Submitted:
13 June 2023
Posted:
14 June 2023
You are already at the latest version
Abstract
The yeast Saccharomyces cerevisiae is widely used in food and nonfood industries. During industrial fermentations yeast are exposed to fluctuations in oxygen concentration, osmotic pressure, pH, ethanol concentration, nutrient availability and temperature. Fermentation performance depends on the ability of different yeast strains to adapt to these changes. Suboptimal growth conditions trigger responses to these external stimuli to allow cellular homeostasis to be maintained. Stress-specific signaling pathways are activated to coordinate changes in tran-scription, translation, protein function, and metabolic fluxes while a transient arrest of growth and cell cycle progression occur. cAMP-PKA, HOG-MAPK and CWI signalling pathways are signal transduction pathways turned on during stress re-sponse. Comprehension of the mechanisms involved in the responses and in the adaptation to these stresses during fermentation is key to improving this industrial process. The scope of this review is to outline the advancement of knowledge about the cAMP-PKA signalling and the crosstalk of this pathway with the CWI and HOG-MAPK cascades in response to the environmental challenges heat and hy-perosmotic stress.
Keywords:
Saccharomyces cerevisiae
; stress
; cAMP-PKA
; CWI
; HOG-MAPK
; crosstalk
1. INTRODUCTION
Microorganisms have evolved responses that allow them to survive stressful challenges in constantly fluctuating external environment. Cellular responses to external stress are rapid, highly dynamic, plastic, complex, and involve the coordinated stimulation of many different pathways for the regulation of the gene expression at different levels [1]. The readjustments allow equilibrate the effects of stress with the physiological requirements of the cell, guarantee that critical cell parameters are fine tuned to ensure cell survival.
The yeast Saccharomyces cerevisiae, a single-celled microorganism used to produce alcoholic drinks and bread, has also been widely used as genetic model system [2]. Yeast cells suffer the exposure to several types of stress as environmental conditions change, both in natural situations and during industrial processes. Both the damage caused by stress and the yeast response depend on the type and degree of stress and the developmental stage of the yeast at the time of the stimulus [3,4]. Regardless of the type of stress exerted on the cells, a general stress response is induced. Therefore, when yeast cells are exposed to a mild stress, an increased tolerance to other stresses is achieved and restoration of cellular homeostasis is facilitated [5,6]. When the buffering capacity fails to recover cellular homeostasis, cell death programs are stimulated to eliminate irreversibly damaged cells [7,8].
S. cerevisiae has evolved mechanisms to sense, respond and adapt to these environmental changes. These mechanisms include several signal transduction pathways. Yeasts are one of the pioneer organisms used to study in detail the feedback mechanisms, the structure, organization and cellular responses through several signalling pathways to different stresses. The signalling pathways, usually conformed by kinase cascades, allow a tight control of the response to a specific signal [9,10,11]. The compartmentalization of intracellular effectors, via adaptors or anchor proteins, is critical to the temporal and spatial control of signal transduction. Although several types of stress have been studied in yeast, the complete stress-activated network and the principles that control signal integration remain incomplete [12].
When S. cerevisiae grows in optimum environmental and nutrient conditions, expression of growth-related genes is high and expression genes involved in stress defense is low. One of the transduction pathaws involved in the regulation of this balance is that of cAMP-protein kinase A (PKA) [4,13,14]. Unfavourable conditions turn off this pathway and, at the same time, stress-specific signalling networks are activated and allow coordinated changes at the level of transcription, translation, post-translational modifications, and metabolic fluxes. This results in an appropriate response to each stress situation.
The well-known cAMP-PKA pathway responds to external stimuli through the modulation of the second messenger cAMP, which activates the PKA [15,16]. S. cerevisiae PKA is a tetrameric holoenzyme consisting of a regulatory subunit (Bcy1) dimer and two catalytic subunits (Tpk1, Tpk2 and Tpk3). A single gene BCY1 encode the regulatory subunit, while there are three genes, TPK1, TPK2 and TPK3, encoding the catalytic subunits [17]. When PKA is in its inactive state, the Bcy1 dimer is bound to two catalytic subunits (Tpk). In response to different stimuli, cAMP increases, and the Bcy1 dimer undergoes conformational changes that promotes the catalytic subunits release, which phosphorylate their target substrates [18,19,20]. The output is a wide variety of specific responses. The cAMP-PKA signalling pathway in S. cerevisiae has also been associated with the regulation of ageing, budding, actin repolarization, glycogen accumulation, stress resistance, sporulation, pseudohyphal differentiation, fermentative growth, stationary phase entry, and transcriptional regulation in response to different stimuli [21,22,23,24,25].
This article reviews the current state of knowledge of the cAMP-PKA pathway involvement and the crosstalk with the CWI and MAPK signallig pathways in the response to environmental challenges focusing in heat and hyperosmotic stress in S. cerevisiae.
2. ROLE OF cAMP-PKA PATHWAY IN THE CELLULAR RESPONSE TO STRESS
Under stressful growth conditions, S. cerevisiae activates both transcriptional and physiological protective mechanisms. The stressed yeast cells activate specific transcription changes; thus, the expression of specialized genes is modulated to address the particular stress condition [26,27].
Genomic expression and global phosphoproteome studies shed light on the modulation of genes and protein phosphorylation involved in carbohydrate metabolism, protein folding degradation and processing [28,29,30]. The expression patterns of these genes during the adaptation to diverse stressful environments were termed as “Environmental Stress Response” (ESR) [3,31,32]. Actively growing cells are more sensitive to stress than quiescent cells [33].
In S. cerevisiae, one of the central controls of the ESR is the cAMP-PKA signalling, which transduces the changes in environmental conditions. The cAMP-PKA pathway is repressed in response to stress, and signalling pathways are activated to coordinate the transcriptional and translational modifications as well as the changes in the metabolic flux along with cell cycle arrest. The importance of this pathway in the adaptive response to stress is evident in mutants with hyperactive cAMP-PKA pathway. These mutants show very low tolerance to stress, decreased viability in stationary phase, and no trehalose and glycogen accumulation. On the other hand, mutations that decrease the PKA result in phenotypes with high tolerance to stress, increased accumulation of glycogen and trehalose, even in actively proliferating cells [34]. Furthermore, under many conditions, cAMP levels are high, resulting in the activation of PKA and accordingly the fermentative growth is promoted. However, in stationary phase, cAMP levels are low [35]. On the other hand, under stressful con ditions, the cAMP levels decrease and, thus, the low PKA activity re sults in the inhibition of programs of genes that regulate growth and, at the same time, in the upregulation of stress responsive genes [14]
The promoters of most genes induced by the ESR contain the STRE element, the binding site for the non-redundant Msn2 and Msn4 transcription factors. Regulation by one or the other of these transcriptional factors depends on the promoter context and the type of stress. PKA regulates the nu clear localization and therefore the activity of Msn2/4. High PKA activity induces Msn2/4 phosphorylation, which maintains their cytoplasmic localization and thus suppresses their activity. On the contrary, when the PKA activity is low, Msn2 localization is predominantly nuclear and it is active [36,37,38,39]. The subcellular localization of Msn2 in yeast is dynamic occurring in bursts in response to rapid pulses of PKA activity [40,41,42]. Later results indicate that the phosphorylation of Msn2 by Tpk1 and Tpk3 isoforms leads to the inhibition of its activity, while Tpk2 seems to function as a partial activator of Msn2 [43]
2.1. Osmotic Stress and PKA
An increased extracellular osmolarity generates hyperosmotic stress. The addition of high concentrations of salts as NaCl or KCl to S. cerevisiae cell cultures generates osmotic stress and ionic stress [44]. During the response to this stress, the concentration gradient promotes ion movement into the cell and the diffusion of water out of the cell to balance the osmotic pressure across the plasma membrane. The result is the sudden reduction in cellular volume and the cell cycle arrest. The cell responds rapidly by increasing the intracellular glycerol concentration, which causes water re-entering the cell. So, the original cell volume and turgor are restored [45].
S. cerevisiae responds to osmotic stress through two main mechanisms. One of them involves the osmolyte exporter Fps1. This channel remains closed upon hyperosmotic conditions preventing glycerol from exiting the cell [46,47]. The other mechanism involves the bona fide sensors, Sln1p and Sho1p, that control the HOG-MAPK (High Osmolarity Glycerol-Mitogen Activated Protein Kinase) pathway [48,49]. The MAPK of this pathway, Hog1, acts on cytoplasmic and nuclear targets to modify cellular metabolism to increase glycerol synthesis [50,51,52]. The HOG pathway includes a three sequentially acting protein kinases named MAPK, MAPK kinase (MAPKK, MAP2K), and MAPKK kinase (MAPKKK, MAP3K) [45,52]. There are two sensors that conform two signalling branches, SHO1 and SLN, which detect osmostress independently and activate MAP3Ks. Both Sln1 and Sho1 activate Pbs2 MAP2K by phosphorylation [53,56,57]. The activated Pbs2 can phosphorylate Hog1, and phosphorylated Hog1 translocates to the nucleus [58]. Hog1 allows the adaptive responses to osmostress of yeast cells, inducing the modulation of intracellular glycerol levels, metabolism, ion transporters, and translation. In addition, Hog1 regulates gene expression of osmostress-responsive genes [52,54,59]. The severity of the stress modulates Hog1 activation, which is negatively regulated by protein phosphatases [52].
The transcriptional regulation of Hog1 target genes occurs through diverse mechanisms, involving physical interaction with transcription factors as Msn2/4, Hot1, Tup1-Ssn6 and other transcriptional regulatory proteins [60,61]. Hog1 can also bind to the coding regions of stress-responsive genes and activates by phosphorylation the transcription elongation factors, Spt4 and Spt5 [62]. Recently, it has been proposed another mechanism by which Hog1 regulates the expression of genes by modulating the activity of the 5'-3' exoribonuclease Xrn1 [63]. Finally, it has been described the association of Hog1 to promoter regions of stress-responsive genes to facilitate the recruitment of RNA Pol II and the chromatin remodeling complexes SWI/SNF or INO80, allowing gene activation or repressing respectively [64,65,66].
PKA also regulates gene expression under osmotic stress in addition to doing so through the HOG pathway. It was demonstrated that PKA activity levels affect osmotolerance and modulate the expression of osmo-responsive genes in S. cerevisiae [67]. However, Hohmann et al proposed that PKA mediates ESR not only upon osmostress but also under several other stress conditions as high ethanol levels, thermal stress, oxidative stress, or nutrient starvation. Therefore, regulation by PKA is likely not specific to osmotic changes [68]. However, other results indicate that the regulation of ESR genes depends on the modulation of Msn2/4 activity by nuclear translocation [39,42,69,70], phosphorylation and degradation [38,71,72]. At some of these regulation levels, the signalling pathways cAMP/ PKA and HOG-MAPK have important roles [39,41,73,74,75,76].
In stress conditions, several protein kinases regulate gene expression through the binding to chromatin in either promoters or coding regions and through phosphorylation of histones, transcription factors, chromatin remodeler complexes and transcription machinery. PKA [77] and Hog1 [64,78] have been described as chromatin associated kinases. Baccarini et al demonstrated the importance of PKA chromatin association in the regulation of osmo-stress responsive genes. During osmotic stress Tpk1 accumulates in the nucleus, while Tpk2 and Bcy1 maintain the nuclear-cytoplasmic localization. The authors also demonstrated that in response to osmotic stress, PKA subunits bind to different gene regions of osmo-inducible genes. Both Tpk1 and Tpk2 subunits are recruited to the coding regions, and Tpk2 is also bound to the promoters of ribosomal protein genes. Tpk1 and Tpk2 mutant versions without catalytic activity do not bind the genes analysed so far. A mutant strain containing a deletion of BCY1 gene which has a deregulated PKA activity, shows an increased Tpk1 but not Tpk2 recruitment. Furthermore, this mutant strain, shows a higher binding rate of the remodeler complexes SWI/SNF and INO80, and also, an upregulated gene expression under hyperosmotic conditions [79].
2.2. Crosstalk between cAMP-PKA and HOG MAPK pathway during osmostress
On several occasions, the same stimulus can be processed by different signaling pathways in the cells. Likewise, there are diverse stimuli that lead to interaction and cross-activation between different signalling pathways. In this way, a specific response will be successful according to stress intensity, and modularity and hierarchical organization of the signalling pathways. The signalling pathways may interact through crosstalk or cooperative processes in response to a single stimulus [80]. In S. cerevisiae there are several examples in which multiple signalling pathways function in a coordinated manner to respond to stimuli.
The specific response to a signal of the different MAPK pathways described [70,81,82], which share several components, requires both insulation mechanisms and the coordinated communication among them. For instance, high osmolarity glycerol (HOG MAPK) pathways, mating programs (pMAPK) and filamentous growth (fgMAPK) can maintain the fidelity of the responses by restricting signalling complexes to discrete subcellular compartments and by switching on mechanisms to avoid crosstalk between MAPK cascades. In fact, in hog1Δ cells subjected to high osmolarity conditions, the pMAPK pathway is activated in contrast to wild-type cells. This way, the activation of the mating pathway is inhibited by Hog1 activity [83,84]. In response to osmotic stress, Hog1 also prevents the activation of the fgMAPK pathway by inhibiting the MAPKKK Ste11 of the SHO1 branch [52,84].
Among the MAPK pathways present in S. cerevisiae, the Cell Wall Integrity (CWI) pathway is key to overcoming cell wall damage caused by stressful conditions as chemical agents affecting cell wall biogenesis [85]. Several other stressors as heat stress, ethanol, hypo- and hyperosmotic shock, oxidative stress, among others that affect secondary the cell wall structure also activate CWI signalling [85,86]. A more detailed description of this signaling cascade is developed in the following section. Another example of crosstalk occurs during polarised growth in mating and in pseudohyphal development, where the activity of fMAPK and pMAPK pathways in coordination with the CWI to allow cell wall remodelling are required [87,88,89].
The relationship between the HOG and cAMP-PKA pathways in S. cerevisiae has also been described. The negative regulation of the HOG pathway mediated by PKA is required by yeast with defects in sphingolipid synthesis [90]. In addition, our own unpublished results indicate that the HOG MAPK and cAMP-PKA pathways interact during osmotic stress [91,92].
PKA catalytic isoforms, Tpk1 and Tpk2, show different roles in the adaptive response to osmotic stress. The lack of TPK2 gene improves the defective cell growth of HOG1-deficient strains under osmotic stress. Also, there is a negative correlation between TPK2 expression and processes such as growth rate during the exponential phase, glucose consumption, and trehalose accumulation in osmotic stress condition in mutant yeast strains with a deletion in the HOG1 gene. In contrast to TPK2, TPK1 expression has a smaller effect on restoring the defective cellular response under osmotic stress in cells with an inactive HOG MAPK pathway. In addition, the phosphorylation of Hog1 induced by osmotic stress and Hog1 nuclear accumulation were unaffected by TPK1 or TPK2 deletions. Thus, the cAMP-PKA signalling is controlling the effectors that are downstream targets of the HOG-MAPK pathway [91] (Figure 1).
Pheromone stimulation initiates yeast mating, which triggers a MAPK cascade made up of Ste11, Ste7, and finally the MAPKs Fus3 and Kss1. During mating, cell cycle is arrested by high concentrations of pheromone,and polarized cell growth is induced to form cellular projections called “shmoos” morphology [93,94,95]. Yeast cells lacking HOG1 gene show a "shmoo-like" morphology in response to osmotic stress due to the crosstalk between the HOG-MAPK and pMAPK pathways [84,96]. Our findings showed that the PKA catalytic subunits Tpk2 and, to a lesser extent, Tpk1, can reduce the crosstalk between the pheromone MAPK pathway and HOG-MAPK in a hog1∆ strain [91,92] (Figure 1).
In S. cerevisiae, filamentous growth is regulated by nutrient availability and the conserved filamentous MAPK the pathway (fgMAPK) [97]. However, the cAMP-PKA pathway activation is also required for this type of growth. Invasive growth is positively regulated by the cAMP-PKA pathway in response to glucose sensing and by the fgMAPK pathway in response to nitrogen-free medium [98,99]. Deletion of TPK2, but not TPK1, prevents filamentous growth. In addition, deletion of TPK3 produces hyperfilamentous growth, indicating that Tpk3 is an inhibitor of this growth [99,100]. In hyperosmotic conditions, a hog1Δ strain exhibits invasive growth which is regulated by a crosstalk between the HOG1-MAPK and fgMAPK pathways [84].
The role of Tpk2 subunit on the crosstalk between the fgMAPK and HOG MAPK pathways was analyzed in a strain with deficiencies in the expression of FLO8 gene. This strain is prevented from pseudohyphal growth [97]. In a hog1∆ mutant and under high osmolarity, the Tpk1 isoform is a positive regulator, but the Tpk2 isoform is a negative regulator of crosstalk between the fgMAPK and HOG MAPK pathways (Figure 1).
Hog1 and PKA can be chromatin-associated kinases (Pokholok et al., 2006). It was shown that Tpks and Bcy1 bind both coding regions and promoters of the osmoresponsive genes upon stress. The recruitment of Tpk1 and Tpk2 is completely prevented in yeast mutant strains carrying catalytic inactive versions [79]. The expression of TPK2 and HOG1 has a reciprocal impact on the binding kinetics of Tpk2 and Hog1 to the chromatin in response to osmotic stress. In addition, Tpk2 and Hog1 affect the association of Snf2 (SWI/SNF complex) and Mns2 to the promoters of osmostress responsive genes [91,92].
Overall, we suggest that when the cells fail to activate the HOG MAPK pathway, they might downregulate the cAMP-PKA pathway to produce a better adaptive response to osmostress. The lack of the HOG1 gene leads to the inactivation of Tpk2, resulting in the insulation between MAPK pathways. Furthermore, this adaptive mechanism would involve changes in the dynamics of Tpk2 association to chromatin and, consequently, in the regulation of gene expression in response to osmotic stress [91,92] (Figure 1).
Two independent osmosensing mechanisms, the Sln1 and Sho1 branches, lead to the activation of specific kinases Ssk2/22 and Ste11 (MAPKKK) that converge on the common MAPKK Pbs2 which activates the Hog1 MAPK. HOG1 deletion (red cross with dashed lines) promotes a reduction in cellular upon osmotic stress (NaCl). Deletion of TPK2 restores the defective response of the hog1∆ strain. Hog1 inhibits the crosstalk with the fgMAPK pathway in response to osmotic stress by inhibiting MAPKKK Ste11 of the Sho branch. In a hog1∆ background, Tpk2 negatively regulates the crosstalk between the HOG-MAPK and fgMAPK pathways in response to osmostress. However, Tpk1 positively regulates the crosstalk between the HOG-MAPK and fgMAPK pathways in response to osmotic stress. In strains with the genetic background BY474, the transcription factor Flo8 is not expressed. In this strain, the invasive growth signalling by the cAMP-PKA pathway is impaired in the presence of glucose. The deletion of HOG1 in the BY4741 strain promotes the crosstalk with the fgMAPK pathway upon osmotic stress. Tpk1 and Tpk2 catalytic subunits of PKA induce or inhibit the crosstalk between HOG and fMAPK pathways, respectively. Black arrows represent positive regulation, T symbols represent inhibition, the red cross with dashed lines indicates HOG1 deletion, green arrows show activated pathways and grey dashed arrows represent off state.
2.3. Thermal Stress and PKA
At suboptimal temperatures, different protective mechanisms are activated in S. cerevisiae, including a transcriptional gene expression program known as the Heat Shock Response (HSR) [27,101]. During this response the expression of genes involved in protein biosynthesis pathways are downregulated and heat-shock proteins genes are upregulated [102]. The HSR is also activated by other stresses such as heavy metals exposure, oxidative and alterations in protein conformation [103]. In addition, yeast cells modify the membrane composition and their metabolism [104]. The upregulation of heat-shock genes is driven by the transcription factor Hsf1 (Heat Shock Factor) [103]. This factor is inactive under non-stress conditions but active when misfolded proteins are accumulated in the cell. All these changes induced by thermal stress ensure the maintenance of proteostasis and metabolism in the cell [105]. In S. cerevisiae, the above mentioned Msn2/4 is a second kind of transcription factors that regulate the heat-shock gene expression. The expression regulation by Msn2/4 transcription factors is much extensive than transcripts induced by heat shock, since includes genes induced by other stresses in the general Environmental Stress Response. The shifts in the transcription levels of the HSR genes are the result of transcriptional changes and also differences in mRNA stability [106].
As it was mentioned before, PKA inhibits the Msn2/4 function, but in addition, other signal transduction pathways also regulate their activity in response to different environmental conditions, through factors as Mck1, Rim15, Yak1, Snf1, and Hog1 [43]. PKA activity is dispensable in the double deletion mutant strain of MSN2 and MSN4 genes. Therefore, the targets regulated by Msn2 and Msn4 stimulate genes that inhibit growth antagonizing the PKA dependent growth [36]. There are evidences that suggest that Yak1 kinase would fulfill this role [36,74].
In response to heat shock, Msn2/4, like Hsf1, is hyperphosphorylated; however, this modification is inhibited by cAMP. Therefore, the hyperphosphorylation might not be mediated by PKA [72]. However, heat shock slightly decreases cAMP levels through Ras activator, Cdc25, destabilization. Thus, cAMP/PKA could be the nexus between stimulus and response in HSR signalling although additional phosphorylation events may also act as regulating this response [107].
The assembling of the ribonucleoprotein (mRNP) composed of mRNAs and RNA-binding proteins (RBPs) is critical in the mRNA fate. During stress conditions, some mRNPs aggregate into larger complexes assembling membraneless organelles named RNP granules. There are many different types of cytoplasmic RNP granules; Stress Granules (SG) and Processing Bodies (PB) are two examples of them. Both types of granules participate in several aspects of mRNA metabolism as storage, localization, translation and decay [108,109,110].
However, how different types of stress impact in the formation of RNP granules is an unresolved question. PBs and SGs contain several groups of proteins as well as mRNAs, and these proteins participate in the biological activities definition of the granules. Among these proteins different protein kinases and phosphatases, have also been found associated to P-bodies [111,112,113,114,115]. PKA has a key role in the regulation of PBs and SGs assembling in response to glucose deprivation and stationary phase entry [114,116]. In addition, PKA regulates the assembly of PBs and SGs and protein translation upon heat stress in S. cerevisiae. It was shown that Tpk2, Tpk3 and Tpk1 isoforms have different roles in the assembling of SGs and PBs induced by thermal stress [117]. In conditions of moderate heat stress, Tpk3 aggregates and induces the assembly of proteins implicated in translation as eIF4G, Pab1 and eIF4E. However, these Tpk3 granules are neither PB nor SG. By contrast, upon severe heat stress the assembling of PBs and SGs containing both Tpk2 and Tpk3 and the 48S translation initiation complex are induced. The deletion of TPK2 elicits a strong translational arrest and an increment in the number and size of SGs and PBs. On the contrary, the deletion of TPK3 inhibits the assembling of SGs and PBs as well as the general protein translation.
Finally, TPK1 has no effect on the SGs and PBs evoked by heat shock. The localization of Tpk2 is dependent on its kinase activity, but Tpk3 kinase activity is not necessary for its aggregation, indicating that each catalytic subunit isoform would play different roles in granule assembly in response to severe heat stress. Therefore, Tpk2 and Tpk3 have opposite roles on the general protein translation in response to heat stress showing that the same signalling pathway can generate different physiological responses [117].
2.4. Crosstalk between cAMP-PKA and CWI pathways during heat stress
As mentioned before, high temperatures induce to the activation of the HSR and the CWI pathways in yeast cells. HSR response is regulated by the action of the transcription factors Hsf1 and Msn2/4. It was demonstrated that TPK1 expression in response to heat shock depends on Msn2/4 but not on Hsf1 [118].
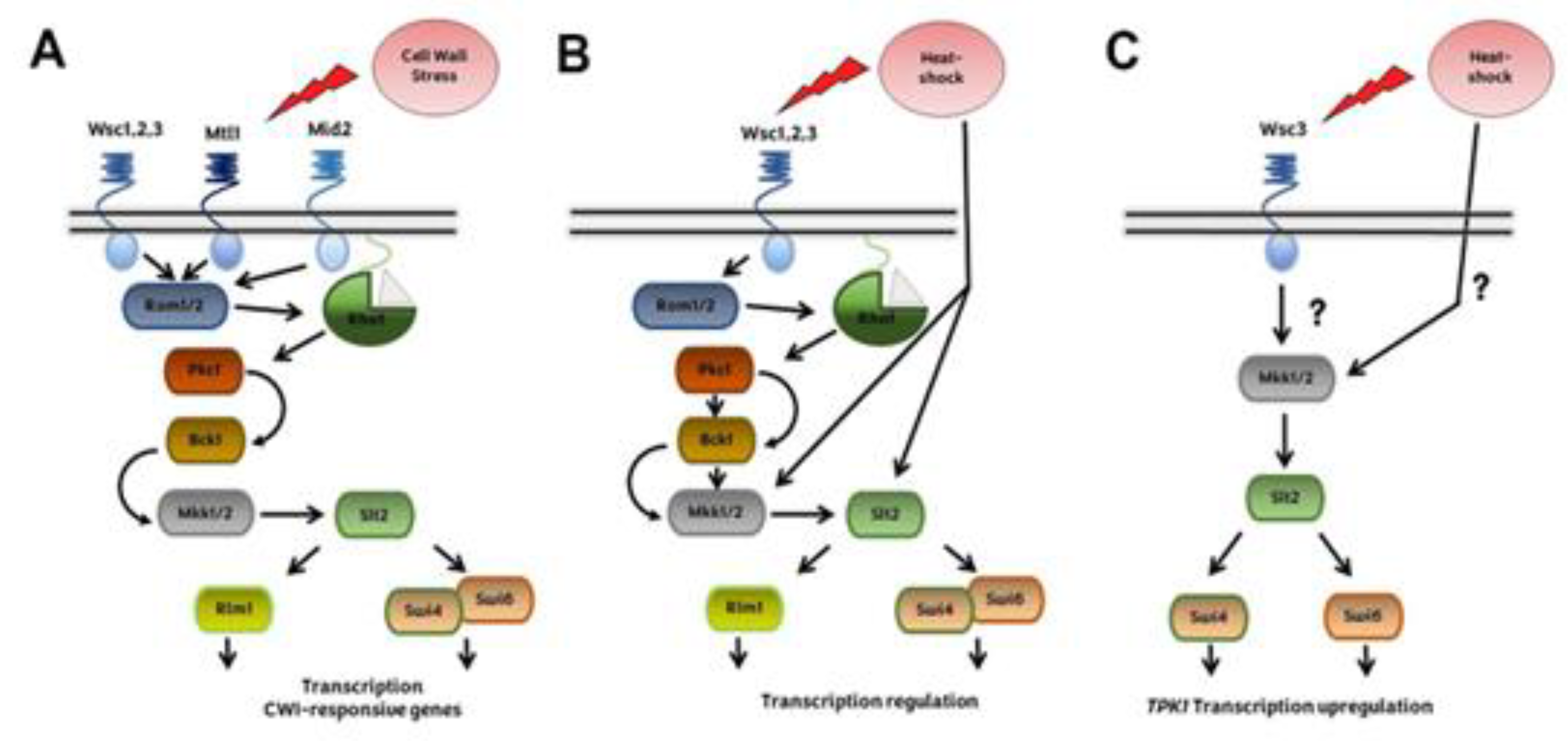
Heat-shock activates CWI pathway [119]. The first environmental condition related to the MAPK Slt2 activation was growth under thermal stress conditions [85,120]. Although the molecular mechanisms involved are not fully understood, changes in plasma membrane composition could participate to CWI activation by thermal stress [86]. There are different CWI sensors described, namely Wsc1–3, Mid2 and Mtl1 [121,122]. The participation of the CWI receptors in the sensing of this stimulus is not fully undestood. Yeast strains carrying single deletion mutations, wsc1Δ, wsc2Δ, or wsc3Δ, are thermo-tolerant, while the double mutants are thermosensitive [123]. The overexpression of Mid2 partially overcome the lack of WSC1 [124]. Therefore, the sensors have overlapping functions although they are also specific. Subsequently, it was stablished that upon heat shock the Wsc receptors have an additive effect [125]. Downstream of the membrane sensors (Wsc1-3, Mid2 and Mtl), the signal is amplified by a MAPK cascade [121,122]. Through Rom1/ Rom2 and the small G-protein Rho1, these sensors stimulate the downstream kinase Pkc1, which activates the MAPK cascade conformed by Bck1 and Mkk1/2. Finally, Mkk1/2 kinases activate the MAPK Slt2, and this kinase regulates the activity of Rlm1 and Swi4/6 transcription factors. The final result is the regulation of genes involved in cell wall biogenesis [85,102] (Figure 2).
CWI pathway is usually activated as a hierarchic top-down cascade; however, some stress stimuli can regulate this pathway at different steps of the cascade downstream Rho1. Some reports show that the activation of Slt2, the last kinase of the cascade, may come from another step in this MAPK cascade. Indeed, upon thermal stress, Slt2 is phosphorylated in a CWI sensor independent manner [126,127,128]. Thus, heat shock can activate the CWI signalling at the Mkk1/2 and/or Stl2 cascade steps [127] (Figure 2).
The crosstalk between the CWI and PKA signalling pathways was also studied. Yeast cells deficient in IRA2 are not thermotolerant; however, the deletion of Wsc1 reverses this phenotype. The authors proposed that Wsc1 negatively regulates targets of RAS. Indeed, the deletion of Ras2 rescues the heat shock sensitivity of a wsc1∆ strain. Thus, Ras and Wsc1 have opposing effects on any downstream target [123]. Later, it was demonstrated that the Wsc1 sensor also contributes to the crosstalk between CWI with the cAMP-PKA pathway at the level of Slt2. It was described that Sdp1, a phosphatase that regulates negatively Slt2, is transcriptionally regulated by the transcription factors Msn2/Msn4 [129].
CWI signalling also plays a role in the regulation of TPK1 expression during heat shock [130]. Previously, it was described that Tpk1 protein levels remain unchanged upon heat shock although TPK1 mRNA is upregulated and the half-life of TPK1 mRNA increases. This mRNA is localized in cytoplasmic foci that are not disassembled after cycloheximide treatment. The fact that these foci are resistant cycloheximide and results from the polysome profiling analysis indicate that TPK1 mRNA is impaired for entry into translation. Therefore, in response to heat shock, Tpk1 levels are regulated by a post-transcriptional mechanism that involves the assembling of TPK1 mRNA granules that are translationally silent. In this regulation the CWI components Wsc3 sensor and Mkk1 are necessary for TPK1 expression upon heat-shock. However, the participation of Stl2 is not absolutely defined. The TPK1 mRNA foci evoked upon thermal stress depends on Wsc3 but not on the other sensors. The levels of Tpk1 protein are lower in a wsc3Δ mutant than in a wild-type strain, and consequently PKA levels are also lower, as was demonstrated by phenotypes analysis. Regarding the participation of the transcription factors Swi4 and Swi6, it was published that apparently only Swi4 seems to be necessary for the regulation of TPK1 expression [130]. It has been reported very little overlapping between the gene profiles of mutant strains swi4Δ and slt2Δ upon heat shock. Genes dependent on Swi4 but independent on both Swi6 and Slt2, such as TPK1 [130], were described [127]. Therefore, the expression of Tpk1 subunit isoform in response to heat stress requires a crosstalk between CWI and cAMP-PKA signalling pathways (Figure 2).
A- Canonical CWI pathway. Damage in cell wall is sensed via wsc1-3, Mtl1 and Mid2 sensors that trigger the MAPK cascade conformed by Bck1, Mkk1/Mkk2, and Slt2 through Rho1 and the activation of Pkc1 the. B- CWI pathway and heat shock. The signalling pathway is activated by a different mechanism, regulating the kinases Mkk1,2 and Stl2 rather than Rho1 or Pkc1. Heat stress would be acting as a lateral input rather than operating in a linear “top-down” manner. C- Tpk1 expression regulation during heat shock. Tpk1 upregulation depends on the Wsc3 membrane sensor Mkk1 and the transcription factor Swi4.
3. EFFECT OF STRESS ON THE SPECIFICITY REGULATION OF cAMP-PKA PATHWAY
Different external signals trigger the production of cAMP as the only second messenger in the cAMP-PKA signalling. Considering the multiple functions of this pathway in S. cerevisiae, an important question is how this kinase achieves specificity, that is, how the cell accomplishes the accurate substrate phosphorylation in response to different stimuli. The three Tpk isoforms are functionally redundant for cell viability despite each one performs specific functions [99,100,131,132,133,134].
The specificity of PKA signalling in S. cerevisiae is regulated by several mechanisms. We will describe these mechanisms highlighting those related to thermal and osmotic stress.
3.1. PKA anchoring through Bcy1 interacting proteins
Yeast PKA localization appears to be different from that described for mammals. Bcy1 localization is variable and responsive to environmental and nutritional conditions [114,135]. Bcy1 N-terminus structure is similar to the canonical mammal RIIα domain (DD domain) as it has a helix-turn-helix motif and the critical amino acids for dimerization [136,137]. However, the binding domain of proteins described as Bcy1 interactors in S. cerevisiae displays different molecular features than the canonical domain of their mammalian counterparts, AKAPs (A-Kinase Anchoring Proteins, DD-AKAP), which contain essential hydrophobic residues [138,139,140].
The N-terminal domain of Bcy1 and two clusters of serine residues which are phosphorylated located at this domain have been reported to be critical for Bcy1 cytoplasmic localization in cells deprived of glucose [141]. Several Bcy1 interacting proteins have been described. Zds1, the first one, participates in the cytoplasmic localization of Bcy1 [141]. Other Bcy1-interacting proteins as Hsp60 (mitochondrial chaperonin), Eno2 (enolase II), and Ira2 (RAS GTPase-activating protein) were identified [142]. However, no interacting proteins were described in thermal or osmotic stress conditions.
3.2. Subcellular localization of Tpk1, Tpk2 and Tpk3 catalytic isoforms
In S. cerevisiae, the localization of each PKA subunit in different subcellular compartments and structures is affected by several environmental conditions, as glucose deprivation, thermal and osmostress or quiescent arrest [135,143]. When the yeast cells were grown in the presence of glucose, both Bcy1 and Tpk2 are localized in the nucleus; however, Tpk1 and Tpk3 subunits are equally distributed in nucleus and cytoplasm [114]. On the other hand, when yeast cells were grown in the presence of glycerol or when enter in stationary phase, both Tpks and Bcy1 subunits are localized mainly in cytoplasm [114].
As mentioned above, Tpk1 accumulates in the nucleus, whereas the localization of Tpk2 and Bcy1 does not change in response to osmostress [79]. On the other hand, upon heat stress, during glucose starvation or in quiescent cells, Tpk1 and Bcy1 display a diffuse cytoplasmic localization. However, Tpk2 and Tpk3 subunits are assembled in PBs and SGs [111,143,144]. The severity of the heat stress also regulates the localization of PKA subunits. In response to a mild heat stress, Tpk2 localization is cytoplasmic instead nuclear, and Tpk3 condensates in cytoplasmic foci which are different to classical SGs or PBs. On the other hand, both Tpk2 and Tpk3 subunits are assembled in SGs under severe heat stress [117]. When the cells are treated with cycloheximide and then subject to heat stress the foci containing Tpk2 and Tpk3 are not detected, indicating that these foci are dependent on the translation initiation repression [117]. The granular localization of Tpk2, but not that of Tpk3, depends on its catalytic activity. All these results suggest that different mechanisms are involved in the assembling of each catalytic subunit in response to severe heat stress [117]. A breakthrough in this topic is the demonstration that the N-terminus of Tpk2 subunit has a prion like domain necessary to localise this catalytic isoform to PBs and SGs upon heat stress, under glucose depletion and after quiescent arrest [145].
Therefore, there is a correlation between isoform specificity, subcellular localization and stress response. The differential subcellular localization of each catalytic isoform of PKA contribute to the specificity control of cAMP-PKA pathway as each isoform may interact with different proteins and potential substrates.
3.3. Transcriptional regulation of PKA subunits
Pioneer high-throughput transcriptomic studies indicate that, in response to heat shock and saline stress, TPK1, TPK2, TPK3 and BCY1 gene expression is upregulated [26,146,147,148,149,150].
However, later published evidence demonstrated that the expression of each PKA subunit is differentially regulated under different growth conditions such as carbon source availability or growth phase [143,151]. In addition, PKA activity regulates the transcription of the three catalytic isoforms and Bcy1 subunits that compose the holoenzyme [118].
Tpk2 catalytic subunit is the isoform with the highest inhibitory effect on the activity of TPK1 and TPK3 promoters but fails to inhibit its own promoter [118]. Of all the subunits that compose PKA, only the expression of catalytic isoform Tpk1 is modulated during heat shock and osmostress. Under these conditions, both mRNA levels and half-life increase. In response to heat shock, the upregulation of TPK1 depends on the transcription factors Msn2/4, Gis1, Sok2, and the kinase Rim15. During the TPK1 promoter activation three positioned nucleosomes are evicted [118,152]. The chromatin remodeling involves the activity of the remodelers RSC and INO80 to maintain the repression of TPK1 promoter under normal growth conditions, and the complex SWI/SNF to allow the activation after thermal stress [152]. Msn2/4 is necessary for the recruitment of the SWI/SNF complex. Strikingly, the catalytic subunits Tpk1 and Tpk2 are both recruited to the TPK1 promoter upon heat shock but with opposite temporal patterns [152]. Furthermore, Tpk1 and Tpk2 catalytic activities have opposite effects on the chromatin remodeling of this promoter [152]. Therefore, a complex regulation mechanism involves the activity of Tpk subunits on the TPK1 promoter. Finally, after thermal stress, the increased level of Tpk1 allows the conformation of PKA holoenzymes containing a higher proportion of the catalytic Tpk1 isoform. This holoenzyme might phosphorylate Tpk1 specific substrates improving the overall cellular fitness when normal environmental conditions are restored [130].
These results uncover a special mechanism involved in the regulation of the Tpk1 subunit expression by thermal stress that contributes to define the specificity of cAMP-PKA.
4. CONCLUDING REMARKS
To respond adequately to stressors, S. cerevisiae employs different signalling pathways. Each pathway is fine-tuned through mechanisms that allow the specificity of the response. The complexity of the inputs to which the yeast may be exposed suggests that several pathways should be interconnected to process environmental signals and to achieve a specific response. Two important crosstalk interactions couple the signalling cAMP-PKA and CWI pathways in response to heat shock, and HOG-MAPK and cAMP-PKA pathways upon osmotic stress.
The regulation of the expression of each PKA subunit is one of the important mechanisms that allows signal transduction specificity. In response to heat shock and saline stress, TPK1 is the only catalytic subunit of PKA upregulated, and the cAMP-PKA/CWI crosstalk coordinates Tpk1 expression.
The interaction of HOG-MAPK and cAMP-PKA pathways highlights the differential roles of the catalytic isoforms of PKA, Tpk1 and Tpk2, in the adaptive response to osmotic stress. The deletion of TPK2 gene, but not TPK1, improves the defective cell growth of HOG1 deficient strains under osmotic stress. PKA catalytic subunits Tpk2 and, to a lesser extent, Tpk1, can reduce the crosstalk between the pMAPK and the HOG-MAPK pathways in a deficient HOG1 strain. The cAMP-PKA pathway activation is required for filamentous growth and each catalytic isoform has a different role in this process. The invasive growth of a hog1Δ strain under hyperosmotic conditions is regulated by a crosstalk between the HOG1-MAPK and fgMAPK pathways. Tpk1 is a positive regulator, while Tpk2 is a negative one in this crosstalk. Finally, there is also an interaction between PKA and HOG1 at the level of transcriptional regulation of osmostress responsive genes. TPK2 and HOG1 have a reciprocal impact on the chromatin-binding kinetics of Tpk2 and Hog1. Also, both kinases regulate the binding of SWI/SNF complex and Mns2 to the promoters of osmostress-responsive genes.
In conclusion, intricated regulatory networks that include the crosstalk between different signalling pathways take place in response to stress. The complementation of signalling pathways, the fine tuning of the signals, and the specificity in the response to different stressors are key to produce a precise and timely gene expression output to overcome the stressful conditions.
Author Contributions
Conceptualization, P.P and S.R; review and editing, M.C.O-M, F.G. and M.B-M; supervision, P.P. and S.R.; project administration, P.P and S.R.; funding acquisition, P.P and S.R. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by grants from the Agencia Nacional de Promoción Científica y Tecnológica (PICT 2014-2937, PICT 2017-02240 and PICT 2018-0378), from the University of Buenos Aires (UBACyT 2016–2018, 20020150100035BA, UBACyT 2020-2023 20020190100122BA and Ministerio de Ciencia, Tecnología E Innovación, Consejo Nacional De Investigaciones Científicas Y Técnicas, PIP 2021-2023 GI).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bleuven, C.; Landry, C.R. Molecular and Cellular Bases of Adaptation to a Changing Environment in Microorganisms. Proc Biol Sci 2016, 283. [CrossRef]
- Vanderwaeren, L.; Dok, R.; Voordeckers, K.; Nuyts, S.; Verstrepen, K.J. Saccharomyces cerevisiae as a Model System for Eukaryotic Cell Biology, from Cell Cycle Control to DNA Damage Response. International Journal of Molecular Sciences 2022, Vol. 23, Page 11665 2022, 23, 11665. [CrossRef]
- Gasch, A.P. The Environmental Stress Response: A Common Yeast Response to Diverse Environmental Stresses. Yeast Stress Responses 2003, 11–70. [CrossRef]
- Chasman, D.; Ho, Y.-H.; Berry, D.B.; Nemec, C.M.; MacGilvray, M.E.; Hose, J.; Merrill, A.E.; Lee, M.V.; Will, J.L.; Coon, J.J.; et al. Pathway Connectivity and Signaling Coordination in the Yeast Stress-Activated Signaling Network. Mol Syst Biol 2014, 10, 759. [CrossRef]
- Święciło, A. Cross-Stress Resistance in Saccharomyces cerevisiae Yeast--New Insight into an Old Phenomenon. Cell Stress Chaperones 2016, 21, 187–200. [CrossRef]
- Guan, Q.; Haroon, S.; Bravo, D.G.; Will, J.L.; Gasch, A.P. Cellular Memory of Acquired Stress Resistance in Saccharomyces cerevisiae. Genetics 2012, 192, 495–505. [CrossRef]
- Falcone, C.; Mazzoni, C. External and Internal Triggers of Cell Death in Yeast. Cellular and Molecular Life Sciences 2016, 73, 2237–2250. [CrossRef]
- Grosfeld, E. V.; Bidiuk, V.A.; Mitkevich, O. V.; Ghazy, E.S.M.O.; Kushnirov, V. V.; Alexandrov, A.I. A Systematic Survey of Characteristic Features of Yeast Cell Death Triggered by External Factors. J Fungi (Basel) 2021, 7. [CrossRef]
- Li, G.; Qian, H. Sensitivity and Specificity Amplification in Signal Transduction. Cell Biochem Biophys 2003, 39, 45–60. [CrossRef]
- Hynes, N.E.; Ingham, P.W.; Lim, W.A.; Marshall, C.J.; Massagué, J.; Pawson, T. Signalling Change: Signal Transduction through the Decades. Nat Rev Mol Cell Biol 2013, 14, 393–398. [CrossRef]
- Lee, M.J.; Yaffe, M.B. Protein Regulation in Signal Transduction. Cold Spring Harb Perspect Biol 2016, 8. [CrossRef]
- Pawson, C.T.; Scott, J.D. Signal Integration through Blending, Bolstering and Bifurcating of Intracellular Information. Nat Struct Mol Biol 2010, 17, 653–658. [CrossRef]
- González, A.; Hall, M.N. Nutrient Sensing and TOR Signaling in Yeast and Mammals. EMBO J 2017, 36, 397–408. [CrossRef]
- Chasman, D.; Ho, Y.; Berry, D.B.; Nemec, C.M.; MacGilvray, M.E.; Hose, J.; Merrill, A.E.; Lee, M.V.; Will, J.L.; Coon, J.J.; et al. Pathway Connectivity and Signaling Coordination in the Yeast Stress-Activated Signaling Network. Mol Syst Biol 2014, 10, 759. [CrossRef]
- Zhang, P.; Kornev, A.P.; Wu, J.; Taylor, S.S. Discovery of Allostery in PKA Signaling. Biophys Rev 2015, 7, 227–238. [CrossRef]
- Taylor, S.S.; Wu, J.; Bruystens, J.G.H.; Del Rio, J.C.; Lu, T.W.; Kornev, A.P.; Ten Eyck, L.F. From Structure to the Dynamic Regulation of a Molecular Switch: A Journey over 3 Decades. J Biol Chem 2021, 296. [CrossRef]
- Toda, T.; Cameron, S.; Sass, P.; Zoller, M.; Wigler, M. Three Different Genes in S. cerevisiae Encode the Catalytic Subunits of the CAMP-Dependent Protein Kinase. Cell 1987, 50, 277–287.
- Rubio-Texeira, M.; Van Zeebroeck, G.; Voordeckers, K.; Thevelein, J.M. Saccharomyces cerevisiae Plasma Membrane Nutrient Sensors and Their Role in PKA Signaling. FEMS Yeast Res 2010, 10, 134–149. [CrossRef]
- Conrad, M.; Schothorst, J.; Kankipati, H.N.; Van Zeebroeck, G.; Rubio-Texeira, M.; Thevelein, J.M. Nutrient Sensing and Signaling in the Yeast Saccharomyces cerevisiae. FEMS Microbiol Rev 2014, 38, 254–299. [CrossRef]
- Thevelein, J.M.; Geladé, R.; Holsbeeks, I.; Lagatie, O.; Popova, Y.; Rolland, F.; Stolz, F.; Van de Velde, S.; Van Dijck, P.; Vandormael, P.; et al. Nutrient Sensing Systems for Rapid Activation of the Protein Kinase A Pathway in Yeast. Biochem Soc Trans 2005, 33, 253–256. [CrossRef]
- Berry, D.; Gasch, A. Stress-Activated Genomic Expression Changes Serve a Preparative Role for Impending Stress in Yeast. Mol Biol Cell 2008, 19, 4580 – 4587. [CrossRef]
- Gancedo, J.M. The Early Steps of Glucose Signalling in Yeast. FEMS Microbiol Rev 2008, 32, 673–704. [CrossRef]
- Palecek, S.P.; Parikh, A.S.; Kron, S.J. Sensing, Signalling and Integrating Physical Processes during Saccharomyces cerevisiae Invasive and Filamentous Growth. Microbiology (Reading) 2002, 148, 893–907.
- Portela, P.; Rossi, S. CAMP-PKA Signal Transduction Specificity in Saccharomyces cerevisiae. Curr Genet 2020, 66, 1093–1099. [CrossRef]
- Santangelo, G.M. Glucose Signaling in Saccharomyces cerevisiae. Microbiol Mol Biol Rev 2006, 70, 253–282. [CrossRef]
- Causton, H.C.; Ren, B.; Koh, S.S.; Harbison, C.T.; Kanin, E.; Jennings, E.G.; Lee, T.I.; True, H.L.; Lander, E.S.; Young, R.A. Remodeling of Yeast Genome Expression in Response to Environmental Changes. Mol Biol Cell 2001, 12, 323–337.
- Gasch, A.P.; Werner-Washburne, M. The Genomics of Yeast Responses to Environmental Stress and Starvation. Funct Integr Genomics 2002, 2, 181–192. [CrossRef]
- Zhang, L.; Winkler, S.; Schlottmann, F.P.; Kohlbacher, O.; Elias, J.E.; Skotheim, J.M.; Ewald, J.C. Multiple Layers of Phospho-Regulation Coordinate Metabolism and the Cell Cycle in Budding Yeast. Front Cell Dev Biol 2019, 7, 338. [CrossRef]
- Grossbach, J.; Gillet, L.; Cl Ement-Ziza, M.; Schmalohr, C.L.; Schubert, O.T.; Sch€ Utter, M.; Mawer, J.S.P.; Barnes, C.A.; Bludau, I.; Weith, M.; et al. The Impact of Genomic Variation on Protein Phosphorylation States and Regulatory Networks. Mol Syst Biol 2022, 18, e10712. [CrossRef]
- Kanshin, E.; Kubiniok, P.; Thattikota, Y.; Amours, D.D.’; Thibault, P. Phosphoproteome Dynamics of Saccharomyces cerevisiae under Heat Shock and Cold Stress. Mol Syst Biol 2015, 11, 813. [CrossRef]
- Hohmann, S.; Mager, W.H. Yeast Stress Responses; Springer, 2003; ISBN 9783540456117.
- Ruis, H.; Schüller, C. Stress Signaling in Yeast. Bioessays 1995, 17, 959–965. [CrossRef]
- Saini, P.; Beniwal, A.; Kokkiligadda, A.; Vij, S. Response and Tolerance of Yeast to Changing Environmental Stress during Ethanol Fermentation. Process Biochemistry 2018, 72, 1–12. [CrossRef]
- Thevelein, J.M.; De Winde, J.H. Novel Sensing Mechanisms and Targets for the CAMP–Protein Kinase A Pathway in the Yeast Saccharomyces cerevisiae. Mol Microbiol 1999, 33, 904–918. [CrossRef]
- Russell, M.; Bradshaw-Rouse, J.; Markwardt, D.; Heideman, W. Changes in Gene Expression in the Ras/Adenylate Cyclase System of Saccharomyces cerevisiae: Correlation with CAMP Levels and Growth Arrest. Mol Biol Cell 1993, 4, 757–765. [CrossRef]
- Smith, A.; Ward, M.P.; Garrett, S. Yeast PKA Represses Msn2p/Msn4p-Dependent Gene Expression to Regulate Growth, Stress Response and Glycogen Accumulation. EMBO Journal 1998, 17, 3556–3564. [CrossRef]
- Boy-Marcotte, E.; Garreau, H.; Jacquet, M. Cyclic AMP Controls the Switch between Division Cycle and Resting State Programs in Response to Ammonium Availability in Saccharomyces cerevisiae. Yeast 1987, 3, 85–93. [CrossRef]
- Durchschlag, E.; Reiter, W.; Ammerer, G.; Schüller, C. Nuclear Localization Destabilizes the Stress-Regulated Transcription Factor Msn2. J Biol Chem 2004, 279, 55425–55432. [CrossRef]
- Görner, W.; Durchschlag, E.; Martinez-Pastor, M.T.; Estruch, F.; Ammerer, G.; Hamilton, B.; Ruis, H.; Schüller, C. Nuclear Localization of the C2H2 Zinc Finger Protein Msn2p Is Regulated by Stress and Protein Kinase A Activity. Genes Dev 1998, 12, 586–597. [CrossRef]
- Cai, L.; Dalal, C.K.; Elowitz, M.B. Frequency-Modulated Nuclear Localization Bursts Coordinate Gene Regulation. Nature 2008 455:7212 2008, 455, 485–490. [CrossRef]
- Garmendia-Torres, C.; Goldbeter, A.; Jacquet, M. Nucleocytoplasmic Oscillations of the Yeast Transcription Factor Msn2: Evidence for Periodic PKA Activation. Curr Biol 2007, 17, 1044–1049. [CrossRef]
- Jacquet, M.; Renault, G.; Lallet, S.; De Mey, J.; Goldbeter, A. Oscillatory Behavior of the Nuclear Localization of the Transcription Factors Msn2 and Msn4 in Response to Stress in Yeast. ScientificWorldJournal 2003, 3, 609–612. [CrossRef]
- Sadeh, A.; Movshovich, N.; Volokh, M.; Gheber, L.; Aharoni, A. Fine-Tuning of the Msn2/4-Mediated Yeast Stress Responses as Revealed by Systematic Deletion of Msn2/4 Partners. Mol Biol Cell 2011, 22, 3127–3138. [CrossRef]
- Zaman, S.; Lippman, S.I.; Zhao, X.; Broach, J.R. How Saccharomyces Responds to Nutrients. Annu Rev Genet 2008, 42, 27–81. [CrossRef]
- Hohmann, S. An Integrated View on a Eukaryotic Osmoregulation System. Curr Genet 2015, 61, 373–382. [CrossRef]
- Tamás, M.J.; Karlgren, S.; Bill, R.M.; Hedfalk, K.; Allegri, L.; Ferreira, M.; Thevelein, J.M.; Rydström, J.; Mullins, J.G.L.; Hohmann, S. A Short Regulatory Domain Restricts Glycerol Transport through Yeast Fps1p. J Biol Chem 2003, 278, 6337–6345. [CrossRef]
- Tamás, M.J.; Luyten, K.; Sutherland, F.C.W.; Hernandez, A.; Albertyn, J.; Valadi, H.; Li, H.; Prior, B.A.; Kilian, S.G.; Ramos, J.; et al. Fps1p Controls the Accumulation and Release of the Compatible Solute Glycerol in Yeast Osmoregulation. Mol Microbiol 1999, 31, 1087–1104. [CrossRef]
- Tanaka, K.; Tatebayashi, K.; Nishimura, A.; Yamamoto, K.; Yang, H.Y.; Saito, H. Yeast Osmosensors Hkr1 and Msb2 Activate the Hog1 MAPK Cascade by Different Mechanisms. Sci Signal 2014, 7. [CrossRef]
- Macia, J.; Regot, S.; Peeters, T.; Conde, N.; Solé, R.; Posas, F. Dynamic Signaling in the Hog1 MAPK Pathway Relies on High Basal Signal Transduction. Sci Signal 2009, 2. [CrossRef]
- Raitt, D.C.; Posas, F.; Saito, H. Yeast Cdc42 GTPase and Ste20 PAK-like Kinase Regulate Sho1-Dependent Activation of the Hog1 MAPK Pathway. EMBO J 2000, 19, 4623–4631. [CrossRef]
- Saito, H.; Tatebayashi, K. Regulation of the Osmoregulatory HOG MAPK Cascade in Yeast. J Biochem 2004, 136, 267–272. [CrossRef]
- Vázquez-Ibarra, A.; Rodríguez-Martínez, G.; Guerrero-Serrano, G.; Kawasaki, L.; Ongay-Larios, L.; Coria, R. Negative Feedback-Loop Mechanisms Regulating HOG- and Pheromone-MAPK Signaling in Yeast. Curr Genet 2020, 66, 867–880. [CrossRef]
- Brewster, J.L.; De Valoir, T.; Dwyer, N.D.; Winter, E.; Gustin, M.C. An Osmosensing Signal Transduction Pathway in Yeast. Science 1993, 259, 1760–1763. [CrossRef]
- Maeda, T.; Takekawa, M.; Saito, H. Activation of Yeast PBS2 MAPKK by MAPKKKs or by Binding of an SH3-Containing Osmosensor. Science 1995, 269, 554–558. [CrossRef]
- Posas, F.; Saito, H. Osmotic Activation of the HOG MAPK Pathway via Ste11p MAPKKK: Scaffold Role of Pbs2p MAPKK. Science 1997, 276, 1702–1708. [CrossRef]
- Saito, H.; Posas, F. Response to Hyperosmotic Stress. Genetics 2012, 192, 289–318. [CrossRef]
- Tatebayashi, K.; Saito, H. Two Activating Phosphorylation Sites of Pbs2 MAP2K in the Yeast HOG Pathway Are Differentially Dephosphorylated by Four PP2C Phosphatases Ptc1-Ptc4. J Biol Chem 2023, 299, 104569. [CrossRef]
- Ferrigno, P.; Posas, F.; Koepp, D.; Saito, H.; Silver, P.A. Regulated Nucleo/Cytoplasmic Exchange of HOG1 MAPK Requires the Importin Beta Homologs NMD5 and XPO1. EMBO J 1998, 17, 5606–5614. [CrossRef]
- Hoffmann, E.K.; Lambert, I.H.; Pedersen, S.F. Physiology of Cell Volume Regulation in Vertebrates. Physiol Rev 2009, 89, 193–277. [CrossRef]
- Alepuz, P.M.; Jovanovic, A.; Reiser, V.; Ammerer, G. Stress-Induced Map Kinase Hog1 Is Part of Transcription Activation Complexes. Mol Cell 2001, 7, 767–777. [CrossRef]
- Gomar-Alba, M.; Alepuz, P.; Del Olmo, M. Dissection of the Elements of Osmotic Stress Response Transcription Factor Hot1 Involved in the Interaction with MAPK Hog1 and in the Activation of Transcription. Biochim Biophys Acta 2013, 1829, 1111–1125. [CrossRef]
- Rep, M.; Krantz, M.; Thevelein, J.M.; Hohmann, S. The Transcriptional Response of Saccharomyces cerevisiae to Osmotic Shock. Hot1p and Msn2p/Msn4p Are Required for the Induction of Subsets of High Osmolarity Glycerol Pathway-Dependent Genes. J Biol Chem 2000, 275, 8290–8300. [CrossRef]
- Fischer, J.; Song, Y.S.; Yosef, N.; Iulio, J. di; Stirling Churchman, L.; Choder, M. The Yeast Exoribonuclease Xrn1 and Associated Factors Modulate RNA Polymerase II Processivity in 5’ and 3’ Gene Regions. J Biol Chem 2020, 295, 11435–11454. [CrossRef]
- Proft, M.; Struhl, K. Hog1 Kinase Converts the Sko1-Cyc8-Tup1 Repressor Complex into an Activator That Recruits SAGA and SWI/SNF in Response to Osmotic Stress. Mol Cell 2002, 9, 1307–1317. [CrossRef]
- Huang, C.J.; Lu, M.Y.; Chang, Y.W.; Li, W.H. Experimental Evolution of Yeast for High-Temperature Tolerance. Mol Biol Evol 2018, 35, 1823–1839. [CrossRef]
- Klopf, E.; Schmidt, H.A.; Clauder-Münster, S.; Steinmetz, L.M.; Schüller, C. INO80 Represses Osmostress Induced Gene Expression by Resetting Promoter Proximal Nucleosomes. Nucleic Acids Res 2017, 45, 3752–3766. [CrossRef]
- Norbeck, J.; Blomberg, A. The Level of CAMP-Dependent Protein Kinase A Activity Strongly Affects Osmotolerance and Osmo-Instigated Gene Expression Changes in Saccharomyces cerevisiae. Yeast 2000, 16, 121–137. [CrossRef]
- Hohmann, S. Osmotic Adaptation in Yeast-Control of the Yeast Osmolyte System. Int Rev Cytol 2002, 215, 149–187. [CrossRef]
- W, G.; E, D.; MT, M.-P.; F, E.; G, A.; B, H.; H, R.; C, S. Nuclear Localization of the C2H2 Zinc Finger Protein Msn2p Is Regulated by Stress and Protein Kinase A Activity. Genes Dev 1998, 12, 586–597. [CrossRef]
- González-Rubio, G.; Sellers-Moya, A.; Martín, H.; Molina, M. A Walk-through MAPK Structure and Functionality with the 30-Year-Old Yeast MAPK Slt2. Int Microbiol 2021. [CrossRef]
- Lallet, S.; Garreau, H.; Garmendia-Torres, C.; Szestakowska, D.; Boy-Marcotte, E.; Quevillon-Chéruel, S.; Jacquet, M. Role of Gal11, a Component of the RNA Polymerase II Mediator in Stress-Induced Hyperphosphorylation of Msn2 in Saccharomyces cerevisiae. Mol Microbiol 2006, 62, 438–452. [CrossRef]
- Garreau, H.; Hasan, R.N.; Renault, G.; Estruch, F.; Boy-Marcotte, E.; Jacquet, M. Hyperphosphorylation of Msn2p and Msn4p in Response to Heat Shock and the Diauxic Shift Is Inhibited by CAMP in Saccharomyces cerevisiae. Microbiology (Reading) 2000, 146 ( Pt 9), 2113–2120. [CrossRef]
- Boy-Marcotte, E.; Perrot, M.; Bussereau, F.; Boucherie, H.; Jacquet, M. Msn2p and Msn4p Control a Large Number of Genes Induced at the Diauxic Transition Which Are Repressed by Cyclic AMP in Saccharomyces cerevisiae. J Bacteriol 1998, 180, 1044–1052. [CrossRef]
- Lee, P.; Cho, B.R.; Joo, H.S.; Hahn, J.S. Yeast Yak1 Kinase, a Bridge between PKA and Stress-Responsive Transcription Factors, Hsf1 and Msn2/Msn4. Mol Microbiol 2008, 70, 882–895. [CrossRef]
- Rep, M.; Krantz, M.; Thevelein, J.M.; Hohmann, S. The Transcriptional Response of Saccharomyces cerevisiae to Osmotic Shock. Hot1p and Msn2p/Msn4p Are Required for the Induction of Subsets of High Osmolarity Glycerol Pathway-Dependent Genes. J Biol Chem 2000, 275, 8290–8300. [CrossRef]
- Capaldi, A.P.; Kaplan, T.; Liu, Y.; Habib, N.; Regev, A.; Friedman, N.; O’shea, E.K. Structure and Function of a Transcriptional Network Activated by the MAPK Hog1. Nat Genet 2008, 40, 1300–1306. [CrossRef]
- Pokholok, D.K.; Zeitlinger, J.; Hannett, N.M.; Reynolds, D.B.; Young, R.A. Activated Signal Transduction Kinases Frequently Occupy Target Genes. Science 2006, 313, 533–536. [CrossRef]
- De Nadal, E.; Zapater, M.; Alepuz, P.M.; Sumoy, L.; Mas, G.; Posas, F. The MAPK Hog1 Recruits Rpd3 Histone Deacetylase to Activate Osmoresponsive Genes. Nature 2004, 427, 370–374. [CrossRef]
- Baccarini, L.; Martínez-Montañés, F.; Rossi, S.; Proft, M.; Portela, P. PKA-Chromatin Association at Stress Responsive Target Genes from Saccharomyces cerevisiae. Biochimica et Biophysica Acta (BBA) - Gene Regulatory Mechanisms 2015, 1849, 1329–1339. [CrossRef]
- Vert, G.; Chory, J. Crosstalk in Cellular Signaling: Background Noise or the Real Thing? Dev Cell 2011, 21, 985. [CrossRef]
- González-Rubio, G.; Fernández-Acero, T.; Martín, H.; Molina, M. Mitogen-Activated Protein Kinase Phosphatases (MKPs) in Fungal Signaling: Conservation, Function, and Regulation. Int J Mol Sci 2019, 20. [CrossRef]
- Chavel, C.A.; Caccamise, L.M.; Li, B.; Cullen, P.J. Global Regulation of a Differentiation MAPK Pathway in Yeast. Genetics 2014, 198, 1309–1328. [CrossRef]
- Baltanás, R.; Bush, A.; Couto, A.; Durrieu, L.; Hohmann, S.; Colman-Lerner, A. Pheromone-Induced Morphogenesis Improves Osmoadaptation Capacity by Activating the HOG MAPK Pathway. Sci Signal 2013, 6. [CrossRef]
- O’Rourke, S.M.; Herskowitz, I. The Hog1 MAPK Prevents Cross Talk between the HOG and Pheromone Response MAPK Pathways in Saccharomyces cerevisiae. Genes Dev 1998, 12, 2874–2886. [CrossRef]
- Levin, D.E. Regulation of Cell Wall Biogenesis in Saccharomyces cerevisiae: The Cell Wall Integrity Signaling Pathway. Genetics 2011, 189, 1145–1175. [CrossRef]
- Jiménez-Gutiérrez, E.; Alegría-Carrasco, E.; Sellers-Moya, A.; Molina, M.; Martín, H. Not Just the Wall: The Other Ways to Turn the Yeast CWI Pathway On. Int Microbiol 2020, 23, 107–119. [CrossRef]
- Birkaya, B.; Maddi, A.; Joshi, J.; Free, S.J.; Cullen, P.J. Role of the Cell Wall Integrity and Filamentous Growth Mitogen-Activated Protein Kinase Pathways in Cell Wall Remodeling during Filamentous Growth. Eukaryot Cell 2009, 8, 1118–1133. [CrossRef]
- Buehrer, B.M.; Errede, B. Coordination of the Mating and Cell Integrity Mitogen-Activated Protein Kinase Pathways in Saccharomyces cerevisiae. Mol Cell Biol 1997, 17, 6517–6525. [CrossRef]
- Zarzov, P.; Mazzoni, C.; Mann, C. The SLT2(MPK1) MAP Kinase Is Activated during Periods of Polarized Cell Growth in Yeast. EMBO J 1996, 15, 83–91. [CrossRef]
- Urita, A.; Ishibashi, Y.; Kawaguchi, R.; Yanase, Y.; Tani, M. Crosstalk between Protein Kinase A and the HOG Pathway under Impaired Biosynthesis of Complex Sphingolipids in Budding Yeast. FEBS J 2022, 289, 766–786. [CrossRef]
- Ojeda LE Interaction between the CAMP-PKA and HOG-MAPK Pathways in the Adaptive Cellular Response to Osmotic Stress in S. cerevisiae., Departamento Química Biológica, Facultad de Ciencias Exactas,UBA, 2022.
- Ojeda LE; Gulias F; Ortola M; Galello FA; Rossi SG; Bermudez Moretti M and Portela, P. Crosstalk between CAMP-PKA and Hog-MAPK Pathways in the Regulation of the Osmotic Stress Response in S. cerevisiae. In Proceedings of the BIOCELL 46 (Suppl. 1); 2022; p. 63.
- Erdman, S.; Snyder, M. A Filamentous Growth Response Mediated by the Yeast Mating Pathway. Genetics 2001, 159, 919–928. [CrossRef]
- Hao, N.; Nayak, S.; Behar, M.; Shanks, R.H.; Nagiec, M.J.; Errede, B.; Hasty, J.; Elston, T.C.; Dohlman, H.G. Regulation of Cell Signaling Dynamics by the Protein Kinase-Scaffold Ste5. Mol Cell 2008, 30, 649–656. [CrossRef]
- Sieber, B.; Coronas-Serna, J.M.; Martin, S.G. A Focus on Yeast Mating: From Pheromone Signaling to Cell-Cell Fusion. Semin Cell Dev Biol 2023, 133, 83–95. [CrossRef]
- Van Drogen, F.; Dard, N.; Pelet, S.; Lee, S.S.; Mishra, R.; Srejić, N.; Peter, M. Crosstalk and Spatiotemporal Regulation between Stress-Induced MAP Kinase Pathways and Pheromone Signaling in Budding Yeast. Cell Cycle 2020, 19, 1707–1715. [CrossRef]
- Kumar, A. The Complex Genetic Basis and Multilayered Regulatory Control of Yeast Pseudohyphal Growth. Annu Rev Genet 2021, 55, 1–21. [CrossRef]
- Cullen, P.J.; Sprague, G.F. The Regulation of Filamentous Growth in Yeast. Genetics 2012, 190, 23–49. [CrossRef]
- Pan, X.; Heitman, J. Protein Kinase A Operates a Molecular Switch That Governs Yeast Pseudohyphal Differentiation. Mol Cell Biol 2002, 22, 3981–3993.
- Robertson, L.S.; Fink, G.R. The Three Yeast A Kinases Have Specific Signaling Functions in Pseudohyphal Growth. Proc Natl Acad Sci U S A 1998, 95, 13783–13787.
- Gasch, A.P.; Spellman, P.T.; Kao, C.M.; Carmel-Harel, O.; Eisen, M.B.; Storz, G.; Botstein, D.; Brown, P.O. Genomic Expression Programs in the Response of Yeast Cells to Environmental Changes. Mol Biol Cell 2000, 11, 4241–4257. [CrossRef]
- Verghese, J.; Abrams, J.; Wang, Y.; Morano, K.A. Biology of the Heat Shock Response and Protein Chaperones: Budding Yeast (Saccharomyces cerevisiae) as a Model System. Microbiol Mol Biol Rev 2012, 76, 115–158. [CrossRef]
- Morimoto, R.I.; Westerheide, S.D. The Heat Shock Response and the Stress of Misfolded Proteins. Handbook of Cell Signaling, Second Edition 2010, 3, 2231–2239. [CrossRef]
- Keuenhof, K.S.; Berglund, L.L.; Hill, S.M.; Schneider, K.L.; Widlund, P.O.; Nyström, T.; Höög, J.L. Large Organellar Changes Occur during Mild Heat Shock in Yeast. J Cell Sci 2022, 135. [CrossRef]
- Mühlhofer, M.; Berchtold, E.; Stratil, C.G.; Csaba, G.; Kunold, E.; Bach, N.C.; Sieber, S.A.; Haslbeck, M.; Zimmer, R.; Buchner, J. The Heat Shock Response in Yeast Maintains Protein Homeostasis by Chaperoning and Replenishing Proteins. Cell Rep 2019, 29, 4593-4607.e8. [CrossRef]
- Castells-Roca, L.; García-Martínez, J.; Moreno, J.; Herrero, E.; Bellí, G.; Pérez-Ortín, J.E. Heat Shock Response in Yeast Involves Changes in Both Transcription Rates and MRNA Stabilities. PLoS One 2011, 6, e17272. [CrossRef]
- Wang, L.; Renault, G.; Garreau, H.; Jacquet, M. Stress Induces Depletion of Cdc25p and Decreases the CAMP Producing Capability in Saccharomyces cerevisiae. Microbiology (Reading) 2004, 150, 3383–3391. [CrossRef]
- Ivanov, P.; Kedersha, N.; Anderson, P. Stress Granules and Processing Bodies in Translational Control. Cold Spring Harb Perspect Biol 2019, 11. [CrossRef]
- Grousl, T.; Vojtova, J.; Hasek, J.; Vomastek, T. Yeast Stress Granules at a Glance. Yeast 2022, 39, 247–261. [CrossRef]
- Jeon, P.; Ham, H.J.; Park, S.; Lee, J.A. Regulation of Cellular Ribonucleoprotein Granules: From Assembly to Degradation via Post-Translational Modification. Cells 2022, 11. [CrossRef]
- Shah, K.H.; Zhang, B.; Ramachandran, V.; Herman, P.K. Processing Body and Stress Granule Assembly Occur by Independent and Differentially Regulated Pathways in Saccharomyces cerevisiae. Genetics 2013, 193, 109–123. [CrossRef]
- Kozubowski, L.; Aboobakar, E.F.; Cardenas, M.E.; Heitman, J. Calcineurin Colocalizes with P-Bodies and Stress Granules during Thermal Stress in Cryptococcus Neoformans. Eukaryot Cell 2011, 10, 1396–1402. [CrossRef]
- Mitchell, S.F.; Jain, S.; She, M.; Parker, R. Global Analysis of Yeast MRNPs. Nat Struct Mol Biol 2013, 20, 127–133. [CrossRef]
- Tudisca, V.; Recouvreux, V.; Moreno, S.; Boy-Marcotte, E.; Jacquet, M.; Portela, P. Differential Localization to Cytoplasm, Nucleus or P-Bodies of Yeast PKA Subunits under Different Growth Conditions. Eur J Cell Biol 2010, 89, 339–348. [CrossRef]
- Youn, J.Y.; Dyakov, B.J.A.; Zhang, J.; Knight, J.D.R.; Vernon, R.M.; Forman-Kay, J.D.; Gingras, A.C. Properties of Stress Granule and P-Body Proteomes. Mol Cell 2019, 76, 286–294.
- Ramachandran, V.; Shah, K.H.; Herman, P.K. The CAMP-Dependent Protein Kinase Signaling Pathway Is a Key Regulator of P Body Foci Formation. Mol Cell 2011, 43, 973–981. [CrossRef]
- Barraza, C.E.; Solari, C.A.; Marcovich, I.; Kershaw, C.; Galello, F.; Rossi, S.; Ashe, M.P.; Portela, P. The Role of PKA in the Translational Response to Heat Stress in Saccharomyces cerevisiae. PLoS One 2017, 12, e0185416. [CrossRef]
- Pautasso, C.; Rossi, S. Transcriptional Regulation of the Protein Kinase A Subunits in Saccharomyces cerevisiae: Autoregulatory Role of the Kinase A Activity. Biochim Biophys Acta Gene Regul Mech 2014, 1839, 275–287. [CrossRef]
- Engelberg, D.; Perlman, R.; Levitzki, A. Transmembrane Signaling in Saccharomyces cerevisiae as a Model for Signaling in Metazoans: State of the Art after 25 Years. Cell Signal 2014, 26, 2865–2878. [CrossRef]
- Torres, L.; Martín, H.; García-Saez, M.I.; Arroyo, J.; Molina, M.; Sánchez, M.; Nombela, C. A Protein Kinase Gene Complements the Lytic Phenotype of Saccharomyces cerevisiae Lyt2 Mutants. Mol Microbiol 1991, 5, 2845–2854. [CrossRef]
- Kock, C.; Dufrêne, Y.; Heinisch, J. Up against the Wall: Is Yeast Cell Wall Integrity Ensured by Mechanosensing in Plasma Membrane Microdomains? Appl Environ Microbiol 2015, 81, 806–811. [CrossRef]
- Rodicio, R.; Heinisch, J.J. Together We Are Strong - Cell Wall Integrity Sensors in Yeasts. Yeast 2010, 27, 531–540. [CrossRef]
- Verna, J.; Lodder, A.; Lee, K.; Vagts, A.; Ballester, R. A Family of Genes Required for Maintenance of Cell Wall Integrity and for the Stress Response in Saccharomyces cerevisiae. Proc Natl Acad Sci U S A 1997, 94, 13804–13809. [CrossRef]
- Ketela, T.; Green, R.; Bussey, H. Saccharomyces cerevisiae Mid2p Is a Potential Cell Wall Stress Sensor and Upstream Activator of the PKC1-MPK1 Cell Integrity Pathway. J Bacteriol 1999, 181, 3330–3340. [CrossRef]
- Zu, T.; Verna, J.; Ballester, R. Mutations in WSC Genes for Putative Stress Receptors Result in Sensitivity to Multiple Stress Conditions and Impairment of Rlm1-Dependent Gene Expression in Saccharomyces cerevisiae. Molecular Genetics and Genomics 2001, 266, 142–155. [CrossRef]
- Harrison, J.C.; Bardes, E.S.G.; Ohya, Y.; Lew, D.J. A Role for the Pkc1p/Mpk1p Kinase Cascade in the Morphogenesis Checkpoint. Nat Cell Biol 2001, 3, 417–420. [CrossRef]
- Harrison, J.C.; Zyla, T.R.; Bardes, E.S.G.; Lew, D.J. Stress-Specific Activation Mechanisms for the “Cell Integrity” MAPK Pathway. Journal of Biological Chemistry 2004, 279, 2616–2622. [CrossRef]
- Krause, S.A.; Gray, J. V. The Protein Kinase C Pathway Is Required for Viability in Quiescence in Saccharomyces cerevisiae. Current Biology 2002, 12, 588–593. [CrossRef]
- Hahn, J.S.; Thiele, D.J. Regulation of the Saccharomyces cerevisiae Slt2 Kinase Pathway by the Stress-Inducible Sdp1 Dual Specificity Phosphatase. Journal of Biological Chemistry 2002, 277, 21278–21284. [CrossRef]
- Cañonero, L.; Pautasso, C.; Galello, F.; Sigaut, L.; Pietrasanta, L.; Javier, A.; Bermúdez-Moretti, M.; Portela, P.; Rossi, S. Heat Stress Regulates the Expression of TPK1 Gene at Transcriptional and Post-Transcriptional Levels in Saccharomyces cerevisiae. Biochim Biophys Acta Mol Cell Res 2022, 1869. [CrossRef]
- Robertson, L.S.; Causton, H.C.; Young, R.A.; Fink, G.R. The Yeast A Kinases Differentially Regulate Iron Uptake and Respiratory Function. Proceedings of the National Academy of Sciences 2000, 97, 5984–5988. [CrossRef]
- Chevtzoff, C.; Vallortigara, J.; Avéret, N.; Rigoulet, M.; Devin, A. The Yeast CAMP Protein Kinase Tpk3p Is Involved in the Regulation of Mitochondrial Enzymatic Content during Growth. Biochimica et Biophysica Acta (BBA) - Bioenergetics 2005, 1706, 117–125. [CrossRef]
- Palomino, A.; Herrero, P.; Moreno, F. Tpk3 and Snf1 Protein Kinases Regulate Rgt1 Association with Saccharomyces cerevisiae HXK2 Promoter. Nucleic Acids Res 2006, 34, 1427–1438. [CrossRef]
- Yu, Q.; Gong, X.; Tong, Y.; Wang, M.; Duan, K.; Zhang, X.; Ge, F.; Yu, X.; Li, S. Phosphorylation of Jhd2 by the Ras-CAMP-PKA(Tpk2) Pathway Regulates Histone Modifications and Autophagy. Nat Commun 2022, 13. [CrossRef]
- Griffioen, G.; Anghileri, P.; Imre, E.; Baroni, M.D.; Ruis, H. Nutritional Control of Nucleocytoplasmic Localization of CAMP-Dependent Protein Kinase Catalytic and Regulatory Subunits in Saccharomyces cerevisiae. J Biol Chem 2000, 275, 1449–1456. [CrossRef]
- Griffioen, G.; Thevelein, J.M. Molecular Mechanisms Controlling the Localisation of Protein Kinase A. Curr Genet 2002, 41, 199–207. [CrossRef]
- González Bardeci, N.; Caramelo, J.J.; Blumenthal, D.K.; Rinaldi, J.; Rossi, S.; Moreno, S. The PKA Regulatory Subunit from Yeast Forms a Homotetramer: Low-Resolution Structure of the N-Terminal Oligomerization Domain. J Struct Biol 2016, 193. [CrossRef]
- Newlon, M.G.; Roy, M.; Morikis, D.; Carr, D.W.; Westphal, R.; Scott, J.D.; Jennings, P.A. A Novel Mechanism of PKA Anchoring Revealed by Solution Structures of Anchoring Complexes. EMBO J 2001, 20, 1651–1662. [CrossRef]
- Gold, M.G.; Lygren, B.; Dokurno, P.; Hoshi, N.; McConnachie, G.; Taskén, K.; Carlson, C.R.; Scott, J.D.; Barford, D. Molecular Basis of AKAP Specificity for PKA Regulatory Subunits. Mol Cell 2006, 24, 383–395. [CrossRef]
- Sarma, G.N.; Kinderman, F.S.; Kim, C.; von Daake, S.; Chen, L.; Wang, B.-C.; Taylor, S.S. Structure of D-AKAP2:PKA RI Complex: Insights into AKAP Specificity and Selectivity. Structure 2010, 18, 155–166. [CrossRef]
- Griffioen, G.; Branduardi, P.; Ballarini, A.; Anghileri, P.; Norbeck, J.; Baroni, M.D.; Ruis, H. Nucleocytoplasmic Distribution of Budding Yeast Protein Kinase A Regulatory Subunit Bcy1 Requires Zds1 and Is Regulated by Yak1-Dependent Phosphorylation of Its Targeting Domain. Mol Cell Biol 2001, 21, 511–523. [CrossRef]
- Galello, F.; Moreno, S.; Rossi, S. Interacting Proteins of Protein Kinase A Regulatory Subunit in Saccharomyces cerevisiae. J Proteomics 2014, 109, 261–275. [CrossRef]
- Tudisca, V.; Recouvreux, V.; Moreno, S.; Boy-Marcotte, E.; Jacquet, M.; Portela, P. Differential Localization to Cytoplasm, Nucleus or P-Bodies of Yeast PKA Subunits under Different Growth Conditions. Eur J Cell Biol 2010, 89, 339–348. [CrossRef]
- Barraza, C.E.; Solari, C.A.; Rinaldi, J.; Ojeda, L.; Rossi, S.; Ashe, M.P.; Portela, P. A Prion-like Domain of Tpk2 Catalytic Subunit of Protein Kinase A Modulates P-Body Formation in Response to Stress in Budding Yeast. Biochim Biophys Acta Mol Cell Res 2021, 1868. [CrossRef]
- Barraza, C.E.; Solari, C.A.; Rinaldi, J.; Ojeda, L.; Rossi, S.; Ashe, M.P.; Portela, P. A Prion-like Domain of Tpk2 Catalytic Subunit of Protein Kinase A Modulates P-Body Formation in Response to Stress in Budding Yeast. Biochim Biophys Acta Mol Cell Res 2021, 1868. [CrossRef]
- Rep, M.; Krantz, M.; Thevelein, J.M.; Hohmann, S. The Transcriptional Response of Saccharomyces cerevisiae to Osmotic Shock. Hot1p and Msn2p/Msn4p Are Required for the Induction of Subsets of High Osmolarity Glycerol Pathway-Dependent Genes. J Biol Chem 2000, 275, 8290–8300.
- Posas, F.; Chambers, J.R.; Heyman, J.A.; Hoeffler, J.P.; de Nadal, E.; Ariño, J. The Transcriptional Response of Yeast to Saline Stress. J Biol Chem 2000, 275, 17249–17255. [CrossRef]
- Gasch, A.P.; Spellman, P.T.; Kao, C.M.; Carmel-Harel, O.; Eisen, M.B.; Storz, G.; Botstein, D.; Brown, P.O. Genomic Expression Programs in the Response of Yeast Cells to Environmental Changes. Mol Biol Cell 2000, 11, 4241–4257. [CrossRef]
- Yale, J.; Bohnert, H.J. Transcript Expression in Saccharomyces cerevisiae at High Salinity. J Biol Chem 2001, 276, 15996–16007. [CrossRef]
- Castells-Roca, L.; Mühlenhoff, U.; Lill, R.; Herrero, E.; Bellí, G. The Oxidative Stress Response in Yeast Cells Involves Changes in the Stability of Aft1 Regulon MRNAs. Mol Microbiol 2011, 81, 232–248. [CrossRef]
- Galello, F.; Pautasso, C.; Reca, S.; Cañonero, L.; Portela, P.; Moreno, S.; Rossi, S. Transcriptional Regulation of the Protein Kinase a Subunits in Saccharomyces cerevisiae during Fermentative Growth. Yeast 2017, 34, 495–508. [CrossRef]
- Reca, S.; Galello, F.; Ojeda, L.; Pautasso, C.; Cañonero, L.; Moreno, S.; Portela, P.; Rossi, S. Chromatin Remodeling and Transcription of the TPK1 Subunit of PKA during Stress in Saccharomyces cerevisiae. Biochim Biophys Acta Gene Regul Mech 2020, 1863, 194599. [CrossRef]
Figure 1.
Model of the crosstalk between cAMP-PKA and HOG pathways in response to osmotic stress.
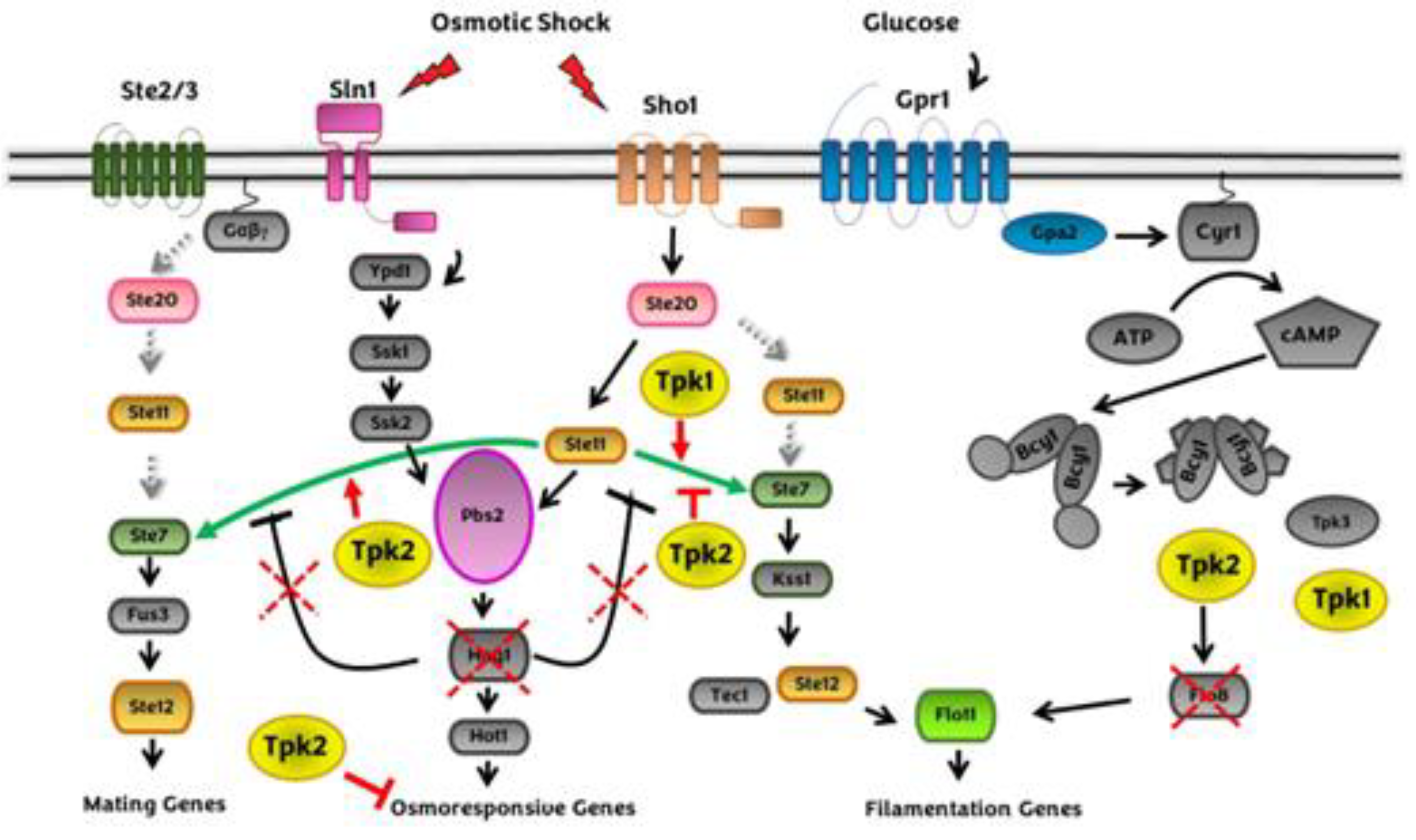
Figure 2.
Crosstalk between cAMP-PKA and CWI pathway in response to heat stress.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



