Submitted:
07 June 2023
Posted:
08 June 2023
You are already at the latest version
Abstract
Development of non-conventional hybrids (NCHs) responds to the demand for the Elotes Occidentales 'EO' race for production of pozole. The effect of: crop cycle (2019, 2020 and 2021) on physical characteristics, flowered grain quality and phytochemical content of two HNC (Pz1 and Pz2) of pozolero maize, as well as the effect of the presence or absence of pedicel, type of pollination (open and controlled, 2019), and progenitor lines (female and male, 2020) on flowered grain quality and content of phytochemical compounds were evaluated. Size, hardness, color, total phenols and anthocyanins in unprocessed grain were determined, and yield, volume and puncture strength in flowered grain. Results were analyzed with a factorial arrangement in a completely randomized design. There were significant differences (p≤0.05) in most of the variables studied by effect of crop cycle and genotype. NCHs had large grains (40 g 100 grains-1), soft endosperm (IF> 60), pink-purple color, and phenol and anthocyanin contents similar to those reported for the 'EO' race. The presence or absence of the pedicel did not affect flowered grain quality. Controlled pollination favored anthocyanin synthesis. The anthocyanin content of NCHs was determined by the female line. Thermal processing reduced anthocyanins by 60%; however, they leached into the flowering broth, so that the pozole made from NCHs improves the nutraceutical value of the pozole, relative to that of pozole made with Cacahuacintle maize.
Keywords:
Elotes Occidentales
; total anthocyanin
; total soluble phenols
; volume of flowered grain.
1. Introduction
Of the 64 maize races identified in Mexico [1], the races specialized for pozole preparation stand out. Pozole is dish that is only surpassed by ‘mole’ in volume of consumption [2]. The base of pozole is flowered grain, which is preferably prepared with Cacahuacintle, Ancho and Elotes Occidentales races, the latter characterized by its elongated cobs with a low number of rows of large grains, and floury texture, with colors ranging from pink to purple through purplish and red [3,4]. These races are produced in specific ecological niches, limiting their production. They have low yields (3.6 t ha-1) associated with problems of lodging, asynchrony between female and male flowering, grain rot, etc. [5], which have led to the development of improved varieties to overcome these disadvantages.
Such is the case of non-conventional hybrids (NCHs), which can be formed with two, three or four parental lines, at least one of which derived from native maize. The NCHs in question include a line from the Elotes Occidentales ‘EO’ race [4]. The heterosis present in these NCHs favors an increase in yield up to 12.4 t ha-1 and reduces the limitations of the original native, while retaining the early cycle and grain quality characteristics [4]. However, it is necessary to determine the effect of different years of cultivation on the quality of unprocessed grain as well as flowered grain and its phytochemical compounds.
The purple, red and blue tones in grains of the ‘EO’ race present in new hybrids are due to anthocyanins in aleurone layer [3]. Anthocyanins are pigments that have gained great relevance due to their antimicrobial, antimutagenic, anticancer and anti-inflammatory properties [6]. Currently, there is a trend towards preference for consumption of pigmented maize over white maize; thus, NCHs can be a better option in terms of nutraceutical quality than unpigmented maize. The objectives were (1) to evaluate the effect of crop cycle on physical characteristics, flowered grain quality and phytochemical content of two NCHs of pozolero maize, (2) presence or absence of pedicel on flowered grain quality and phytochemical content, and (3) type of pollination (open and controlled, 2019) and progenitor lines (female and male, 2020) on phytochemical content.
2. Results and Discussion
2.1. Physical characterization of grain
The analysis of variance (ANOVA) showed significant statistical difference (p≤0.05) in most of the variables evaluated due to the effect of crop cycles and hybrid (Table 1). The test weight of Pz2 was very stable during evaluation cycles; its value ranged from 73.1 to 73.9 kg hL-1. The hundred grains weight (HGW) was 36.4-44.5 g, values that qualify them as large grains (> 38.0 g) [7]. In both variables the highest values were found in hybrid Pz2. In the ‘EO’ race HGW values between 27 and 64 g have been reported [3,8], which is consistent with what was found in this research. The flotation index (FI) varied from 58 to 93 floating grains. In the 2019 cycle the grains had intermediate texture, while in 2020 and 2021 their endosperm had a very soft texture. The FI correlated significantly with the percentage of floury endosperm (EH) (r = 0.72; p> 0.01), corroborating the soft texture of these grains. The proportions of vitreous endosperm decreased (Table 1).
In the pedicel (PE) and pericarp (PER) fractions, crop year and hybrid did not cause significant differences, possibly indicating that these variables are less influenced by the production environment, except for year 2020 when Pz1 had a lower % of pedicel than Pz2 (Table 1). PE and PER values for Pz1 and Pz2 hybrid agree with Preciado-Ortiz et al. [4] for NCHs. The reduced amount of PE (0.85-1.32%) in NCHs, relative to that in Cacahuacintle (2.4%) [9], could be due to genetic and environmental aspects as well as smaller grain size of NCHs.
The % of germ had few changes over the evaluation years with values from 10.4 to 11.4% (Table 1), results that were similar to those reported by Vázquez-Carrillo et al. [10] for a collection of the ‘EO’ race but lower than those reported by Preciado-Ortiz et al. [4] in NCHs.
Grain color was pink-purple with luminosity values between 41.87 and 50.43%, hue from 51.4 to 69.1°, and chroma from 11.7 to 14.2, similar to those reported by Ballesteros Martínez et al. [3] for ‘EO’ race.
2.2. Flowered grain quality
The ANOVA of the technological variables showed statistical difference (p ≤ 0.05) in the flowered factor (with and without pedicel), significant differences were present in flowered grain moisture (FGM) and color variables (L*, hue and chroma), while in the genotype factor there were differences in all variables, except in dry matter loss (DML) (Table 2).
2.3. Effect of type of flowering
Flowered grains without pedicel (FGWOP) had higher moisture content (62.2 % ± 3.4) than FG with pedicel (FGWP) (61.1 % ± 2.7), suggesting that elimination of the pedicel may have facilitated the entry of water into the endosperm. However, it may also have been due to a concentration effect since with the pedicel present, there is a greater amount of dry matter, resulting in a lower moisture content. It was found that after flowering (almost three hours) the aleurone layer remained attached to the endosperm, which favors the retention of the grain components and preserves the pinkish color of this structure (Figure 1). Santiago-Ramos et al. [11] mention that the aleurone layer persists attached to the endosperm even after the alkaline and thermal conditions of nixtamalization and prevents losses of protein and starch. The color of FGWOP was lighter (48.0 ± 6.4 %) but with less tone and purity (52.1 ± 15.8 and 14.28 ± 1.5), while lightness in the color of FGWP was reduced, and tone and purity of color increased (45.1 ± 6.4; 56.6 ± 13.4 and 16.7 ± 1.6, respectively), a difference that is attributed to the dark color of the pedicel. The rest of the FG technological variables were not affected by the presence or absence of pedicel (Table 2), results that could explain the use of de-headed 'EO' maize in the region where it is consumed. On the other hand, Preciado-Ortiz (2023; pers. comm.) reported that the presence of the PE in 'EO' flowered grain does not affect the palatable characteristics of flowered grain.2. 4 Effect of genotypes
The hybrids grown in 2019 required the least time to flower (FT); the rest of the genotypes, including the Cacahuacintle control (CACC), required 160 and up to 205 minutes to flower (Table 3). Similar values were reported by Ballesteros Martínez et al. [3]. The loss of dry matter (LDM), which includes solids in nejayote and those in washing and flowering water, was 6.88 to 7.71%, not statistically different from CACC. The high alkalinity of nejayote (pH> 12) promotes pericarp solubility; thus, it was in nixtamalization where the highest LDM was recorded, as Méndez-Lagunas et al. [12] also reported. Flowered grain moisture (FGM) ranged from 57.6 to 64.1%. The NCHs in the 2020 and 2021 cycles and CACC had the highest moisture content and were statistically different from genotypes Pz2-19 and Pz1-19C (Table 3). Flowered grain yield (FGY) ranged from 2.24 to 2.55 kg of flowered grain per kg of maize. Pz1-19 and Pz1-19C, with open and controlled pollination, were the lowest yielding and statistically different from the rest of the ‘EO’ genotypes and CACC. This difference was associated with a greater hardness of their grains (FI<68). The largest volume of flowered grain (VFG) was found in the female line of ♀Pz1-20 (385.0 cm3 100 g-1 maize) and was statistically equal to hybrids Pz1-20, Pz2-21, female ♀Pz2-20 and CACC. Genotypes grown in 2019 had the lowest volumes, they were also among those that required less FT and more force to be perforated, which is attributed to the greater hardness of their grains (FI<70).
In the NCHs of 2019, which were the hardest grains, the flowered grain quality was diminished, due to a decrease in FGM, FGY and VFG; they also required the greatest puncture force (PFFG). Controlled pollination (C) did not affect the technological characteristics of Pz1 maize, while Pz2-19C, with an average FI of 99, required little FT (150 min) and less PFFG (1.59 N). In addition, it reached a good FGM (60.3%), while the rest of the variables were similar to those of its homologous Pz2-19 with open pollination,with values between 1.14 and 1.61 N (Table 3). With the female parent of Pz1-20, the numerically highest VGF was produced (385 cm3 100 g-1 maize). Similar values were reported for the Cacahuacintle race (388 cm3 100 g-1 maize) [13], which required the least PFFG (1.14 N). These characteristics are associated with starch and the formation of the amylose-lipid complex, which gives greater stability to hydrated starch granules, imparting a spongy structure, associated with a soft texture and easy chewiness [14].
The color parameters L*, hue and chroma, NCHs were statistically different from CACC. This was expected since CACC maize is white, while NCHs have pigmentation in the aleurone layer, which remained in the flowered grain (Figure 1), a result that supports the added value of NCHs over white grains. The processing of NCHs (nixtamalization and flowering) reduced the original grain color, especially of those harvested in 2019 (Figure 1).
2.5. Phytochemical components
2.5.1. Total soluble phenols (TSP) and total anthocyanins (TA) in whole grain
In the 2019 cycle, Pz1 had the highest TSP content (187.1 mg FAE 100 g-1 ds), higher than Pz2 (145.3 mg FAE 100 g-1 ds). In TA the behavior was reversed, Pz2 had the highest content (12.9 mg PCE 100 g-1 ds) (Figure 2B). With controlled pollination, TSP was similar in both hybrids (Figure 2A). When comparing pollination types (cycle 2019), the TSP content in hybrid Pz1-19C (controlled pollination) decreased by 16.43%, while in Pz2 it remained at 145 mg FAE 100 g-1 ds. TA in hybrids Pz1-19C and Pz2-19C, produced with controlled pollination, increased by 12.2 and 25.4%, respectively. The effect of pollination type on anthocyanin content has not been evaluated; however, Kahriman et al. [15] found that carotenoid content increases significantly when produced with controlled pollination relative to that with open pollination.
In the 2020 cycle, the highest TSP contents were found in the male parents of both hybrids (215.3 and 180.32 mg FAE 100 g-1 ds), while the lowest values were found in the female parents (134.8 and 160.2 mg FAE 100 g-1 ds), and intermediate contents (145.0 and 174.5 mg FAE 100 g-1 ds) in the hybrids (Figure 2A). The results obtained in the hybrids could have been influenced by both parents since Darrah et al. [16] mention that the pericarp has a maternal origin, while the germ has both female and male influence. Studies have determined that the highest concentration of TSP is found in pericarp and germ [17]. In contrast, Ruiz-Torres et al. [18] found that the expression of phenolic compounds in white maize crosses is mainly determined by the genetic effects of the female lines. In TA content, the highest concentration was found in the female line of ♀Pz2 (17.48 mg PCE 100 g-1 ds); in the Pz2 hybrid it decreased to 15.9 mg PCE 100 g-1 ds, behavior that was reversed in the Pz1 hybrid, in which heterobeltiosis was observed with respect to this variable, since it registered the highest TA content (17.04 mg PCE 100 g-1 ds). In both hybrids their respective male parents had the lowest values (Figure 2B). The results found could be due to the fact that pigments were present only in the aleurone layer, and this structure has triploid cells (3n) in which one paternal and two maternal genes participate [19]; so, as previously reported, anthocyanin content in hybrids is determined by the line that participates as female in the cross [4,18].
Regarding the behavior of hybrids in the evaluated cycles, in TSP content, the hybrid Pz2 was more stable, with contents of 145.3, 174.3 and 174.2 mg FAE 100 g-1 ds for the 2019, 2020 and 2021 cycles, respectively. In hybrid Pz1, TSP contents were 187.01, 145.0 and 198.0 mg FAE 100 g-1 ds for the same years (Figure 2A). Little has been reported on TSP content in ‘EO’ race samples. In this regard, Ballesteros Martínez et al. [3] reported values lower than those found in this work, ranging from 100 to 130 mg FAE g-1 ds, while Rocandio-Rodríguez et al. [21] reported 344 mg GAE 100 g-1 ds in a cross of Elotes Occidentales x Tuxpeño. In TA content, a trend of interaction with the environment was observed across years, with values of 10.5, 17.0 and 15.2 mg PCE 100 g-1 ds for Pz1 in cycles 2019, 2020 and 2021, respectively, and 12.9, 15.9 and 18.1 mg PCE 100 g-1 ds for Pz2 in the same years (Figure 2 B). In the 2019 and 2021 cycles, the Pz2 hybrid was superior to Pz1. The values found were similar to those reported by Broa Rojas et al. [22] in some EO crosses, and Preciado-Ortiz et al. [4] in NCHs of the EO race. The behavior of both hybrids in TSP and TA content in the evaluation cycles indicated good adaptability to the production area for which they were developed; in both cases they have shown upward behavior, which could indicate that better climatic conditions favored the synthesis of these phytochemicals. This contrasts with Mansilla et al. [23], who observed a decrease in phenol and anthocyanin contents in purple corn over four growing cycles, possibly due to poor adaptation of the seed to the growing area, as they used a mixture of introduced seeds from several countries.2.5.2 TSP and TA content in flowered grain, nejayote, washing water and flowering broth.
A statistically significant difference (p≤0.05) was found between flowered grain with and without pedicel in all evaluated variables. The highest concentration of TSP was for flowered grains with pedicel (Figure 3A). The TSP content in FG ranged from 64.96 to 116.99 mg FAE 100 g-1 dm.
This behavior can be explained since phenolic acids derived from hydroxycinnamic acid are bound to lignocellulosic material [24]. Little information is available on phenolic content in flowered grain. In this regard, Peralta-Veran et al. [25] found contents of 50-71 mg GAE 100 g-1 ds, which are lower than those found in present study. However, Ballesteros Martínez et al. [3] found higher contents in flowered grain of maize populations of the Elotes Occidentales race. The phenolic content of unprocessed grain decreased in flowered grain from 16.2 to 49%, with the genotypes that had the pedicel in the flowered grain suffering the losses due to temperature and prolonged flowering times. On average, in flowered grains without pedicel (FGWOP) there was a decrease of 48%, while in FGWP the reduction was 36.9%. In both types of flowered grain, TSP losses were lower than those reported by Peralta-Veran et al. [25] who reported losses of 70%. It is interesting to mention that the genotypes with the lowest TSP content in grain (Pz1-20 and ♀Pz1-20) were those that retained the most of these compounds in flowered grain either with or without pedicel, Peralta-Veran et al. [25] observed similar behavior.
Several studies mention that the highest content of phenolic compounds is found in the pericarp followed by the germ, while a minimal amount is found in the endosperm [17,26]; however, they have also been reported in the cell wall of aleurone cells [27]. In the flowered grain, the pericarp was no longer present, so it is probably the TSP of the aleurone layer and the germ that is being quantified, as well as those in the pedicel when present.
The total anthocyanin (TA) content in flowered grain ranged from 3.84 to 9.23 mg PCE 100 g-1 ds, with no statistical difference (p≤0.05) between the grains with or without pedicel (Figure 3). The differences between one type of grain or another may be due to a dilution effect, since the presence of the pedicel translates into a higher proportion of dry matter. The same pattern in raw grain was also observed. The 2019 hybrids had lower concentrations than the 2020 and 2021 hybrids, possibly due harder grains (IF of 58 and 68%) since it has been reported that anthocyanin content decreases in hybrid maize due to an increase in hardness compared to native maize, as TA content is related to endosperm softness [28]. During processing from raw grain to flowered grain, TA losses ranged from 47.8 to 68%. Once again, it was evident that genotypes with higher TA content had greater losses than those with lower TA contents. TA contents were slightly higher than those found by Ballesteros Martínez et al. [3], who reported values of 4 to 6.37 mg EPC 100 g-1 ds in flowered grain, with average losses of 71%.
The correlation between FI and TA showed that softer grains had higher anthocyanin content (Table 5). This suggests that the aleurone layer adheres more strongly to the vitreous endosperm, which could favor its retention, in contrast to what was observed in grains with floury endosperm, where these phytochemicals were lost in smaller amounts (Figure 2 and Figure 3). However, more studies are needed in this regard. TA in FGWP and FGWOP showed a high negative correlation with hue and L of flowered grain with and without pedicel (Table 5), which demonstrates that they are responsible for grain coloration since the higher the TA values, the lower the grain lightness, and hue values will be closer to zero, where the red is located in the L*, C, h color space. In unprocessed grain, no negative correlation of TA with L and hue was observed, but this is possibly because the reading was on the pericarp layer, which interferes with the expression of the color of the aleurone layer.
Complete elimination of the pericarp and subsequent flowering significantly reduced phenolic compounds in flowered grain (Figure 2A). The alkaline conditions and nixtamalization temperature favor the leaching of phenolic compounds into the nejayote (TSPn), where the highest concentration of TSPn was found, with values from 297.2 to 324.9 mg FAE L-1, while in the washing water (TSPl) values were from 179.8 to 230.0 mg FAE L-1 (Table 4). These results were lower than those reported by other authors [12,29], possibly due to the type of maize and low TSP content in raw grain. In broth of FG without pedicel, (TSPc) phenol contents were between 156.6-209.3 mg FAE L-1. It was observed that in broth of FG with pedicel there was a higher concentration of these metabolites, 197.6-263.9 mg FAE L-1, which could be due to the fact that phenols present in the lignocellulosic material of the pedicel were leached into the flowering broth, since the samples with the highest TSP content were those that had the highest proportion of pedicel. Flowering time was not modified by the presence or absence of the pedicel (Figure 2, Table 1).
The anthocyanin content in flowering broth ranged from 4.25 to 12.76 mg PCE L-1. The effect of flowering time was evident, since hybrids that required more FT (2020 and 2021) and presented the greatest leaching of anthocyanins from grain to broth (Table 4). It was also observed that FGWOP leached more anthocyanins into the flowering broth than their counterparts with pedicel. However, this could also be due to flowering time since FG without pedicel required more time than FG with pedicel (Table 4). In flowered grains, endosperm flowering explosion occurs at the apical or lateral part. Phytochemical compounds that are leached into flowered broth are not considered losses since in the preparation of the dish called pozole, the flowering broth is consumed as well as flowered grains. Therefore, making pozole with pigmented grains could be an option for consuming foods with nutraceutical value since it has been reported that maize anthocyanins have anticancer activity, cardioprotective effect, antimicrobial, antimutagenic, antidiabetic, to mention a few [6,30].
3. Materials and Methods
3.1. Genotypes.
The non-conventional maize hybrids (NCH) Pz1 and Pz2, which include the ‘EO’ race in their genealogy were studied. In the 2019, 2020 and 2021 crop cycles at the Campo Experimental Bajío, Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), NCHs with open pollination were evaluated. In 2019, NCHs with controlled pollination were also evaluated, and in the 2020 cycle the male and female parent lines of the two hybrids were included. As a control to evaluate the quality of flowered grain, a commercial Cacahuacintle (CACC) white grain maize was used, as it is that most commonly used for this purpose in central Mexico [2].
3.2. Physical characteristics of grain
The variables test weight (TW), the hundred grains weight (HGW) and grain hardness measured with the flotation index (FI) and the percentages of pedicel, pericarp, germ, floury and vitreous endosperm were determined by manual dissection according to the standard NMX-034/1 [7]. Color in unprocessed and flowered grain was measured with a Hunter Lab MiniScanXEPlus® colorimeter (Reston, VA, USA) from which L*, a* and b* parameters were obtained, and hue angle (and saturation index ( were determined [31].
3.3. Nixtamalization and obtaining flowered grain
A 100 g sample of maize free of pests and damage was nixtamalized with 250 mL of water and 1% Ca(OH)2 with respect to the amount of grain. The nixtamalization time was assigned according to FI, following NMX-034/1 [7]. After cooking, it was left to rest for 16 h. After this time, it was strained to remove the nejayote and the nixtamal was washed to remove pericarp. The pedicel was manually removed (de-headed). The washed and de-headed nixtamal was subjected to a second cooking to obtain flowered grain, for which 250 mL of water was added. Flowering time was measured from when the samples started to boil until 60% of a random sample of 10 grains had flowered. The grains were separated from the cooking liquor and the volume of flowered grain (VGF) was measured; the result is reported in cm3 100 g-1 maize. A 50 mL sample was taken of nejayote (maize cooking liquor), washing water, and flowered broth and evaporated in an oven at 130 °C. The percentage of dry matter loss (DML) was obtained by adding the solids from the three liquids. The technological variables of flowered grain were studied in grain with and without pedicel.
3.4. Quality of flowered grain
In flowered grain (FG) was quantified the following: yield (YGF) expressed in kg FG per kg maize, moisture (FGM) following method 44-15.02 of the American Association of Cereal Chemistry [32], and puncture force (PF) obtained on a Brook-field® texturometer model CT3 (Middleboro, MA, USA), using a needle-shaped strut. Results are reported in Newtons.
3.5. Phytochemical components
The content of total soluble phenols (TSP) and total anthocyanins (TA) in raw maize, flowered grain, nejayote, washing water and flowered broth were analyzed.
3.5.1. Obtaining the extract.
In the case of solid samples, 100 mg of flour previously defatted with petroleum ether for 8 h in Goldfisch Labconco® (Kansas, Missouri, USA) equipment was weighed. In the case of nejayote, washing water and flowering broth, 1 mL of a homogenized sample was taken and placed in a 2 mL capacity Eppendorf® tube. In both cases it was gauged to 2 mL with 1% TFA prepared in 80% methanol. The samples were sonicated for 15 min in a Branson® sonicator bath (model 2510-MTH, Danbury, USA), followed by rest for 105 min in refrigeration. Subsequently, the samples were centrifuged at 10000 rpm for 10 min in a centrifuge (Hermle model Z 200 M/H, Germany), decanted, and finally gauged to 2 mL with the same solvent.
3.5.2. Total soluble phenol content.
Phenols were quantified with the Folin-Ciocalteu reagent [33] following the method adapted to microplates, onto which 25 µL of extract was placed, and 125 µL of distilled water was added. Subsequently, 20 µL of Folin-Ciocalteu reagent 0.2 N and 30 µL of 20% sodium carbonate (Na2CO3) were added. The mixture was left to rest for 30 min in the dark, and absorbance was then measured at 760 nm in a plate reader (Epoch, Biotek). A standard curve of ferulic acid (R2=0.995), the most abundant phenolic compound in maize [34], was prepared, and the results were expressed as mg of ferulic acid equivalents (FAE) per 100 g of dry sample (ds).
3.5.3. Total anthocyanin content.
Anthocyanins were quantified by spectrophotometry, and absorbance was read at 520 nm in an Epoch (Biotek) plate reader. A standard curve of pelargonidin chloride (R2=0.997) (Sigma Aldrich) was prepared since it has been seen that, due to the color of the analyzed genotypes, this compound predominates [35]. A scan was also performed to identify the wavelength of maximum absorbance. The results were expressed in mg of pelargonidin chloride equivalents (PCE) per 100 g of dry sample.
3.6. Statistical analysis
The physical characterization variables of the grains were analyzed in a completely randomized design with two factors of variation: genotype and years of cultivation. In the case of flowered grain quality variables, the analysis was done with a 2x11 factorial design, where one factor was the type of flowered grain (with or without pedicel) and the other factor the eleven genotypes evaluated. An analysis of variance followed by comparison of means with the Tukey test (p≤0.05) was performed in the case of grain characteristics and phytochemical content. For pozole quality variables, we compared means with the Dunnett statistic (p≤0.05), using the Cacahuacintle race as a control since it has been the most studied to determine pozole quality [36]. Additionally, we explored correlations between FI and the variables of color and total anthocyanin with the Pearson correlation coefficient (p≤0.05). We duplicated all measurements.
4. Conclusions
Grain hardness and size of the hybrids Pz1 and Pz2 were modified by effect of crop cycles, while the pink-purple color remained. In the 2019 cycle, controlled pollination increased total anthocyanin content relative to that of hybrids with open pollination, but not the total content of soluble phenols. In the 2020 and 2021 cycles, the total anthocyanin content was higher than that of hybrids produced with controlled pollination.
In the 2020 cycle where hybrids were studied with their two parents, anthocyanin content was determined by the line that participated as female in the cross.
The quality of flowered grain of hybrids was not affected by the presence or absence of the pedicel and was similar to the quality of flowered Cacahuacintle maize grain. However, the presence of pedicel favored a higher phenolic content in flowered grain and flowering broth. In flowered grain, the presence of the aleurone layer was confirmed, which contributes significant amounts of phytochemical compounds to pozole. Hybrids Pz1 and Pz2 have a higher nutraceutical value than white maize.
Author Contributions
R. E. P. O. developed the hybrids and generated the experiments in the field; L. G. C. and M. G. V. C processed the samples in the laboratory. All authors interpreted the data, and approved the final version. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by GRUMA througth CONALSA, grant number 1053635249 and INIFAP.
Data Availability
The data presented in this study are available within the article.
Acknowledgments
We would like to thank the technicians of the maize Laboratory of INIFAP-CEVAMEX, for their support and dedication, and CONALSA and INIFAP for funding the project.
Conflicts of Interest
The authors declare no conflict of interest.
References
- ReferencesCONABIO (Comisión Nacional para el Conocimiento y Uso de la Biodiversidad). Proyecto global de maíces nativos. https://biodiversidad.gob.mx/diversidad/proyectoMaices (accessed 2023-04-23).
- Vázquez-Carrillo, M. G.; Santiago-Ramos, D.; Palacios-Rojas, N. Calidad Industrial y Nutricional de Razas Mexicanas de Maíz Pozolero, 1st ed.; Biblioteca Básica de Agricultura: México, 2016. [Google Scholar]
- Ballesteros Martínez, G.; Zarazúa Villaseñor, P.; Salinas Moreno, Y.; De la Cruz Larios, L. Fijación del color en grano y características físicas, tecnológicas y nutracéuticas en maíz Elotes Occidentales. Rev. Mex. Ciencias Agrícolas 2019, 10 (3), 585–599. [Google Scholar] [CrossRef]
- Preciado-Ortiz, R. E.; Ochoa-Centeno, N. J.; Vázquez-Carrillo, M. G.; Santiago-Ramos, D.; Terrón-Ibarra, A. D. Grain yield, physical and pasting properties, and anthocyanins of non-conventional pigmented corn hybrids for pozole end-use adapted to subtropical regions. Appl. Food Res. 2022, 2, 100180. [Google Scholar] [CrossRef]
- Gómez-Montiel, N. O.; Cantú-Almaguer, M. Á.; Hernández-Galeno, C. del Á.; Vázquez Carrillo, M. G.; Aragón Cuevas, F.; Palemón Alberto, F. V-237 AN, Cultivar mejorado de maíz “Ancho Pozolero” para la región semicálida de Guerrero. Rev. Mex. Ciencias Agrícolas 2014, 5 (7), 1315–1319. [Google Scholar] [CrossRef]
- Kim, H. Y.; Lee, K. Y.; Kim, M.; Hong, M.; Deepa, P.; Kim, S. A review of the biological properties of purple corn ( Zea Mays L.). Sci. Pharm. 2023, 91 (1), 6. [Google Scholar] [CrossRef]
- Secretaría de Economía, S. Productos Alimenticios Para Uso Humano No Industrializados-Cereales-Maíz-Parte1: Granos Para Tortillas y Productos Nixtamalizados-Especificaciones y Métodos de Prueba, NMX-FF-034/1-SCFI-2020; Dirección General de Normas; Ciudad de México, México. 2020.
- Salinas-Moreno, Y.; Aragón-Cuevas, F.; Ybarra-Moncada, C.; Aguilar-Villarreal, J.; Altunar-López, B.; Sosa-Montes, E. Caracterización física y composición química de razas de maíz de grano azul/morado de las regiones tropicales y subtropicales de Oaxaca. Rev. Fitotec. Mex. 2013, 36 (1), 23–31. [Google Scholar] [CrossRef]
- Vázquez-Carrillo, M. G.; Santiago-Ramos, D.; Domínguez-Rendón, E.; Audelo-Benites, M. A. Effects of Two different pozole preparation processes on quality variables and pasting properties of processed maize grain. J. Food Qual. 2017, 8627363. [Google Scholar] [CrossRef]
- Vázquez-Carrillo, M. G.; Pérez-Camarrilo, J. P.; Hernández-Casillas, J. M.; Marrufo-Díaz, M. de la L.; Martínez-Ruiz, E. Calidad de grano y tortillas de maíces criollos del Altiplano y Valle del Mezquital, México. Rev. Fitotec. Mex. 2010, 33 (4), 49–56. [Google Scholar]
- Santiago-ramos, D.; Figueroa-Cárdenas, J. D. D.; Mariscal-Moreno, R. M.; Escalante-aburto, A.; Ponce-García, N.; Véles-Medina, J. J. Physical and chemical changes undergone by pericarp and endosperm during corn nixtamalization-A review. J. Cereal Sci. 2018, 81, 108–117. [Google Scholar] [CrossRef]
- Méndez-Lagunas, L. L.; Cruz-Gracida, M.; Barriada-Bernal, L. G.; Rodríguez-Méndez, L. I. Profile of phenolic acids, antioxidant activity and total phenolic compounds during blue corn tortilla processing and its bioaccessibility. J. Food Sci. Technol. 2020, 57 (12), 4688–4696. [Google Scholar] [CrossRef] [PubMed]
- Flores-Hernández, L. A.; Castillo-González, F.; Vázquez-Carrillo, M. G.; Livera-Muñoz, M.; Benítez-Riquelme, I.; Nieto-Sotelo, J.; Ramírez-Hernández, A. Composition and flowering quality of Cacahuacintle maize populations from the High Valleys of Mexico. Plant Foods Hum. Nutr. 2023. [Google Scholar] [CrossRef] [PubMed]
- Dorantes-Campuzano, M. F.; Cabrera-Ramírez, A. H.; Rodríguez-García, M. E.; Palacios-Rojas, N.; Preciado-Ortíz, R. E.; Luzardo-Ocampo, I.; Gaytán Martínez, M. Effect of maize processing on amylose-lipid complex in pozole, a traditional mexican dish. Appl. Food Res. 2022, 2 (1). [Google Scholar] [CrossRef]
- Kahrıman, F.; Egesel, C. Ö.; Aydın, T.; Subaşı, S. The Role of artificial pollination and pollen effect on ear development and kernel structure of different maize genotypes. J. Pollinat. Ecol. 2015, 15 (2), 6–14. [Google Scholar] [CrossRef]
- Darrah, L. L.; McMullen, M. D.; Zuber, M. S. Breeding, genetics and seed corn production. In Corn: Chemistry and Technology; Serna-Saldívar, S. O., Ed.; Elsevier Inc., 2018; pp 19–41. [CrossRef]
- Das, A. K.; Singh, V. Antioxidative free and bound phenolic constituents in botanical fractions of indian specialty maize (Zea Mays L.) genotypes. Food Chem. 2016, 201, 298–306. [Google Scholar] [CrossRef]
- Ruiz-Torres, N. A.; Rincón-Sánchez, F.; Hernández-López, V. M.; Figueroa-Cárdenas, J. D. D.; Loarca-Piña, M. G. F. Determinación de compuestos fenólicos y su actividad antioxidante en granos de maíz. Rev. Fitotec. Mex. 2008, 31 (3), 29–34. [Google Scholar] [CrossRef]
- Paulsmeyer, M. N.; Juvik, J. A. Increasing aleurone layer number and pericarp yield for elevated nutrient content in maize. G3/Genes/Genomics/Genetics 2023, 00. [Google Scholar] [CrossRef]
- Espinosa Trujillo, E.; Mendoza Castillo, M. D. C.; Castillo González, F.; Ortiz Cereceres, J.; Delgado Alvarado, A.; Carrillo Salazar, A. Anthocyanin accumulation in pericarp and aleurone layer of maize kernel and their genetic effects on native pigmented varieties. Rev. Fitotec. Mex. 2009, 32 (4), 303–309. [Google Scholar]
- Rocandio-Rodríguez, M.; Torres-Castillo, J. A.; Juárez-Aragón, M. C.; Chacón-Hernández, J. C.; Moreno-Ramírez, Y. D. R.; Mora-Ravelo, S. G.; Delgado-Martínez, R.; Hernández-Juárez, A.; Heinz-Castro, R. T. Q.; Reyes-Zepeda, F. Evaluation of resistance of eleven maize races (Zea Mays L.) to the red spider mite (Tetranychus Merganser, Boudreaux). Plants 2022, 11 (11). [Google Scholar] [CrossRef] [PubMed]
- Broa Rojas, E.; Vázquez Carrillo, M. G.; Estrella Chulím, N. G.; Hernández Salgado, J. H.; Ramírez Valverde, B.; Bahena Delgado, G. Características fisicoquímicas y calidad de la proteína de maíces nativos pigmentados de Morelos en dos años de cultivo. Rev. Mex. Ciencias Agrícolas 2019, 10 (3), 683–697. [Google Scholar] [CrossRef]
- Mansilla, P. S.; Bongianino, N. F.; Nazar, M. C.; Pérez, G. T. Agronomic and chemical description of open-pollinated varieties of opaque-2 and purple maize (Zea Mays L.) adapted to semiarid region of Argentina. Genet. Resour. Crop Evol. 2021, 68 (6), 2351–2366. [Google Scholar] [CrossRef]
- Liu, T.; Yang, L.; Liu, B.; Tan, L. Hydroxycinnamic acids release during bioconversion of corn stover and their effects on lignocellulolytic enzymes. Bioresour. Technol. 2019, 294, 122116. [Google Scholar] [CrossRef]
- Peralta-Veran, L.; Espinosa-Leal, C.; Escalante-Aburto, A.; Preciado-Ortiz, R. E.; Puente-Garza, C. A.; Serna-Saldivar, S. O.; García-Lara, S. Effects of pozole broth production on phenolic acids and antioxidant activity of specialty maize landraces. J. Cereal Sci. 2022, 107 (June), 1–8. [Google Scholar] [CrossRef]
- Das, A. K.; Singh, V. Antioxidative free and bound phenolic constituents in pericarp, germ and endosperm of indian dent (Zea Mays var. indentata) and flint (Zea Mays var. indurata) maize. J. Funct. Foods 2015, 13, 363–374. [Google Scholar] [CrossRef]
- Razgonova, M.; Zinchenko, Y.; Pikula, K.; Tekutyeva, L.; Son, O.; Zakharenko, A.; Kalenik, T.; Golokhvast, K. Spatial distribution of polyphenolic compounds in corn grains (Zea Mays L. var. pioneer) studied by laser confocal microscopy and high-Resolution mass spectrometry. Plants 2022, 11, 630. [Google Scholar] [CrossRef]
- Salinas-Moreno, Y.; Soria-Ruiz, J.; Espinosa Trujillo, E. Aprovechamiento y Distribución de Maíz Azul En El Estado de México, 1ra ed.; Impresos Lebam: Texcoco, Edo. México. México, 2010. [Google Scholar]
- Gutiérrez-Uribe, J. A.; Rojas-García, C.; García-Lara, S.; Serna-Saldivar, S. O. Phytochemical analysis of wastewater (nejayote) obtained after lime-cooking of different types of maize kernels processed into masa for tortillas. J. Cereal Sci. 2010, 52 (3), 410–416. [Google Scholar] [CrossRef]
- Petroni, K.; Pilu, R.; Tonelli, C. Anthocyanins in corn: a wealth of genes for human health. Planta 2014, 240 (5), 901–911. [Google Scholar] [CrossRef] [PubMed]
- Sant’Anna, V.; Gurak, P. D.; Ferreira Marczak, L. D.; Tessaro, I. C. Tracking bioactive compounds with colour changes in foods – a review. Dye. Pigment. 2013, 98 (3), 601–608. [Google Scholar] [CrossRef]
- AACC Approved Methods of Analysis. Method 44-15.02. Moisture-Air Oven Methods, 11th ed.; Cereals & Grains Association: St. Paul, MN, USA, 2020. [Google Scholar]
- Singleton, V. L.; Rossi, J. A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16 (3), 144–158. [Google Scholar] [CrossRef]
- López-Vásquez, V. M.; Salinas-Moreno, Y.; Alemán-de la Torre, I.; Morales-Hernández, N.; Bautista-Ramírez, E. Effect of adding anthocyanins to blue maize dough on color, texture and antioxidant capacity of maize tortillas. Ing. Agrícola y Biosist. 2020, 12 (2), 183–200. [Google Scholar] [CrossRef]
- Peniche-Pavía, H. A.; Tiessen, A. Anthocyanin profiling of maize grains using DIESI-MSQD reveals that cyanidin-based derivatives predominate in purple corn, whereas pelargonidin-based molecules occur in red-pink varieties from Mexico. J. Agric. Food Chem. 2020, 68 (21), 5980–5994. [Google Scholar] [CrossRef]
- Hernández Galeno, C. del Á.; Salinas Moreno, Y.; Antonio López, P.; Santacruz Varela, A.; Castillo González, F.; Corona Torres, T. Calidad pozolera en poblaciones de maíz Cacahuacintle de Los Valles Altos de Puebla, México. Rev. Mex. Ciencias Agrícolas 2014, 5 (4), 703–716. [Google Scholar] [CrossRef]
Figure 1.
Appearance of raw grain, nixtamal, flowered grain without pedicel (FGWOP) and flowered grain with pedicel (FGWP) of two maize hybrids of the Elotes occidentales race evaluated in three crop cycles, and a genotype of the Cacahuacintle race.
Figure 1.
Appearance of raw grain, nixtamal, flowered grain without pedicel (FGWOP) and flowered grain with pedicel (FGWP) of two maize hybrids of the Elotes occidentales race evaluated in three crop cycles, and a genotype of the Cacahuacintle race.

Figure 2.
Total soluble phenols (A) and total anthocyanins (B) contend of two hybrids of the Elotes Occidentales race and their parents, developed for the Mexican subtropics.
Figure 2.
Total soluble phenols (A) and total anthocyanins (B) contend of two hybrids of the Elotes Occidentales race and their parents, developed for the Mexican subtropics.
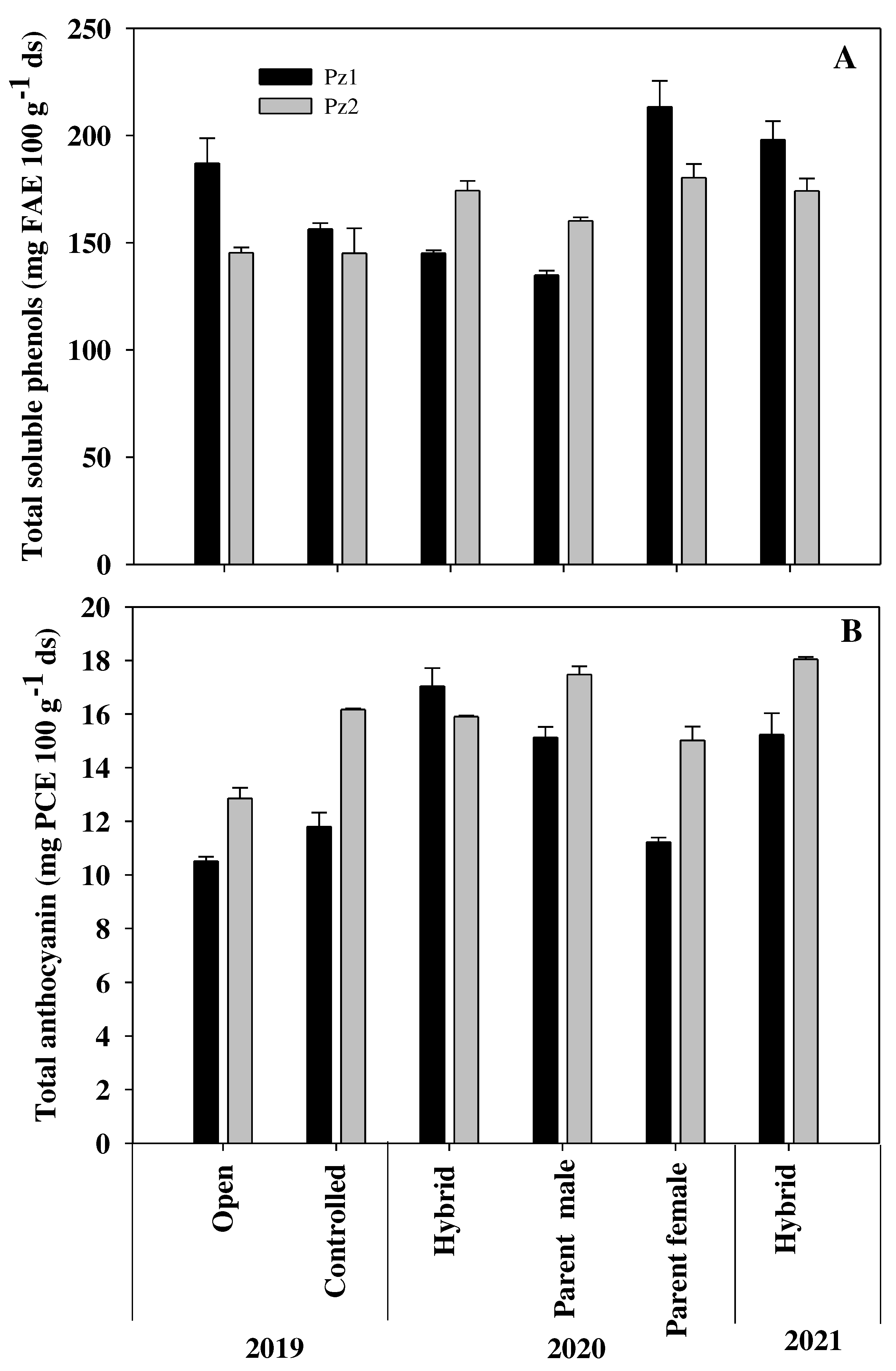
Figure 3.
Contents of total soluble phenols (A) and total anthocyanins (B) in flowered grain without pedicel (FGWOP) and with pedicel (FGWP) of HNC of 'EO' maize with open and controlled pollination (2019) and its female parents (2020), evaluated in the 2019, 2020 and 2021 crop cycles in Celaya, Guanajuato, Mexico.
Figure 3.
Contents of total soluble phenols (A) and total anthocyanins (B) in flowered grain without pedicel (FGWOP) and with pedicel (FGWP) of HNC of 'EO' maize with open and controlled pollination (2019) and its female parents (2020), evaluated in the 2019, 2020 and 2021 crop cycles in Celaya, Guanajuato, Mexico.
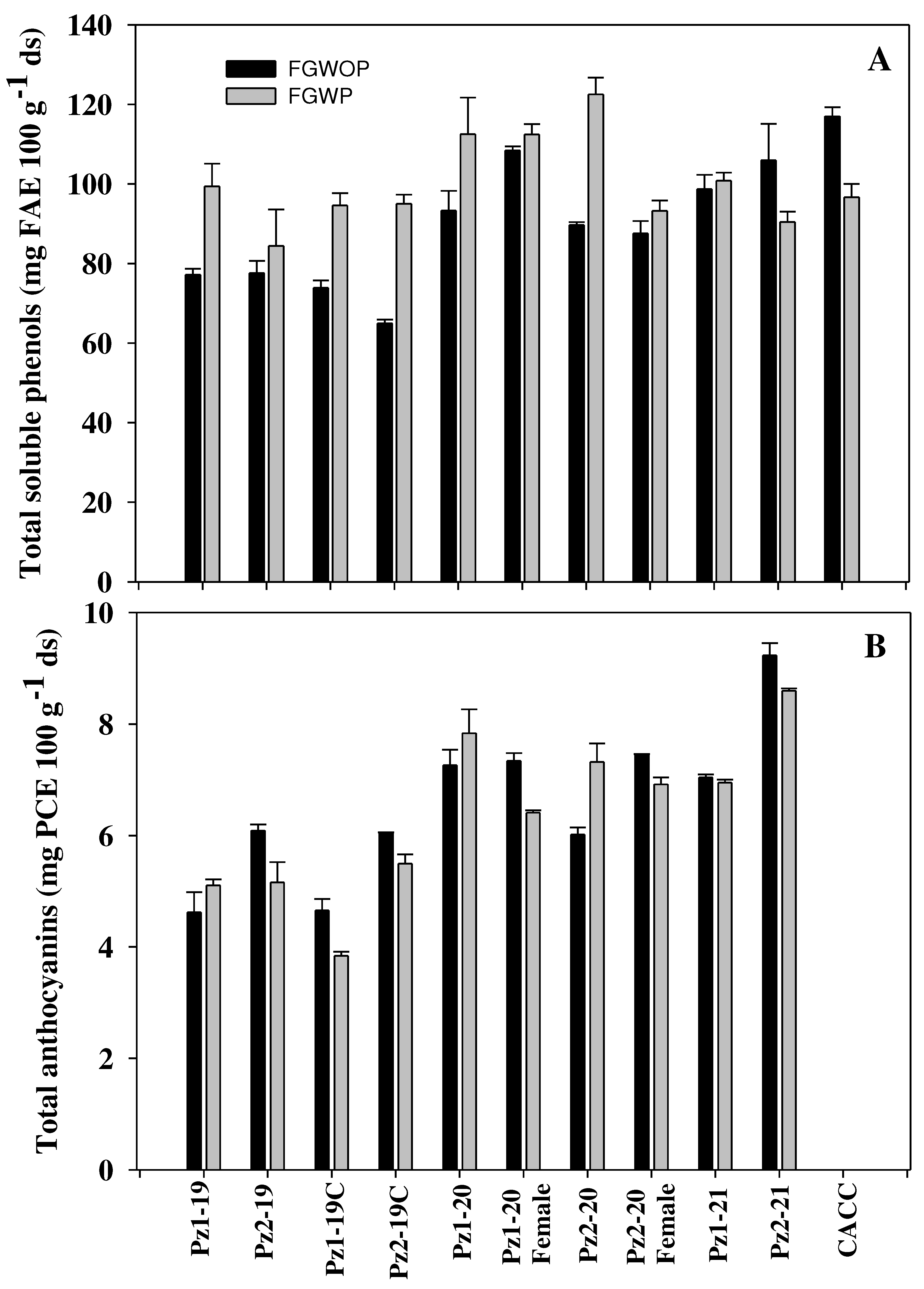

Table 1.
Grain physical characteristics of pozolero maize NCHs evaluated in the 2019, 2020 and 2021 crop cycles in Celaya, Guanajuato, Mexico.
Table 1.
Grain physical characteristics of pozolero maize NCHs evaluated in the 2019, 2020 and 2021 crop cycles in Celaya, Guanajuato, Mexico.
| Genotype | Year | |||
|---|---|---|---|---|
| 2019 | 2020 | 2021 | HSD | |
| Test weight (kg hL-1) | ||||
| Pz1 | 74.75 ± 0.7 Aa | 71.65 ± 0.35 Ba | 68.10 ±7.35 Aa | 17.763 |
| Pz2 | 73.05 ±0.07 Bb | 73.25 ± 0.35 Aab | 73.95 ± 0.07 Aa | 0.886 |
| HSD | 0.304 | 1.521 | 22.375 | |
| Hundred grain weight (g) | ||||
| Pz1 | 41.92 ± 0.21 Ba | 41.04 ± 0.01 Ab | 36.38 ± 0.01 Bc | 0.512 |
| Pz2 | 44.51 ± 0.30 Aa | 38.59 ± 0.02 Bc | 43.24 ± 0.01 Ab | 0.736 |
| HSD | 1.13 | 0.068 | 0.0481 | |
| Flotation index(%) | ||||
| Pz1 | 58.0 ± 1.4 Bb | 88.5 ± 0.7 Aa | 93.0 ± 4.2 Aa | 10.923 |
| Pz2 | 68.0 ± 1.4 Aa | 79.0 ± 7.1 Aa | 82.0 ± 2.8 Aa | 18.688 |
| HSD | 6.085 | 21.62 | 15.513 | |
| Pedicel (%) | ||||
| Pz1 | 1.32 ± 0.05 Aa | 0.85 ± 0.05 Ba | 1.37 ± 0.29 Aa | 0.719 |
| Pz2 | 1.23 ± 0.67 Aa | 1.23 ± 0.04 Aa | 1.19 ± 0.002 Aa | 0.199 |
| HSD | 0.263 | 0.198 | 0.882 | |
| Pericarp (%) | ||||
| Pz1 | 4.3 ± 0.06 Aa | 4.08 ± 0.13 Aa | 4.51 ± 0.13 Aa | 0.467 |
| Pz2 | 4.0 ± 0.13 Aa | 4.51 ± 0.23 Aa | 4.12 0.21 Aa | 0.828 |
| HSD | 0.444 | 0.838 | 0.734 | |
| Germ (%) | ||||
| Pz1 | 11.35 ± 0.04 Ba | 11.49 ± 0.23 Aa | 10.39 ± 0.01 Ab | 0.572 |
| Pz2 | 11.77 ± 0.07 Aa | 10.99 ± 0.49 Aa | 11.13 ± 0.69 Aa | 2.072 |
| HSD | 0.251 | 1.684 | 2.108 | |
| Floury endosperm (%) | ||||
| Pz1 | 42.5 ± 0.31 Ab | 62.26 ± 5.43 Aa | 70.67 ± 0.74 Aa | 13.245 |
| Pz2 | 44.13 ± 0.86 Ab | 53.11 ± 5.71 Aab | 64.78 ± 0.57 Ba | 13.992 |
| HSD | 2.79 | 23.967 | 2.853 | |
| Vitreous endosperm (%) | ||||
| Pz1 | 40.53 Aa | 16.34 Ab | 13.07 Bb | 13.559 |
| Pz2 | 38.88 Aa | 30.16 Aab | 18.78 Ab | 13.776 |
| HSD | 3.732 | 23.412 | 5.668 | |
| Lightness (%) | ||||
| Pz1 | 48.90 ± 0.04 Aa | 41.87 ± 0.28 Bb | 48.18 ± 0.82 Aa | 2.095 |
| Pz2 | 46.64 ± 0.35 Bb | 46.67 ± 0.16 Ab | 50.43 ± 0.74 Aa | 1.997 |
| HSD | 1.059 | 0.982 | 3.352 | |
| Hue (°) | ||||
| Pz1 | 51.41 ± 0.63 Bc | 61.16 ± 0.64 Ab | 65.0 ± 1.27 Aa | 3.753 |
| Pz2 | 61.21 ± 0.92 Ab | 55.59 ± 1.97 Ab | 69.1 ± 2.12 Aa | 7.327 |
| HSD | 3.372 | 6.306 | 7.527 | |
| Chroma | ||||
| Pz1 | 11.76 ± 0.19 Bb | 12.08 ± 10.43 Ab | 14.20 ± 0.28 Aa | 1.346 |
| Pz2 | 14.21 ± 0.51 Aa | 13.33 ± 0.41 Aa | 13.45 ± 0.35 Aa | 1.793 |
| HSD | 1.662 | 1.826 | 1.378 | |
* HSD honest significant difference, capital letters indicate significant statistical difference between hybrids, lower case letters indicate significant statistical differences between crop years according to Tukey test (p≤0.05).
Table 2.
Mean squares and significance in nixtamalization and flowered variables of two NCHs of pozolero maize evaluated in 2019, 2020 and 2021 in Celaya, Gto. Mexico.
Table 2.
Mean squares and significance in nixtamalization and flowered variables of two NCHs of pozolero maize evaluated in 2019, 2020 and 2021 in Celaya, Gto. Mexico.
| Mean squares of technological variables | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Variation source |
gl | RGF | VGF | PMS | TF | HGF | Dureza | L | Hue | Croma |
| Genotype | 10 | 0.044* | 1607.95* | 0.29 | 2042.49* | 25.46* | 0.35* | 158.17* | 838.74* | 4.41* |
| Flowered | 1 | 0.014 | 205.11 | 0.22 | 9.09 | 12.767* | 0.039 | 92.10* | 217.78* | 63.62* |
| FXG | 10 | 0.0037 | 121.36 | 0.24 | 409.84 | 7.61* | 0.099* | 12.36* | 35.21* | 4.56* |
| Error | 22 | 0.0043 | 116.48 | 0.23 | 209.95 | 2.94 | 0.024 | 0.26 | 3.87 | 0.35 |
| Mean | 2.4 | 354.43 | 7.33 | 171.14 | 61.62 | 1.56 | 46.59 | 54.37 | 15.49 | |
| CV(%) | 2.75 | 3.05 | 6.48 | 8.47 | 2.78 | 9.99 | 1.09 | 3.62 | 3.82 | |
| R | 0.84 | 0.87 | 0.52 | 0.84 | 0.84 | 0.89 | 0.99 | 0.99 | 0.95 | |
Table 3.
Flowered quality variables (with and without pedicel) of NCHs of pozolero maize 'EO' race evaluated in 2019, 2020 and 2021 in Celaya, Gto. Mexico.
Table 3.
Flowered quality variables (with and without pedicel) of NCHs of pozolero maize 'EO' race evaluated in 2019, 2020 and 2021 in Celaya, Gto. Mexico.
| FGY (kg kg-1maize) | VFG (cm3) |
FT (min) |
FGM (%) |
DML (%) |
FGPF (N) |
L (%) |
Hue (°) |
Chroma | |
|---|---|---|---|---|---|---|---|---|---|
| Pz1-19 | 2.26 ± 0.08* | 332.5 ± 6.45* | 147.5 ± 5.0* | 58.93 ± 1.2 | 7.23 ± 0.23 | 1.84 ± 0.07* | 51.92 ± 3.6* | 63.17 ± 3.4* | 16.72 ±2.9 |
| Pz2-19 | 2.32 ± 0.07 | 338.8 ± 33.15* | 151.3 ± 10.3* | 57.71 ± 2.9* | 7.71 ± 0.43 | 1.99 ± 0.2* | 47.53 ± 1.9* | 63.19 ± 3.3* | 14.54 ± 1.9* |
| Pz1-19C | 2.24 ± 0.11* | 320.0 ± 10.8* | 153.8 ± 7.5* | 57.61 ± 2.5* | 7.32 ± 0.53 | 1.91 ± 0.15* | 46.21 ± 3.2* | 61.08 ± 3.5* | 15.77 ± 0.5* |
| Pz2-19C | 2.32 ± 0.07 | 345.0 ± 10.8* | 150.0 ± 0.0* | 60.29 ± 2.0 | 7.21 ± 0.49 | 1.59 ± 0.12 | 47.52 ± 0.4* | 56.77 ± 3.2* | 15.34 ± 2.4* |
| Pz1-20 | 2.50 ± 0.05 | 375.0 ± 19.6 | 196.8 ± 22.6 | 62.71 ± 0.5 | 6.88 ± 0.49 | 1.19 ± 0.11 | 42.58 ± 1.2* | 40.83 ± 1.9* | 15.11 ± 0.6* |
| ♀Pz1-20 | 2.55 ± 0.05 | 385.0 ± 12.9 | 194.8 ± 17.04 | 63.55 ± 0.8 | 7.70 ± 0.20 | 1.14 ± 0.21 | 43.85 ± 1.2* | 46.68 ± 4.1* | 15.74 ± 1.3* |
| Pz2-20 | 2.51 ± 0.07 | 352.5 ± 8.7* | 161.3 ± 26.3* | 63.73 ± 1.9 | 7.23 ± 0.48 | 1.23 ± 0.12 | 46.76 ±1.1* | 49.34 ± 9.6* | 15.34 ± 0.5* |
| ♀Pz2-20 | 2.43 ± 0.02 | 365.0 ± 10.0 | 205.0 ± 13.21 | 62.79 ± 1.2 | 7.56 ± 0.42 | 1.61 ± 0.24 | 40.85 ± 0.4* | 39.08 ± 6.4* | 15.82 ± 0.3* |
| Pz1-21 | 2.38 ± 0.07 | 351.3 ± 2.5* | 160.0± 14.1* | 63.97 ± 4.0 | 7.32 ± 0.53 | 1.52 ± 0.30 | 42.38 ± 2.3* | 47.68 ± 2.7* | 15.55 ± 0.3* |
| Pz2-21 | 2.41 ± 0.06 | 356.3 ± 8.5 | 163.5 ± 16.3* | 64.17 ± 2.9 | 6.96 ± 0.80 | 1.76 ± 0.41* | 40.4 ± 5.6* | 41.53 ± 1.9* | 13.21 ± 2.5* |
| CACC | 2.45 ± 0.05 | 377.5 ± 8.7 | 198.8 ± 22.5 | 62.38 ± 1.0 | 7.50 ± 0.39 | 1.41 ± 0.19 | 62.48 ± 2.4 | 88.72 ± 2.8 | 17.21 ± 2.6 |
* FGY flowered grain yield, VFG flowered grain volume, FT flowering time, FGM flowered grain moisture, DML dry matter loss, FGPF flowered grain puncture force, L lightness, Hue hue angle, Chroma color purity. Means with (*) are statistically different from the control genotype CACC (Dunnett, 0.05).
Table 5.
Correlation between grain hardness (FI) with color characteristics (L, hue, chroma) and total anthocyanins (TA) in raw maice, flowered grain without and with pedicel.
Table 5.
Correlation between grain hardness (FI) with color characteristics (L, hue, chroma) and total anthocyanins (TA) in raw maice, flowered grain without and with pedicel.
| Raw grain | Flowered grain wihtout pedicel | Flowered grain wiht pedicel | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FI | L | Hue | Chroma | TA | L | Hue | Chroma | TA | L | Hue | Chroma | TA | |
| FI | 1 | 0.304 | 0.750* | -0.279 | 0.748* | -0.757* | -0.819* | -0.053 | 0.754* | -0.506* | -0.748* | -0.150 | 0.730* |
| L | 1 | 0.274 | 0.202 | 0.015 | 0.022 | -0.125 | -0.264 | 0.225 | -0.192 | -0.027 | -0.150 | 0.052 | |
| Hue | 1 | -0.317 | 0.734* | -0.678* | -0.539* | -0.603* | 0.896* | -0.612* | -0.692* | -0.243 | 0.776* | ||
| Chroma | 1 | -0.252 | -0.066 | 0.222 | 0.323 | -0.278 | -0.064 | 0.263 | -0.253 | -0.542* | |||
| TA | 1 | 0.760* | -0.830* | -0.214 | 0.860* | -0.658* | -0.840* | -0.488* | 0.831* | ||||
| LWOP | 1 | 0.722* | -0.034 | -0.719* | 0.586* | 0.716* | 0.554* | -0.562* | |||||
| HueWOP | 1 | -0.142 | -0.747* | 0.683* | 0.829* | 0.434 | -0.804* | ||||||
| ChromaWOP | 1 | -0.428 | 0.166 | 0.082 | -0.184 | -0.343 | |||||||
| ATWOP | 1 | -0.793* | -0.882* | -0.437 | 0.902* | ||||||||
| LWP | 1 | 0.833* | 0.477* | -0.713* | |||||||||
| HueWP | 1 | 0.431 | -0.852* | ||||||||||
| ChromaWP | 1 | -0.351 | |||||||||||
| TAWP | 1 | ||||||||||||
* Statistically significant (p≤0.05). WOP without pedicel, WP with pedicel.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



