
Submitted:
27 June 2023
Posted:
28 June 2023
You are already at the latest version
Abstract
The ability of some bacteria to perform Extracellular Electron Transfer (EET) has been explored in bioelectrochemical systems (BES) to obtain energy or chemicals from pure substances or residual substrates. Here, a new pyoverdine-producing Pseudomonas aeruginosa strain was isolated from a MFC biofilm oxidizing glycerol, a by-product of biodiesel production. Strain EL14 was investigated to assess its electrogenic ability and products. In an open circuit system (fermentation system) EL14 was able to consume glycerol and produce 1,3-propanediol, an unusual product from glycerol oxidation in P. aeruginosa. The microbial fuel cell (MFC), EL14 reached a current density of 82.4 mA m-2, during the first feeding cycle, then drops sharply as the biofilm falls off. Cyclic voltammetry suggests electron transfer to the anode occurrs indirectly, i.e., through a redox substance, with redox peaks at 0.22 and -0.45 V (vs Ag/AgCl) and directly probably by membrane redox-proteins with redox peak at 0.05 V (vs Ag/AgCl). EL14 produced added-value bioproducts, acetic and butyric acids, as well as 1,3 propanediol, in both fermentative and anodic conditions. However, the yield of 1,3-PDO from glycerol was enhanced from 0.57 to 0.89 (mol of 1,3-PDO mol-1 of glycerol) under MFC conditions compared with the fermentation. This result was unexpected since successful 1,3-PDO production is not usually associated with the P. aeruginosa glycerol metabolism. By comparing EL14 genomic sequences related to the 1,3-PDO biosynthesis with reference P. aeruginosa strains, we observed that strain EL14 has three copies of dhaT gene (1,3-propanediol dehydrogenase a different arrangement compared to other Pseudomonas isolates). Thus, this work functionally characterizes a bacterium never before associated to 1,3-PDO biosynthesis, indicating its potential for converting a by-product of the biodiesel industry into an emerging chemical product.
Keywords:
Pseudomonas aeruginosa
; bioelectrochemical systems
; fermentation
; 1
; 3-propanediol
; microbial fuel cells
1. Introduction
Microbial Fuel Cells (MFCs) have emerged as an alternative technology for renewable energy generation. MFCs follow a similar concept to traditional fuel cells, but MFCs utilize the catalytic abilities of microorganisms and can use a variety of substrates, converting the energy stored in chemical bonds into electrical current [1]. In this scenario, glycerol is an attractive fuel to be oxidized in MFCs, as it is a versatile and abundant by-product of the biodiesel industry, corresponding to ca. 10% (w w-1) of total biodiesel produced [2]. Residual glycerol production for the year 2024 is estimated at 680.000 tonnes [3].
Several microorganisms can metabolize glycerol. The usual glycerol metabolism observed in Klebsiella, Clostridium, and Citrobacter genus regenerates NAD+ from the oxidative pathway coupled to 1,3 propanediol synthesis [4,5,6]. The reductive biochemical pathway converts glycerol to 3-hydroxipropionaldeyde (3-HPA) catalysed by glycerol dehydratase, which is then reduced to 1,3-PDO by 1,3-PDO dehydrogenase [4,5,6].
1,3-PDO is used as a monomer in the formation of polymers and directly in end-use products and, therefore, has a high commercial value [7]. Different 1,3-PDO production processes use petroleum derivatives, glucose, or glycerol [8]. However, most 1,3-PDO is synthesized from acrolein or ethylene oxide, both derived from fossil fuels. In the search to increase the competitiveness of biological 1,3-PDO production [4], efforts have been made to improve microbial efficiency in 1,3-PDO-producing microorganisms by metabolic engineering [9,10]. The use of glycerol as a substrate for obtaining 1,3-PDO is an attractive alternative, as it represents the use of a by-product of the biodiesel industry, contributing to the biorefinery concept of this production chain [3].
In previous work, the anodic compartment of a MFC inoculated with a mixed culture from a mining pond, was fed with glycerol [11]. After anode biofilm formation, the microbial community of the biofilm was mostly composed of Citrobacter, Pseudomonas, and Klebsiella, at average relative abundances of 45%, 31%, and 6%, respectively. During the MFC feed cycle, 1,3-PDO was produced in the anodic compartment, which was subsequently oxidized and converted to electricity. Both Citrobacter and Klebsiella are known to form 1,3-PDO from glycerol and it was assumed that the role of Pseudomonas was restricted to aiding in the external electrons transfer (EET) to the electrode. Indeed, Pseudomonas species demonstrated a significant role in providing electron shuttles to the anodic microbial communities in MFCs [11,12]. Pseudomonas produce phenazines that act as electron carriers and help other microorganis perform EET, and also transfer electrons from their own respiratory chain to the electrode [13].
In this work, the electrochemical and fermentative performances of a Pseudomonas aeruginosa strain EL14 isolated from an MFC anode fed with glycerol [11] were compared with reference strains. EL14 was used as the sole biocatalyst of an MFC bioanode with glycerol as substrate. The products profile and concentration generated in the anodic compartment of MFC were compared with a fermentative system (FE). Furthermore, genome sequencing of the isolate allowed the investigation of genes that could contribute to the formation of 1,3-PDO, an unusual product of P. aeruginosa glycerol metabolism.
2. Methodology
2.1. Bacteria isolation
Briefly, the isolation of potentially electrogenic bacteria was performed from an electrogenic biofilm formed on the anode of an MFC fed with glycerol [11]. The carbon cloth (9 cm2) anode was washed with 10 mL Lovley and Phillips [14] culture medium replacing acetate with 11mM glycerol and vortexed with glass beads. This suspension was serially diluted to 10-2 and streaked onto solid media of the same composition. The dishes were placed in anaerobic jars (Anaerogen®) until the appearance of colonies. One colony was picked and named EL14. A CFU resulting from the isolation step was cultured in Lovley liquid medium for DNA extraction.
2.2. Genome sequencing and identification of 1,3-PDO-associated genes
Total DNA of P. aeruginosa EL14 strain was extracted using the Wizard Genomic DNA Purification Kit (Promega®) according to the manufacturer’s instructions. The concentration of DNA was determined fluorometrically using the Qubit® 3.0 (Qubit® dsDNA Broad Range Assay Kit, Life Technologies, Carlsbad, CA, USA). DNA library was prepared using the Nextera XT DNA Library Prep Kit (Illumina, San Diego, CA, USA), assessed for quality using the 2100 Bioanalyzer (Agilent Genomics, Santa Clara, CA, USA), and subsequently submitted to sequencing using the Illumina HiSeq (2×150 bp) platform (Illumina, San Diego, CA, USA). Paired-end short reads were trimmed and assembled using SPAdes v.3.15.4 (Bankevich et al., 2012, Prjibelski et al., 2020). The completeness, quality, and contamination were assessed using Type Strain Genome Server (TYGS) [15]. Draft genomes were annotated using Prokka v.1.14.5. The P. aeruginosa EL14 genome is available at NCBI under GenBank accession number JASMRB000000000.
The identification of 1,3-PDO-associated genes (dhaT and dhaB) was first performed by BLASTn. Subsequently, the sequences were extracted, translated to protein, aligned, and compared to the DhaT amino acids sequence of P. aeruginosa localized into pathogenicity island PAGI-1 region (GenBank accession number AF241171) using Geneious Prime® v. 2023.1.2 (Biomatters Ltd.).
2.3. Cultivation of the isolate and culture medium
The EL14 strain was stored as a liquid stock, containing 15% (v v-1) glycerol, in Luria Bertani (LB) medium. In the fermentative and bioelectrochemical systems, the culture medium described by Lovley and Philips [14] was used, containing (in mM): 30 NaHCO3, 4.2 Na2HPO4, 5.4 NaH2PO4.H2O, 0.68 CaCl2.2H2O, 1.3 KCl, 28 NH4Cl, 1.7 NaCl, 0.5 MgCl2.6H2O, 0.41 MgSO4.7H2O, 0.025 MnCl2.4H2O, 0.0041 Na2Mo4.2H2O, yeast extract 0.05 g L-1, and glycerol at 22 mM, as carbon source.
2.4. Microbial fuel cell design and operation
L-shaped dual-chamber MFC was made of glass, with an anode compartment volume of 35 mL, containing the working electrode, which consisted of a 9 cm2 carbon cloth, fixed with a platinum wire for electric connection. The 16 cm2 cathode (gas-diffusing layer-type carbon cloth, 0.3 mg cm-2 PtC 40%) was hot-pressed at 130°C and 35 kgf cm-2 for 180 s with a proton exchanger membrane (Nafion 117), creating an air-breathing cathode. The external circuit was connected to a 2200 Ω resistor and an Arduino Mega 2560 microcontroller board (C++ language) coupled to a PC for the voltage monitoring. Dissolved oxygen was removed from the anodic compartment by bubbling nitrogen.
For the kinetic assays samples were collected at regular times, to determine pH, glycerol and the by-product concentrations by Gas Chromatography (GC).
2.5. Fermentation system
The fermentative assays were carried out in triplicate in 100-mL flasks containing 50 mL of medium. The culture medium Lovley and Philips [14] was used as described, containing 22 mM of glycerol as a carbon source. To remove dissolved oxygen from the culture medium and inside the flasks, nitrogen gas was bubbled for 10 min prior to autoclaving. After inoculation, flasks were incubated at 37 °C and 150 rpm.
Samples were collected at regular times, from 0 to 25 h after inoculation, to determine pH, glycerol, and the by-product concentrations by Gas Chromatography (GC).
2.6. Substrate and by-product analysis
The samples collected from the fermentative and MFC assays were centrifuged at 9,000 rpm for 2 min (Universal Centrifuge 320 – Hettich), filtered (PTFE Hydrophilic 0.22 µm – Analítica), and vortexed for 20 seconds. Glycerol the by-products of its metabolism, such as acetic acid, butyric acid, and 1,3-PDO were quantified using a SHIMADZU Gas Chromatograph with flame ionization detector (FID) and N(g) as carrier gas [16]. 250µL sample was mixed to 750 of ethanol (Ethyl alcohol, pure, 200 proof, HPLC/spectrophotometric – SIGMA-ALDRICH) in an Eppendorf flask, and vortexed for 1 min. Next, the diluted sample was centrifuged at 9,000 rpm for 4 min and filtered. Analysis was performed on a Stabilwax®-DA column with 30-m length, 0.25-mm diameter, and 0.25-µm bonded film (RESTEK). The method used a temperature of 185 °C for 3 min, followed by heating to 220 °C at 40 °C min-1, and 220 °C for 1 min. The carrier gas was N2, used at a linear flow rate of 2.5 mL min-1. For the analysis, 2 µL of sample was injected into the equipment at 300 °C, and a 30:1 split was performed. A flame ionization detector (FID) at 290 °C was also employed.
2.7. Conversion factor (YP/S)
YP/S corresponds to the conversion factor of the substrate into the product in a given fermentation time interval and is calculated as [17],
where P0 and P are the concentrations of the product (M) at times t = 0 and t of the fermentation, respectively. S0 and S are the substrate concentrations (M) at times t = 0 and t of fermentation, respectively.
2.8. Electrochemical measurements
Cyclic voltammetry was performed between -0.6 and 0.6V against an Ag/AgCl reference electrode, at a scan rate of 2 mV s−1, and data was recorded using AUTOLAB PGSTAT 30 Potentiostat controlled with NOVA 1.11 software.
2.9. Coulombic Efficiency (CE)
The actual Coulombs transferred per mol of substrate oxidized is determined by integrating the current over time, so that the Coulombic efficiency for an MFC, evaluated over a period of time tb, is calculated as [18],
where Cp (C) is the total Coulombs calculated by integrating the current over time. CTi (C) is the theoretical number of Coulombs that can be produced from glycerol,
where F is Faraday’s constant (98 485 C/mol of electrons), bi is the number of mol of electrons produced per mol of substrate (14), Si (g L-1) the substrate concentration, v (L) the liquid volume, and Mi the molecular weight of the substrate (92.09 g mol-1).
3. Results and discussion
3.1. Electrochemical activity of the isolate
Initially, the oxidation of glycerol on the anodic chamber of the MFC by the P. aeruginosa EL14 strain was tested with different external resistances: 500, 1000, 2200, and 3200Ω. Most investigations employ external resistor of the MFC in the range of 500 - 1000 Ω (Table 1) [19,20,21]. Here, only the MFC under 2200 and 3200Ω produced detectable current. No current was recorded in the abiotic MFC, confirming the electrogenic capacity of the isolate. The current was monitored over 14 days under 2200 Ω, (Figure 1) and a lag-phase of 6 h generally could be detected. During its growth, Pseudomonas spp. produce molecules with redox activity associated with mediated electron transfer, such as pyoverdine (PYOv) and pyocyanin (PYOc). The first cycle of the MFC reached a maximum current at 82.4 ± 5.7 mA m-2 and maintained it for 16.3 h, followed by a gradual current decay. However, the subsequent feeding glycerol cycles produced lower current outputs compared with the first one, 37.6 ± 7.8, and 17.7 ± 4.2 mA m-2 in cycles 2 and 3, respectively. The decrease in the current after the first feeding cycle has also been observed with other Pseudomonas species [21]. This effect can be attributed to biofilm ripeness followed by its detachment [22].
Several factors influence the performance of MFCs, including the electron transfer mechanism of the biocatalysts, substrate type and concentration, and reactor design [23]. The literature reports that MFCs containing P. aeruginosa cultures fed with glycerol reached currents ranging from 0.63 to 418.3 mA m-2 (Table 1). For example, Dantas et al., (2013) reported currents above 300 mA m-2, but the glycerol load was 271.5 mM, much higher than that used in our work (22 mM). In addition, in the aforementioned work, the anode and cathode products were continuously recirculated. However, our results are in agreement with MFC operating under similar conditions. Zani et al., (2023) reached 91.3 mA m-2 by operating a P. aeruginosa MFC, fed with glycerol at 11 mM. Similarly, Gomes et al. (2011) employed P. aeruginosa ATCC27583 and recorded 110 mA m-2 (Table 1).
The Coulombic Efficiency (CE), furnishes a measurement of the efficiency of charge transfer from the substrate to the anode. CE around 4.5% is similar to other P. aeruginosa strains investigated in our group (Table 1). Nevertheless, improvement in the MFC design and cathode efficiency might enhance the CE.
In order to investigate the electrochemical activity and the possible EET mechanism, cyclic voltammetry of the anodic compartment was performed under two conditions: A) An abiotic anode in the presence of planktonic cells (Figure 2, green line); and B) The bioanode of the MFC was transferred to a fresh and sterile medium (Figure 2, blue line). In the first case, an oxidation and reduction peaks were detected at 0.22 and - 0.45 V (vs. Ag/AgCl), respectively. Thus, planktonic cells and their mediators act transfering electrons to the electrode and EET occurs through the mediated electron transfer (MET). P. aeruginosa is known to produce phenazines that act as redox mediators in bioelectrochemical systems. Within this class of molecules, pyocyanin (PYO), and phenazine-1-carboxamide (PCA) are the well-known phenazine derivatives. Furthermore, pyoverdine (PYOv), a yellowish-green siderophore, which consists of a peptide chain (6-12 amino acids) and a chromophore group, can also exhibit redox properties [24]. PYOv oxidation occurs at around 0.2 to 0.3V vs. Ag/AgCl, at pH 7 [25] which is very close to observed in Figure 2 (green line). Thus , suggesting that PYOv acted as a redox mediator on EET of El14. On the other hand, the oxidation peak observed for the biofilm is shifted towards lower potentials (0.05 V vs Ag/AgCl) (Figure 2, blue line). This signal should correspond to the redox species located at the cell membrane promoting a direct electron transfer (DET) via redox-active proteins, such as c-type cytochromes associated, in the outer cellular membrane [26], or even in the extracellular matrix [27].
3.2. Glycerol oxidation and by-products formation during MFC and fermentative system operation
The P. aeruginosa isolate (EL14) was tested as the biocatalyst in a bioelectrochemical system (MFC) and also in a traditional fermentation system. We used glycerol at 22 mM as substrate, and the products formed were monitored over time in both systems (Figure 3). EL14 produced acetic acid, butyric acid, and surprisingly, 1,3-PDO, in both MFC and fermentation systems.
In the MFC, the by-products were monitored during the first operating cycle with an external resistance of 2200 Ω (Figure 3A). Glycerol was completely consumed in 7 hours of operation with concomitant generation of 1,3-PDO, acetic acid, and butyric acid at 19.5 mM, 7.15 mM, and 1.60 mM, respectively. After that, the by-products were consumed over time, accompanied by an increase in electric current. When organic acids and 1,3-PDO were depleted, electricity could no longer be detected. Thus, the by-products of glycerol metabolism are responsible for the maintenance of the current in the MFC furnishing 82.4 ± 5.7 mA m-2.
Operating at fermentative condition P. aeruginosa EL14 produces 1,3-PDO; acetic acid; and butyric acid at concentrations of 16.1, 9.55, and 3.79 mM, respectively (Figure 3B). However, contrary to the MFC system, the concentration of 1,3-PDO and acetic acid remained constant, after 20 h of fermentation. This was not surprising, because, in the absence of an electron acceptor (electrode), cells cannot perform EET to gain energy. Therefore, the oxidation of its by-products occurs to a minor extent, compared with the MFC. In the fermentative system, only butyric acid is consumed (Figure 3B).
Although the glycerol oxidation by-products detected in the fermentative and the MFC were similar, the systems differ mainly in their yield of glycerol conversion (YP/S) (Figure 3C and 3D). Fermentative system, shows a higher conversion of glycerol to organic acids (acetic acid 0.86 mol mol-1 and butyric acid 0.33 mol mol-1) in the MFC (Figure 3C) (acid acetic acid 0.33 mol mol-1 and butyric acid 0.073 mol mol-1). On the other hand, the yield of 1,3-PDO was higher in the MFC (0.89 mol mol-1) compared with the fermentation one (0.57 mol mol-1). Thus, in the fermentative system, the highest YP/S was associated with the formation of oxidized by-products (acetic and butyric acid) in the MFC the major product was 1,3-PDO, a reduced product. Interestingly, this might be related to the polarization of the anode. When electrons from bacterial metabolism are transferred to the anode, the high external resistance (2200Ω) makes it difficult for electrons to pass through the external circuit to the cathode, which can lead to the polarization of the working electrode, giving it a partial negative charge. The polarized electrode can alter the intracellular oxidation-reduction potential (ORP) or act as an external source of electrons for the bacteria.
It is known that ORP levels affect many biological functions of cells through gene expression [28,29] and enzyme synthesis, which consequently affect the detection and transduction of signals and, finally, the metabolic profiles [30,31]. So, if the transfer of electrons from the electrode to the cells is involved, the electrode can function as an electron donor, and a surplus of electrons beyond those derived from substrate oxidation is available to the cells. This triggers an increase in the intracellular NADH content that can enhance the formation of the reduced final product [32]. In fact, the production of 1,3-PDO was higher in our MFC system than in the fermentative one. Another interesting observation is that in MFC the OD at 600 nm was higher (Figure 3E) than in the fermentation system (Figure 3F), 2.7 and 2.0, respectively. The higher cell growth in MFC also relies on an electron surplus for energy generation in MFC, compared to the fermentation alone.
Moscoviz et al (2017) [8] reported yields between 0.50 and 0.72 mol for 1,3-PDO/mol of glycerol during fermentation system. Our P. aeruginosa isolate showed promising results, as we obtained maximum yields of 0.57 and 0.89 mol mol-1 in fermentative and MFC, respectively. The high conversion of glycerol into 1,3-PDO in the MFC is probably related to the polarization of the electrode in the MFC.
Another interesting feature is that P. aeruginosa has a limited fermentative capacity to produce short-chain organic acids [33,34]. The pyruvate fermentation pathway, which is converted to acetate, lactate, and a small amount of succinate, confers the metabolic ability to survive in oxygen-limited environments [35]. However, 1,3-PDO is rarely associated with the metabolism of Pseudomonas species [36,37]. The most common bacteria naturally producing 1,3-PDO include Citrobacter freundii [38], Klebsiella pneumoniae [39], Klebsiella oxytoca [40,41], Clostridium pasteurianum [42,43], Clostridium butyricum [44], and Clostridium beijerinckii , which metabolize glycerol to 1,3-PDO through a reductive pathway. In the first step of the reductive pathway, glycerol is dehydrated as a result of the action of the enzyme glycerol dehydratase (GDHt) forming 3-hydroxy propionaldehyde (3-HPA). Next, the NADH+H+-dependent 1,3-PDO dehydrogenase (1,3-PDDH) reduces 3-HPA to 1,3-PDO, generating NAD+ again [45,46]. The genomic features of P. aeruginosa isolate EL14 suggest that 1,3-PDO production might occurs through alternative routes that do not involve the glycerol dehydratase enzyme, which will be discussed in the next section.
3.3. Identification of the genes responsible for the production of 1,3-PDO
Considering that strain EL14 is a promising 1,3-PDO microbial producer, we decided to gain further insight into its potential biochemical features by sequencing its genome. According to whole genome sequencing analysis, strain EL14 clustered with P. aeruginosa DSM 50071 with a 93% bootstrap (Figure S1). The highest genomic similarities lay with P. aeruginosa, with 95.2 % digital DNA-DNA hybridization (dDDH d4) for the EL14 strain.
Next, we wanted to comprehend the genomic context in strain EL14 and its association with 1,3-PDO production. Two key enzymes participate in 1,3-PDO metabolism via reductive pathway: (i) glycerol dehydratase (dhaB gene), which carries out the conversion of glycerol into 3-HPA, and (ii) glycerol dehydrogenase (dhaT gene), which catalyzes the 3-HPA conversion into 1,3-PDO. In this context, we identified three copies of the dhaT gene distributed in different loci of the EL14 genome. Interestingly, we could not find the dhaB gene in the EL14 genome in comparison with other P. aeruginosa annotated genomes and even when comparing it with 1,3-PDO microbial producers such as K. pneumoniae and Clostridium spp [47]. At the protein level, the identity matrix in alignments showed that all DhaT proteins have between 29.6 to 32.3 % of identity, reinforcing the idea that these proteins are highly diverse (Figure 4).
When aligned with DhaT sequences derived from other P. aeruginosa isolates that presented in their genomes the 1,3-PDO production pathway, DhaT_3 showed the highest protein percentage of identity (Figure 4). Also, all DhaT proteins from EL14 were smaller than the other 1,3-PDO and shortened at the N-terminal portion (around 28 aa). In addition, we found that 1,3-PDO dehydrogenases were conserved among strains of P. aeruginosa available on NCBI, but these were distinct from dhaT_1-3 found in EL14 strain. Gathering these features, we suggested that the new DhaT-related proteins identified in EL14 might have potential to produce 1,3-PDO in a synergistic way which needs to be explored in future efforts. Moreover, we aim to investigate alternative routes associated with the intermediary 3-HPA [47] that could lead to the production of 1,3-PDO without the involvement of the DhaB protein.
4. Conclusions
We reported an isolate of P. aeruginosa (EL14) used as a biocatalyst in fermentative and MFC systems. In both configurations, EL14 produced value-added products from glycerol: organic acids and 1,3-PDO, a product rarely associated with the metabolism of Pseudomonas species. In the bioelectrochemical system, 1,3-PDO yields was higher than that obtained in the fermentative system, this is associated with the polarization of the bioanode by the EET of the electroactive bacteria and the high external resistance applied to the MFC (2200 W). The unexpected fermentative profile of the isolate highlights a potential producer of 1.3-PDO. Also, new DhaT-related proteins identified in EL14 genome suggest their activity in glycerol and 1,3-PDO metabolism that might act in a synergistic way and in alternative routes without DhaB involvement. EL14 also proved to be a promising biocatalyst for bioelectricity generation, the maximum current obtained is in the same range as others P. aeruginosa strains reported.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1. Taxonomic affiliation of strain EL14 inferred with TYGS (FastME 2.1.6.1) (Leftor and Gascuel, 2015) from GBDP (Genome Blast Distance Phylogeny) distances calculated from strain EL14 genome. The branch lengths are scaled in terms of GBDP distance formula d5. The numbers above branches are GBDP pseudo-bootstrap support values > 60 % from 100 replications, with an average branch support of 95.5 %. The tree was rooted at the midpoint.
Author Contributions
VRS, and M-EG conceived of the project. JPN conducted the fermentative assays, MFC operation and electrochemical characterization. LBKM isolated the bacteria. MP conducted the nucleic acid extractions and provided the bioinformatics support. ARA has helped with the electrochemical results discussions. VRS, M-EG, and LBKM wrote the final. All authors have read and approved the manuscript.
Funding
The authors would like to thank the São Paulo Research Foundation (FAPESP) (Processes: 2014/50924-4, 2021/07294-6, 2021/010134-7, 2021/01748-5, 2021/01655-7, and 2022/04024-0), and the National Council for Scientific and Technological Development (CNPq) (INCT 465571/2014-0, 150712/2022-7).
Acknowledgments
Authors also thank CNPq for the Research Productivity Scholarships (Processes: 308914/2019-8, 302750/2020-7, 306601/2022-2).
Conflicts of Interest
The authors declare that no conflict of interest could be perceived as prejudicial to the impartiality of the reported research.
References
- Slate, A.J.; Whitehead, K.A.; Brownson, D.A.C.; Banks, C.E. Microbial Fuel Cells: An Overview of Current Technology. Renew and Sustain Energy Rev 2019, 101, 60–81. [CrossRef]
- Dasari, M.A.; Kiatsimkul, P.P.; Sutterlin, W.R.; Suppes, G.J. Low-Pressure Hydrogenolysis of Glycerol to Propylene Glycol. Appl Catal A Gen 2005, 281, 225–231. [CrossRef]
- Attarbachi, T.; Kingsley, M.D.; Spallina, V. New Trends on Crude Glycerol Purification: A Review. Fuel 2023, 340, 127485. [CrossRef]
- Zhu, F.; Liu, D.; Chen, Z. Recent Advances in Biological Production of 1,3-Propanediol: New Routes and Engineering Strategies. Green Chemistry 2022, 24, 1390–1403. [CrossRef]
- Przystałowska, H.; Lipiński, D.; Słomski, R. Biotechnological Conversion of Glycerol from Biofuels to 1,3-Propanediol Using Escherichia coli. Acta Biochim Pol 2015, 62, 23–34. [CrossRef]
- Forage, R.G.; Lin, E.C.C. DHA System Mediating Aerobic and Anaerobic Dissimilation of Glycerol in Klebsiella pneumoniae NCIB 418. J Bacteriol 1982, 151, 591–599. [CrossRef]
- da Silva Ruy, A.D.; de Brito Alves, R.M.; Reis Hewer, T.L.; de Aguiar Pontes, D.; Gomes Teixeira, L.S.; Magalhães Pontes, L.A. Catalysts for Glycerol Hydrogenolysis to 1,3-Propanediol: A Review of Chemical Routes and Market. Catal Today 2021, 381, 243–253. [CrossRef]
- Marone, A.; Ayala-Campos, O.R.; Trably, E.; Carmona-Martínez, A.A.; Moscoviz, R.; Latrille, E.; Steyer, J.P.; Alcaraz-Gonzalez, V.; Bernet, N. Coupling Dark Fermentation and Microbial Electrolysis to Enhance Bio-Hydrogen Production from Agro-Industrial Wastewaters and by-Products in a Bio-Refinery Framework. Int J Hydrogen Energy 2017, 42, 1609–1621. [CrossRef]
- Li, Z.; Dong, Y.; Liu, Y.; Cen, X.; Liu, D.; Chen, Z. Systems Metabolic Engineering of Corynebacterium glutamicum for High-Level Production of 1,3-Propanediol from Glucose and Xylose. Metab Eng 2022, 70, 79–88. [CrossRef]
- Zhou, S.; Lama, S.; Sankaranarayanan, M.; Park, S. Metabolic Engineering of Pseudomonas denitrificans for the 1,3-Propanediol Production from Glycerol. Bioresour Technol 2019, 292, 121933. [CrossRef]
- Halfeld, G.G.; de Almeida, E.J.R.; Reginatto, V.; de Andrade, A.R. Acclimatization of a Microbial Consortium into a Stable Biofilm to Produce Energy and 1,3-Propanediol from Glycerol in a Microbial Fuel Cell. Int J Hydrogen Energy 2022, 47, 21241–21252. [CrossRef]
- Allam, F.; Elnouby, M.; Sabry, S.A.; El-Khatib, K.M.; El-Badan, D.E. Optimization of Factors Affecting Current Generation, Biofilm Formation and Rhamnolipid Production by Electroactive Pseudomonas aeruginosa FA17. Int J Hydrogen Energy 2021, 46, 11419–11432. [CrossRef]
- Yong, X.Y.; Yan, Z.Y.; Shen, H.B.; Zhou, J.; Wu, X.Y.; Zhang, L.J.; Zheng, T.; Jiang, M.; Wei, P.; Jia, H.H.; et al. An Integrated Aerobic-Anaerobic Strategy for Performance Enhancement of Pseudomonas aeruginosa-Inoculated Microbial Fuel Cell. Bioresour Technol 2017, 241, 1191–1196. [CrossRef]
- Lovley, D.R.; Phillips, E.J.P. Novel Mode of Microbial Energy Metabolism: Organic Carbon Oxidation Coupled to Dissimilatory Reduction of Iron or Manganese. Appl Environ Microbiol 1988, 54, 1472–1480. [CrossRef]
- Meier-Kolthoff, J.P.; Göker, M. TYGS Is an Automated High-Throughput Platform for State-of-the-Art Genome-Based Taxonomy. Nature Communications 2019 10:1 2019, 10, 1–10. [CrossRef]
- Egoburo, D.E.; Diaz Peña, R.; Kolender, A.; Pettinari, M.J. Optimization and Validation of a GC–FID Method for Quantitative Determination of 1,3-Propanediol in Bacterial Culture Aqueous Supernatants Containing Glycerol. Chromatographia 2017, 80, 1121–1127. [CrossRef]
- Willibaldo Schmidell, Urgel de Almeida Lima, Walter Borzani, E.A. Biotecnologia Industrial - Vol. 2: Engenharia Bioquímica, Volume 2. 8 2001, 2, 1689–1699.
- Liu, H.; Logan, B.E. Electricity Generation Using an Air-Cathode Single Chamber Microbial Fuel Cell in the Presence and Absence of a Proton Exchange Membrane. Environ Sci Technol 2004, 38, 4040–4046. [CrossRef]
- Gomes, A.S.; La Rotta, C.E.; Nitschke, M.; González, E.R. Evaluation of Current Output in Pseudomonas aeruginosa Microbial Fuel Cells Using Glycerol as Susbtrate and Nafion 117 as Proton Exchange Membrane. ECS Trans 2011, 41, 2011–2017. [CrossRef]
- Dantas, P. V.; Peres, S.; Campos-Takaki, G.M.; La Rotta, C.E. Utilization of Raw Glycerol for Pyocyanin Production from Pseudomonas aeruginosa in Half-Microbial Fuel Cells: Evaluation of Two Electrochemical Approaches . J Electrochem Soc 2013, 160, G142–G148. [CrossRef]
- Zani, A.C.B.; Almeida, É.J.R. de; Furlan, J.P.R.; Pedrino, M.; Guazzaroni, M.E.; Stehling, E.G.; Andrade, A.R. de; Reginatto, V. Electrobiochemical Skills of Pseudomonas aeruginosa Species That Produce Pyocyanin or Pyoverdine for Glycerol Oxidation in a Microbial Fuel Cell. Chemosphere 2023, 335, 139073. [CrossRef]
- dos Passos, V.F.; Marcilio, R.; Aquino-Neto, S.; Santana, F.B.; Dias, A.C.F.; Andreote, F.D.; de Andrade, A.R.; Reginatto, V. Hydrogen and Electrical Energy Co-Generation by a Cooperative Fermentation System Comprising Clostridium and Microbial Fuel Cell Inoculated with Port Drainage Sediment. Bioresour Technol 2019, 277, 94–103. [CrossRef]
- Dwivedi, K.A.; Huang, S.J.; Wang, C.T. Integration of Various Technology-Based Approaches for Enhancing the Performance of Microbial Fuel Cell Technology: A Review. Chemosphere 2022, 287, 132248. [CrossRef]
- Bosire, E.M.; Rosenbaum, M.A. Electrochemical Potential Influences Phenazine Production, Electron Transfer and Consequently Electric Current Generation by Pseudomonas aeruginosa. Front Microbiol 2017, 8, 260405. [CrossRef]
- Gandouzi, I.; Tertis, M.; Cernat, A.; Saidane-Mosbahi, D.; Ilea, A.; Cristea, C. A Nanocomposite Based on Reduced Graphene and Gold Nanoparticles for Highly Sensitive Electrochemical Detection of Pseudomonas aeruginosa through Its Virulence Factors. Materials 2019, Vol. 12, Page 1180 2019, 12, 1180. [CrossRef]
- Jia, R.; Yang, D.; Xu, D.; Gu, T. Anaerobic Corrosion of 304 Stainless Steel Caused by the Pseudomonas aeruginosa Biofilm. Front Microbiol 2017, 8, 298487. [CrossRef]
- Lovley, D.R. Electromicrobiology. https://doi.org/10.1146/annurev-micro-092611-150104 2012, 66, 391–409. [CrossRef]
- Murray, D.B.; Haynes, K.; Tomita, M. Redox Regulation in Respiring Saccharomyces cerevisiae. Biochimica et Biophysica Acta (BBA) - General Subjects 2011, 1810, 945–958. [CrossRef]
- Vemuri, G.N.; Altman, E.; Sangurdekar, D.P.; Khodursky, A.B.; Eiteman, M.A. Overflow Metabolism in Escherichia coli during Steady-State Growth: Transcriptional Regulation and Effect of the Redox Ratio. Appl Environ Microbiol 2006, 72, 3653–3661. [CrossRef]
- Mason, J.T.; Kim, S.K.; Knaff, D.B.; Wood, M.J. Thermodynamic Basis for Redox Regulation of the Yap1 Signal Transduction Pathway. Biochemistry 2006, 45, 13409–13417. [CrossRef]
- Riondet, C.; Cachon, R.; Waché, Y.; Alcaraz, G.; Diviès, C. Extracellular Oxidoreduction Potential Modifies Carbon and Electron Flow in Escherichia coli. J Bacteriol 2000, 182, 620–626. [CrossRef]
- Vassilev, I.; Gießelmann, G.; Schwechheimer, S.K.; Wittmann, C.; Virdis, B.; Krömer, J.O. Anodic Electro-Fermentation: Anaerobic Production of L-Lysine by Recombinant Corynebacterium glutamicum. Biotechnol Bioeng 2018, 115, 1499–1508. [CrossRef]
- Williams, H.D.; Zlosnik, J.E.A.; Ryall, B. Oxygen, Cyanide and Energy Generation in the Cystic Fibrosis Pathogen Pseudomonas aeruginosa. Adv Microb Physiol 2006, 52, 1–71. [CrossRef]
- Price-Whelan, A.; Dietrich, L.E.P.; Newman, D.K. Pyocyanin Alters Redox Homeostasis and Carbon Flux through Central Metabolic Pathways in Pseudomonas aeruginosa PA14. J Bacteriol 2007, 189, 6372–6381. [CrossRef]
- Eschbach, M.; Schreiber, K.; Trunk, K.; Buer, J.; Jahn, D.; Schobert, M. Long-Term Anaerobic Survival of the Opportunistic Pathogen Pseudomonas aeruginosa via Pyruvate Fermentation. J Bacteriol 2004, 186, 4596–4604. [CrossRef]
- Poblete-Castro, I.; Wittmann, C.; Nikel, P.I. Biochemistry, Genetics and Biotechnology of Glycerol Utilization in Pseudomonas Species. Microb Biotechnol 2020, 13, 32–53. [CrossRef]
- Head, N.E.; Yu, H. Cross-Sectional Analysis of Clinical and Environmental Isolates of Pseudomonas aeruginosa: Biofilm Formation, Virulence, and Genome Diversity. Infect Immun 2004, 72, 133–144. [CrossRef]
- Malinowski, J.J. Evaluation of Liquid Extraction Potentials for Downstream Separation of 1,3-Propanediol. Biotechnology Techniques 1999, 13, 127–130. [CrossRef]
- Biebl, H.; Zeng, A.P.; Menzel, K.; Deckwer, W.D. Fermentation of Glycerol to 1,3-Propanediol and 2,3-Butanediol by Klebsiella pneumoniae. Appl Microbiol Biotechnol 1998, 50, 24–29. [CrossRef]
- Homann, T.; Tag, C.; Biebl, H.; Deckwer, W.D.; Schink, B. Fermentation of Glycerol to 1,3-Propanediol by Klebsiella and Citrobacter Strains. Appl Microbiol Biotechnol 1990, 33, 121–126. [CrossRef]
- Cheng, K.K.; Zhang, J.A.; Liu, D.H.; Sun, Y.; Liu, H.J.; Yang, M. De; Xu, J.M. Pilot-Scale Production of 1,3-Propanediol Using Klebsiella pneumoniae. Process Biochemistry 2007, 42, 740–744. [CrossRef]
- Nakas, J.P.; Schaedle, M.; Parkinson, C.M.; Coonley, C.E.; Tanenbaum, S.W.; Tertiolecta, D.; Primolecta, D.; Parva, D.; Bardawil, D.; Salina, D. System Development for Linked-Fermentation Production of Solvents from Algal Biomass. Appl Environ Microbiol 1983, 46, 1017–1023. [CrossRef]
- Biebl, H. Fermentation of Glycerol by Clostridium pasteurianum — Batch and Continuous Culture Studies. J Ind Microbiol Biotechnol 2001, 27, 18–26. [CrossRef]
- Papanikolaou, S.; Fick, M.; Aggelis, G. The Effect of Raw Glycerol Concentration on the Production of 1,3-Propanediol by Clostridium butyricum. Journal of Chemical Technology & Biotechnology 2004, 79, 1189–1196. [CrossRef]
- Ahrens, K.; Menzel, K.; Zeng, A.-P.; Deckwer, W.-D. Kinetic, Dynamic, and Pathway Studies of Glycerol Metabolism by Klebsiella pneumoniae in Anaerobic Continuous Culture: III. Enzymes and Fluxes of Glycerol Dissimilation and 1,3-Propanediol Formation. 1998. [CrossRef]
- Skraly, F.A.; Lytle, B.L.; Cameron, D.C. Construction and Characterization of a 1,3-Propanediol Operon. Appl Environ Microbiol 1998, 64, 98–105. [CrossRef]
- Liu, H.; Xu, Y.; Zheng, Z.; Liu, D. 1,3-Propanediol and Its Copolymers: Research, Development and Industrialization. Biotechnol J 2010, 5, 1137–1148. [CrossRef]
Figure 1.
Current density (black line) produced in the MFC with an external resistor of 2200Ω and glycerol concentration of 22 mM. The shadows represent the standard deviation in the replicates.
Figure 1.
Current density (black line) produced in the MFC with an external resistor of 2200Ω and glycerol concentration of 22 mM. The shadows represent the standard deviation in the replicates.
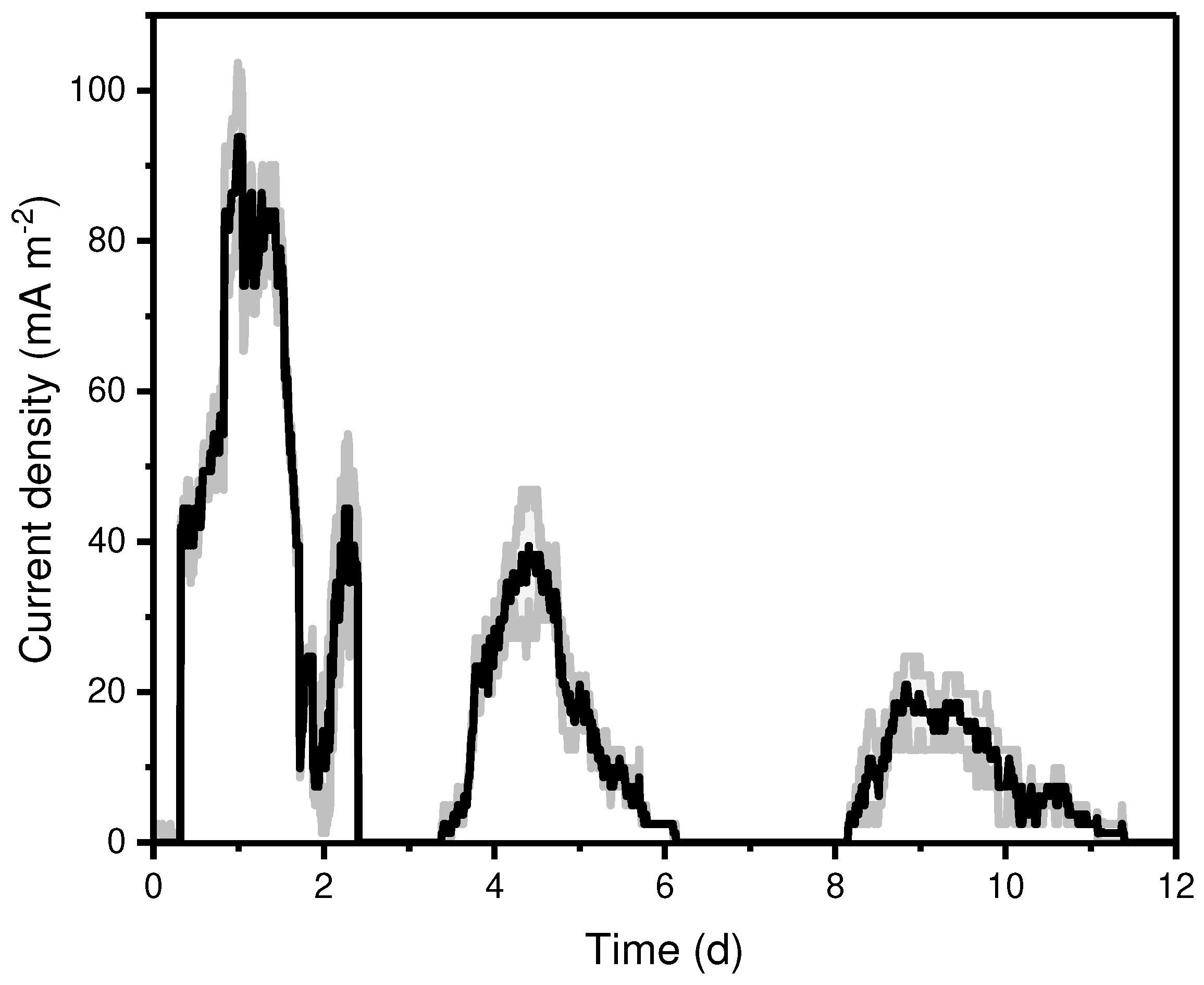
Figure 2.
Cyclic voltammograms of the anode with planktonic cells (green line) and of the anode with biofilm in fresh and sterile medium (blue line). Abiotic control is represented by black line. ν = 2 mV s-1, pH 6.9, SE = Lovely and Philips medium [14].
Figure 2.
Cyclic voltammograms of the anode with planktonic cells (green line) and of the anode with biofilm in fresh and sterile medium (blue line). Abiotic control is represented by black line. ν = 2 mV s-1, pH 6.9, SE = Lovely and Philips medium [14].
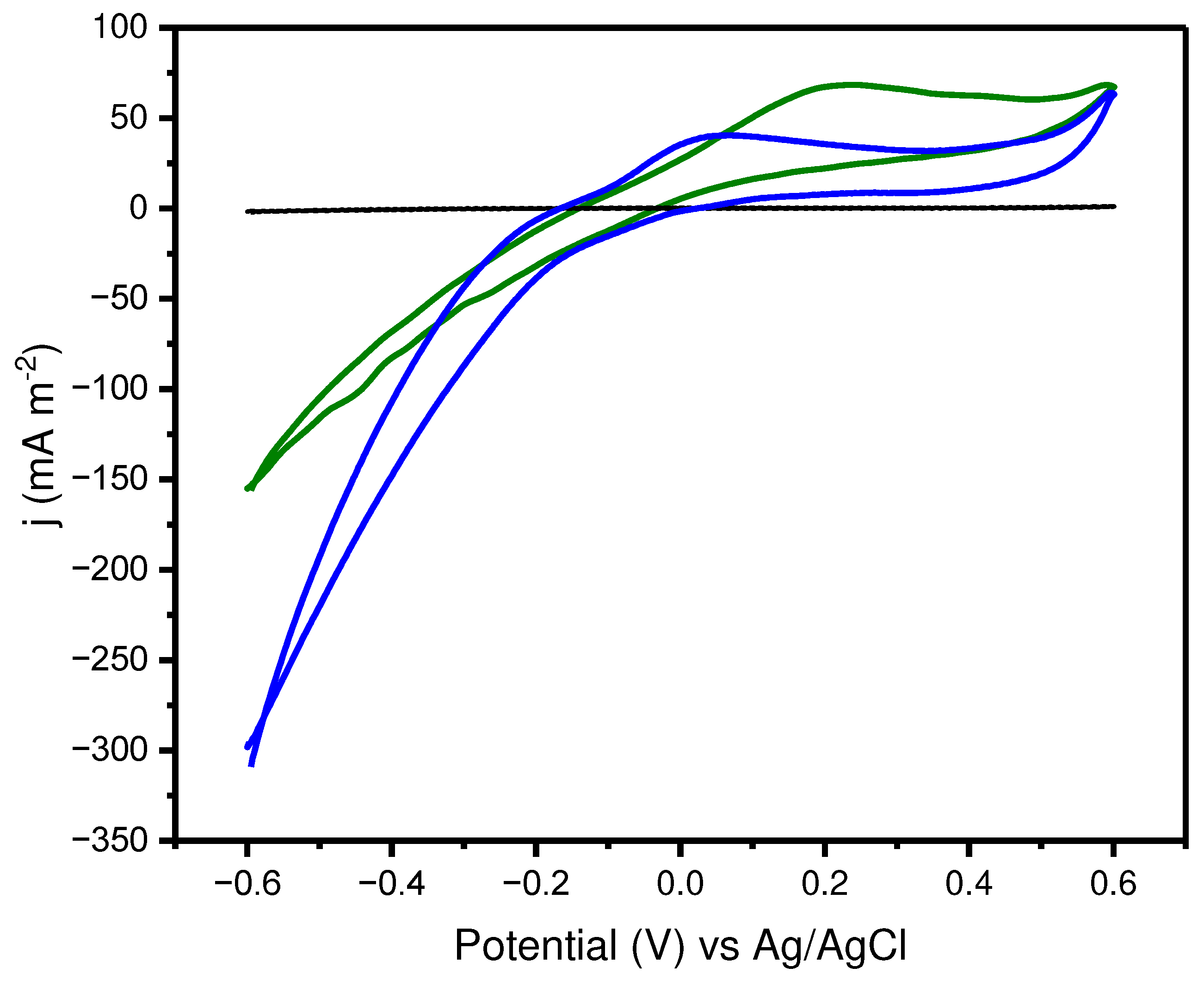
Figure 3.
Concentration of glycerol (grey line), 1,3-PDO (red line), acetic acid (blue line) and butyric acid (green line) and the current density (black line) produced by the MFC with an external 2200Ω resistor (A) and in the fermentative system over time (B). YP/S conversion factor from glycerol to 1,3-PDO (orange line), acetic acid (purple line), and butyric acid (pink line) during MFC operation (C); and in the fermentation (D); Optical density at 600 nm (gray line) and pH (red line) in MFC (E) and fermentation (F).
Figure 3.
Concentration of glycerol (grey line), 1,3-PDO (red line), acetic acid (blue line) and butyric acid (green line) and the current density (black line) produced by the MFC with an external 2200Ω resistor (A) and in the fermentative system over time (B). YP/S conversion factor from glycerol to 1,3-PDO (orange line), acetic acid (purple line), and butyric acid (pink line) during MFC operation (C); and in the fermentation (D); Optical density at 600 nm (gray line) and pH (red line) in MFC (E) and fermentation (F).
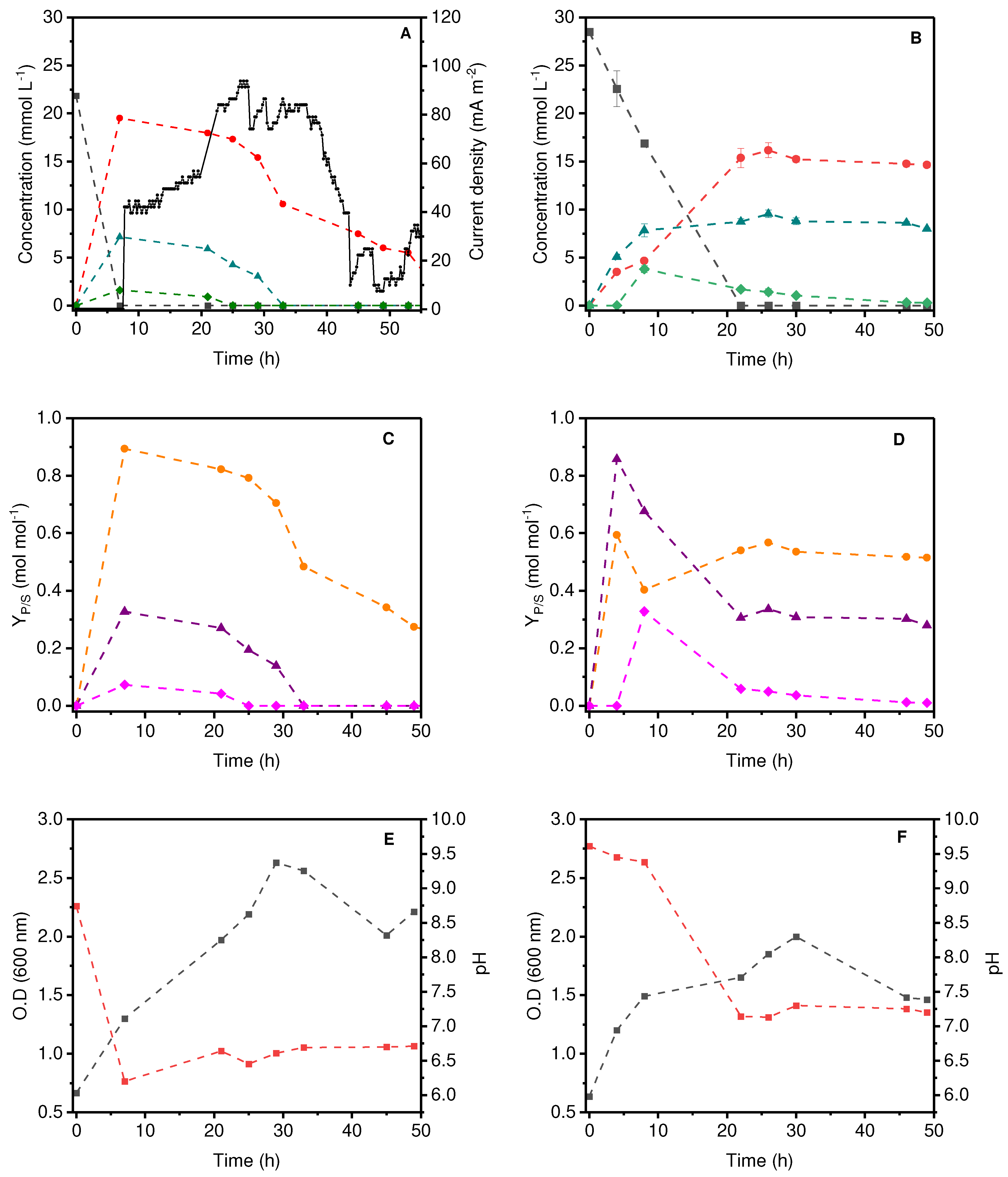
Figure 4.
Protein sequence alignment of the three 1,3-propanediol dehydrogenases variants (DhaT) identified in the strain EL14 genome, and compared with DhaT from P. aeruginosa PAO1 (brownish bar). All mismatches among the protein sequences are represented by blank spaces. The upper black bar represents the number of amino acids in the protein sequences.
Figure 4.
Protein sequence alignment of the three 1,3-propanediol dehydrogenases variants (DhaT) identified in the strain EL14 genome, and compared with DhaT from P. aeruginosa PAO1 (brownish bar). All mismatches among the protein sequences are represented by blank spaces. The upper black bar represents the number of amino acids in the protein sequences.
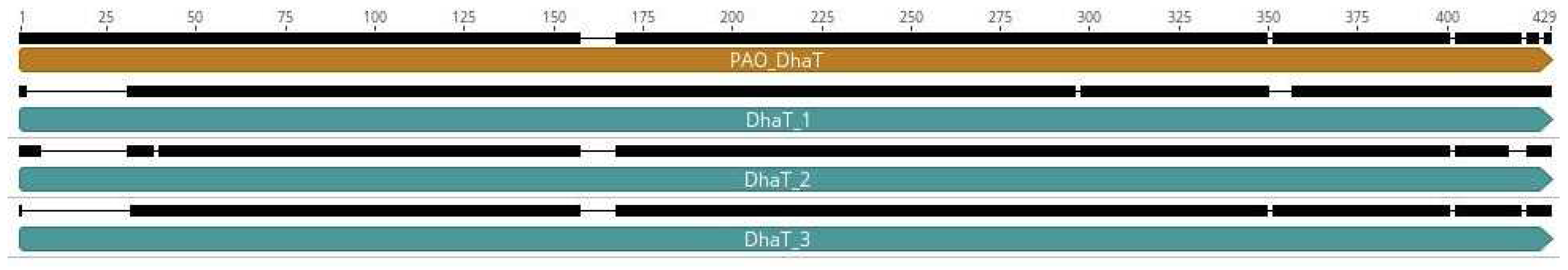

Table 1.
Maximum current density and experimental parameters for MFC fed with glycerol as substrate and P. aeruginosa as biocatalyst.
Table 1.
Maximum current density and experimental parameters for MFC fed with glycerol as substrate and P. aeruginosa as biocatalyst.
| Strain | iMAX (mA m-2) |
CE (%) |
Glycerol (mM)* |
REXT (ohms) | Anode | Salt bridge + cathode |
Reference |
|---|---|---|---|---|---|---|---|
| EW819 | 91.3 | 1.49 | 11 | 1000 | Carbon cloth | PEM + Pt-Carbon cloth (air) | [21] |
| ATCC27853 | 110 | --- | 101 | 500 | Carbon cloth | PEM + Pt-Carbon black (air) | [19] |
| EW603 | 141.2 | 5.16 | 11 | 1000 | Carbon cloth | PEM + Pt-Carbon cloth (air) | [21] |
| ATCC27853 | 153 | --- | 101 | 500 | Carbon cloth | PEM + Pt-Carbon black (air) | [19] |
| ATCC27853 | 399.0 | 46.70 | 271.5 | 1000 | Carbon felt | SB + Carbon felt (ferrocyanide) | [20] |
| ATCC27853 | 418.3 | 48.63 | 271.5 | 1000 | Carbon felt | PEM + Pt-Black on a carbon felt with carbon vulcan powder and Teflon (air) | [20] |
| EL14 | 82.4 | 4.54 | 22 | 2200 | Carbon cloth | PEM + Pt-Carbon cloth (air) | This work |
--- Not reported. * Concentrations given in g L-1 were converted to mM.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



