Submitted:
17 May 2023
Posted:
18 May 2023
You are already at the latest version
Abstract
The concept of gray-zone lymphoma (GZL) has been progressively refined since its in-troduction in the literature in 1998. For several years, it was applied to a rather broad spectrum of conditions, posing the problem of the differential diagnosis between any type of Hodgkin lymphoma (HL) and diffuse large B-cell lymphoma, with special reference to primary mediasti-nal forms (PMBL). Officially recognized as a provisional entity in the 4th and revised 4th editions of the WHO Classification of Tumour of Haematopoietic and Lymphoid Tissues with the term “B-cell lymphoma unclassifiable with features intermediate between diffuse large B-cell lym-phoma and classical Hodgkin lymphoma”, it was limited to tumours, which showed either morphologic features reminiscent of classical HL (CHL) but carrying a complete B-cell pheno-type or conversely provided with a PMBL morphology but revealing CHL phenotypic charac-teristics. The definition of GZL has been further revised in the recently published International Lymphoma Classification and 5th Edition of the WHO Classification of Haemato-lymphoid Tu-mours, which have limited it to mediastinal neoplasms based on emerging molecular evidences. The aim of this review is to critically discuss the issue of GZL, also in the light of the suboptimal response to current therapies.
Keywords:
mediastinal gray-zone lymphoma
; classical Hodgkin lymphoma
; primary mediastinal B-cell lymphoma
; diffuse large B-cell lymphoma
; NOS
; EBV-positive diffuse large B-cell lymphoma
; morphology
; phenotype
; gene expression profile
; mutational landscape
1. Introduction
Gray-zone lymphoma (GZL) does represent a problem for both pathologists and clinicians, due to the rarity of the condition, quite subjective diagnostic criteria, lack of standard therapy, and aggressive clinical course. The aim of the present review is to discuss: a) the evolution of the concept of GZL, b) the potential limitations of tissue sampling, c) the more restrictive diagnostic criteria introduced by the most recent classification proposals, d) the novel pathobiological information provided by high-throughput technologies, and e) therapeutic perspectives.
2. Evolution of the Concept of Gray-Zone Lymphoma
The term gray-zone lymphoma (GZL) was originally proposed by Rüdiger et al. in 1998 as referred to a “distinct subset of cases representing mediastinal large B-cell lymphomas with features of Hodgkin’s lymphoma” [1]. In 2005, Poppema et al. underlined that “classical Hodgkin lymphoma (CHL) and at least a proportion of primary mediastinal B-cell lymphoma (PMBL) are derived from B cells at similar stages of differentiation and share common pathogenetic features” [2]. Such statement was supported by the evidence published two years before by Rosenwald et al., showing that the gene expression profile of PMBL was similar to the profile of CHL, with the exception of the preservation of the B-cell programme, repressed in CHL [3]. Through time, the concept of GZL was further expanded to tumours showing border-line characteristics between HL and large B-cell lymphoma (LBCL), by including also lymphocyte-rich CHL, nodular lymphocyte predominant Hodgkin lymphoma (NLPHL), and T-cell /histiocyte-rich LBCL, as well as cases of ambiguous morphology and phenotype located at extra-mediastinal sites and/or EBV-positive [4].
In 2008 and 2017, the fourth and revised fourth editions of the WHO Classification of tumours of the Haematopoietic and Lymphoid Tissues (IARC, Lyon) introduced the provisional entity “B-cell lymphoma unclassifiable with features intermediate between diffuse large B-cell lymphoma and classical Hodgkin lymphoma”, which included two subtypes, characterized by morphologic features reminiscent of CHL but carrying a complete B-cell phenotype or conversely provided with a PMBL morphology but revealing a CHL phenotypic profile (with strong CD30 and CD15 positivity, together with lack of most if not all B-cell markers as well as of CD45) [5,6].
During the last few years, several important contributions have been produced thanks to the extensive application of high-throughput molecular techniques as well as to the attempt to better understand the morphologic and phenotypic spectrum of primarily of mediastinal GZLs [7,8,9].
Currently, both the International Consensus Classification and the 5th Edition of the WHO Classification agree in limiting the concept of GZL to a mediastinal B-cell lymphoma with overlapping features between PMBL and CHL, especially nodular sclerosis CHL (NSCHL) [10,11]. Current evidence indicates that cases with morphologic and immunophenotypic features similar to mediastinal GZL (MGZL), but occurring outside and without involvement of the mediastinum, harbor different gene expression profiles and DNA alterations [12]. Hence, these cases are better classified as DLBCL, NOS. Last but not least, cases with composite or sequential CHL and PMBL, which are rare but do exist, are not currently included in the setting of GZL, since they need further studies to understand their biology, as they may represent examples of tumors with high cell plasticity [13].
3. Clinical Characteristics
The median age of patients with GZL is 33 years and approximately two thirds are males. Of these patients, nearly half have a bulky mediastinal mass >10 cm, while a minority have extranodal involvement and only 13% stage IV disease. The mediastinal mass can cause superior vena cava syndrome or respiratory distress. Supra-clavicular lymph nodes may be involved. There may be spread to lung by direct extension, as well as to liver, spleen, and bone-marrow, in particular in relapsing or resistant (R/R) cases [14].
4. The Biopsy Issues
The complexity of the diagnostic approach to GZL is further hampered by the amount of bioptic material available. Mediastinal masses, which fulfil the criteria of bulky disease, are often sampled by needle biopsies. Compatibly with the condition of the patient, a wedge biopsy in the course of a mini-thoracotomy would be much more useful to solve the problems of the differential diagnosis. This is a key point because therapies used for PMBL, CHL or MGZL are different as are the results they produce. In particular, MGZL have shown the worst outcome even when treated with intensive regimens like DA-EPOCH-R (see below) [15]. Finally, a fine needle aspirate cytology is totally inadequate [16].
5. Morphology and Phenotype
In spite of the criteria given by lymphoma classifications (WHO 4th, Revised 4th, 5th, and ICC), the diagnosis of MGZL remains rather subjective, a fact that can lead to use the term as a convenient basket category [5,6,10,11]. In a still ongoing study promoted by the Italian Lymphoma Foundation, most diagnoses of GZL rendered at peripheral Italian Hospitals were not confirmed on central review by an expert panel (unpublished data). Some attempts have been made to define the different morphologic and phenotypic conditions, which the term MGZL can be applied to. Sarkozy et al. identified four different groups (which they graded from zero to three) [7]. Group 0 shows a Hodgkin-like morphology with a scleronodular architecture and Reed-Sternberg cells (RSCs) in an inflammatory background. Tumoral cells express CD30 and show strong and diffuse positivity for CD20 on 100% of them along with other B-cell markers, such as CD79a, PAX5/BSAP, and OCT2. By comparison with the previous one, group 1 reveals a Hodgkin-like morphology with a more intermediate architecture, less fibrosis and numerous sheets of large mononucleated cells in an inflammatory background. Besides positivity for CD20 and other B-cell markers, such as BOB1 and OCT2, there is partial expression of CD15. Group 2 has a large B-cell morphology with some RSCs in an inflammatory background. Tumoral cells strongly express CD30 and CD15. PAX5 is moderately positive, while CD20 is negative. Other B-cell markers are variably expressed. Finally, Group 3 has a large B-cell morphology with fibrosis producing compartmentalization as observed in PMBL. Phenotypically, neoplastic cells strongly express CD20, CD30 and CD15. The stains for CD23 and MAL did not influence the subdivision into the four groups. The majority of cases express PD-L1/PD-L2 and carry CIITA and CD274 loci abnormalities on FISH analysis. One of the major limitations of the subclassification proposed by Sarkozy and coworkers was the inclusion of some non-mediastinal cases as well as of EBV-positive tumours, which the ICC and 5th edition of the WHO Classification take separate from GZLs.
The view of Sarkozy et al. is not shared by Egan and Pittaluga, who underlined that GZL is characterized by varying degrees of discordance between morphology and immunophenotype making its classification difficult [17]. Tumor cells are usually abundant and although traditionally categorized as ‘PMBL-like’ or ‘CHL-like’, the cytomorphological spectrum is in between that seen in PMBL or CHL. Neoplastic cells may have a centroblastic or immunoblastic appearance but can be larger and more pleomorphic. Lacunar cells or RS-like cells can be detected in variable numbers and turn to be more intermediate in size. The inflammatory milieu of CHL-like is often less apparent, with fewer eosinophils and plasma cells. Necrosis is variable but may be scarce or absent. Tumor architecture is usually diffuse, but occasionally a nodular growth pattern and coarse fibrosis may be found. Expression of PAX-5/BSAP, BOB.1, and OCT-2 is variable among GZLs, but usually there is positivity for at least one B-cell marker in a panel of CD20, CD79a, and PAX5. Cases rich in centroblastic-appearing cells may lack or only partially express B-cell markers (such as CD20) but show positivity for CD30 or moderate- strong staining for CD15. Tumors with an abundance of RS-like cells may have a strong expression of multiple B-cell markers/transcription factors and CD45, and lack CD15 positivity. CD30 is usually expressed, although it may be weak and positivity for CD23 may also occur. MAL may also be positive irrespective of histology. EBER positivity has occasionally been reported, although its interpretation is controversial (Figure 1 and Figure 2).
The tumour microenvironment (TME) of MGZL would merit more in-depth studies, taking into consideration, however, that it can significantly vary based on the classification proposed by Sarkozy and co-workers. For instance, it would be useful to assess whether there is any predictive test, which can herald therapeutic responses, as recently proposed for T-cell/histiocyte-rich large B-cell lymphoma [18].
6. Molecular features
As in PMBL and CHL, alterations have been reported at the JAK2/CD274/PDCD1LG2 locus at 9p24.1 and the CIITA locus at 16p13.13, including gains, amplifications, and rearrangements [19,20]. Alterations of the PD-L1 locus can represent the rational for the successful usage of immune check-point inhibitors, as discussed later. Gains at the REL locus at 2p.16.1 have been described in 33% of cases and gains at the MYC locus at 8q24 in 27% [20].
Methylation profiling of MGZL showed a distinct epigenetic profile intermediate between CHL and DLBCL but remarkably different from DLBCL [20]. By utilizing genes such as HOXA5, MMP9, EPHA7 and DAPK1 a final combined prediction of 100% was achieved between MGZL, CHL and PMBL [20]. Sarkozy and coworkers carried out a gene expression profiling study in a large series of GZLs (mediastinal and extra-mediastinal), CHLs, PMBLs, and polymorphic EBV+ DLBCLs of the NOS type (poly-EBV-DLBCLs) [8]. In an unsupervised principal component analysis, GZLs showed intermediate scores in a spectrum between CHL and PMBCL, whereas poly-EBV-DLBCLs clustered distinctly. The main biological pathways underlying the GZL spectrum were related to the cell cycle, which reflected the tumor cell content, and extracellular matrix signatures that were related to TME [8]. Differential expression analysis and phenotypic characterization of TME highlighted the predominance of regulatory macrophages in GZL as compared to CHL and PMBCL [8]. Notably, two distinct subtypes of GZL were distinguished and phenotypically reminiscent of PMBCL and DLBCL, respectively [8]. The former (PMBCL-type GZL) was characterized by clinical presentation in the “thymic” anatomic niche. Sarkozy and coworkers have also performed the first extensive next generation sequencing (NGS) study of GZL and related entities [9]. In particular, they studied coding sequence mutations of 50 EBV-negative GZLs and 20 poly-EBV-DLBCLs and compared them to examples of CHL, PMBL, and DLBCL [9]. Exomes of 21 GZL and 7 poly-EBV-L cases, along with paired constitutional DNA, were analyzed in a discovery cohort, followed by targeted sequencing of 217 genes in an extension cohort of 29 GZLs and 13 poly-EBV-DLBCLs. GZL cases with thymic niche involvement (anterior mediastinal mass) exhibited a mutation profile closely resembling CHL and PMBCL, SOCS1 (45%), B2M (45%), TNFAIP3 (35%), GNA13 (35%), LRRN3 (32%), and NFKBIA (29%) being the most recurrently mutated genes [9]. In contrast, GZL cases without thymic niche involvement (n = 18) had a significantly distinct pattern that was enriched in mutations related to apoptosis defects [TP53 (39%), BCL2 (28%), BIRC6 (22%)] and relatively depleted of mutations in GNA13, XPO1, or NF-kB signaling pathway genes (TNFAIP3, NFKBIE, IKBKB, NFKBIA) [9]. They also exhibited more BCL2/BCL6 rearrangements compared with thymic GZL. Poly-EBV+ DLBCLs presented a distinct mutational profile, including STAT3 mutations and a significantly lower coding mutation load in comparison with EBV- GZLs [9].
The two above mentioned contributions of the French Group, nicely commented by Elias Campo and Elaine S. Jaffe in Blood, have guided the decision to limit the concept of GZL to EBV-negative mediastinal tumours in both the ICC and 5th edition of the WHO Classification [10].
Preliminary data of a study carried out by our Group on a series of GZLs, which underwent NGS at the time of diagnosis and/or at relapse (unpublished), suggest that GZL harbors a complex clonal structure. Mutations affecting epigenetic controllers appear to be neutral events that have likely arisen early in tumour evolution. Conversely, mutations affecting genes under positive selection are sub-clonal events at diagnosis that subsequently expand their estimated clone size after therapy, possibly underpinning mechanisms of drug-resistance. Thus, further studies are required to unravel the molecular characteristics of GZL in the light of its poor response to current therapies.
7. Therapeutic Strategies
There is no standard therapy for MGZL [21,22]. Among 112 patients treated across 19 North American centers over a ten-year period, two regimens were predominately used: A) cyclophosphamide, doxorubicin, oncovin, and prednisone (CHOP) +/- rituximab; B) doxorubicin, bleomycin, vinblastine, and dacarbazine (ABVD) +/- rituximab. The overall response (ORR) and complete response (CR) rates were 82% and 73% for the patients who received rituximab, respectively, vs. 59% and 43% for those who did not, respectively [23]. Notably, progression-free survival (PFS) rate was significantly inferior for patients treated with ABVD +/- rituximab compared to patients treated with CHOP +/- rituximab [23]. Recently, DA-EPOCH-R has been proposed as front-line therapy, based on the efficacy shown in PMBL [24]. The results are better than the ones recorded with R-CHOP but not as brilliant as in PMBL with a 5-year event-free survival (EFS) of 62% (PMBL 93%) and an overall survival (OS) of 74% (PMBL 97%) [24]. Thus, MGZL appears to be a relatively chemotherapy-resistant disease with a worse prognosis than PMBL and CHL [21,22,23,24]. The role of radiotherapy is debated [21]. It has at times been used as a consolidative tool after chemoimmunotherapy for localized and/or bulky disease. However, no conclusion can be drawn due to the relatively small case number. The majority of patients with R/R disease have been taken to hematopoietic stem cell transplantation (HSCT), more often autologous (ASCT). The 2-year OS of these patients was 88% vs. 67% for those, who did not receive HSCT. However, caution should be used in analyzing such results, since fit-patients with chemo-sensitive disease were likely more frequently brought to HSCT [21,22,23]. Based on the results observed in PMBL [25], a very recent publication has reported that the combination of Brentuximab Vedotin (BV) and Nivolumab is effective in about 70% of patients with R/R MGZL [26]. BV is an anti-CD30 humanized monoclonal antibody conjugated with monomethyl auristatin E and the expression of CD30 by tumor cells represents the rationale of its usage [25,26]. BV was found effective also as maintenance after ASCT in R/R GZL [27]. Besides the target effect, BV may also induce an immunogenic environment contributing to depletion of regulatory T-cells that can potentiate the effect of immune-checkpoint inhibitors, such as Nivolumab [26]. Nivolumab is an anti-PD1 monoclonal antibody that acts by restoring T-cell- mediated antitumor responses via blockade of PD-1/ligand interactions, a mechanism thought to be critical in GZL, as supported by the findings of frequent copy-number alterations of 9p24.1 and expression of PD-L1 and – less frequently – PD-L2 (see above) [20,25,26].
8. Conclusions and Future Directions
From what above discussed, GZL does still represent a condition, which the title of Pirandello’s drama “Six characters in search of an author” does quite well apply to. The rarity of the disease, the diagnostic difficulties and the suboptimal response to most therapies underline the need for internationally shared guidelines. The complexity is further increased by the possibility that MGZL might not represent a single entity but rather a spectrum of diseases that require an individual tailoring of therapies. The 5th edition of the WHO Classification and ICC incorporate in their criteria an increased understanding of MGZL. Nonetheless, current studies are insufficient and more in depth molecular characterization is needed to further understand the pathobiology of MGZL, It is predictable that an improved understanding of genetic aberrations and microenvironmental characteristics of MGZL will lead to more effective targeted therapeutic approaches.
Author Contributions
All authors have read and agreed to the published version of the manuscript.
Funding
The manuscript was supported by the AIRC 5x1000 Grant number 21198 (AIRC: Italian Association for Cancer Research, Milan, Italy).
Institutional Review Board Statement
The “unpublished data” were obtained in the course of two studies conducted according to the guidelines of the Declaration of Helsinki and approved by the Institutional Review Board of the European Institute of Oncology (protocol code UID 2701) and the Ethical Committee of the Region Tuscany (CEAVC Em. 2022-263 Studio 18236_oss), respectively. The remaining parts of the manuscript represent a review of the literature, which does not require any approval.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the studies, whose results are quoted as “unpublished data”.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rüdiger. T., Jaffe, E.S., Delsol, G., deWolf-Peeters, C., Gascoyne, R.D., Georgii, A., Harris, N.L., Kadin, M.E., MacLennan, K.A., Poppema, S., et al. Workshop report on Hodgkin’s disease and related diseases (‘gray zone’ lymphoma. Ann. Oncol. 1998, Suppl. 5, 31–38.
- Poppema, S. , Kluiver, J.L., Atayar, C., van den Berg, A., Rosenwald, A., Hummel, M., Lenze, D., Lammert, H., Stein, H., Joos, S., et al. Report: workshop on mediastinal gray zone lymphoma. Eur. J. Haematol. 2005, Suppl. 66, 45–52. [Google Scholar] [CrossRef]
- Rosenwald, A. , Wright, G., Leroy, K., Yu, X., Gaulard, P., Gascoyne, R.D., Chan, W.C., Zhao, T., Haioun, C., Greiner, T.C., et al. Molecular diagnosis of primary mediastinal B cell lymphoma identifies a clinically favorable subgroup of diffuse large B cell lymphoma related to Hodgkin lymphoma. J. Exp. Med. 2003, 198, 851–862. [Google Scholar] [PubMed]
- Harris, N.L. Shades of gray between large B-cell lymphomas and Hodgkin lymphomas: differential diagnosis and biological implications. Mod. Pathol. 2013, Suppl. 26, 57–70. [Google Scholar] [CrossRef]
- Jaffe, E.S. , Harris, N.L., Stein, H., Campo, E., Stefano Pileri, S.A., Swerdlow, S.H. B-cell lymphoma, unclassifiable, with features intermediate between diffuse large B-cell lymphoma and classic Hodgkin lymphoma. In: WHO Classification of tumours of haematopoietic and lymphoid tissues. Fourth edition. Editors: Swerdlow, S.H., Campo, E., Harris, N.L., Jaffe, E.S., Pileri, S.A., Stein, H., Thiele, J., Vardiman, J.W. IARC press, Lyon, France; 2008. pp. 267–268.
- Jaffe, E.S. , Stein, H., Swerdlow, S.H., Campo E., Pileri, S.A., Harris, N.L. B-cell lymphoma, unclassifiable, with features intermediate between diffuse large B-cell lymphoma and classic Hodgkin lymphoma. In: WHO Classification of tumours of haematopoietic and lymphoid tissues. Revised fourth edition. Editors: Swerdlow, S.H., Campo, E., Harris, N.L., Jaffe, E.S., Pileri, S.A., Stein, H., Thiele, J., Arber, D.A., Hasserjian, R.P., Le Beau, M.M., Orazi, A., Siebert, R. IARC press, Lyon, France; 2017. pp. 342–344.
- Sarkozy, C. , Copie-Bergman, C., Damotte, D., Ben-Neriah, S., Burroni, B., Cornillon, J., Lemal, R., Golfier, C., Fabiani, B., Chassagne-Clément, C., et al. Gray-zone Lymphoma Between cHL and Large B-Cell Lymphoma: A Histopathologic Series From the LYSA. Am. J. Surg. Pathol. 2019, 43, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Sarkozy, C. , Chong, L., Takata, K., Chavez, E.A., Miyata-Takata, T., Duns, G., Telenius, A., Boyle, M., Slack, G.W., Laurent, C., et al. Gene expression profiling of gray zone lymphoma. Blood Adv. 2020, 4, 2523–2535. [Google Scholar] [CrossRef] [PubMed]
- Sarkozy, C. , Hung, S.S., Chavez, E.A., Duns, G., Takata, K., Chong, L.C., Aoki, T., Jiang, A., Miyata-Takata, T., Telenius, A., Slack, G.W, et al. Mutational landscape of gray zone lymphoma. Blood 2021, 137, 1765–1776. [Google Scholar] [CrossRef]
- Campo E, Jaffe ES, Cook JR, Quintanilla-Martinez L, Swerdlow SH, Anderson KC, Brousset P, Cerroni L, de Leval L, Dirnhofer S, et al. The International Consensus Classification of Mature Lymphoid Neoplasms: a report from the Clinical Advisory Committee. Blood 2022, 140, 1229–1253. [Google Scholar] [CrossRef]
- Alaggio, R. , Amador, C., Anagnostopoulos, I., Attygalle, A.D., Araujo, I.B.O., Berti, E., Bhagat, G., Borges, A.M., Boyer, D., Calaminici, M., et al. The 5th edition of the World Health Organization Classification of Haematolymphoid Tumours: Lymphoid Neoplasms. Leukemia 2022, 36, 1720–1748. [Google Scholar] [CrossRef]
- Campo, E. , Jaffe, E.S. Taking gray zone lymphomas out of the shadows. Blood 2021, 137, 1703–1704. [Google Scholar] [CrossRef]
- Aussedat, G. , Traverse-Glehen, A., Stamatoullas, A., Molina, T., Safar, V., Laurent, C., Michot, J.M., Hirsch, P., Nicolas-Virelizier, E., Lamure, S., et al. Composite and sequential lymphoma between classical Hodgkin lymphoma and primary mediastinal lymphoma/diffuse large B-cell lymphoma, a clinico-pathological series of 25 cases. Br. J. Haematol. 2020, 189, 244–256. [Google Scholar] [PubMed]
- Kritharis, A. , Pilichowska, M., Evens, A.M. How I mange patients with gray zone lymphoma. Br. J. Haematol. 2016, 174, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Chihara, D. , Westin, J.R., Miranda, R.N., Cheah, C.Y., Oki, Y., Turturro, F., Romaguera, J.E., Neelapu, S.S., Nastoupil, L.J., Fayad, L.E., et al. Dose adjusted-EPOCH-R and mediastinal disease may improve outcomes for patients with gray-zone lymphoma. Br. J. Haematol. 2017, 179, 503–506. [Google Scholar] [CrossRef] [PubMed]
- Vitagliano, G. , Cretella, P., Zeppa, P., Caputo, A. Large-cell lymphoma with features intermediate between Hodgkin’s, primary mediastinal B-cell and gray-zone lymphoma: a conundrum on fine needle aspiration cytology. Cytopathology 2020, 31, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Egan, C. , Pittaluga, S. Into the gray-zone: update on the diagnosis and classification of a rare lymphoma. Expert Rev. Hematol. 2020, 13, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Tabanelli, V. , Melle, F., Motta, G., Mazzara, S., Fabbri, M., Agostinelli, C., Calleri, A., Del Corvo, M., Fiori, S., Lorenzini, D., et al. The identification of TCF1+ progenitor exhausted T cells in THRLBCL may predict a better response to PD-1/PD-L1 blockade. Blood Adv. 2022, 6, 4634–4644. [Google Scholar] [CrossRef] [PubMed]
- Steidl, C. , Gascoyne, R.D. The molecular pathogenesis of primary mediastinal large B-cell lymphoma. Blood 2011, 118, 2659–2669. [Google Scholar] [CrossRef]
- Eberle, F.C. , Rodriguez-Canales, J., Wei, L., Hanson, J.C., Killian, J.K., Sun, H.W., Adams, L.G., Hewitt, S.M., Wilson, W.H., Pittaluga, S., et al. Methylation profiling of mediastinal gray zone lymphoma reveals a distinctive signature with elements shared by classical Hodgkin’s lymphoma and primary mediastinal large B-cell lymphoma. Haematologica 2011, 96, 558–566. [Google Scholar]
- Kritharis, A. , Pilichowska, M., Evens, A.M. How I manage patients with gray zone lymphoma. Br. J. Haematol. 2016, 174, 345–350. [Google Scholar] [CrossRef]
- Dunleavy, K. Primary mediastinal B-cell lymphoma: biology and evolving therapeutic strategies. Hematology Am. Soc, Hematol. Educ. Program 2017, 1, 298–303. [Google Scholar]
- Evens, A.M. , Kanakry, J.A., Sehn, L.H., Kritharis, A., Feldman, T., Kroll, A., Gascoyne, R.D., Abramson, J.S., Petrich, A.M., Hernandez-Ilizaliturri, F.J., et al. Gray zone lymphoma with features intermediate between classical Hodgkin lymphoma and diffuse large B-cell lymphoma: characteristics, outcomes, and prognostication among a large multicenter cohort. Am. J. Hematol. 2015, 90, 778–83. [Google Scholar] [PubMed]
- Wilson, W.H. , Pittaluga, S., Nicolae, A., Camphausen, K., Shovlin, M., Steinberg, S.M., Roschewski, M., Staudt, L.M., Jaffe, E.S., Dunleavy, K. A prospective study of mediastinal gray-zone lymphoma. Blood 2014, 124, 1563–1569. [Google Scholar] [CrossRef] [PubMed]
- Zinzani, P.L. , Santoro, A., Gritti, G., Brice, P., Barr, P.M., Kuruvilla, J., Cunningham, D., Kline, J., Johnson, N.A., Mehta-Shah, N., et al. Nivolumab Combined With Brentuximab Vedotin for Relapsed/Refractory Primary Mediastinal Large B-Cell Lymphoma: Efficacy and Safety From the Phase II CheckMate 436 Study. J. Clin. Oncol. 2019, 37, 3081–3089. [Google Scholar] [PubMed]
- Santoro, A. , Moskowitz, A.J., Ferrari, S., Carlo-Stella, C., Lisano, J.M., Francis, S., Wen, R., Akyol, A., Savage, K.J. Nivolumab combined with brentuximab vedotin for relapsed/refractory mediastinal gray zone lymphoma. Blood online ahead of print. 2023. [Google Scholar] [CrossRef]
- Terao, T. , Yuda, J., Yamauchi, N., Guo, Y.-M., Shimada, K., Sugano, M., Ishii, G., Minami, Y. Brentuximal vedotin after autologous stem cell transplantation for refractory gray zone lymphoma with long term remission. Mol. Clin. Oncol. 2021, 14, 125–127. [Google Scholar] [CrossRef]
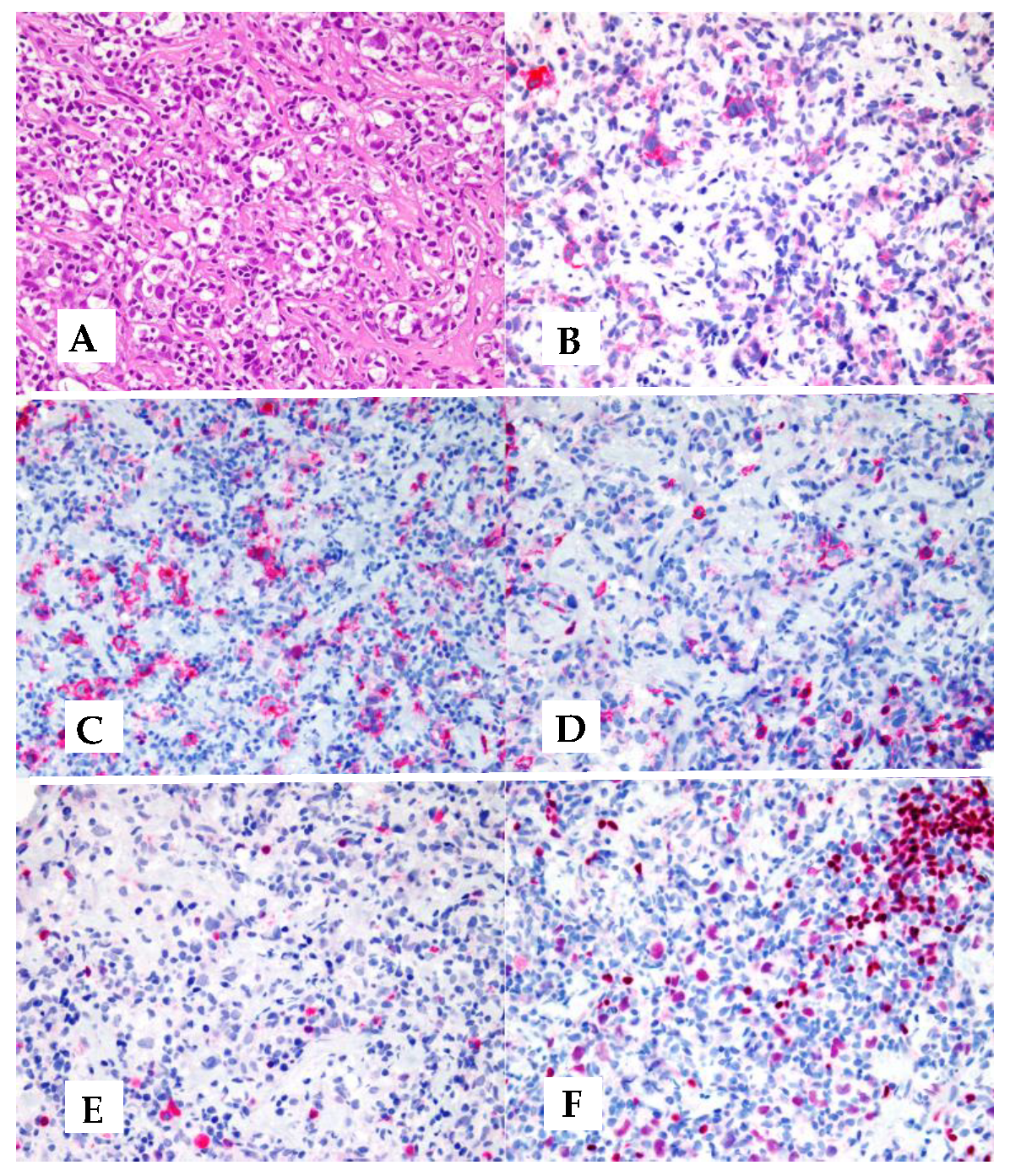
Figure 1.
Example of MGZL with CHL-like morphology: A) Fibrosis with nodular growth, reminiscent of nodular sclerosis CHL. B) Reed-Sternberg-like cells as well as mononuclear neoplastic cells are comprised within a reach inflammatory milieu. C) An area with a sheet of neoplastic cells centered by necrosis. D) CD30 is partly and variably expressed by neoplastic cells. D) CD15 turns negative. F-L) Strong positivity of neoplastic cells for CD20, CD79a, PAX5/BSAP, and BOB1. (Original magnification: x4 for Figure 1A, x40 for all the remaining ones. A-C: hematoxylin and eosin; D – L: immunoalkaline phosphatase technique, Gill’s hematoxylin nuclear counterstaining).
Figure 1.
Example of MGZL with CHL-like morphology: A) Fibrosis with nodular growth, reminiscent of nodular sclerosis CHL. B) Reed-Sternberg-like cells as well as mononuclear neoplastic cells are comprised within a reach inflammatory milieu. C) An area with a sheet of neoplastic cells centered by necrosis. D) CD30 is partly and variably expressed by neoplastic cells. D) CD15 turns negative. F-L) Strong positivity of neoplastic cells for CD20, CD79a, PAX5/BSAP, and BOB1. (Original magnification: x4 for Figure 1A, x40 for all the remaining ones. A-C: hematoxylin and eosin; D – L: immunoalkaline phosphatase technique, Gill’s hematoxylin nuclear counterstaining).

Figure 2.
A MGZL with PMBL-like morphology: A) The tumour consists of large cell with clear cytoplasm, which are subdivided into cluster by a fibrotic reaction. B, C) Neoplastic cells express CD30 and CD15. D, E) Only a few tumoral cells turn positive for CD20, while CD79a staining is confined to scattered reactive plasmacells. F) The positivity for PAX5/BSAP varies from negative to moderate, being always lower than the one of reactive small B-lymphocytes (Original magnification: x40. A: hematoxylin and eosin; B-F: immunoalkaline phosphatase technique, Gill’s hematoxylin nuclear counterstaining).
Figure 2.
A MGZL with PMBL-like morphology: A) The tumour consists of large cell with clear cytoplasm, which are subdivided into cluster by a fibrotic reaction. B, C) Neoplastic cells express CD30 and CD15. D, E) Only a few tumoral cells turn positive for CD20, while CD79a staining is confined to scattered reactive plasmacells. F) The positivity for PAX5/BSAP varies from negative to moderate, being always lower than the one of reactive small B-lymphocytes (Original magnification: x40. A: hematoxylin and eosin; B-F: immunoalkaline phosphatase technique, Gill’s hematoxylin nuclear counterstaining).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



