
Submitted:
24 July 2023
Posted:
24 July 2023
You are already at the latest version
Abstract
Trichinella spiralis (T. spiralis), a nematode parasite, is the major cause of Trichinellosis, a zoonotic disease. A key role of MAPR in the reproductive system is to maintain pregnancy. Previous studies found antihormone (P4 and RU486) drug design and vaccine therapy of recombinant protein (rTs-MAPRC2) to control T. spiralis infection. The current study investigates the inhibitory effects of different ratios of antibody against Ts-MAPRC2 on the development of muscle larvae (ML) and newborn larvae (NBL). First, we performed indirect immunofluorescence assays (IIFA) and examined the effects of rTs-MAPRC2-Ab on ML and NBL in vitro as well as in vivo. After-ward, siRNA-Ts-MAPRC2 was transfected into T. spiralis muscle larvae. Following that, Ts-MAPRC2 protein was detected by Western Blotting and mRNA levels were determined by qPCR. Also, we assessed whether siRNA-treated NBLs were infective by analyzing muscle larvae burden (MLs). Our Result showed rTs-MAPRC2-Ab greatly inhibited the activity of the Ts-MAPRC2 gene in ML and NBL of T. spiralis. rTs-MAPRC2-Ab reduced larval infectivity and survival in the host in a dose-dependent manner (1:50, 1:200, 1:800 dilutions). Further, siR-NA-Ts-MAPRC2 effectively silenced the Ts-MAPRC2 gene in muscle larvae (ML) by in vitro, as well as in new-born larvae (NBL) of T. spiralis by in vivo. In addition, siRNA-Ts-MAPRC2 (siR-NA180, siRNA419, siRNA559) reduced host larval survival and infectivity significantly. The effect of siRNA on larval development, survival, and infectivity was significantly reduced when the Ts-MAPRC2 gene was knocked down. This study, therefore, suggests that Ts-MAPRC2 might be a novel molecular target that might be useful in the development of vaccines against T. spiralis infection.
Keywords:
inhibitory effect
; knockdown
; rTs‐MAPRC2‐Ab
; Trichinella spiralis
; worm burden
1. Introduction
Trichinellosis is a zoonotic disease that is mainly caused by Trichinella spiralis (T. spiralis), a nematode parasite [1]. Mostly infection source with T. spiralis in humans is pork, and pork byproducts or undercooked meat are often eaten raw or undercooked [2,3,4]. China has a high morbidity rate linked to this disease because of the prevalence of naturalized animal reserves and the consumption of pork and pork products in the country [5,6,7]. The primary means of survival of T. spiralis nematodes is their ability to transmit directly from host to host. The immune system and normal cellular functions are adjusted to these infections at all stages [8,9]. Despite the widespread use of antihelminthic agents against Trichinellosis, excessive usage leads to drug residues in meat, parasite resistance, and other environmental issues. To prevent the spread of Trichinellosis, it is critical to developing a vaccine that is effective for humans and pigs [7,10]. There have recently been discoveries of proteins that inhibit parasite viability, inhibit parasite invasion, and, thus, reduce resistance to parasite infection. Additionally, their resistance to T. spiralis larvae inoculation has been studied [11,12,13,14]. In most cases, these vaccines have shown some success against T. spiralis infection. To date, no vaccine equivalent to the one currently available for the T. spiralis infection currently exists [1]. A group of membrane-associated progesterone receptor (MAPR) proteins known as progesterone receptor membrane component 1 (PGRMC-1), as well as progesterone receptor membrane component 2 (PGRMC-2), belong to the same family [15,16,17,18]. Similarly, several studies reported the presence of PGRMC receptors, progesterone-induced proteins, progesterone receptor-associated proteins (p48 protein), and small androgen receptor-interacting proteins in S. japonicum [19,20]. Previously, we characterized and cloned the membrane-associated progesterone receptor component-2 (Ts-MAPRC2) gene from T. spiralis. As part of the process, several experiments were conducted, including expression, purification, immunoblotting, binding ability against progesterone antibody, and immunofluorescence assay (IFA). Additionally, we evaluated the direct effects of progesterone (P4) and mifepristone (RU486) on Ts-MAPRC2 gene expression using in vitro cell culture tests, which revealed that expression levels varied at each stage of development (muscle larvae, female adult worms, male adult worms, and newborn larvae). After that, the in vivo phenotypic effects and relative mRNA effects of mifepristone were assessed in relation to the F-AL stage [21,22,23]. Small interfering RNA (siRNA) is double-stranded RNA and carries a sequence of 21 to 25 nucleotides. In C. elegans, these small RNAs were found [24]. The siRNAs are synthesized artificially and are used to study host-parasite interactions. In fact, they negatively modulate gene expression. In the last decade, scientists have begun to study siRNA synthesized from double-stranded RNA to silence gene expression and are using siRNA as an effective tool for studying and identifying gene functions [25,26]. It can be used to silence and suppress genes necessary for the development of parasites and the molting processes of the T. spiralis, thus reducing parasite attacks on humans and other mammals [25,27]. In the current study, we investigate the inhibitory effect of rTs-MAPRC2-Ab ratios in ML and NBL stages. Afterward, we performed indirect immunofluorescence assays (IIFA) at the ML and NBL stages. We also investigated the in vitro effects of rTs-MAPRC2-Ab on ML and NBL. Further, we examined the infectivity of ML and NBL treated with rTs-MAPRC2-Ab in vivo. This study also focuses on preparing different groups treated with siRNA-Ts-MAPRC2 (siRNA-180, siRNA-419, siRNA-559, and siRNA-control, along with PBS control), and its transfection into T. spiralis muscle larvae. Afterward, we performed the Western blotting of the Ts-MAPRC2 protein as well as the quantification of mRNA expression for Ts-MAPRC2. Additionally, we conducted an in vitro phenotyping study on siRNA effects on ML stages. Moreover, we evaluated the infectivity of NBLs treated with siRNA by examining the muscle larvae burden (MLs).
2. Materials and Methods
2.1. Preservation of Animals and Parasites
Sprague-Dawley rats (body weight 220–250 g) and BALB/c mice (body weight 18–20 g) were bought from Qinglongshan Animal Breeding Farm in Nanjing, Jiangsu, China (SXK 2008–0004). At the Animal House of Nanjing Agricultural University, the animals were maintained under supervised conditions. In Nanyang, Henan Province, China, T. spiralis (ISS534) was isolated from a pig and nurtured in BALB/c mice using serial passage after every six to eight months. A standardized HCl-pepsin digestion protocol was used by 40 days’ post-infection to recover T. spiralis muscle larvae (ML) from BALB/c mice [28]. In this study, adult worms (AL) were collected at 6 days’ post-ingestion, and newborn larvae (NBL) were recovered from an adult female (F-AL) at 6 days’ post-ingestion from an RPMI-1640 culture media at 37 °C for 24 hours as reported previously [29,30]. Several development phases of parasites were collected and homogenized in liquid nitrogen.
2.2. Molecular cloning of Ts-MAPRC2
Among its 234 amino acids, Ts-MAPRC2 (705 bp) has a full-length sequence of 705 bases. In the current study, a fragment with a size of 97–234 amino acids (225–705bp) was expressed from the Ts-MAPRC2 with conserved domain (104–173 amino acids). Gene Ts-MAPRC2 in T. spiralis was amplified using qRT-PCR techniques. To clone our gene, we used specific primers (5′-GAATTC AATAGATTTCGTATAAAATGGACATCT-3′) and (5′-AAGCTT TCACTGATCATCAACATCACAATCAGAG-3′) with restriction enzymes EcoR I and Hind III. PCR products were purified with Gel-Extraction Kits (Omega, Biotech, Norcross, GA, USA) and ligated into a pMD19-T cloning vector (TaKaRa, China). Further processing was performed on the recombinant plasmid (pMD19-T/Ts-MAPRC2) and the strain was converted into E. coli strain (DH5α) (Invitrogen, China) and cultured in LB (Luria Bertani) [31] medium containing ampicillin (100 μg/mL). Through the digestion of restriction enzymes and sequencing (Invitrogen, China), the presence of positive bacteria (pMD19-T/Ts-MAPRC2) clones was assured.
2.3. rTs-MAPRC2 protein development
As part of the restriction digestion, plasmid (pMD19-T/ Ts-MAPRC2) was digested with EcoRI and Hind III enzymes and then cloned into the prokaryotic expression vector pET-32a (+) (Novagen, Beijing, China). In subsequent experiments, the recombinant plasmid (pET32a (+)/Ts-MAPRC2) was carried out in BL21 (DE3) cells with 1 mM IPTG (Isopropyl-β-D-thiogalactopyranoside; Sigma-Aldrich, Shanghai, China). Moreover, lysozyme (10 mg/mL) (Sigma-Aldrich, USA) was used to lyse the cells. SDS-PAGE (12% w/v) was used to confirm the sonicated outputs. We purified recombinant Ts-MAPRC2 (rTs-MAPRC2) using the His-TrapTM FF column as directed by the manufacturer (GE Healthcare, Piscataway, NJ, USA). Protein concentrations were determined using the PierceTM BCA-Protein Assay Kit (Thermo Scientific, Waltham, MA, USA). In order to identify the strategy for filtering and expression of the empty pET32a (histidine-tagged) protein, we followed the same protocol that was used to purify and express Ts-MAPRC2, and vector protein was used as a negative control. Photographs of the rTs-MAPRC2 purified protein SDS-PAGE were taken. For subsequent experiments, rTs-MAPRC2 protein stock was prepared and maintained at -80 °C [32].
2.4. Developing an anti-rat polyclonal antibody against rTs-MAPRC2
SD rats (n = 6) were immunized with equal amounts of rTs-MAPRC2 protein (300 μg) and Freund’s complete adjuvant (Sigma-Aldrich, Darmstadt, Germany) subcutaneously for preparation of antisera. In the second week, Freund’s Incomplete Adjuvant (Sigma-Aldrich) was used to inoculate the second dose of 200 μg rTs-MAPRC2 protein. As before, we administered the two booster injections at a one-week interval. For further experiments, serum samples were collected one week after the last dose and stored at -80 °C. Negative controls included serum collected from rats who were not treated (n = 3).In a previous study, we successfully characterized, cloned the Ts-MAPRC2 gene (480 bp), expressed it into pET-32a (expression vector), confirmed restriction enzyme digestion with EcoR I and Hind III, purified rTs-MAPRC2, prepared polyclonal antibodies, and performed immunoblot analysis [21,22].
2.5. Detection of the natural protein Ts-MAPRC2 by indirect immunofluorescence assay (IIFA)
As a first step, whole worms of ML and NBL were soaked in 5% BSA (Bovine serum albumin) to block nonspecific binding, followed by incubation with primary antibodies (rat antisera against rTs-MAPRC2), pET32a rat serum (control group) and PBS (control group) at 37 °C for 2 h (1:50, 1:200 and 1:800 dilution). For Ts-MAPRC2 native protein detection, different time hours (2h, 4h, 8h, and 24h) of ML were used. Each group of worms (ML, NBL) was washed with PBS and incubated with FITC-anti-rat IgG conjugate (1:100; Santa Cruz, Dallas, Texas, USA). After another wash in PBS, the complete worms (ML, NBL) were observed under an Olympus fluorescent microscope (Tokyo, Japan) [11,33].
2.6. In vitro phenotyping of rTs-MAPRC2-Ab effects on ML and NBL
At the ML and NBL stages, the selected concentration rTs-MAPRC2-Ab ratios (1:50, 1:200, and 1:800 dilution) were used in conjunction with controls pET32a rat serum (control group) and PBS (control group). A 6-well precondition was used with 2000 worms/well of both stages (ML, NBL). RPMI-1640, 10% heat-inactivated FBS (Fetal Bovine Serum), and 2% antibiotics (100 U/mL penicillin and 100 mg/mL streptomycin) (Gibco, Paisley, UK) were used as the culture medium, which was incubated at 37 °C and 5% CO2. ML stage parasites were tested at various time intervals (0h, 4h, 8h,24h) for motility and ecdysis (molting process). NBL motility was checked after 24 hours. An inverted bright field microscope (Olympus, Shibuya, Japan) was used to examine the phenotypic appearance at both stages (ML and NBL) [21].
2.7. In vivo assessment of the infectivity of ML treated with rTs-MAPRC2-Ab
A total of fifty BALB/c mice (body weight 18–20 g) were acquired at Qinglongshan Animal Breeding Farm in Nanjing, Jiangsu, China (SXK 2008–0004) and kept in animal housing at Nanjing Agricultural University. In order to determine the infectivity of effective rTs-MAPRC2-Ab T. spiralis, mice were randomly divided into five groups (n = 10 per group). To find out the infectivity of ML and efficacy of rTs-MAPRC2-Ab, three experimental groups (1:50, 1:200 and 1:800 dilution), one pET32a rat serum (control group) and one PBS (control group) were infected orally with 500 MLs treated with rTs-MAPRC2-Ab (1:50, 1:200 and 1:800 dilution), pET32a rat serum (control group) and PBS (control group) respectively. A total of five mice from each group were euthanized six days’ post-infection (6dpi) via head dislocation. Adult worms were collected from the mice intestines using the method discussed above, and they were counted as follows: adult female (F-AL) length, adult male (M-AL), and total adult worm (AL) [21]. The number of NBL was also assessed and collected by culturing 50 adult worms per group in RPMI-1640 with FBS (20%), penicillin (500 units/mL), and streptomycin (500 mg/mL) for 24 hours at 37˚C with 5% CO2. MLs were collected from all five remaining mice in each group by digestion method after euthanasia at 35 DPI [34].
2.8. Assessment of the infectivity of NBL treated with rTs-MAPRC2-Ab
Thirty mice were divided into 5 groups (n = 6 per group) and injected through tail vein injection with 20000 NBLs treated with rTs-MAPRC2-Ab (1:50, 1:200 and 1:800 dilution), pET32a rat serum (control group) and PBS (control group) respectively. The mice of every group were euthanized by head dislocation at 35 DPI, and the MLs were collected by artificial digestion as described above. Based on treatment with rTs-MAPRC2-Ab (1:50, 1:200, and 1:800 dilution), and in comparison, to controls pET32a rat serum (control group) and PBS (control group), ML reductions were calculated [34].
2.9. Preparation of siRNA
To design siRNA sequences, full-length cDNA encoding Ts-MAPRC2 (705 bp) was used (Gene Bank accession XM_003375886.1) [35]. This study used Ts-MAPRC2-specific siRNA oligos created by Gene Pharma (Shanghai, China) (Stealth TM RNAi duplexes). As shown in Table 1, the sequences of these three siRNAs and control siRNAs were used in this study.

Table 1.
The siRNAs used in the study.
| siRNA name | Sense (5′–3′) | Antisense (5′–3′) |
|---|---|---|
| siRNA-180 | 5’-CUGGGAUUCUUGCGGUAAUTT-3 | 5’-AUUACCGCAAGAAUCCCAGTT-3 |
| siRNA-419 | 5’-GGUGGACCAUAUGGCUUAUTT-3’ | 5’-AUAAGCCAUAUGGUCCACCTT-3’ |
| siRNA-559 | 5’- GGCUAUGCAUGAGCUGAAATT-3’ | 5’-UUUCAGCUCAUGCAUAGCCTT-3’ |
| siRNA-Control | 5’-UUCUCCGAACGUGUCACGUTT-3’ | 5’- ACGUGACACGUUCGGAGAATT-3’ |
2.10. Transfection of siRNA to T. spiralis Worms
The soaking method was used to knock down Ts-MAPRC2 gene expression [25,26]. The muscle larvae were cultured in RPMI-1640 (supplemented with penicillin 500 units/mL and streptomycin 500 mg/mL) with siRNA-180, siRNA-419, and siRNA-559 in 12-well culture plates (500 MLs/500 ul) at 37 °C with 5% CO2. After 36 hours, 20% FBS was added and incubated for another 24 hours. As controls, we used PBS and siRNA-Control. After incubation with Lipofectamine 2000 Reagent (Invitrogen, US) for 20 minutes, control siRNA or siRNA-Ts-MAPRC2 were added to larvae in a final concentration of 2 μM [36].
2.11. Protein expression of Ts-MAPRC2 as determined by Western blotting
In order to observe the effect of siRNA on the expression of Ts-MAPRC2, worms were harvested after 3 days of incubation. Using RIPA solution (Thermo Scientific, Waltham, MA, USA) and a proteinase inhibitor (Thermo Scientific, Waltham, MA, USA) for complete lysis, the total soluble protein of T. spiralis larvae (muscle larvae) parasites were collected from all treated groups (siRNA-180, siRNA-419, siRNA-559, siRNA-Control, and PBS control). The total soluble protein of T. spiralis, was collected by high-speed centrifugation at 12,000 r/min for 30 minutes at 4 °C, and the supernatant was analyzed by the bicinchoninic acid method (BCA) kit previously described [32]. After that, the total soluble proteins of the T. spiralis (muscle larvae) parasites were separated by SDS-PAGE (12%) and transferred to polyvinylidene difluoride (PVDF) membranes (Millipore, Bedford, MA, USA). A subsequent block of nonspecific binding was performed with 5% skim milk in Tris-buffered saline containing 0.1% Tween-20 (TBST). The membranes were washed three times with TBST before incubation with the primary antibody (rat antisera against rTs-MAPRC2,1:300 dilution) at 4 °C overnight. Afterward, the membranes were washed three times and incubated for 1 h at 37 °C with HRP-conjugated goat-antirat IgG (diluted 1:5000 in TBST). Additionally, a mouse antibody against GAPDH (1:1000) (Proteintech, US) was used as a quantitative protein control to detect GAPDH expression. A secondary antibody was made up of goat anti-mouse IgG (1:5000) HRP-conjugated. To detect bound antibodies, the Tanon™ High-sig ECL Western Blotting Substrate kit was used as instructed by the manufacturer [37].
2.12. Expression of Ts-MAPRC2 mRNA by RNA extraction and qPCR
In brief, the Trizol method was used to extract RNA from ML worms. A prime script RT reagent kit (Takara, CA, USA) was used for RNA extraction from siRNA-treated groups (siRNA-180, siRNA-419, siRNA-559, siRNA-Control, and PBS control) of ML worms. In order to reverse transcribe RNA isolated from each group at the ML stage, HiScript II Q RT SuperMix kit (Vazyme, Nanjing, China) was used. To analyze the Ts-MAPRC2 gene, the following primers were used: forward (5′-GCTGGGATTCTCGGAATAATGT-3′) and reverse (5′-CGGTGGACAATCTCCTGACAGTTG-3′). BI 7500 Fast Real-time PCR System (Applied Biosystems, USA) and Cham-QTM SYBR RT-qPCR master mix-Kit (Vazyme, Nanjing, China) were used for quantitative amplification. As an internal control, GenBank Accession No. AF452239 GADPH (Glyceraldehyde-3-phosphate dehydrogenase) of Trichinella was used. GADPH primers were designed as follows: forward (5′-GTCGTGGCTGTGAATGATC-3′) and reverse (5′-GCTGCCCCACTTAATTGCTT-3′), and data were computed using the comparative Ct (2−ΔΔCt) method [38].
2.13. In vitro phenotyping of siRNA effects on ML
In 12-well culture plates (500 ML/500 ul) at 37 °C with 5% CO2, we cultured muscle larvae in RPMI-1640 (supplemented with penicillin 500 units/mL and streptomycin 500 mg/mL) with siRNA-180, siRNA-419, siRNA-559, siRNA-Control, and PBS control as described above [39]. ML stage parasites were tested for motility and ecdysis (molting process). An inverted bright field microscope (Olympus, Shibuya, Japan) was used to examine the phenotypic appearance [21].
2.14. Evaluation of siRNA-treated NBL for infectivity
Twenty-five mice were divided into five groups (n = 5 per group), and each group was infected using 20000 NBLs treated with siRNA-180, siRNA-419, siRNA-559, siRNA-Control, and PBS control as described above. At 35 DPI, the mice of each group were euthanized by head dislocation, and the MLs were collected by artificial digestion as described above. The reductions in MLs were calculated based on treatment with siRNAs (siRNA-180, siRNA-419, siRNA-559) compared to siRNA-Control and the PBS control [34].
2.15. Statistical Analysis
Analyzing the data using analytical statistics software (Statistix 8.1, 2003) included a one-way analysis of variance (ANOVA), followed by Tukey’s analysis and LSD (least significant difference). Standardized RT-qPCR data in Microsoft Excel 2010 (Redmond, Washington, USA) using 2-ΔΔCt. Triplicates of all tests were performed. These figures were created using Origin software (Origin Pro 2021, Origin Lab Corporation, Northampton, MA, USA). Data were expressed as mean ± SD (n = 3). p ≤ 0.05 was considered significant.
3. Results
3.1. Indirect immunofluorescence assay (IIFA) in Muscle larvae (ML) and new borne larvae (NBL)
Anti-rTs-MAPRC2 rat serum was tested in different concentrations (1:50, 1:200, and 1:800) and different time intervals (2h, 4h, 8h, and 24h) on the cuticle at ML stage with pET32a rat serum (control group) as well as PBS (control group), respectively (Figure 1). The results indicated that anti-rTs-MAPRC2 rat serum (1:50) was brighter than other concentrations (1:200, and 1:800). Time intervals 8h and 24 h lead to higher FITC fluorescence protein recognition. Furthermore, we also found native protein in new borne larvae (NBL) by using anti-rat sera against rTs-MAPRC2 at different concentrations (1:50, 1:100, and 1:800) after 24h in comparison with pET32a rat serum (control group) and PBS (control group), respectively (Figure 2). rTs-MAPRC2-Ab reduced larval infectivity and survival in the host in a dose-dependent manner (1:50, 1:200, 1:800 dilutions).
Figure 1.
Identification of native Ts-MAPRC2 protein on the surface of T. spiralis muscle larvae (ML) were analyzed by IIFT using anti-rat sera against rTs-MAPRC2 at different concentrations (1:50, 1:200, and 1:800) with different time intervals (2h, 4h, 8h, and 24h) compared with pET32a rat serum (control group) and PBS (control group) respectively. Scale bar = 200 μm.
Figure 1.
Identification of native Ts-MAPRC2 protein on the surface of T. spiralis muscle larvae (ML) were analyzed by IIFT using anti-rat sera against rTs-MAPRC2 at different concentrations (1:50, 1:200, and 1:800) with different time intervals (2h, 4h, 8h, and 24h) compared with pET32a rat serum (control group) and PBS (control group) respectively. Scale bar = 200 μm.
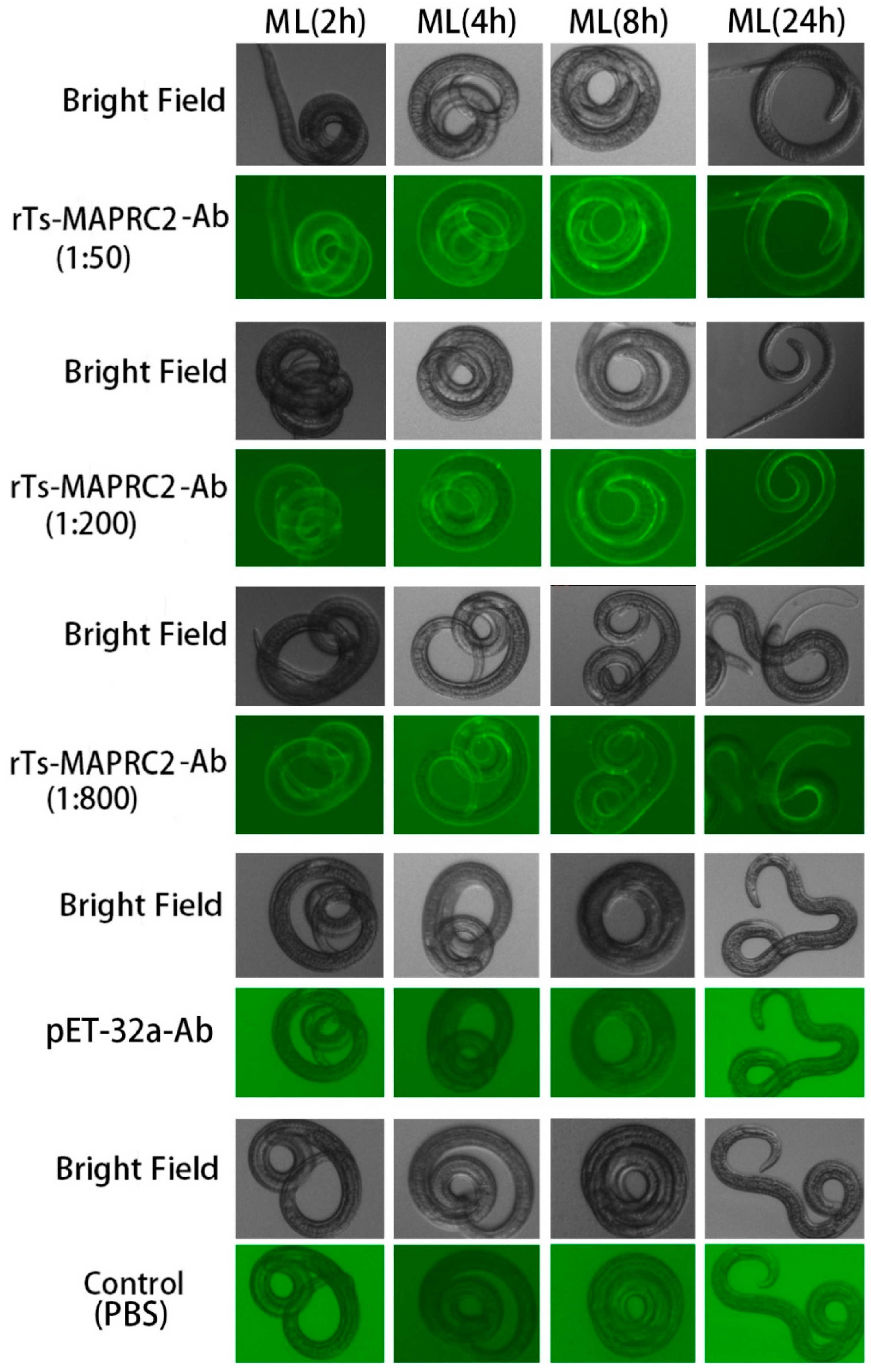
Figure 2.
Identification of native Ts-MAPRC2 protein in T. spiralis new borne larvae (NBL) were analyzed by IIFT using anti-rat sera against rTs-MAPRC2 at different concentrations (1:50, 1:200, and 1:800) after 24h compared with pET32a rat serum (control group) and PBS (control group) respectively. Scale bar = 200 μm.
Figure 2.
Identification of native Ts-MAPRC2 protein in T. spiralis new borne larvae (NBL) were analyzed by IIFT using anti-rat sera against rTs-MAPRC2 at different concentrations (1:50, 1:200, and 1:800) after 24h compared with pET32a rat serum (control group) and PBS (control group) respectively. Scale bar = 200 μm.
3.2. Effect of rTs-MAPRC2-Ab on ML and NBL in vitro
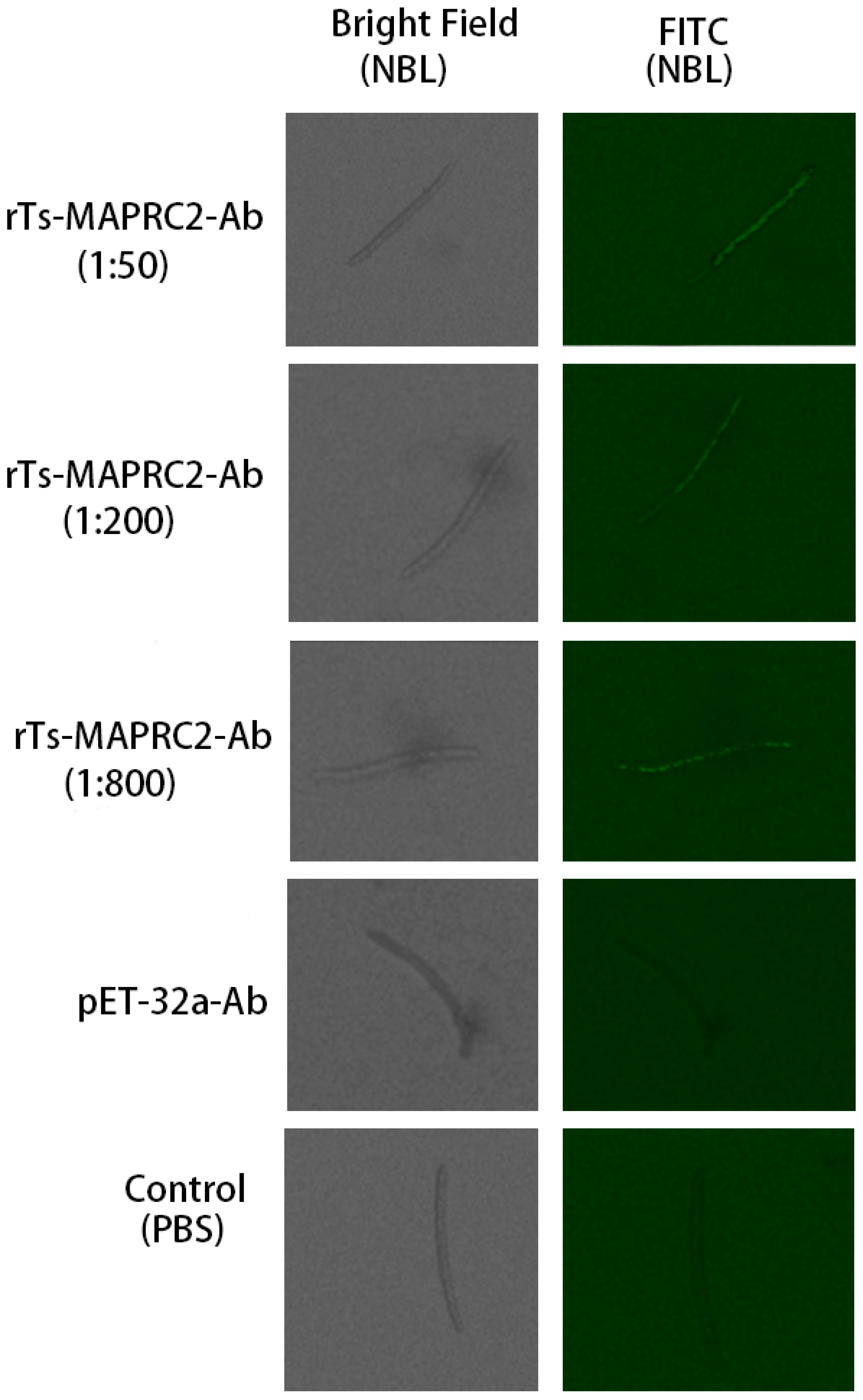
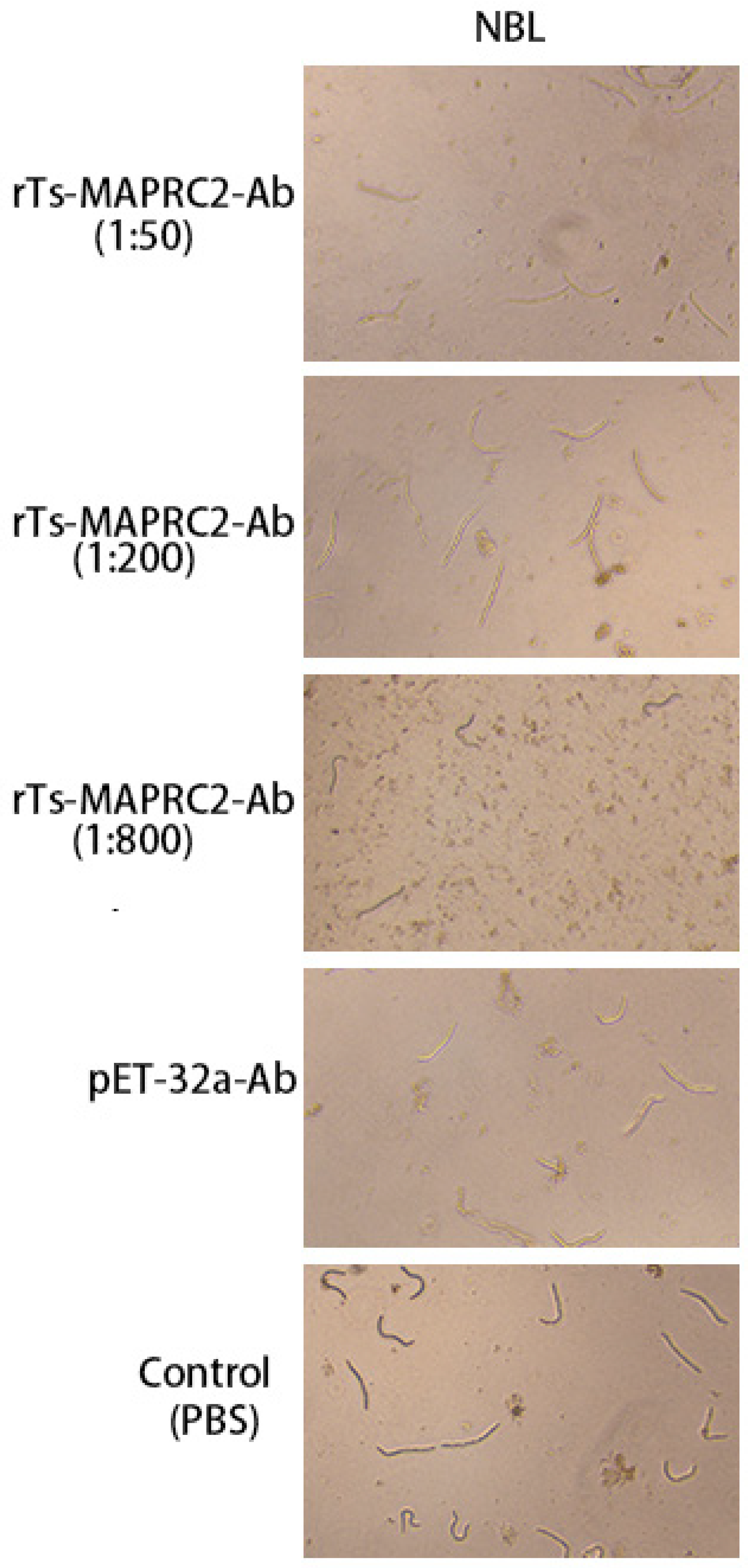
At the ML stage, the selected concentration rTs-MAPRC2-Ab ratios (1:50, 1:200, and 1:800 dilution) with various time intervals (0h, 4h, 8h,24h) were used in conjunction with controls pET32a rat serum (control group) and PBS (control group). At all-time intervals (0h, 4h, 8h, 24h) motility was present in ML stage. The comparison of ecdysis (molting process) showed that after 24 hours, the ecdysis process was slower in 1:50 (rTs-MAPRC2-Ab ratio) as compared to 1:200 and 1:800 (rTs-MAPRC2-Ab ratio). In both pET32a rat serum (control group) and PBS-control, the process of ecdysis is faster (Figure 3). In addition, we observed motility in new-born larvae (NBL) using anti-rat sera against rTs-MAPRC2, at different concentrations (1:50, 1:100, and 1:800) following 24h of treatment, when compared with pET32a rat serum (control group) and PBS (control group), respectively (Figure 4). The results showed that NBL worms were more motile in the control groups (pET32a-control and PBS-control). In 1:50 (rTs-MAPRC2-Ab ratio), motility was slower than at 1:200 and 1:800 (rTs-MAPRC2-Ab ratio).
Figure 3.
The selected concentration ratios of rTs-MAPRC2-Ab (1:50, 1:200, and 1:800) with varying time intervals (0h, 4h, 8h, 24h), in conjunction with controls of pET32a rat serum (control group) and PBS (control group), were used at the ML stage to observed ecdysis (molting process) and motility at objective 10×.
Figure 3.
The selected concentration ratios of rTs-MAPRC2-Ab (1:50, 1:200, and 1:800) with varying time intervals (0h, 4h, 8h, 24h), in conjunction with controls of pET32a rat serum (control group) and PBS (control group), were used at the ML stage to observed ecdysis (molting process) and motility at objective 10×.
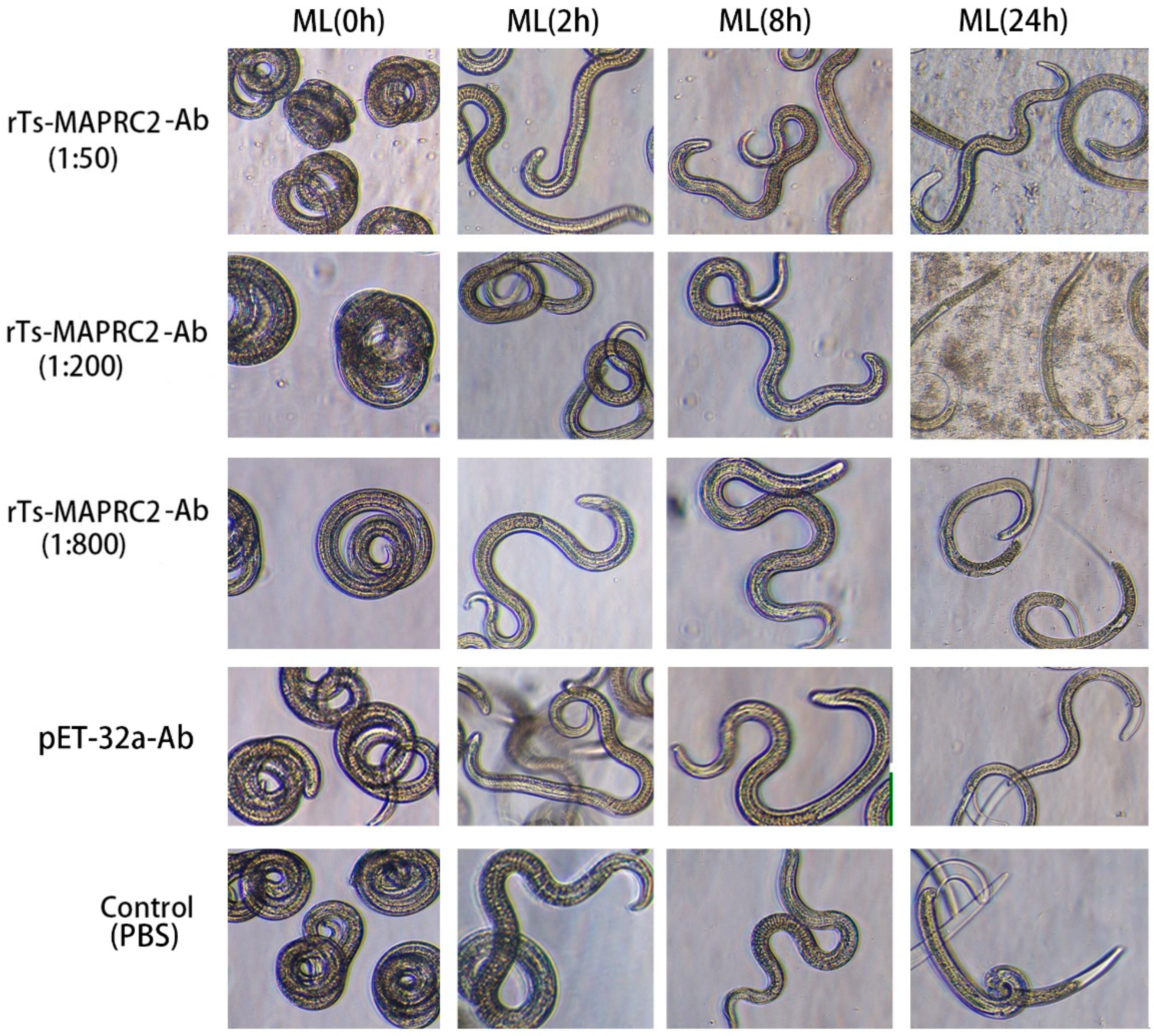
Figure 4.
The concentration ratio of rTs-MAPRC2-Ab after 24h, as well as controls of pET32a rat serum (control group) and PBS (control group), were used at the NBL stage to determine motility at objective 10×.
Figure 4.
The concentration ratio of rTs-MAPRC2-Ab after 24h, as well as controls of pET32a rat serum (control group) and PBS (control group), were used at the NBL stage to determine motility at objective 10×.
3.3. Assessing the infectivity of ML treated with rTs-MAPRC2-Ab in vivo
The effects of rTs-MAPRC2-Ab on the infectivity and efficacy of ML were tested by infecting 500 MLs orally treated with rTs-MAPRC2-Ab (1:50, 1:200 and 1:800 dilution), pET32a rat serum (control group) and PBS (control group), respectively. Using head dislocation, five mice from each group were euthanized at 6 days’ post-infection. Female adult worms collected at 6 days post-immunization from immunized with rTs-MAPRC2-Ab (1:50, 1:200 and 1:800 dilutions) were consistently shorter than rTs-MAPRC2-Ab-immunized from pET32a rat serum and PBS groups (p ≤ 0.05). Furthermore, the adult female length treated with anti-rTs-MAPRC2 rat serum (1:50) was smaller than the other concentrations (1:200, and 1:800) (p ≤ 0.05) (Figure 5A). (Figure 5B) None of the three groups (1:50, 1:200 and 1:800 dilution) of adult males showed statistically more significant variation in length compared with pET32a rat serum and PBS groups (p ≤ 0.05).
Figure 5.
Length of T. spiralis from mice treated with rTs-MAPRC2-Ab (1:50, 1:200 and 1:800 dilution), pET32a rat serum (control group) or PBS (control group) at six days post-infection (6dpi). (A) Female adult worms’ measurement (µm). (B) Male adult worms’ measurement (µm). Statistical data were presented as mean ± SD. p ≤ 0.05 was considered significant. Different letters mean significant and the same letters mean nonsignificant.
Figure 5.
Length of T. spiralis from mice treated with rTs-MAPRC2-Ab (1:50, 1:200 and 1:800 dilution), pET32a rat serum (control group) or PBS (control group) at six days post-infection (6dpi). (A) Female adult worms’ measurement (µm). (B) Male adult worms’ measurement (µm). Statistical data were presented as mean ± SD. p ≤ 0.05 was considered significant. Different letters mean significant and the same letters mean nonsignificant.
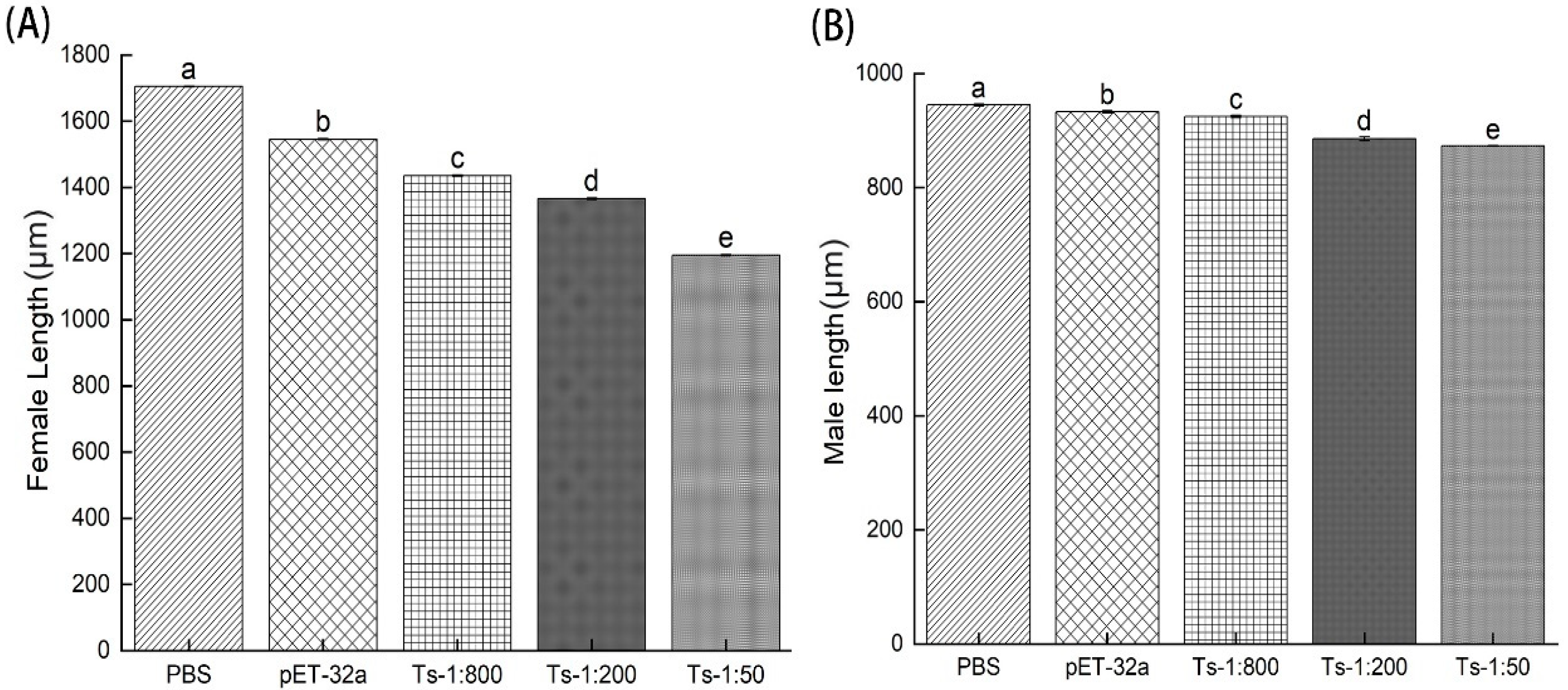
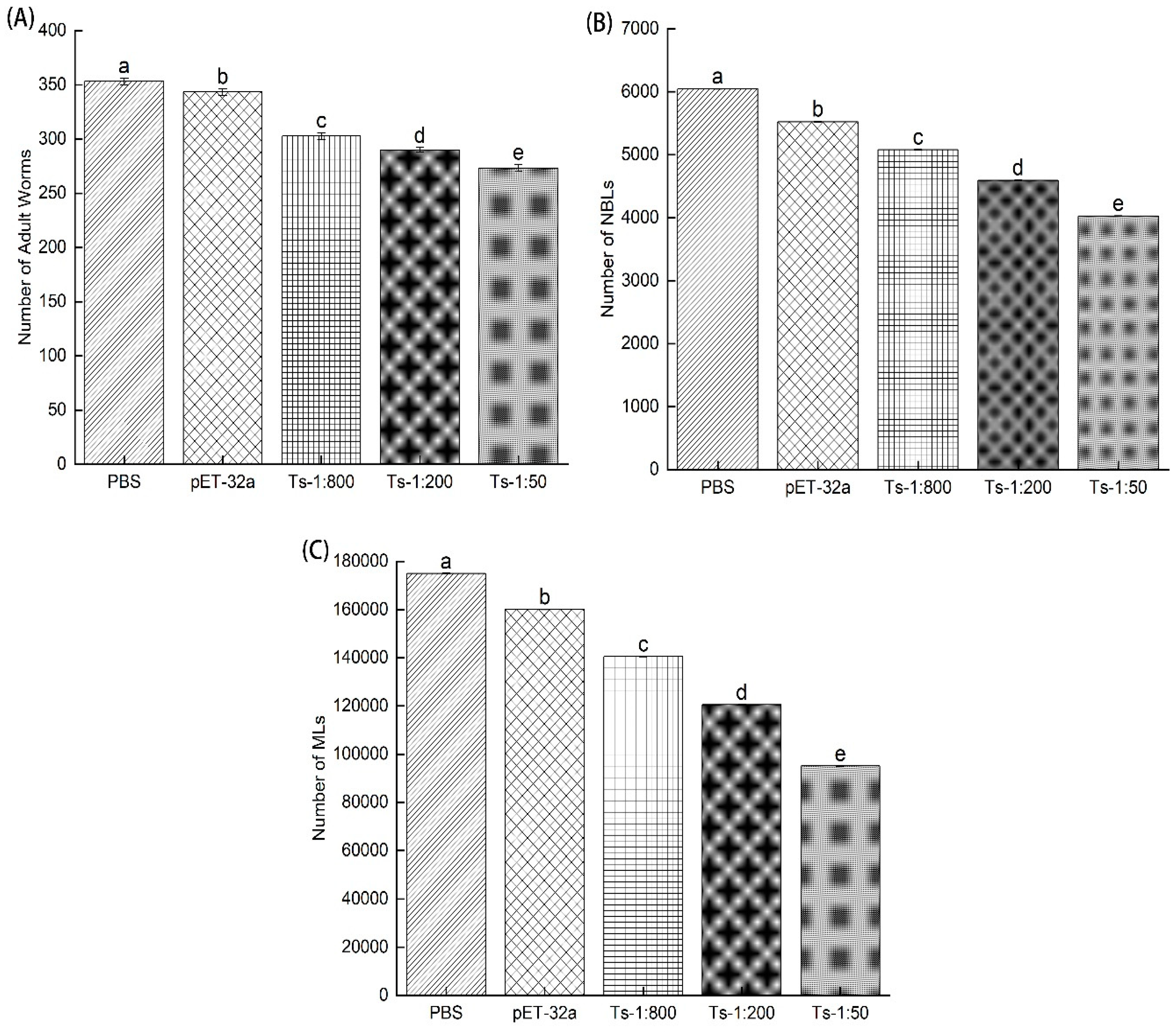
Figure 6. showed the count of the total adult worm after 6dpi, collection of total new borne larvae (NBL) after 24h and the total number of muscle larvae (MLs) after 35 days of post-infection (35dpi) treated with rTs-MAPRC2-Ab (1:50, 1:200 and 1:800 dilution), pET32a rat serum (control group) and PBS (control group), respectively. Figure 6A indicated that adult worm burden was less in 1:50 (rTs-MAPRC2-Ab ratio) than in 1:200 and 1:800 (rTs-MAPRC2-Ab ratio), as well as in comparison to pET32a rat serum (control group) and PBS (control group), respectively (p ≤ 0.05). Figure 6B showed the total number of newly borne larvae collected from adult worms due to in vitro culture after 24 hours. Results indicated that the total new borne larvae (NBLs) were lower in 1:50 (rTs-MAPRC2-Ab ratio) than in 1:200 and 1:800 (rTs-MAPRC2-Ab ratio), as well as in comparison with pET32a rat serum (control group) and PBS (control group), respectively (p ≤ 0.05). Figure 6C showed the total number of muscle larvae collected (MLs) after 35 days of post-infection (35dpi). According to the results, the total number of muscle larvae collected (MLs) was lower in 1:50 (rTs-MAPRC2-Ab ratio) compared to 1:200 and 1:800 (rTs-MAPRC2-Ab ratio), as well as compared to pET32a rat serum (control group) and PBS (control group), respectively (p ≤ 0.05).
Figure 6.
Results include the (A) number of adult worms collected after 6dpi, (B) the total number of new borne larvae collected after 24h, and (C) the total number of muscle larvae collected after 35 days of post-infection (35dpi) treated with rTs-MAPRC2-Ab (1:50, 1:200, and 1:800 dilution), pET32a rat serum (control group), and PBS (control group). Statistical data were presented as mean ± SD. p ≤ 0.05 was considered significant. Different letters mean significant and the same letters mean nonsignificant.
Figure 6.
Results include the (A) number of adult worms collected after 6dpi, (B) the total number of new borne larvae collected after 24h, and (C) the total number of muscle larvae collected after 35 days of post-infection (35dpi) treated with rTs-MAPRC2-Ab (1:50, 1:200, and 1:800 dilution), pET32a rat serum (control group), and PBS (control group). Statistical data were presented as mean ± SD. p ≤ 0.05 was considered significant. Different letters mean significant and the same letters mean nonsignificant.
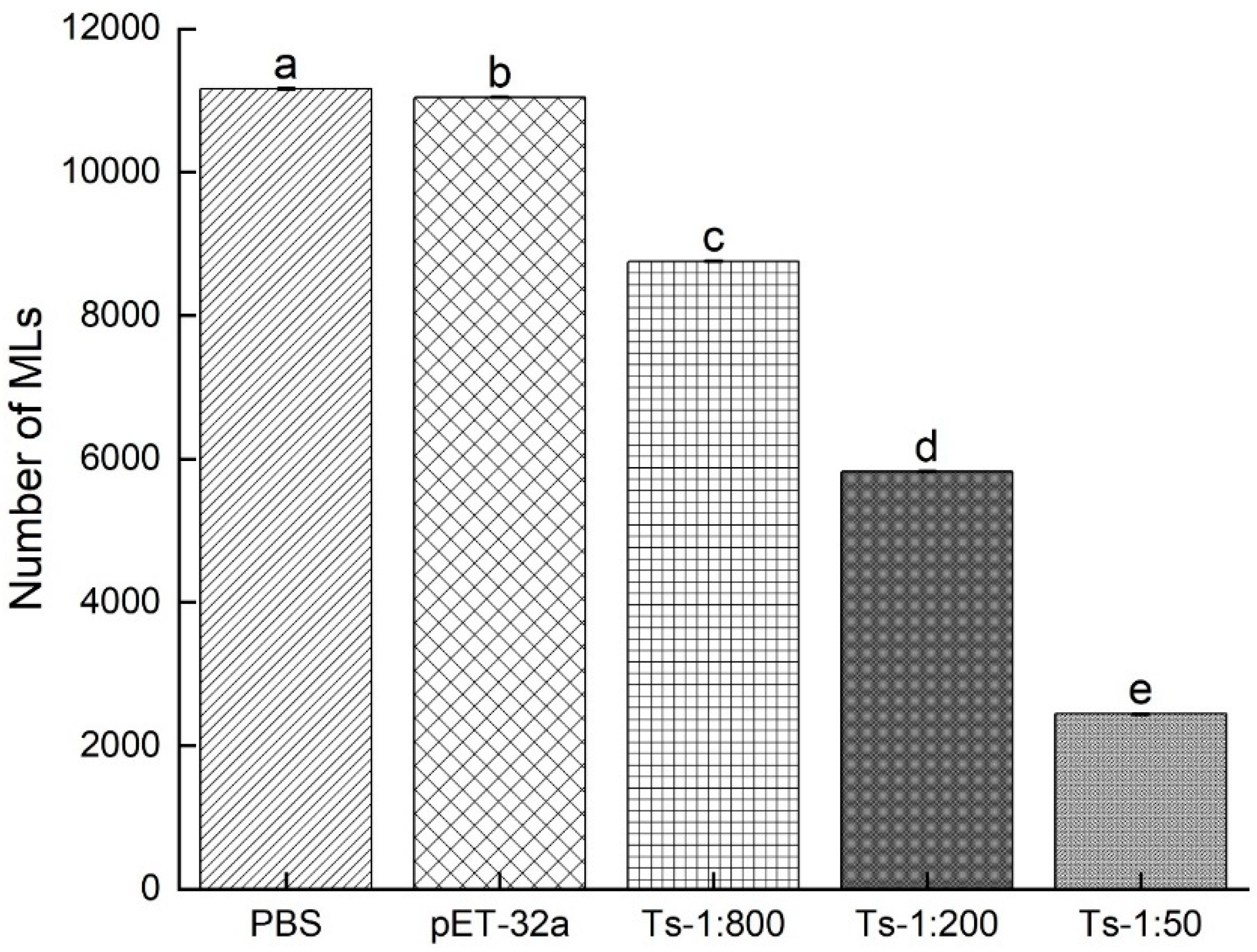
3.4. Infectivity of NBL treated with rTs-MAPRC2-Ab
The NBLs were treated with rTs-MAPRC2-Ab (1:50, 1:200, and 1:800 dilutions), pET32a rat serum (control group), and PBS (control group). The total number of muscle larvae (MLs) were counted at 35 days post-infection (DPI). As shown in Figure 7, in the results of the study, the total number of muscle larvae collected (MLs) was significantly lower in 1:50 (rTs-MAPRC2-Ab ratio) as compared to 1:200, 1:800, and pET32a rat serum (control group) and PBS (control group), respectively (p ≤ 0.05).
Figure 7.
The total number of muscle larvae collected after 35 days of post-infection (35dpi) treated NBL with rTs-MAPRC2-Ab (1:50, 1:200, and 1:800 dilution), pET32a rat serum (control group), and PBS (control group). Statistical data were presented as mean ± SD. p ≤ 0.05 was considered significant. Different letters mean significant and the same letters mean nonsignificant.
Figure 7.
The total number of muscle larvae collected after 35 days of post-infection (35dpi) treated NBL with rTs-MAPRC2-Ab (1:50, 1:200, and 1:800 dilution), pET32a rat serum (control group), and PBS (control group). Statistical data were presented as mean ± SD. p ≤ 0.05 was considered significant. Different letters mean significant and the same letters mean nonsignificant.
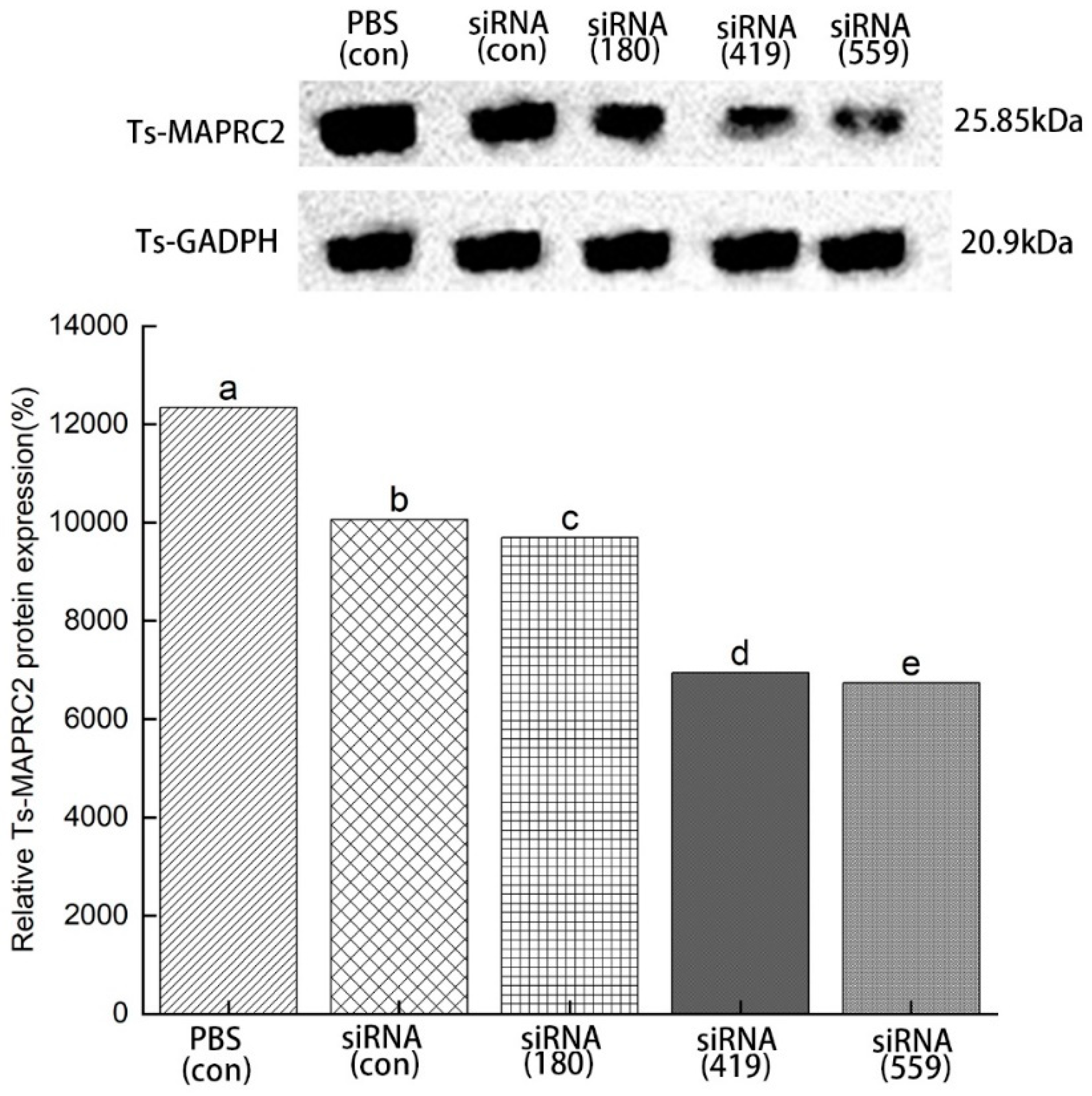
3.5. Specific siRNA-mediated suppression of Ts-MAPRC2 protein expression
ML were incubated in 2 μM siRNA- Ts-MAPRC2 of all treated groups (siRNA-180, siRNA-419, siRNA-559, siRNA-Control, and PBS control) for 3 days to determine the level of inhibition expression. The result showed siRNA-559 was found to have a greater inhibitory effect on Ts-MAPRC2 protein expression compared with siRNA-180 and siRNA-419. Comparing siRNA-559 and siRNA-419, siRNA-559 has more effect on silencing Ts-MAPRC2 protein. There was no more significant difference between control siRNA and control PBS (Figure 8) (p ≤ 0.05).
Figure 8.
Western blot analysis of Ts-MAPRC2 protein expression induced by siRNA. Western blot with anti-rTs-MAPRC2 antibodies showed the specific inhibition of Ts-MAPRC2 protein expression of T. spiralis muscle larvae (ML) with different siRNAs treated groups (siRNA-180, siRNA-419, siRNA-559, siRNA-Control, and PBS control) for 3 days. The tests were conducted in triplicate. Data was presented as the mean ± SD. p ≤ 0.05 was considered significant. Different letters indicate significance, while the same letters indicate nonsignificantly.
Figure 8.
Western blot analysis of Ts-MAPRC2 protein expression induced by siRNA. Western blot with anti-rTs-MAPRC2 antibodies showed the specific inhibition of Ts-MAPRC2 protein expression of T. spiralis muscle larvae (ML) with different siRNAs treated groups (siRNA-180, siRNA-419, siRNA-559, siRNA-Control, and PBS control) for 3 days. The tests were conducted in triplicate. Data was presented as the mean ± SD. p ≤ 0.05 was considered significant. Different letters indicate significance, while the same letters indicate nonsignificantly.
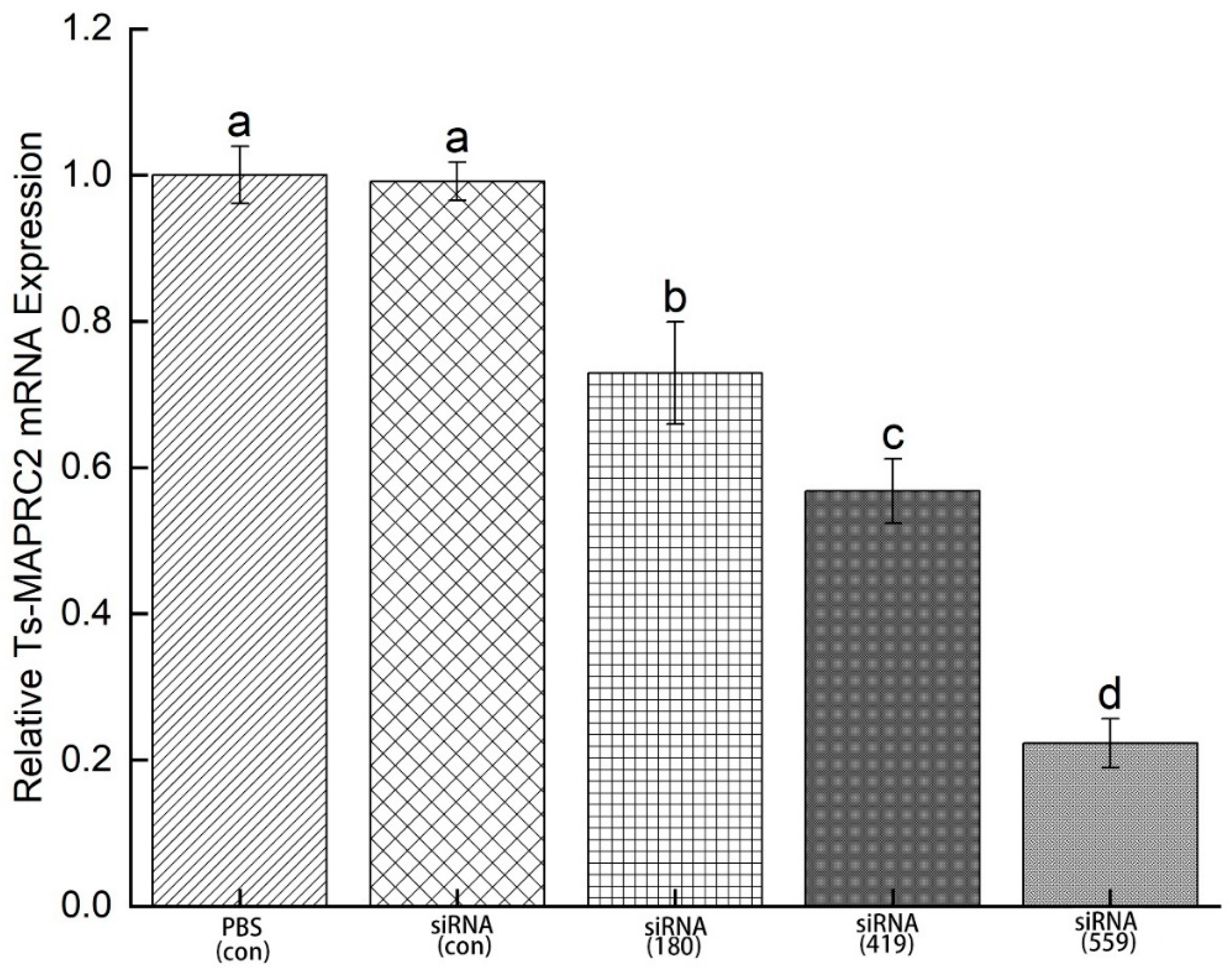
3.6. Specific siRNA-mediated suppression of Ts-MAPRC2 mRNA expression
In order to determine mRNA expression, ML were incubated in 2 μM siRNA- Ts-MAPRC2 of the treated groups (siRNA-180, siRNA-419, siRNA-559, siRNA-Control, and PBS control) for 3 days. Study results revealed that siRNA-559 had a greater downregulation of the Ts-MAPRC2 gene compared to siRNA-419 and siRNA-180. In comparison with siRNA-419, siRNA-559 has more downregulation of the Ts-MAPRC2 gene (p ≤ 0.05). Control siRNA and control PBS did not significantly differ in terms of mRNA expression (Figure 9) (p ≤ 0.05).
Figure 9.
mRNA expression of Ts-MAPRC2 gene induced by siRNAs. mRNA expression Ts-MAPRC2 gene of T. spiralis muscle larvae (ML) treated with different siRNAs (siRNA-180, siRNA-419, siRNA-559, siRNA-Control, and PBS control) for 3 days. Triplicates of the tests were performed. The data were presented as the mean ± SD. p ≤ 0.05 was considered significant. The same letter indicates insignificant results, but a different letter indicates significance.
Figure 9.
mRNA expression of Ts-MAPRC2 gene induced by siRNAs. mRNA expression Ts-MAPRC2 gene of T. spiralis muscle larvae (ML) treated with different siRNAs (siRNA-180, siRNA-419, siRNA-559, siRNA-Control, and PBS control) for 3 days. Triplicates of the tests were performed. The data were presented as the mean ± SD. p ≤ 0.05 was considered significant. The same letter indicates insignificant results, but a different letter indicates significance.
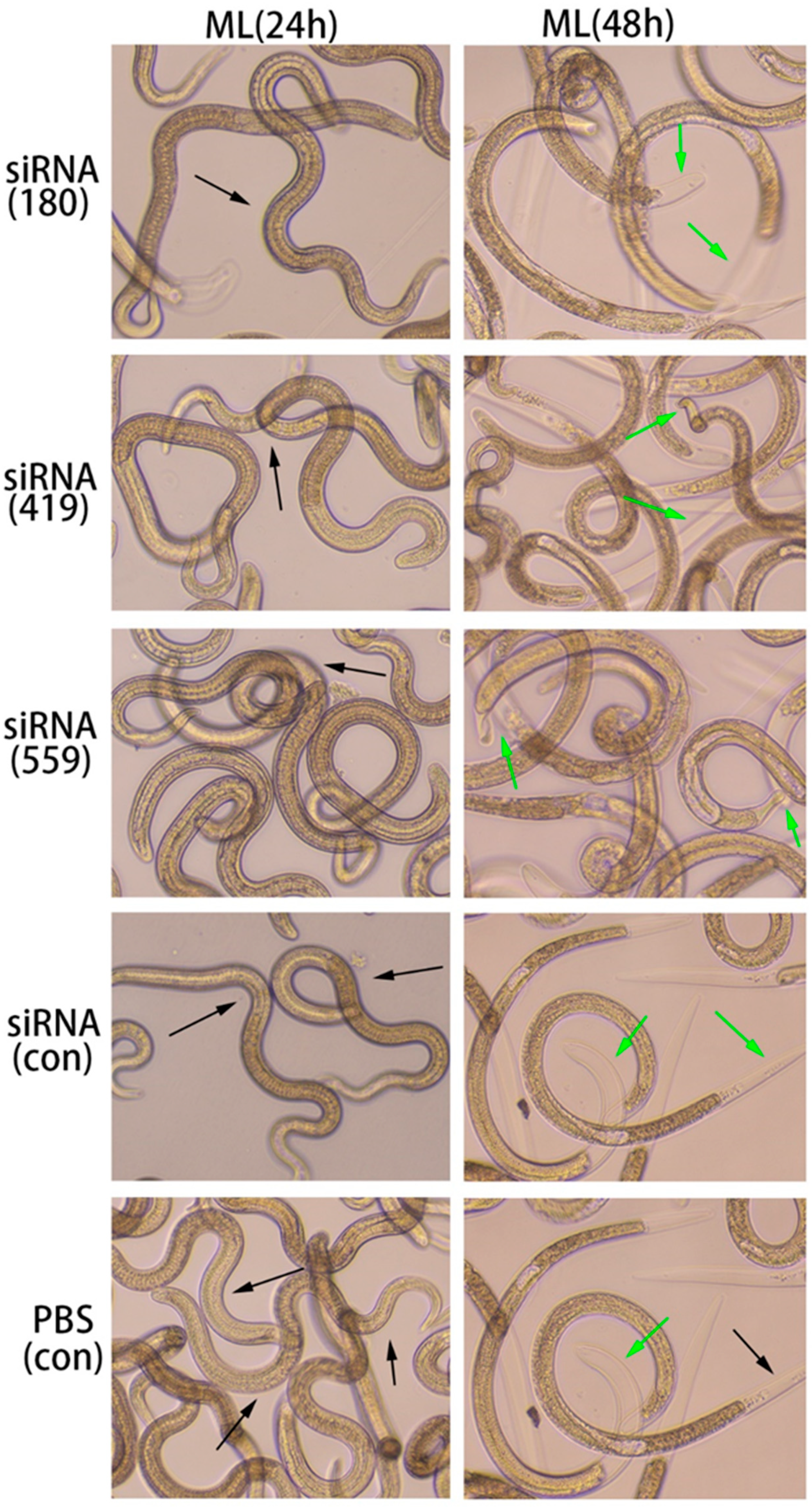
3.7. In vitro phenotyping of siRNA effects on ML
In the ML stage, the selected siRNAs (siRNA-180, siRNA-419, siRNA-559) were treated for 24 hours and 48 hours along with the controls (siRNA-control, and PBS control). After 24 hours, ecdysis (molting process) was slower in all treated groups (siRNA180, siRNA419, and siRNA559) as well as controls (siRNA-Control, and PBS control). However, at 48h, there was a difference in the ecdysis of all treated groups (siRNA180, siRNA419, siRNA559) as well as in the control groups (siRNA-Control, and PBS control). The study results showed that siRNA-559 had lesser ecdysis than siRNA-419 and siRNA-180. Compared to siRNA-419, siRNA-559 was a slower ecdysis process. The ecdysis of siRNA-180, control siRNA and control PBS were normal (Figure 10).
Figure 10.
Various siRNAs targeting the Ts-MAPRC2 gene (siRNA-180, siRNA-419, siRNA-559) were used with varying time intervals (24h, 48h), in conjunction with siRNA-Control and PBS controls at the ML stage to observe motility and ecdysis (molting process) at objective 10×. The green arrow pointed to ecdysis (molting process) and the black arrow represented motility.
Figure 10.
Various siRNAs targeting the Ts-MAPRC2 gene (siRNA-180, siRNA-419, siRNA-559) were used with varying time intervals (24h, 48h), in conjunction with siRNA-Control and PBS controls at the ML stage to observe motility and ecdysis (molting process) at objective 10×. The green arrow pointed to ecdysis (molting process) and the black arrow represented motility.
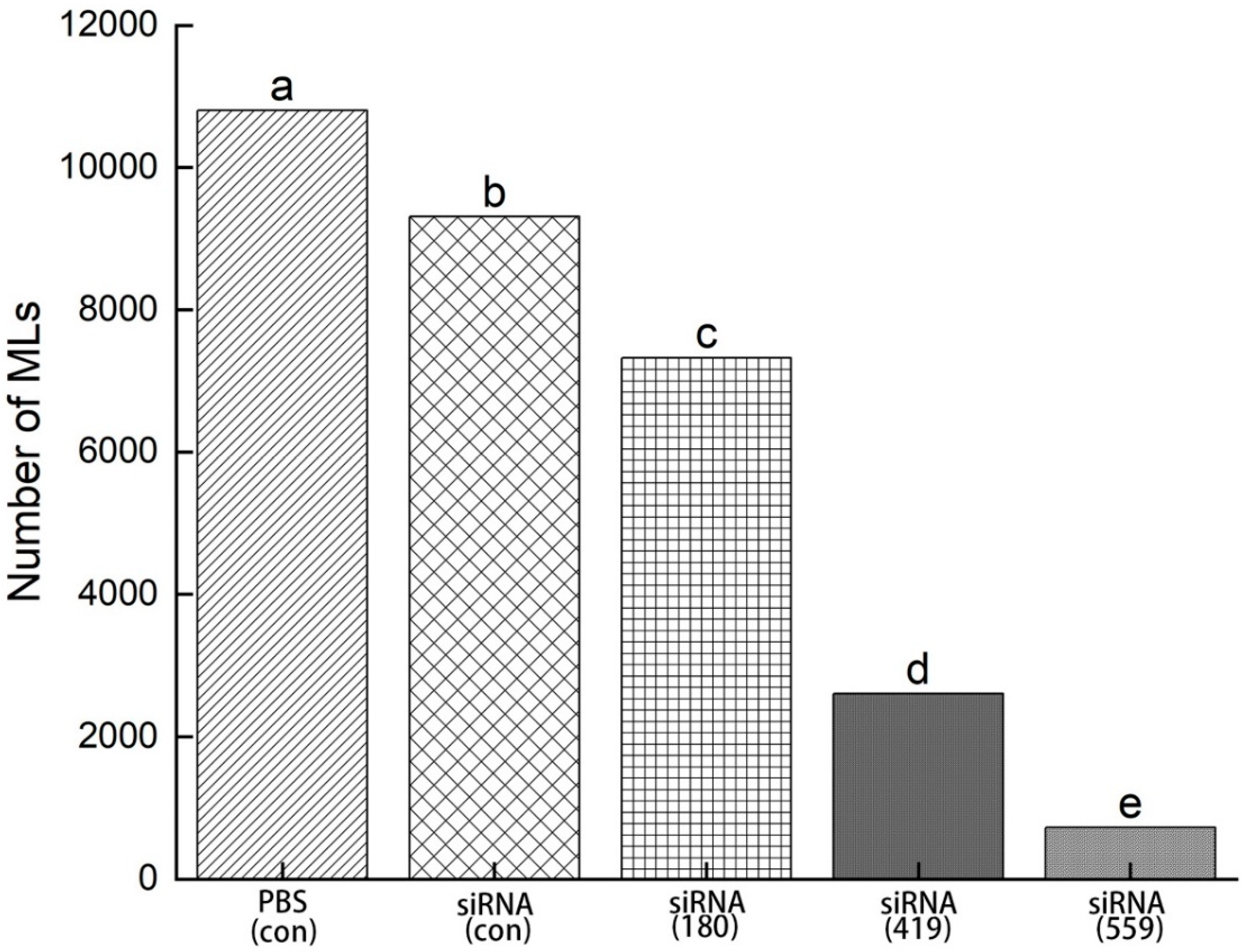
3.8. Evaluation of siRNA-treated NBL for infectivity
NBLs treated with siRNAs (siRNA-180, siRNA-419, siRNA-559, siRNA-Control, and PBS control) were injected into mice through the tail vein and the total number of muscle larvae (MLs) were counted at 35 DPI. There was a significant decrease in muscle larvae (MLs) collected with siRNA-559 when compared with siRNA-419 and siRNA-180. Compared to siRNA-419, siRNA-559 had a significantly lower total number of muscle larvae (MLs). Control siRNA compared to control PBS had no more significant difference in the total number of muscle larvae (MLs) (Figure 11) (p ≤ 0.05).
Figure 11.
After 35 days of post-infection (35dpi), total muscle larvae were collected from NBLs treated with siRNAs (siRNA-180, siRNA-419, siRNA-559, siRNA-Control, and PBS). Statistical data were presented as mean ± SD. p ≤ 0.05 was considered significant. The same letters indicate significance, while different letters indicate non-significantly.
Figure 11.
After 35 days of post-infection (35dpi), total muscle larvae were collected from NBLs treated with siRNAs (siRNA-180, siRNA-419, siRNA-559, siRNA-Control, and PBS). Statistical data were presented as mean ± SD. p ≤ 0.05 was considered significant. The same letters indicate significance, while different letters indicate non-significantly.
4. Discussion
PGRMC1 and PGRMC2 are members of the same family of membrane-associated progesterone receptors (MAPR) [40,41]. First, PGRMC-1 protein was collected from pork smooth muscle at 28 kDa [42,43]. In our previous study we Characterization of MAPRC2 from T. spiralis and its Interaction with Progesterone and Mifepristone [21,22]. This study was performed IIFT to find out the native protein on the cuticle at ML, a difference of 2 hours, 4 hours, 8 hours, and 24 hours was observed (Figure 1). FITC fluorescence protein recognition increases at time intervals of 8 hours and 24 hours. Additionally, the native protein was also detected in the new borne larvae (NBL) using rat antisera against the rTs-MAPRC2 at different concentrations after 24h. (Figure 2). As a second group of studies, immunofluorescence assays were also performed to examine membrane-bound progesterone proteins in the T. solium cysticerci as well as T. spiralis [20,21,44].
Moreover, motility and ecdysis (molting process) were observed with various concentrations of rTs-MAPRC2-Ab. The motility was present in the ML stage at all four-time intervals (0h, 4h, 8h, 24h). The study of ecdysis (molting process) revealed that 1:50 (rTs-MAPRC2-Ab ratio) had a slower rate of molting compared to 1:200 (rTs-MAPRC2-Ab ratio) and 1:800 (rTs-MAPRC2-Ab ratio). (Figure 3). We also observed motility in new-born larvae (NBL) after treatments with different concentrations of anti-rat sera against rTs-MAPRC2, after 24 h of treatment (Figure 4). Motility was slower in 1:50 (rTs-MAPRC2-Ab ratio) than in 1:200 and 1:800 (rTs-MAPRC2-Ab ratio). The result of motility and ecdysis (molting process) were correspondingly supported by the finding of Aleem et al. as well as Gagliardo et al. [21,45].
Further, testing the infectivity of ML in vivo after treatment with rTs-MAPRC2-Ab. Adult female worms collected at 6 days post-immunization from those immunized with rTs-MAPRC2-Ab (1:50, 1:200 and 1:800 dilutions) were consistently shorter than those from groups immunized with pET32a rat serum and PBS (p ≤ 0.05). It is noted that the efficacy of anti-rTs-MAPRC2 rat serum (1:50) is superior to other concentrations (1:200 and 1:800) (Figure 5B) (p ≤ 0.05). In some respects, these findings are similar to those of Cui et al. [46].
Figure 6A shows adult worm burden was lower in 1:50 (rTs-MAPRC2-Ab ratio) than in 1:200 and 1:800 (rTs-MAPRC2-Ab ratio) respectively (p ≤ 0.05). According to figure 6B, the total number of new larvae borne from adult worms due to in vitro culture was shown after 24 hours. Overall, the number of new borne larvae (NBLs) was lower at 1:50 (rTs-MAPRC2-Ab ratio) than at 1:200 and 1:800 (rTs-MAPRC2-Ab ratio), respectively (p ≤ 0.05). There was a lower number of muscle larvae collected (MLs) in the ratio of 1:50 (rTS-MAPRC2-Ab) compared to 1:200 (rTS-MAPRC2-Ab) and 1:800 (rTS-MAPRC2-Ab), respectively (Figure 6C) (p ≤ 0.05). Based on these results, it is evident that anti-rTs-MAPRC2 rat serum (1:50) provides a better result than other concentrations (1:200 and 1:800) in accordance with these findings [32,46].
We subsequently analyzed the infectivity of NBLs treated with rTs-MAPRC2Ab. Based upon the results of the study shown in Figure 7, the number of muscle larvae collected (MLs) was significantly lower in 1:50 (rTs-MAPRC2-Ab ratio) (p ≤ 0.05). Several other studies also support these results, including Fei et al., Gagliardo et al., and Cui et al. [34,45,46].
siRNAs are artificially synthesized to study the host-parasite interaction and its application was an effective tool for identifying as well as studying gene functions [25,26]. Genes needed for parasite development and the growth process of T. spiralis can be silenced or suppressed, which reduces or prevents parasite attacks on humans as well as on other mammals [27]. According to Figure 8 results, siRNA-559 inhibits the expression of Ts-MAPRC2 more effectively than siRNA-180 and siRNA-419. Simultaneously, siRNA-559 inhibits the Ts-MAPRC2 protein more effectively than siRNA-419 (p ≤ 0.05). The findings support the hypothesis that Ts-MAPRC2 have specific steroid-binding domains that might mediate the steroid cycle siRNA-Ts-MAPRC2 effect on worm reproduction these findings [21,34,40]. In subsequent experiments, mRNA levels in ML were determined for all the treated groups. As compared to siRNA-419 and siRNA-180, siRNA-559 showed a greater downregulation of the Ts-MAPRC2 gene. Simultaneously with siRNA-419, siRNA-559 was found to downregulate the Ts-MAPRC2 gene more than siRNA-419 (p ≤ 0.05) (Figure 9). It indicated that suppression of Ts-MAPRC2 expression would have a detrimental effect on T. spiralis within the host based on these findings [21,34,36]. Moreover, the selected siRNAs (siRNA-180, siRNA-419, siRNA-559) were exposed for 24 hours and 48 hours in order to observe motility and ecdysis (molting process). All three treated groups exhibited a slower ecdysis (molting process) after 24 hours. At 48 hours, all the treated groups showed a difference in the ecdysis. Based on the results of the study, siRNA-559 showed a lower ecdysis rate than both siRNA-419 and siRNA-180. As shown in Figure 10, siRNA-180, control siRNA, and control PBS all gave normal results. Accordingly, the results of Aleem et al. and Gagliardo et al. were also in support of the results of motility and ecdysis (molting process) [21,45]. Further, we evaluated the infectivity of NBLs treated with siRNAs. According to the results shown in Figure 11, siRNA-559 significantly decreased the number of muscle larvae (MLs) collected as compared with siRNA-419 and siRNA-180(p ≤ 0.05). Other studies have supported these findings, including Fei et al., Gagliardo et al., and Cui et al. [21,34,45].
5. Conclusion
It is concluded from the present study that antibody against Ts-MAPRC2 exerted a profound inhibitory effect on the development in both muscle larvae (ML) and new-born larvae (NBL) of T. spiralis. Further, siRNA-Ts-MAPRC2 strongly silenced the Ts-MAPRC2 gene in both muscle larvae (ML) and new-born larvae (NBL) of T. spiralis. By knocking down the Ts-MAPRC2 gene by siRNA, we found that larval survival, development, and infectivity were significantly reduced. Based on these results, Ts-MAPRC2 could be considered as a novel molecular target for the development of vaccines against T. spiralis infection.
Author Contributions
Conceptualization, R.Y.; methodology, M.T.A.; validation, M.T.A. and R.Y.; formal analysis, M.T.A., and Z.W.; investigation, M.T.A., Y.Z., C.C. and L.M.; writing—original draft preparation, M.T.A.; writing—review and editing, M.T.A. and R.Y.; project administration, X.L., X.S., L.X. and R.Y.; funding acquisition, R.Y. and X.L. All authors read and approved the final manuscript.
Funding
This work was supported by the Social Development Project of Jiangsu Province (BE2022848) and a project funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Institutional Review Board Statement
Regarding the animals included in our research, this study followed the guidelines of the Animal Ethics Committee, Nanjing Agricultural University, China. The approval ID was PZ2020057.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated or analyzed during this study are included within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yang, Y.; Bai, X.; Li, C.; Tong, M.; Zhang, P.; Cai, W.; Liu, X.; Liu, M. Molecular Characterization of Fructose-1,6-Bisphosphate Aldolase from Trichinella Spiralis and Its Potential in Inducing Immune Protection. Front. Cell. Infect. Microbiol. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Cui, J.; Wang, Z.Q. An Epidemiological Overview of Swine Trichinellosis in China. Vet. J. 2011, 190, 323–328. [Google Scholar] [CrossRef]
- Darwin Murrell, K.; Pozio, E. Worldwide Occurrence and Impact of Human Trichinellosis, 1986-2009. Emerg. Infect. Dis. 2011, 17, 2194–2202. [Google Scholar] [CrossRef] [PubMed]
- U.N. Food & Agriculture Organization; World Health Organization Multicriteria-Based Ranking for Risk Management of Foodborne Parasites [Preliminary Report]. 2012, 47.
- Cui, J.; Jiang, P.; Liu, L.N.; Wang, Z.Q. Survey of Trichinella Infections in Domestic Pigs from Northern and Eastern Henan, China. Vet. Parasitol. 2013, 194, 133–135. [Google Scholar] [CrossRef]
- Jiang, P.; Zhang, X.; Wang, L.A.; Han, L.H.; Yang, M.; Duan, J.Y.; Sun, G.G.; Qi, X.; Liu, R.D.; Wang, Z.Q.; et al. Survey of Trichinella Infection from Domestic Pigs in the Historical Endemic Areas of Henan Province, Central China. Parasitol. Res. 2016, 115, 4707–4709. [Google Scholar] [CrossRef]
- Bai, X.; Hu, X.; Liu, X.; Tang, B.; Liu, M. Current Research of Trichinellosis in China. Front. Microbiol. 2017, 8, 1472. [Google Scholar] [CrossRef]
- Zhang, N.; Li, W.; Fu, B. Vaccines against Trichinella Spiralis: Progress, Challenges and Future Prospects. Transbound. Emerg. Dis. 2018, 65, 1447–1458. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Wu, X.; Wang, X.; Bai, X.; Shi, H.; Tang, B.; Liu, X.; Song, Y.; Boireau, P.; Wang, F.; et al. Vaccination of Mice with an Antigenic Serine Protease-like Protein Elicits a Protective Immune Response against Trichinella Spiralis Infection. J. Parasitol. 2013, 99, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Sun, X.; Li, B.; Huang, J.; Zhan, B.; Zhu, X. Vaccination with a Paramyosin-Based Multi-Epitope Vaccine Elicits Significant Protective Immunity against Trichinella Spiralis Infection in Mice. Front. Microbiol. 2017, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.Y.; Zhang, Y.; Yang, D.; Ren, H.N.; Sun, G.G.; Jiang, P.; Liu, R.D.; Zhang, X.; Cui, J.; Wang, Z.Q. The Immune Protection Induced by a Serine Protease Inhibitor from the Foodborne Parasite Trichinella Spiralis. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Li, W.; Yang, Z.; Pan, A.; Liao, W.; Zhou, X. A Novel Antigenic Cathepsin B Protease Induces Protective Immunity in Trichinella-Infected Mice. Vaccine 2018, 36, 248–255. [Google Scholar] [CrossRef]
- Strauss, J.F.; Barbieri, R.L.; Gargiulo, A.R. Yen & Jaffe’s Reproductive Endocrinology: Physiology, Pathophysiology, and Clinical Management: Eighth Edition. Yen Jaffe’s Reprod. Endocrinol. Physiol. Pathophysiol. Clin. Manag. Eighth Ed. 2018, 1–1006. [Google Scholar] [CrossRef]
- Piccinni, M.P.; Giudizi, M.G.; Biagiotti, R.; Beloni, L.; Giannarini, L.; Sampognaro, S.; Parronchi, P.; Manetti, R.; Annunziato, F.; Livi, C. Progesterone Favors the Development of Human T Helper Cells Producing Th2-Type Cytokines and Promotes Both IL-4 Production and Membrane CD30 Expression in Established Th1 Cell Clones. J. Immunol. 1995, 155. [Google Scholar] [CrossRef]
- Miyaura, H.; Iwata, M. Direct and Indirect Inhibition of Th1 Development by Progesterone and Glucocorticoids. J. Immunol. 2002, 168, 1087–1094. [Google Scholar] [CrossRef]
- Anzaldúa, S.R.; Camacho-Arroyo, I.; Cerbón, M.A. Histomorphological Changes in the Oviduct Epithelium of the Rabbit during Early Pregnancy. Anat. Histol. Embryol. 2002, 31, 308–312. [Google Scholar] [CrossRef]
- Nuñez, G.G.; Gentile, T.; Costantino, S.N.; Sarchi, M.I.; Venturiello, S.M. In Vitro and in Vivo Effects of Progesterone on Trichinella Spiralis Newborn Larvae. Parasitology 2005, 131, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Hlaka, L.; Chitanga, S.; Masola, B.; Mukaratirwa, S. Host Pregnancy Influences the Establishment of Trichinella Zimbabwensis in Balb C Mice. J. Parasit. Dis. 2017, 41, 799–804. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, J.; Shao, J.; Gao, Y.; Xu, J.; Yu, S.; Liu, Z.; Jia, L. The Unique Pharmacological Characteristics of Mifepristone (RU486): From Terminating Pregnancy to Preventing Cancer Metastasis. Med. Res. Rev. 2014, 34, 979–1000. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-díaz, H.; Nava-castro, K.E.; Escobedo, G.; Domínguez-ramírez, L.; García-varela, M.; Río-araiza, V.H.; Palacios-arreola, M.I.; Morales-montor, J. A Novel Progesterone Receptor Membrane Component ( PGRMC ) in the Human and Swine Parasite Taenia Solium : Implications to the Host-Parasite Relationship. 2019. [CrossRef]
- MT, A.; J, S.; Z, Y.; Z, W.; Y, Z.; M, L.; SA, L.; M, H.; H, A.; MW, H.; et al. Characterization of Membrane-Associated Progesterone Receptor Component-2 (MAPRC2) from Trichinella Spiralis and Its Interaction with Progesterone and Mifepristone. Vaccines 2021, 9. [Google Scholar] [CrossRef]
- Aleem, M.T.; Khan, A.; Wen, Z.; Yu, Z.; Li, K.; Shaukat, A.; Chen, C.; Rehman, T.U.; Lu, M.; Xu, L.; et al. Molecular Docking and in Silico Simulation of Trichinella Spiralis Membrane-Associated Progesterone Receptor Component 2 (Ts-MAPRC2) and Its Interaction with Human PGRMC1. Biomed Res. Int. 2022, 2022. [Google Scholar] [CrossRef] [PubMed]
- Aleem, M.T.; Yan, R.; Khan, A.; Asrar, R.; Shakoor, A.; Asif, A.; Wen, Z.; Yu, Z.; Malik, M.A.; Tauseef-ur-Rehman; et al. Advances in the Development of Anti- Trichinella Spiralis Vaccine, Challenges, and Future Prospective. Parasit. Helminths Zoonoses - From Basic to Appl. Res. [Working Title] 2022. [CrossRef]
- Britton, C.; Laing, R.; Devaney, E. Small RNAs in Parasitic Nematodes – Forms and Functions. Parasitology 2020, 147, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.Q.; Zeng, J.; Sun, X.Y.; Yue, X.; Hu, C.X.; Jiang, P.; Liu, R.D.; Cui, J.; Wang, Z.Q. Trichinella Spiralis: RNAi-Mediated Silencing of Serine Protease Results in Reduction of Intrusion, Development and Fecundity. Trop. Biomed. 2020, 37, 932–946. [Google Scholar] [CrossRef]
- Britton, C.; Winter, A.D.; Marks, N.D.; Gu, H.; McNeilly, T.N.; Gillan, V.; Devaney, E. Application of Small RNA Technology for Improved Control of Parasitic Helminths. Vet. Parasitol. 2015, 212, 47–53. [Google Scholar] [CrossRef]
- Zhuo, T.X.; Wang, Z.; Song, Y.Y.; Yan, S.W.; Liu, R.D.; Zhang, X.; Wang, Z.Q.; Cui, J. Characterization of a Novel Glutamine Synthetase From Trichinella Spiralis and Its Participation in Larval Acid Resistance, Molting, and Development. Front. cell Dev. Biol. 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Kapp, N.; Whyte, P.; Tang, J.; Jackson, E.; Brahmi, D. A Review of Evidence for Safe Abortion Care. Contraception 2013, 88, 350–363. [Google Scholar] [CrossRef] [PubMed]
- YR, Y.; YF, Q. Progress in Treatment and Prevention of Trichinellosis. J. Infect. Dis. Ther. 2015, 3. [Google Scholar] [CrossRef]
- Wang, L.; Cui, J.; Hu, D.D.; Liu, R.D.; Wang, Z.Q. Identification of Early Diagnostic Antigens from Major Excretory-Secretory Proteins of Trichinella Spiralis Muscle Larvae Using Immunoproteomics. Parasites and Vectors 2014, 7, 40. [Google Scholar] [CrossRef]
- Yang, Y.; Lacour, S.A.; Lainé-Prade, V.; Versillé, N.; Grasset-Chevillot, A.; Feng, S.; Liu, M.Y.; Boireau, P.; Vallée, I. Trichinella Spiralis Newborn Larvae: Characterization of a Stage Specific Serine Proteinase Expression, NBL1, Using Monoclonal Antibodies. Parasitology 2015, 142, 783–790. [Google Scholar] [CrossRef]
- Yu, Z.; Lu, Y.; Liu, Z.; Aleem, M.T.; Liu, J.; Luo, J.; Yan, R.; Xu, L.; Song, X.; Li, X. Recombinant Toxoplasma Gondii Ribosomal Protein P2 Modulates the Functions of Murine Macrophages In Vitro and Provides Immunity against Acute Toxoplasmosis In Vivo. Vaccines 2021, 9, 357. [Google Scholar] [CrossRef]
- Liu, C.Y.; Ren, H.N.; Song, Y.Y.; Sun, G.G.; Liu, R.D.; Jiang, P.; Long, S.R.; Zhang, X.; Wang, Z.Q.; Cui, J. Experimental Parasitology Characterization of a Putative Glutathione S-Transferase of the Parasitic Nematode Trichinella Spiralis. Exp. Parasitol. 2018, 187, 59–66. [Google Scholar] [CrossRef]
- Fei, H.; Naqvi, M.A.U.H.; Naqvi, S.Z.; Xu, L.; Song, X.; Li, X.; Yan, R. Trichinella Spiralis: Knockdown of Gamma Interferon Inducible Lysosomal Thiol Reductase (Gilt) Results in the Reduction of Worm Burden. PLoS Negl. Trop. Dis. 2021, 15, 1–16. [Google Scholar] [CrossRef]
- Naito, Y.; Yoshimura, J.; Morishita, S.; Ui-Tei, K. SiDirect 2.0: Updated Software for Designing Functional siRNA with Reduced Seed-Dependent off-Target Effect. BMC Bioinformatics 2009, 10, 1–8. [Google Scholar] [CrossRef]
- Yi, N.; Yu, P.; Wu, L.; Liu, Z.; Guan, J.; Liu, C.; Liu, M.; Lu, Y. RNAi-Mediated Silencing of Trichinella Spiralis Serpin-Type Serine Protease Inhibitors Results in a Reduction in Larval Infectivity. Vet. Res. 2020, 51, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Aimulajiang, K.; Cao, M.; Liao, S.; Naqvi, M.A.-H.; Tian, X.; Li, Z.; Lu, M.; Lakho, S.A.; Li, X.; Xu, L.; et al. Development and Potential Application of Ras Domain Containing Protein from Haemonchus Contortus for Diagnosis of Goat Infection. Anim. 2020, Vol. 10, Page 138 2020, 10, 138. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.D.; Wang, Z.Q.; Wang, L.; Long, S.R.; Ren, H.J.; Cui, J. Analysis of Differentially Expressed Genes of Trichinella Spiralis Larvae Activated by Bile and Cultured with Intestinal Epithelial Cells Using Real-Time PCR. Parasitol. Res. 2013, 112, 4113–4120. [Google Scholar] [CrossRef]
- Zou, X.; Jin, Y.M.; Liu, P.P.; Wu, Q.J.; Liu, J.M.; Lin, J.J. RNAi Silencing of Calcium-Regulated Heat-Stable Protein of 24 kDa in Schistosoma Japonicum Affects Parasite Growth. Parasitol. Res. 2011, 108, 567–572. [Google Scholar] [CrossRef] [PubMed]
- Cahill, M.A. Progesterone Receptor Membrane Component 1: An Integrative Review. J. Steroid Biochem. Mol. Biol. 2007, 105, 16–36. [Google Scholar] [CrossRef]
- Lösel, R.M.; Besong, D.; Peluso, J.J.; Wehling, M. Progesterone Receptor Membrane Component 1--Many Tasks for a Versatile Protein. Steroids 2008, 73, 929–934. [Google Scholar] [CrossRef] [PubMed]
- Falkenstein, E.; Meyer, C.; Eisen, C.; Scriba, P.C.; Wehling, M. Full-Length cDNA Sequence of a Progesterone Membrane-Binding Protein from Porcine Vascular Smooth Muscle Cells. Biochem. Biophys. Res. Commun. 1996, 229, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Peluso, J.J. Multiplicity of Progesterone’s Actions and Receptors in the Mammalian Ovary. Biol. Reprod. 2006, 75, 2–8. [Google Scholar] [CrossRef]
- Hu, C.X.; Jiang, P.; Yue, X.; Zeng, J.; Zhang, X.Z.; Song, Y.Y.; Liu, R.D.; Zhang, X.; Wang, Z.Q.; Cui, J. Molecular Characterization of a Trichinella Spiralis Elastase-1 and Its Potential as a Diagnostic Antigen for Trichinellosis. Parasites and Vectors 2020, 13, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Gagliardo, L.F.; McVay, C.S.; Appleton, J.A. Molting, Ecdysis, and Reproduction of Trichinella Spiralis Are Supported in Vitro by Intestinal Epithelial Cells. Infect. Immun. 2002, 70, 1853–1859. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Han, Y.; Yue, X.; Liu, F.; Song, Y.Y.; Yan, S.W.; Lei, J.J.; Zhang, X.; Jiang, P.; Wang, Z.Q. Vaccination of Mice with a Recombinant Novel Cathepsin B Inhibits Trichinella Spiralis Development, Reduces the Fecundity and Worm Burden. Parasites and Vectors 2019, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



