
Submitted:
07 August 2023
Posted:
07 August 2023
You are already at the latest version
Abstract
Pancreatic ductal adenocarcinoma (PDAC) is a ravaging disease whose poor prognosis requires a more detailed understanding of its biology to foster the development of effective therapies. The unsatisfactory results of treatments targeting cell proliferation and its related mechanisms suggested to rather focus on the inflammatory tumor microenvironment (TME). Here, we discuss the role of cancer secreted proteins in the complex TME tumor-stroma crosstalk, to sched lights on druggable molecular targets for the development of innovative, safer and more efficient therapeutic strategies.
Keywords:
Pancreatic Ductal Adenocarcinoma
; secretome
; cell signaling
; tumor microenvironment
; small molecules
; monoclonal antibodies.
1. Pancreatic cancer Fact Sheet
The most common type of pancreatic cancer is the Pancreatic Ductal Adenocarcinoma (PDAC), which arises from the ductal epithelium of the organ. Around 70% of pancreatic cancers begin in the organ’s head, and the majority start from the ducts that transport digestive enzymes (pancreatic ductal adenocarcinoma) [1,2]. The prevalence rate is 49.8 cases per million people, while the predicted global incidence rate is 58.6 cases per million people annually. Annual mortality is projected to be 57.7 per million persons. In the past 25 years, there has been a 55%, 63% and 53% increase in incidence, prevalence and death, respectively. While representing 1.8% of all malignancies, pancreatic cancer is responsible for 4.6% of cancer-related fatalities. Men have a somewhat greater incidence, prevalence, and fatality rate. By 2060, it is anticipated that pancreatic cancer deaths would have increased almost 1.97-fold.
Despite recent advances in surgical techniques and medical therapies, the median survival time for a pancreatic cancer patient at diagnosis is 4-6 months, with an 8% 5-year survival rate [3,4]. Pancreatic cancer is predicted to overtake colorectal, breast, and prostate cancer as the second leading cause of cancer death in the United States by 2030, surpassing colorectal, breast, and prostate cancer. In 2040, gastrointestinal cancers (pancreatic, liver, and colorectal cancer) are expected to be three of the four leading causes of cancer death. [3].
The main reasons for such a poor prognosis can be found in the particularly complex anatomical region in which the tumor grows, as well as the fact that this tumor is usually diagnosed at an advanced stage in most patients. In fact, the survival curve of patients falls dramatically in the first year of follow-up from the moment of diagnosis; for this neoplasm, there have been no significant improvements in the last three decades, and the poor prognosis is substantially uniform at the international level [5,6].
Pancreatic cancer is a complex malignant tumor with a poor prognosis because it does not exhibit specific symptoms at an early stage, and even when they do appear, they are rather vague disorders that can be misinterpreted. For these reasons, the diagnosis is frequently made after the disease has spread. The only potentially curative treatment is surgical removal, but radical surgical resection of the tumor is indicated only in cases of intrapancreatic disease that does not extend to the retroperitoneum or transverse mesocolon and does not involve infiltration of the superior mesenteric artery, celiac tripod, or spleno-mesenteric-portal axis. For this reason, pancreatic cancer is only resectable in 10-20% of patients at the time of diagnosis, while it is locally advanced in 30-35% of patients due to infiltration of the large abdominal vessels, and metastases are already present in more than 50% of cases. However, even in the early stages, the prognosis is poor, as the median survival rates with surgery alone in this group of patients are in the order of 12 months, and the 5-year survival rate is between 5 and 10%. Median survival for stages III and IV is 10 and 6 months, respectively [7,8].
Differently from metastatic diseases in other tumor types (e.g. colon [9], gastroesophageal [10] head and neck [11] breast [12], lung [13] cancers) first-line treatment for metastatic pancreatic cancer is still chemotherapy [14]. Indeed, even though immunotherapies and/or targeted therapies for the majority of solid tumors have advanced quickly, progress in the treatment of pancreatic ductal adenocarcinoma has been unusually slow [15,16]. In this regard, researchers recently discovered novel players within the TME that can help improve therapeutic actions for various cancer therapies. A special emphasis here, has been placed on pancreatic cancer- secreted proteins, which are the primary source in developing and maintaining a cancer-friendly environment.
2. Proteins secreted by pancreatic cancer cells: messages sent to the neighborhood.
Cell communication in multicellular organisms allows cells to adapt their phenotypes and function. A number of secreted factors, soluble or associated to membranes, mediates critical molecular mechanisms involved in tissue and organism homeostasis. Proteins typically follow the conventional protein secretion pathway, which involves the endoplasmic reticulum (ER) and the Golgi complex. However, some proteins use alternative routes, such as unconventional protein secretion (UPS) pathways, induced by cellular stress such as nutrient deficiency, mechanical stress, inflammation, and ER stress. When the pathways leading to protein secretion - which mediate both short- and long-range signals - are dysregulated, disease pathogenesis is accelerated. In this context it is known that transformed cells are the source of distinct extracellular signals [17] that are captured by neighboring non-transformed stromal cells, influencing tumor development, metastasis, and even drug efficacy [18]. Moreover, tumoral secretomes provide a promising source of potential biomarkers useful in-patient management, and, in some cancers, there is a growing interest in intracellular proteins that have distinct and different functions when secreted, demonstrating how UPS pathways are still not fully understood [19]. In pancreatic cancer, mutated epithelial, acinar or ductal cells account for a small portion of the tumor mass, with the remaining made up of stromal cells and components. This reflects the significance of cell communication being hampered by pancreatic cancer cells, which collaborate in shaping a fibrotic and inflammatory microenvironment that promotes tumor establishment and progression [20].
Here, a literature search was conducted to find a comprehensive dataset that could illustrate the main biochemical pathway in which pancreatic cancer cells direct such a dramatic tissue transformation. We used PubMed to look for PDAC secretome studies. To maximize search specificity and sensitivity, the following keywords were used: “Pancreatic cancer,” “Pancreatic ductal adenocarcinoma,” and “secretome, extracellular protein and pancreatic carcinoma” and “tumor microenvironment”. Only studies involving pancreatic cancer- secreted proteins were included after further screening based on title and abstract. There were no restrictions on the type of study. Our studies primarily employ two methods for secretome analysis of pancreatic adenocarcinoma: mass spectrometry and the Enzyme-Linked ImmunoSorbent Assay (ELISA). Most mass spectrometry-based studies include secretome or proteome analysis of human [21,22,23,24,25,26], murine [27] or hamster [28] cell lines. On the other hand, most studies using ELISA analyzed serum from patients with hereditary predisposition [29] as well as advanced disease [30,31], and pancreatic cancer cell supernatant [22]. In addition, we included a recent review [32] resuming the use of novel protein biomarkers in early PDAC diagnosis, prognosis, and treatment response prediction, as well as the utility of possible biomarker combinations in diagnostic panels. Finally, an approach integrating meta-analysis of PDAC proteome and secretome MS data, used to identify potential disease biomarkers [33], was further considered (Table 1).
The obtained dataset represented and summarized the coexistence of cytokines, growth factors, extracellular matrix proteins, proteases and protease inhibitors, membrane- and extracellular vesicle- associated proteins, and metabolic enzymes in the neoplastic milieu (Figure 1).
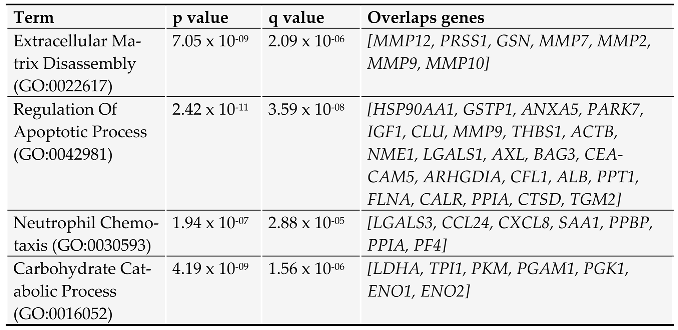
To define protein clusters associated with main cellular processes, the dataset was analyzed in the GO biological processes 2023 database by using Enrichr (https://appyters.maayanlab.cloud/ accessed in June 2023) (Table 2). Proteins with a known function when secreted are then briefly discussed.
As a result of the analysis, it was possible to put in evidence a set of proteases responsible for extracellular matrix and cellular component disassembly. As previously reported, the acellular components of the pancreatic tumor mass, as well as their changes over time, drive the tumor’s progression. In this regard, MMP-2 and MMP-9 (Matrix MetalloProteinases) gelatinases are abnormally and contemporarily up-regulated in pancreatic cancer [31,34], but the clinical relevance measured by the correlation between their expression and survival, metastasis, or tumor stage is debatable [35]. Instead, the expression of the matrilysin MMP-7 in tumor samples was linked to a poorer prognosis in patients [36] and an unfavorable pathologic response to neoadjuvant therapy [37]. Furthermore, the multifunctional zinc finger transcription factor YY1 (Yin Yang 1) has been shown to suppress stromelysin-2 (MMP-10), whose overexpression is a favorable independent prognostic factor in PDAC patients [38]. Finally, the secreted gelsolin, a scavenger of extracellular actin, has been recently reported for its involvement in attenuating DNGR-1-dependent dendritic cell-mediated anti-tumor immunity [39].
Some apoptosis-regulating proteins with very different biochemical functions have been also identified, and some of them have been linked to a role in pancreatic cancer. E.g., Hsp90AA1 (Heat Shock Protein 90 Alpha Family Class A Member 1) is one of the most abundant proteins expressed in the cells, which interacts with several secreted client proteins. Hsp90AA1 promotes tumor aggressiveness and chemoresistance by activating AKT through LRP-1 (Low-density lipoprotein Receptor-related Protein 1) [40]. Among other chaperones, PARK7 (Parkinson protein 7) [41] and PPIA (Peptidylprolyl Isomerase A) [42] are upregulated and secreted by cancer cells. While PARK7 has been described for its ability to counteract environmental oxidative stress [41], PPIA is known to act through CD47 and NF-kB pathway, thus promoting cell proliferation [43]. In addition, the extracellular chaperone Clusterin (CLU) has been shown to be a mediator of chemoresistance in pancreatic cancer [44]. The overexpression of the co-chaperone BAG3 (BCL2 Associated Athanogene 3) has also been described to be associated with pancreatic cancer aggressiveness [45], and its sera levels are measurable in pancreatic cancer patients [31,46]. Furthermore, a distinct function was described for secreted BAG3. Indeed, its interaction with IFITM2 (Interferon Induced Transmembrane Protein 2) on the plasma membrane can induce pro-tumoral cytokine release by macrophages [47] and fibroblasts [48], thus sustaining tumor growth. Another relevant protein in this context is the soluble GSTP1 (Glutathione S-Transferase Pi 1) able to catalyze the conjugation of many hydrophobic and electrophilic compounds with reduced glutathione. This activity plays a role in regulating oxidative stress, thus negatively affecting proliferation and upregulating apoptosis in PDAC cells, as demonstrated by knockdown experiments [49]. The transglutaminase-2 gene (TGM2) has been also identified as an important extracellular cross-linking enzyme involved in ECM turnover, and its levels were associated to a poor patient survival in pancreatic cancer patients [50]. Annexins dysregulation has also been reported as a common feature in several cancers [51], and, among them, soluble ANXA5 (Annexin A5) has been described for its activity in cell membrane resealing [52], an essential repair machinery which promotes survival in invasive cancer cells [53]. Insulin-like growth factor (IGF)-axis, which mediates survival signals essential for pancreatic cancer development and progression, belongs to traditional signaling molecules, and its crucial role was evidenced by the strong association between SNPs (Single Nucleotide Polymorphisms) in correlated IGF1R, IGF2R, and IRS1 genes with tumor response to therapy and disease stage [54]. Finally, AXL (a member of the TAM tyrosine kinase receptors) is present in our dataset. The AXL/Gas6 (Growth Arrest Specific 6) signaling pathway plays a role in tumor cell proliferation, and plasma detection of the AXL soluble form, obtained by an ADAM10/ADAM17 (ADAM Metallopeptidase Domain)-specific shedding mechanism, has been described as an early signature of PDAC [55], while its functions, likely relying on attenuating Gas6 functions in the TME, remain unknown [56].
An additional functional cluster of secreted proteins were linked to neutrophil chemotaxis. Tumor-infiltrating neutrophils indicate poor prognosis for patients and activated neutrophils can generate neutrophil extracellular traps (NETs) which are emerging in several cancers as a marker of cancer progression and immunosuppression [57,58]. As a first example, extracellular Galectin-3 (LGALS3), detected in the blood of PDAC patients [58], has been associated with neutrophils recruitment and inflammation exacerbation in several infectious diseases [59]. Indeed, given the similarities in the pathogenesis of inflammatory diseases and cancer, it is not surprising to find in the described pancreatic cancer- secreted proteins dataset molecules such as IL8 (CXCL8) [61], Eotaxin-2 (CCL24) [62], PPBP (Pro-Platelet Basic Protein) [63], and PF4 (Platelet Factor 4) [64] which are potent neutrophil chemotactic factors. Serum Amyloid A1 (SAA1) [65], another protein induced in pancreatic cancer cells, attracts neutrophils to the tumor nest by interacting with TLR2 (Toll-Like Receptor 2) [66].
Finally, altered carbohydrate catabolism has been recognized as the major metabolic alteration in pancreatic cancer [67], but the role of those enzymes in the pancreatic cancer milieu has not been fully elucidated yet. The evidence that secretory PKM (Pyruvate Kinase M1/2) promotes tumor angiogenesis by facilitating endothelial cell proliferation and new vessel formation via the PI3K/AKT and Wnt/-catenin signaling pathways provides some hints [68]. On the other hand, secreted PGK1 (Phosphoglycerate Kinase 1) has been shown to act on angiostatin levels, resulting in an anti-angiogenic and tumor suppressive function [69]. Finally, enolases, specifically the cell surface associated ENO1, have been identified as a pancreatic cancer neoantigen, promoting cancer cell survival and migration by coordinating with integrins and uPAR (Plasminogen Activator, Urokinase Receptor) [70].
Other proteins relevant in this context, but not included in Table 2, are: GDF15 (Growth Differentiation Factor 15), whose expression was positively correlated with poor survival in PDAC patients and whose downregulation inhibited PDAC tumor growth in vivo [71]; LIF (LIF Interleukin 6 Family Cytokine), a pleiotropic cytokine that regulates cell survival by interacting with its receptor LIFR/GP130 expressed on surrounding fibroblasts, thus promoting pro invasive phenotype [72]; VEGF (Vascular Endothelial Growth Factor) [73], which functions as an endothelial cell mitogen and is strongly linked to angiogenesis in pancreatic cancer; and TF (Tissue Factor), which, in its alternatively spliced form asTF, is released by cancer cells thereby triggering the activation of PI3K/Akt, MAPK, and FAK pathways through its interactions with α6β1 and αvβ3 integrins [74].
The list of proteins resulting from the analysis was used to screen the presence of pharmacologic molecules designed to interfere with their activity, to describe available candidates for therapeutic interventions in pancreatic cancer.
3. Communications breakdown operated by small drugs.
The DGIdb (The Drug Gene Interaction Database, accessed in June 2023) was queried, screened and integrated with literature search for available molecules possibly having inhibitory activity on pancreatic cancer- secreted proteins illustrated above; obtained search results are thereby described.
The response to synthetic inhibitors of MMPs (MMPIs) were studied in the past decades in several solid tumors. However, despite the promising preclinical data, all trials were unsuccessful in tumor mass reduction or overall survival improvement [75].
Clusterin expression was challenged using the drug OGX-011, an antisense oligonucleotide that showed a potentiating effect of various FDA-approved anticancer chemotherapeutics during clinical trials [76]; any trial result for pancreatic cancer is still available [77].
Ganetespib (STA-9090) is a small molecule that interferes with HSP90 client protein binding, thus promoting the inactivation and degradation of the signaling proteins that regulate cancer progression. Unfortunately, a Phase II study carried out in refractory metastatic pancreatic cancer failed to prove its clinical efficacy [78]. More clinical trials as a neoadjuvant treatment and/or in combination with chemotherapy are expected [79].
The extracellular galectin-3 functions were shown to be targeted by RN1, an arabinogalactan polysaccharide isolated from the flowers of the Chinese ginseng plant (Panax notoginseng) which displayed antitumoral activity in PDAC in vitro and in vivo [80]. While the use of galectin inhibitors, such as belapectin (GR-MD-02), has been accepted as a novel therapeutic tool in liver antifibrotic therapy [81], clinical trials are ongoing to explore its utility in lung and head and neck cancer diseases (clinical trial: NCT02575404).
4. Communications breakdown operated by monoclonal antibodies.
The target specificity of monoclonal antibodies (mAbs) makes them powerful tools for a wide spectrum of biomedical and clinical application. As previously stated, the use of DGIdb was supported and integrated by a literature search to identify available mAbs able to bind and neutralize the secreted proteins here selected for discussion.
Xentuzumab, an IgG1 monoclonal antibody with high affinity for IGF-1 and IGF-2 currently tested in preclinical models for the treatment of advanced solid tumors, allowed the collection of several interesting data [82,83]. A phase I trial enrolling patients affected by different advanced solid tumors, including PDAC, allowed to verify its safety, tolerability and clinical manageability. On the other hand, in a phase II study in metastatic breast cancer, treatments with the Xentuzumab combined to everolimus and exemestane did not show significant impact on PFS (Progression Free Survival) [84].
Another strategy, aimed at neutralizing VEGF with the monoclonal antibody Bevacizumab, showed promising results in preclinical studies [85], but it did not show appreciable benefit when combined with gemcitabine in clinical trials [86]. Thanks to its high safety profile, trials were further extended to a third compound, erlotinib, but still without satisfying results [87].
Recently LIF, which seems to directly fuel oncogenic KRAS signaling, has been proposed as a therapeutic target in PDAC, thus providing a chance to develop a new tool able to overcome chemotherapy resistance in KRAS- targeted protocols. Indeed, in syngeneic mouse models of oncogenic KrasG12D-driven pancreatic adenocarcinoma, using LIF neutralizing antibodies, or only gemcitabine, had no effect on tumor growth, while the combined treatment was able to repress the tumor growth and improve the animal survival [88]. A recently completed first-in-human trial with a humanized anti-LIF mAb (MSC-1) in advanced solid tumors showed that the mAb was well tolerated and was able to extend the PFS of some patients [89]; a phase II study in PDAC patients is currently underway (D8151C00001).
In this context, the neutralization of extracellular BAG3 is another promising strategy supported by studies carried out in several murine preclinical models treated with an anti-BAG3 mAb, that showed its ability in reducing PDAC tumor growth as monotherapy [47,90]. but even more striking results were observed in combined treatments with the ICIs (Immune Check-point Inhibitors) anti-PD-1 [91] and anti-SIRP-alpha [92]. The ex vivo studies on mice tissues showed that combo treatments were able to promote an immunostimulatory microenvironment along with a significant reduction of cancer fibrosis [91,92,93]. Further refinements and developments of the anti-BAG3 mAb- based therapies could hopefully contribute additional evidence of efficacy and safety of combined treatments and could allow to tailor and diversify the protocols also for other solid tumors [94].
An anti-IL-8 antibody was also tested in a humanized mouse model of PDAC in combination with anti-PD-1. The treatment results in a significantly reduced tumor growth, as well as an increased innate immune response and type I cytokine expression in myeloid cells, that reveals a novel function of IL-8 antibody in myeloid cell “re-education” [95]. HuMax-IL8 was tested in a Phase I trial on solid tumors showing its safety and tolerability, while further studies are ongoing to evaluate the efficacy of anti-IL-8 treatments combined with other immunotherapies [96].
Another around the corner strategic perspective aims at targeting the asTF protein by First-In-Class humanized antibody which exerted a significant effect on tumor growth in an animal model, downregulating several gene function categories including focal adhesion, cell motility, cell proliferation, cytoskeleton, regulatory proteases and cell death, many of which are known to be TF-associated [97]. In this case, XB002, a novel, investigational ADC (Antibody Drug Conjugate), is currently being tested as single-agent and combination therapy in subjects with inoperable locally advanced, or metastatic, solid tumors in a Phase I trial; results are expected in late 2024 (NCT04925284) (Figure 2).
5. Conclusion and Future directions
The abnormal production of secreted factors in malignant cells, via canonical and non-canonical pathways, has long been the key mechanism through which metabolic rewiring of cancer cells and neighboring non-malignant cells directs tumor progression [99]. With a focus on targeting pancreatic cancer- secreted proteins, we tried to summarize current and hopefully promising novel therapeutic approaches for the treatment of pancreatic cancer. The study, carried out by analyzing the data available in the literature, confirms the need for further efforts in selecting new molecular targets with lower toxicity risks along with the design of more appropriate, selective and specific therapeutic tools. Furthermore, a deep profiling of the tumor tissue proteome, as well as circulating proteins, using high-throughput technologies during the pre-treatment stage could help in tailoring personalized therapies, thus favoring the development of more effective therapeutic strategies for this fatal disease.
Funding
This research was funded by Regione Campania, POR CAMPANIA FESR 2014-2020 “SYSTEM INNOVATION FOR CANCER EARLY DIAGNOSIS SICED”.
Conflicts of Interest
AR, MDM, LM and MCT are shareholders of FIBROSYS s.r.l., an academic spin-off developing diagnostic and therapeutic tools related to BAG3 and its receptor. The remaining authors have nothing to disclose.
References
- Khalaf N, El-Serag HB, Abrams HR, Thrift AP. Burden of Pancreatic Cancer: From Epidemiology to Practice. Clinical Gastroenterology and Hepatology. 2021;19(5):876-884. [CrossRef]
- Park W, Chawla A, O’Reilly EM. Pancreatic Cancer. JAMA. 2021;326(9):851. [CrossRef]
- Lippi G, Mattiuzzi C. The global burden of pancreatic cancer. Archives of Medical Science. 2020;16(4):820-824. [CrossRef]
- Siegel RL, Miller KD, Wagle NS, Jemal A. Cancer statistics, 2023. CA: A Cancer Journal for Clinicians. 2023;73(1):17-48. [CrossRef]
- Rahib L, Wehner MR, Matrisian LM, Nead KT. Estimated Projection of US Cancer Incidence and Death to 2040. JAMA Network Open. 2021;4(4):e214708. [CrossRef]
- Kindler HL. A Glimmer of Hope for Pancreatic Cancer. New England Journal of Medicine. 2018;379(25):2463-2464. [CrossRef]
- Galon J, Bruni D. Approaches to treat immune hot, altered and cold tumours with combination immunotherapies. Nature Reviews Drug Discovery. 2019;18(3):197-218. [CrossRef]
- Schepis T, De Lucia SS, Pellegrino A; et al. State-of-the-Art and Upcoming Innovations in Pancreatic Cancer Care: A Step Forward to Precision Medicine. Cancers. 2023;15(13):3423. [CrossRef]
- Morris VK, Kennedy EB, Baxter NN; et al. Treatment of Metastatic Colorectal Cancer: ASCO Guideline. Journal of Clinical Oncology. 2023;41(3):678-700. [CrossRef]
- Shah MA, Kennedy EB, Alarcon-Rozas AE; et al. Immunotherapy and Targeted Therapy for Advanced Gastroesophageal Cancer: ASCO Guideline. Journal of Clinical Oncology. 2023;41(7):1470-1491. [CrossRef]
- Moy B, Rumble RB, Come SE; et al. Chemotherapy and Targeted Therapy for Patients With Human Epidermal Growth Factor Receptor 2–Negative Metastatic Breast Cancer That is Either Endocrine-Pretreated or Hormone Receptor–Negative: ASCO Guideline Update. Journal of Clinical Oncology. 2021;39(35):3938-3958. [CrossRef]
- Yilmaz E, Ismaila N, Bauman JE; et al. Immunotherapy and Biomarker Testing in Recurrent and Metastatic Head and Neck Cancers: ASCO Guideline. Journal of Clinical Oncology. 2023;41(5):1132-1146. [CrossRef]
- Mamdani H, Matosevic S, Khalid AB, Durm G, Jalal SI. Immunotherapy in Lung Cancer: Current Landscape and Future Directions. Frontiers in Immunology. 2022;13. [CrossRef]
- Di Costanzo F, Di Costanzo F, Antonuzzo L, Mazza E, Giommoni E. Optimizing First-Line Chemotherapy in Metastatic Pancreatic Cancer: Efficacy of FOLFIRINOX versus Nab-Paclitaxel Plus Gemcitabine. Cancers. 2023;15(2):416. [CrossRef]
- Principe DR, Underwood PW, Korc M, Trevino JG, Munshi HG, Rana A. The Current Treatment Paradigm for Pancreatic Ductal Adenocarcinoma and Barriers to Therapeutic Efficacy. Frontiers in Oncology. 2021;11. [CrossRef]
- Di Federico A, Mosca M, Pagani R; et al. Immunotherapy in Pancreatic Cancer: Why Do We Keep Failing? A Focus on Tumor Immune Microenvironment, Predictive Biomarkers and Treatment Outcomes. Cancers. 2022;14(10):2429. [CrossRef]
- Robinson JL, Feizi A, Uhlén M, Nielsen J. A Systematic Investigation of the Malignant Functions and Diagnostic Potential of the Cancer Secretome. Cell Reports. 2019;26(10):2622-2635.e5. [CrossRef]
- De Morais JA, Zelanis A. Bioinformatic reanalysis of public proteomics data reveals that nuclear proteins are recurrent in cancer secretomes. Traffic. 2021;23(2):98-108. [CrossRef]
- Padgaonkar M, Shendre S, Chatterjee P, Banerjee S. Cancer secretome: Finding out hidden messages in extracellular secretions. Clinical and Translational Oncology. 2022;25(5):1145-1155. [CrossRef]
- Sherman MH, Beatty GL. Tumor Microenvironment in Pancreatic Cancer Pathogenesis and Therapeutic Resistance. Annual Review of Pathology: Mechanisms of Disease. 2023;18(1):123-148. [CrossRef]
- Chung JC, Oh MJ, Choi SH, Bae CD. PROTEOMIC ANALYSIS TO IDENTIFY BIOMARKER PROTEINS IN PANCREATIC DUCTAL ADENOCARCINOMA. ANZ Journal of Surgery. 2008;78(4):245-251. [CrossRef]
- Brandi J, Pozza ED, Dando I; et al. Secretome protein signature of human pancreatic cancer stem-like cells. Journal of Proteomics. 2016;136:1-12. [CrossRef]
- Li X, Liu H, Dun MD; et al. Proteome and secretome analysis of pancreatic cancer cells. PROTEOMICS. 2022;22(13-14):2100320. [CrossRef]
- Liu P, Kong L, Jin H, Wu Y, Tan X, Song B. Differential secretome of pancreatic cancer cells in serum-containing conditioned medium reveals CCT8 as a new biomarker of pancreatic cancer invasion and metastasis. Cancer Cell International. 2019;19(1). [CrossRef]
- Liu P, Weng Y, Sui Z; et al. Quantitative secretomic analysis of pancreatic cancer cells in serum-containing conditioned medium. Scientific Reports. 2016;6(1). [CrossRef]
- Jo JH, Kim SA, Lee JH; et al. GLRX3, a novel cancer stem cell-related secretory biomarker of pancreatic ductal adenocarcinoma. BMC Cancer. 2021;21(1). [CrossRef]
- Kelly KA, Bardeesy N, Anbazhagan R; et al. Targeted Nanoparticles for Imaging Incipient Pancreatic Ductal Adenocarcinoma. PLoS Medicine. 2008;5(4):e85. [CrossRef]
- Liu P, Kong L, Liang K; et al. Identification of dissociation factors in pancreatic Cancer using a mass spectrometry-based proteomic approach. BMC Cancer. 2020;20(1). [CrossRef]
- Levink IJM, Visser IJ, Koopmann BDM; et al. Protein biomarkers in pancreatic juice and serum for identification of pancreatic cancer. Gastrointestinal Endoscopy. 2022;96(5):801-813.e2. [CrossRef]
- Chang YT, Wu CC, Shyr YM; et al. Secretome-Based Identification of ULBP2 as a Novel Serum Marker for Pancreatic Cancer Detection. PLoS ONE. 2011;6(5):e20029. [CrossRef]
- Firpo MA, Boucher KM, Bleicher J; et al. Multianalyte Serum Biomarker Panel for Early Detection of Pancreatic Adenocarcinoma. JCO Clinical Cancer Informatics. 2023;(7). [CrossRef]
- Kapszewicz M, Małecka-Wojciesko E. Simple Serum Pancreatic Ductal Adenocarcinoma (PDAC) Protein Biomarkers—Is There Anything in Sight? Journal of Clinical Medicine. 2021;10(22):5463. [CrossRef]
- de Oliveira G, Paccielli Freire P, Santiloni Cury S; et al. An Integrated Meta-Analysis of Secretome and Proteome Identify Potential Biomarkers of Pancreatic Ductal Adenocarcinoma. Cancers. 2020;12(3):716. [CrossRef]
- Xu T, Xu X, Liu PC, Mao H, Ju S. Transcriptomic Analyses and Potential Therapeutic Targets of Pancreatic Cancer With Concomitant Diabetes. Frontiers in Oncology. 2020;10. [CrossRef]
- Slapak EJ, Duitman J, Tekin C, Bijlsma MF, Spek CA. Matrix Metalloproteases in Pancreatic Ductal Adenocarcinoma: Key Drivers of Disease Progression? Biology. 2020;9(4):80. [CrossRef]
- Jones LE, Humphreys MJ, Campbell F, Neoptolemos JP, Boyd MT. Comprehensive Analysis of Matrix Metalloproteinase and Tissue Inhibitor Expression in Pancreatic Cancer. Clinical Cancer Research. 2004;10(8):2832-2845. [CrossRef]
- Shoucair S, Chen J, Martinson JR; et al. Association of Matrix Metalloproteinase 7 Expression With Pathologic Response After Neoadjuvant Treatment in Patients With Resected Pancreatic Ductal Adenocarcinoma. JAMA Surgery. 2022;157(7):e221362. [CrossRef]
- Zhang JJ, Zhu Y, Xie KL; et al. Yin Yang-1 suppresses invasion and metastasis of pancreatic ductal adenocarcinoma by downregulating MMP10 in a MUC4/ErbB2/p38/MEF2C-dependent mechanism. Molecular Cancer. 2014;13(1). [CrossRef]
- Giampazolias E, Schulz O, Lim KHJ; et al. Secreted gelsolin inhibits DNGR-1-dependent cross-presentation and cancer immunity. Cell. 2021;184(15):4016-4031.e22. [CrossRef]
- Xue N, Du T, Lai F, Jin J, Ji M, Chen X. Secreted HSP90α-LRP1 Signaling Promotes Tumor Metastasis and Chemoresistance in Pancreatic Cancer. International Journal of Molecular Sciences. 2022;23(10):5532. [CrossRef]
- Chen Y, Kang M, Lu W; et al. DJ-1, a novel biomarker and a selected target gene for overcoming chemoresistance in pancreatic cancer. Journal of Cancer Research and Clinical Oncology. 2012;138(9):1463-1474. [CrossRef]
- Li M, Zhai Q, Bharadwaj U; et al. Cyclophilin A is overexpressed in human pancreatic cancer cells and stimulates cell proliferation through CD147. Cancer. 2006;106(10):2284-2294. [CrossRef]
- Han JM, Jung HJ. Cyclophilin A/CD147 Interaction: A Promising Target for Anticancer Therapy. Int J Mol Sci. 2022 Aug 19;23(16):9341. [CrossRef]
- Chen Q, Wang Z, Zhang K; et al. Erratum to: Clusterin confers gemcitabine resistance in pancreatic cancer. World Journal of Surgical Oncology. 2013;11(1). [CrossRef]
- Rosati A, Bersani S, Tavano F; et al. Expression of the Antiapoptotic Protein BAG3 Is a Feature of Pancreatic Adenocarcinoma and Its Overexpression Is Associated With Poorer Survival. The American Journal of Pathology. 2012;181(5):1524-1529. [CrossRef]
- Falco A, Rosati A, Festa M; et al. BAG3 Is a Novel Serum Biomarker for Pancreatic Adenocarcinomas. American Journal of Gastroenterology. 2013;108(7):1178-1180. [CrossRef]
- Rosati A, Basile A, D’Auria R; et al. BAG3 promotes pancreatic ductal adenocarcinoma growth by activating stromal macrophages. Nature Communications. 2015;6(1). [CrossRef]
- Dufrusine B, Damiani V, Capone E; et al. BAG3 induces fibroblasts to release key cytokines involved in pancreatic cell migration. Journal of Cellular Biochemistry. 2021;123(1):65-76. [CrossRef]
- Singh RR, Mohammad J, Orr M, Reindl KM. Glutathione S-Transferase pi-1 Knockdown Reduces Pancreatic Ductal Adenocarcinoma Growth by Activating Oxidative Stress Response Pathways. Cancers. 2020;12(6):1501. [CrossRef]
- Zhang S, Yao HF, Li H; et al. Transglutaminases Are Oncogenic Biomarkers in Human Cancers and Therapeutic Targeting of TGM2 Blocks Chemoresistance and Macrophage Infiltration in Pancreatic Cancer. Research Square Platform LLC; 2023. Accessed August 2, 2023. [CrossRef]
- Prieto-Fernández L, Menéndez ST, Otero-Rosales M; et al. Pathobiological functions and clinical implications of annexin dysregulation in human cancers. Frontiers in Cell and Developmental Biology. 2022;10. [CrossRef]
- Bouter A, Gounou C, Bérat R; et al. Annexin-A5 assembled into two-dimensional arrays promotes cell membrane repair. Nature Communications. 2011;2(1). [CrossRef]
- Bouvet F, Ros M, Bonedeau E; et al. Defective Membrane Repair Machinery Impairs Survival of Invasive Cancer Cells. Cold Spring Harbor Laboratory; 2020. Accessed August 2, 2023. [CrossRef]
- Dong X, Javle M, Hess KR, Shroff R, Abbruzzese JL, Li D. Insulin-Like Growth Factor Axis Gene Polymorphisms and Clinical Outcomes in Pancreatic Cancer. Gastroenterology. 2010;139(2):464-473.e3. [CrossRef]
- Martínez-Bosch N, Cristóbal H, Iglesias M; et al. Soluble AXL is a novel blood marker for early detection of pancreatic ductal adenocarcinoma and differential diagnosis from chronic pancreatitis. eBioMedicine. 2022;75:103797. [CrossRef]
- Miller MA, Sullivan RJ, Lauffenburger DA. Molecular Pathways: Receptor Ectodomain Shedding in Treatment, Resistance, and Monitoring of Cancer. Clinical Cancer Research. 2017;23(3):623-629. [CrossRef]
- Jiang W, Li X, Xiang C, Zhou W. Neutrophils in pancreatic cancer: Potential therapeutic targets. Frontiers in Oncology. 2022;12. [CrossRef]
- Jin W, Xu HX, Zhang SR; et al. Tumor-Infiltrating NETs Predict Postsurgical Survival in Patients with Pancreatic Ductal Adenocarcinoma. Annals of Surgical Oncology. 2018;26(2):635-643. [CrossRef]
- Yi N, Zhao X, Ji J; et al. Serum galectin-3 as a biomarker for screening, early diagnosis, prognosis and therapeutic effect evaluation of pancreatic cancer. Journal of Cellular and Molecular Medicine. 2020;24(19):11583-11591. [CrossRef]
- Guo Y, Shen R, Yu L; et al. Roles of galectin-3 in the tumor microenvironment and tumor metabolism (Review). Oncology Reports. Published online September 22, 2020. [CrossRef]
- Teijeira A, Garasa S, Ochoa MC; et al. IL8, Neutrophils, and NETs in a Collusion against Cancer Immunity and Immunotherapy. Clinical Cancer Research. 2020;27(9):2383-2393. [CrossRef]
- Menzies-Gow A, Ying S, Sabroe I; et al. Eotaxin (CCL11) and Eotaxin-2 (CCL24) Induce Recruitment of Eosinophils, Basophils, Neutrophils, and Macrophages As Well As Features of Early- and Late-Phase Allergic Reactions Following Cutaneous Injection in Human Atopic and Nonatopic Volunteers. The Journal of Immunology. 2002;169(5):2712-2718. [CrossRef]
- Sabrkhany S, Kuijpers MJE, van Kuijk SMJ; et al. A combination of platelet features allows detection of early-stage cancer. European Journal of Cancer. 2017;80:5-13. [CrossRef]
- Lecot P, Ardin M, Dussurgey S; et al. Gene Signature of Circulating Platelet-Bound Neutrophils Is Associated with Poor Prognosis in Cancer Patients. Cold Spring Harbor Laboratory; 2021. Accessed August 2, 2023. [CrossRef]
- Takehara M, Sato Y, Kimura T; et al. Cancer-associated adipocytes promote pancreatic cancer progression through SAA1 expression. Cancer Science. 2020;111(8):2883-2894. [CrossRef]
- Niu X, Yin L, Yang X; et al. Serum amyloid A 1 induces suppressive neutrophils through the Toll-like receptor 2–mediated signaling pathway to promote progression of breast cancer. Cancer Science. 2022;113(4):1140-1153. [CrossRef]
- Liu YH, Hu CM, Hsu YS, Lee WH. Interplays of glucose metabolism and KRAS mutation in pancreatic ductal adenocarcinoma. Cell Death & Disease. 2022;13(9). [CrossRef]
- Yang P, Li Z, Wang Y, Zhang L, Wu H, Li Z. Secreted pyruvate kinase M2 facilitates cell migration via PI3K/Akt and Wnt/β-catenin pathway in colon cancer cells. Biochemical and Biophysical Research Communications. 2015;459(2):327-332. [CrossRef]
- Lay AJ, Jiang XM, Kisker O; et al. Phosphoglycerate kinase acts in tumour angiogenesis as a disulphide reductase. Nature. 2000;408(6814):869-873. [CrossRef]
- Principe M, Borgoni S, Novelli F. Commentary: “Alpha-enolase (ENO1) controls alpha v/beta 3 integrin expression and regulates pancreatic cancer adhesion, invasion, and metastasis.” Journal of Rare Diseases Research & Treatment. 2017;2(5):18-21. [CrossRef]
- Zhao Z, Zhang J, Yin L; et al. Upregulated GDF-15 expression facilitates pancreatic ductal adenocarcinoma progression through orphan receptor GFRAL. Aging. Published online November 17, 2020. [CrossRef]
- Albrengues J, Bourget I, Pons C; et al. LIF Mediates Proinvasive Activation of Stromal Fibroblasts in Cancer. Cell Reports. 2014;7(5):1664-1678. [CrossRef]
- Xelwa N, Candy GP, Devar J, Omoshoro-Jones J, Smith M, Nweke EE. Targeting Growth Factor Signaling Pathways in Pancreatic Cancer: Towards Inhibiting Chemoresistance. Frontiers in Oncology. 2021;11. [CrossRef]
- van den Berg YW, van den Hengel LG, Myers HR; et al. Alternatively spliced tissue factor induces angiogenesis through integrin ligation. Proceedings of the National Academy of Sciences. 2009;106(46):19497-19502. [CrossRef]
- Winer A, Adams S, Mignatti P. Matrix Metalloproteinase Inhibitors in Cancer Therapy: Turning Past Failures Into Future Successes. Molecular Cancer Therapeutics. 2018;17(6):1147-1155. [CrossRef]
- Praharaj PP, Patra S, Panigrahi DP, Patra SK, Bhutia SK. Clusterin as modulator of carcinogenesis: A potential avenue for targeted cancer therapy. Biochimica et Biophysica Acta (BBA) - Reviews on Cancer. 2021;1875(2):188500. [CrossRef]
- Zhang Y, Lu L, Song F; et al. Research progress on non-protein-targeted drugs for cancer therapy. Journal of Experimental & Clinical Cancer Research. 2023;42(1). [CrossRef]
- Cardin DB, Thota R, Goff LW; et al. A Phase II Study of Ganetespib as Second-line or Third-line Therapy for Metastatic Pancreatic Cancer. American Journal of Clinical Oncology. 2018;41(8):772-776. [CrossRef]
- Lang JE, Forero-Torres A, Yee D; et al. Safety and efficacy of HSP90 inhibitor ganetespib for neoadjuvant treatment of stage II/III breast cancer. npj Breast Cancer. 2022;8(1). [CrossRef]
- Zhang L, Wang P, Qin Y; et al. RN1, a novel galectin-3 inhibitor, inhibits pancreatic cancer cell growth in vitro and in vivo via blocking galectin-3 associated signaling pathways. Oncogene. 2016;36(9):1297-1308. [CrossRef]
- Kram M. Galectin-3 inhibition as a potential therapeutic target in non-alcoholic steatohepatitis liver fibrosis. World Journal of Hepatology. 2023;15(2):201-207. [CrossRef]
- Friedbichler K, Hofmann MH, Kroez M; et al. Data from Pharmacodynamic and Antineoplastic Activity of BI 836845, a Fully Human IGF Ligand-Neutralizing Antibody, and Mechanistic Rationale for Combination with Rapamycin. American Association for Cancer Research (AACR); 2023. Accessed August 3, 2023. [CrossRef]
- Weyer-Czernilofsky U, Hofmann MH, Friedbichler K; et al. Data from Antitumor Activity of the IGF-1/IGF-2–Neutralizing Antibody Xentuzumab (BI 836845) in Combination with Enzalutamide in Prostate Cancer Models. American Association for Cancer Research (AACR); 2023. Accessed August 3, 2023. [CrossRef]
- Schmid P, Cortes J, Joaquim A; et al. XENERA-1: A randomised double-blind Phase II trial of xentuzumab in combination with everolimus and exemestane versus everolimus and exemestane in patients with hormone receptor-positive/HER2-negative metastatic breast cancer and non-visceral disease. Breast Cancer Research. 2023;25(1). [CrossRef]
- Sullivan LA, Carbon JG, Roland CL; et al. r84, a Novel Therapeutic Antibody against Mouse and Human VEGF with Potent Anti-Tumor Activity and Limited Toxicity Induction. PLoS ONE. 2010;5(8):e12031. [CrossRef]
- Kindler HL, Niedzwiecki D, Hollis D; et al. Gemcitabine Plus Bevacizumab Compared With Gemcitabine Plus Placebo in Patients With Advanced Pancreatic Cancer: Phase III Trial of the Cancer and Leukemia Group B (CALGB 80303). Journal of Clinical Oncology. 2010;28(22):3617-3622. [CrossRef]
- Van Cutsem E, Vervenne WL, Bennouna J; et al. Phase III Trial of Bevacizumab in Combination With Gemcitabine and Erlotinib in Patients With Metastatic Pancreatic Cancer. Journal of Clinical Oncology. 2009;27(13):2231-2237. [CrossRef]
- Wang MT, Fer N, Galeas J; et al. Blockade of leukemia inhibitory factor as a therapeutic approach to KRAS driven pancreatic cancer. Nature Communications. 2019;10(1). [CrossRef]
- Borazanci E, Schram AM, Garralda E; et al. Phase I, first-in-human study of MSC-1 (AZD0171), a humanized anti-leukemia inhibitory factor monoclonal antibody, for advanced solid tumors. ESMO Open. 2022;7(4):100530. [CrossRef]
- Basile A, De Marco M, Festa M; et al. Development of an anti-BAG3 humanized antibody for treatment of pancreatic cancer. Molecular Oncology. 2019;13(6):1388-1399. [CrossRef]
- Iorio V, Rosati A, D’Auria R; et al. Combined effect of anti-BAG3 and anti-PD-1 treatment on macrophage infiltrate, CD8+ Tcell number and tumour growth in pancreatic cancer. Gut. Published online August 11, 2017:gutjnl-2017-314225. [CrossRef]
- De Marco M, Gauttier V, Pengam S; et al. Concerted BAG3 and SIRPα blockade impairs pancreatic tumor growth. Cell Death Discovery. 2022;8(1). [CrossRef]
- Iorio V, De Marco M, Basile A; et al. CAF-Derived IL6 and GM-CSF Cooperate to Induce M2-like TAMs–Letter. Clinical Cancer Research. 2019;25(2):892-893. [CrossRef]
- Rosati A, Marzullo L, De Marco M, De Laurenzi V, D’Amico MF, Turco MC. Toxicity in combined therapies for tumours treatments: A lesson from BAG3 in the TME? Frontiers in Immunology. 2023;14. [CrossRef]
- Li P, Rozich N, Wang J; et al. Anti-IL-8 antibody activates myeloid cells and potentiates the anti-tumor activity of anti-PD-1 antibody in the humanized pancreatic cancer murine model. Cancer Letters. 2022; 539:215722. [CrossRef]
- Bilusic M, Heery CR, Collins JM; et al. Phase I trial of HuMax-IL8 (BMS-986253), an anti-IL-8 monoclonal antibody, in patients with metastatic or unresectable solid tumors. Journal for ImmunoTherapy of Cancer. 2019;7(1). [CrossRef]
- .Lewis CS, Karve A, Matiash K; et al. A First-In-Class, Humanized Antibody Targeting Alternatively Spliced Tissue Factor: Preclinical Evaluation in an Orthotopic Model of Pancreatic Ductal Adenocarcinoma. Frontiers in Oncology. 2021;11. [CrossRef]
- Ganguly K, Kimmelman AC. Reprogramming of tissue metabolism during cancer metastasis. Trends in Cancer. 2023;9(6):461-471. [CrossRef]
Figure 1.
Proteins Functions- A Global Core Biodata Resource, Panther Classification System (https://pantherdb.org/, accessed in June 2023), classified the gene dataset for functions. Proteins secreted by pancreatic cancer cells (right column) were classified as having a general function (center column) or a specific function (left column). An Alluvial plot (https://www.mapequation.org/alluvial/, accessed in June 2023) was used to depict the classifications.
Figure 1.
Proteins Functions- A Global Core Biodata Resource, Panther Classification System (https://pantherdb.org/, accessed in June 2023), classified the gene dataset for functions. Proteins secreted by pancreatic cancer cells (right column) were classified as having a general function (center column) or a specific function (left column). An Alluvial plot (https://www.mapequation.org/alluvial/, accessed in June 2023) was used to depict the classifications.
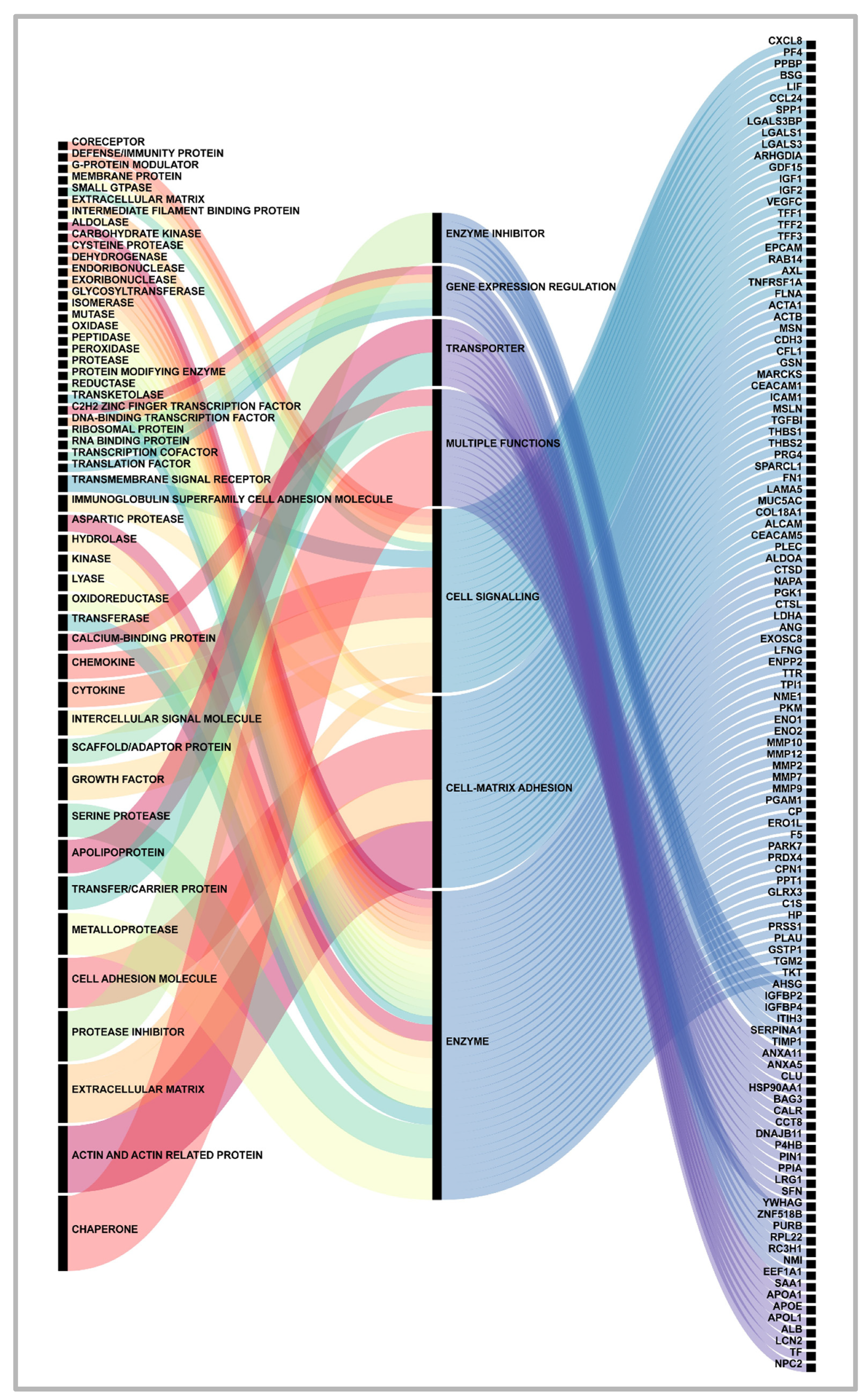
Figure 2.
Druggable secreted proteins landscape in pancreatic cancer cells. Image was realized with Biorender.
Figure 2.
Druggable secreted proteins landscape in pancreatic cancer cells. Image was realized with Biorender.
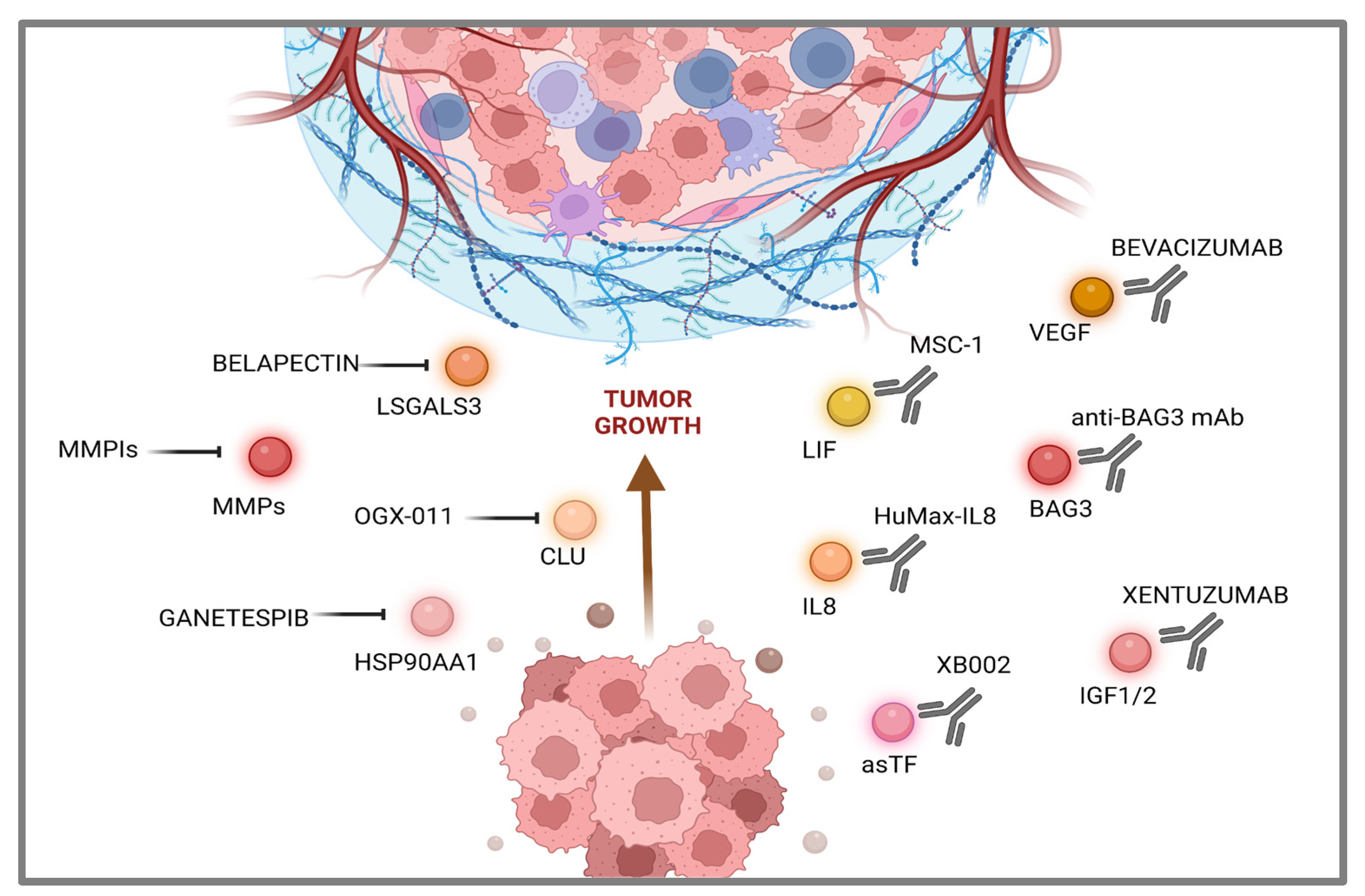

Table 1.
Pancreatic cancer cells- secreted proteins dataset.
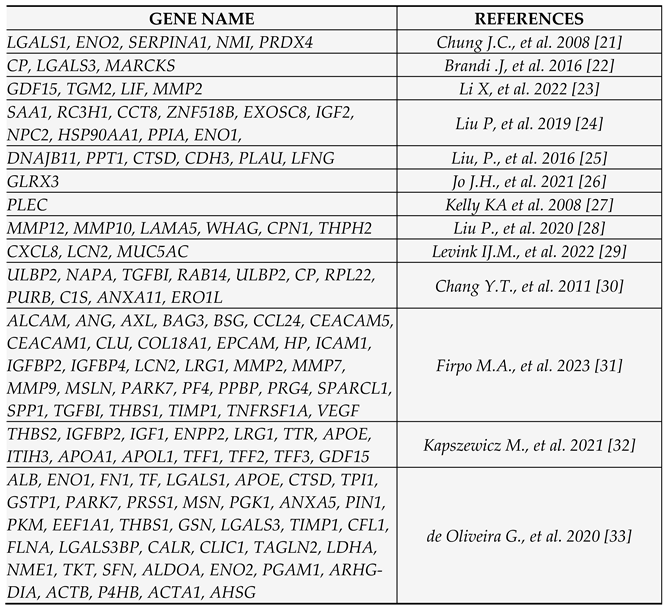
Table 2.
Pancreatic cancer cells secreted proteins dataset Enrichr-Appyter online applications generated a downloadable table. The names of the genes were reported along with the p-values and q-values of significant terms in the chosen library. The q-value is an adjusted p-value calculated using the Benjamini-Hochberg method for multiple hypothesis testing correction.
Table 2.
Pancreatic cancer cells secreted proteins dataset Enrichr-Appyter online applications generated a downloadable table. The names of the genes were reported along with the p-values and q-values of significant terms in the chosen library. The q-value is an adjusted p-value calculated using the Benjamini-Hochberg method for multiple hypothesis testing correction.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



