Submitted:
18 July 2023
Posted:
19 July 2023
You are already at the latest version
Abstract
Invasive macrophytes are considered problematic in natural environments and hydroelectric reservoirs. Climate changes, the occurrence of watercourses, and biotic interactions influence the biological invasion of macrophytes. Native species can correlate with invasives positively or negatively. Urochloa subquadripara is an invasive in natural or disturbed habitats co-occurring with the natives Eichhornia crassipes and Salvinia minima. Aquatic plant communities can be altered by climate change, so species distribution models (SDMs) are important tools for predicting in-vaded areas. This work aimed to make an SDM for U. subquadripara correlating with the poten-tial distribution of native species E. crassipes and S. minima. Occurrence data for U. subquadripara, E. crassipes, and S. minima were collected from databases and in consultation with the published literature. Parameters encompassing biological information of the species were entered into the CLIMEX software and used to generate the Ecoclimatic Index (EI). The species co-occurrence was performed based on multicriteria decision-making (MCDM), and weights were assigned using the analytical hierarchy process (AHP). It was observed that U. subquadripara, E. crassipes, and S. minima have a higher occurrence in tropical and subtropical regions; however, it is predicted that, from climatic changes, these species may move to high latitudes. According to climate change, the risk of invasion by U. subquadripara in the northern hemisphere is mainly in lakes, while in the southern hemisphere, the areas conducive to invasions are rivers and reservoirs. In general, emerging and floating macrophyte species such as U. subquadripara, E. crassipes, and S. minima will be favored, causing suppression of submerged species. Therefore, identifying the potential distribution of these species allows the creation of pre-invasion intervention strategies.
Keywords:
Biological invasion
; Eichhornia crassipes
; Multicriteria decision making
; Salvinia minima
; Ten-ner-grass
1. Introduction
Macrophytes are the primary producers of freshwater ecosystems and are essential for several biotic interactions [1]. These ecosystems have endemic species that move between watercourses, but they are highly invasive due to the high pressure of propagules [2]. In-vasive exotic species can cause changes in the native community, such as the extinction of macrophytes, amphibians, and fish [1]; it is estimated that spending on biological inva-sions since 1970 in the world has been approximately US$ 1.3 trillion [3].
Urochloa subquadripara (Trin) R.D. Webster, “tenner-grass” (synonymous with Brachiaria subquadripara, Brachiaria arrecta, and Urochloa arrecta) [4,5,6] is an emergent macro-phyte, rooted near the margins, which has long, floating branches, forming mats on the surface of the water [7,8]. It is a Poaceae native to Africa and invasive in tropical and sub-tropical regions [9].
Due to the easy propagation by stolons, rhizomes, or fragments that can be trans-ported in water, the invasion of U. subquadripara raises concerns about water use and the suppression of biodiversity [8,10]. In addition, the species can grow as an epiphytic life form, rooted on the banks but extending on the surface of the water using floating macro-phytes such as Pistia stratiotes and Eichhornia crassipes as support [11].
Urochloa subquadripara colonizes natural and artificial water bodies [12], occurring in Brazil in areas such as the Pantanal [13], Cerrado and Atlantic Forest [14]. In addition, it is problematic in hydroelectric reservoirs such as Barra Bonita (SP) [15], Funil (MG) [16] and Santana (RJ) [17]. The invasive potential of U. subquadripara varies according to biotic and abiotic factors [18].
The invasion of U. subquadripara correlates with the composition of the native com-munity, varying in different spatial scales [6]. At more minor scales, the similarity be-tween native and invasive species can result in competition; however, at larger spatial scales, the probability of occurrence increases with the richness of native macrophytes [19,20,21]. The co-occurrence between native and invasive species is explained by the "Theory of biotic acceptance", which suggests that the presence of native species is positively cor-related with the occurrence of invasive species [22,23].
The occurrence of U. subquadripara is associated, on a large scale, with emerging and floating native macrophytes, of which the genus Eichhornia and Salvinia stand out [7,19]. Eichhornia crassipes is a Pontederiaceae native to the Amazon basin in Brazil and Ecuador, considered an invasive of water bodies and reservoirs worldwide [24]. Salvinia minima is a macrophyte native to Mexico, Central, and South America [25], and due to its rapid growth, it is also considered a problematic weed [26].
The distribution of macrophytes is largely related to climatic regions, limited by in-creasing latitude and altitude [27,28]. Climate changes, mainly temperature, and precipi-tation, will influence the regime of water masses, which may reduce the volume and in-crease the water temperature of lakes and reservoirs [28,29,30]. Thus, climate changes in-terfere with the habitat of macrophytes, altering growth, reproduction, development [31], phenology, distribution, and species migration [32]. Emerging and floating macrophytes such as U. subquadripara, E. crassipes, and S. minima are more prone to impacts related to changes in temperature since they are more exposed than submerged macrophytes [33].
Some approaches, such as species distribution models (SDMs), can be adopted for ecological niche projection considering the climate [34]. SDMs allow for predicting the po-tential distribution of a species using occurrence and climate data [35], being an important tool in preventing the invasion of exotic species in new environments. The CLIMEX soft-ware generates SDMs from the Ecoclimatic Index (EI) based on the growth and stress pa-rameters of the species under study [36]. This methodology has been widely used for modeling the ecological niche of invasive species and weeds [37,38,39,40,41].
Thus, the potential distribution of an invasive species can be determined by paying attention only to the climate, but the integrated understanding of the factor that influences the invasive potential, such as the co-occurrence of species, allows the filtering of suitable places [35]. Multicriteria decision-making (MCDM) is an alternative tool combining several criteria in a single index [42]. Within MCDM, the analytical hierarchy process (AHP) is widely used and consists of comparing the criteria, which ensures greater relia-bility in the judgment of assigned weights [43].
Therefore, due to the invasive capacity of U. subquadripara, the risk to ecosystems and natural communities, and disturbances in reservoirs, it is necessary to study the potential distribution of the species to develop control strategies to minimize possible impacts. This study aimed to develop a potential distribution model for U. subquadripara using the CLIMEX software. In addition, to determine the potential distribution of E. crassipes and S. minima and, based on multicriteria decision-making, correlate the climatically suitable areas for native species with areas suitable for the occurrence of the invasive species.
2. Materials and Methods
2.1. Global Distribution of Urochloa subquadripara, Eichhornia crassipes e Salvinia minima
The global distributions of U. subquadripara, E. crassipes, and S. minima were defined from occurrence points collected in the Global Biodiversity Information Facility [44,45,46] and Invasive Species Compendium [47,48] online databases. New occurrence points were searched in the literature in databases such as Web of Science and Google Scholar and added to the global distribution of the species. Incomplete or duplicate information has been omitted.
The occurrence points of U. subquadripara encompassed searches with synonyms of the species, totaling 478 points after filtering 286. The global distribution of E. crassipes resulted in 8803 occurrence points and, after data filtering, 2970. S. minima resulted in 1594 occurrence points and, after data filtering, 534 (Table S1).
2.2. CLIMEX
CLIMEX software is used for climate-based species ecological niche modeling [49]. From the known distribution and biological information of the species, it is possible to generate the potential climate distribution [49]. The CLIMEX software is based on Shelford's Law of Tolerance, which determines that the growth of a species has an optimal value according to a given environmental resource, in addition to an upper and lower limit [50]. Climate suitability is defined according to the Ecoclimatic Index, which combines functions adjusted for growth and stress (Equation 1) [38,39]. The Ecoclimatic Index ranges from 0 to 100. Areas with EI = 0 were considered unsuitable; 0>EI>30 with moderate adequacy; and EI>30 highly adequate [39].
Where: EI: Ecoclimatic Index; GIA: annual growth index; SI: annual stress index; SX: interaction between stress indices.
2.3. Parameter Adjustments and Model Validation in the CLIMEX Software
The ecological niche models for U. subquadripara, E. crassipes, and S. minima were generated considering biological information of the species and known distributions. In the CLIMEX software, the "Compare location" function was used, and the biological parameters searched were adjusted so that most of the species occurrence points were inserted in locations considered by the model as highly suitable (EI>30) [39].
Urochloa subquadripara had the African continent and South America as validation regions; e, E. crassipes, and S. minima in the United States of America, Mexico, and South America; considering native areas and in places invaded by the species.
2.3.1. Subsubsection
2.3.1.1 Growth Indices
Edaphoclimatic characteristics that affect U. subquadripara seed germination have already been described [51]. The seed germination begins at 20 – 21ºC, with the optimum at 25ºC [51]. In addition, U. subquadripara has been reported to occur in places such as Paragominas (Brazil) [8] and Bantul Yogyakarta (Indonesia) [52], where the minimum and maximum average temperatures are 21 and 34ºC. Therefore, the parameters related to the ideal temperature for the growth of U. subquadripara (DV1, DV2) were set at 22 and 35ºC. Research carried out with the species also adopted temperature values within the range established as optimal [53,54,55].
The lower and upper-temperature limits (DV0, DV3) for the growth of U. subquadripara were established at 4 and 39ºC. The lower temperature was defined as reported for species of the same genus, U. panicoides [37], and from the best fit to the global distribution of U. subquadripara. The upper temperature was set at 39ºC because the species has a priority distribution in tropical and subtropical regions, with occurrence reported in Autazes (Brazil) [56] and Córdoba (Colombia) [57], where the maximum temperatures reach 38 and 39ºC, respectively.
The lower limit, upper limit, and ideal soil moisture adopted in making the model were defined considering that the species grow in different soil moisture and according to the best fit in the global distribution, being SM0, SM1, SM2, and SM3, 0; 0.1; 8 and 10 respectively (Table 1).
2.3.1.2. Stress Parameters
The temperature limit of cold stress (TTCS) was defined according to the DV0, 4ºC, limiting the potential distribution of U. subquadripara in temperate regions without reports of the species. Furthermore, U. subquadripara occurs in Naples (United States of America) [51] and Alto Paraná (Brazil) [58], regions with minimum temperatures of up to 7°C. The cold stress accumulation rate (THCS), cold stress degrees threshold (DTCS), and the day cold stress degrees rate (DHCS) were defined at -0.001 week-1, 4ºC, and -0, 01 week-1, respectively.
The temperature limit of heat stress (TTHS) was set at 40ºC because, from that value on, seed germination of U. subquadripara is reduced [51](Teuton et al., 2004). The threshold of degrees of heat stress (DTHS) was set at 39°C according to the upper-temperature limit (DV3). The heat stress accumulation rate (THHS) and the degree of heat stress-day rate (DHHS) was set to 0.01 week-1.
Under drought conditions, seeds of U. subquadripara show dormancy, and when soil moisture is restored, germination occurs [51]. In modeling the ecological niche, the drought stress threshold (SMDS) and the drought stress accumulation rate (HDS) were considered at 0.1 and 0.005 week-1.
2.3.2. Eichhornia crassipes
2.3.2.1. Stress Parameters
Ecological niche modeling for E. crassipes was performed, but the authors considered only places where the species is considered naturalized [39]; in addition, new occurrences were reported [45]. Therefore, the new modeling was generated with adjustments in the parameters of the old one to encompass most of the current distribution points. The modeling proposed by Kriticos and Brunel [39] and the current distribution of E. crassipes is presented in Figure S1.
The ideal lower (DV1) and higher (DV2) temperatures were maintained at 25 and 30ºC, as defined by [39]. The lower (DV0) and upper (DV3) temperature limits have been changed. The DV0 was defined at 0.5ºC because the species is sensitive to frost [39], and from air temperatures close to 0.5ºC, damage to the leaf blades occurs [59].
The DV3 was defined at 36ºC because research carried out in tanks suggests that, in water temperatures above 34ºC, the death of E. crassipes occurs [59]. However, other reports demonstrate that the species can withstand temperatures up to 43ºC [60,61]; therefore, an intermediate value was established.
In addition, to create the new model, humidity and degree days (PDD) parameters were inserted. The maximum leaf area of E. crassipes occurs between 1800 and 2000 ºC days; therefore, the defined PDD value was 1916 ºC days [62].
Eichhornia crassipes is a macrophyte; thus, the species is highly dependent on water in the soil [39]. However, other factors such as water salinity [63,64] and pH [65] influence the occurrence. The lower limit, upper limit, and ideal soil moisture adopted in making the model were defined considering the suitability for high moisture values and according to the best fit in the global distribution, with SM0, SM1, SM2, and SM3, 0; 0.1; 8 and 10 respectively (Table 1).
2.3.2.2. Stress Parameters
The temperature limits of cold stress (TTCS) and heat stress (TTHS) were maintained at 0.5 and 37ºC according to the methodology proposed by [39]. The accumulation rate of cold stress (THCS) and heat stress (THHS) were defined as -0.001 week -1 and 0.001 week -1, respectively.
The drought stress threshold (SMDS) (0.02) and the drought stress accumulation rate (HDS) (-0.005 week-1); were adopted in this model because, under progressive drought, there are reductions in the photosynthesis of E. crassipes; however, the defined values were low because the species tolerates conditions of up to 6% of the volumetric content of water in the soil [66] ((Table 1).
2.3.3. Salvinia minima
2.3.3.1. Growth Indicess
Research on the influence of climatic factors on the development of S. minima is scarce, so the minimum and maximum temperature values were based on studies carried out with Salvinia molesta. The species have a similar global distribution [46,67], so such a comparison is valid.
The development of S. minima is maximized at temperatures of 23 and 35ºC, and at 15ºC, there is a decline in growth and chlorophyll production [68]. In addition, experiments with S. minima adopted the average temperature range of 20 – 25ºC [69,70]. Therefore, the temperature range established as ideal for the growth of S. minima (DV1, DV2) was 23 – 30ºC.
The minimum temperature values for S. molesta vary between 5 and 10.7ºC [71,72]; and those with maximum temperature between 39 and 43ºC [61,71,72,73]. The definition of the lower limit (DV0) and the upper-temperature limit (DV3) was carried out considering the best adjustment to the occurrence data of S. minima, being 0.5 and 39ºC, respectively.
The lower and upper limits and the ideal soil moisture adopted in the making of the model were defined considering the best fit in the global distribution, being SM0, SM1, SM2, and SM3, 0.1; 0.2; 8, and 10, respectively (Table 1).
2.3.3.2. Stress Parameters
The cold stress temperature threshold (TTCS) was set at 5ºC per the DV0, and the cold stress accumulation rate (THCS) was established at -0.0003 week-1.
The temperature limit of heat stress (TTHS) was 39ºC, according to the established DV3, and because the maximum temperature for the growth of S. molesta is 39.6ºC [72]. Values for heat stress accumulation rate (THHS), heat stress degree threshold (DTHS), and heat stress degree rate-day (DHHS) were defined as 0.1 week-1, 35°C, and -0.1 week-1, considering the global distribution of S. minima.
Salvinia minima, when exposed to periods of drought, manages to return to its primary state due to its morphology that forms a mat, protecting the apical meristem and allowing the re-establishment of the species when the humidity conditions are adequate [74,75]. This tolerance occurs because, in the native area of the species, there is exposure to floods but also long periods of drought [75]. Therefore, drought stress was not considered in modeling the ecological niche of S. minima (Table 1).
2.4. Climate Data, Models, and Scenarios
Modeling of U. subquadripara, E. crassipes, and S. minima were carried out using climatic data on a Climond 10' grid. The files contain weather data on average minimum and maximum temperatures, precipitation, and monthly relative humidity. Data from 1961 to 1990, centered on 1975, represented the historical climate [76].
2.5. Multicriteria Decision-Making and Analytical Hierarchy Process
The use of multicriteria decision making (MCDM) based on GIS (Geographic Information Systems) has as basic units layers represented by polygons (vectors) or pixels (raster) (Figure 1); it allows the combination of weights of a large number of criteria [77] and obtaining results on spatial scales [42,78].
The criteria for making the multicriteria decision were based on the modeling results for the invasive species (U. subquadripara) and native species (E. crassipes and S. minima). Each criterion was divided into classes considering the Ecoclimatic Index; the values of the classes were normalized so that they were on a scale between 0 and 1 (Table 2).
The analytical hierarchy process (AHP) is widely used in MCDM to determine the weight of each criterion [78,79,80]. The AHP method uses a square preference matrix, in which all criteria are compared to each other based on the intensity of importance, where value 1 expresses “equal importance” and value 9 “extreme importance” over another factor (Table 3) [81,82].
Considering the theory of biotic acceptance, the foundation used to determine the importance of each criterion was the relevance in the potential distribution of U. subquadripara. For example, the ecological niche for U. subquadripara (criterion 1) was considered of substantial or essential importance to the ecological niche for E. crassipes (criterion 2); therefore, the value 5 was assigned to the corresponding position in the matrix. The transposition position obtained the reciprocal value, in this case, 1/5, equivalent to 0.20 (Table 3).
The criteria weights were obtained by calculating the eigenvectors of the paired comparison matrix [77]. Thus, each criterion assumed a weight based on its importance compared to the others [83] (Table 4).
The calculation of the consistency ratio (CR) was carried out to measure the consistency of the judgments and avoid inconsistencies in the definition of the values of the paired comparison matrix [77,81,83].
The consistency ratio is obtained as described in equations 2 and 3 [83]. RC values < 0.1 indicate that the pairwise comparison was properly performed [81].
Where: CI: Consistency index; λmax.: maximum eigenvalue of the paired comparison matrix; n: array order; CR: consistency ratio (default value); and; RI: Resultant Average Consistency Index.
The consistency ratio calculated in this study was equal to 0; therefore, the judgments are considered consistent.
2.6. Global Lakes and Wetlands Database (GLWD)
To compare regions considered highly suitable with the existence of bodies of water for the establishment of species, we present a map with the global database of lakes and wetlands (GLWD). Data used include level 1 (lakes with surface area ≥50 km2 and reservoirs with storage capacity ≥0.5 km3) and level 2 (lakes, reservoirs, and rivers with surface area ≥0.1 km2) [84].
3. Results
3.1. Invasive Exotic Species (Urochloa subquadripara)
The global distribution of U. subquadripara generated 276 points of occurrence in 30 countries, with Australia (48.20%), Brazil (14.35%), and Taiwan (11.53%) being the regions with the most reports (Figure 2a). Approximately 28.74% of the world area was considered very suitable for the occurrence of the species (EI>30); 5.28% moderately adequate (30 ≤EI ≤ 100); and 65.98% inadequate (EI=0). The model predicts high climate suitability in tropical regions and does not predict suitability in temperate regions such as European and American countries (Figure 2b).
The ecological niche model for U. subquadripara was validated using the native region (African continent) [7], considering that the species has high adaptation to the climatic conditions of that area. Furthermore, South America was chosen for validation because U. subquadripara is found as an invader of hydroelectric reservoirs in Brazil [7,12,85], in environments natural in Argentina [86], and as pasture in Ecuador [87] and Venezuela [88,89]. In the validation regions, about 98.6% of the points reported are in areas considered very suitable and 1.4% with moderate suitability; thus, there is a high correlation between the occurrence of the species and the generated model (Figure 3).
3.2. Native Species
3.2.1. Eichhornia crassipes
Distribution data for Eichhornia crassipes generated 2970 occurrence points, distributed in 110 countries. The countries with the highest occurrences of the species are the United States of America (35.6%), Mexico (12.7%), Australia (5.8%), and India (4.1%) (Figure 4a).
The modeling of the ecological niche of E. crassipes is in agreement with the species' current distribution, with most of the points of occurrence inserted in moderately and very suitable places. Compared to the model proposed by Kriticos and Brunel [39], the modeling covers occurrence points in European countries, the United States of America, and Japan, in addition to restricting regions of Africa, where there is no occurrence of the species. About 45.72% of the world area was considered unsuitable (EI=0) climatically for the occurrence of E. crassipes; 20.82% moderately adequate (30 ≤EI ≤ 100) and 33.46% very adequate (EI>30) (Figure 4b).
South America is the region of origin of Eichhornia crassipes, so it was one of the areas chosen for validation, along with invaded areas (United States of America and Mexico). The invaded areas were defined due to the high occurrence of the species; in addition, E. crassipes is problematic in at least thirteen states of the United States of America [90], being considered a critical invasive macrophyte in Florida [91]. About 94.8% of the occurrence points in the validation regions are found in very suitable areas, 4.7% in regions with moderate adequacy, and 0.5% in inadequate regions, demonstrating high reliability in the final model (Figure 5 ).
3.2.2. Salvinia minima
The occurrence data of S. minima generated 534 points, distributed in African (0.56%), American (94.95%), Asian (2.62%), and European (1.87%) continents in 34 countries (Figure 6a).
Approximately 60.00; 14,19, and 25.81% of the world area were considered inadequate, moderately adequate, and very adequate for establishing S. minima, respectively (Figure 6b). Around 98.7% of the occurrence points of S. minima are in regions considered very suitable (EI>30), and 1.3% in moderately suitable places (0>EI>30). The occurrence of the species in inappropriate regions was not observed (EI=0).
South America and North American regions were defined for model validation. About 79.5% of the occurrence points are located in the United States of America or Mexico; in addition, S. minima are present in fourteen states of the United States of America [92], colonizing natural habitats in Florida [93], Texas and Louisiana [94], so this region was chosen for validation. Approximately 98.9% of the points in the areas are inserted in very suitable regions, and 1.1% have moderate climatic suitability (Figure 7).
3.3. Co-Occurrence of Urochloa subquadripara with Native Species
Combining the modeling of U. subquadripara, E. crassipes, and S. minima from the multicriteria decision-making generated a raster whose pixel values ranged from 0 to 1, where 0 are areas considered unsuitable for the occurrence of U. subquadripara and 1 very suitable areas (Figure 8a).
The suitability of U. subquadripara remains limited in northern temperate regions of the US, China, Canada, and Russia, but there was an increase in suitability compared to modeling alone (Figure 8a). The global database of lakes and wetlands shows a prevalence of lakes in the northern hemisphere; however, due to low temperatures, they are subject to freezing (Figure 8b).
Regions such as Brazil, the African continent, and Asia show high suitability, and desert areas in Africa and Australia remained unsuitable for U. subquadripara (Figure 8a). In the southern hemisphere, the greatest risk of biological invasion by macrophytes is about reservoirs and large rivers (Figure 8b).
4. Discussion
4.1. Invasive EXOTIC species (Urochloa subquadripara)
There are about 3457 macrophyte species in the world, and Brazil has the highest number [95]. The distribution of U. subquadripara follows the general pattern, with few occurrences in temperate climates. The species is found in 38.7% of the world's area [95] and is restricted by cold, heat, and drought stress. Distribution limits are compatible with climatic regions, with restrictions observed in arid climates (BWh and BWk); Mediterranean and temperate (Cs and Cw); wet continental (D); and glacial (E) [96].
The occurrence of U. subquadripara is highly dependent on water availability and quality [97]. However, when compared to other macrophytes, such as Hymenachne pernambucensis, U. subquadripara is more resistant to periods of drought [98], which may be related to greater accumulation of biomass, efficiency in the use of N and P [99], and root and stem regrowth [10]. In addition, U. subquadripara is also terrestrial, being used as pasture [54]. Therefore, the species does not occur in arid climates but in semi-arid climate regions (BSh). In addition, species of the genus Urochloa have cold stress as the main distribution limiting factor [37].
The regions considered suitable in India and the African continent are classified as tropical savanna climate (Aw), monsoon climate (Am), and equatorial climate (Af), which are also predominant in Brazil. Regions of Argentina with high climatic suitability have humid subtropical climates (Cfa) and oceanic climates (Cfb) [96], similar to regions of Australia, the country with the highest number of occurrences. Sub-Saharan Africa concentrates the largest number of endemic macrophytes [28,95]; therefore, the dissemination of U. subquadripara, whose invasion success mechanism consists of a competition, mainly shading of submerged species [97], would pose a risk to these species.
4.2. Native Species
4.2.1. Eichhornia crassipes
The worldwide distribution of E. crassipes is the widest of the species studied, occurring in tropical and subtropical regions and extending into Mediterranean and temperate climates. Eichhornia crassipes are reported in 57.28% of the world, and only 1.2% of cataloged macrophytes have this vast distribution [95].
The occurrence of the species is limited by cold, heat [39], and drought; however, its distribution shifts to higher latitudes than U. subquadripara, colonizing regions with humid continental climate (Dfb and Dfa) [96]. The model proposed by Kriticos and Brunel [39] highlights that, with climate change, the species may colonize higher latitudes due to reduced cold stress and be limited by increased heat stress in Africa, India, Brazil, and Australia. [39]. This pattern is already observed in the current model, mainly about the displacement to the north.
In the current model, occurrences of E. crassipes in temporary, permanent, or ephemeral water courses were considered [100]. However, the model by Kriticos and Brunel [39] does not include reports in ephemeral watercourses in the United States of America (Colorado, Connecticut, Illinois, Maryland, Seattle, New England) and casual occurrences in European countries. Whereas under periodic drought conditions, E. crassipes present phenotypic plasticity [66]. Progressive drought can reduce the photosynthesis of the species; however, morphological adaptations are strategies of E. crassipes that survive both as an emergent macrophyte and as a terrestrial [66,101].
Control strategies after species establishment are difficult and costly. Biological, chemical, and mechanical control methods have been used to reduce the density of E. crassipes plants [102]; however, the main measure adopted is the prevention of dissemination, with the creation of laws that prevent the commercialization and distribution of the species [39,103].
4.2.2. Salvinia minima
The distribution of S. minima is the narrowest of the studied species, occurring in 14.54% of the world area [95]. The species occurs in tropical (Af, Am, Aw), humid subtropical (Cfa), and oceanic (Cfb) climatic regions [96]. The generated model highly correlates with the S. minima distribution data, with no occurrence points inserted in inappropriate regions.
Regions of Europe and China considered suitable have climatic conditions similar to the United States of America and southern South America [96]. Meanwhile, regions of the African continent and Australia have similar climatic characteristics to Brazil and India [96]. Salvinia minima were first identified in South Africa in 2022 at the Hartbeespoort dam, where it already causes damage to recreational activities, fishing, and boat traffic [104]. The presence of the macrophyte is worrying because, according to the model obtained, sub-Saharan Africa is suitable for the occurrence of S. minima; thus, laws that require its control have been applied [104].
Salvinia minima are highly prevalent in North America. The species was introduced to the United States through the aquarium trade [105], first identified in St. John's in Florida in 1928 [106], and expanded in the country due to high propagation, with a leaf doubling time of 3.5 days [26]. It is widely found in the coastal region of the country and problematic in the states of Florida and Louisiana, where biological control is carried out with Cyrtobagous salviniae Calder and Sands (Coleoptera: Curculionidae) [92,105].
Despite causing economic and biodiversity damage, S. minima have a high potential to be used in phytoremediation due to its rapid growth, adaptation to different climatic conditions, and tolerance to contaminants [68]. The species has already been identified as a phytoremediator of sewage treatment water, reducing the content of total suspended solids [107], heavy metals such as chromium [69,108], lead [109], cadmium, nickel and zinc [110]; and herbicides such as atrazine [111].
4.3. Co-Occurrence of Urochloa subquadripara with Native Species
Species distribution models generated by CLIMEX have limitations because they only consider climatic factors, not considering, for example, biotic interactions [112]. In addition to climate and the occurrence of watercourses, biological invasion by macrophytes depends on the composition of the community, the existence of natural enemies, and correlations with native species [23].
The invasion of U. subquadripara is positively correlated with the occurrence of E. crassipes and S. minima [7]. Furthermore, as it occurs in the epiphytic life form, the presence of native macrophytes such as E. crassipes increases the pressure of U. subquadripara propagules, favoring local colonization and the invasive expansion [11]. Therefore, considering the co-occurrence between species allows for identifying the potential distribution of the invader, minimizing model limitations.
The result of the co-occurrence of U. subquadripara with E. crassipes and S. minima has tropical and subtropical regions as very suitable areas. However, there is an increase in moderately suitable regions with humid subtropical (Cfa) and oceanic (Cfb) climates in the United States of America, China, and European countries [96]. This occurs because the modeling of both native species identified these locations as climatically suitable, therefore, likely for biological invasion by U. subquadripara.
These results align with climate forecasts, which estimate that northern latitudes will have higher warming rates than the rest of the world [113], which causes an increase in lake temperatures and a more extended melting period [114]. In addition, higher precipitation rates may occur [28], which, associated with surface runoff, transports more nutrients to the lakes, favoring the occurrence of emerging macrophytes [115]. Therefore, U. subquadripara can move to temperate climates [116].
The northern hemisphere encompasses the most significant number of lakes in the world [84] and the largest number of lakes larger than 10 ha [117]. Thus, the risk of invasion is aggravated because the surface area of the watercourse is positively correlated with the richness of macrophytes [27]. It is predicted that, with climate change, the number of invasive macrophytes in Europe and North America will increase [118], which would suppress native macrophytes, mainly underwater [28].
Brazil, sub-Saharan Africa, and the Asian continent were classified as very suitable. These locations are also subject to climate change; however, the impact on macrophytes' occurrence depends on the lake's depth [28]. Shallow lakes subject to warming [119] and decreases in precipitation [120] can reduce water volume and colonization by invasive macrophytes such as U. subquadripara. However, in the southern hemisphere, the concern with the invasion of macrophytes is mainly in hydroelectric reservoirs and large watercourses.
Anthropogenic changes, such as the creation of dams and hydroelectric plants, can interfere with natural geographic barriers and encourage the invasion of exotic species [10]. Generally, hydroelectric reservoirs are more invaded than natural lakes [121] due to the increase of nutrients in the water, sedimentation, and changes in biotic interactions [122,123]. Macrophytes obstruct the water intake in the reservoirs, reducing efficiency in energy generation and causing economic losses [124]. There are around 391 large hydroelectric plants in Brazil [125]; therefore, the modeling indicates a high risk of invasion since the country is climatically very suitable for U. subquadripara, E. crassipes, and S. minima.
5. Conclusions
Urochloa subquadripara has a high invasive potential; in addition, E. crassipes and S. minima, considered native, are also considered exotic invasive in other regions and are problematic even in native areas. With climate change favoring the growth and dissemination of floating and emerging macrophytes, the displacement of these species to the northern hemisphere is worrying since it is the region with the most significant number of lakes in the world.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Global distribution points of Urochloa subquadripara, Eichhornia crassipes and Salvinia minima; Figure S1: Current distribution of Eichhornia crassipes plants and Ecoclimatic Index (EI), modeled by Kriticos and Brunel, 2016, using CLIMEX. Unsuitable areas in white (EI = 0), low suitable in light red (0 <EI <30), and high suitable in red (30 <EI <100).
Author Contributions
Tayna Sousa Duque: Conceptualization, Formal analysis, Investigation, Writing - Original Draft, Writing - Review & Editing. Iasmim Marcella Souza: Formal analysis, Investigation, Writing - Original Draft, Writing- Review & Editing. Débora Sampaio Mendes: Formal analysis, Investigation, Writing - Original Draft. Ricardo Siqueira da Silva: Conceptualization, Methodology, Resources, Writing - Review & Editing. Danielle Piuzzana Mucida: Resources, Writing - Original Draft, Writing - Review & Editing, Supervision. Francisca Daniele da Silva: Writing - Review & Editing. Daniel Valadão Silva: Writing - Review & Editing, Resources. José Barbosa dos Santos: Conceptualization, Methodology, Resources, Writing - Original Draft, Writing - Review & Editing.
Funding
No funding was received to assist with the preparation of this manuscript.
Data Availability Statement
Data is contained within the article.
Acknowledgments
To the “Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)”, “Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) - Código Financeiro 001” and “Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG)”, and “Syngenta Crop Protection” for financial support.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Tasker, S. J.; Foggo, A.; Bilton, D. T. Quantifying the ecological impacts of alien aquatic macrophytes: A global meta-analysis of effects on fish, macroinvertebrate and macrophyte assemblages. Freshwater Biology, v. 67, n. 11, p. 1847-1860, 2022.
- Dudgeon, D. Freshwater biodiversity. Cambridge University Press, 2020.
- Diagne, et al. High and rising economic costs of biological invasions worldwide. Nature, v. 592, p. 571-576, 2021.
- Diamante, N. A. et al. Molecular analysis of the invasive populations of Urochloa (Poaceae) in a large Neotropical reservoir. Aquatic Botany, v. 161, p. 103183, 2020. [CrossRef]
- Carniatto, N. et al. Effects of an invasive alien Poaceae on aquatic macrophytes and fish communities in a Neotropical reservoir. Biotropica, v. 45, n. 6, p. 747-754, 2013.
- Thomaz, S. M. et al. Temporal trends and effects of diversity on occurrence of exotic macrophytes in a large reservoir. Acta Oecologica, v. 35, n. 5, p. 614-620, 2009. [CrossRef]
- Thomaz, S. M.; Silveira, M. J.; Michelan, T. S. The colonization success of an exotic Poaceae is related to native macrophyte richness, wind disturbance and riparian vegetation. Aquatic Sciences, v. 74, n. 4, p. 809-815, 2012. [CrossRef]
- Fares, A. L. B.; Nonato, F. A. S.; Michelan, T. S. New records of the invasive macrophyte, Urochloa arrecta extend its range to eastern Brazilian Amazon altered freshwater ecosystems. Acta Amazonica, v. 50, p. 133-137, 2020. [CrossRef]
- Amorim, S. R.; Umetsu, C. A.; Camargo, A. F. M. Effects of a non-native species of Poaceae on aquatic macrophyte community composition: a comparison with a native species. Journal of Aquatic Plant Management, v. 53, n. 1, p. 191-196, 2015.
- Michelan, T. S. et al. Effects of an exotic invasive macrophyte (tropical signalgrass) on native plant community composition, species richness and functional diversity. Freshwater Biology, v. 55, n. 6, p. 1315-1326, 2010. [CrossRef]
- Michelan, T. S.; Dainez Filho, M. S.; Thomaz, S. M. Aquatic macrophyte mats as dispersers of one invasive plant species. Brazilian Journal of Biology, v. 78, p. 169-171, 2017.
- Martins, D. et al. Caracterização da comunidade de plantas aquáticas de dezoito reservatórios pertencentes a cinco bacias hidrográficas do estado de São Paulo. Planta daninha, v. 26, p. 17-32, 2008.
- Pott, V. J. et al. Aquatic macrophyte diversity of the Pantanal wetland and upper basin. Brazilian Journal of Biology, v. 71, p. 255-263, 2011. [CrossRef]
- Flora do Brasil. Disponível em: <http://floradobrasil.jbrj.gov.br/reflora/floradobrasil/FB26027> . Acesso em 13 de julho de 2022.
- Domingos, V. D. et al. Initial growth of Brachiaria subquadripara (Trin.) Hitchc. plants under different nutritional conditions. Revista Brasileira de Engenharia Agrícola e Ambiental, v. 19, p. 560-566, 2015.
- Moraes, I. C. et al. Karyotype analysis and mode of reproduction of two species of Urochloa P. Beauv. Crop Science, v. 61, n. 5, p. 3415-3424, 2021. [CrossRef]
- Pitelli, R. et al. Dinâmica da comunidade de macrófitas aquáticas no reservatório de Santana, RJ. Planta daninha, v. 26, p. 473-480, 2008.
- Michelan, T.S.; Thomaz, S. M.; Bini, L. M. Native macrophyte density and richness affect the invasiveness of a tropical Poaceae species. PLoS One, v. 8, n. 3, p. e60004, 2013. [CrossRef]
- Thomaz, S. M.; Michelan, T.S. Associations between a highly invasive species and native macrophytes differ across spatial scales. Biological Invasions, v. 13, n. 8, p. 1881-1891, 2011. [CrossRef]
- Capers, R. S. et al. Aquatic plant community invasibility and scale-dependent patterns in native and invasive species richness. Ecology, v. 88, n. 12, p. 3135-3143, 2007. [CrossRef]
- Fridley, J. D. et al. The invasion paradox: reconciling pattern and process in species invasions. Ecology, v. 88, n. 1, p. 3-17, 2007.
- Stohlgren, T. J. et al. Scale and plant invasions: a theory of biotic acceptance. Preslia, v. 78, n. 4, p. 405-426, 2006.
- Thomaz, S. M.; Mormul, R. P.; Michelan, T.S. Propagule pressure, invasibility of freshwater ecosystems by macrophytes and their ecological impacts: a review of tropical freshwater ecosystems. Hydrobiologia, v. 746, n. 1, p. 39-59, 2015. [CrossRef]
- Patel, S. Threats, management and envisaged utilizations of aquatic weed Eichhornia crassipes: an overview. Reviews in Environmental Science and Bio/Technology, v. 11, n. 3, p. 249-259, 2012. [CrossRef]
- Sanchez-Galvan, G. et al. Assessment of the hyperaccumulating lead capacity of Salvinia minima using bioadsorption and intracellular accumulation factors. Water, Air, and Soil Pollution, v. 194, n. 1, p. 77-90, 2008. [CrossRef]
- Nichols, P. B.; Couch, J. D.; Al-Hamdani, S. H. Selected physiological responses of Salvinia minima to different chromium concentrations. Aquatic Botany, v. 68, n. 4, p. 313-319, 2000. [CrossRef]
- Alahuhta, J. et al. Macroecology of macrophytes in the freshwater realm: Patterns, mechanisms and implications. Aquatic Botany, v. 168, p. 103325, 2021. [CrossRef]
- Lind, L.; Eckstein, R. L.; Relyea, R. A. Direct and indirect effects of climate change on distribution and community composition of macrophytes in lentic systems. Biological Reviews, 2022. [CrossRef]
- Hintz, W. D., Schuler, M. S., Borrelli, J. J., Eichler, L. W., Stoler, A. B., Moriarty, V. W., ... & Relyea, R. A. (2020). Concurrent improvement and deterioration of epilimnetic water quality in an oligotrophic lake over 37 years. Limnology and Oceanography, 65(5), 927-938. [CrossRef]
- Gubiani, E. A. et al. Metapopulation models predict the temporal response of two macrophytes to drought in a subtropical water reservoir. Ecological Engineering, v. 100, p. 1-7, 2017. [CrossRef]
- Chaudhry, S; Sidhu, G.P.S. Climate change regulated abiotic stress mechanisms in plants: a comprehensive review. Plant Cell Reports, p. 1-31, 2021. [CrossRef]
- Battarbee, R. W. et al. Comparing palaeolimnological and instrumental evidence of climate change for remote mountain lakes over the last 200 years. Journal of Paleolimnology, v. 28, n. 1, p. 161-179, 2002. [CrossRef]
- Santamaría, L. Why are most aquatic plants widely distributed? Dispersal, clonal growth and small-scale heterogeneity in a stressful environment. Acta oecologica, v. 23, n. 3, p. 137-154, 2002. [CrossRef]
- Tourinho, L.; Vale, M.M. Choosing among correlative, mechanistic and hybrid models of species’ niche and distribution. Integrative Zoology, 2021.
- Silva, R. S. Impactos do aquecimento global na distribuição espaço-temporal do tomateiro e do inseto praga Neoleucinodes elegantalis. 2016.
- Jae-Min, J. et al. Prediction for potential distribution of yellow crazy ant (Anoplolepis Gracilipes) in response to climate change scenario. p. 313-313, 2016.
- Duque, T. S. et al. Potential Distribution of and Sensitivity Analysis for Urochloa panicoides Weed Using Modeling: An Implication of Invasion Risk Analysis for China and Europe. Plants, v. 11, n. 13, p. 1761, 2022. [CrossRef]
- Barroso, G. M. et al. Spatio-Temporal Distribution of Digitaria insularis: Risk Analysis of Areas with Potential for Selection of Glyphosate-Resistant Biotypes in Eucalyptus Crops in Brazil. Sustainability, v. 13, n. 18, p. 10405, 2021. [CrossRef]
- Kriticos, D. J.; Brunel, S. Assessing and managing the current and future pest risk from water hyacinth, (Eichhornia crassipes), an invasive aquatic plant threatening the environment and water security. PloS one, v. 11, n. 8, p. e0120054, 2016.
- Araújo, F. H. V. et al. Modelling climate suitability for Striga asiatica, a potential invasive weed of cereal crops. Crop Protection, v. 160, p. 106050, 2022. [CrossRef]
- Ferreira, S. R.; Dos Santos, J. C. B.; Ferreira, A. Análise de risco da invação biologica de Amaranthus palmeri no mundo frente às mudanças climáticas. Congresso Técnico Científico da Engenharia e da Agronomia – CONTECC, 2021.
- Chen, Y.; Yu, J.; Khan, S. Spatial sensitivity analysis of multi-criteria weights in GIS-based land suitability evaluation. Environmental modelling & software, v. 25, n. 12, p. 1582-1591, 2010. [CrossRef]
- Tavana, M.; Soltanifar, M.; Santos-Arteaga, F. J. Analytical hierarchy process: revolution and evolution. Annals of Operations Research, p. 1-29, 2021. [CrossRef]
- GBIF.org, Global Biodiversity Information Facility. Urochloa subquadripara (Trin.) R.D.Webster. Disponível em: <https://www.gbif.org/pt/species/2705869>. Acesso em: 20 de agosto de 2022a. [CrossRef]
- GBIF.org, Global Biodiversity Information Facility. Eichhornia crassipes (Mart.) Solms. Disponível em: <https://www.gbif.org/pt/species/2705869>. Acesso em: 21 de agosto de 2022b. [CrossRef]
- GBIF.org, Global Biodiversity Information Facility. Salvinia minima var. minima. Disponível em: <https://www.gbif.org/pt/species/2705869>. Acesso em: 20 de agosto de 2022c. [CrossRef]
- CABI, Invasive Species Compendium. Salvinia minima var. minima. Disponível em:< https://www.cabi.org/isc/datasheet/55773>. Acesso em: 20 de agosto de 2022a.
- CABI, Invasive Species Compendium. Urochloa subquadripara (Trin.) R.D.Webster. Disponível em:< https://www.cabi.org/isc/datasheet/55773>. 20 de agosto de 2022b.
- Kriticos, D. J. et al. Exploring the effects of climate on plants, animals and diseases. CLIMEX Version, v. 4, p. 184, 2015.
- Shelford, V. E. The ecology of North America. Urbana, IL. 1963. 10, p. 1237-1243, 2018.
- Teuton, T. C. et al. Factors affecting seed germination of tropical signalgrass (Urochloa subquadripara). Weed Science, v. 52, n. 3, p. 376-381, 2004. [CrossRef]
- Nahdi, M. S.; Darsikin, D. Distribusi dan Kemelimpahan Spesies Tumbuhan Bawah pada Naungan Pinus mercusii, Acacia auriculiformis dan Eucalyptus alba di Hutan Gama Giri Mandiri, Yogyakarta. Jurnal Natur Indonesia, v. 16, n. 1, p. 33-41, 2014. https://natur.ejournal.unri.ac.id/index.php/JN/article/view/2753/2696.
- St. Luce, M.; Gouveia, G. G.; Eudoxie, G. D. Comparative effects of food processing liquid slurry and inorganic fertilizers on tanner grass (Brachiaria arrecta) pasture: grass yield, crude protein and P levels and residual soil N and P. Grass and Forage Science, v. 72, n. 3, p. 401-413, 2017. https://doi-org.ez36.periodicos.capes.gov.br/10.1111/gfs.12240.
- Perozo-Bravo, A.; González, B.; Ortega-Alcalá, J. Efecto de la presión de pastoreo y la suplementación estratégica sobre la composición de la materia seca del pasto tanner (Brachiaria arrecta) antes y después del pastoreo. Revista de la Facultad de Agronomía, v. 26, n. 1, p. 39-58, 2009.
- Jack, H. et al. The mineral content of some tropical forages commonly used in small ruminant production systems in the Caribbean–Part 2. Tropical Agriculture, v. 97, n. 1, 2020.
- Galvão, A. K. L. et al. Levantamento fitossociológico em pastagens de várzea no Estado do Amazonas. Planta daninha, v. 29, p. 69-75, 2011. [CrossRef]
- Botero, S., & Martínez, S. (2017). Análise produtiva de dois modelos de suplementação de pastagem para mitigar o impacto da estação seca no gado de engorda em Montería, Colômbia (Dissertação de doutorado, Zamorano, Escuela Agricola Panamericano, 2017.).
- Evangelista, H. B. et al. Shade provided by riparian plants and biotic resistance by macrophytes reduce the establishment of an invasive Poaceae. Journal of Applied Ecology, v. 54, n. 2, p. 648-656, 2017. https://doi-org.ez36.periodicos.capes.gov.br/10.1111/1365-2664.12791.
- Penfound, W. T., & Earle, T. T. (1948). The biology of the water hyacinth. Ecological Monographs, 447-472.
- Yan, S. H., Song, W., & Guo, J. Y. (2017). Advances in management and utilization of invasive water hyacinth (Eichhornia crassipes) in aquatic ecosystems–a review. Critical reviews in biotechnology, 37(2), 218-228. [CrossRef]
- Howard, G. W.; Harley, K. L. S. How do floating aquatic weeds affect wetland conservation and development? How can these effects be minimised?. Wetlands Ecology and Management, v. 5, n. 3, p. 215-225, 1997. [CrossRef]
- Spencer, D. F.; Ksander, G. G. Seasonal growth of waterhyacinth in the Sacramento/San joaquin Delta, California. 2005.
- De Lacerda, L. P. et al. Growth and differential salinity reduction between Portulaca oleracea and Eichhornia crassipes in experimental hydroponic units. Environmental technology, v.40, n.17, p. 2267-2275, 2019. [CrossRef]
- Imchen, T.; Sawant, S. S.; Ezaz, W. Exposure of Eichhornia crassipes (Mart.) Solms to salt water and its implications. Current Science, p. 439-443. 2017. [CrossRef]
- Albano Pérez, E.; Ruiz Téllez, T.; Sánchez Guzmán, J. M. Influence of physico-chemical parameters of the aquatic medium on germination of Eichhornia crassipes seeds. Plant Biology, v.13, n.4, p.643-648, 2011.
- Venter, N. et al. The amphibious invader: Rooted water hyacinth’s morphological and physiological strategy to survive stranding and drought events. Aquatic botany, v. 143, p. 41-48, 2017. [CrossRef]
- GBIF.org, Global Biodiversity Information Facility. Salvinia ×molesta D.S.Mitch.. Disponível em: <https://www.gbif.org/pt/species/2705869>. Acesso em: 18 de janeiro de 2023d. [CrossRef]
- Al-Hamdani, S. H.; Ghazal, J. J. Selected physiological responses of Salvinia minima to various temperatures and light intensities. American Fern Journal, v. 99, n. 3, p. 155-161, 2009. [CrossRef]
- Ponce, S. C. et al. Effect of solution pH on the dynamic of biosorption of Cr (VI) by living plants of Salvinia minima. Ecological Engineering, v. 74, p. 33-41, 2015. [CrossRef]
- Prado, C. et al. Differential physiological responses of two Salvinia species to hexavalent chromium at a glance. Aquatic Toxicology, v. 175, p. 213-221, 2016. [CrossRef]
- Room, P. M. Equations relating growth and uptake of nitrogen by Salvinia molesta to temperature and the availability of nitrogen. Aquatic Botany, v. 24, n. 1, p. 43-59, 1986. [CrossRef]
- Van Der Heide, T. et al. A simple equation for describing the temperature dependent growth of free-floating macrophytes. Aquatic Botany, v. 84, n. 2, p. 171-175, 2006. [CrossRef]
- Whiteman, J. B.; Room, P. M. Temperatures lethal to Salvinia molesta Mitchell. Aquatic Botany, v. 40, n. 1, p. 27-35, 1991. [CrossRef]
- Oliver, J. D. A review of the biology of giant Salvinia. 1993.
- Paudel, S.; Milleville, A.; Battaglia, L. L. Responses of native and invasive floating aquatic plant communities to salinity and desiccation stress in the southeastern US coastal floodplain forests. Estuaries and Coasts, v. 41, n. 8, p. 2331-2339, 2018. [CrossRef]
- Kriticos, D. J. et al. CliMond: global high-resolution historical and future scenario climate surfaces for bioclimatic modelling. Methods in Ecology and Evolution, v. 3, n. 1, p. 53-64, 2012. [CrossRef]
- Hossain, M. S.; Das, N. G. GIS-based multi-criteria evaluation to land suitability modelling for giant prawn (Macrobrachium rosenbergii) farming in Companigonj Upazila of Noakhali, Bangladesh. Computers and electronics in agriculture, v. 70, n. 1, p. 172-186, 2010. [CrossRef]
- Marinoni, O. Implementation of the analytical hierarchy process with VBA in ArcGIS. Computers & Geosciences, v. 30, n. 6, p. 637-646, 2004. https://doi-org.ez36.periodicos.capes.gov.br/10.1016/j.cageo.2004.03.010.
- Ohta, K. et al. Analysis of the geographical accessibility of neurosurgical emergency hospitals in Sapporo city using GIS and AHP. International journal of geographical information science, v. 21, n. 6, p. 687-698, 2007. [CrossRef]
- Marinoni, O. et al. The multiple criteria analysis tool (MCAT): A new software tool to support environmental investment decision making. Environmental Modelling & Software, v. 24, n. 2, p. 153-164, 2009. [CrossRef]
- Saaty, T. L. A scaling method for priorities in hierarchical structures. Journal of mathematical psychology, v. 15, n. 3, p. 234-281, 1977. [CrossRef]
- Saaty, T. L.; Vargas, L. G. Prediction, projection and forecasting: applications of the analytic hierarchy process in economics, finance, politics, games and sports. Boston: Kluwer Academic Publishers, 1991.
- Kumar, M.; Shaikh, V. R. Site suitability analysis for urban development using GIS based multicriteria evaluation technique. Journal of the Indian Society of Remote Sensing, v. 41, n. 2, p. 417-424, 2013. https://doi-org.ez36.periodicos.capes.gov.br/10.1007/s12524-012-0221-8.
- Lehner, B.; Döll, P. Development and validation of a global database of lakes, reservoirs and wetlands. Journal of hydrology, v. 296, n. 1-4, p. 1-22, 2004. [CrossRef]
- Carniatto, N., Cunha, E. R., Thomaz, S. M., Quirino, B. A., & Fugi, R. (2020). Feeding of fish inhabiting native and non-native macrophyte stands in a Neotropical reservoir. Hydrobiologia, 847(6), 1553-1563. [CrossRef]
- Morrone, O., & Zuloaga, F. O. (2006). Novedades para la flora del nordeste de la Argentina. Hickenia, 3, 9-29.
- Gaibor Litardo, J. R. (2020). Análisis de fuentes nitrogenadas en los pastos Brachiaria en el cantón Babahoyo (Bachelor's thesis, BABAHOYO: UTB, 2020).
- Herrera, A. M., Martínez, N., Herrera, P., Colmenares, O., & Birbe, B. (2009). Consumo de heno y producción de leche en vacas doble propósito suplementadas con bloques multinutricionales. Zootecnia Tropical, 27(4), 465-473.
- Perozo-Bravo, A., González, B., Ortega-Alcalá, J., Fuenmayor, A., & Pirela, M. (2013). Evaluación de la presión de pastoreo en tanner (Urochloa arrecta) y la suplementación estratégica en mautas mestizas en bosque húmedo tropical y suelos ácidos. Revista Científica, 23(2), 150-156.
- Vonbank, J. A. et al. Water hyacinth (Eichhornia crassipes) invasion and establishment in a temperate river system. River Research and Applications, v. 34, n. [CrossRef]
- Gettys, L. A.; Thayer, K. L.; Sigmon, J. W. Phytotoxic Effects of Acetic Acid and d-limonene on Four Aquatic Plants. HortTechnology, v. 32, n. 2, p. 110-118, 2022. [CrossRef]
- Parys, K. A.; Johnson, S. J. Biological control of common Salvinia (Salvinia minima) in Louisiana using Cyrtobagous salviniae (Coleoptera: Curculionidae). Florida Entomologist, v. 96, n. 1, p. 10-18, 2013. [CrossRef]
- Rowe, C. A.; Hauber, D. P.; Wolf, P. G. Genomic variation of introduced Salvinia minima in southeastern United States. Aquatic Botany, v. 151, p. 38-42, 2018. [CrossRef]
- Jacono, C.C.; Davern, T. R.; Center, T. D. The adventive status of Salvinia minima and S. molesta in the southern United States and the related distribution of the weevil Cyrtobagous salviniae. Castanea, p. 214-226, 2001.
- Murphy, K. et al. World distribution, diversity and endemism of aquatic macrophytes. Aquatic Botany, v. 158, p. 103127, 2019. [CrossRef]
- Köppen, W.; Geiger, R. Klimate der Erde. Gotha: Verlag Justus Perthes. 1928.
- Fleming, J. P.; Dibble, E. D. Ecological mechanisms of invasion success in aquatic macrophytes. Hydrobiologia, v. 746, n. 1, p. 23-37, 2015. [CrossRef]
- Leal, R. P. et al. The success of an invasive Poaceae explained by drought resilience but not by higher competitive ability. Environmental and Experimental Botany, v. 194, p. 104717, 2022. [CrossRef]
- Bianco, S.et al. Crescimento e nutrição mineral de Urochloa arrecta. Planta Daninha, v. 33, p. 33-40, 2015.
- Lukács, B. A., Sramkó, G., & Molnár, A. (2013). Plant diversity and conservation value of continental temporary pools. Biological Conservation, 158, 393-400. [CrossRef]
- Lawlor, D. W. Genetic engineering to improve plant performance under drought: physiological evaluation of achievements, limitations, and possibilities. Journal of experimental botany, v. 64, n. 1, p. 83-108, 2013. [CrossRef]
- Villamagna, A. M.; Murphy, B. R. Ecological and socio-economic impacts of invasive water hyacinth (Eichhornia crassipes): a review. Freshwater biology, v. 55, n. 2, p. 282-298, 2010. [CrossRef]
- EPPO. European and mediterranean plant protection organization organisation europeenne et mediterraneenne pour la protection des plantes. Report of a Pest Risk Analysis: Eichhornia crassipes. OEPP/EPPO Boletim 38: 441–449, 2008.
- Coetzee, J. A. Into Africa: Salvinia minima Baker (Salviniaceae) invades South Africa. BioInvasions Records, v. 11, n. 4, p. 1011-1018, 2022. [CrossRef]
- Tipping, P. W. et al. Ecology of common salvinia, Salvinia minima Baker, in southern Florida. Aquatic botany, v. 102, p. 23-27, 2012. [CrossRef]
- Long, R.W.; Lakela, O. Flora of tropical Florida. Fla., University of Miami Press, 1971.
- Abdul Aziz, N. I. H. et al. Phytoremediation of TSS, NH3-N and COD from Sewage Wastewater by Lemna minor L., Salvinia minima, Ipomea aquatica and Centella asiatica. Applied Sciences, v. 10, n. 16, p. 5397, 2020.
- Prado, C. et al. Uptake of chromium by Salvinia minima: effect on plant growth, leaf respiration and carbohydrate metabolism. Journal of Hazardous Materials, v. 177, n. 1-3, p. 546-553, 2010. [CrossRef]
- Castro-Longoria, E. et al. Biosynthesis of lead nanoparticles by the aquatic water fern, Salvinia minima Baker, when exposed to high lead concentration. Colloids and Surfaces B: Biointerfaces, v. 114, p. 277-283, 2014. [CrossRef]
- Iha, D. S., E Bianchini Jr, I. Phytoremediation of Cd, Ni, Pb and Zn by Salvinia minima. International journal of phytoremediation, v. 17, n. 10, p. 929-935, 2015.
- Guimarães, F. P. et al. Potential of macrophytes for removing atrazine from aqueous solution. Planta daninha, v. 29, p. 1137-1147, 2011. [CrossRef]
- Shabani, F.; Kumar, L.; Esmaeili, A. Future distributions of Fusarium oxysporum f. spp. in European, Middle Eastern and North African agricultural regions under climate change. Agriculture, ecosystems & environment, v. 197, p. 96-105, 2014. [CrossRef]
- IPCC, 2018. Framing and Context. In: Global Warming of 1.5°C. An IPCC Special Report on the impacts of global warming of 1.5°C above pre-industrial levels and related global greenhouse gas emission pathways, in the context of strengthening the global response to the threat of climate change, sustainable development, and efforts to eradicate poverty. Disponível em: <https://www.ipcc.ch/sr15/>. Acesso em: 22 de dezembro de 2022.
- Hondzo, M., & Stefan, H. G. (1991). Three case studies of lake temperature and stratification response to warmer climate. Water Resources Research, 27(8), 1837-1846. [CrossRef]
- Alahuhta, J. et al. Climate change and the future distributions of aquatic macrophytes across boreal catchments. Journal of Biogeography, v. 38, n. 2, p. 383-393, 2011. [CrossRef]
- Chandrasena, N. et al. How will weed management change under climate change? Some perspectives. Journal of Crop and Weed, v. 5, n. 2, p. 95-105, 2009.
- Maberly, S. C., O’Donnell, R. A., Woolway, R. I., Cutler, M. E., Gong, M., Jones, I. D., ... & Tyler, A. N. (2020). Global lake thermal regions shift under climate change. Nature Communications, 11(1), 1232. [CrossRef]
- Bellard, C. et al. Will climate change promote future invasions?. Global change biology, v. 19, n. 12, p. 3740-3748, 2013. [CrossRef]
- Lacoul, P.; Freedman, B. Environmental influences on aquatic plants in freshwater ecosystems. Environmental Reviews, v. 14, n. 2, p. 89-136, 2006. [CrossRef]
- Kundzewicz, Z. W. et al. The implications of projected climate change for freshwater resources and their management. Hydrological Sciences Journal, v. 53, n. 1, p. 3-10, 2008. [CrossRef]
- Johnson, P. T. J.; Olden, J. D.; Vander Zanden, M. J. Dam invaders: Impoudments facilitate biological invasions into freshwaters. Frontiers in Ecology and the Environment, n. 7, p. 357-363, 2008.
- Ruhi, A. et al. Understanding the nexus between hydrological alteration and biological invasions. In: Multiple stressors in river ecosystems. Elsevier, 2019. p. 45-64. [CrossRef]
- Havel, J. E.; Lee, C. E.; Vander Zanden, J. M. Do reservoirs facilitate invasions into landscapes?. BioScience, v. 55, n. 6, p. 518-525, 2005. [CrossRef]
- CEMIG. Macrófitas Aquáticas. Caracterização e importância em reservatórios hidrelétricos. Disponível em: < https://www.cemig.com.br/wp-content/uploads/2021/03/livro-macrofitas-cemig-2021.pdf>. Acesso em 28 de setembro de 2022.
- ANA. Catálogo de metadados da Agência nacional de Águas. Disponível em: <https://metadados.snirh.gov.br/geonetwork/srv/api/records/56ac7cb5-aa22-4081-a135-c7fc0938a449>. Acesso em: 15 de novembro de 2022.
Figure 1.
Weighted combination of criteria matrix datasets.
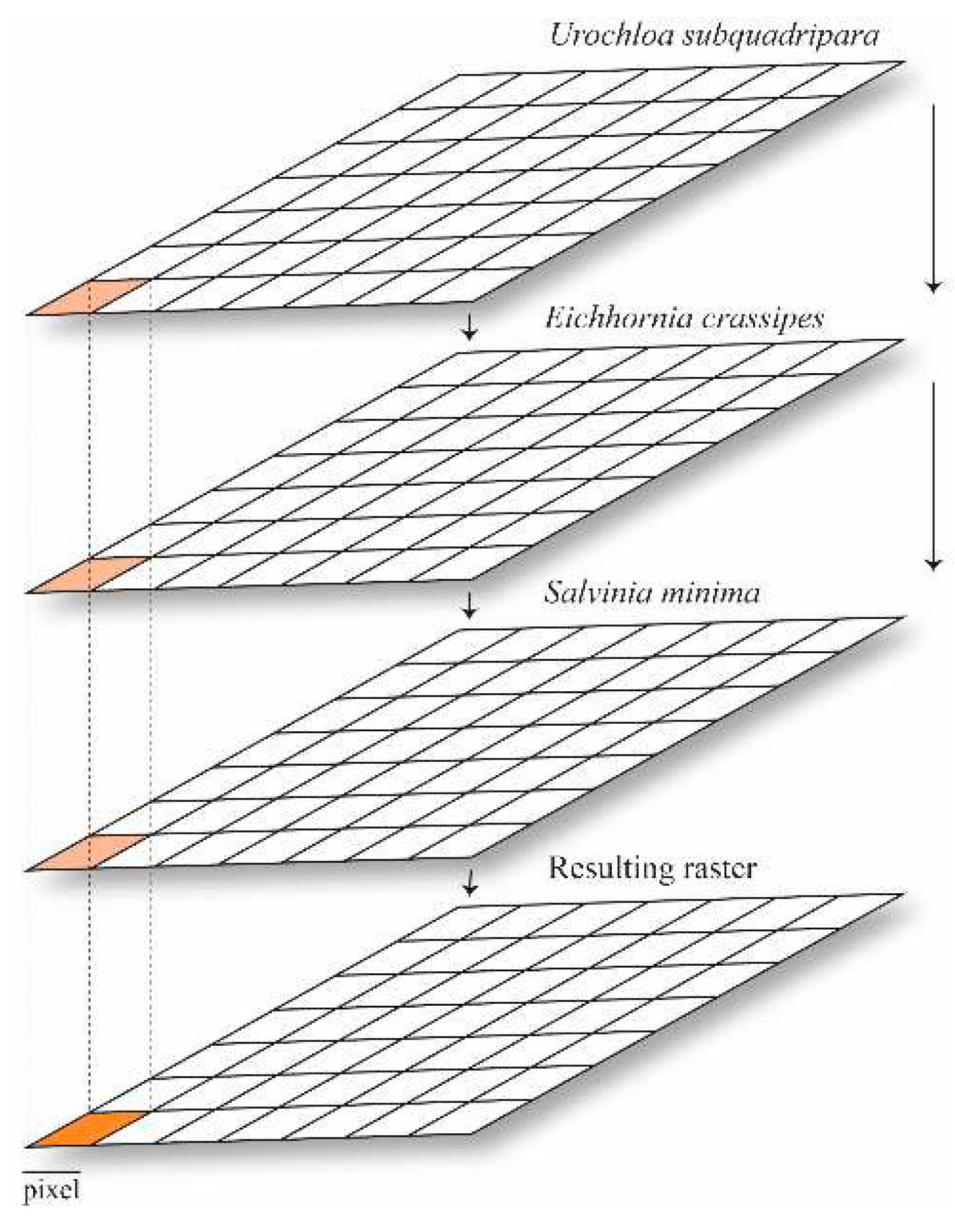
Figure 2.
(a) Known global distribution of Urochloa subquadripara plants and (b) Ecoclimatic Index (EI) for U. subquadripara, modeled using CLIMEX. Unsuitable areas in white (EI = 0), low suitable in light orange (0 <EI <30), and high suitable in orange (30 ≤EI ≤ 100).
Figure 2.
(a) Known global distribution of Urochloa subquadripara plants and (b) Ecoclimatic Index (EI) for U. subquadripara, modeled using CLIMEX. Unsuitable areas in white (EI = 0), low suitable in light orange (0 <EI <30), and high suitable in orange (30 ≤EI ≤ 100).
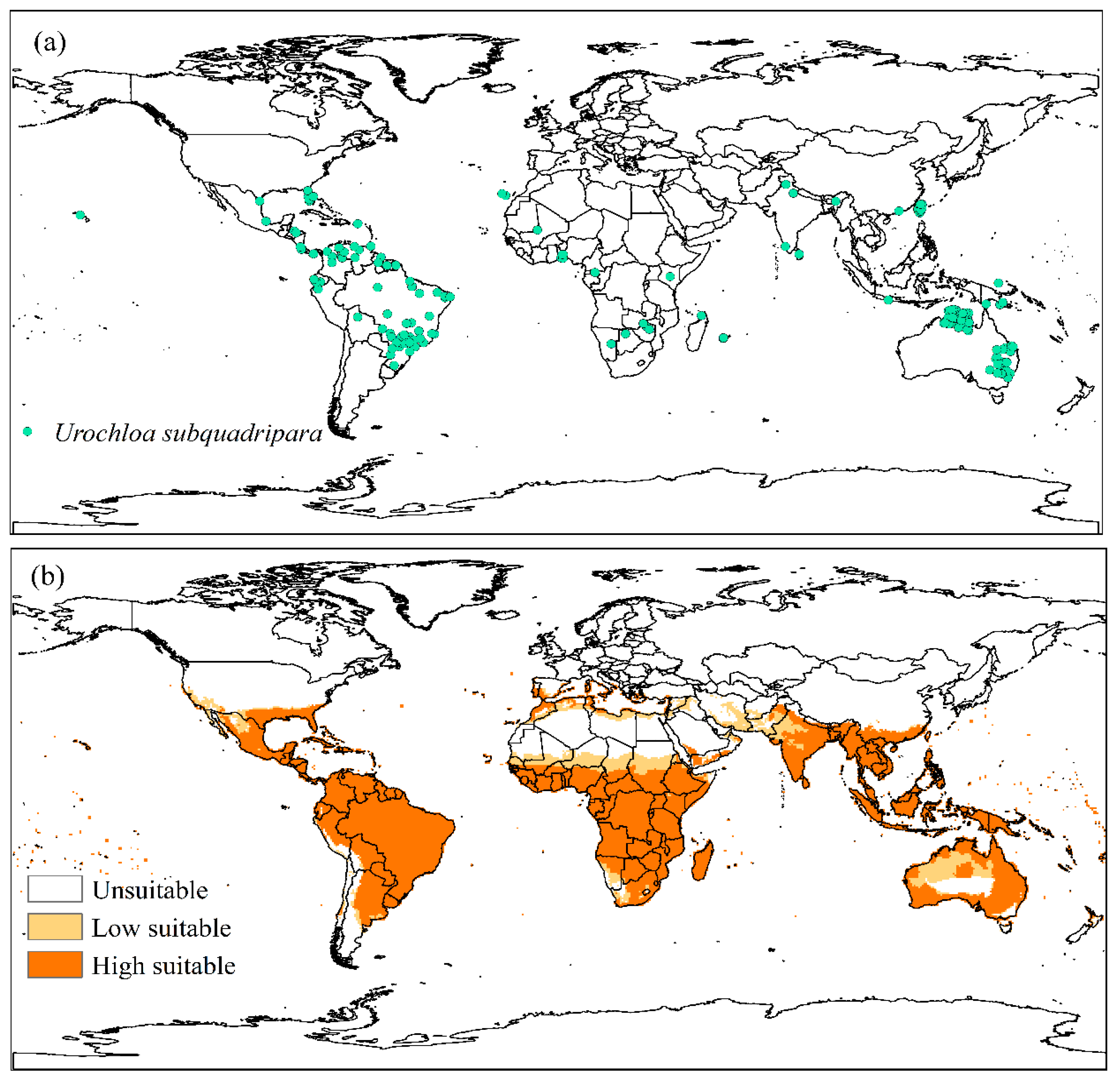
Figure 3.
Current distribution of Urochloa subquadripara in validation regions, native (African continent) and invaded area (South America), based on the Ecoclimatic Index (EI). Unsuitable areas in white (EI = 0), low suitable in light orange (0 <EI <30), and high suitable in orange (30 ≤EI ≤ 100).
Figure 3.
Current distribution of Urochloa subquadripara in validation regions, native (African continent) and invaded area (South America), based on the Ecoclimatic Index (EI). Unsuitable areas in white (EI = 0), low suitable in light orange (0 <EI <30), and high suitable in orange (30 ≤EI ≤ 100).
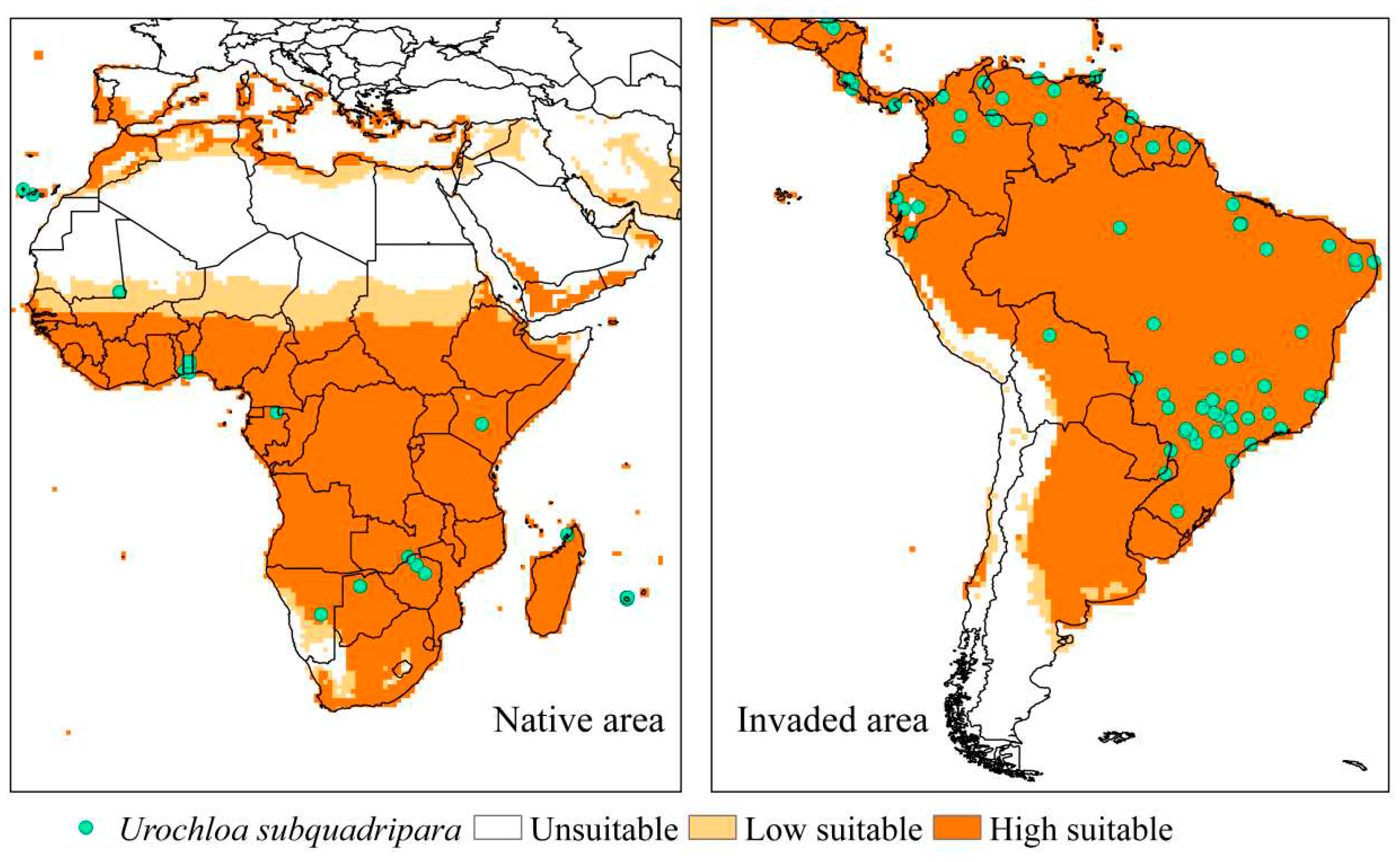
Figure 4.
(a) Known global distribution of Eichhornia crassipes plants and (b) Ecoclimatic Index (EI) for E. crassipes, modeled using CLIMEX. Unsuitable areas in white (EI = 0), low suitable in light orange (0 <EI <30), and high suitable in orange (30 ≤EI ≤ 100).
Figure 4.
(a) Known global distribution of Eichhornia crassipes plants and (b) Ecoclimatic Index (EI) for E. crassipes, modeled using CLIMEX. Unsuitable areas in white (EI = 0), low suitable in light orange (0 <EI <30), and high suitable in orange (30 ≤EI ≤ 100).
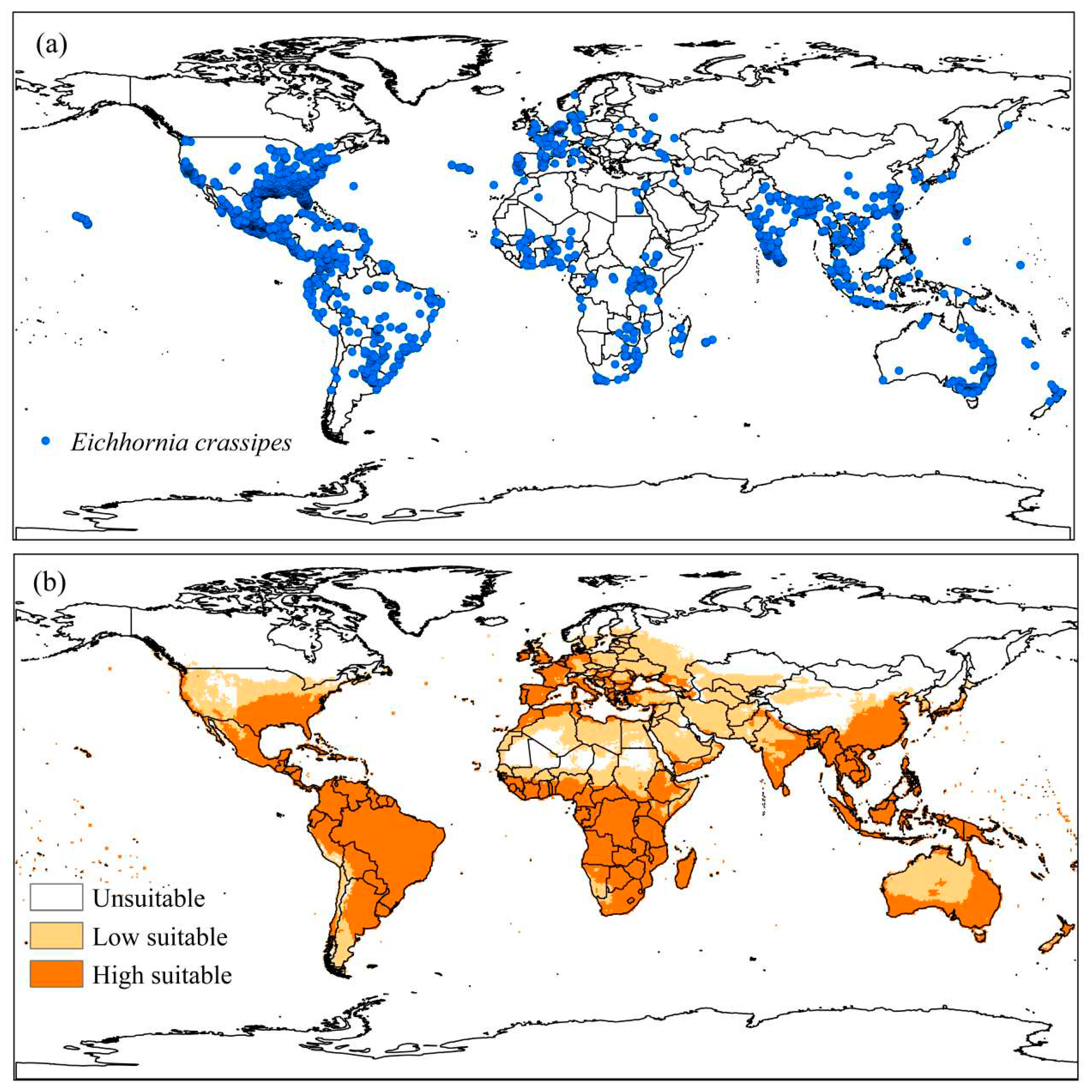
Figure 5.
Current distribution of Eichhornia crassipes in validation regions, native area (South America), and invaded area (United States of America and Mexico), based on the Ecoclimatic Index (EI). Unsuitable areas in white (EI = 0), low suitable in light orange (0 <EI <30), and high suitable in orange (30 ≤EI ≤ 100).
Figure 5.
Current distribution of Eichhornia crassipes in validation regions, native area (South America), and invaded area (United States of America and Mexico), based on the Ecoclimatic Index (EI). Unsuitable areas in white (EI = 0), low suitable in light orange (0 <EI <30), and high suitable in orange (30 ≤EI ≤ 100).
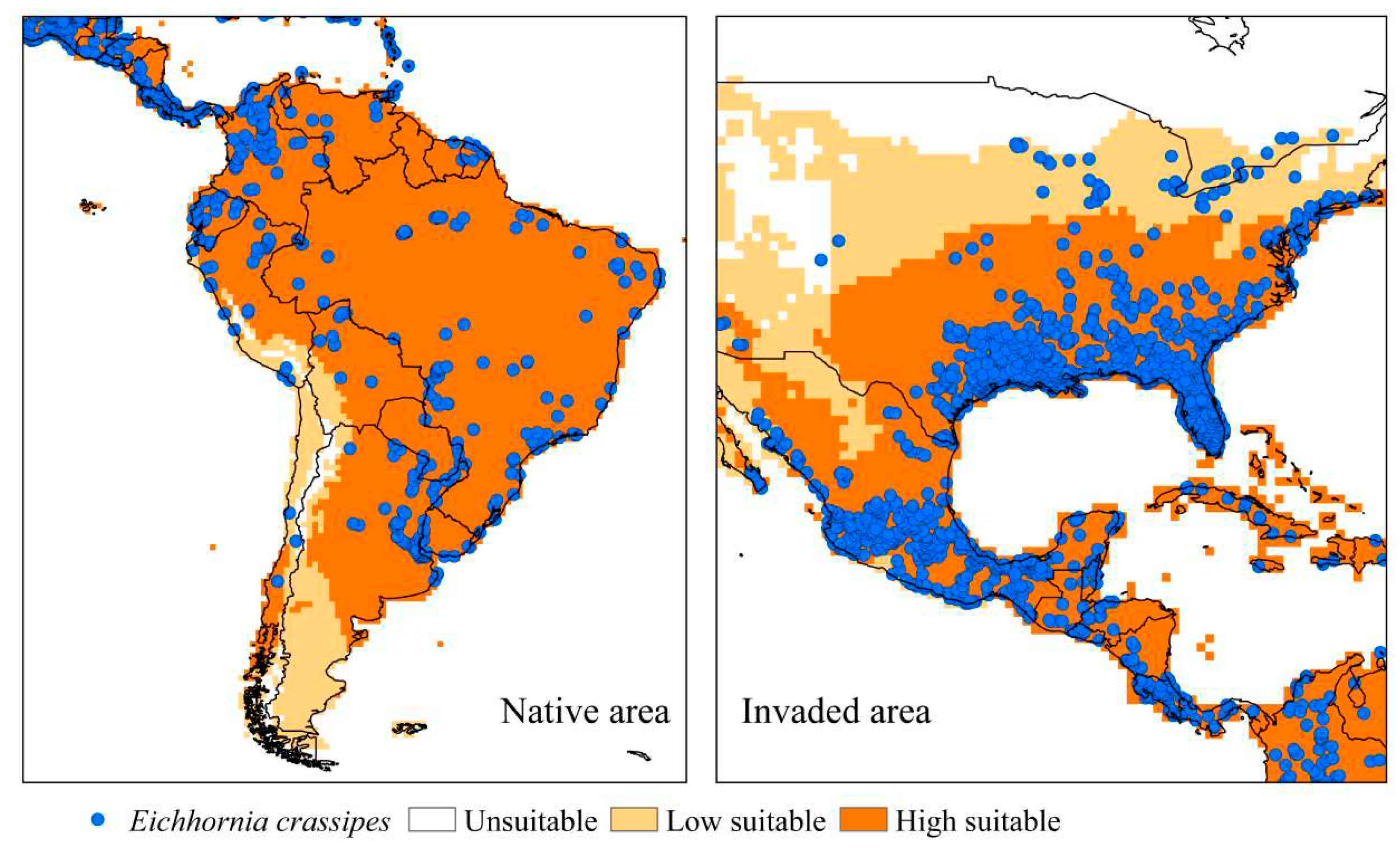
Figure 6.
(a) Known global distribution of Salvinia minima plants and (b) Ecoclimatic Index (EI) for S. minima, modeled using CLIMEX. Unsuitable areas in white (EI = 0), low suitable in light orange (0 <EI <30), and high suitable in orange (30 ≤EI ≤ 100).
Figure 6.
(a) Known global distribution of Salvinia minima plants and (b) Ecoclimatic Index (EI) for S. minima, modeled using CLIMEX. Unsuitable areas in white (EI = 0), low suitable in light orange (0 <EI <30), and high suitable in orange (30 ≤EI ≤ 100).
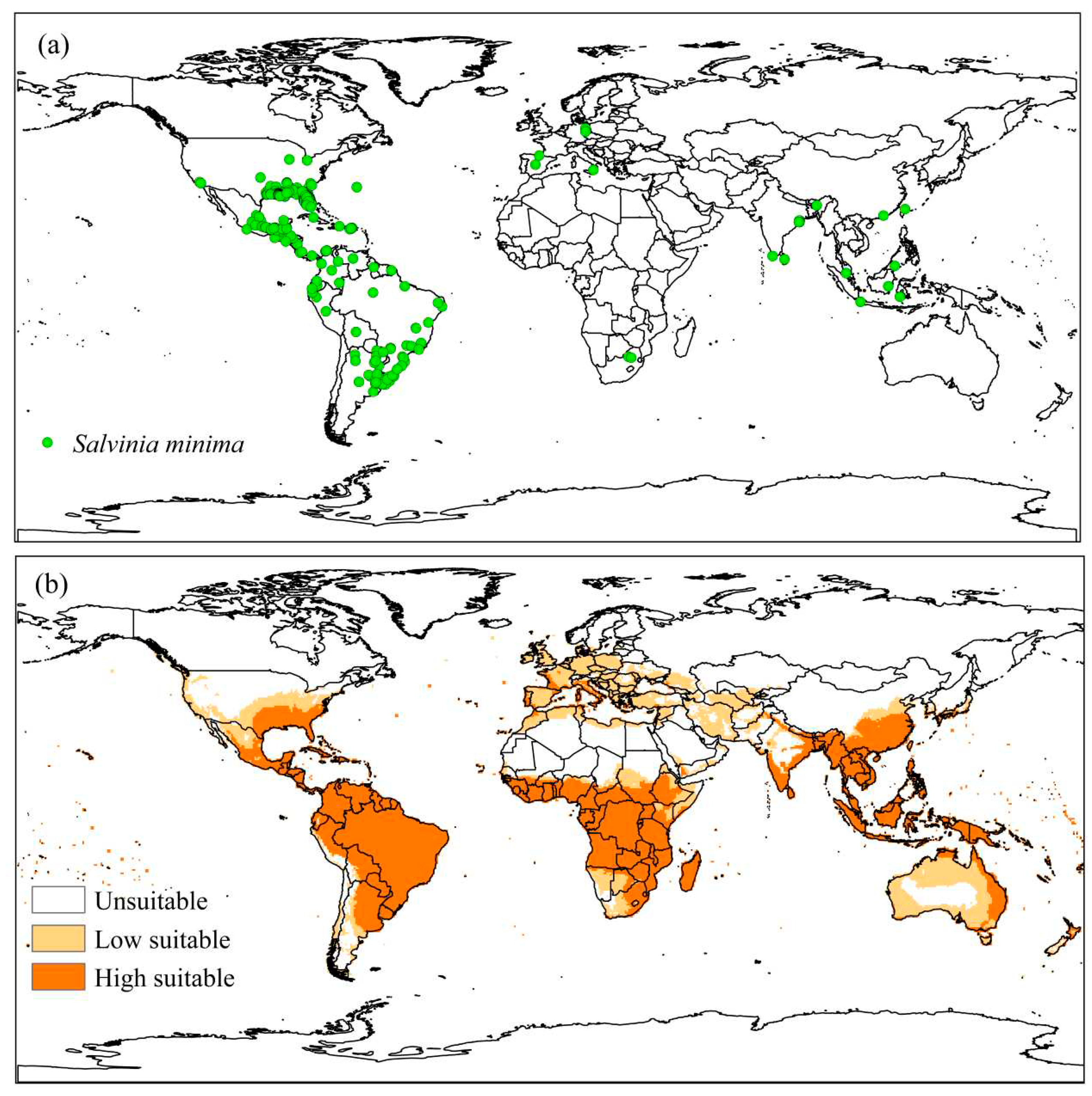
Figure 7.
Current distribution of Salvinia minima in validation regions, native area (South America), and invaded area (United States of America and Mexico), based on the Ecoclimatic Index (EI). Unsuitable areas in white (EI = 0), low suitable in light orange (0 <EI <30), and high suitable in orange (30 ≤EI ≤ 100).
Figure 7.
Current distribution of Salvinia minima in validation regions, native area (South America), and invaded area (United States of America and Mexico), based on the Ecoclimatic Index (EI). Unsuitable areas in white (EI = 0), low suitable in light orange (0 <EI <30), and high suitable in orange (30 ≤EI ≤ 100).
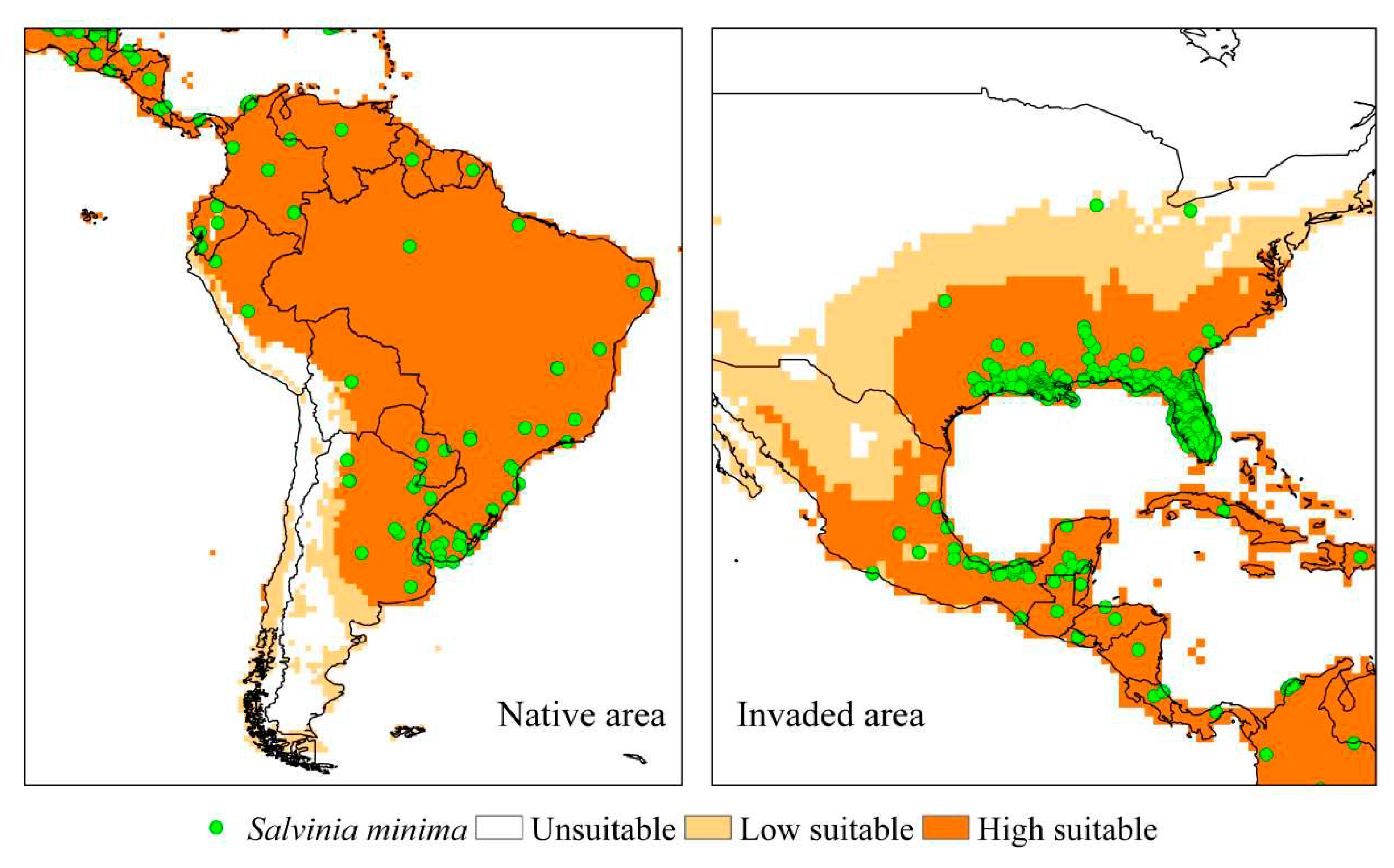
Figure 8.
(a) Climate suitability for Urochloa subquadripara considering co-occurrence with native species (Eichhornia crassipes and Salvinia minima), considering Ecoclimatic Indices (EI) modeled using CLIMEX. Inappropriate areas in white (0) and very suitable areas in red (1); (b) global lakes and wetlands database (GLWD), including tier 1 (lakes with surface area ≥50 km2 and reservoirs with storage capacity ≥0.5 km3) and tier 2 (lakes, reservoirs and rivers with surface area ≥0.1.
Figure 8.
(a) Climate suitability for Urochloa subquadripara considering co-occurrence with native species (Eichhornia crassipes and Salvinia minima), considering Ecoclimatic Indices (EI) modeled using CLIMEX. Inappropriate areas in white (0) and very suitable areas in red (1); (b) global lakes and wetlands database (GLWD), including tier 1 (lakes with surface area ≥50 km2 and reservoirs with storage capacity ≥0.5 km3) and tier 2 (lakes, reservoirs and rivers with surface area ≥0.1.
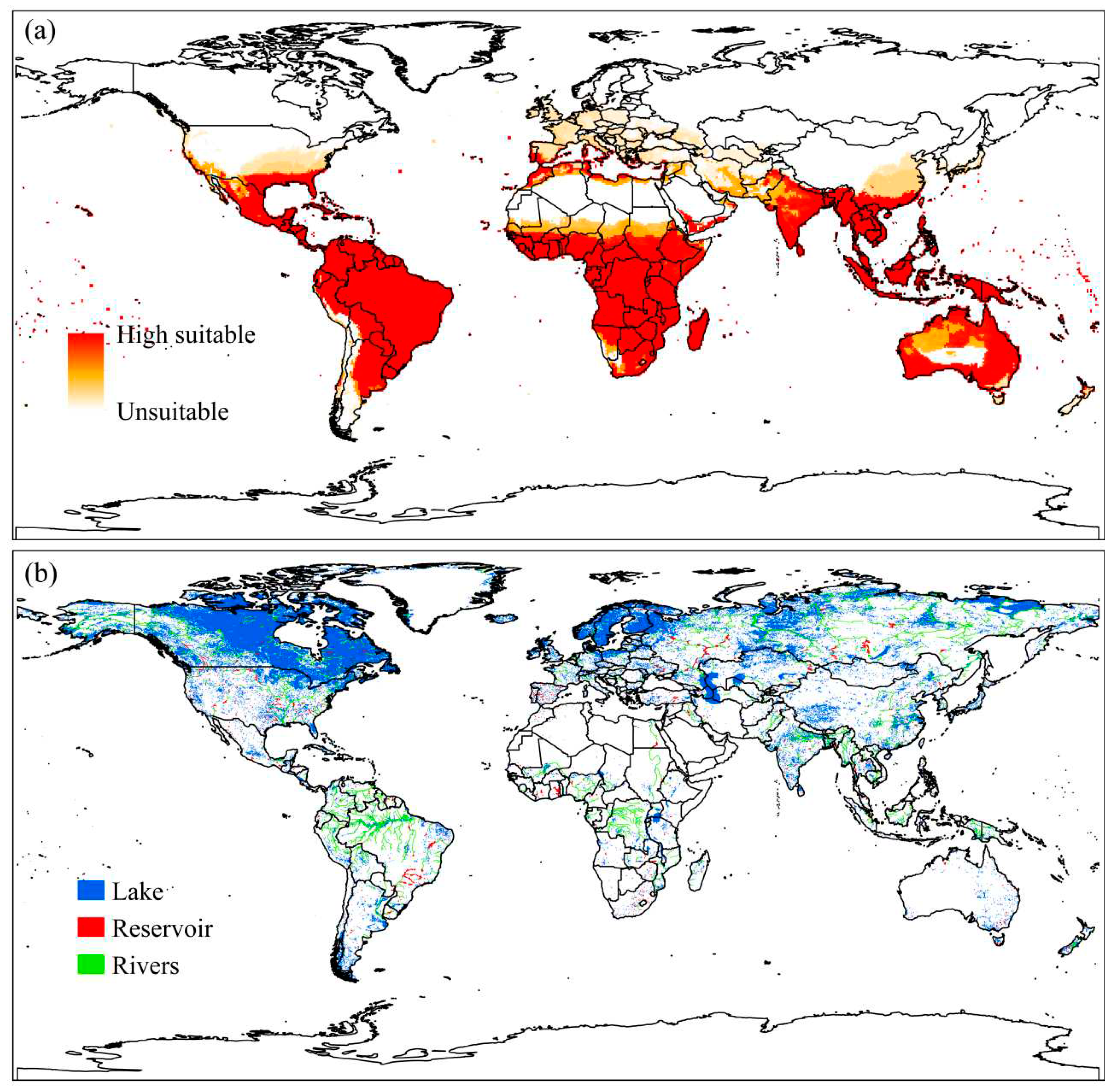

Table 1.
Adjusted parameter values for modeling the invasive species Urochloa subquadripara (US) and native Eichhornia crassipes (EC) and Salvinia minima (SM) using CLIMEX.
Table 1.
Adjusted parameter values for modeling the invasive species Urochloa subquadripara (US) and native Eichhornia crassipes (EC) and Salvinia minima (SM) using CLIMEX.
| Parameters | Index | Unit. | US | EC | SM |
|---|---|---|---|---|---|
| Lower temperature threshold | DV0 | ºC | 4 | 0.5 | 5 |
| Lower optimum temperature | DV1 | ºC | 22 | 25 | 23 |
| Upper optimum temperature | DV2 | ºC | 35 | 30 | 30 |
| Upper optimum threshold | DV3 | ºC | 39 | 36 | 39 |
| Lower soil moisture threshold | SM0 | -- | 0 | 0 | 0.1 |
| Lower optimum soil moisture | SM1 | -- | 0.1 | 0.1 | 0.2 |
| Upper optimum soil moisture | SM2 | -- | 8 | 8 | 8 |
| Upper soil moisture threshold | SM3 | -- | 10 | 10 | 10 |
| Cold stress temperature threshold | TTCS | ºC | 4 | 0.5 | 5 |
| Cold stress temperature rate | THCS | Week-1 | -0.001 | -0.0003 | -0.0003 |
| Cold stress degree-day threshold | DTCS | ºC day | 4 | ---- | ---- |
| Cold stress degree-day rate | DHCS | Week-1 | -0.01 | ---- | ---- |
| Heat Stress Temperature threshold | TTHS | ºC | 40 | 37 | 39 |
| Heat Stress Temperature rate | THHS | week-1 | 0.01 | 0.001 | 0.1 |
| Heat Stress Threshold | DTHS | ºC dia | 39 | ---- | 35 |
| Heat Stress Degree-day rate | DHHS | week-1 | 0.01 | ---- | 0.1 |
| Dry Stress Threshold | SMDS | -- | 0.1 | 0.02 | ---- |
| Dry Stress rate | HDS | week-1 | 0.005 | -0.005 | |
| Degree-days threshold | PPD | ºC dia | ---- | 1916 | ---- |
Table 2.
Criteria used in multicriteria decision-making, classes, and normalized values.
| Criteria | ||
|---|---|---|
| Criterion | Description | |
| Criterion 1 | Ecological niche for U. subquadripara | |
| Criterion 2 | Ecological niche for E. crassipes | |
| Criterion 3 | Ecological niche for S. mínima | |
| Criteria Classes | ||
| Description | Classe | Normalized value |
| EI = 0 | 0 | 0 |
| 0>EI>30 | 1 | 0.5 |
| EI>30 | 2 | 1 |
Table 3.
Paired comparison matrix describing preferences between the criteria identified in table 2.
Table 3.
Paired comparison matrix describing preferences between the criteria identified in table 2.
| Criterion 1 | Criterion 2 | Criterion 3 | |
|---|---|---|---|
| Criterion 1 | 1 | 5 | 5 |
| Criterion 2 | 0.2 | 1 | 1 |
| Criterion 3 | 0.2 | 1 | 1 |
The intensity of importance: 1 (equal importance); 3 (moderate importance of one factor over the other); 5 (strong or essential importance); 7 (very strong importance); 9 (extreme importance); 2,4,6,8 (intermediate values); reciprocals (values for inverse comparison) (Adapted from Saaty and Vargas [82]).
Table 4.
Weights used in multicriteria decision making.
| Criterion | Description | Weight* |
|---|---|---|
| Criterion 1 | Ecological Niche para U. subquadripara | 0.714 |
| Criterion 2 | Ecological Niche para E. crassipes | 0.143 |
| Criterion 3 | Ecological Niche para S. minima | 0.143 |
* Eigenvectors resulting from the paired comparison matrix.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



