (A) and berries (B) and severity on leaves (C) and berries (D) of the cv. Touriga Franca during the season 2020. Values are presented as mean ± SE; different letters mean significant differences (P < 0.05, Duncan’s test) between treatments, within each phenological stage (numbers in letters refer to each phenological stage). CT – Conventional Treatment; JKE – Japanese Knotweed Extract; NE – Nettle Extract; C – Control.
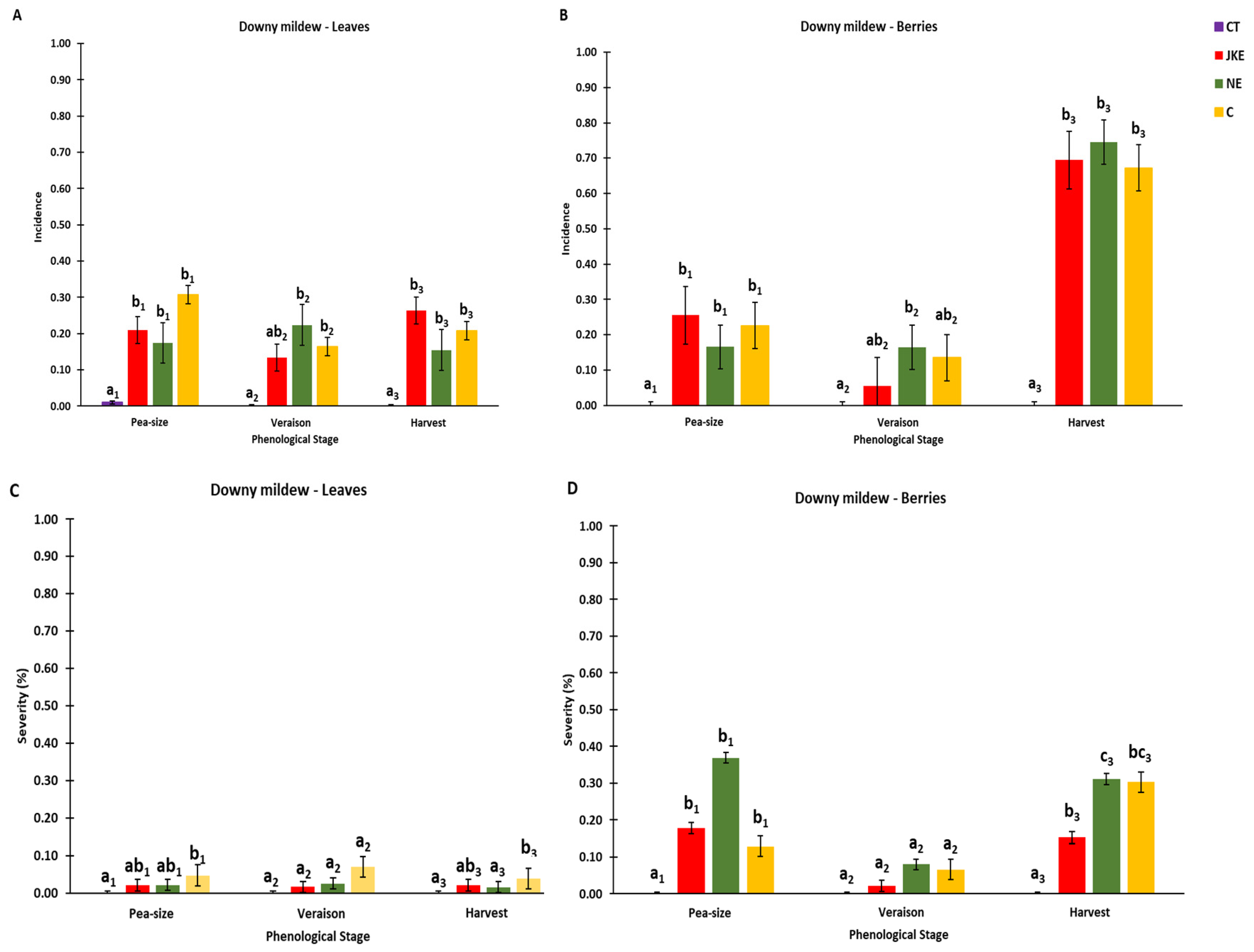
Figure 3.
Total carotenoids concentration content in leaves (A) (µg.g-1 FW) and berries (B) (µg.g-1 DW) of cv. Touriga Franca with different foliar treatments at veraison and harvest of 2019 and 2020. Values are presented as mean ± SE. Different letters mean significant differences (P < 0.05, Duncan’s test) between treatments within each phenological stage: veraison or harvest of each year (lowercase, year: 1 - 2019; 2 - 2020); “ns” indicates non-significant differences. C – Control, CT – Conventional Treatment; JKE – Japanese Knotweed Extract; NE – Nettle Extract.
Figure 3.
Total carotenoids concentration content in leaves (A) (µg.g-1 FW) and berries (B) (µg.g-1 DW) of cv. Touriga Franca with different foliar treatments at veraison and harvest of 2019 and 2020. Values are presented as mean ± SE. Different letters mean significant differences (P < 0.05, Duncan’s test) between treatments within each phenological stage: veraison or harvest of each year (lowercase, year: 1 - 2019; 2 - 2020); “ns” indicates non-significant differences. C – Control, CT – Conventional Treatment; JKE – Japanese Knotweed Extract; NE – Nettle Extract.
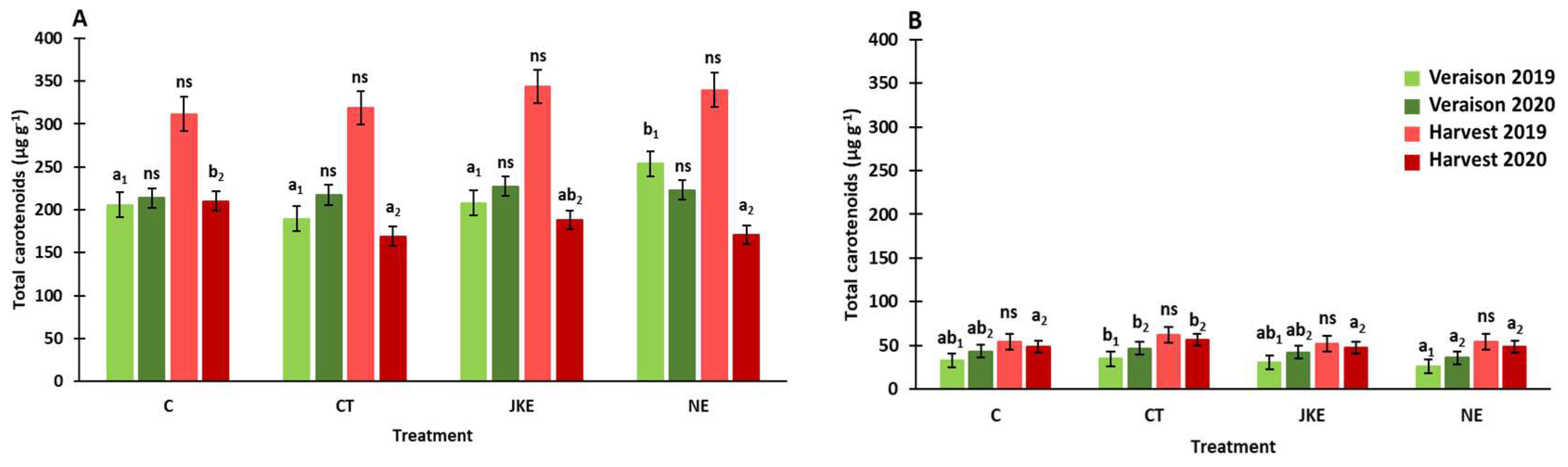

Table 1.
Total phenolic, anthocyanin and flavonoid concentrations in leaves and berries of cv. Touriga Franca with different foliar treatments, at veraison and harvest of 2019 and 2020. C – Control; CT – Conventional Treatment; JKE – Japanese Knotweed Extract; NE – Nettle Extract. Values are presented as mean ± SE. Different letters mean significant differences (P < 0.05, Duncan’s test) between treatments within each phenological stage: veraison or harvest of each year (lowercase, year: 1 - 2019; 2 - 2020); no letters indicate non-significant differences.
Table 1.
Total phenolic, anthocyanin and flavonoid concentrations in leaves and berries of cv. Touriga Franca with different foliar treatments, at veraison and harvest of 2019 and 2020. C – Control; CT – Conventional Treatment; JKE – Japanese Knotweed Extract; NE – Nettle Extract. Values are presented as mean ± SE. Different letters mean significant differences (P < 0.05, Duncan’s test) between treatments within each phenological stage: veraison or harvest of each year (lowercase, year: 1 - 2019; 2 - 2020); no letters indicate non-significant differences.
| Bioactive Compound | Plant organ | Growth stage | C | CT | JKE | NE |
|---|---|---|---|---|---|---|
|
Total Phenolics (mg.g-1) |
Leaf | Veraison 2019 | 13.33 ± 1.35 | 13.51 ± 1.32 | 11.25 ± 1.01 | 11.02 ± 1.06 |
| Veraison 2020 | 22.19 ± 1.69a2 | 27.72 ± 0.94b2 | 28.09 ± 2.14b2 | 27.33 ± 1.64b2 | ||
| Harvest 2019 | 11.36 ± 0.57 | 14.86 ± 1.31 | 12.90 ± 1.30 | 12.97 ± 1.20 | ||
| Harvest 2020 | 23.70 ± 0.85 | 26.67 ± 1.28 | 26.91 ± 1.02 | 24.14 ± 1.29 | ||
| Berry | Veraison 2019 | 18.34 ± 3.16ab1 | 14.14 ± 2.15a1 | 17.86 ± 1.73ab1 | 26.31 ± 4.41b1 | |
| Veraison 2020 | 50.38 ± 1.32 | 50.60 ± 4.82 | 63.58 ± 5.35 | 59.33 ± 4.81 | ||
| Harvest 2019 | 16.99 ± 1.52 | 16.87 ± 2.11 | 13.61 ± 1.20 | 17.58 ± 1.94 | ||
| Harvest 2020 | 31.74 ± 4.15 | 32.54 ± 2.82 | 32.88 ± 3.55 | 28.30 ± 1.81 | ||
| Total Anthocyanins (mg.g-1) | Berry | Veraison 2019 | 1.87 ± 0.58 | 6.39 ± 0.33 | 3.01 ± 2.42 | 2.86 ± 1.12 |
| Veraison 2020 | 3.26 ± 0.50 | 4.00 ± 1.57 | 1.27 ± 0.28 | 3.34 ± 0.70 | ||
| Harvest 2019 | 2.20 ± 0.33a1 | 6.97 ± 1.30b1 | 2.46 ± 0.85a1 | 3.05 ± 1.88ab1 | ||
| Harvest 2020 | 6.94 ± 0.94 | 9.95 ± 2.33 | 7.05 ± 1.48 | 6.39 ± 0.47 | ||
|
Flavonoids (mg.g-1) |
Leaf | Veraison 2019 | 2.54 ± 0.21a1 | 3.63 ± 0.25b1 | 2.54 ± 0.19a1 | 2.70 ± 0.17a1 |
| Veraison 2020 | 3.59 ± 0.50a2 | 5.54 ± 0.70b2 | 4.27 ± 0.57ab2 | 4.05 ± 0.42ab2 | ||
| Harvest 2019 | 2.85 ± 0.37a1 | 4.11 ± 0.35b1 | 3.22 ± 0.32ab1 | 3.51 ± 0.54ab1 | ||
| Harvest 2020 | 4.19 ± 0.22a2 | 5.35 ± 0.19b2 | 5.54 ± 0.34b2 | 4.30 ± 0.32a2 | ||
| Berry | Veraison 2019 | 4.60 ± 0.62a1 | 4.33 ± 0.80a1 | 7.19 ± 0.44b1 | 5.67± 0.86ab1 | |
| Veraison 2020 | 18.32 ± 2.31 | 14.27 ± 1.38 | 18.90 ± 2.57 | 15.72 ± 1.64 | ||
| Harvest 2019 | 6.59 ± 1.86 | 4.03 ± 0.85 | 6.17 ± 3.73 | 5.43 ± 0.85 | ||
| Harvest 2020 | 8.51 ± 0.99 | 9.33 ± 1.45 | 9.61 ± 0.99 | 9.03 ± 1.97 |
Table 2.
Antioxidant activity in leaves and berries, using three different assays (ABTS•+, DPPH, and FRAP), in cv. Touriga Franca with different foliar treatments, at veraison and harvest of 2019 and 2020. C – Control; CT – Conventional Treatment; JKE – Japanese Knotweed Extract; NE – Nettle Extract. Values are presented as mean ± SE. Different letters mean significant differences (P < 0.05, Duncan’s test) between treatments within each phenological stage: veraison or harvest of each year (lowercase, year: 1 - 2019; 2 - 2020); no letters indicate non-significant differences.
Table 2.
Antioxidant activity in leaves and berries, using three different assays (ABTS•+, DPPH, and FRAP), in cv. Touriga Franca with different foliar treatments, at veraison and harvest of 2019 and 2020. C – Control; CT – Conventional Treatment; JKE – Japanese Knotweed Extract; NE – Nettle Extract. Values are presented as mean ± SE. Different letters mean significant differences (P < 0.05, Duncan’s test) between treatments within each phenological stage: veraison or harvest of each year (lowercase, year: 1 - 2019; 2 - 2020); no letters indicate non-significant differences.
| Antioxidant activity assay | Plant organ | Growth stage | C | CT | JKE | NE |
|---|---|---|---|---|---|---|
|
ABTS•+ (µmol Trolox µg-1) |
Leaf | Veraison 2019 | 0.46 ± 0.03b1 | 0.31 ± 0.02a1 | 0.41 ± 0.03b1 | 0.45 ± 0.04b1 |
| Veraison 2020 | 0.31 ± 0.02b2 | 0.13 ± 0.02a2 | 0.19 ± 0.04a2 | 0.21 ± 0.03a2 | ||
| Harvest 2019 | 0.47 ± 0.04b1 | 0.27 ± 0.04a1 | 0.40 ± 0.04b1 | 0.41 ± 0.03b1 | ||
| Harvest 2020 | 0.17 ± 0.02b2 | 0.12 ± 0.03ab2 | 0.08 ± 0.02a2 | 0.15 ± 0.03ab2 | ||
| Berry | Veraison 2019 | 65.19 ± 9.71ab1 | 84.40 ± 4.60b1 | 77.18 ± 8.39ab1 | 56.32 ± 8.76a1 | |
| Veraison 2020 | 59.36 ± 6.79b2 | 56.48 ± 11.51b2 | 38.29 ± 9.73ab2 | 27.89 ± 7.03a2 | ||
| Harvest 2019 | 79.01 ± 7.16 | 78.62 ± 5.49 | 73.49 ± 7.49 | 80.59 ± 6.51 | ||
| Harvest 2020 | 63.08 ± 4.72 | 75.81 ± 9.30 | 59.71 ± 7.78 | 54.82 ± 9.96 | ||
|
DPPH (µmol Trolox µg-1) |
Leaf | Veraison 2019 | 0.79 ± 0.05 | 0.73 ± 0.03 | 0.79 ± 0.05 | 0.80 ± 0.04 |
| Veraison 2020 | 1.09 ± 0.04 | 1.03 ± 0.04 | 1.08 ± 0.07 | 1.02 ± 0.05 | ||
| Harvest 2019 | 0.74 ± 0.04b1 | 0.62 ± 0.03a1 | 0.70 ± 0.03ab1 | 0.69 ± 0.03ab1 | ||
| Harvest 2020 | 1.24 ± 0.05 | 1.28 ± 0.03 | 1.22 ± 0.03 | 1.24 ± 0.04 | ||
| Berry | Veraison 2019 | 59.74 ± 2.52ab1 | 69.53 ± 2.44b1 | 66.07 ± 4.52b1 | 54.61 ± 3.12a1 | |
| Veraison 2020 | 49.16 ± 2.14 | 49.42 ± 2.13 | 45.14 ± 2.40 | 47.02 ± 2.87 | ||
| Harvest 2019 | 65.48 ± 1.89 | 66.34 ± 3.58 | 68.23 ± 2.54 | 70.52 ± 1.73 | ||
| Harvest 2020 | 57.57± 1.49 | 58.99 ± 3.98 | 53.04 ± 2.94 | 61.74 ± 5.64 | ||
|
FRAP (µmol Trolox µg-1) |
Leaf | Veraison 2019 | 0.70 ± 0.07a1 | 0.92 ± 0.05b1 | 0.74 ± 0.06a1 | 0.70 ± 0.06a1 |
| Veraison 2020 | 0.38 ± 0.03 | 0.53 ± 0.05 | 0.54 ± 0.07 | 0.48 ± 0.06 | ||
| Harvest 2019 | 0.58 ± 0.03a1 | 0.97 ± 0.11b1 | 0.66 ± 0.06a1 | 0.62 ± 0.05a1 | ||
| Harvest 2020 | 0.48 ± 0.04 | 0.56 ± 0.05 | 0.58 ± 0.04 | 0.50 ± 0.04 | ||
| Berry | Veraison 2019 | 33.82 ± 4.80b1 | 19.22 ± 1.05a1 | 24.87 ± 2.75ab1 | 34.58 ± 4.15b1 | |
| Veraison 2020 | 55.40 ± 3.30a2 | 64.68 ± 4.25ab2 | 66.21 ± 1.66b2 | 69.77 ± 3.99b2 | ||
| Harvest 2019 | 23.70 ± 1.18ab1 | 20.17 ± 1.97a1 | 19.29 ± 2.50a1 | 25.86 ± 1.27b1 | ||
| Harvest 2020 | 43.10 ± 2.19ab2 | 43.82 ± 2.80b2 | 42.55 ± 3.31ab2 | 35.32 ± 1.31a2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



