
Submitted:
31 July 2023
Posted:
31 July 2023
You are already at the latest version
Abstract
Most fungal species are commensals, not pathogenic for plants, humans, or animals. However, several species from genera Alternaria, Aspergillus, Trichophyton and Microsporum are common causes of disease, even for immunocompetent individuals, contributing to skin barrier impairment. A total of 68 allergic dogs were selected from a veterinary dermatology and allergy outpatient consultation, for conditions related to Malassezia overgrowth or other fungal complications. Allergy diagnosis was made through anamnestic and clinical criteria and allergy-implicated species were identified by intradermal testing (IDT) and serum determination of specific immunoglobulin E (sIgE). Dermatophagoides farinae, Dactylis glomerata and Malassezia pachydermatis showed as the higher sensitization species from each group. Significant correlations at p<0.05 were found between Dactylis glomerata and Phleum pratense grass pollens, Dermatophagoides farinae and Dermatophagoides pteronyssinus, Acarus siro, Tyrophagus putrescentiae and Lepidoglyphus destructor dust/storage mites, and between fungi like Aspergillus mix and Penicillium or Alternaria alternata. A significant correlation was also found between sensitization to Aspergillus mix and D. farinae, D. pteronyssinus or A. siro. Rather severe dermatitis was observed where positive IDT to Malassezia pachydermatis was found, regardless of the presence of circulating sIgE, allowing us to perceive the usefulness of both sIgE and IDT for allergy diagnosis to fungi.
Keywords:
Dermatomycosis
; allergy
; Alternaria
; Aspergillus
; dermatophytes
; fungal allergy
; Malassezia
; Penicillium
1. Introduction
Most fungi, which evolved for around 1.5 billion years [1], belong to saprophytic species, not pathogenic to plants, humans, or animals. Nevertheless, a small part may become pathogenic, either by producing toxins, infecting, or sensitizing with subsequent allergy [2]. Species from the fungal kingdom are ubiquitous, with some of them, like those from the genera Alternaria, Aspergillus, Fusarium, Mucor [2], Trichophyton and Microsporum [3] being a possible cause of disease in people and animals, mostly in face of immune-compromised conditions [2].
As organic matter decomposers, Fungi secrete enzymes into the surrounding environment, digesting other organisms’ molecules like carbohydrates, proteins and lipids and then absorbing the resulting nutrients (e.g. carbohydrate metabolytes) in a heterotrophic way. Enrichment of the surrounding environment in fungi metabolic leftovers eventually occurs with the airborne spreading of spores, hyphae and their fragments [4].
Different species of fungi have been recognized as sensitizers, hence possibly allergenic. Besides respiratory infection [5], several species of the Aspergillus genus have been implicated in respiratory allergies like bronchopulmonary, sinusitis and IgE-mediated asthma or hypersensitivity pneumonitis [6,7]. Sensitization to Alternaria fungus has been also reported, mostly in warm climates, associated with type I hypersensitivity, in either indoor or outdoor environments [8,9]. Fusarium species, a common cereal contaminant, may lead to immune system impairment and sensitization with allergy. Allergic alveolitis, atopic conjunctivitis, bronchial asthma and rhinitis are among the most common manifestations [10]. Fusarium allergens are known for cross-reacting each other [11] and may also cross-react with other species [12]. Curvularia, a genus with over 40 mostly saprophytic species, may also sensitize with mainly respiratory signs in humans [13]. It also cross-reacts with Alternaria alternata and Epicoccum nigrum [14,15]. Curvularia infection [16] and allergy have been also reported in dogs [17]. Cladosporium is another ubiquitous genus with reported infection in humans and dogs [18], horses [19] and cats [20,21] as well as allergy in humans [2,22] and dogs [23]. Mucor and Rhizopus, two genera from the Mucorales group, are also related to allergy in humans [8] and animals [17,24,25].
Dermatophytosis, a zoonosis mainly caused by fungi from the genera Microsporum, Trichophyton and Epidermophyton, is a common condition in human and animal immunocompetent individuals [26,27] (Figure 1). Allergy to Dermatophyte is a known condition, with several allergens identified from Trichophyton and associated IgE-mediated asthma in humans. Those allergens may either trigger immediate or delayed hypersensitivity. Delayed responses seem to confer protection, while immediate not. Trichophyton allergens may play a simultaneous role in fungal pathogenesis and allergenicity as suggested by their amino-acid sequence. This dual condition may be associated with particular T-cell epitopes, which may play a key role in new peptide vaccines, with better efficiency regarding Trichophyton infection and allergy [28].
Candida is a genus with over 200 species, most of them commensals of human and animal microbiota and only facultatively pathogenic, in which 15 species have been isolated from human and animal infections, affecting several organs besides mucosa or skin [29]. Candidiasis is not an often cause of infection in animals but may occur associated with atopy [30,31].
Sensitization to fungi reaches roughly 5% of the global population, however, this rate stands higher in the atopic group. Exposure to fungal allergens may vary according to the environment, occurring by contact with intact spores, mycelia, or their fragments. Germinating spores present a higher allergen variety. Hence, the kind of environment in which individuals live will play a crucial role in the contact with fungal particles, with fungal structure-derived particles becoming aerosolized in concentrations 300 to 500 times greater than the original spores [32]. Being certainly high, contact with fungi may result in underestimated subclinical sensitization. Moreover, contact with primary sensitization to a small number of fungal species may result in sensitization to a high variety of species as sensitization to fungi is considered highly cross-reactive. In fact, when a group of 6565 individuals with positive IgE in at least one fungal test was tested with a larger battery of fungal species, 1208 showed positive to all [33]. Fungal proteins sharing homologous structures and similar functions have shown marked cross-reactivity [34,35].
Fungi commonly fill a high concentration of airborne allergens, but increased exposure to indoor microbial diversity may play a protective role for atopy [8]. However, studies on the Alternaria genus, one the most associated with allergy, have shown that even low-level fungal exposure for long periods is not necessarily associated with sensitization, unless in atopic individuals. Fungi-related respiratory disease is frequently associated with either atopy [8] or immune-impairment [2].
Recurrent airway obstruction (RAO), a common condition in equine, has been associated with frequent exposure to moldy hay, but only non-IgE-mediated mechanisms have been implicated in the pathogenesis. However, sensitization to fungi with clinical aggravation in moldy environments and by challenging with mold extracts have been observed as well as higher scores of basophil histamine-releasing test upon stimulation with fungal allergens in horses with RAO, when compared with healthy individuals [36]. Regarding Aspergillus fumigatus, sIgE and IgG response was found in bronchoalveolar lavage (BAL) fluid of RAO-affected horses, following in vitro provocation with fungal extracts [37]. Despite no difference in specific IgE to fungal extracts between healthy and affected horses, sIgE to fungal allergens, like Alt a 1, Asp f 7, 8 and 9, were mainly detected in BAL and serum from RAO-affected patients. Difference in sIgE to Asp f 7 was also observed between RAO-affected and healthy individuals [38,39].
Despite the lack of identification of the major allergens implicated in sensitization, mold allergome to horses has already started to be disclosed, and that course is also needed regarding dogs and cats [40].
Skin testing is currently performed with whole-allergen extracts, which may vary in the allergen content, compromising inter-brand and even inter-batch reproducibility. After the clinical diagnosis of atopy, IDT (Figure 2) stands as the veterinary allergy first-line diagnosis method for implicated species. Currently, most suppliers present allergen extracts in well-defined concentrations, which have been presented for most groups of allergenic species [41]. However, standardization still needs improvement to assure better reproducibility [42].
Malassezia is another relevant genus from a lipophilic group of yeasts [43], in veterinary medicine. Malassezia pachydermatis, a common commensal inhabitant of dog and cat skin and mucosa, is the most reported species out of 18 [44].
Despite the commensal condition, different factors like the host’s innate/adaptive immune impairment and secreted virulence factors, Malassezia populations may overgrow with severe dermatitis [44], a frequent condition observed in allergic dogs, with or without sensitization to Malassezia [41] (Figure 3).
Skin inflammatory conditions, mostly associated with impaired epidermal surface, sebum production, or excessive moisture may favor Malassezia overgrowth, in a faulty context of the complex cutaneous equilibrium. In fact, besides keratinization disorders, several endocrinopathies, metabolic diseases and neoplasia, allergies like atopic dermatitis, food hypersensitivity, or flea allergy frequently comprise a set of conditions commonly associated with Malassezia pachydermatis overgrowth with dermatitis [45]. Malassezia overgrowth in dogs usually shows as a ceruminous pruritic external otitis or intertriginous dermatitis. Skin becomes erythematous, evolving to kerato-sebaceous skin thickening [46].
Not many studies have been published in veterinary medicine, regarding Malassezia-derived animal conditions, despite its clinical relevance [42]. In 2021, Di Tomaso et al. reported the results of sera evaluation from 45 dogs, in which 11 showed positive for Malassezia [47]. Another study, evaluating both IDT and serum specific-IgE to Malassezia, revealed 24% positivity in IDT, despite no positivity in sera [48]. Already in 2009, Furiani et al. had reported a percentage of positivity of 35% in IDT [49].
In a retrospective study with 111 allergic dogs, mainly atopic, including 33 from predisposed breeds, mostly with seborrheic disruptive skin barrier, 49.6% showed Malassezia overgrowth-associated dermatitis [50]. In another retrospective study with 84 allergic dogs, mainly atopic, including 31 from predisposed breeds, 58,3% presented with Malassezia overgrowth-associated dermatitis [51].
Specifically regarding Malassezia, several dog breeds, like American Cocker Spaniel, Australian Silky Terrier, Basset Hound, Boxer, Dachshund, English Poodle, Setter, Shih Tzu and West Highland White Terrier have been pointed as being at higher risk for overgrowth, leading to dermatitis. Towards cats, Devon Rex and Sphynx are the two breeds recognized as more predisposed to Malassezia overgrowth [52], but the condition has been mostly diagnosed in individuals suffering from visceral paraneoplastic syndromes [46]. Zoonotic risk is assumed low, especially in immunocompetent individuals [44]
2. Materials and Methods
2.1. Dog population
From a veterinary dermatology and allergy outpatient consultation, at the University of Évora Veterinary Hospital (Évora, Portugal) a total of 68 allergic dogs were selected for conditions related to Malassezia overgrowth and/or other fungal complications. Primary allergy diagnosis/selection was made through a comprehensive query for anamnestic and clinical criteria [42,53].
2.2. Methods
The further diagnostic evaluation followed IDT by inoculation of 50 µL of commercial allergen extracts from a set of relevant allergenic species, such as Dactylis glomerata and Phleum pratense grass pollens, Dermatophagoides farinae, Dermatophagoides pteronyssinus, Acarus siro, Tyrophagus putrescentiae and Lepidoglyphus destructor dust/storage mites, and fungi as Alternaria alternada, Aspergillus mix and Malassezia pachidermatis. Positive (0.01% histamine phosphate solution) and negative (physiological saline solution) commercial controls were also administered (Diater and Nextmune, Madrid, Spain). Dermal wheal and flare reactions were evaluated after 15 minutes. Reactions were considered positive when the resulting wheals were at least equal to or higher than halfway between the negative and the positive control reaction, and then scored increasingly from 0 (negative) to 4 (maximum positive) [42]. All dogs were sedated with commercial medetomidine (Orion Pharma, Espoo, Finland) (roughly 0.03 mg/kg body weight, administered subcutaneously) and sedation was reversed post-testing with the correspondent dose of atipamezole (Orion Pharma, Espoo, Finland) administered intramuscularly. Allergen-sIgE to an environmental panel of allergenic species, including the ones used for IDT, were determined in macELISA (LETI Animal Health Laboratories, Barcelona, Spain) and results expressed in ELISA Absorbance Units (EAU). Thirty dogs were simultaneously submitted to IDT and sIgE determination, 21 only to sIgE and 17 only to IDT. Differences between intradermal wheal scores and between sIgE ELISA EAU were compared using the Pearson correlation coefficient. Statistical significance was set at p<0.05.
3. Results
3.1. General observations
IDT and sIgE scores varied according to individual sensitization patterns, skin reactivity as well as according to each extract potency. sIgE and IDT positive rates are shown in Table 1. Decreasing sensitization rate, observed in positive sIgE, was: D. farinae > T. putrescentiae > A. siro > D. pteronyssinus > D. glomerata > P. pratense > L. desctructor > M. pachidermatis > Aspergillus mix. No circulating sIgE was found to Alternaria alternata in this population. Regarding IDT, decreasing positive rate was: D. farinae and L. destructor > A. siro > T. putrescentiae > D. pteronyssinus > A. alternata > Aspergilus mix > M. pachidermatis > D. glomerata > Penicillium. Penicillium was not assessed in IDT.
House dust and storage mites clearly showed as the most allergenic species for this population, followed by the two grass pollen species regarding circulating sIgE, besides the higher rate of positive IDT to the fungal species. Only 3 dogs sensitized to grass pollens did not present with sIgE to both D. glomerata and Phleum pratense, while 9 were without positive IDT to both.
The highest rate of positive sIgE to fungi was found to M. pachidermatis (10 out of 51), followed by Aspergilus mix (6 out of 51) and Penicilum mix (4 out of 51). No dog presented with circulating sIgE to A. alternata, despite 9 out of 47 having presented positive IDT, a rate close to the observed to the other fungal species intradermally tested. No dog revealed simultaneously sensitized to the four fungal species. The highest positivity to fungi was found to M. pachidermatis. However, from the 10 dogs with sIgE and 7 with positive IDT, only one showed positive for both tests. In the same way, none of the tested dogs presented with simultaneous positive sIgE and IDT to Aspergillus mix. M. pachidermatis was found as the only sensitizing fungal species in 9 dogs, while Aspergillus mix in 5 and Alternaria alternata in 4. No dog revealed Penicillium as a single fungal sensitizer. Twenty-three dogs revealed reactive to at least one fungal species/group of species and 14, most of the fungi-reactive patients, revealed reactive to at least two or three species/group of species.
3.2. Associations observed
Regarding house dust-mite and storage-mite sensitization, significant correlation was found between sIgE to D. farinae and D. pteronyssinus (r=0.587; p<0.00001), A. siro (r=0.958; 0.00001), T. putrescentiae (r=0.967; p<0.00001), and L. destructor (r=0.343; p=0.01). In terms of intradermal reactivity, significant correlation was also found between D. farinae and D. pteronyssinus (r=0.384; p=0.007), A. siro (r=0.446; p=0.001) and L. destructor (r=0.336; p=0.02).
Regarding grass pollen, significant correlation was found between D. glomerata and P. pratense, either in terms of sIgE (r=0.885; p<0.00001) or intradermal reactivity (r=0.31; p=0.027).
For simultaneous sensitization to fungi and house dust or storage mites, a significant correlation was found between sIgE to Aspergillus mix and D. farinae (r=0.283; p=0.046) or A. siro (r=0.288; p=0.042). Sensitization between Aspergillus mix and Penicillium mix also correlated (r=0.356; p=0.011) as well as positive IDT to Alternaria alternata and Aspergillus mix (r=0.599; p<0.00001). Correlation between Aspergillus mix and D. pteronyssinus was also observed (0.494; p=0.0002).
4. Discussion
As ubiquitous forms of life, fungi and their particles are common components of the environment, contacting with mucosal and cutaneous barriers. Hence, deep penetration will depend on the quality of those barriers, with a low skin barrier effect, which is commonly seen in allergic dogs [54] allowing higher penetration with sensitization, in dependence of individual genetic predisposition [55].
House dust and storage mites have been shown as the most allergenic species in this population, where sensitization and IDT scores were also high. Pollinosis was not suspected as clinically relevant for most patients, despite their sensitization to D. glomerata and P. pratense. Many of these dogs presented sensitization to those pollens, without leading to pollinosis.
Regarding fungal species, sensitization was found low, despite an expressive rate of positive IDT, associated with a compatible clinical frame, with consistent improvement following directed antifungal treatment and environmental sanitation measures.
Associations found regarding sIgE or IDT, between dust and storage mites [56,57,58] and between grass pollen species [59,60,61] were expected as genetic predisposition to develop sensitization as well as cross-reaction occur. Interestingly, the highest significant correlation (r=0.967; p<0.00001) observed in sIgE was found between D. farinae and T. putrescentiae as suggested by ELISA inhibition studies recently performed by Song et. al. (2022) [58]. Sometimes, those underlying sensitizations conduce to allergy and sometimes not [41,62,63]. Only three dogs sensitized to grass pollens did not present with sIgE to both D. glomerata and P. pratense, while nine were found without positive IDT for both, which may be related to a higher sensitization rate, without allergy, as atopic dermatitis pollinosis would demand not only circulating sIgE, but the presence of sIgE coupled to skin mastocytes, leading to positive IDT. Similar condition was found regarding M pachidermtis, for instance, where ten dogs presented with sIgE but only seven positives in IDT. The associations found between sensitization or IDT to dust and storage mites and fungi (e.g. D. farinae, A. siro and D. pteronyssinus regarding Aspergillus mix), besides individual predisposition, may be also related to environmental living conditions, favoring the prevalence of both groups of allergenic species. That may be also suspected by the simultaneous sensitization found to more than one fungal species. However, some of the positive correlations found as the one between D. pteronyssinus, a dust mite recognized as less sensitizer to dogs, and Aspergillus mix, for instance, may decay with sample enlargement. About fungi, cross-sensitization has been also reported [11,12,14,15,64], even with food [65].
Despite sensitization to the different fungi species tested, the highest rate was observed to M. pachidermatis, with 16 positive cases (sIgE and IDT) and being the only sensitizing species in 9 individuals across this group of dogs, which may be explained by the fact that it was an atopic population, where Malassezia overgrowth is highly frequent. Despite the presence of sIgE to M. pachidermatis, more severe related inflammatory skin conditions were observed when positive IDT was found. Moreover, as with other allergen species, and probably even more, because of the proteolytic capacity inherent to mold extracts, with consequent self-disruption, specific diagnosis of allergy to fungi should also undergo both sIgE and IDT, allowing filling the flaws of each method.
Acknowledgments
The author thanks Ana Martins for proof-reading the text.
Conflicts of interest
The author declares no conflict of interest.
References
- Wang, D.Y.; Kumar, S.; Hedges, S.B. Divergence time estimates for the early history of animal phyla and the origin of plants, animals and fungi. Proc. Royal Soc. 1999, 266, 163–171. [Google Scholar] [CrossRef] [PubMed]
- De Lucca, A.J. Harmful fungi in both Agriculture and Medicine. Rev. Iberoam Micol. 2007, 24, 3–13. [Google Scholar] [CrossRef]
- Moriello, K.A.; Coyner, K.; Paterson, S.; Mignon, B. Diagnosis and treatment of dermatophytosis in dogs and cats. Clinical Consensus Guidelines of the World Association for Veterinary Dermatology. Vet Dermatol. 2017, 28, 266–268. [Google Scholar] [CrossRef]
- Yafetto, L.; Carroll, L.; Cui, Y.; Davis, D.J.; Fischer, M.W.; Henterly, A.C.; Kessler, J.D.; Kilroy, H.A.; Shidler, J.B.; Stolze-Rybczynski, J.L.; et al. The fastest flights in nature: Highspeed spore discharge mechanisms among fungi. PLoS ONE 2008, 3, e3237. [Google Scholar] [CrossRef] [PubMed]
- Akan, M.; Hazirogiu, R.; Ihan, Z.; Sareyyupogiu, B.; Tunca, R. A case of aspergillosis in a broiler breeder flock. Avian Dis. 2002, 46, 497–501. [Google Scholar] [CrossRef]
- Maurya, V.; Gugnami, H.C.; Sarnia, P.U.; Madan, T.; Shah, A. Sensitization to Aspergillus antigens and occurrence of allergic bronchopulmonary aspergillosis in patients with asthma. Chest 2005, 127, 1252–1259. [Google Scholar] [CrossRef]
- Shah, A.; Panjabi, O. Allergic bronchopulmonary aspergillosis: A review of a disease with a worldwide distribution. J. Asthma. 2002, 39, 273–289. [Google Scholar] [CrossRef] [PubMed]
- Barnes, C. Fungi and Atopy. Clin. Rev. Allergy Immunol. 2019, 57, 439–448. [Google Scholar] [CrossRef]
- Cˇ elakovská, J.; Bukacˇ, J.; Ettler, K.; Vaneckova, J.; Ettlerova, K.; Krejsek, J. Sensitisation to outdoor and indoor fungi in atopic dermatitis patients and the relation to the occurrence of food allergy to peanuts and walnuts. Mycoses 2018, 61, 698–703. [Google Scholar] [CrossRef]
- Lacey, J.; Dutkiewicz, J. Bioaerosols and occupational lung disease. J. Aerosol. Sci. 1994, 25, 1371–1404. [Google Scholar] [CrossRef]
- Verma, J.; Singh, B.P.; Sridhara, S.; Gaur, S.N.; Arora, N. Purification and characterization of a cross-reactive 45-kD major allergen of Fusarium solani. Int. Arch. Allergy. Immunol. 2003, 130, 193–199. [Google Scholar] [CrossRef]
- O’Neil, O.E.; McOants, M.L.; Salvaggio, J.E.; Lehrer, S.B. Fusarium solani: Prevalence of skin reactivity and antigenic allergenic analysis. J. Allergy Clin. Immunol. 1986, 77, 842–849. [Google Scholar] [CrossRef]
- Gupta, R.; Singh, B.P.; Sridhara, S.; Gaur, S.N.; Chaudhary, V.K.; Arora, N. Allergens of Curvularia lunata during cultivation in different media. J. Allergy Clin. Immunol. 1999, 4, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Singh, B.P.; Sridhara, S.; Gaur, S.N.; Kunar, R.; Chaudhary, V.K.; Arora, N. Identification of cross-reactive proteins amongst different Curvularia species. Int. Arch. Allergy Immunol. 2002, 127, 38–46. [Google Scholar] [CrossRef]
- Gupta, R.; Singh, B.P.; Sridhara, S.; Gaur, S.N.; Kumar, R.; Ghaudhary, V.K.; Arora, N. Allergenic cross-reactivity of Curvularia lunata with other airborne fungal species. Allergy 2002, 57, 636–640. [Google Scholar] [CrossRef] [PubMed]
- Strzok, E.; Siepker, C.; Armwood, A.; Howerth, E.; Smith, J.; Banovic, F. Successful treatment of cutaneous Curvularia geniculata, Nocardia niigatensis, and viral papillomatosis in a dog during the therapeutic management of immune-mediated hemolytic anemia. Front Vet. Sci. 2019, 6, 249. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.; Kim, H.; Jang, H.; Park, H. Sensitization rates of causative allergens for dogs with atopic dermatitis: Detection of canine allergen-specific IgE. J. Vet. Med. Sci. 2014, 15, 545–550. [Google Scholar] [CrossRef]
- Spano, M.; Zuliani, D.; Peano, A.; Bertazzolo, W. Cladosporium cladosporioides-complex infection in a mixed-breed dog. Vet. Clin. Pathol. 2018, 47, 150–153. [Google Scholar] [CrossRef]
- Dauvillier, J.; TerWoort, F.; van Erck-Westergren, E. Fungi in respiratory samples of horses with inflammatory airway disease. J. Vet. Intern. Med. 2019, 33, 968–975. [Google Scholar] [CrossRef]
- Jarrah, S.A.; Zanetti, C.C.; Maruyama, F.H.; Ito, A.T.; Rosa, J.M.A.; Colodel, E.M.; Lima, S.R.; Nakazato, L.; Dutra, V. Cladosporium cladosporioides isolated from a cat with squamous cell carcinoma. Arq. Bras. Med. Vet. Zootec. 2017, 69, 377–380. [Google Scholar] [CrossRef]
- Mariani, C.L.; Platt, S.R.; Scase, T.J.; Howerth, E.W.; Chrisman, C.L.; Clemmons, R.M. Cerebral phaeohyphomycosis caused by Cladosporium spp. in two domestic shorthair cats. J. Am. Anim. Hosp. Assoc. 2002, 38, 225–230. [Google Scholar] [CrossRef]
- Ozdemir, O. Molds and Respiratory Allergy–Part 1. MOJ Immunol. 2015, 2, 00045. [Google Scholar] [CrossRef]
- Meason-Smith, C.; Diesel, A.; Patterson, A.P.; Older, C.E.; Mansell, J.M.; Suchodolski, J.S.; Hoffmann, A.R. What is living on your dog’s skin? Characterization of the canine cutaneous mycobiota and fungal dysbiosis in canine allergic dermatitis. FEMS Microbiol. Ecol. 2015, 91, fiv139. [Google Scholar] [CrossRef]
- Cafarchia, C.; Figueredo, L.A.; Otranto, D. Fungal diseases of horses. Vet. Microbiol. 2013, 167, 215–234. [Google Scholar] [CrossRef]
- Youn, H.Y.; Kang, H.S.; Bhang, D.H.; Kim, M.K.; Hwang, C.Y.; Han, H.R. Allergens causing atopic diseases in canine. J. Vet. Sci. 2002, 3, 335–341. [Google Scholar] [CrossRef]
- Lund, A.; DeBoer, D.J. Immunoprophylaxis of dermatophytosis in animals. Mycopathologia 2008, 166, 407–424. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, F.; Crepaldi, N.; Tognoli, L.; Pereira, R.E.P. Dermatófitos–Revisão de literatura. Rev. Cient Eletrônica. Med. Vet. 2009, 12. [Google Scholar]
- Woodfolk, J. Allergy and dermatophytes. Clin. Microbiol. Rev. 2005, 18, 30–43. [Google Scholar] [CrossRef]
- Seyedmousavi, S.; Bosco, S.d.G.; de Hoog, S.; Ebel, F.; Elad, D.; Gomes, R.R.; Jacobsen, I.D.; Jensen, H.E.; Martel, A.; Mignon, B.; et al. Fungal infections in animals: A patchwork of different situations. Med. Mycol. 2018, 56, S165–S187. [Google Scholar] [CrossRef]
- Yurayart, C.; Chindamporn, A.; Suradhat, S.; Tummaruk, P.; Kajiwara, S.; Prapasarakul, N. Comparative analysis of the frequency, distribution and population sizes of yeasts associated with seborrheic dermatitis and healthy skin. Vet. Microbiol. 2011, 148, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Mueller, R.S.; Bettenay, S.V.; Shipstone, M. Cutaneous candidiasis in a dog caused by Candida guilliermondii. Vet. Rec. 2002, 150, 728–730. [Google Scholar] [CrossRef] [PubMed]
- Green, B.J.; Tovey, E.R.; Sercombe, J.K.; Blachere, F.M.; Beezhold, D.H.; Schmechel, D. Airborne fungal fragments and allergenicity. Med. Mycol. 2006, 44 (Suppl. 1), S245–S255. [Google Scholar] [CrossRef] [PubMed]
- Amado, M.; Portnoy, J.M.; Barnes, C. Fungal cross-allergenicity in specific IgE testing. J. Allergy Clin. Immunol. 2014, 133 (Suppl. 2), AB92. [Google Scholar] [CrossRef]
- Williams, P.B.; Barnes, C.S.; Portnoy, J.M. Environmental allergens workgroup. Innate and adaptive immune response to fungal products and allergens. J. Allergy Clin. Immunol. Pract. 2016, 4, 386–395. [Google Scholar]
- Crameri, R.; Weichel, M.; Fluckiger, S.; Glaser, A.G.; Rhyner, C. Fungal allergies: A yet unsolved problem. Chem. Immunol. Allergy 2006, 91, 121–133. [Google Scholar]
- Dirscherl, P.; Grabner, A.; Buschmann, H. Responsiveness of basophil granulocytes of horses suffering from chronic obstructive pulmonary disease to various allergens. Vet. Immunol. Immunopathol. 1993, 38, 217–227. [Google Scholar] [CrossRef]
- Schmallenbach, K.H.; Rahman, I.; Sasse, H.H.; Dixon, P.M.; Halliwell, R.E.; McGorum, B.C.; Crameri, R.; Miller, H.R. Studies on pulmonary and systemic Aspergillus fumigatus-specific IgE and IgG antibodies in horses affected with chronic obstructive pulmonary disease (COPD). Vet. Immunol. Immunopathol. 1998, 66, 245–256. [Google Scholar] [CrossRef]
- Kunzle, F.; Gerber, V.; Van Der Haegen, A.; Wampfler, B.; Straub, R.; Marti, E. IgE-bearing cells in bronchoalveolar lavage fluid and allergen-specific IgE levels in sera from RAO-affected horses. J. Vet. Med. A Physiol. Pathol. Clin. Med. 2007, 54, 40–47. [Google Scholar] [CrossRef]
- Eder, C.; Crameri, R.; Mayer, C.; Eicher, R.; Straub, R.; Gerber, H.; Lazary, S.; Marti, E. Allergen-specific IgE levels against crude mould and storage mite extracts and recombinant mould allergens in sera from horses affected with chronic bronchitis. Vet. Immunol. Immunopathol. 2000, 73, 241–253. [Google Scholar] [CrossRef]
- Mueller, R.S.; Janda, J.; Jensen-Jarolim, E.; Rhyner, C.; Marti, E. Allergens in veterinary medicine. Allergy 2016, 71, 27–35. [Google Scholar] [CrossRef]
- Martins, L.; Bento, O.; Inacio, F. Veterinary allergy diagnosis: Past, present and future perspectives. Allergo J. Int. 2016, 25, 20–32. [Google Scholar] [CrossRef]
- Hensel, P.; Santoro, D.; Favrot, C.; Hill, P.; Griffin, C. Canine atopic dermatitis: Detailed guideline for diagnosis and allergen identification. BMC Vet. Res. 2015, 11, 196. [Google Scholar] [CrossRef] [PubMed]
- Matricardi, P.M.; Kleine-Tebbe, J.; Hoffmann, H.J.; Valenta, R.; Hilger, C.; Hofmaier, S.; Aalberse, R.C.; Agache, I.; Asero, R.; Ballmer-Weber, B.; et al. EAACI Molecular Allergology User’s Guide. Pediatr. Allergy Immunol. 2016, 27 (Suppl. 23), 1–250. [Google Scholar] [PubMed]
- Bond, R.; Morris, D.O.; Guillot, J.; Bensignor, E.J.; Robson, D.; Mason, K.V.; Kano, R.; Hill, P.B. Biology, diagnosis and treatment of Malassezia dermatitis in dogs and cats. Clinical Consensus Guidelines of the World Association for Veterinary Dermatology. Vet. Dermatol. 2020, 31, 27.e4. [Google Scholar] [CrossRef]
- Balappanavar, B.R.; Vasanth, M.S. Clinico-diagnostic and therapeutic management of canine malasseziosis. Intas. Polivet. 2013, 14, 353–357. [Google Scholar]
- Guillot, J.; Bond, R. Malassezia yeasts in veterinary dermatology: An updated overview. Front. Cell Infect Microbiol. 2020, 10, 79. [Google Scholar] [CrossRef] [PubMed]
- Di Tommaso, M.; Luciani, A.; Crisi, P.E.; Beschi, M.; Rosi, P.; Rocconi, F.; Miglio, A. Detection of serum allergen-specific IgE in atopic dogs tested in northern Italy: Preliminary study. Animals 2021, 11, 358. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Chan, W.Y.; Hill, P.B. Prevalence of positive reactions in intradermal and IgE serological allergy tests in dogs from south Australia, and the subsequent outcome of allergen-specific immunotherapy. Aust. Vet. J. 2020, 98, 17–25. [Google Scholar] [CrossRef]
- Furiani, N.; Scarampella, F.; Noli, C.; Ordeix, L. A retrospective study of 486 intradermal tests performed on atopic dogs in Northern Italy. Veterinaria (Cremona) 2009, 23, 41–46. [Google Scholar]
- Cecci, G.M.; Cardoso, M.L.; Bento, O.; Martins, L.M. Allergy approach to a dog population from a veterinary dermatology consultation at the tropical inland city of Londrina, Paraná, Brazil. In Proceedings of the European Academy of Allergy and Clinical Immunology (EAACI) Congress, Lisbon, Portugal, 1–5 June 2019. [Google Scholar]
- Esteves de Campos, I.; Ferreiro Pinto, C.; Munhoz Severino, A.C.; Bento, O.; Antunes, C.; Costa, A.R.; Martins, L.M. Dermatological and allergy approach to a dog population from a veterinary consultation at the tropical coastal city of São Paulo, Brazil. In Proceedings of the European Academy of Allergy and Clinical Immunology (EAACI) Congress, Lisbon, Portugal, 1–5 June 2019. [Google Scholar]
- Fujimura, K.E.; Sitarik, A.R.; Havstad, S.; Lin, D.L.; Levan, S.; Fadrosh, D.; Panzer, A.R.; LaMere, B.; Rackaityte, E.; Lukacs, N.W.; et al. Neonatal gut microbiota associates with childhood multi–sensitized atopy and T-cell differentiation. Nat. Med. 2016, 22, 1187–1191. [Google Scholar] [CrossRef]
- Olivry, T.; Mueller, R.S. Critically appraised topic on adverse food reactions of companion animals (9): time to flare of cutaneous signs after a dietary challenge in dogs and cats with food allergies. BMC Vet. Res. 2020, 16, 158. [Google Scholar] [CrossRef] [PubMed]
- Pin, D.; Pendaries, V.; Alassane, KS.; Froment, C.; Amalric, N.; Cardiergues, M.; et al. Refined immunochemical characterization in healthy dog skin of the epidermal cornification proteins, filaggrin, and corneodesmosin. J Histochem Cytochem. 2019, 67, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Paixão, A.; Caldeira, J.; Leocádio, J.; Martins, L. A importância da integridade da barreira cutânea na prevenção da alergia veterinária – The importance of skin barrier integrity for the prevention of veterinary allergy. Rev Port Imunoalergologia. 2022, 30, 9–20. [Google Scholar] [CrossRef]
- Masuda, K.; Tsujimoto, H.; Fujiwara, S.; Kurata, K.; Hasegawa, A.; Yasueda, H.; Yamashita, K.; DeBoer, D.J.; de Weck, A.L.; Sakaguchi, M. IgE sensitivity and cross-reactivity to crude and purified mite allergens (Der f 1, Der f 2, Der p 1, Der p 2) in atopic dogs sensitive to Dermatophagoides mite allergens. Vet Immunol Immunopathol. 1999, 72, 303–13. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Caldas, E.; Iraola Calvo, V. Mite allergens. Curr Allergy Asthma Rep. 2005, 5, 402–10. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Lee, J.; Jeong, K.Y.; Cheon, D.S.; Park, J.W. Comparison of sensitization patterns to dust mite allergens between atopic dermatitis patients and dogs, and non-specific reactivity of canine IgE to the storage mite Tyrophagus putrescentiae. Exp Appl Acarol. 2022, 88, 41–55. [Google Scholar] [CrossRef]
- Weber, R.W. Patterns of pollen cross-allergenicity. J Allergy Clin Immunol. 2003, 112, 229–39. [Google Scholar] [CrossRef] [PubMed]
- Buckley, L.; Schmidt, V.; McEwan, N.; Nuttall, T. Cross-reaction and co-sensitization among related and unrelated allergens in canine intradermal tests. Vet Dermatol. 2013, 24, 422–7. [Google Scholar] [CrossRef] [PubMed]
- Li, J.D.; Gu, J.Q.; Xu, Y.Y.; Cui, L.; Li, L.S.; Wang, Z.X.; Yin, J.; Guan, K. Serum IgE profiles in Chinese pollinosis patients with grass pollen sensitisation. World Allergy Organ J. 2022, 11, 100624. [Google Scholar] [CrossRef]
- Martins, L.M.; Marques, A.G.; Pereira, L.M.; Goicoa, A.; Semião-Santos, S.J. , Bento, O. P.; House-dust mite allergy: mapping of Dermatophagoides pteronyssinus allergens for dogs by two-dimensional immunoblotting. Postepy Dermatol Alergol. 2015, 32, 73–81. [Google Scholar] [PubMed]
- Martins, L.M.; Marques, A.G.; Pereira, L.M.; Semião-Santos, S.J.; Bento, O.P. Allergy to grass pollen: mapping of Dactylis glomerata and Phleum pratense allergens for dogs by two-dimensional immunoblotting. Postepy Dermatol Alergol. 2017, 34, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Bacher, P.; Hohnstein, T.; Beerbaum, E.; Röcker, M.; Blango, M.G.; Kaufmann, S.; Röhmel, J.; Eschenhagen, P.; Grehn, C.; Seidel, K.; et al. Human Anti-fungal Th17 Immunity and Pathology Rely on Cross-Reactivity against Candida albicans. Cell 2019, 176, 1340–1355.e15. [Google Scholar] [CrossRef] [PubMed]
- Xing, H.; Wang, J.; Sun, Y.; Wang, H. Recent Advances in the Allergic Cross-Reactivity between Fungi and Foods. J Immunol Res. 2022, 2022, 7583400. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Common dermatomycosis lesion observations in dog. . (a) Spots of alopecia; (b) Pruritic inflammatory spots of alopecia (in previously clipped area); (c) Pruritic alopecic and depigmented spot; (d) Surrounding hair debris with refringent spores (400x).
Figure 1.
Common dermatomycosis lesion observations in dog. . (a) Spots of alopecia; (b) Pruritic inflammatory spots of alopecia (in previously clipped area); (c) Pruritic alopecic and depigmented spot; (d) Surrounding hair debris with refringent spores (400x).
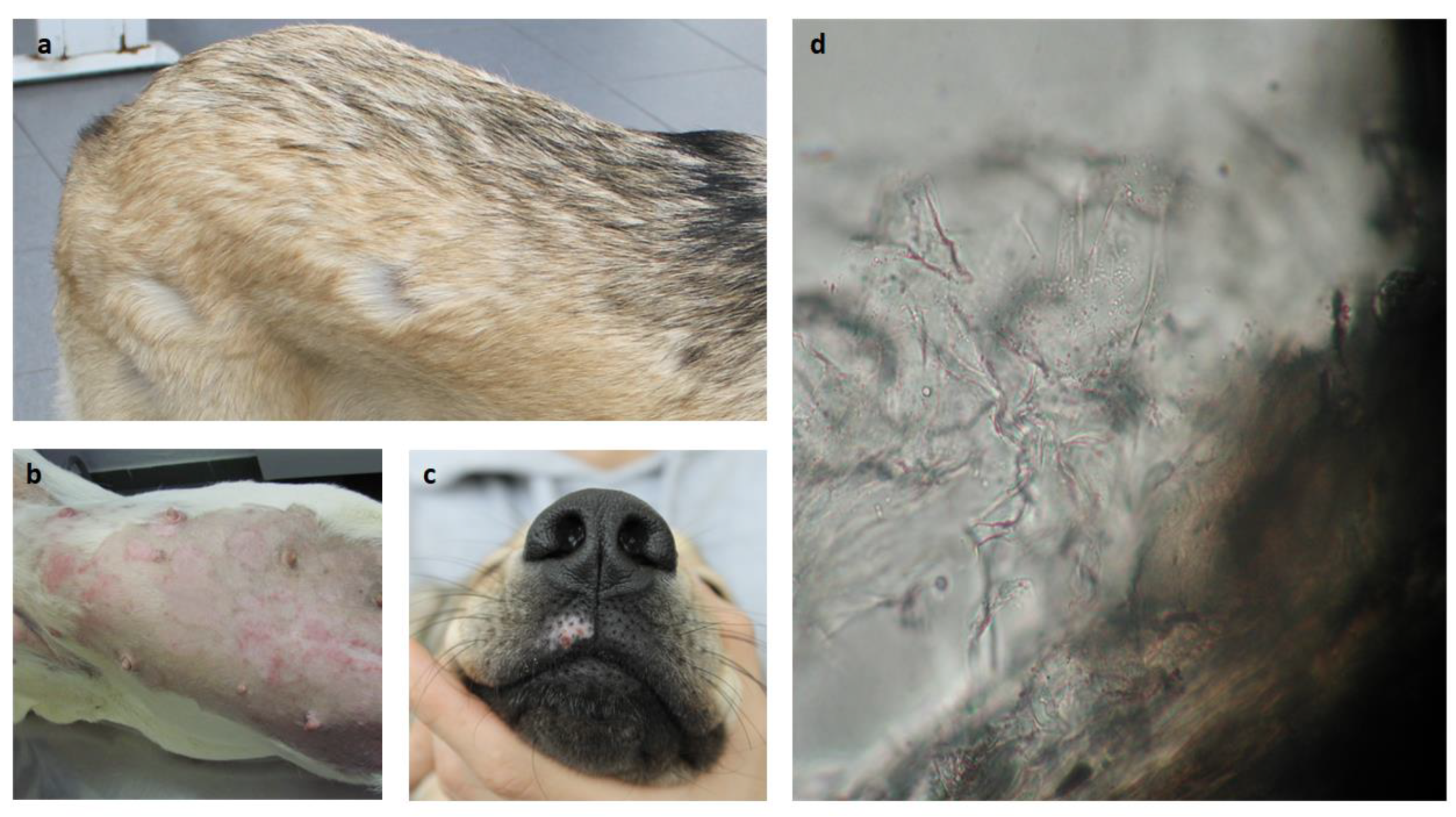
Figure 2.
Common IDT reactions in a 0–4 score range. (a) Negative control; (b) Positive control; (c) Phleum pratense; (d) Dactylis glomerata; (e) Malassezia pachydermatis; (f1-6) House dust and storage mites.
Figure 2.
Common IDT reactions in a 0–4 score range. (a) Negative control; (b) Positive control; (c) Phleum pratense; (d) Dactylis glomerata; (e) Malassezia pachydermatis; (f1-6) House dust and storage mites.
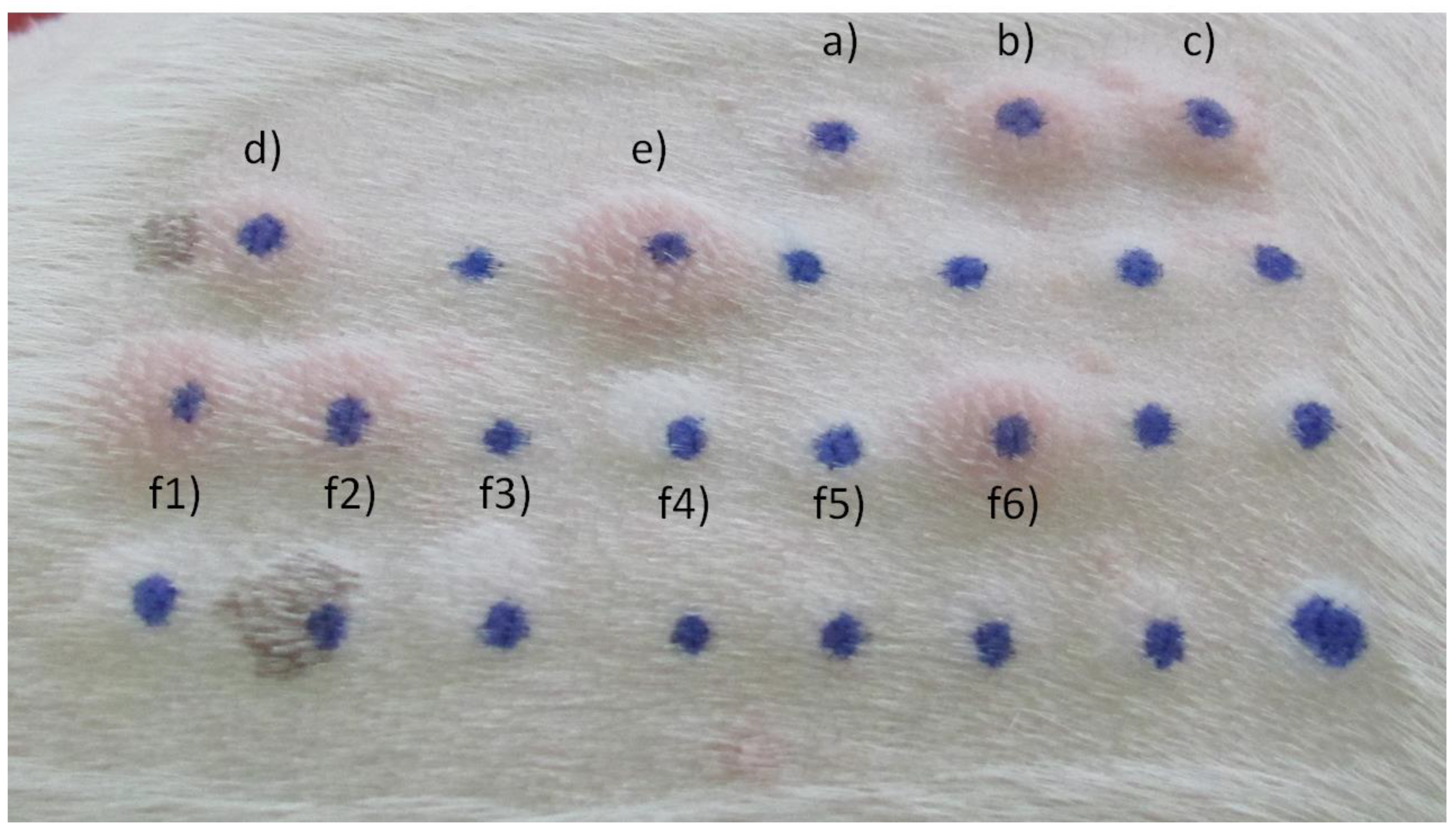
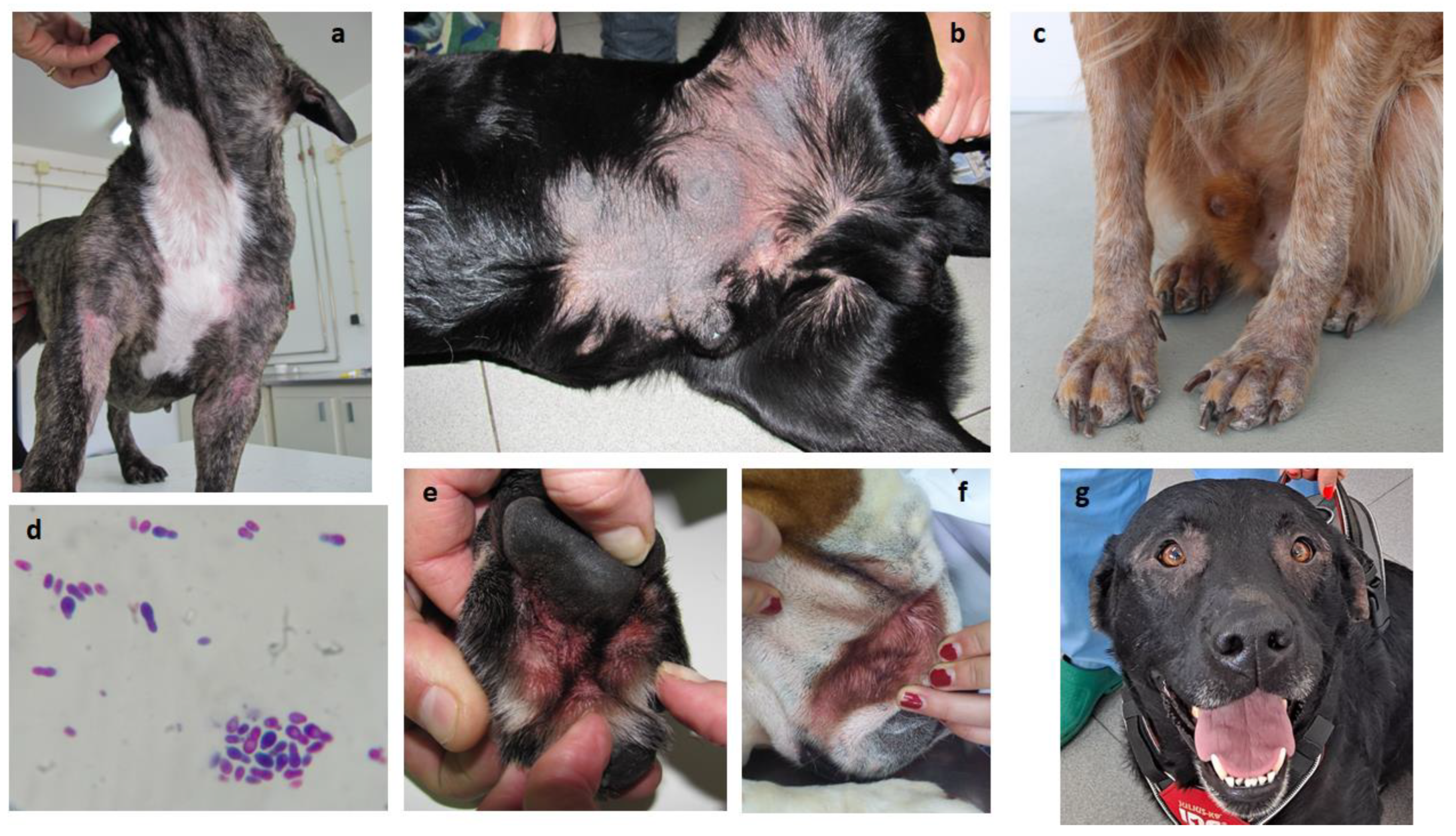
Figure 3.
Pruritic dermatitis in allergic dogs presenting Malassezia overgrowth. (a) Common areas of alopecic acute allergic dermatitis; (b) Hyperkeratosis and lichenification in chronic dermatitis; (c) Alopecia and early hyperkeratosis; (d) Malassezia overgrowth in skin cytology (400x); (e) Chronic interdigital dermatitis; (f) Facial intertriginous dermatitis; (g) Periocular alopecia in allergic dermatitis.
Figure 3.
Pruritic dermatitis in allergic dogs presenting Malassezia overgrowth. (a) Common areas of alopecic acute allergic dermatitis; (b) Hyperkeratosis and lichenification in chronic dermatitis; (c) Alopecia and early hyperkeratosis; (d) Malassezia overgrowth in skin cytology (400x); (e) Chronic interdigital dermatitis; (f) Facial intertriginous dermatitis; (g) Periocular alopecia in allergic dermatitis.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



