
Submitted:
24 June 2023
Posted:
25 June 2023
You are already at the latest version
Abstract
Background: Numerous investigations have shed light on the intriguing interplay between gut microbiota(GM) and psoriasis (Ps) as well as psoriatic arthritis (PsA). However, the precise nature of the causal relationship between them remains an area of active investigation.
Methods: For the purpose of our investigation, we meticulously curated a collection of genetic variants (P < 1 × 10−5) associated with GM (n = 18,340) derived from the MiBioGen study. To explore the intricate relationship between GM and Ps as well as PsA, we harnessed the comprehensive resources of the FinnGen database, encompassing a vast cohort of individuals, including 4,510 Ps cases and 212,242 controls, and 1,637 PsA cases and 212,242 controls. Even complementary MR methods were implemented, including an inverse variance weighting method, followed by a sensitivity analysis to verify the robustness of the results.
Results: In this study, we found that certain bacterial taxa, such as Lactococcus, Ruminiclostridium 5, and Eubacterium fissicatena, were identified as risk factors. Conversely, Alloprevotella and Odoribacter demonstrated a protective effect against Ps. In the case of PsA, our results revealed a distinct set of risk factors and protective factors among the gut bacterial taxa. Lactococcus, Verrucomicrobiales, Akkermansia, Coprococcus 1, Lachnospiraceae, and Verrucomicrobiaceae were identified as risk factors for PsA. On the other hand, Odoribacter, Rikenellaceae, Clostridium innocuum, and Marvinbryantia exhibited a protective effect against the development of PsA.
Conclusion: Our findings shed light on the distinctive disease characteristics and onset features between Ps and PsA. Notably, certain intestinal flora were implicated in the pathogenesis of PsA, suggesting their potential role as early diagnostic indicators. Furthermore, we identified several bacterial flora that exhibit a potential protective effect against the occurrence of Ps and PsA. These discoveries lay a solid foundation for future endeavors in the prevention and treatment of these conditions.
Keywords:
Gut Microbiota (GM)
; Psoriasis (Ps)
; Psoriatic Arthritis (PsA)
; Mendelian randomization(MR)
; Causal effect
Introduction
Psoriasis (Ps), a chronic inflammatory skin disorder, is characterized by aberrant proliferation of keratinocytes and infiltration of immune cells into the epidermis [1,2]. The condition affects a significant portion of the population, with varying prevalence among different ethnic groups. Psoriatic arthritis(PsA), a severe comorbidity of Ps, manifests as joint pain, swelling, and rigidity [2]. Both Ps and PsA exhibit a strong genetic predisposition, with heritability estimates ranging from 60% to 100% [1,3]. Furthermore, the prevalence of these conditions is on the rise [3].
Ps and psoriatic arthritis (PsA) have garnered significant research attention to investigate their intricate relationship and potential implications in disease pathogenesis, progression, and management [4]. Observations have revealed distinct alterations in the gut microbiome of individuals with Ps and PsA, presenting a unique dysbiosis pattern [5,6] . Several hypotheses have been put forth to elucidate the role of the gut microbiome in the pathogenesis of Ps and PsA, encompassing factors such as intestinal permeability, perturbed immune homeostasis, and imbalances in specific bacteria producing short- and medium-chain fatty acids [5,6].Notably, interventions aimed at restoring the microbiome have exhibited promise as preventive and therapeutic strategies for Ps and PsA [7]. For instance, in murine models displaying Ps-like skin inflammation, oral administration of a broad-spectrum antibiotic effectively mitigated the severity of skin inflammation by downregulating the Th17 immune response [8]. However, it is crucial to acknowledge that substantial heterogeneity exists between studies investigating alterations in gut microbial composition and Ps, necessitating unified methodological standards and large-scale investigations to comprehensively comprehend the microbiota's contribution to Ps pathogenesis and explore its potential as a therapeutic target [5,6,7].
Mendelian randomization (MR) is a statistical method used in genetic epidemiology research to investigate causal relationships between exposures and outcomes by utilizing genetic variants as instrumental variables [9]. MR analyses provide valuable insights into the potential causal relationships between the gut microbiome and various health conditions. By utilizing genetic variants as instrumental variables, researchers can explore the role of the gut microbiome in disease development and identify specific microbial taxa that may be causally linked to certain conditions. These findings contribute to a better understanding of the complex interactions between the gut microbiome and human health.
In this study, we have conducted a two-sample MR investigation using large-scale genome-wide association study (GWAS) data of gut microbiome and Ps and PsA. The objective of our study was to uncover potential causal effects of 196 gut microbiota taxa on Ps and PsA.
Materials and Methods
Data Sources and Filter Instrumental Variables
The study design framework is depicted in Figure 1, illustrating the structure of our investigation. Notably, the MiBioGen group conducted an extensive genome-wide meta-analysis of gut microbiota composition, incorporating genetic variation data pertaining to the gut microbiota [10]. This remarkable research endeavor encompassed a cohort of 18,340 individuals hailing from diverse regions such as the United States, the United Kingdom, Finland, Sweden, Denmark, the Netherlands, and other countries. The comprehensive dataset employed in this study encompassed 16SrRNA gene sequencing profiles and genotyping information. Through meticulous analysis, we identified and classified bacteria at various taxonomic levels, including 9 phyla, 16 classes, 20 orders, 35 families, and 131 genera. Subsequently, three unidentified families and 12 unknown genera were excluded from the dataset, resulting in the inclusion of 9 phyla, 16 classes, 20 orders, 31 families, and 120 genera for further analysis in the subsequent Mendelian randomization (MR) investigation (Supplementary Table S1).
To ensure the selection of high-quality genetic data, we chose the genome-wide association study (GWAS) dataset with the most comprehensive coverage of single nucleotide polymorphisms (SNPs) published in 2021, sourced from the FinnGen project [11].This specific GWAS dataset focused on the phenotypes "Ps" and "Psoriatic Arthritis" and incorporated Finnish adult subjects, consisting of 4,510 cases and 212,242 controls for Ps and 1,637 cases and 212,242 controls for PsA.
To investigate potential causal links and associations between gut microbiota and Ps and PsA, it is essential to select valid instrumental variables (IVs) that satisfy three key assumptions: (1) the correlation hypothesis, (2) the exclusivity hypothesis, and (3) the independence assumption [12] (Figure 1). We selected single nucleotide polymorphisms (SNPs) as instrumental variables (IVs) up to the genome-wide significance threshold (P < 5 × 10−8). To ensure the independence of each SNP, we applied a linkage disequilibrium (LD) factor (r2) of 0.01 and a clumping window width (kb) of 5,000 [13]. Subsequently, we extracted information on SNPs associated with the aforementioned intestinal flora from the summary genome-wide association study (GWAS) data on Ps and PsA. We eliminated missing SNPs and set the minor allele frequency (MAF) at 0.01 [14]. Additionally, we excluded all SNPs with palindromic structures to mitigate the influence of alleles on the results. To examine the presence of bias in the causal relationship between intestinal flora and Ps and PsA due to weak IVs, we employed the F value. When the F-statistic was less than 10, we considered the used SNP a weak IV and excluded it from the analysis [15]. To evaluate the potential influence of confounding factors, we utilized the PhenoScanner online tool [16] and referred to the European Dermatological Association Guidelines on Ps and PsA [17]. SNPs that showed associations with known confounders of Ps and PsA were subsequently excluded from the analysis.
Mendelian Randomization Analysis
In this study, we utilized four different approaches, namely MR-Egger, weighted median, random-effect inverse variance weighted (IVW), and weighted mode, to perform the MR analysis and calculate causal estimates between gut microbiota composition and the risk of Ps and PsA. Each approach has its specific requirements and assumptions [18]. In the MR analysis conducted in this study, a causal relationship between gut microbiota composition and the risk of Ps and PsA was considered when a significant p-value (p < 0.05) derived from any of these four methods was detected. By employing these approaches, the study aimed to assess the potential causal effects of gut microbiota composition on Ps and PsA and provide insights into their relationship.
Sensitivity Analysis
The sensitivity analysis encompassed a heterogeneity test and a multiplicity of validity test. To confirm IV heterogeneity, Cochran's Q-test was employed, and a p-value of less than 0.05 was considered indicative of the absence of heterogeneity [19]. For evaluating the magnitude of horizontal pleiotropy, MR-PRESSO aggregated the residuals for each SNP. The MR-PRESSO outlier test facilitated the identification of outlier SNPs that contributed to pleiotropy at the overall level [20]. The impact of individual outliers on the overall results was assessed using a leave-one-out analysis, calculating the remaining SNP effects after iteratively removing each SNP. Both MR-PRESSO and the leave-one-out analysis methods were employed to identify and eliminate SNPs exhibiting pleiotropy or heterogeneity [20]. Furthermore, the causal direction was investigated using the MR Steiger test [21]. Additionally, a reverse Mendelian randomization analysis was performed. The MR analyses were conducted using the R (version 4.3.0) computational environment, utilizing the “TwoSampleMR” and “MR-PRESSO” packages. The R package ' forestploter' was employed for generating certain figures. Statistical significance for causal effects was determined using a p-value threshold of less than 0.05.
Results
By applying genome-wide significance threshold screening (p < 1×10-5), LD tests, harmonization, and verification of F-statistics, multiple SNPs are identified as IVs for each of the 196 bacterial taxa. The F-statistics of all the retained SNPs demonstrate a correlation strength above 10, indicating a sufficient association between the instrumental variables and their corresponding bacterial taxa. Consequently, our study is free from weak instrument bias.
Two-Sample MR Analysis of Intestinal Flora Taxa to Ps and PsA
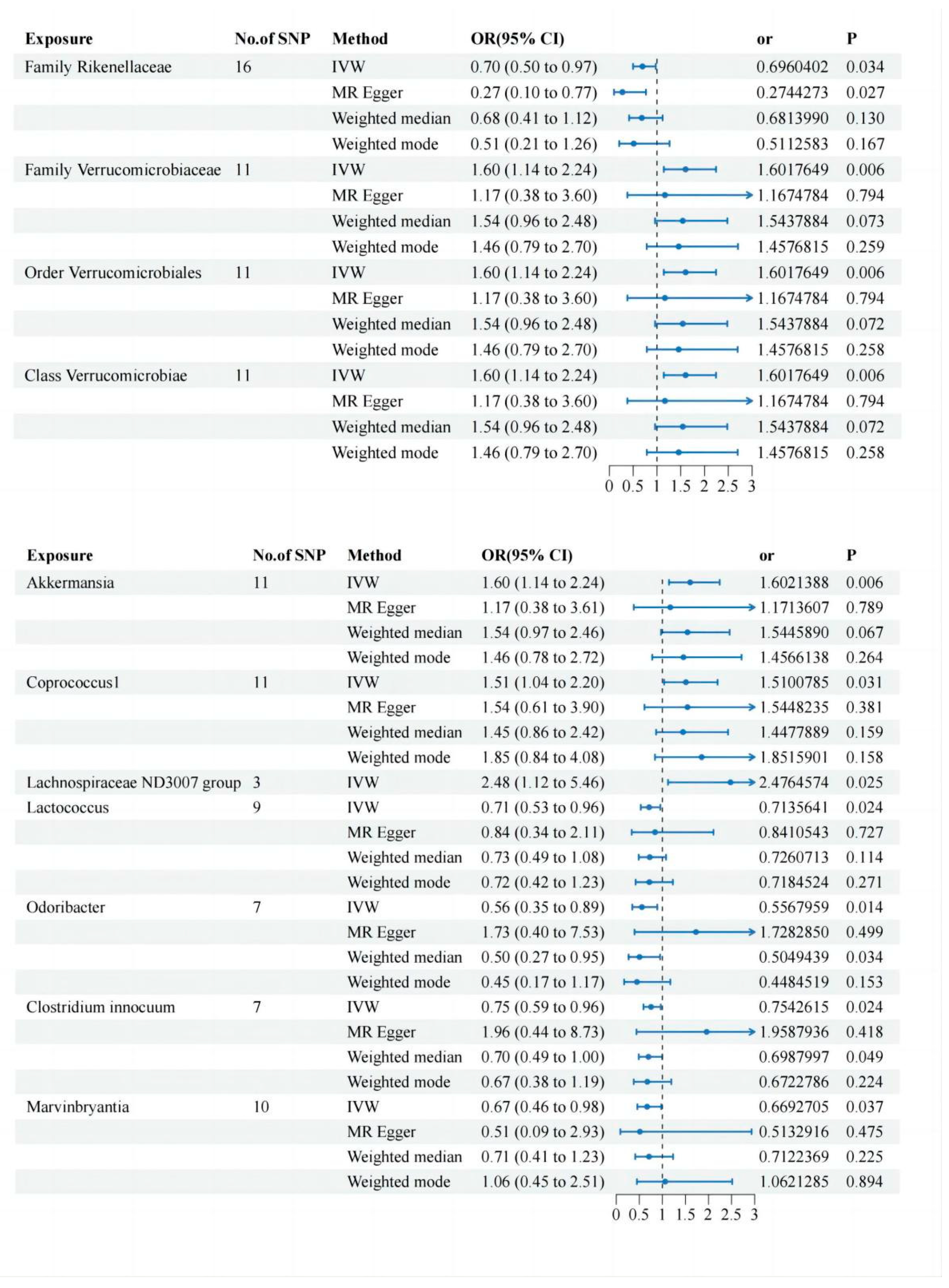
We screened 2,033 SNPs as instrumental variables from 196 gut microbiota. The results of the MR analysis for IVs are shown in a circus plot (Figure 2 and Figure 3) and detailed in Supplementary Tables S2 and S3. Our study demonstrate the presence of nine taxa of gut microbiota that accelerate the onset of Ps and PsA, while five taxa of gut microbiota have a protective effect, reducing the risk of these conditions. In Ps, the results of IVW indicated suggestive causal effects of genetically predicted increased abundance of Alloprevotella (OR, 0.84; 95% CI, 0.71–0.99; p = 0.047) and Odoribacter (OR, 0.71; 95% CI, 0.53–0.96; p = 0.024), which exhibited protective effects on Ps risk (Figure 4). Conversely, Eubacterium fissicatena at the group level (OR, 1.14; 95% CI, 0.99–1.30; p = 0.001), Ruminiclostridium5 (OR, 1.31; 95% CI, 1.01–1.70; p = 0.043), and Lactococcus (OR, 1.25; 95% CI, 1.08–1.44; p = 0.003) were associated with a higher risk of Ps. For PsA, the IVW analyses demonstrated suggestive causal effects of genetically predicted increased abundance of Rikenellaceae at the family level (OR, 0.70; 95% CI, 0.50–0.97; p = 0.034) and Clostridium innocuum (OR, 0.75; 95% CI, 0.59–0.96; p = 0.024), which exhibited protective effects on PsA risk. Conversely, Verrucomicrobiales at the order level (OR, 1.60; 95% CI, 1.14–1.24; p = 0.006), Coprococcus1 (OR, 1.51; 95% CI, 1.03–2.19; p = 0.03), Lachnospiraceae ND3007 group (OR, 2.48; 95% CI, 1.12–5.46; p = 0.025), Akkermansia (OR, 1.60; 95% CI, 1.14–1.24; p = 0.006), Verrucomicrobiae at the class level (OR, 1.60; 95% CI, 1.14–1.24; p = 0.006), and Verrucomicrobiaceae at the family level (OR, 1.60; 95% CI, 1.14–1.24; p = 0.006) were associated with a higher risk of PsA (Figure 5). It is worth noting that the order Verrucomicrobiales, along with the families Verrucomicrobiaceae and Rikenellaceae, belongs to the subcategory of the class Verrucomicrobiae. As a result, there may be significant overlap in the SNPs present in these four sets, as detailed in Supplementary Table S3. However, due to the Lachnospiraceae ND3007 group having only 3 positive SNPs, OR values obtained from the other three analysis methods are considerably higher than the upper limit of the interval, so only the IVW analysis was kept. Figure 4 and Figure 5 showed the results of the four analytical methods (MR-Egger, weighted median, IVW, and weighted mode), including P values, OR and OR(95%CI).
Sensitivity Analysis
The statistical analysis revealed that all P values indicating heterogeneity among the bacterial taxa mentioned above were greater than 0.05. Furthermore, MR Egger's test found no evidence of horizontal pleiotropy, indicating that the genetic variants used as instruments for Mendelian randomization did not have pleiotropic effects. Additionally, the reliability of the results was assessed using MRPRESSO which identified no outliers. The forest plots and the leave-one-out test revealed that no strong single SNP drives the MR estimation (Supplementary Figures S1–4, Supplementary Figures S4 and S5). This suggests that the IVs analysis based on the IVW provides reliable results for the assessment of pleiotropy.
Reverse MR Analysis
To mitigate the influence of reverse causality on the aforementioned findings, we conducted a reverse MR analysis with significant gut flora as the outcome and Ps and PsA as the exposure variables. The IVW model showed that the genus Marvinbryantia (OR = 0.96, 95% CI 0.91–1.00, p = 0.040) had a reverse causal relationship with PsA, showing PsA may lead to a decrease in the abundance of Marvinbryantia. Our analysis did not provide any additional evidence supporting a causal effect of Ps and PsA on the other 13 genera (Supplementary Tables S6 and S7).
Discussion
Ps, a chronic autoimmune disease characterized by arthritis, often leads to the development of PsA as a common complication. PsA is estimated to occur in 7% to 42% of individuals with Ps, and its prevalence gradually increases as the duration of Ps persists [22]. Previous studies have suggested a potential association between gut microbiota and the development of Ps and PsA [5]. However, direct evidence establishing a causal correlation is currently lacking. In this study, we conducted a comprehensive investigation using MR analysis to explore the relationship between 196 gut bacterial taxa and the occurrence of Ps and PsA. Regarding Ps, we found that certain bacterial taxa, such as Lactococcus, Ruminiclostridium 5, and Eubacterium fissicatena, were identified as risk factors. Conversely, Alloprevotella and Odoribacter demonstrated a protective effect against Ps. In the case of PsA, our results revealed a distinct set of risk factors and protective factors among the gut bacterial taxa. Lactococcus, Verrucomicrobiales, Akkermansia, Coprococcus 1, Lachnospiraceae, and Verrucomicrobiaceae were identified as risk factors for PsA. On the other hand, Odoribacter, Rikenellaceae, Clostridium innocuum, and Marvinbryantia exhibited a protective effect against the development of PsA.
Interestingly, among these risk factors, only one species of Lactococcus is shared between Ps and PsA, while the remaining bacteria differ. Similarly, the protective factor flora shows a striking similarity, with Odoribacter being the common protective bacterium. These findings suggest that while PsA is a complication of Ps, its pathogenesis does not completely align with that of Ps. Several of the risky bacteria identified in our study align with previous research findings. For instance, the abundance of Lactococcus and Lachnospiraceae has been shown to increase in the gut microbiota of patients with Ps [23,24]. Ruminiclostridium 5, another risky bacterium, exhibited increased abundance in an experimental model of Ps induced by imiquimod [25]. As for Odoribacter, a co-protective bacterium for both Ps and PsA, it has been reported to be more abundant in healthy individuals compared to patients [26].Furthermore, studies have demonstrated that resveratrol, a compound, can increase the abundance of Odoribacter groups, thereby restoring intestinal ecology in mice [27]. Interestingly, both oral and topical administration of resveratrol have shown potential in alleviating imiquimod-induced Ps-like dermatitis [28,29], suggesting that Odoribacter might serve as a probiotic for Ps.
Additionally, among the protective flora associated with PsA, Rikenellaceae has been found to decrease in abundance in the gut microbiota of Ps patients [30]. Another study observed a decrease in the abundance of Marvinbryantia over time in imiquimod-induced Ps-like mice [30]. Intriguingly, an observational study revealed a lower abundance of Clostridium innocuum in PsA patients (n=9) compared to non-PsA patients (n=10) [31]. These findings underscore the shared characteristics and differences between Ps and PsA. The results concerning Verrucomicrobiae, Akkermansia, and Coprococcus in our study further support this notion. Previous reports have shown a decreased abundance of these bacterial types in the intestinal flora of Ps patients [32,33,34,35], indicating their potential as protective factors for Ps. However, our findings indicate that they act as risky flora in PsA, further emphasizing the distinct disease characteristics and pathogenesis of PsA compared to Ps.
Currently, there is a prevailing belief that Ps and PsA share common pathogenic factors, including genetic risk alleles, environmental triggers, and cytokine pathways. However, it is important to note that the resident cells in the skin and joints differ significantly, and the clinical manifestations of musculoskeletal disorders and skin lesions exhibit substantial variation among individuals [33,36,37]. Despite the common involvement of tumor necrosis factor (TNF) and the IL-23-IL-17 axis in the pathogenesis of both Ps and PsA, monotherapy targeting IL-17 or IL-23 has demonstrated high efficacy in Ps but not in PsA. Although the effectiveness in PsA is less pronounced, these observations further underscore the distinct pathogenic mechanisms underlying skin and joint diseases. One potential mechanism contributing to these differences lies in the microbiome and mucosal immunity. Studies have reported significant dysregulation of intestinal mucosal immune function in PsA patients [38,39,40]. Moreover, compared to Ps, PsA patients exhibit lower overall intestinal diversity [35], suggesting that alterations in intestinal immune dynamics may contribute to synovial entheses inflammation. Notably, a specific subset of osteoclast precursors, CD14+CD16+, has been identified in PsA patients but not in Ps [41].Typically, patients with Ps experience skin lesions first, followed by the onset of PsA. However, it is worth noting that approximately 15% of cases present with arthritis and skin lesions occurring simultaneously or with arthritis preceding the skin lesions [42]. A recent study revealed that patients who develop PsA as the initial symptom often face increased delays in seeking medical attention and initiating treatment, which can significantly impact long-term prognosis. Previous research has identified potential predictors of Ps progressing to PsA. For instance, CXCL10 has been proposed as a predictive marker for the development of PsA [43]. Another case-control study demonstrated independent associations between PsA and serum levels of ITGb5, M2BP, and CRP [44]. Furthermore, evaluation of skin proteome and serum samples has revealed the presence of ITGb5 and periostin in PsA patients, distinguishing them from those with Ps alone [45].Our study suggests that specific gut flora analysis may aid in early diagnosis of PsA among patients presenting with joint inflammation. Moreover, targeting the shared pathogenic bacteria, such as Lactococcus, or considering probiotic supplementation with Odoribacter, could potentially serve as treatment options for individuals with Ps and concomitant PsA.
In this study, we conducted a comprehensive investigation to explore the causal relationship between GM and Ps and PsA, utilizing publicly available GWAS summary statistics. By employing a robust two-sample MR analysis approach, we uncovered specific bacterial groups that hold the potential to influence the development of Ps and PsA. Our findings shed light on the distinctive disease characteristics and onset features between psoriasis and psoriatic arthritis. Notably, certain intestinal flora were implicated in the pathogenesis of PsA, suggesting their potential role as early diagnostic indicators. Furthermore, we identified several bacterial flora that exhibit a potential protective effect against the occurrence of Ps and PsA. These discoveries lay a solid foundation for future endeavors in the prevention and treatment of these conditions.
One of the key strengths of our study lies in the rigorous utilization of MR method, which effectively mitigates the impact of reverse causal associations and confounding factors. This methodological approach adds considerable robustness to our findings and enhances the validity of our causal inferences. Notably, our MR study encompassed a remarkably broad population, leveraging publicly available data at a minimal cost. This extensive coverage not only enhances the generalizability of our results but also augments the practicality and persuasiveness of our findings when compared to conventional observational studies. By employing a MR approach, our study offers valuable insights into the field of gut microbiota and its implications in the context of Ps and PsA.
Naturally, it is essential to acknowledge the limitations of our research.Firstly, we extracted publicly available pooled data on exposure (gut microbiota) and outcome (Ps and PsA) for the GWAS analysis. However, it was challenging to ascertain whether there was any overlap in subjects between the MR analyses conducted on the two samples. Secondly, inconsistencies were observed in the analysis of certain bacterial populations, which could potentially be attributed to the utilization of MR Egger's method for estimating causality. It is plausible that this method introduces bias by altering the Type 1 error rate, leading to inflated rates of Type 1 errors and subsequently influencing the odds ratio (OR) [46].Thirdly, we particularly the exclusive focus on the European population, which restricts the generalizability of our findings to other ethnicities or regions. Therefore, caution should be exercised in extrapolating our results to populations beyond the scope of our study. To establish a more comprehensive understanding of the topic, it is imperative that future investigations encompass diverse populations and account for potential variations in genetic, environmental, and lifestyle factors. Moreover, we emphasize the importance of conducting further observational studies and laboratory-based investigations to validate and expand upon our current findings. By consolidating evidence from multiple research approaches, we can advance the knowledge base and provide a more robust understanding of the intricate relationship between GM and the development of Ps and PsA.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
NY and YG designed the study. JW and YL analyzed the data and drew the figures. All authors critically revised the manuscript. All authors read and approved the final manuscript.
Funding
Natural Science Foundation of Changsha (KQ2208389) and The Youth Science Foundation of Xiangya Hospital (2022Q12).
Data Availability Statement
The data that support the findings of this study are available on request from the corresponding author. The data are not publicly available due to privacy or ethical restrictions.
Acknowledgments
This study was conducted using the GWAS and FinnGen database resource. We would like to thank all participants. Table 1 was created by biorender.
Conflicts of Interest
The authors declare no conflict of interest.
Ethics Statement
Ethical review and approval were not required for the study on human participants in accordance with the local legislation and institutional requirements. Written informed consent for participation was not required for this study in accordance with the national legislation and the institutional requirements.
Disclosure Statement of Financial Interest
There are no financial conflicts of interest to disclose.
Abbreviations
GM, Gut Microbiota; Ps, Psoriasis; PsA, Psoriatic Arthritis; MR, Mendelian randomization; GWAS, Genome-wide association study; IV, Instrumental variable; SNP, Single nucleotide polymorphism; IVW, Inverse variance weighted; MR-PRESSO, MR polymorphism residual sum and outlier; ICU, Intensive care unit; FDR, False discovery rate; OR, Odds ratio; CI, Confidence interval.
References
- Ni, X.; Lai, Y. Keratinocyte: A trigger or an executor of psoriasis? J Leukoc Biol 2020, 108, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, C.; Armstrong, A.; Gudjonsson, J.; Barker, J. Psoriasis. Lancet (London, England) 2021, 397, 1301–1315. [Google Scholar] [CrossRef]
- Mou, Y.; Li, F.; Xu, Y.; Jin, X.; Dong, S.; Xia, J. Global trends in the incidence of psoriasis from 1990 to 2019. Eur J Dermatol 2022, 32, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Coates, L.; Soriano, E.; Corp, N.; Bertheussen, H.; Callis Duffin, K.; Campanholo, C.; Chau, J.; Eder, L.; Fernández-Ávila, D.; FitzGerald, O.; et al. Group for Research and Assessment of Psoriasis and Psoriatic Arthritis (GRAPPA): updated treatment recommendations for psoriatic arthritis 2021. Nature reviews. Rheumatology 2022, 18, 465–479. [Google Scholar] [CrossRef] [PubMed]
- Myers, B.; Brownstone, N.; Reddy, V.; Chan, S.; Thibodeaux, Q.; Truong, A.; Bhutani, T.; Chang, H.W.; Liao, W. The gut microbiome in psoriasis and psoriatic arthritis. Best Pract Res Clin Rheumatol 2019, 33, 101494. [Google Scholar] [CrossRef]
- Olejniczak-Staruch, I.; Ciazynska, M.; Sobolewska-Sztychny, D.; Narbutt, J.; Skibinska, M.; Lesiak, A. Alterations of the Skin and Gut Microbiome in Psoriasis and Psoriatic Arthritis. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Sikora, M.; Stec, A.; Chrabaszcz, M.; Knot, A.; Waskiel-Burnat, A.; Rakowska, A.; Olszewska, M.; Rudnicka, L. Gut Microbiome in Psoriasis: An Updated Review. Pathogens 2020, 9. [Google Scholar] [CrossRef]
- Stehlikova, Z.; Kostovcikova, K.; Kverka, M.; Rossmann, P.; Dvorak, J.; Novosadova, I.; Kostovcik, M.; Coufal, S.; Srutkova, D.; Prochazkova, P. , et al. Crucial Role of Microbiota in Experimental Psoriasis Revealed by a Gnotobiotic Mouse Model. Front Microbiol 2019, 10, 236. [Google Scholar] [CrossRef]
- Zuber, V.; Grinberg, N.; Gill, D.; Manipur, I.; Slob, E.; Patel, A.; Wallace, C.; Burgess, S. Combining evidence from Mendelian randomization and colocalization: Review and comparison of approaches. American journal of human genetics 2022, 109, 767–782. [Google Scholar] [CrossRef]
- Kurilshikov, A.; Medina-Gomez, C.; Bacigalupe, R.; Radjabzadeh, D.; Wang, J.; Demirkan, A.; Le Roy, C.I.; Raygoza Garay, J.A.; Finnicum, C.T.; Liu, X. , et al. Large-scale association analyses identify host factors influencing human gut microbiome composition. Nat Genet 2021, 53, 156–165. [Google Scholar] [CrossRef]
- Kurki, M.I.; Karjalainen, J.; Palta, P.; Sipila, T.P.; Kristiansson, K.; Donner, K.M.; Reeve, M.P.; Laivuori, H.; Aavikko, M.; Kaunisto, M.A.; et al. FinnGen provides genetic insights from a well-phenotyped isolated population. Nature 2023, 613, 508–518. [Google Scholar] [CrossRef] [PubMed]
- Zuber, V.; Grinberg, N.F.; Gill, D.; Manipur, I.; Slob, E.A.W.; Patel, A.; Wallace, C.; Burgess, S. Combining evidence from Mendelian randomization and colocalization: Review and comparison of approaches. Am J Hum Genet 2022, 109, 767–782. [Google Scholar] [CrossRef]
- Arnold, M.; Raffler, J.; Pfeufer, A.; Suhre, K.; Kastenmuller, G. SNiPA: an interactive, genetic variant-centered annotation browser. Bioinformatics 2015, 31, 1334–1336. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fu, R.; Li, R.; Zeng, J.; Liu, T.; Li, X.; Jiang, W. Causality of gut microbiome and hypertension: A bidirectional mendelian randomization study. Front Cardiovasc Med 2023, 10, 1167346. [Google Scholar] [CrossRef]
- Xiang, K.; Zhang, J.J.; Xu, Y.Y.; Zhong, X.; Ni, J.; Pan, H.F. Genetically Predicted Causality of 28 Gut Microbiome Families and Type 2 Diabetes Mellitus Risk. Front Endocrinol (Lausanne) 2022, 13, 780133. [Google Scholar] [CrossRef]
- Kamat, M.A.; Blackshaw, J.A.; Young, R.; Surendran, P.; Burgess, S.; Danesh, J.; Butterworth, A.S.; Staley, J.R. PhenoScanner V2: an expanded tool for searching human genotype-phenotype associations. Bioinformatics 2019, 35, 4851–4853. [Google Scholar] [CrossRef]
- Nast, A.; Smith, C.; Spuls, P.; Avila Valle, G.; Bata-Csörgö, Z.; Boonen, H.; De Jong, E.; Garcia-Doval, I.; Gisondi, P.; Kaur-Knudsen, D.; et al. EuroGuiDerm Guideline on the systemic treatment of Psoriasis vulgaris - Part 2: specific clinical and comorbid situations. Journal of the European Academy of Dermatology and Venereology : JEADV 2021, 35, 281–317. [Google Scholar] [CrossRef] [PubMed]
- Bowden, J.; Davey Smith, G.; Burgess, S. Mendelian randomization with invalid instruments: effect estimation and bias detection through Egger regression. International journal of epidemiology 2015, 44, 512–525. [Google Scholar] [CrossRef]
- Burgess, S.; Thompson, S. Avoiding bias from weak instruments in Mendelian randomization studies. International journal of epidemiology 2011, 40, 755–764. [Google Scholar] [CrossRef]
- Verbanck, M.; Chen, C.; Neale, B.; Do, R. Detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nature genetics 2018, 50, 693–698. [Google Scholar] [CrossRef]
- Lu, L.; Wan, B.; Sun, M. Mendelian randomization identifies age at menarche as an independent causal effect factor for gestational diabetes mellitus. Diabetes, obesity & metabolism 2023, 25, 248–260. [Google Scholar] [CrossRef]
- Ogdie, A.; Weiss, P. The Epidemiology of Psoriatic Arthritis. Rheum Dis Clin North Am 2015, 41, 545–568. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Gao, R.; Yu, N.; Zhu, Y.; Ding, Y.; Qin, H. Dysbiosis of gut microbiota was closely associated with psoriasis. Sci China Life Sci 2019, 62, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, J.; Cohen, N.A.; Shalev, V.; Uzan, A.; Koren, O.; Maharshak, N. Psoriatic patients have a distinct structural and functional fecal microbiota compared with controls. J Dermatol 2019, 46, 595–603. [Google Scholar] [CrossRef]
- Chen, X.; Chen, Y.; Stanton, C.; Ross, R.P.; Zhao, J.; Chen, W.; Yang, B. Dose-Response Efficacy and Mechanisms of Orally Administered Bifidobacterium breve CCFM683 on IMQ-Induced Psoriasis in Mice. Nutrients 2023, 15. [Google Scholar] [CrossRef]
- Xiao, S.; Zhang, G.; Jiang, C.; Liu, X.; Wang, X.; Li, Y.; Cheng, M.; Lv, H.; Xian, F.; Guo, X. , et al. Deciphering Gut Microbiota Dysbiosis and Corresponding Genetic and Metabolic Dysregulation in Psoriasis Patients Using Metagenomics Sequencing. Front Cell Infect Microbiol 2021, 11, 605825. [Google Scholar] [CrossRef]
- Cai, T.T.; Ye, X.L.; Li, R.R.; Chen, H.; Wang, Y.Y.; Yong, H.J.; Pan, M.L.; Lu, W.; Tang, Y.; Miao, H. , et al. Resveratrol Modulates the Gut Microbiota and Inflammation to Protect Against Diabetic Nephropathy in Mice. Front Pharmacol 2020, 11, 1249. [Google Scholar] [CrossRef]
- Kjaer, T.N.; Thorsen, K.; Jessen, N.; Stenderup, K.; Pedersen, S.B. Resveratrol ameliorates imiquimod-induced psoriasis-like skin inflammation in mice. PLoS One 2015, 10, e0126599. [Google Scholar] [CrossRef]
- Elgewelly, M.A.; Elmasry, S.M.; Sayed, N.S.E.; Abbas, H. Resveratrol-Loaded Vesicular Elastic Nanocarriers Gel in Imiquimod-Induced Psoriasis Treatment: In Vitro and In Vivo Evaluation. J Pharm Sci 2022, 111, 417–431. [Google Scholar] [CrossRef]
- Pinget, G.V.; Tan, J.K.; Ni, D.; Taitz, J.; Daien, C.I.; Mielle, J.; Moore, R.J.; Stanley, D.; Simpson, S.; King, N.J.C.; et al. Dysbiosis in imiquimod-induced psoriasis alters gut immunity and exacerbates colitis development. Cell Rep 2022, 40, 111191. [Google Scholar] [CrossRef]
- Lin, C.Y.; Hsu, C.Y.; He, H.R.; Chiang, W.Y.; Lin, S.H.; Huang, Y.L.; Kuo, Y.H.; Su, Y.J. Gut microbiota differences between psoriatic arthritis and other undifferentiated arthritis: A pilot study. Medicine (Baltimore) 2022, 101, e29870. [Google Scholar] [CrossRef]
- Tan, L.; Zhao, S.; Zhu, W.; Wu, L.; Li, J.; Shen, M.; Lei, L.; Chen, X.; Peng, C. The Akkermansia muciniphila is a gut microbiota signature in psoriasis. Exp Dermatol 2018, 27, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Ritchlin, C.T.; Colbert, R.A.; Gladman, D.D. Psoriatic Arthritis. N Engl J Med 2017, 376, 957–970. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Ho, H.J.; Tseng, C.H.; Lai, Z.L.; Shieh, J.J.; Wu, C.Y. Intestinal microbiota profiling and predicted metabolic dysregulation in psoriasis patients. Exp Dermatol 2018, 27, 1336–1343. [Google Scholar] [CrossRef] [PubMed]
- Scher, J.U.; Ubeda, C.; Artacho, A.; Attur, M.; Isaac, S.; Reddy, S.M.; Marmon, S.; Neimann, A.; Brusca, S.; Patel, T.; et al. Decreased bacterial diversity characterizes the altered gut microbiota in patients with psoriatic arthritis, resembling dysbiosis in inflammatory bowel disease. Arthritis Rheumatol 2015, 67, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Barnas, J.L.; Ritchlin, C.T. Etiology and Pathogenesis of Psoriatic Arthritis. Rheum Dis Clin North Am 2015, 41, 643–663. [Google Scholar] [CrossRef]
- Ocampo, D.V.; Gladman, D. Psoriatic arthritis. F1000Res 2019, 8. [Google Scholar] [CrossRef]
- Ciccia, F.; Guggino, G.; Ferrante, A.; Raimondo, S.; Bignone, R.; Rodolico, V.; Peralta, S.; Van Tok, M.; Cannizzaro, A.; Schinocca, C.; et al. Interleukin-9 Overexpression and Th9 Polarization Characterize the Inflamed Gut, the Synovial Tissue, and the Peripheral Blood of Patients With Psoriatic Arthritis. Arthritis Rheumatol 2016, 68, 1922–1931. [Google Scholar] [CrossRef]
- Mielants, H.; Veys, E.M.; Cuvelier, C.; De Vos, M.; Goemaere, S.; De Clercq, L.; Schatteman, L.; Elewaut, D. The evolution of spondyloarthropathies in relation to gut histology. II. Histological aspects. J Rheumatol 1995, 22, 2273–2278. [Google Scholar]
- Scarpa, R.; Manguso, F.; D'Arienzo, A.; D'Armiento, F.P.; Astarita, C.; Mazzacca, G.; Ayala, F. Microscopic inflammatory changes in colon of patients with both active psoriasis and psoriatic arthritis without bowel symptoms. J Rheumatol 2000, 27, 1241–1246. [Google Scholar]
- Ritchlin, C.T.; Haas-Smith, S.A.; Li, P.; Hicks, D.G.; Schwarz, E.M. Mechanisms of TNF-alpha- and RANKL-mediated osteoclastogenesis and bone resorption in psoriatic arthritis. J Clin Invest 2003, 111, 821–831. [Google Scholar] [CrossRef] [PubMed]
- Luelmo, J.; Gratacos, J.; Moreno Martinez-Losa, M.; Ribera, M.; Romani, J.; Calvet, J.; Leal, L.; Larrosa, M. A report of 4 years of experience of a multidisciplinary unit of psoriasis and psoriatic arthritis. Reumatol Clin 2014, 10, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Abji, F.; Pollock, R.A.; Liang, K.; Chandran, V.; Gladman, D.D. Brief Report: CXCL10 Is a Possible Biomarker for the Development of Psoriatic Arthritis Among Patients With Psoriasis. Arthritis Rheumatol 2016, 68, 2911–2916. [Google Scholar] [CrossRef]
- Cretu, D.; Gao, L.; Liang, K.; Soosaipillai, A.; Diamandis, E.P.; Chandran, V. Differentiating Psoriatic Arthritis From Psoriasis Without Psoriatic Arthritis Using Novel Serum Biomarkers. Arthritis Care Res (Hoboken) 2018, 70, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Cretu, D.; Liang, K.; Saraon, P.; Batruch, I.; Diamandis, E.P.; Chandran, V. Quantitative tandem mass-spectrometry of skin tissue reveals putative psoriatic arthritis biomarkers. Clin Proteomics 2015, 12, 1. [Google Scholar] [CrossRef] [PubMed]
- Burgess, S.; Thompson, S.G. Interpreting findings from Mendelian randomization using the MR-Egger method. Eur J Epidemiol 2017, 32, 377–389. [Google Scholar] [CrossRef]
Figure 1.
Mendelian randomization concept and assumptions. Schematic illustration depicting the causal relationship between gut microbiota and Ps and PsA through MR analyses. The illustration demonstrated the presence of nine taxa of gut microbiota that accelerate the onset of Ps and PsA, while five taxa of gut microbiota have a protective effect, reducing the risk of these conditions. Abbreviations: Ps, Psoriasis; PsA, Psoriatic Arthritis, MR, Mendelian randomization; SNPs, single nucleotide polymorphisms.
Figure 1.
Mendelian randomization concept and assumptions. Schematic illustration depicting the causal relationship between gut microbiota and Ps and PsA through MR analyses. The illustration demonstrated the presence of nine taxa of gut microbiota that accelerate the onset of Ps and PsA, while five taxa of gut microbiota have a protective effect, reducing the risk of these conditions. Abbreviations: Ps, Psoriasis; PsA, Psoriatic Arthritis, MR, Mendelian randomization; SNPs, single nucleotide polymorphisms.
Figure 2.
The circus plot showing four method results of all gut microbiota. The circular representation depicted the estimates obtained through IVW, weighted media, and MR-Egger methods, moving from the outer to the inner circle, respectively. The classification of gut microbiota was based on order, phylum, class, family, and genus. The varying shades of color in the circle represented the magnitude of the p-values, with the corresponding label inside the circle. Abbreviations: Ps, Psoriasis; MR, Mendelian randomization; IVW, inverse variance-weighted; WM, weighted median). Statistical significance: p < 0.05.
Figure 2.
The circus plot showing four method results of all gut microbiota. The circular representation depicted the estimates obtained through IVW, weighted media, and MR-Egger methods, moving from the outer to the inner circle, respectively. The classification of gut microbiota was based on order, phylum, class, family, and genus. The varying shades of color in the circle represented the magnitude of the p-values, with the corresponding label inside the circle. Abbreviations: Ps, Psoriasis; MR, Mendelian randomization; IVW, inverse variance-weighted; WM, weighted median). Statistical significance: p < 0.05.
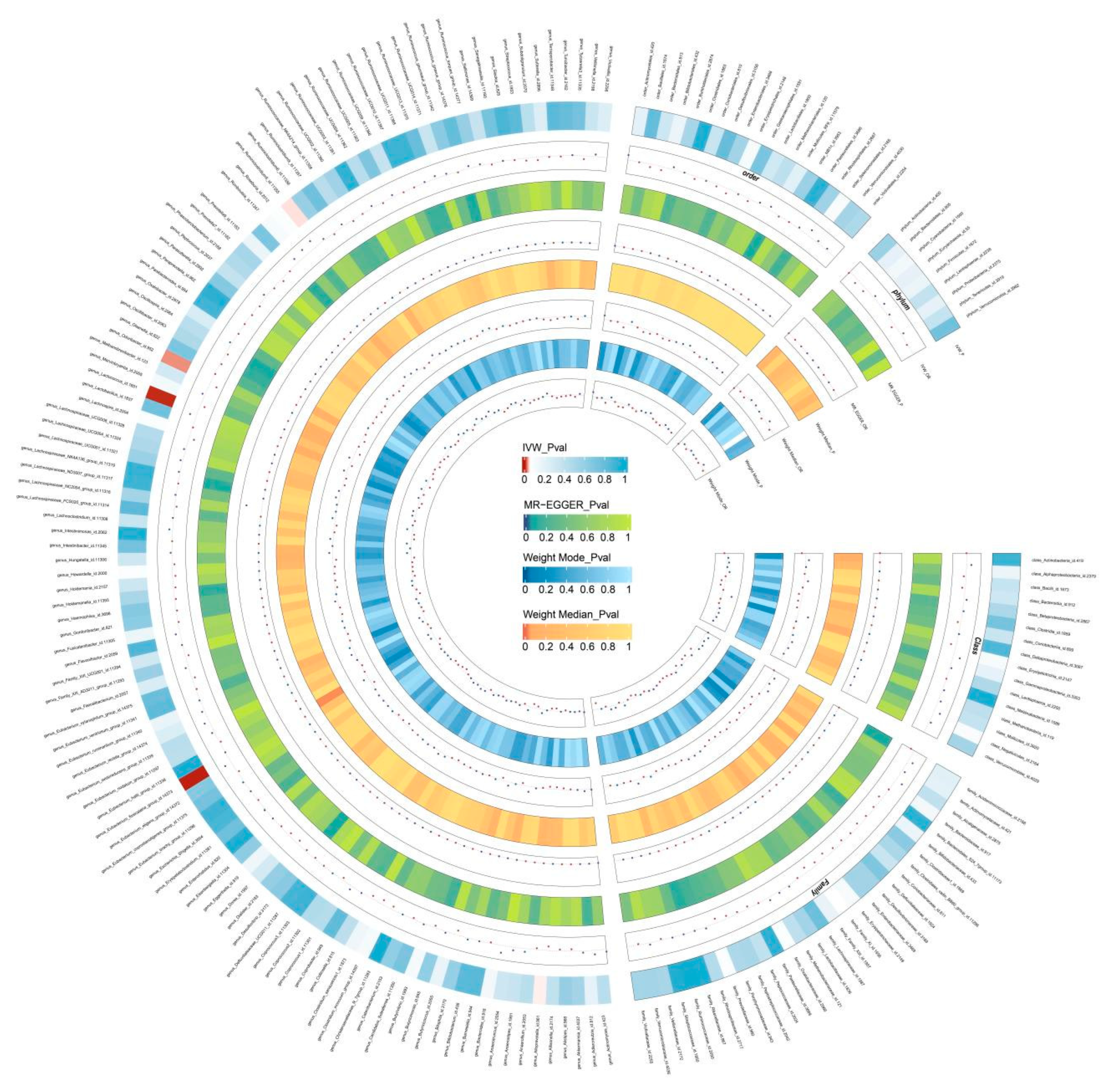
Figure 3.
The circus plot showing four method results of all gut microbiota. The circular representation depicted the estimates obtained through IVW, weighted media, and MR-Egger methods, moving from the outer to the inner circle, respectively. The classification of gut microbiota was based on order, phylum, class, family, and genus. The varying shades of color in the circle represented the magnitude of the p-values, with the corresponding label inside the circle. Abbreviations: PsA, Psoriatic Arthritis; MR, Mendelian randomization; IVW, inverse variance-weighted; WM, weighted median; Statistical significance: p < 0.05.
Figure 3.
The circus plot showing four method results of all gut microbiota. The circular representation depicted the estimates obtained through IVW, weighted media, and MR-Egger methods, moving from the outer to the inner circle, respectively. The classification of gut microbiota was based on order, phylum, class, family, and genus. The varying shades of color in the circle represented the magnitude of the p-values, with the corresponding label inside the circle. Abbreviations: PsA, Psoriatic Arthritis; MR, Mendelian randomization; IVW, inverse variance-weighted; WM, weighted median; Statistical significance: p < 0.05.
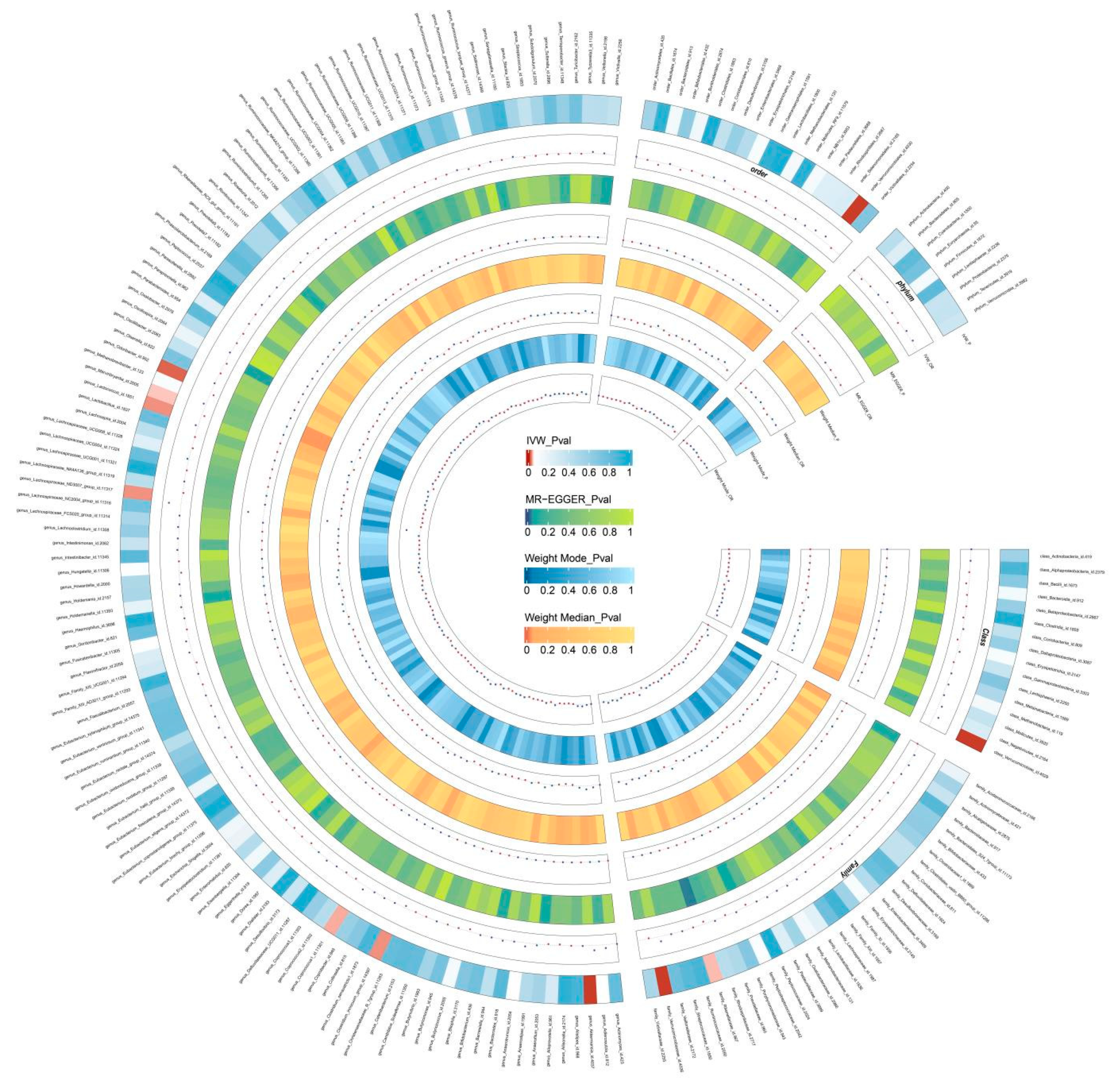
Figure 4.
Forest plots for the associations of genetic susceptibility to GM with different Mendelian randomizations of Ps. Abbreviations: Ps, Psoriasis; GM, Gut Microbiota; OR, odds ratio; CI, confidence interval; Statistical significance: p < 0.05.
Figure 4.
Forest plots for the associations of genetic susceptibility to GM with different Mendelian randomizations of Ps. Abbreviations: Ps, Psoriasis; GM, Gut Microbiota; OR, odds ratio; CI, confidence interval; Statistical significance: p < 0.05.
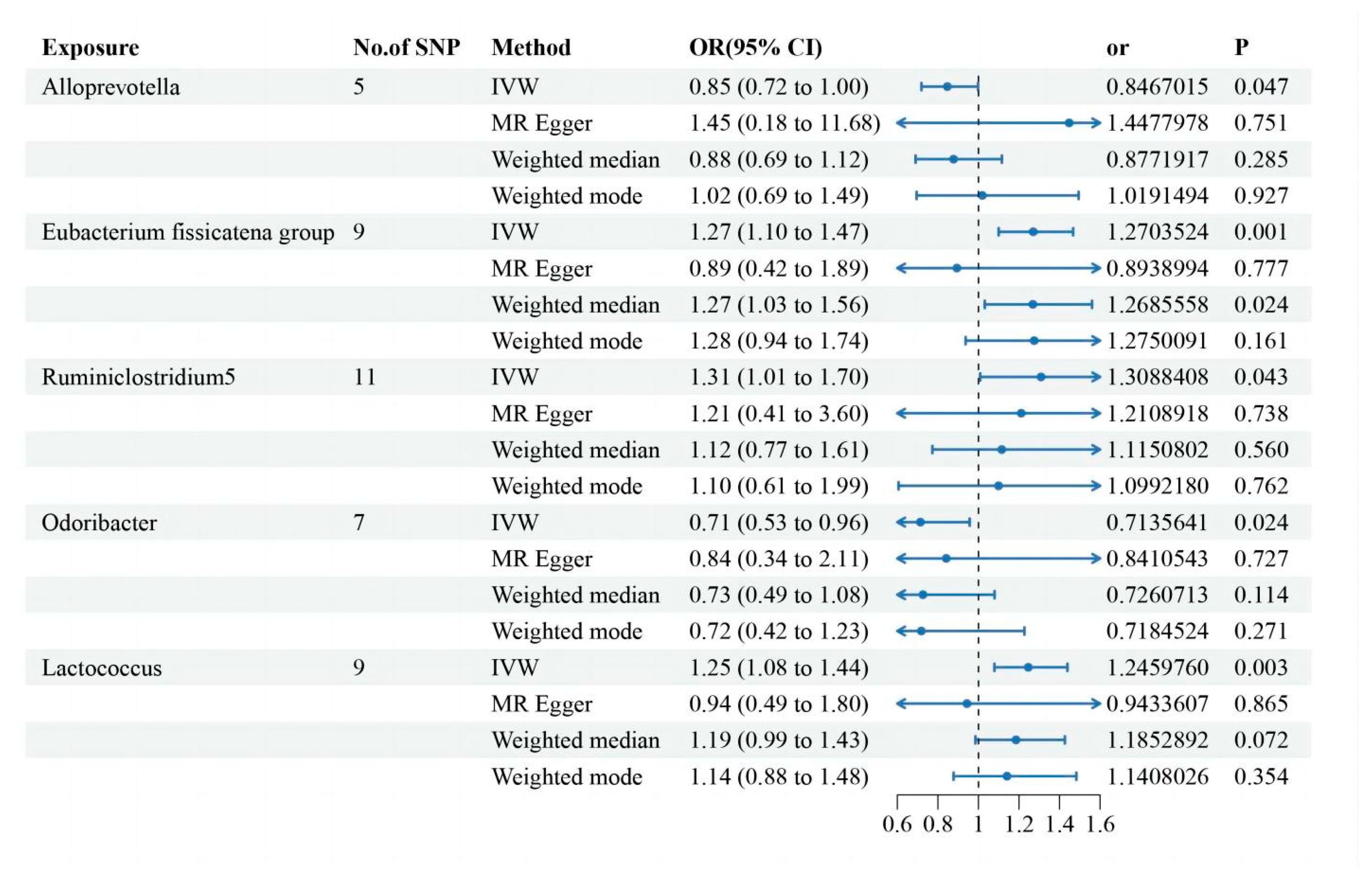
Figure 5.
Forest plots for the associations of genetic susceptibility to GM with different Mendelian randomizations of PsA. Abbreviations: PsA, Psoriatic Arthritis, GM, Gut Microbiota, OR, odds ratio; CI, confidence interval; Statistical significance: p < 0.05.
Figure 5.
Forest plots for the associations of genetic susceptibility to GM with different Mendelian randomizations of PsA. Abbreviations: PsA, Psoriatic Arthritis, GM, Gut Microbiota, OR, odds ratio; CI, confidence interval; Statistical significance: p < 0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



