
Submitted:
12 July 2023
Posted:
14 July 2023
You are already at the latest version
Abstract
In the past decades, one of the most widely reported phenomena on Caribbean reefs is the general fall in coral cover and rise in macroalgae. Reefs with low coral cover and high macroalgal abun-dances are often presumed to provide poorer ecosystem functions and services. In this study, we assessed the condition of coral reefs on the Caribbean Coast of Costa Rica and determined how eight key ecosystem metrics varied in response to differential coral and fleshy macroalgae covers. Most reefs surveyed had high fleshy macroalgae and low live coral covers, with an average (± SD) of 31 ± 28% and 14 ± 13% per site. We found that the rugosity, urchin density, fish richness, total fish biomass, large fish density, and the potential fishery value of the reef were higher in sites with low fleshy macroalgae covers (>10%). The value of many of the ecosystem metrics estimated for coral reefs of the region appears to be lower than what’s been reported for other areas in the Caribbean, which may be related to the high macroalgae covers found on the reefs. Our results concur with the prevailing paradigm that an increase in macroalgae abundance could reduce the ecosystem services provided by coral reefs.
Keywords:
coral reefs
; macroalgae
; ecosystem services
; ecosystem functions
; phase-shifts
1. Introduction
Coral reefs are in decline worldwide due to the effects of multiple global and local stressors [1]. In the Caribbean, warming events, disease outbreaks, hurricanes, overfishing, and water pollution due to inadequate coastal development have severely impacted coral reefs [2,3,4,5,6]. Because of these stressors, coral cover declined and was replaced by a high abundance of macroalgae in many Caribbean reefs [6,7,8,9]. As coral cover declined, the structural complexity of many reefs also decreased, which caused significant changes in the associated fish communities [10,11,12]. As coral reefs shift towards macroalgae-dominated states, it’s believed that their ability to support ecosystem services will be compromised [13,14,15]. Nonetheless, further studies are needed to understand the full implications of coral-algal phase shifts in the diverse range of ecosystem services provided by coral reefs, including food provision, coastal protection, tourism, recreational opportunities, and aesthetic and cultural values [16].
Recent evidence suggests that live coral cover may not be as important as previously thought in sustaining the functionality of coral reefs [17,18]. In the Caribbean, Lester et al. [17] documented poor relationships between coral cover and different ecosystem metrics relating to reef ecosystem functions and services and found numerous bright spots where the herbivorous fish biomass, large fish density, fishery value, and fish species richness were high despite the low coral cover. Thus, there appears to be high variability in the ability of reefs with low coral cover to support important reef functions [17]. One factor that may be conditioning this variability, is that various ecological states are possible in the absence of high coral cover, as the reef's benthos could be dominated by fleshy macroalgae, filamentous turf algae, calcareous coralline algae, soft corals, sponges, and other organisms [15,17,19,20]. Improving the state of knowledge of the ecosystem services provided by non-coral dominated reefs may prove useful to foster effective management strategies to support reef resilience [17,21]. Especially considering that, as the impact of global stressors like climate change and ocean acidification increases, it’s unrealistic to expect that coral reefs would recover to their past coral-dominated configurations [1].
Coral reefs in the Southwestern Caribbean (SWC), which includes the Caribbean coasts of southern Honduras, Nicaragua, Costa Rica, Panama and Colombia, have distinct coral assemblages that tend to be less diverse and complex than in other regions of the Caribbean [22,23]. The reefs of this region have been historically exposed to high levels of river discharge, which can limit the development of coral-dominated reefs [24,25,26,27]. Thereby, the SWC represents an interesting area to study the functionality of non-coral dominated reefs, as many of the reefs found in this region have low coral and high algae covers [28,29,30].
In the Caribbean Coast of Costa Rica, three areas of reef development are recognized: (1) between Moín and Limón (1), (2) Cahuita, and (3) between Puerto Viejo and Punta Mona (Figure 1)[31]. The first reports on the status of coral reefs on the Caribbean coast of Costa Rica were conducted in the 1970s and early 1980s, which described the reef and associated organisms at Cahuita [32,33]. These studies noted high levels of terrigenous sediments on the reefs [32,33]. Further studies showed that sediment loading on Cahuita reefs appeared to have increased since the 1950s and identified that the source of the sediments were banana plantations and deforested uplands north of the reef [25]. The increased sediment loading was correlated with a reduction in the growth rates of the coral colonies [25]. During the 1980s and 1990s, other natural and anthropogenic disturbances also impacted the reefs, including warming events, disease outbreaks, earthquakes, water pollution, and tourism impacts [31,34,35,36,37,38]. Because of these stressors, coral cover in Cahuita reefs decreased from 40% in the early 1980s to 10% in the early 1990s [34]. As a part of the regional program CARICOMP, the reefs in Cahuita were monitored from 1999 to 2008 [39,40]. Coral cover increased from 13% in 1999 to 28 % in 2003, but decreased during the next five years to around 17% [39]. In contrast, macroalgae cover increased significantly from 37% in 2003 to 61% in 2008 [39].
The coral reefs of the Moín – Limón and Puerto Viejo – Punta Mona areas have been historically less studied than the reefs in Cahuita, but the existing assessments also report low coral and high algae covers [28,31]. Scleractinian and fire corals covered around 23 % of the reef benthos around Puerto Viejo in 1988, which declined to just 14% in 1993; while macroalgae cover was around 33% in 1988, and 38% in 1993 [31]. In Punta Cocles, coral cover was 5% in 1983 and increased to 16% in 2002 [41,42]. Algae covers was around 59%, mostly due to the high abundance of fleshy brown macroalgae [42]. In Manzanillo, coral cover was only around 1.5% in 1993 and about 7% in 2003; while non-coralline algae cover was around 79% in 1999 and 69% in 2003 [28,43]. On the west side of Isla Uvita, between Moín – Limón, sponges dominated the reef benthos and coral cover was low (< 5%) in 2005 [28].
Since 2010, there’s only a few published reports on the status of coral reefs along the Caribbean Coast of Costa Rica [44,45]. Williams et al. [45] compared the status of Orbicella reefs and gorgonian plains between multiple countries in the Caribbean, and recorded a mean coral cover of 7.7 % in 2012 around Cahuita and Puerto Viejo – Punta Mona. Overall, there’s a lack of monitoring records on the status of coral reefs on the Caribbean coast of Costa Rica, and the existing records mostly focus on Cahuita. Also, the spatial and methodological disparity between existing assessments makes it difficult to determine regional trends in the cover of benthic organisms.
We report on the status of coral reefs of the Caribbean coast of Costa Rica based on reef survey data collected between 2019 – 2022. To our knowledge, this is the most spatially representative coral reef assessment ever recorded for the Caribbean coast of Costa Rica until now, as we surveyed 24 reef sites located among the three reefs areas. We examined how eight key ecosystem metrics —including the coral richness, substrate rugosity, urchin density, fish richness, total fish biomass, herbivore fish biomass, large fish density, and the potential fishery value of the reef— varied in response to differential coral and macroalgae cover. Our results concur with the prevailing paradigm that an increase in macroalgae abundance could reduce the ecosystem services provided by coral reefs.
2. Materials and Methods
2.1. Study area
The Caribbean coast of Costa Rica consist of high energy sandy beaches that are interrupted by carbonate promontories on the Southern section. Between Moín and Limón, carbonate platforms and small fringing and patch reefs are found along the coast, including Isla Uvita [31] (Figure 1B). Cahuita is the largest fringing reef, with three barriers; the outer barrier, stretching for 5km from the Western side of Punta Cahuita, the inner barrier, which is about 500 m long and is located between the coast and the eastern part of outer barrier; and a 100 m long barrier located at the western end of the reef [31]. Between Puerto Viejo and Punta Mona, fringing reefs, patch reef, carbonate banks and algal ridges can be found [31]. Two marine protected areas (MPA) with coral reefs are in the Caribbean coast of Costa Rica; Cahuita National Park (“Parque Nacional Cahuita”), founded in 1978; and Gandoca – Manzanillo Wildlife Refuge (“Refugio Nacional de Vida Silvestre Gandoca-Manzanillo”), founded in 1982 (Figure 1). Within the protected areas artisanal fishing and tourism activities are permitted following specific regulations.
2.2. Survey sites and data collection
Between 2019 and 2022, we conducted reef surveys at 24 sites along the Caribbean Coast of Costa Rica. Survey sites were selected with an effort to span the regions from Moín to Punta Mona. Only sites between 2 and 10 m where there was a base structure of calcium carbonate present were considered. We surveyed nine reef sites between Moín and Limón, five in Cahuita National Park, and ten between Puerto Viejo and Punta Mona, of which eight were inside the Gandoca – Manzanillo Wildlife Refuge (Figure 1). Survey site coordinates and mean depth can be found in the supplementary material Table S1.
On each reef survey, benthic, substrate rugosity, motile invertebrate, and fish community data were collected via SCUBA using standard methodologies. During every survey, three 10 m linear transects were arranged parallel to the cost. Benthic community data were collected using a photo-quadrat methodology. Ten 0.25 m2 quadrats were placed at each meter of the transect line, accounting for a total surface area of 2.5 m2 per transect. A photograph of each quadrat was taken at a 90° angle to the reef benthos. Photographs from all surveys were analyzed using the software photoQuad v1.4 [46] to quantify the cover percentage per benthic taxa and the coral species richness.
The substrate cover percentage was estimated by overlaying 50 stratified random points on each image and identifying the taxa that occurred under each point. We classified the organisms into ten functional groups: hard coral (all scleractinian taxa and the hydrozoan Millepora), soft coral, sponge, non-calcified Hexacorallia (anemones, zoanthids and corallimorphs), turf algae (defined in Smith et al. [15] as the “epilithic algal matrix” generally composed of a mixed community of filamentous algae and cyanobacteria usually less than 2 cm tall), fleshy macroalgae (Dictyota, Dictyoteris, Sargassum, Laurencia, Lobophora, etc.), calcareous macroalgae (Halimeda, Amphiroa, Galaxaura, etc.), crustose coralline algae or CCA (defined in Smith et al. [15] as all encrusting, non-geniculate members of the Corallinales), crustose non-coralline algae (Peysonelia), and non-biological substrate (sand, basalt). Hard corals were identified to species, fleshy and calcareous macroalgae to genus, and other organisms to their functional group. We estimated the coral richness per transect by counting the number of corals that appeared in the photographs of the ten quadrats surveyed. In total, we analyzed 820 photos from 82 transects.
To quantify the substrate rugosity, we fitted a 10 m long fine-linked chain (link length: 1 cm) to the contours of the reef substrate along each linear transect and measured the total distance in a straight line from the start to the end point of the chain. We calculated the rugosity index by subtracting one from the ratio of chain length to the linear distance between the chain ends; where a value of 0 indicates a flat surface and values >1 indicate greater rugosity [47,48]. Motile invertebrate data was collected by identifying and counting all the motile invertebrates found within one meter of each side of the 10 m transect line [48]. Motile invertebrates were defined as conspicuous animals of more than 2.5 cm in body size, and included echinoids, asteroids, gastropods, cephalopods, and crustaceans [48].
Fish community data was collected by counting and estimating the total length of each fish found within a 5 m wide and 10 m long belt transect [48]. Total fish length was estimated based on the following categories: <5, 5-10,10-15, 15-20, 20-25, 25-30, 30-40, 40-50, 50-100, >100 cm. Due to poor environmental conditions, we were only able to conduct fish community surveys in 15 of the 24 survey sites. For each transect, species-specific biomass was estimated according to their length to mass ratio [48].
2.3. Ecosystem metrics
Based on the reef survey data, we quantified eight ecosystem metrics relating to reef functions and services, including coral richness, substrate rugosity, urchin density, fish richness, total fish biomass, herbivore fish biomass, large fish density, and potential fishery value of the reef. Metric selection was based on existing reviews of the ecosystem services provided by coral reefs, and empirical studies on coral reef functionality [14,16,17,49]. Many of the ecosystem metrics were selected following Lester et al. [17], which studied the relationship between coral cover and different ecosystem metrics relating to functions and ecosystem services provided by Caribbean coral reefs. Each of the ecosystem metrics considered is described in the following section:
- Coral richness: Number of coral species encountered at each benthic transect (spp. 2.5 m-2). This metric is a component of biodiversity and acts as an important supporting service [16]. Thus, coral richness can enhance coral growth and reef resilience and may influence dive tourism as it’s related to the aesthetic value of the reef [50,51,52,53].
- Fish richness: Number of fish species encountered at each transect (spp. 50 m-2). This metric is a component of biodiversity, acting as an important supporting service [16]. Thus, fish richness can enhance reef resilience, support fisheries productivity, and influence dive tourism as it’s related to the aesthetic value of the reef [50,52,57,58].
- Large fish density: Number of fishes greater than 20 cm in total length counted at each transect per area (ind 100 m-2). Lester et al. [17] used a > 40 cm cutoff to measure this metric. However, we didn’t count any individual with an estimated total length greater than 40 cm, so we decided to use the > 20 cm cutoff. Larger fishes may increase the fishery value, improve the aesthetic value of the reef, and influence dive tourism [57,58,61,62].
- Potential fishery value: To quantify the potential fishery value of the reef fish assemblage, we multiplied the species-specific biomass estimates for each transect by the species-specific price estimates reported by Lester et al. [17] (USD 100 m-2). The price estimates are based on fishery statistics from Puerto Rico that provided an estimate of value in USD per pound of fish landed by species or by family Lester et al. [17]. For species that didn’t appeared in the Lester et al. [17] list, we first determined if they had or not a fishery value based on the information listed in FishBase under the human uses category [63]. If “commercial fisheries” was listed as a human use of the specific species, we then assigned it the price estimate of a similar species included in the Lester et al. [17] list (i.e. of the same genus or family). Of the total fish species encountered on the surveys, four species of commercial fishery importance were not listed in the species-specific price estimates reported by Lester et al. [17] (Supplementary material S4).
2.4. Data analysis
We estimated the general and per-site average cover of the ten benthic functional groups previously described, including hard corals, soft corals, sponges, non-calcified Hexacorallia, turf algae, fleshy macroalgae, calcareous macroalgae, CCA, crustose non-coralline algae, and non-biological substrate. The percentual contribution of each coral species to the total hard coral cover was determined by adding the estimated mean cover of the specific specie at each site and dividing it by the summation of the total hard coral cover estimated for each site [48]. The percentual contribution of each macroalgae genus to the total fleshy macroalgae was also quantified. Following Smith et al. [15], we used Pearson correlations to examine whether there was a negative relationship between mean fleshy macroalgal and coral cover, and between the cover of reef builders (hard corals + CCA) and that of fleshy algae (fleshy macroalgae + turf algae) among sites. Based on the mean cover of these benthic groups, we determined which of the 24 reef sites surveyed were dominated (> 50% cover) by hard corals, fleshy macroalgae, reef builders and fleshy algae. Also, to evaluate variations in the reef benthic composition between the Moín-Limón, Cahuita, and Puerto Viejo-Punta Mona areas, NMDS and PERMANOVA tests were performed. A SIMPER was performed to identify the benthic taxa that contributed most to the differences between regions.
We examine whether the ecosystem metrics quantified per transect varied between reefs with different levels of coral and fleshy macroalgae covers. For the analysis, we binned the coral cover values into three categories following Lester et al. [17]: low (<10% cover), medium (10-20 % cover), and high (>20% cover). We also binned Fleshy macroalgae covers into three categories: low (<10 % cover), medium (10-50 % cover), and high (>50 % cover). We determined these fleshy macroalgae cover categories based on the variation of our data and literature information. The lower cover category was mostly based on reef health indicators for Caribbean Mesoamerican reefs, which consider reefs with fleshy macroalgae covers around 12 to 0% to be in a regular to a very good state. While a >50 % cutoff was used to define the higher cover category, as to only include reefs where fleshy macroalgae were the dominant benthic organisms. Kruskal-Wallis and Mann–Whitney tests were used to determine significant differences in the ecosystem metrics —including coral richness, substrate rugosity, urchin density, fish richness, total fish biomass, herbivore fish biomass, large fish density, and the potential fishery value of the reef— between the coral and fleshy macroalgae cover categories. To assess if the data meted the homogeneity of variance assumption, we computed Leven’s tests using the “car” R package [64].
Lastly, among the reef surveyed, we identified “bright spots” and “dark spots” of coral cover, fleshy macroalgae cover, coral richness, substrate rugosity, urchin density, fish richness, total fish biomass, herbivore fish biomass, large fish density, and the potential fishery value of the reef. A site was identified as a bright spot or a dark spot when its mean was significantly different than the general mean estimated among all sites. To determine if the means were significantly different, we compared their 95% confidence intervals (CI); if the range of the CI didn´t overlap, then significant differences were assumed. All statistical tests were run using R (v.4.2.3) [65].
3. Results
The reefs surveyed were dominated by turf algae and fleshy macroalgae, with an average (± SD) cover of 42±19 % and 31±28 %, respectively (Figure 1A). Brown macroalgae of the genus Dictyota/Dictyopteris were the predominant fleshy macroalgae species found on the reefs (Figure 2B). Hard live corals covered 14 ±13 % of the reef substrate on average (Figure 2A). We identified 14 species of reef-building corals among all survey sites (Supplementary material, Table S2). The lettuce coral Agaricia agaricites and the fire coral Millepora coplanata were the predominant coral species found on the reefs, followed by the elkhorn coral Acropora palmata and the massive starlet coral Siderastrea siderea (Figure 2C). The fleshy macroalgae cover was higher than the coral cover in 15 of the 24 reefs surveyed, of which six presented fleshy macroalgae covers superior to 50%. None of the reefs surveyed presented covers of reef builders (hard corals + CCA) higher than 50%, and almost all were dominated by fleshy algae (fleshy macroalgae + turf algae) (Figure 3). We recorded significant negative relationships between coral and fleshy macroalgae cover (cor = -0.43, p < 0.001) and between reef builders and fleshy algae cover (cor = -0.83, p < 0.001).
The composition of the benthic community of the reefs varied significantly between the regions of Moín - Limón, Cahuita, and Puerto Viejo - Punta Mona (Figure 4, PERMANOVA, p = 0.001). According to SIMPER, the taxa that contributed the most to the dissimilarity between sites were the macroalgae of the genus Dictyota / Dictyopteris, which were more abundant in the reefs of the Puerto Viejo - Punta Mona area (Supplementary material, Table S3). We recorded bright spots of high coral and low fleshy macroalgae in Cahuita and Moín – Limón, while most of the reefs of Puerto Viejo – Punta Mona were categorized as dark spots of high fleshy macroalgae cover (Figure 5A-B). The reef with the highest average coral cover was located near Punta Cahuita (pe17: 39% cover), where fire corals were the dominant coral specie. We identified another bright spot of coral cover on Isla Uvita (uv20: 39% cover), where elkhorn corals were the main reef-forming species. On average, we recorded 3±2 coral species per 2.5 m2 of reef area. Despite their high macroalgae covers, many reefs in Puerto Viejo – Punta Mona appeared to be bright spots of coral richness (Figure 5C). Regarding substrate rugosity, the reefs were not very structurally complex and tended to be flat, with an averaged linear rugosity index of 0.2±0.1. The two reef locations with the highest average coral covers were categorized as bright spots of substrate rugosity (Figure 5D).
Urchin density was low among the surveyed reefs, with an average of 0.7±2.0 ind. m-2 (Figure 5E). The most common urchin species were Echinometra lucunter, Echinometra viridis, and Diadema antillarum, with average densities of 0.5, 0.1, and 0.05 ind. m-2. Concerning the fish community, we recorded 56 species belonging to 11 families were among the 16 sites where fish surveys were conducted (Supplementary material, Table S4). On average, we recorded only 5±1 fish species per 50 m2 of reef area (Figure 5F). The estimated values of the fish community metrics were low across the reef of the region, with an average biomass of 3.7±7.3 kg 100 m-2, herbivore biomass of 1.6±5.6 kg 100 m-2, large fish density (>20cm) of 15±34 ind 100 m-2, and a fishery value of only 5.1±9.6 $ 100 m-2 (Figure 5G-J). Overall, the fish families with the highest average biomass were Acanthuridae (surgeonfishes) and Pomacentridae (damselfishes), with 1.4±5.2 and 0.7±1.0 kg 100 m-2, respectively. Between all survey sites only one reef was identified as a bright spot of fish biomass, which was the same reef on Isla Uvita categorized as a bright spot of high coral and low macroalgae cover.
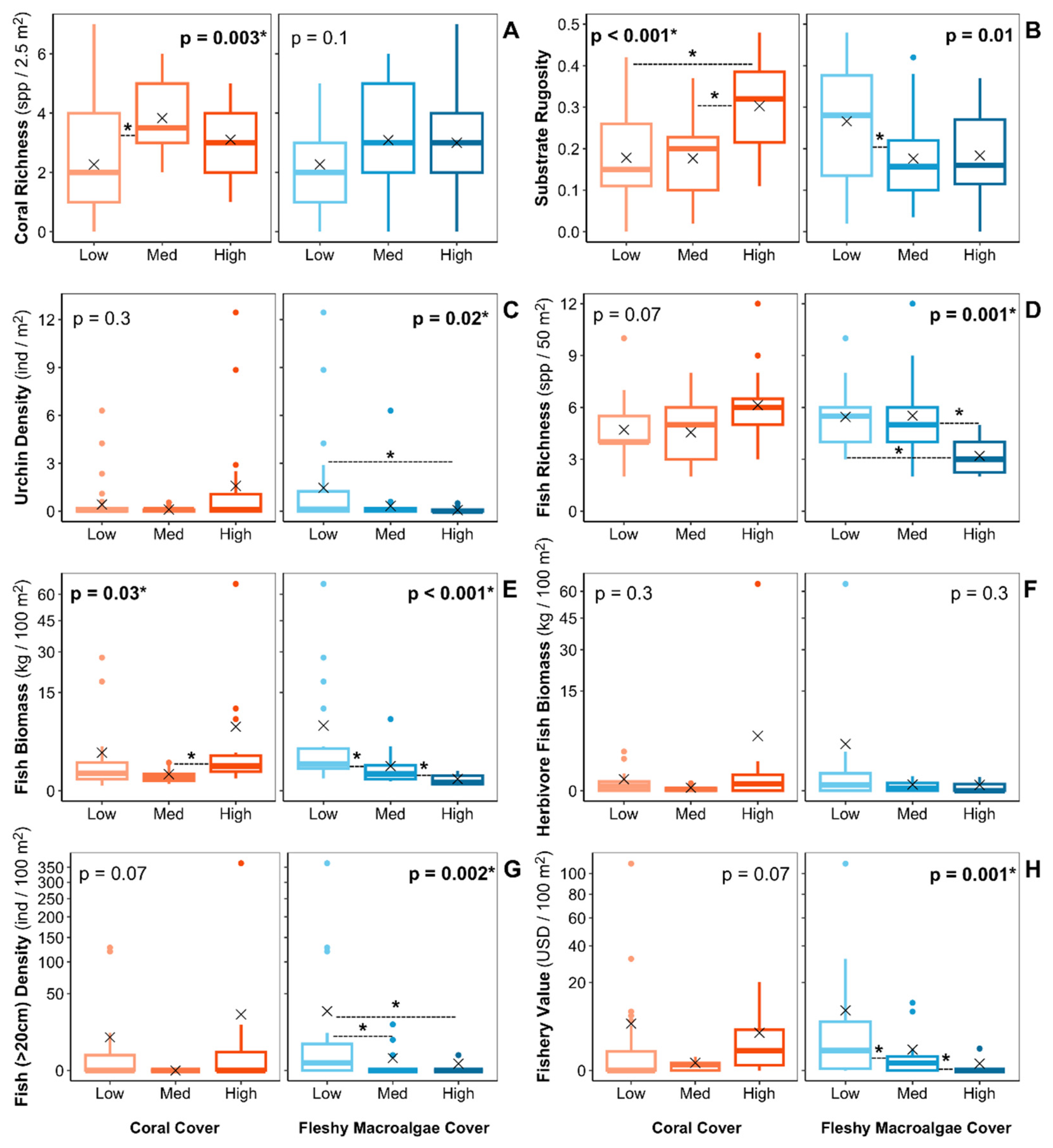
The estimated value of some ecosystem metrics relating to reef functions and ecosystem services varied between sites with different coral and macroalgae cover levels. Reefs with low coral covers (<10%) were associated with significantly lower coral richness per unit area (Figure 6A). Substrate rugosity was higher on reefs with high coral (>20%) and low macroalgae covers (<10%) (Figure 6B). Urchin density was also significantly higher on reefs with low macroalgae cover (<10%) (Figure 6C). Metrics relating to the fish community— fish species richness, biomass, large fish density (>20cm), and fishery value —were significantly higher on reefs with low macroalgae cover (Figure 6D-H). Although not statistically significant, we also recorded a tendency of lower herbivore fish richness in reefs with higher macroalgae covers. Fish community metrics appeared higher on reefs with high coral cover. However, apart from fish biomass, we didn’t find statistically significant differences of the fish community metrics between the coral cover categories.
4. Discussion
This study represents the first report on the status of coral reef communities on the Caribbean Coast of Costa Rica since Williams et al. [45] and Araya-Vargas and Nova-Bustos [44]. Between all 24 sites surveyed, this report is also the most spatially representative coral reef assessment ever recorded for the region, as it includes many reefs whose status has never been reported in the literature. The 13% average coral cover estimated for the surveyed reefs falls in a similar range to the 15.9% regional average reported for the Great Caribbean in 2019 [66]. However, the 77% total algae cover, including all algae functional groups, appears to be at the higher end of the values reported for coral reefs of the Great Caribbean, which averaged 52% for 2019 [66]. Compared to the Mesoamerican region, the 31% average fleshy macroalgae cover estimated for Costa Rican reefs is similar to the one reported in 2022 for Guatemalan reefs (30%) and higher than the ones reported for Mexico (24%), Belize (18%), and Honduras (25%) [67].
The high fleshy macroalgae and low coral cover found on the coral reefs of the Caribbean Coast of Costa Rica may indicate that these ecosystems faced phase shifts from hard corals towards macroalgae domination. For the reefs around Puerto Viejo, coral cover was around 23 % in 1988 and 14% in 1993 [31], which are lower than the 5.6 to 12 % average coral cover recorded for the reefs surveyed near the area in 2021 (site codes: pv7 and sb8). In contrast, total algae cover appears to have increased from 38% in 1993 to around 85 % in 2021 [31]. For reefs around Puerto Vargas in Cahuita National Park (site codes: pv15 and bi16), we estimated coral covers between 12 and 25%, which are much lower than the 40% covers recorded in the early 1980s [34]. Also, the increasing algae cover tendencies previously reported between 2003 (37%) and 2008 (61%) [39] seem to have continue until now, as we recorded total algae covers of around 70 to 73% for the Puerto Vargas area. Given these comparisons, it's important to consider that the specific site and methodologies used to quantify the coral and algae covers differ between studies. So, although the impression that macroalgae cover has substantially increased on the reefs of the Caribbean Coast of Costa Rica seems like a fair assumption, the lack of monitoring records and the spatial and methodological disparity between existing assessments makes it difficult to state this a fact. Nonetheless, whether macroalgae cover has increased or not, this study shows that macroalgae cover were high across most reefs of the region, and that the high macroalgae levels were associated with poorer ecosystem functions and services provided by the reefs.
We found that reefs with lower fleshy macroalgae covers were associated with lower substrate rugosity, urchin density, fish richness, fish biomass, density of large fishes and fishery values. Our results align with the prevailing paradigm that phase shift from coral to macroalgae dominated reefs could reduce the ecosystem services provided by coral reefs [13,14,15]. In contrast, most ecosystem metrics didn't vary significantly with coral cover. These results discord with past studies that found stronger linkages between coral cover and ecosystem services, and align with Lester et al. [17], who reported poor correlations between coral cover and numerous non-coral ecosystem metrics for Caribbean reefs. As discussed by Lester et al. [17], this results may suggest that, following the dramatic losses of coral cover in the 1980s and the subsequent changes in the coral species composition [8,10], some of the ecological relationships that once existed for Caribbean reefs may no longer hold. In the past, branching Acropora species were the dominant corals in many Caribbean reefs and may have played a crucial role in supporting the fish community [12,68]. Currently, the dominant coral species in the Caribbean tend to be smaller and less structurally complex [10]. Thus, the relationships between coral cover and fish community metrics in the Caribbean may depend more on the cover of specific species, like corals of the Acropora genus, than on the general coral cover [17]. This is reflected in our results, as A. palmata was the dominant coral species in the only reef categorized as a bright spot for fish community metrics.
Lester et al. [17] found numerous coral reefs in the Caribbean where metrics relating to reef functions and services were high despite low coral (>10%). Among the reefs of the Caribbean coast of Costa Rica, we didn’t find bright spots of high ecosystem metrics despite low coral cover. Also, the estimated values of most of the ecosystem metrics fell in the lower range of the values reported for other regions of the Caribbean, especially the metrics related to the fish community [6,17,69]. Lester et al. [17] suggested that the high variability they found in the ecosystem metrics for reefs with low coral cover was related to the existing range of distinct low-coral community types in the Caribbean, including those typified by sponges, gorgonians, macroalgae, or CCA [15,20]. Based on what's known about the functionality of low-coral communities, it is likely that reef communities with a greater abundance of CCA, soft corals, or sponges will be able to support a greater variety of the ecosystem services provided by coral reefs than communities dominated by turf and/or macroalgae [15,70,71,72]. This concurs with the poor ecosystem metrics we recorded, as most reefs with low coral cover were dominated by turf and/or fleshy macroalgae.
The fleshy algae dominance on the coral reefs of the Caribbean coast of Costa Rica may be due to a combination of natural and anthropogenic stressors. Loss of keystone herbivores is commonly cited as a key underlying driver of coral-algal phase shifts [9,73]. Multiple studies identify the 1983 – 1984 massive die-off of the herbivorous urchin Diadema antillarum as the prime cause of the proliferation of macroalgae in Caribbean reefs [4,8]. Before 1983, D. antillarum was common on coral reefs of the Caribbean Coast of Costa Rica, with estimated densities of 3.6 – 8.8 ind. m-2 for reefs in Cahuita [74]. After the massive die-off in 1983, Diadema densities reduced to 0.2 – 2 ind. m-2 [38]. In 1992, very low densities (0.01 ind. m-2) were observed and between 1999 and 2003 densities ranged between 0.3 and 0.7 ind. m-2 [34,75]. Almost 40 years later, we found that Diadema densities for the whole region are very low (0.05 ind. m-2), and the population doesn’t seem to have recovered. The lack of recovery of this keystone herbivore may be one of the principal factors driving the high macroalgae covers found in Costa Rican’s Caribbean reefs, which concurs with the significantly higher urchin densities found on reefs with low fleshy macroalgae covers. Further evidence is that macroalgae cover declined and coral recruitment increased on Caribbean reefs where D. antillarum densities recovered [76,77,78,79]. Also, D. antillarum seem to have an affinity for brown macroalgae of the genus Dictyota, which may also be evidence that their low abundance is driving the algae dominance found on the reefs, because Dictyota / Dictyopteris spp. were the algae species that contributed the most to the reported fleshy macroalgae covers [80].
The lack of recovery of the populations of D. antillarum seems to be affecting the whole Caribbean basin [81,82]. Lessios [81] reports that the current population densities are approximately 12% of those before the die-off. The factors constraining the recovery are unclear, but it’s believed to be associated with recruitment limitations [81]. As a response to the low recovery, multiple restoration studies of D. antillarum have been attempted, and most have shown positive results in reducing macroalgae. Restoration of keystone herbivores could be an interesting managing strategy to try to control the high macroalgae abundance found on the reefs of the Caribbean coast of Costa Rica. But first, further studies are needed to assess current settlement, survival, and herbivory rates of not only D. antillarum but other herbivorous invertebrates like Maguimithrax crabs and Tripneustes, and Eucidaris sea urchins, which could also contribute to the recovery of coral reefs [56,83].
The herbivore fish biomass estimated for Costa Rican Caribbean reefs appear to be among the lower values reported for the Caribbean region [17,59]. Reduce populations of herbivorous fishes due to overfishing has been cited as a major factor driving the algae dominance in coral reefs [8,84,85]. If this is true, coral reefs with high herbivore abundance would be expected to have lower macroalgae and higher coral covers [73]. However, we didn’t fine significant differences in fish herbivore biomass between reefs with different levels of coral and macroalgae cover. Moreover, if overfishing is a prime factor driving the high macroalgae covers, we would expect to see a significantly higher abundance of fish herbivores inside MPA, which wasn’t the case. In addition, although there is not much information about the status of reefs fisheries in the Caribbean of Costa Rica, herbivorous fishes doesn’t appear to be main fishing targets [86]. Our results concur with recent studies that indicate that coral-algae phase shifts are not driven simply by declines in herbivores, which have been increasing or stable given the use of marine protected areas and fishing bans [59,87].
Mounting evidence suggests that the high macroalgae abundance in Caribbean reefs is driven by elevated nutrient enrichment in coastal waters due to coastal development [3,88,89,90,91]. Pollution and sediment loading due to coastal development can alter the reef benthos through light attenuation, smothering, and eutrophication [92]. Increased sediments can also deter herbivores from grazing and may displace them entirely [93,94]. Increased terrigenous sediment loading has been previously identified as the most important anthropogenic impact on coral reefs of the Caribbean of Costa Rica [25,31]. It´s believed that the increase in terrigenous sediments was caused by extensive logging and the establishment of banana plantations in the area [25]. Water quality assessments around Cahuita and Isla Uvita [95,96] recorded nitrite and nitrate concentrations which were much higher that the reported values for reefs in Panama and Florida [3,97,98]. In Cahuita, average nitrite concentrations in 2017-2018, were higher than in 2005-2004 and 1997, reflecting an increased nitrogen loading [95,99,100]. Sanitary and water quality assessments of rivers and streams along the Caribbean coast of Costa Rica found that around 68 % of the water bodies surveyed were unsuitable for primary contact activities or as water supply sources [101]. These studies suggest that the water quality in Costa Rica’s Caribbean may not be ideal for coral reef development. Thus, bottom-up control via nutrient enrichment and sediment loading could explain the fleshy algae dominance found on the reefs. To assess that future studies that quantify the water quality and nutrient enrichment along the Caribbean coast of Costa Rica and explore their relationship with the reef communities are needed. Also, as previously discuss by Cortés and Jimenez [31], an integrate management approach that includes the watersheds and the marine environment must be taken in order to control the water quality problem.
In conclusion, fleshy macroalgae exceed coral cover in most coral reefs of the Caribbean coast of Costa Rica. Higher fleshy macroalgae covers were associated with lower substrate rugosity, urchin density, fish richness, total fish biomass, large fish density, and the potential fishery value of the reef. In agreement with existing literature, the high macroalgae cover may be driven by low densities of herbivorous urchins and water pollution from coastal development. The low average values of the ecosystem metrics and the apparent lack of variability of the fish community metrics between sites and regions despite the presence of marine protected areas (MPA) suggest that the current management actions implemented to protect the coral reefs are insufficient. In the future, management actions to assist in the recovery of herbivore populations and to reduce water pollution may prove useful to reduce the fleshy macroalgae cover and increase the resilience of the coral reefs. It is crucial to continue monitoring the status of the reef communities to evaluate the effectiveness of existing and future management actions.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Survey site coordinates and mean depth; Table S2: Coral species registered in the survey sites of the Caribbean coast of Costa Rica; Table S4: Analysis of Similarities (SIMPER) of benthic taxa between regions; Table S4: Fish species registered in the survey sites of the Caribbean coast of Costa Rica.
Author Contributions
All authors had significant contributions to this study. Conceptualization, F.Q., and J.J.A.; methodology, F.Q., S.M., C.F.G., and J.J.A.; software, F.Q., and S.M.; validation, F.Q. and J.J.A ; formal analysis, F.Q.; investigation, F.Q, S.M., C.F.G., and J.J.A.; resources, C.F.G., and J.J.A.; data curation, F.Q., and S.M.; writing—original draft preparation, F.Q., and J.J.A; writing—review and editing, F.Q., S.M., C.F.G., and J.J.A; visualization, F.Q.; supervision, J.J.A.; project administration, J.J.A..; funding acquisition, C.F.G., and J.J.A.. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financed through the Research Vice-Rectory of the University of Costa Rica, through project B9088.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are openly available in figshare at https://doi.org/10.6084/m9.figshare.23528196.v2
Acknowledgments
We gratefully acknowledge the support and collaboration of the staff of the Parque Nacional Cahuita, Refugio Nacional de Vida Silvestre Gandoca-Manzanillo and Área de Conservación La Amistad Caribe. We also would like to thank Andrea Arriaga Madrigal, María José Solana González, Randy Domínguez Miranda, and other assistants that help us with the data collection. This study was conducted under the following research permits ACLAC-DR-006-2019, R-SINAC-PNI-ACLAC-023-2021 and R-SINAC-SE-DT-PI-012-2023.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hughes, T.P.; Barnes, M.L.; Bellwood, D.R.; Cinner, J.E.; Cumming, G.S.; Jackson, J.B.C.; Kleypas, J.; van de Leemput, I.A.; Lough, J.M.; Morrison, T.H.; et al. Coral Reefs in the Anthropocene. Nature 2017, 546, 82–90. [Google Scholar] [CrossRef]
- Aronson, R.B.; Precht, W.F. White-Band Disease and the Changing Face of Caribbean Coral Reefs. In The Ecology and Etiology of Newly Emerging Marine Diseases; Porter, J.W., Ed.; Developments in Hydrobiology; Springer Netherlands: Dordrecht, 2001; pp. 25–38. ISBN 978-94-017-3284-0. [Google Scholar]
- Lapointe, B.E. Nutrient Thresholds for Bottom-up Control of Macroalgal Blooms on Coral Reefs in Jamaica and Southeast Florida. Limnol. Oceanogr. 1997, 42, 1119–1131. [Google Scholar] [CrossRef]
- Mumby, P.J.; Hedley, J.D.; Zychaluk, K.; Harborne, A.R.; Blackwell, P.G. Revisiting the Catastrophic Die-off of the Urchin Diadema Antillarum on Caribbean Coral Reefs: Fresh Insights on Resilience from a Simulation Model. Ecol. Model. 2006, 196, 131–148. [Google Scholar] [CrossRef]
- Muñiz-Castillo, A.I.; Rivera-Sosa, A.; Chollett, I.; Eakin, C.M.; Andrade-Gómez, L.; McField, M.; Arias-González, J.E. Three Decades of Heat Stress Exposure in Caribbean Coral Reefs: A New Regional Delineation to Enhance Conservation. Sci. Rep. 2019, 9, 11013. [Google Scholar] [CrossRef] [PubMed]
- Randazzo-Eisemann, Á.; Garza-Pérez, J.R.; Penié-Rodriguez, I.; Figueroa-Zavala, B. 25 Years of Multiple Stressors Driving the Coral-Algae Phase Shift in Akumal, Mexico. Ocean Coast. Manag. 2021, 214, 105917. [Google Scholar] [CrossRef]
- Contreras-Silva, A.I.; Tilstra, A.; Migani, V.; Thiel, A.; Pérez-Cervantes, E.; Estrada-Saldívar, N.; Elias-Ilosvay, X.; Mott, C.; Alvarez-Filip, L.; Wild, C. A Meta-Analysis to Assess Long-Term Spatiotemporal Changes of Benthic Coral and Macroalgae Cover in the Mexican Caribbean. Sci. Rep. 2020, 10, 8897. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P. Catastrophes, Phase Shifts, and Large-Scale Degradation of a Caribbean Coral Reef. Science 1994, 265, 1547–1551. [Google Scholar] [CrossRef]
- Mumby, P.J. Phase Shifts and the Stability of Macroalgal Communities on Caribbean Coral Reefs. Coral Reefs 2009, 28, 761–773. [Google Scholar] [CrossRef]
- Alvarez-Filip, L.; Dulvy, N.K.; Gill, J.A.; Côté, I.M.; Watkinson, A.R. Flattening of Caribbean Coral Reefs: Region-Wide Declines in Architectural Complexity. Proc. R. Soc. B Biol. Sci. 2009, 276, 3019–3025. [Google Scholar] [CrossRef]
- Alvarez-Filip, L.; Gill, J.A.; Dulvy, N.K.; Perry, A.L.; Watkinson, A.R.; Côté, I.M. Drivers of Region-Wide Declines in Architectural Complexity on Caribbean Reefs. Coral Reefs 2011, 30, 1051–1060. [Google Scholar] [CrossRef]
- Newman, S.P.; Meesters, E.H.; Dryden, C.S.; Williams, S.M.; Sanchez, C.; Mumby, P.J.; Polunin, N.V.C. Reef Flattening Effects on Total Richness and Species Responses in the Caribbean. J. Anim. Ecol. 2015, 84, 1678–1689. [Google Scholar] [CrossRef] [PubMed]
- Chong-Seng, K.M.; Mannering, T.D.; Pratchett, M.S.; Bellwood, D.R.; Graham, N.A.J. The Influence of Coral Reef Benthic Condition on Associated Fish Assemblages. PLOS ONE 2012, 7, e42167. [Google Scholar] [CrossRef]
- Pratchett, M.S.; Hoey, A.S.; Wilson, S.K. Reef Degradation and the Loss of Critical Ecosystem Goods and Services Provided by Coral Reef Fishes. Curr. Opin. Environ. Sustain. 2014, 7, 37–43. [Google Scholar] [CrossRef]
- Smith, J.E.; Brainard, R.; Carter, A.; Grillo, S.; Edwards, C.; Harris, J.; Lewis, L.; Obura, D.; Rohwer, F.; Sala, E.; et al. Re-Evaluating the Health of Coral Reef Communities: Baselines and Evidence for Human Impacts across the Central Pacific. Proc. R. Soc. B Biol. Sci. 2016, 283, 20151985. [Google Scholar] [CrossRef] [PubMed]
- Woodhead, A.J.; Hicks, C.C.; Norström, A.V.; Williams, G.J.; Graham, N.A.J. Coral Reef Ecosystem Services in the Anthropocene. Funct. Ecol. 2019, 33, 1023–1034. [Google Scholar] [CrossRef]
- Lester, S.E.; Rassweiler, A.; McCoy, S.J.; Dubel, A.K.; Donovan, M.K.; Miller, M.W.; Miller, S.D.; Ruttenberg, B.I.; Samhouri, J.F.; Hay, M.E. Caribbean Reefs of the Anthropocene: Variance in Ecosystem Metrics Indicates Bright Spots on Coral Depauperate Reefs. Glob. Change Biol. 2020, 26, 4785–4799. [Google Scholar] [CrossRef]
- Robinson, J.P.W.; Wilson, S.K.; Robinson, J.; Gerry, C.; Lucas, J.; Assan, C.; Govinden, R.; Jennings, S.; Graham, N.A.J. Productive Instability of Coral Reef Fisheries after Climate-Driven Regime Shifts. Nat. Ecol. Evol. 2019, 3, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Bruno, J.F.; Sweatman, H.; Precht, W.F.; Selig, E.R.; Schutte, V.G.W. Assessing Evidence of Phase Shifts from Coral to Macroalgal Dominance on Coral Reefs. Ecology 2009, 90, 1478–1484. [Google Scholar] [CrossRef] [PubMed]
- Norström, A.V.; Nyström, M.; Lokrantz, J.; Folke, C. Alternative States on Coral Reefs: Beyond Coral–Macroalgal Phase Shifts. Mar. Ecol. Prog. Ser. 2009, 376, 295–306. [Google Scholar] [CrossRef]
- Mcleod, E.; Anthony, K.R.N.; Mumby, P.J.; Maynard, J.; Beeden, R.; Graham, N.A.J.; Heron, S.F.; Hoegh-Guldberg, O.; Jupiter, S.; MacGowan, P.; et al. The Future of Resilience-Based Management in Coral Reef Ecosystems. J. Environ. Manage. 2019, 233, 291–301. [Google Scholar] [CrossRef]
- Williams, S.M.; Chollett, I.; Roff, G.; Cortés, J.; Dryden, C.S.; Mumby, P.J. Hierarchical Spatial Patterns in Caribbean Reef Benthic Assemblages. J. Biogeogr. 2015, 42, 1327–1335. [Google Scholar] [CrossRef]
- Spalding, M.D.; Fox, H.E.; Allen, G.R.; Davidson, N.; Ferdaña, Z.A.; Finlayson, M.; Halpern, B.S.; Jorge, M.A.; Lombana, A.; Lourie, S.A.; et al. Marine Ecoregions of the World: A Bioregionalization of Coastal and Shelf Areas. BioScience 2007, 57, 573–583. [Google Scholar] [CrossRef]
- Aronson, R.B.; Hilbun, N.L.; Bianchi, T.S.; Filley, T.R.; McKee, B.A. Land Use, Water Quality, and the History of Coral Assemblages at Bocas Del Toro, Panamá. Mar. Ecol. Prog. Ser. 2014, 504, 159–170. [Google Scholar] [CrossRef]
- Cortés, J.N.; Risk, M.J. A Reef under Siltation Stress: Cahuita, Costa Rica. Bull. Mar. Sci. 1985, 36, 339–356. [Google Scholar]
- Cramer, K.L.; O’Dea, A.; Leonard-Pingel, J.S.; Norris, R.D. Millennial-Scale Change in the Structure of a Caribbean Reef Ecosystem and the Role of Human and Natural Disturbance. Ecography 2020, 43, 283–293. [Google Scholar] [CrossRef]
- Restrepo, J.D.; Park, E.; Aquino, S.; Latrubesse, E.M. Coral Reefs Chronically Exposed to River Sediment Plumes in the Southwestern Caribbean: Rosario Islands, Colombia. Sci. Total Environ. 2016, 553, 316–329. [Google Scholar] [CrossRef]
- Cortés, J.; Jiménez, C.E.; Fonseca, A.C.; Alvarado, J.J. Status and Conservation of Coral Reefs in Costa Rica. Rev. Biol. Trop. 2010, 58, 33–50. [Google Scholar] [CrossRef]
- Guzmán, H.M. Caribbean Coral Reefs of Panama: Present Status and Future Perspectives. In Latin American Coral Reefs; Cortés, J., Ed.; Elsevier Science: Amsterdam, 2003; pp. 241–274. ISBN 978-0-444-51388-5. [Google Scholar]
- Rivas, N.; Gómez, C.E.; Millán, S.; Mejía-Quintero, K.; Chasqui, L. Coral Reef Degradation at an Atoll of the Western Colombian Caribbean. PeerJ 2023, 11, e15057. [Google Scholar] [CrossRef]
- Cortés, J.; Jiménez, C. Past, Present and Future of the Coral Reefs of the Caribbean Coast of Costa Rica. In Latin American Coral Reefs; Cortés, J., Ed.; Elsevier Science: Amsterdam, 2003; pp. 223–239. ISBN 978-0-444-51388-5. [Google Scholar]
- G.m, W. The Benthic Flora of Punta Cahuita: An Annotated List of Species with New Additions to the Costa Rican Atlantic Flora [Algae]. Brenesia 1974. [Google Scholar]
- Risk, M.J.; Murillo, M.M.; Cortés, J. Observaciones biológicas preliminares sobre el arrecife coralino en el Parque Nacional de Cahuita, Costa Rica. Rev. Biol. Trop. 1980, 28, 361–382. [Google Scholar]
- Cortés, J. A Reef under Siltation Stress: A Decade of Degradation. Biol. Conserv. 1996, 76, 215–215. [Google Scholar] [CrossRef]
- Cortés, J.; Soto, R.; Jimenez, C.; Astorga, A. Earthquake Associated Mortality of Intertidal and Coral Reef Organisms (Caribbean of Costa Rica). Proc. 7th Int. Coral Reef Symp. 1992, 1, 235–240. [Google Scholar]
- Guzmán, H.M.; Cortés, J. Mortandad de Gorgonia flabellum Linnaeus (Octocorallia: Gorgoniidae) en la Costa del Caribe de Costa Rica. Rev. Biol. Trop. 1984, 32, 305–308. [Google Scholar]
- Guzmán, H.M.; Jiménez, C.E. Contamination of Coral Reefs by Heavy Metals along the Caribbean Coast of Central America (Costa Rica and Panama). Mar. Pollut. Bull. 1992, 24, 554–561. [Google Scholar] [CrossRef]
- Murillo, M.M.; Cortés, J. Alta mortalidad en la población del erizo de mar Diadema antillarum Philippi (Echinodermata: Echinoidea), en el Parque Nacional Cahuita, Limón,Costa Rica. Rev. Biol. Trop. 1984, 32, 167–169. [Google Scholar]
- Cortés, J.; Fonseca, A.C.; Nivia-Ruiz, J.; Nielsen-Muñoz, V.; Samper-Villarreal, J.; Salas, E.; Martínez, S.; Zamora-Trejos, P. Monitoring Coral Reefs, Seagrasses and Mangroves in Costa Rica (CARICOMP). Rev. Biol. Trop. 2010, 58, 1–22. [Google Scholar] [PubMed]
- Cortés, J.; Oxenford, H.A.; van Tussenbroek, B.I.; Jordán-Dahlgren, E.; Cróquer, A.; Bastidas, C.; Ogden, J.C. The CARICOMP Network of Caribbean Marine Laboratories (1985–2007): History, Key Findings, and Lessons Learned. Front. Mar. Sci. 2019, 5. [Google Scholar] [CrossRef]
- Cortés-Núñez, J.; Guzmán-Espinal, H.M. Arrecifes Coralinos de La Costa Atlántica de Costa Rica. Brenesia. 1985, 275–292. [Google Scholar]
- Fernández, C.; Alvarado, J.J. El arrecife coralino de Punta Cocles, costa Caribe de Costa Rica. Rev. Biol. Trop. 2004, 52, 121–129. [Google Scholar]
- Fonseca, A. A Rapid Assessment at Cahuita National Park, Costa Rica, 1999 (Part 1: Stony Corals and Algae); Smithsonian Institution Press, 2003. [Google Scholar]
- Araya-Vargas, A.; Nova-Bustos, N. Health Status Evaluation of Shallow Coral Reefs in Cahuita and Manzanillo, Costa Rica. Biodivers. Nat. Hist. 2017, 3, 48–55. [Google Scholar]
- Williams, S.; Mumby, P.; Chollett, I.; Cortés, J. Importance of Differentiating Orbicella Reefs from Gorgonian Plains for Ecological Assessments of Caribbean Reefs. Mar. Ecol. Prog. Ser. 2015, 530. [Google Scholar] [CrossRef]
- Trygonis, V.; Sini, M. PhotoQuad: A Dedicated Seabed Image Processing Software, and a Comparative Error Analysis of Four Photoquadrat Methods. J. Exp. Mar. Biol. Ecol. 2012, 424–425, 99–108. [Google Scholar] [CrossRef]
- Aronson, R.B.; Precht, W.F. Landscape Patterns of Reef Coral Diversity: A Test of the Intermediate Disturbance Hypothesis. J. Exp. Mar. Biol. Ecol. 1995, 192, 1–14. [Google Scholar] [CrossRef]
- Alvarado, J.J.; Beita-Jiménez, A.; Mena, S.; Fernández-García, C.; Guzman-Mora, A.G.; Cortés, J. Ecosistemas coralinos del Parque Nacional Isla del Coco, Costa Rica: estructura y comparación 1987-2014. Rev. Biol. Trop. 2016, 64, 153–175. [Google Scholar] [CrossRef]
- Moberg, F.; Folke, C. Ecological Goods and Services of Coral Reef Ecosystems. Ecol. Econ. 1999, 29, 215–233. [Google Scholar] [CrossRef]
- Uyarra, M.C.; Watkinson, A.R.; Côté, I.M. Managing Dive Tourism for the Sustainable Use of Coral Reefs: Validating Diver Perceptions of Attractive Site Features. Environ. Manage. 2009, 43, 1–16. [Google Scholar] [CrossRef]
- Imamura, K.; Takano, K.T.; Kumagai, N.H.; Yoshida, Y.; Yamano, H.; Fujii, M.; Nakashizuka, T.; Managi, S. Valuation of Coral Reefs in Japan: Willingness to Pay for Conservation and the Effect of Information. Ecosyst. Serv. 2020, 46, 101166. [Google Scholar] [CrossRef]
- Rogers, C.S. Coral Reef Resilience through Biodiversity. Int. Sch. Res. Not. 2013, 2013, e739034. [Google Scholar] [CrossRef]
- Clements, C.S.; Hay, M.E. Biodiversity Enhances Coral Growth, Tissue Survivorship and Suppression of Macroalgae. Nat. Ecol. Evol. 2019, 3, 178–182. [Google Scholar] [CrossRef]
- Graham, N.A.J.; Nash, K.L. The Importance of Structural Complexity in Coral Reef Ecosystems. Coral Reefs 2013, 32, 315–326. [Google Scholar] [CrossRef]
- Ferrario, F.; Beck, M.W.; Storlazzi, C.D.; Micheli, F.; Shepard, C.C.; Airoldi, L. The Effectiveness of Coral Reefs for Coastal Hazard Risk Reduction and Adaptation. Nat. Commun. 2014, 5, 3794. [Google Scholar] [CrossRef]
- Francis, F.T.; Filbee-Dexter, K.; Yan, H.F.; Côté, I.M. Invertebrate Herbivores: Overlooked Allies in the Recovery of Degraded Coral Reefs? Glob. Ecol. Conserv. 2019, 17, e00593. [Google Scholar] [CrossRef]
- Grafeld, S.; Oleson, K.; Barnes, M.; Peng, M.; Chan, C.; Weijerman, M. Divers’ Willingness to Pay for Improved Coral Reef Conditions in Guam: An Untapped Source of Funding for Management and Conservation? Ecol. Econ. 2016, 128, 202–213. [Google Scholar] [CrossRef]
- Williams, I.D.; Polunin, N.V.C. Differences between Protected and Unprotected Reefs of the Western Caribbean in Attributes Preferred by Dive Tourists. Environ. Conserv. 2000, 27, 382–391. [Google Scholar] [CrossRef]
- Arias-González, J.E.; Fung, T.; Seymour, R.M.; Garza-Pérez, J.R.; Acosta-González, G.; Bozec, Y.-M.; Johnson, C.R. A Coral-Algal Phase Shift in Mesoamerica Not Driven by Changes in Herbivorous Fish Abundance. PLOS ONE 2017, 12, e0174855. [Google Scholar] [CrossRef]
- Cheal, A.J.; MacNeil, M.A.; Cripps, E.; Emslie, M.J.; Jonker, M.; Schaffelke, B.; Sweatman, H. Coral–Macroalgal Phase Shifts or Reef Resilience: Links with Diversity and Functional Roles of Herbivorous Fishes on the Great Barrier Reef. Coral Reefs 2010, 29, 1005–1015. [Google Scholar] [CrossRef]
- Gill, D.A.; Schuhmann, P.W.; Oxenford, H.A. Recreational Diver Preferences for Reef Fish Attributes: Economic Implications of Future Change. Ecol. Econ. 2015, 111, 48–57. [Google Scholar] [CrossRef]
- Graham, N.; Dulvy, N.; Jennings, S.; Polunin, N. Size-Spectra as Indicators of the Effects of Fishing on Coral Reef Fish Assemblages. Coral Reefs 2005, 24, 118–124. [Google Scholar] [CrossRef]
- Froese, R.; Pauly, D. FishBase. World Wide Web Electronic Publication. Available online: https://www.fishbase.se/summary/citation.php (accessed on 13 June 2023).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression; SAGE Publications, 2018; ISBN 978-1-5443-3648-0. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing 2022.
- Souter, D.; Planes, S.; Wicquart, J.; Logan, M.; Obura, D. Status of Coral Reefs of the World: 2020 Report. Global Coral Reef Monitoring Network (GCRMN)/International Coral Reef Initiative (ICRI) 2021.
- McField, M.; Soto, M.; Craig, N.; Giro, A.; Drysdale, I.; Guerrero, C.; Rueda, M.; Kramer, P.; Canty, S.; Muñiz, I. 2022 Mesoamerican Reef Report Card. Healthy Reefs Initiative 2022.
- Huntington, B.E.; Miller, M.W.; Pausch, R.; Richter, L. Facilitation in Caribbean Coral Reefs: High Densities of Staghorn Coral Foster Greater Coral Condition and Reef Fish Composition. Oecologia 2017, 184, 247–257. [Google Scholar] [CrossRef]
- Newman, M.J.H.; Paredes, G.A.; Sala, E.; Jackson, J.B.C. Structure of Caribbean Coral Reef Communities across a Large Gradient of Fish Biomass. Ecol. Lett. 2006, 9, 1216–1227. [Google Scholar] [CrossRef] [PubMed]
- Santavy, D.L.; Courtney, L.A.; Fisher, W.S.; Quarles, R.L.; Jordan, S.J. Estimating Surface Area of Sponges and Gorgonians as Indicators of Habitat Availability on Caribbean Coral Reefs. Hydrobiologia 2013, 707, 1–16. [Google Scholar] [CrossRef]
- Wulff, J. Sponge Contributions to the Geology and Biology of Reefs: Past, Present, and Future. In Coral Reefs at the Crossroads; Hubbard, D.K., Rogers, C.S., Lipps, J.H., Stanley, Jr., George, D., Eds.; Coral Reefs of the World; Springer Netherlands: Dordrecht, 2016; pp. 103–126. ISBN 978-94-017-7567-0. [Google Scholar]
- Bell, J.J.; Rovellini, A.; Davy, S.K.; Taylor, M.W.; Fulton, E.A.; Dunn, M.R.; Bennett, H.M.; Kandler, N.M.; Luter, H.M.; Webster, N.S. Climate Change Alterations to Ecosystem Dominance: How Might Sponge-Dominated Reefs Function? Ecology 2018, 99, 1920–1931. [Google Scholar] [CrossRef]
- Mumby, P.J.; Hastings, A.; Edwards, H.J. Thresholds and the Resilience of Caribbean Coral Reefs. Nature 2007, 450, 98–101. [Google Scholar] [CrossRef]
- Valdez, M.F.; Villalobos, C.R. Distribución espacial, correlación con el substrato y grado de agregación en Diadema antillarum Philippi (Echinodermata: Echinoidea). Rev. Biol. Trop. 1978, 26, 237–245. [Google Scholar]
- Alvarado, J.J.; Cortes, J.; Salas, E. Population Densities of Diadema Antillarum Philippi at Cahuita National Park (1977-2003), Costa Rica. Carib J Sci 2004, 40, 257–259. [Google Scholar]
- Carpenter, R.C.; Edmunds, P.J. Local and Regional Scale Recovery of Diadema Promotes Recruitment of Scleractinian Corals. Ecol. Lett. 2006, 9, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Edmunds, P.J.; Carpenter, R.C. Recovery of Diadema Antillarum Reduces Macroalgal Cover and Increases Abundance of Juvenile Corals on a Caribbean Reef. Proc. Natl. Acad. Sci. 2001, 98, 5067–5071. [Google Scholar] [CrossRef] [PubMed]
- Idjadi, J.A.; Haring, R.N.; Precht, W.F. Recovery of the Sea Urchin Diadema Antillarum Promotes Scleractinian Coral Growth and Survivorship on Shallow Jamaican Reefs. Mar. Ecol. Prog. Ser. 2010, 403, 91–100. [Google Scholar] [CrossRef]
- Myhre, S.; Acevedo-Gutiérrez, A. Recovery of Sea Urchin Diadema Antillarum Populations Is Correlated to Increased Coral and Reduced Macroalgal Cover. Mar. Ecol. Prog. Ser. 2007, 329, 205–210. [Google Scholar] [CrossRef]
- Williams, S.M. The Reduction of Harmful Algae on Caribbean Coral Reefs through the Reintroduction of a Keystone Herbivore, the Long-Spined Sea Urchin Diadema Antillarum. Restor. Ecol. 2022, 30, e13475. [Google Scholar] [CrossRef]
- Lessios, H. a. The Great Diadema Antillarum Die-Off: 30 Years Later. Annu. Rev. Mar. Sci. 2016, 8, 267–283. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Barreras, R.; Montañez-Acuña, A.; Otaño-Cruz, A.; Ling, S.D. Apparent Stability of a Low-Density Diadema Antillarum Regime for Puerto Rican Coral Reefs. ICES J. Mar. Sci. 2018, 75, 2193–2201. [Google Scholar] [CrossRef]
- Spadaro, A.J.; Butler, M.J. Herbivorous Crabs Reverse the Seaweed Dilemma on Coral Reefs. Curr. Biol. 2021, 31, 853–859.e3. [Google Scholar] [CrossRef]
- Jackson, J.B.C.; Kirby, M.X.; Berger, W.H.; Bjorndal, K.A.; Botsford, L.W.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.; Erlandson, J.; Estes, J.A.; et al. Historical Overfishing and the Recent Collapse of Coastal Ecosystems. Science 2001, 293, 629–637. [Google Scholar] [CrossRef]
- Pandolfi, J.M.; Bradbury, R.H.; Sala, E.; Hughes, T.P.; Bjorndal, K.A.; Cooke, R.G.; McArdle, D.; McClenachan, L.; Newman, M.J.H.; Paredes, G.; et al. Global Trajectories of the Long-Term Decline of Coral Reef Ecosystems. Science 2003, 301, 955–958. [Google Scholar] [CrossRef]
- Laguna Cruz, M.; Pereira-Chaves, J.; Ríos Duarte, R. Influencia del pez León (Pterois miles y Pterois volitans) en la cadena de valor de la pesca artesanal, Caribe sur de Costa Rica. UNED Res. J. 2019, 11. [Google Scholar] [CrossRef]
- Suchley, A.; McField, M.D.; Alvarez-Filip, L. Rapidly Increasing Macroalgal Cover Not Related to Herbivorous Fishes on Mesoamerican Reefs. PeerJ 2016, 4, e2084. [Google Scholar] [CrossRef]
- González-De Zayas, R.; Rossi, S.; Hernández-Fernández, L.; Velázquez-Ochoa, R.; Soares, M.; Merino-Ibarra, M.; Castillo-Sandoval, F.S.; Soto-Jiménez, M.F. Stable Isotopes Used to Assess Pollution Impacts on Coastal and Marine Ecosystems of Cuba and México. Reg. Stud. Mar. Sci. 2020, 39, 101413. [Google Scholar] [CrossRef]
- Lapointe, B.E.; Brewton, R.A.; Herren, L.W.; Porter, J.W.; Hu, C. Nitrogen Enrichment, Altered Stoichiometry, and Coral Reef Decline at Looe Key, Florida Keys, USA: A 3-Decade Study. Mar. Biol. 2019, 166, 108. [Google Scholar] [CrossRef]
- Lapointe, B.E.; Tewfik, A.; Phillips, M. Macroalgae Reveal Nitrogen Enrichment and Elevated N:P Ratios on the Belize Barrier Reef. Mar. Pollut. Bull. 2021, 171, 112686. [Google Scholar] [CrossRef]
- Wakwella, A.; Mumby, P.J.; Roff, G. Sedimentation and Overfishing Drive Changes in Early Succession and Coral Recruitment. Proc. R. Soc. B Biol. Sci. 2020, 287, 20202575. [Google Scholar] [CrossRef] [PubMed]
- Sura, S.A.; Bell, A.; Kunes, K.L.; Turba, R.; Songer, R.; Fong, P. Responses of Two Common Coral Reef Macroalgae to Nutrient Addition, Sediment Addition, and Mechanical Damage. J. Exp. Mar. Biol. Ecol. 2021, 536, 151512. [Google Scholar] [CrossRef]
- Gordon, S.E.; Goatley, C.H.R.; Bellwood, D.R. Low-Quality Sediments Deter Grazing by the Parrotfish Scarus Rivulatus on Inner-Shelf Reefs. Coral Reefs 2016, 35, 285–291. [Google Scholar] [CrossRef]
- Wenger, A.S.; Harvey, E.; Wilson, S.; Rawson, C.; Newman, S.J.; Clarke, D.; Saunders, B.J.; Browne, N.; Travers, M.J.; Mcilwain, J.L.; et al. A Critical Analysis of the Direct Effects of Dredging on Fish. Fish Fish. 2017, 18, 967–985. [Google Scholar] [CrossRef]
- Samper-Villarreal, J.; Sagot-Valverde, J.G.; Gómez-Ramírez, E.H.; Cortés, J. Water Quality as a Potential Factor Influencing Seagrass Change Over Time at Cahuita National Park, Costa Rica. Caribb. J. Sci. 2021, 51, 72–85. [Google Scholar] [CrossRef]
- Alfaro-Sandí, J.; Piedra-Marín, G.; Saravia-Arguedas, A.Y.; Piedra-Castro, L.; Alfaro-Sandí, J.; Piedra-Marín, G.; Saravia-Arguedas, A.Y.; Piedra-Castro, L. Evaluación de los parámetros físicos y químicos del agua de mar en los alrededores de la Isla Uvita, Limón, Costa Rica. Rev. Tecnol. En Marcha 2021, 34, 88–95. [Google Scholar] [CrossRef]
- D’Croz, L.; Rosario, J.B. del; Gondola, P. The Effect of Fresh Water Runoff on the Distribution of Dissolved Inorganic Nutrients and Plankton in the Bocas Del Toro Archipelago, Caribbean Panama. Caribb. J. Sci. 2005. [Google Scholar]
- D’Croz, L.; Robertson, D.R. Coastal Oceanographic Conditions Affecting Coral Reefs on Both Sides of the Isthmus of Panama. In Proceedings of the Proceedings of the 8th International Coral Reef Symposium; 1997; Volume 2, pp. 3–205. [Google Scholar]
- Roder, C.; Cortés, J.; Jiménez, C.; Lara, R. Riverine Input of Particulate Material and Inorganic Nutrients to a Coastal Reef Ecosystem at the Caribbean Coast of Costa Rica. Mar. Pollut. Bull. 2009, 58, 1937–1943. [Google Scholar] [CrossRef]
- Muller-Parker, G.; Cortés, J. Spatial Distribution of Light and Nutrients in Some Coral Reefs of Costa Rica during January 1997. Rev. Biol. Trop. 2001, 49, 251–263. [Google Scholar]
- Badilla-Aguilar, A.; Mora-Alvarado, D. Calidad sanitaria de las aguas superficiales en litorales de Costa Rica: situación del 2012 al 2018. Rev. Tecnol. En Marcha 2019. [Google Scholar] [CrossRef]
Figure 1.
(A) Study area and survey sites across the regions of; (B) Moín – Limón; (C) Cahuita; and (D) Puerto Viejo – Punta Mona. Survey site code can be found in the supplementary material Table S1.
Figure 1.
(A) Study area and survey sites across the regions of; (B) Moín – Limón; (C) Cahuita; and (D) Puerto Viejo – Punta Mona. Survey site code can be found in the supplementary material Table S1.
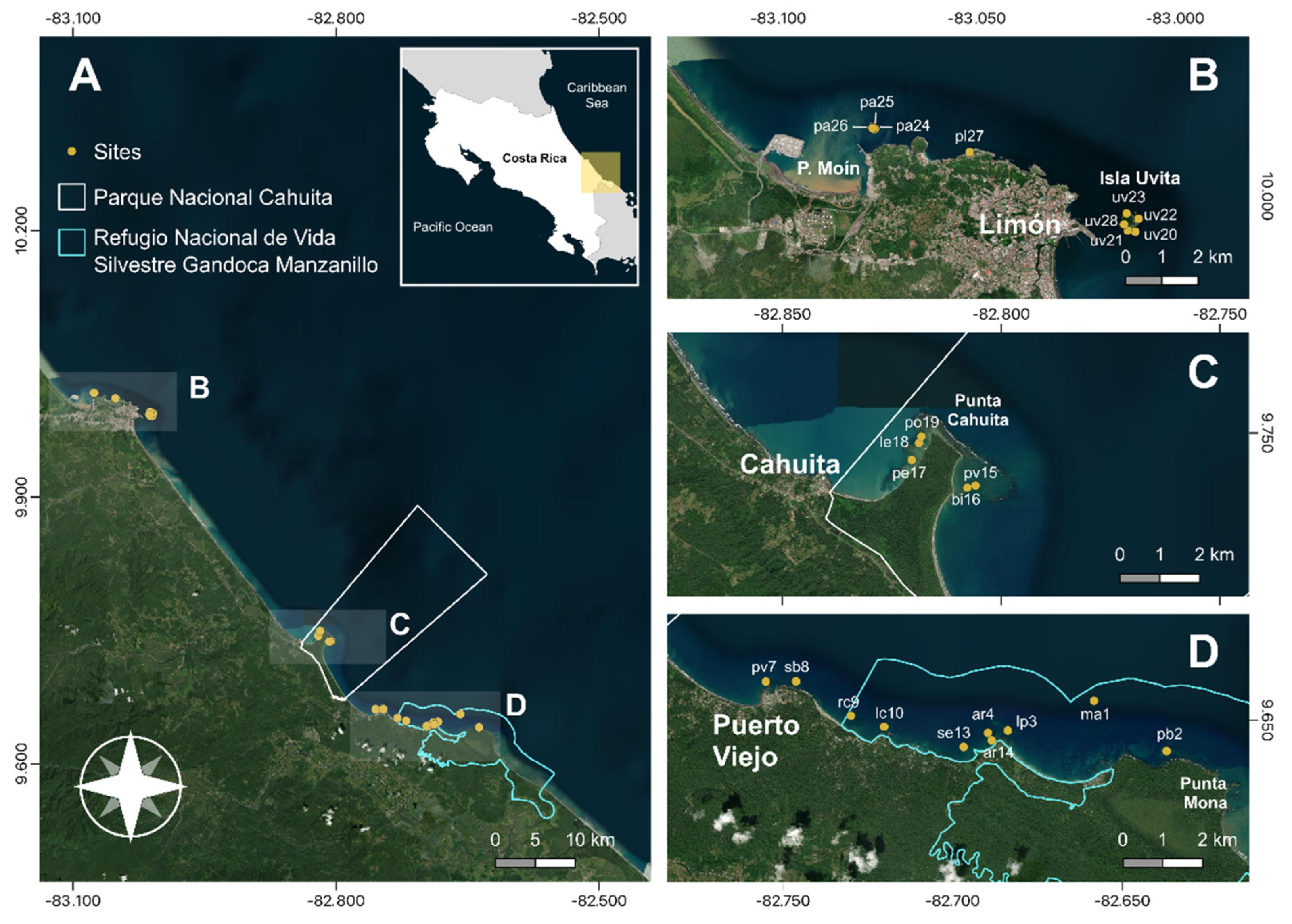
Figure 2.
(A) Percentage cover of benthic organisms in the reefs surveyed on the Caribbean Coast of Costa Rica. Each point represents the mean cover estimated for a survey site. ✕ symbol represents the mean cover between all survey sites. Calcareous macroalgae (Calc. Macroalgae). Crustose coralline algae (CCA); (B) Macroalgae species percentage contribution to the total fleshy macroalgae cover of the reefs surveyed; (C) Coral species percentage contribution to the total live coral cover of the reefs surveyed.
Figure 2.
(A) Percentage cover of benthic organisms in the reefs surveyed on the Caribbean Coast of Costa Rica. Each point represents the mean cover estimated for a survey site. ✕ symbol represents the mean cover between all survey sites. Calcareous macroalgae (Calc. Macroalgae). Crustose coralline algae (CCA); (B) Macroalgae species percentage contribution to the total fleshy macroalgae cover of the reefs surveyed; (C) Coral species percentage contribution to the total live coral cover of the reefs surveyed.
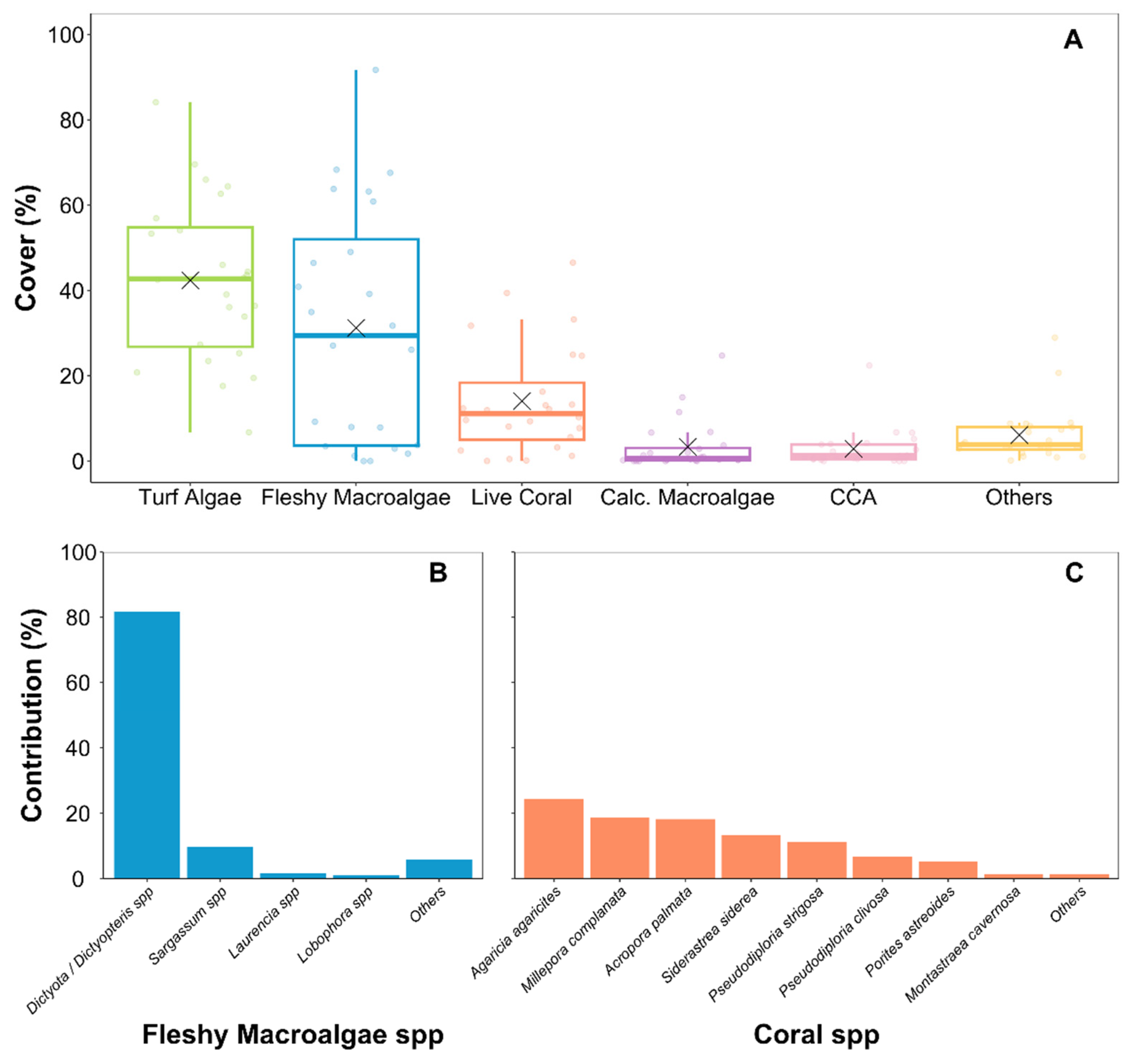
Figure 3.
A) Relationship between mean coral and fleshy macroalgal cover across all survey sites (± SE); (B) Relationship between mean reef builders (hard coral + CCA) and fleshy algae (turf algae + fleshy macroalgae) cover across all survey sites (± SE).
Figure 3.
A) Relationship between mean coral and fleshy macroalgal cover across all survey sites (± SE); (B) Relationship between mean reef builders (hard coral + CCA) and fleshy algae (turf algae + fleshy macroalgae) cover across all survey sites (± SE).
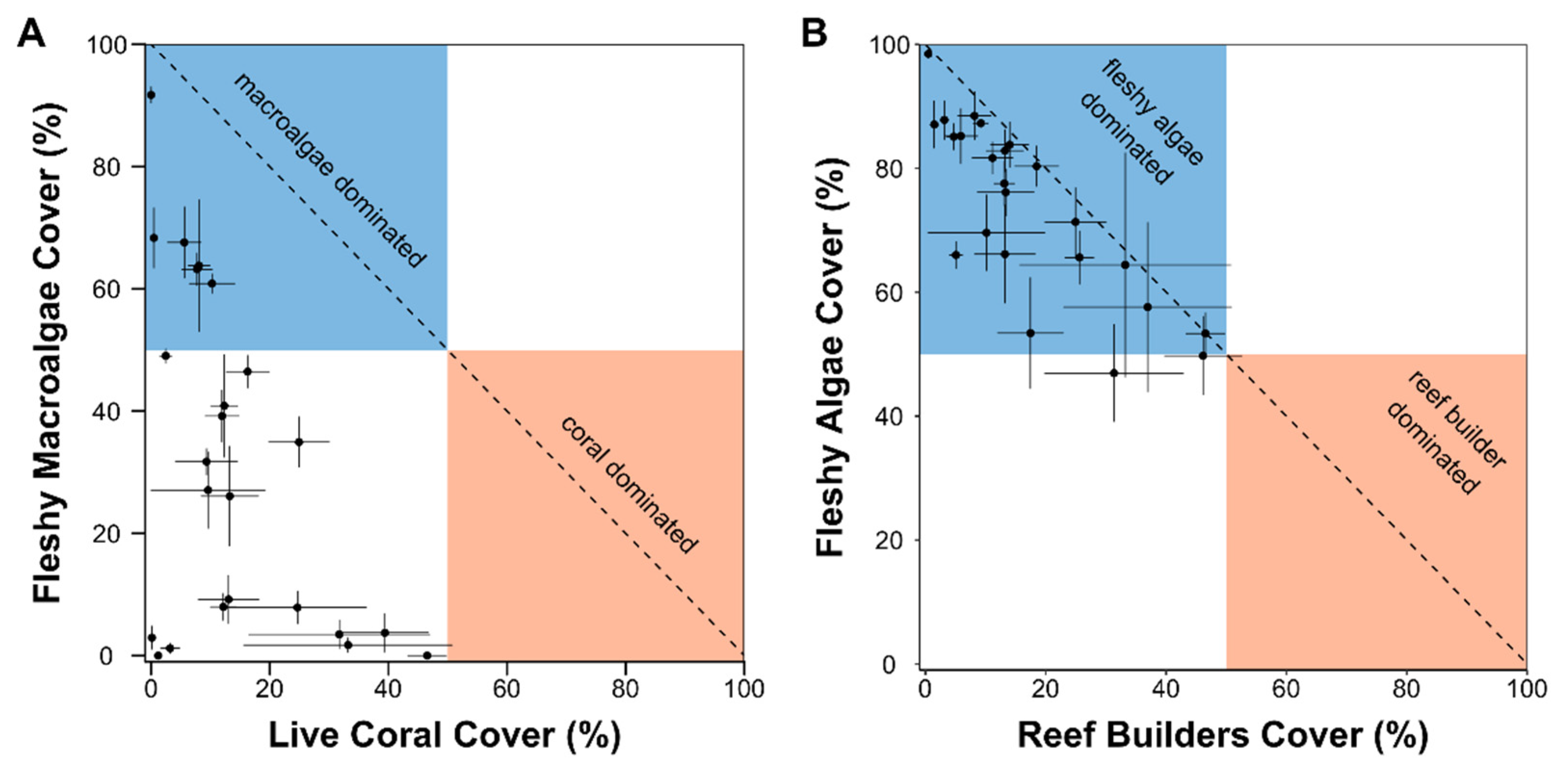
Figure 4.
(A) Estimated average cover of benthic organisms by survey site. Calcareous macroalgae (Calc. Macroalgae). Crustose coralline algae (CCA); (B) NMDS of the reef benthic communities on the three regions of reefs development in the Caribbean Coast of Costa Rica. Stress values and p-value of the PERMANOVA are shown.
Figure 4.
(A) Estimated average cover of benthic organisms by survey site. Calcareous macroalgae (Calc. Macroalgae). Crustose coralline algae (CCA); (B) NMDS of the reef benthic communities on the three regions of reefs development in the Caribbean Coast of Costa Rica. Stress values and p-value of the PERMANOVA are shown.
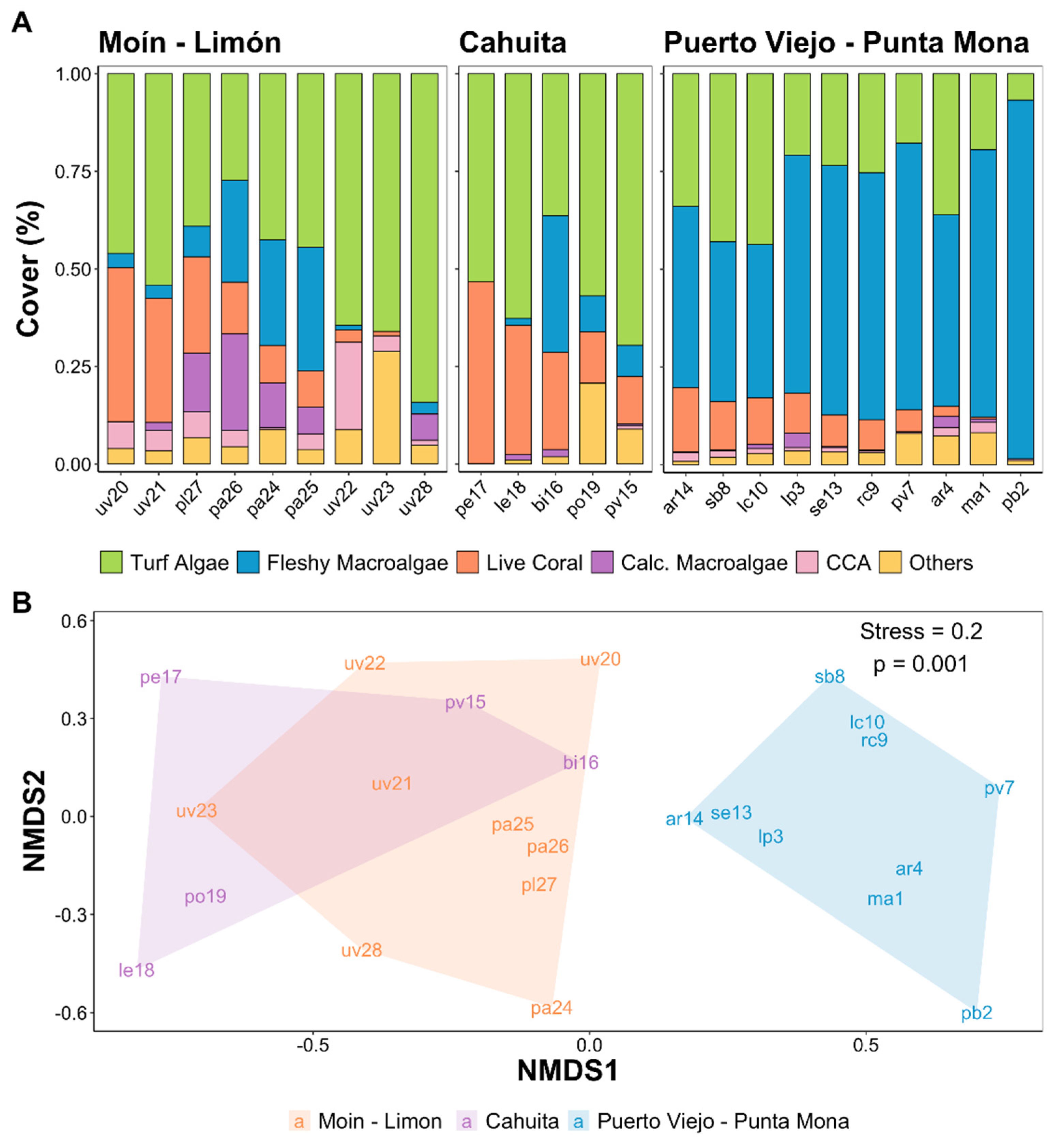
Figure 5.
Estimated ecosystem metrics by survey site. The horizontal solid black line represents the general mean among all sites and the dotted line the 95% confidence intervals. Sites categorized as bright and dark spots are painted orange and blue, respectively.
Figure 5.
Estimated ecosystem metrics by survey site. The horizontal solid black line represents the general mean among all sites and the dotted line the 95% confidence intervals. Sites categorized as bright and dark spots are painted orange and blue, respectively.
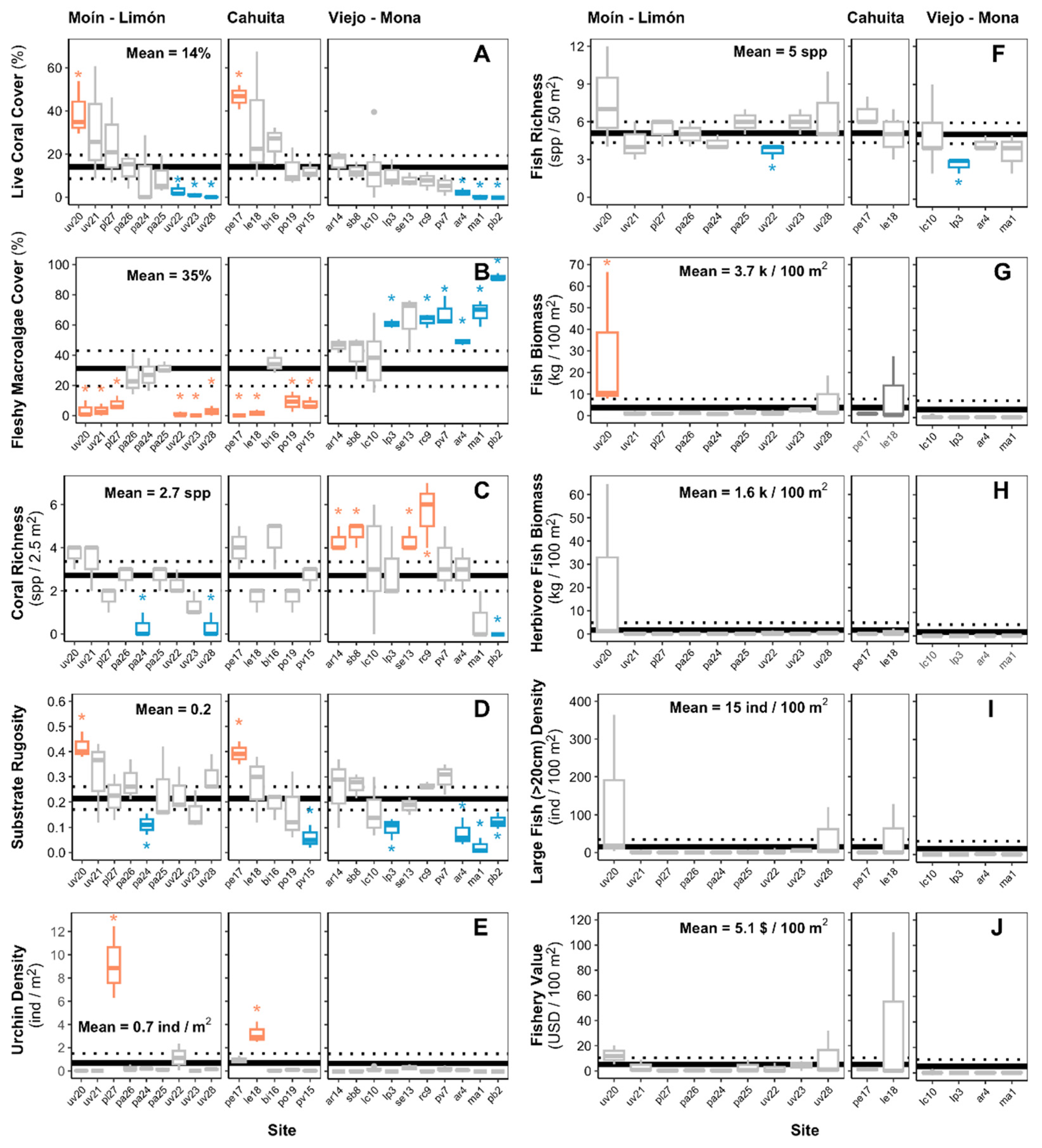
Figure 6.
Estimated ecosystem metrics by three categories of coral and fleshy macroalgae cover. High coral cover is defined as >20%, medium is 10%–20%, and low is <10%. High fleshy macroalgae cover is defined as >50%, medium is 50%–10%, and low is <10%. ✕ symbol represents the mean cover. The p-value from Kruskall-Wallis tests comparing the mean of the three categories are shown in each panel. The dotted lines between boxplots represent significant differences between the two cover categories based on Mann–Whitney tests.
Figure 6.
Estimated ecosystem metrics by three categories of coral and fleshy macroalgae cover. High coral cover is defined as >20%, medium is 10%–20%, and low is <10%. High fleshy macroalgae cover is defined as >50%, medium is 50%–10%, and low is <10%. ✕ symbol represents the mean cover. The p-value from Kruskall-Wallis tests comparing the mean of the three categories are shown in each panel. The dotted lines between boxplots represent significant differences between the two cover categories based on Mann–Whitney tests.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



