
Submitted:
28 April 2023
Posted:
29 April 2023
You are already at the latest version
Abstract
An impaired healing response underlies diabetic foot wounds chronicity, frequently translating in amputation, disability, and early mortality. In addition, diabetics frequently suffer of the underappreciated episode of post-epithelization ulcer recurrence. Recurrence epidemiological data are alarmingly high; so that the ulcer is considered in “remission” and not healed for the time it remains epithelialized. Recurrence may result from the conspiracy of behavioral and biological factors. Although the damaging role of behavioral, clinical predisposing factors is undebatable, the role of endogenous biological signalers that prime the residual scar tissue to recurrence has remained elusive. We propose that ulcer recurrence is deeply impinged by chronic hyperglycemia and its downstream biological effectors, which originate epigenetic drivers that enforce abnormal pathologic phenotypes to dermal fibroblasts, and keratinocytes as memory cells. Hyperglycemia-derived cytotoxic reactants accumulate and modify dermal proteins, reduce scar tissue mechanical tolerance, and disrupt fibroblasts-secretory activity. Accordingly, the combination of epigenetic and local and systemic cytotoxic signalers, induce the onset of “at-risk phenotypes” as premature skin cells aging, dysmetabolism, inflammatory, pro-degradative, and oxidative programs that may ultimately converge to scar cells demise. Unfortunately, post epithelialization recurrence rates data are missing in clinical studies of accepted ulcer healing therapies during follow-up periods. Intra-ulcer infiltration of epidermal growth factor exhibits the most consistent remission data with the lowest recurrences during 12-month follow-up. Recurrence data should be regarded as a valuable clinical endpoint during the investigational period for each emergent healing candidate.
Keywords:
Diabetic foot ulcer
; ulcer remission
; ulcer recurrence
; ulcer relapse
; diabetic complications
; metabolic memory
Introduction
The prevalence of both type 1 and type 2 diabetes (T1DM and T2DM) and the associated complications are globally increasing (1-3). For T2-DM, its incidence has progressively escalated approaching to a pandemic condition that accounts for 90% to 95% of all the diabetic population (4).
Diabetes-affected patients have a significantly shorter life expectancy than nondiabetic individuals (5, 6). Accordingly, the seminal revolution brought about by insulin treatment, did not translate into a significant reduction of the chronic complications that preside morbidity and mortality (7).
Mounting evidences support the existence of a diabetes metabolic memory as a proximal trigger in the perpetuation of multi-organ complications (8), including the torpid healing response (9). Diabetic foot ulceration (DFU) is one of the most frightened diabetic complications, leading to amputation, disability, social exclusion, and early mortality (10). The lifetime incidence of foot ulcers has been estimated to reach up to 34% of diabetic subjects, contributing to 80% of all non-traumatic lower extremities amputations around the world (11, 12).
Although the onset of DFU is associated to predisposing factors as peripheral neuropathy, limb ischemia, cutaneous frailty; diabetic patients are affected by an intrinsic healing impairment that spans from the oral cavity mucosa, to lower extremities peripheral soft tissues (13-15). DFU chronicity phenotype (16) seems to be driven by precocious senescence, proliferative arrest, and apoptosis, as distal effectors of an abnormal hyperglycemia-associated epigenetic code (17, 18).
Aside from the impaired healing response, a parallel conundrum in diabetic ulcer pathology is the high rate of recurrences after the primary reepithelialization (19, 20). In line with this fact, a classic report reveals that roughly 40% of diabetic patients have a recurrence within 1 year after ulcer healing (21, 22). Additionally, an underappreciated risk of DFU recidivism is its ability to “metastize” at anatomical niches away from the primary occurrence, frequently leading to a subsequent amputation (23).
Ulcer recurrence investigation has mostly focused on the identification and validation of predisposing clinical factors as poor glycemic control, previous ulcers, wound healing time, local skin damages, bone deformities, neuropathy, ischemia, diabetes duration, end-stage renal disease, toxic habits, and scar tissue mechanical stress (22, 24, 25). Although the deleterious role of behavioral factors is unquestionable (21, 26), we assume that there are endogenous biological signalers that prime the residual scar tissue to recurrence. Studies addressed to identify molecular pro-recurrence predictors are a contemporary need. We consequently examined potential molecular and cellular drivers, of local or systemic origin, that may concertedly cooperate behind ulcer recurrence. Having reviewed the literature, we may conclude that: (1) Endogenous deterrent factors underpinning the diabetic healing deficit are likely the same that cooperate for ulcer relapse. (2) These endogenous factors may be represented by soluble circulating signalers as by dysfunctional cells that secrete “pathological messages”. (3) These factors are a consequence of hyperglycemia whereas some of them are glucose-derived chemicals. (4) Both wound chronification and recurrence are influenced by an abnormal cell physiology on the bases of an epigenetic code, resulting from the interaction of glucose and its derivatives with expression/transcription regulatory factors. (4) Numerous published clinical trials on innovative treatments for the healing of DFU, omit to mention recurrence rates during post epithelialization follow-up. Intervention with locally infiltrated epidermal growth factor (EGF) appears as a promising tool to achieve prolonged remission times.
The information analyzed for this work was retrieved from Pubmed and Google Scholar data bases, restricted to English language with no date limitation.
II.1- Putative epigenetic drivers in ulcer recurrence
Scar tissue is vulnerable to recurrence, an event that is theoretically propelled by an abnormal cellular physiology with an underlying epigenetic program, and a chemically-modified extracellular matrix by toxic products (17). Thus, ulcer relapses alike the impaired diabetic healing are clinical consequences of poor glycemic control, highlighting the pathogenic participation of endogenous cytotoxic-recurrence primers (20). Blood glucose levels and the hyperglycemia-derived products, constitute an environmental factor that impacts on the plasticity of epigenetic mechanisms, thereby modifying the whole transcriptome of skin cells (27). A primary multi-organ effect originated by deregulated glycemia consists in disrupting three major biological functions: DNA expression, RNA transcription, and protein translation. Studies have indicated that hyperglycemia may increase DNA mutations, DNA breaks, genomic instability, and particularly epigenetic dysregulation (28, 29). Accordingly, hyperglycemia and a constellation of downstream factors transform the cellular native epigenetic architecture, rendering a de novo code that largely alters cellular physiology (30). A clear association exists among hyperglycemia, the pattern of DNA methylation changes, diabetes complications unset and their clinical progression (31). Abnormal DNA methylation predisposes to diabetes susceptibility genes (32) and to perpetuate the diabetic phenotype in ulcer-derived fibroblasts, which is independent to the number of culture passages, and the presence of a normal glucose concentration in the culture medium. The discovery of this epigenotype, supports the role of epigenetic behind the persistent diabetic phenotypic behavior of ulcer’s fibroblasts population (33). Another form of epigenetic modification, histone methylation, has been shown to correlate with glycemia levels and in turn with the formation of advanced glycation end products (AGEs) and the ensued hyperglycemia-related inflammation, oxidative stress, and apoptosis (34). It is known that AGEs as a biochemical hallmark of diabetes and a crucial ingredient within the damage cascade stick to histones, modifying chromatin structure and therefore its interaction capabilities, rendering an abnormal transcriptome (Figure 1) (35, 36).
Figure 1.
Theoretical epigenetic bases of recurrence.
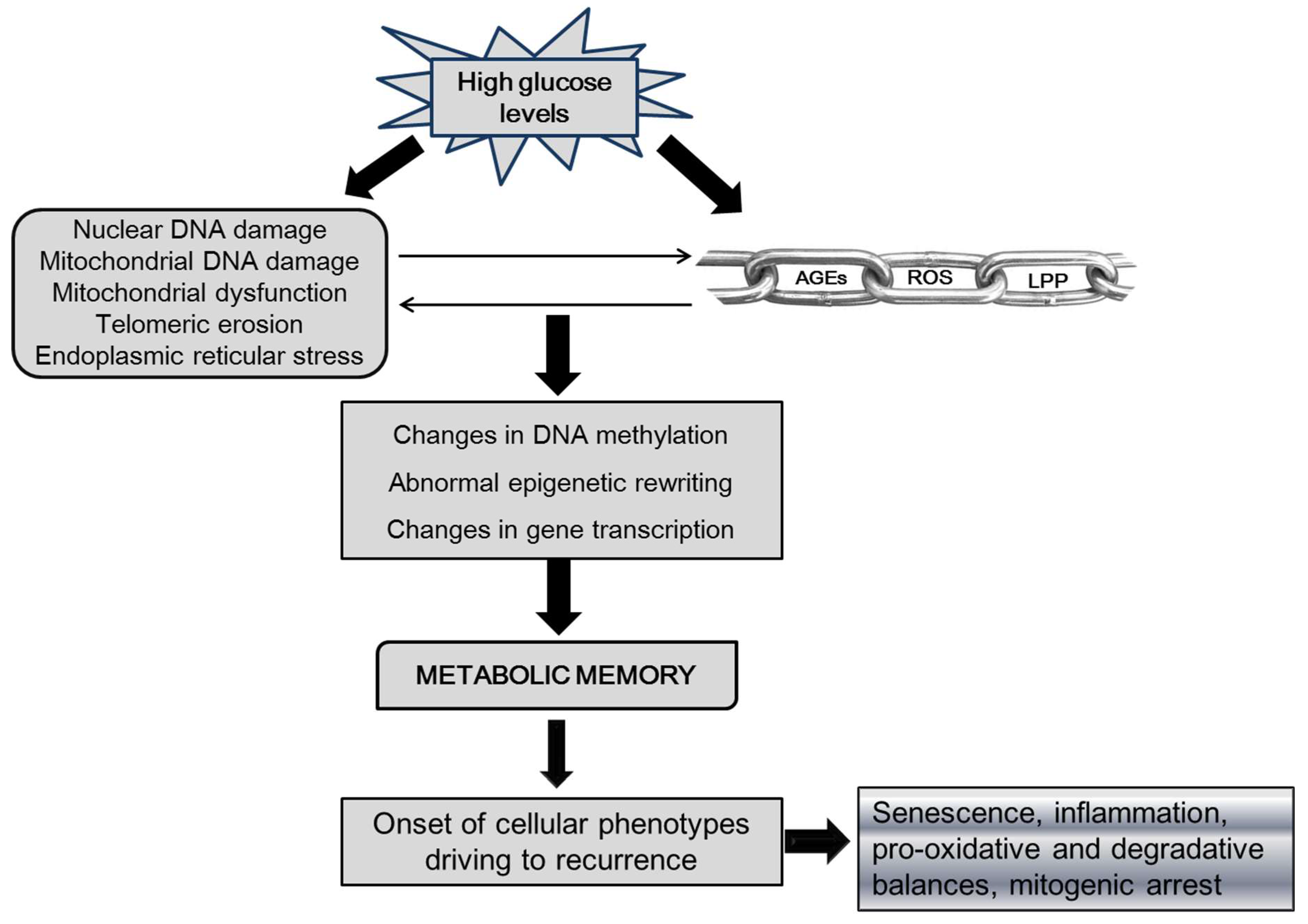
Legend: AGEs-advanced glycation end-products, ROS-reactive oxygen species, LPP-lipoperoxidation products. Chronic high glucose level is the primary precursor of AGEs formation and accumulation in tissues, which is in pathophysiological link to ROS and LPP formation and accumulation. Hyperglycemia, AGEs, and ROS have a direct cytotoxic effect which includes damaging DNA and impairing mitochondrial function, which further amplifies ROS production and hypometabolism. High glucose burden and the above mentioned derivatives, introduce a novel epigenetic imprinting on the bases of a DNA aberrant methylation pattern, and abnormal transcription, and expression profiles. This foundation of metabolic memory perpetuates the diabetic phenotype in fibroblasts and keratinocytes, which includes inflammatory and senescence activators as proliferative arrest.
As above mentioned, hyperglycemia also modifies transcriptional profile by affecting mRNA, transcription factors, the production and extracellular release of microRNAs (miRNA), and long non-coding RNA (lncRNA) (37). El-Osta’s findings inaugurated the contention that exposure to transient hyperglycemia, was sufficient to reprogram cells’ native epigenetic program, and that these modifications largely persisted even beyond the normalization of glucose levels (38). In other words, the acute exposure of cells to high glucose stress, translates in chronic consequences that involves the onset of lasting pro-inflammatory and oxidative programs upon an altered epigenetic code. This concept was further validated in human cultured skin primary fibroblast (27) and vascular cells while showing that high glucose concentrations introduced significant transcriptomic modifications in genes controlling multiple pathways, all involved in wound healing events including angiogenesis (39).
MicroRNAs (miRNA) and non-coding RNAs in general are involved in an extensive array of cellular functions, representing an additional layer of epigenetic control in cell physiology (28). Deficiency of miRNA biogenesis has revealed their biological significance in the skin healing process (40). Interestingly, there is a substantial specificity and differentiation of miRNAs profiles for the specific type of diabetes and the evolving complications, which has encouraged its application as diagnosis and prediction biomarkers (40, 41). A large number of microRNAs have been implicated in pathological diabetic healing (40-45). A recent study identified the enhanced expression of miR-155 in peripheral blood of T2DM patients as a potential predictor for the onset of DFU (46), whereas the expression level of miR-203 in patients with DFU, positively correlated with the severity of the damage (47). High expression levels of miR-34c positively correlated with the amputation rates while negatively with the healing response, being also identified as an independent risk factor for ulceration and osteomyelitis (48). Likewise, long non-coding RNAs (lncRNAs) are an important epigenetic regulator at the level of histone methylation and gene transcription (37, 49). lncRNAs play a substantial role in diabetic wound healing, encompassing from infiltrating macrophages polarization control, to keratinocytes proliferation, and migration. Differential expression of lncRNAs in diabetic patients shows their involvement in impaired diabetic healing, and their putative role as biomarkers for diabetes-mediated damages (49-51). Although miRNAs and lncRNAs are included in the broad collection of hyperglycemic stress-related epigenetic derangements (52), it still remains elusive if some of the non-coding RNA forms are associated to scar tissue relapse, and could therefore be used as predictive biomarker.
Conclusively, genetic and epigenetic cell resources are impacted by chronic hyperglycemia establishing an abnormal epigenetic program that predisposes to the onset and perpetuation of diabetic traits as cellular senescence, proliferative quiescence, inflammation, oxidative imbalance, and apoptosis (39). Thus, scar tissue integrity is intrinsically jeopardized by these underlying silent primers, which may lead to tissue death as the ultimate event in recurrence, validating the concept of ulcer remission instead of ulcer healing.
II.2- Dermal matrix, fibroblasts, and keratinocytes in ulcer recurrence
Diabetes disrupts skin structure and physiology. Chronic and irregular courses of glucose and glucoxidation-derived products undermine skin cells physiology and progressively intoxicates the dermal matrix by the accumulation of AGEs, nitrosilation products, and free radicals byproducts (53, 54). Diabetics’ intact skin shows reduced biomechanical resilience and stress tolerance, diminished elasticity, increased stiffness, and reduction of collagen and elastin contents, all together contributing to healing impairment (41, 55), and predisposing to scar tissue and epithelial coverage fractures. The above described dermal anomalies are largely associated to accumulation of AGEs crosslinked with long half-life proteins as collagen, which irreversibly and progressively affects skin matrix mechanics, induces premature aging, and impairs critical healing events as angiogenesis, fibroblasts attachment, and myofibroblasts-induced contraction (56, 57). Both AGEs and oxidative stress have direct cytotoxic effect on skin fibroblasts physiology, are instrumental ingredients of the ulcer cytopathic milieu, and ultimately contribute to mold the diabetes epigenetic map (Figure 2) (58, 59). It is therefore inferable that scar tissue cells are born embedded within a milieu permeated by cytotoxic products.
Figure 2.
Impact of hyperglycemia and derived cytotoxic products on the dermal matrix and skin cells.
Figure 2.
Impact of hyperglycemia and derived cytotoxic products on the dermal matrix and skin cells.
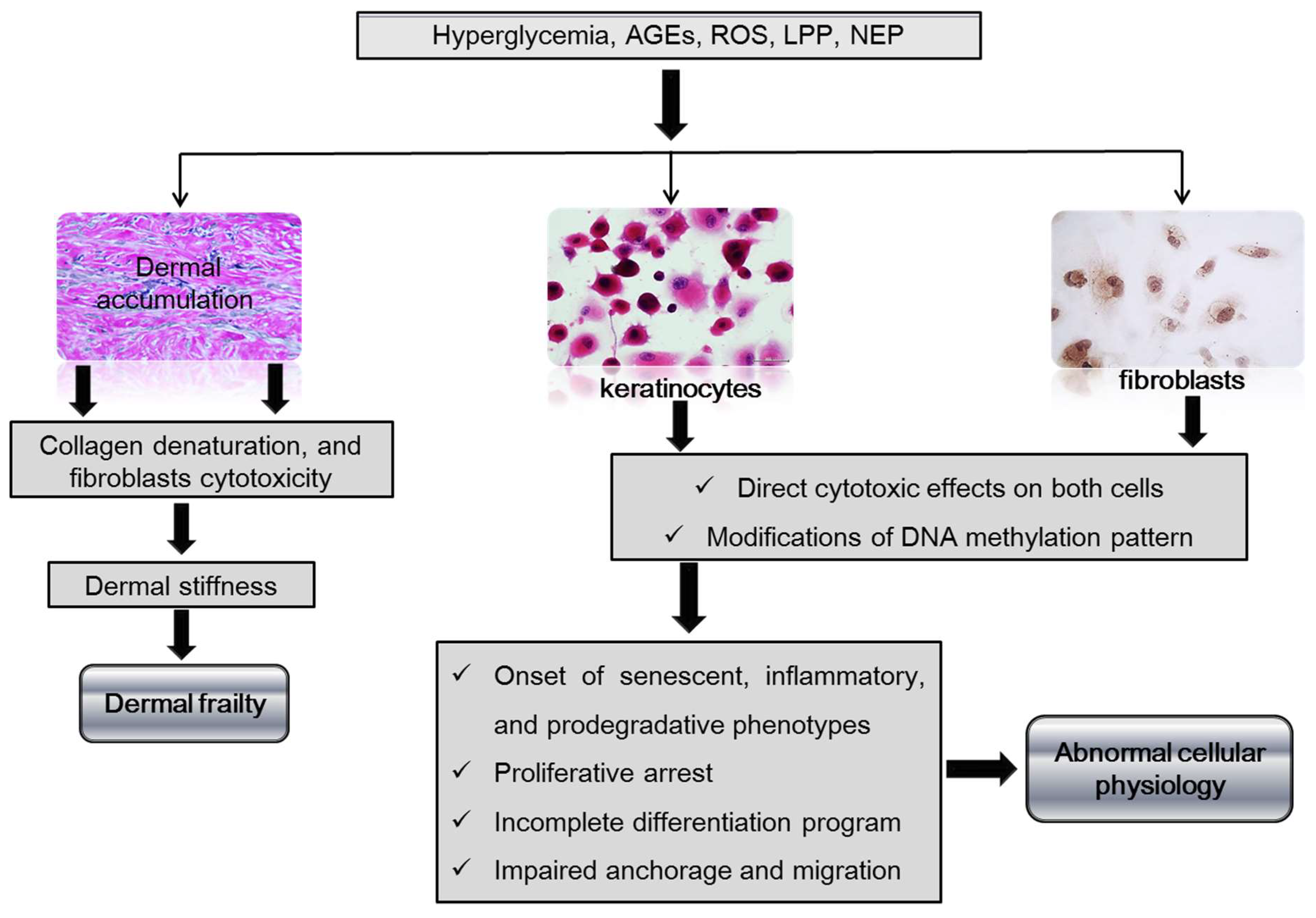
Legend: AGEs-advanced glycation end-products, ROS-reactive oxygen species, LPP-lipoperoxidation products, NEP-nitrosylation end-products. Hyperglycemia and its metabolic associated derivatives as AGEs and ROS accumulate in the dermal collagen, and alter the chemical and physical properties of the skin, becoming vulnerable and frail. In addition these are cytotoxic products that jeopardize dermal fibroblast survival and secretory activity. Hyperglycemia and the downstream cytotoxic products damage the native DNA methylation pattern, subsequently modifying gene transcription. The onset of a diabetic phenotype with an abnormal behavior of keratinocytes and fibroblasts is driven by a de novo re-written epigenetic code.
Another pathogenic ingredient predisposing to ulcer recurrence is the unusual premature skin cell aging brought by hyperglycemia and its derivatives (60-62). Diabetes is a mitochondrial-related disease, and not surprisingly, mitochondrial dysfunction is considered a primary trigger of skin aging and other phenotypic manifestations as impaired healing (62). A variety of mitochondrial and other diabetes disorders as hyperinflammation, high proteolytic activity, defective oxidative phosphorylation, local hypometabolism, excessive ROS generation, and accumulation of AGEs are found in skin fibroblasts and keratinocytes (63, 64) which translate in healing impairment (65). We deem that the persistence of these factors and/or their epigenetic signature may contribute to ulcer recurrence. In line with this notion, magnetic resonance studies confirmed that edema and hypometabolism of lipids and aminoacids, persist during remission time rendering scar tissue vulnerability (66). Decisively, diabetes biochemical derangements are a major and direct skin-aging driving factor, causing cellular dysfunction, and dermal proteins denaturation and decay (67, 68).
Skin fibroblasts are a functionally heterogeneous mesenchymal cell population with a central role in wound repair. These are sensitive cells with a large reserve of plasticity and reprogramming before external clues, which may alter their biological behavior and ultimately the wound healing fate (69). Most importantly, fibroblasts are cells endowed with the ability to retain a memory from their positional location, mechanical, and inflammatory environments, and specially a metabolic memory. Accordingly, fibroblasts can sense intracellular and extracellular metabolic changes in their microenvironment and consequently orchestrate a long-lasting phenotypic response (70). Thus, the society of wound fibroblasts may modify the course of the healing process as the long term fate of the residual scar (33, 59). Short term exposure of cultured, healthy, non-diabetic donor dermal fibroblasts to high glucose burden hampers proliferation, anabolism, and migration signaling pathways and orchestrates senescence (71), thus mirroring the phenotypic pattern detected in diabetic fibroblasts explanted from foot ulcers (59, 72). Hyperglycemia causes apoptosis of dermal fibroblasts, reduces collagen expression, and upregulates RELA/p65 expression, which implicates the onset of pro-inflammatory and pro-degradative profiles (53, 73, 74). Increased repertoire of inflammatory biomarkers is associated with the course of non-healing DFU whereas elevated figures of neutrophil-to-lymphocyte ratio (NLR) show a positive correlation with increased risk of amputation, and ulcer septic complications (75). The presence of endogenous skin-damage predisposing markers was identified by single-cell transcriptome studies of DFU specimens, in which multiple fibroblast cell clusters showed an increased inflammation pattern, changes that were likewise detected in areas of intact skin of diabetic subjects (76). Thus, scar tissue aftermath may depend on fibroblast metabolism, its secretory capability of extracellular proteins, and its control over the quality and duration of the inflammatory reaction (65).
Aside from its critical physiology, keratinocyte is the veteran sentinel cell that initiates the healing cascade after the epidermal integrity is disrupted (77). Like dermal fibroblasts, keratinocytes are also memory cells (78, 79) whose “response to wounding” is seriously affected by high-glucose stress (80). Hyperglycemia introduces alterations of keratinocytes metabolism, adhesion, migration, proliferation, and differentiation (81, 82), all having an epigenetic fundamental through abnormal changes in DNA methylation (83-85). Similarly, the onset of a senescent phenotype by epidermal cells may be a major molecular gear for ulcer recurrence. Hyperglycemia and its chemical derivatives shape an epigenetic landscape in which upregulation and post-translational modifications of p53, p21, and p16 contribute to keratinocytes senescence (17). Hypothetically, significant for ulcer recurrence could be the active expression of matrix metalloproteinase-9 (MMP-9), a type IV collagenase expressed by keratinocytes at the wound's leading edge, which may hinder re-epithelialization when up-regulated by AGEs. MMP-9 levels are elevated by hyperglycemia and glycation products via the upregulation of ten-eleven translocation enzyme 2 (TET2) gene expression. TET2 expression is higher in epidermal cells of diabetic patients than in normal skin, which appeared to be consequent of high levels of α-ketoglutarate. Of note, the levels of α-ketoglutarate correlate with local hypoxia, ischemia, and with poor glycemic control, exemplifying how the local environment and metabolism impacts on wound cells physiology via epigenetic mechanisms (9, 86).
An intrinsic fragility of the epidermal layer may be a predisposing factor for ulcer recurrence. One of the hallmarks of diabetic wounds is the high rate of keratinocytes proliferation versus an unsuccessful differentiation platform. Studies in diabetic mice have demonstrated an abnormal skin differentiation program due to an underlying keratinocyte dysfunction. Human non-diabetic keratinocytes exposed to hyperglycemic stress, and diabetic subjects-derived epidermal cells, exhibit in common a differentiation dysfunction mediated by an overexpressed c-Myc, which blunts differentiation by activating the WNT/β-catenin pathway (87). Previous observations had already indicated that activation of the β -catenin pathway, and an enhanced expression of c-Myc, were implicated in the diabetic torpid re-epithelialization response by disrupting keratinocyte migration and differentiation (88).
We also deem that a successful reciprocal and dynamic communication between epidermal keratinocytes and dermal fibroblasts, is mandatory to ensure scar tissue health and accordingly prevent wound recurrence. This notion is founded on the evidences of signaling crosstalk between these two major cells lineages. Keratinocytes nurture fibroblasts and myofibroblasts activity via the paracrine secretion of vascular endothelial growth factor-A (VEGF-A), transforming growth factor-β1 (TGF-β1), and connective tissue growth factor (CTGF), whereas reciprocally fibroblast-derived TGF-β1 expression, is essential for keratinocytes physiology including migration (89). High glucose levels and AGEs are known to disrupt this dermo-epidermal cells homeostatic circuit by reducing forkhead box O1 transcription factor (FOXO1) expression in keratinocytes, thus hindering keratinocytes ability to produce TGF-β1 (Figure 3) (89). All together these findings converge to highlight: (1) the impact of glycemic control in keratinocytes physiology, (2) the significance of keratinocytes epigenetic imprinting in scar tissue integrity, and (3) the biological significance of a healthful dermo-epidermal axis.
Figure 3.
Remission duration may depend on dermo-epidermal axis homeostasis.
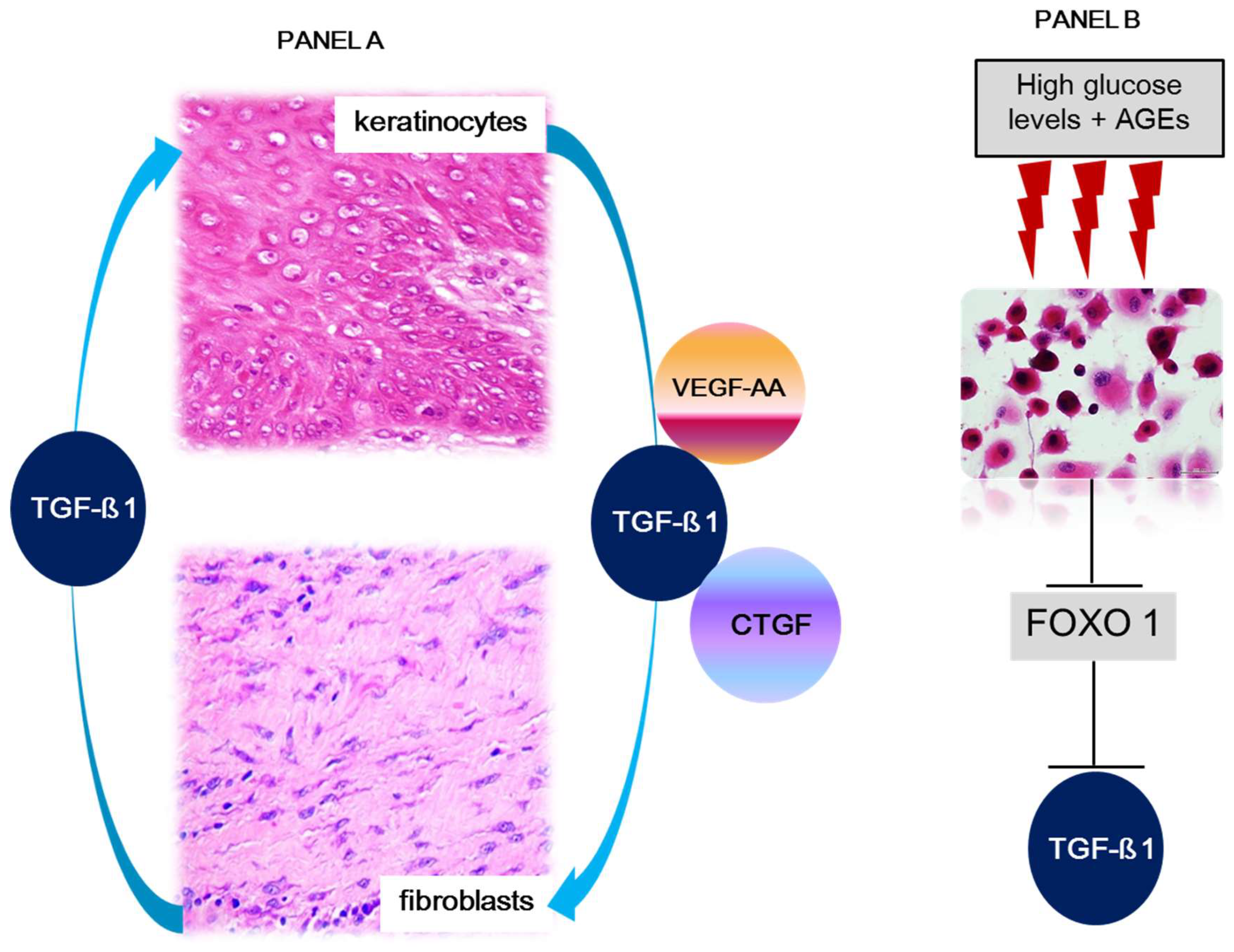
Legend: VEGF-A., vascular endothelial growth factor-A, TGF-β1., transforming growth factor-β1, and CTGF., connective tissue growth factor. FOXO1., forkhead box 1. Panel A: Dermal fibroblasts and epidermal keratinocytes conserve a dynamic and reciprocal crosstalk with the exchange of signalers for both cell populations homeostasis. Under physiological conditions epidermal keratinocytes secrete growth factors involved in fibroblasts physiology, including the synthesis, secretion, and turnover of dermal matrix proteins. The circuit includes the secretion by fibroblasts and utilization by keratinocytes compartment of TGF-β1. In this context, this growth factor appears to participate in keratinocytes stability, migration, turnover, and differentiation. Panel B. In diabetic subjects, and hyperglycemia experimental settings, high glucose levels and AGEs results in significantly reduced TGF-β1 expression by keratinocytes. This event is due to FOXO1 molecular changes which consequently reduces FOXO1 ability to promote TGF-β1 expression.
III- Ulcer recurrences in the clinical arena
Although recurrence rates reports of diabetic foot ulcers appear to broadly differ in the current literature (90), statistical data are alarmingly high despite the variety of healing interventions, and improved multidisciplinary management of the condition (19, 91). The literature on ulcer recurrence quite often misses data on the most specific recurrence time points, and anatomical sites (19); whereas unfortunately, not all the clinical trials examining the healing efficacy of drugs, devices, or management approaches, include information about recurrence rates during a reasonable follow-up period (92). Table 1, summarizes the recurrence data of relevant clinical investigations (clinical trials and meta-analysis) comprising three major groups of recently developed products: recombinant proteins (EGF and PDGF-BB), cellular and/or tissue-based products, and devices (vacuum assisted closure/negative pressure wound therapy). Of a total of 20 articles reviewed, recurrence data were reported in 10 (50%).
According to the data collected, post epithelialization recurrence rates are high, especially those observed in the nation-wide phase III clinical trial in the USA for Regranex/Becaplermin (106). Inversely, EGF treatment based on the intralesional infiltration delivery appears to provide the longest remission times, with the lowest recurrence rates (Table 1). A recent systematic review of randomized controlled trials, investigating different recombinant GFs for the purpose of wound healing, concludes that EGF is the most effective GF to enhance DFU healing (92). Similar conclusion is drawn from another meta-analysis stratified by the types of administration route (intralesional injection and topical administration) in which six studies involving 530 patients were eligible for reviewing (114). It is likely that EGF is the most broadly studied GF in wound healing (115), and interestingly, its healing effects are far more notorious as much the wounds are bigger (96), suggesting that EGF is endowed with a broad therapeutic window. Aside from that, it is likely that the success of the infiltrated EGF in prolonging scar tissue integrity, may reside on its ability to positively impact on the skin cells abnormal epigenetic program, and/or reduce the society of senescent fibroblasts (17). Definitively, durability of ulcer-free time should be regarded as a primary endpoint of major benefit in clinical trials for developing products/treatments (116).
IV- Concluding remarks and future directions
Diabetes is likely the most convincing scenario to illustrate how a trivial exogenous factor such as the level of blood glucose may reshape the native epigenetic program and ultimately build a disease-pathologic memory. The existence of this hyperglycemia-mediated metabolic stress memory explains why prior hyperglycemic exposure is not “forgotten” with time and successive cell generations. On the other hand, diabetes is an exemplary disease in terms of the generation and the progressive accumulation of hyperglycemia-derived cytotoxic products, some of them being cumulative in tissues like the skin. Accordingly, the conjunction of epigenetic and the consequent abnormal cellular behavior, along with the chronic cytotoxicity exerted by the spillover of AGEs and free radicals are biological factors that impair the healing response, and continuously jeopardize scar tissue homeostasis, stability, and viability. It is not surprising therefore that under this environment, societies of senescent and mitosis-refractory cells are found entrenched within the residual scar and the skin of diabetics in general. The fact as stated by Armstrong and co-workers, that recurrence may globally affect up to 40% of the patients on the first year after re-epithelialization, in addition to be distressing may indicate that: (1) the alert sense implicit in the ulcer remission concept has not sensitized enough to patients and wound care providers, (2) glycemia control may remain insufficient in the post-healing period, (3) endogenous, biological drivers remain silently active deteriorating the homeostasis of scar tissue and cells, (4) not all the innovative treatments contemporarily accepted to enhance acute ulcer healing, translate in prolonged remissions and far less in definitive healing, (5) treatments to be considered as effective are called to promote scar tissue resilience and offer a reasonable remission time.
We hold the argument that future innovations for chronic wounds and accordingly prevent ulcer relapses, must ideally entail the ability to target the cell epigenetic core which could erase the chronic hyperglycemic stress memory, reduce the burden of senescent cells, and impose “healthy” re-differentiation programs. Manipulating the diabetic metabolic epigenetic code may indefectibly assist in the control of the all its chronic complications. Of note, however, this therapeutic dream line must be anticipated by the well-deserved glycemic control.
Authors contributions
All the authors enlisted contributed to review and analyze the information of the specialized literature and to write the manuscript.
Funding
No funds were required for this review.
Conflicts of Interest
We declare no conflict of interest.
References
- Cheng, H.T.; Xu, X.; Lim, P.S.; Hung, K.Y. Worldwide Epidemiology of Diabetes-Related End-Stage Renal Disease, 2000-2015. Diabetes Care. 2021, 44, 89–97. [Google Scholar] [CrossRef]
- Liu, J.; Ren, Z.H.; Qiang, H.; Wu, J.; Shen, M.; Zhang, L.; et al. Trends in the incidence of diabetes mellitus: results from the Global Burden of Disease Study 2017 and implications for diabetes mellitus prevention. BMC Public Health. 2020, 20, 1415. [Google Scholar] [CrossRef] [PubMed]
- Mobasseri, M.; Shirmohammadi, M.; Amiri, T.; Vahed, N.; Hosseini Fard, H.; Ghojazadeh, M. Prevalence and incidence of type 1 diabetes in the world: a systematic review and meta-analysis. Health Promot Perspect. 2020, 10, 98–115. [Google Scholar] [CrossRef] [PubMed]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; et al. Pathophysiology of Type 2 Diabetes Mellitus. Int J Mol Sci. 2020, 21. [Google Scholar] [CrossRef]
- Magliano, D.J.; Sacre, J.W.; Harding, J.L.; Gregg, E.W.; Zimmet, P.Z.; Shaw, J.E. Young-onset type 2 diabetes mellitus - implications for morbidity and mortality. Nat Rev Endocrinol. 2020, 16, 321–31. [Google Scholar] [CrossRef]
- Rosengren, A.; Dikaiou, P. Cardiovascular outcomes in type 1 and type 2 diabetes. Diabetologia. 2023, 66, 425–37. [Google Scholar] [CrossRef] [PubMed]
- Mauricio, D.; Alonso, N.; Gratacos, M. Chronic Diabetes Complications: The Need to Move beyond Classical Concepts. Trends Endocrinol Metab. 2020, 31, 287–95. [Google Scholar] [CrossRef]
- Natarajan, R. Epigenetic Mechanisms in Diabetic Vascular Complications and Metabolic Memory: The 2020 Edwin Bierman Award Lecture. Diabetes. 2021, 70, 328–37. [Google Scholar] [CrossRef]
- Deng, J.Y.; Wu, X.Q.; He, W.J.; Liao, X.; Tang, M.; Nie, X.Q. Targeting DNA methylation and demethylation in diabetic foot ulcers. J Adv Res. 2023. [CrossRef]
- Dogruel, H.; Aydemir, M.; Balci, M.K. Management of diabetic foot ulcers and the challenging points: An endocrine view. World J Diabetes. 2022, 13, 27–36. [Google Scholar] [CrossRef]
- Chen, L.; Sun, S.; Gao, Y.; Ran, X. Global mortality of diabetic foot ulcer: A systematic review and meta-analysis of observational studies. Diabetes Obes Metab. 2023, 25, 36–45. [Google Scholar] [CrossRef]
- McDermott, K.; Fang, M.; Boulton, A.J.M.; Selvin, E.; Hicks, C.W. Etiology, Epidemiology, and Disparities in the Burden of Diabetic Foot Ulcers. Diabetes Care. 2023, 46, 209–21. [Google Scholar] [CrossRef]
- Boulton, A.J.M.; Armstrong, D.G.; Londahl, M.; Frykberg, R.G.; Game, F.L.; Edmonds, M.E.; et al. New Evidence-Based Therapies for Complex Diabetic Foot Wounds. Arlington (VA) 2022. [CrossRef] [PubMed]
- Rai, V.; Moellmer, R.; Agrawal, D.K. Clinically relevant experimental rodent models of diabetic foot ulcer. Mol Cell Biochem. 2022, 477, 1239–47. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Rodriguez, N.; Martinez-Jimenez, I.; Garcia-Ojalvo, A.; Mendoza-Mari, Y.; Guillen-Nieto, G.; Armstrong, D.; et al. Wound chronicity, impaired immunity and infection in diabetic patients. MEDICC Review. 2022, 24, 44–58. [Google Scholar] [CrossRef]
- Leal, E.C.; Carvalho, E. Heme Oxygenase-1 as Therapeutic Target for Diabetic Foot Ulcers. Int J Mol Sci. 2022, 23. [Google Scholar] [CrossRef]
- Berlanga-Acosta, J.A.; Guillen-Nieto, G.E.; Rodriguez-Rodriguez, N.; Mendoza-Mari, Y.; Bringas-Vega, M.L.; Berlanga-Saez, J.O.; et al. Cellular Senescence as the Pathogenic Hub of Diabetes-Related Wound Chronicity. Front Endocrinol (Lausanne). 2020, 11, 573032. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, P.; Vasishta, S.; Balakrishnan, A.; Joshi, M. Mechanistic insights into glucose induced vascular epigenetic reprogramming in type 2 diabetes. Life Sciences. 2022, 298, 120490. [Google Scholar] [CrossRef]
- Bouly, M.; Laborne, F.X.; Tourte, C.; Henry, E.; Penfornis, A.; Dardari, D. Post-healing follow-up study of patients in remission for diabetic foot ulcers Pied-REM study. PLoS One. 2022, 17, e0268242. [Google Scholar] [CrossRef]
- Guo, Q.; Ying, G.; Jing, O.; Zhang, Y.; Liu, Y.; Deng, M.; et al. Influencing factors for the recurrence of diabetic foot ulcers: A meta-analysis. Int Wound J. 2023, 20, 1762–75. [Google Scholar] [CrossRef]
- Armstrong, D.G.; Boulton, A.J.M.; Bus, S.A. Diabetic Foot Ulcers and Their Recurrence. N Engl J Med. 2017, 376, 2367–75. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.H.; Li, S.Q.; Kou, Y.; Huang, L.; Yu, T.; Hu, A. Risk factors for the recurrence of diabetic foot ulcers among diabetic patients: a meta-analysis. Int Wound J. 2019, 16, 1373–82. [Google Scholar] [CrossRef] [PubMed]
- Petersen, B.J.; Rothenberg, G.M.; Lakhani, P.J.; Zhou, M.; Linders, D.R.; Bloom, J.D.; et al. Ulcer metastasis? Anatomical locations of recurrence for patients in diabetic foot remission. J Foot Ankle Res. 2020, 13, 1. [Google Scholar] [CrossRef]
- Engberg, S.; Kirketerp-Moller, K.; Ullits Andersen, H.; Rasmussen, A. Incidence and predictors of recurrent and other new diabetic foot ulcers: a retrospective cohort study. Diabet Med. 2019, 36, 1417–23. [Google Scholar] [CrossRef] [PubMed]
- Hicks, C.W.; Canner, J.K.; Mathioudakis, N.; Lippincott, C.; Sherman, R.L.; Abularrage, C.J. Incidence and Risk Factors Associated With Ulcer Recurrence Among Patients With Diabetic Foot Ulcers Treated in a Multidisciplinary Setting. J Surg Res. 2020, 246, 243–50. [Google Scholar] [CrossRef]
- Gazzaruso, C.; Gallotti, P.; Pujia, A.; Montalcini, T.; Giustina, A.; Coppola, A. Predictors of healing, ulcer recurrence and persistence, amputation and mortality in type 2 diabetic patients with diabetic foot: a 10-year retrospective cohort study. Endocrine. 2021, 71, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Pang, L.; Wang, Y.; Zheng, M.; Wang, Q.; Lin, H.; Zhang, L.; et al. Transcriptomic study of high-glucose effects on human skin fibroblast cells. Mol Med Rep. 2016, 13, 2627–34. [Google Scholar] [CrossRef]
- Cugalj Kern, B.; Trebusak Podkrajsek, K.; Kovac, J.; Sket, R.; Jenko Bizjan, B.; Tesovnik, T.; et al. The Role of Epigenetic Modifications in Late Complications in Type 1 Diabetes. Genes (Basel). 2022, 13. [Google Scholar] [CrossRef]
- Ramteke, P.; Deb, A.; Shepal, V.; Bhat, M.K. Hyperglycemia Associated Metabolic and Molecular Alterations in Cancer Risk, Progression, Treatment, and Mortality. Cancers (Basel). 2019, 11. [Google Scholar] [CrossRef]
- Rehman, S.; Aatif, M.; Rafi, Z.; Khan, M.Y.; Shahab, U.; Ahmad, S.; et al. Effect of non-enzymatic glycosylation in the epigenetics of cancer. Semin Cancer Biol. 2022, 83, 543–55. [Google Scholar] [CrossRef]
- Chen, Z.; Miao, F.; Braffett, B.H.; Lachin, J.M.; Zhang, L.; Wu, X.; et al. DNA methylation mediates development of HbA1c-associated complications in type 1 diabetes. Nat Metab. 2020, 2, 744–62. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Stefan-Lifshitz, M.; Tomer, Y. Genetic and environmental factors regulate the type 1 diabetes gene CTSH via differential DNA methylation. J Biol Chem. 2021, 296, 100774. [Google Scholar] [CrossRef]
- Park, L.K.; Maione, A.G.; Smith, A.; Gerami-Naini, B.; Iyer, L.K.; Mooney, D.J.; et al. Genome-wide DNA methylation analysis identifies a metabolic memory profile in patient-derived diabetic foot ulcer fibroblasts. Epigenetics. 2014, 9, 1339–49. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Luan, Y.; Feng, Q.; Chen, X.; Qin, B.; Ren, K.D.; et al. Epigenetics and Beyond: Targeting Histone Methylation to Treat Type 2 Diabetes Mellitus. Front Pharmacol. 2021, 12, 807413. [Google Scholar] [CrossRef] [PubMed]
- Mir, A.R.; Habib, S.; Uddin, M. Recent advances in histone glycation: emerging role in diabetes and cancer. Glycobiology. 2021, 31, 1072–9. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Omans, N.D.; Leicher, R.; Osunsade, A.; Agustinus, A.S.; Finkin-Groner, E.; et al. Reversible histone glycation is associated with disease-related changes in chromatin architecture. Nat Commun. 2019, 10, 1289. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wei, M.; Liu, X.; Xiao, S.; Cai, Y.; Li, F.; et al. The progress, prospects, and challenges of the use of non-coding RNA for diabetic wounds. Mol Ther Nucleic Acids. 2021, 24, 554–78. [Google Scholar] [CrossRef]
- El-Osta, A.; Brasacchio, D.; Yao, D.; Pocai, A.; Jones, P.L.; Roeder, R.G.; et al. Transient high glucose causes persistent epigenetic changes and altered gene expression during subsequent normoglycemia. J Exp Med. 2008, 205, 2409–17. [Google Scholar] [CrossRef]
- Bhamidipati, T.; Kumar, M.; Verma, S.S.; Mohanty, S.K.; Kacar, S.; Reese, D.; et al. Epigenetic basis of diabetic vasculopathy. Front Endocrinol (Lausanne). 2022, 13, 989844. [Google Scholar] [CrossRef]
- Goodarzi, G.; Maniati, M.; Qujeq, D. The role of microRNAs in the healing of diabetic ulcers. Int Wound J. 2019, 16, 621–33. [Google Scholar] [CrossRef]
- Ozdemir, D.; Feinberg, M.W. MicroRNAs in diabetic wound healing: Pathophysiology and therapeutic opportunities. Trends Cardiovasc Med. 2019, 29, 131–7. [Google Scholar] [CrossRef]
- Lou, R.; Chen, J.; Zhou, F.; Wang, C.; Leung, C.H.; Lin, L. Exosome-cargoed microRNAs: Potential therapeutic molecules for diabetic wound healing. Drug Discov Today. 2022, 27, 103323. [Google Scholar] [CrossRef] [PubMed]
- Ross, K. MiR equal than others: MicroRNA enhancement for cutaneous wound healing. J Cell Physiol. 2021, 236, 8050–9. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yang, C.; Wang, X.Y.; Zhou, L.Y.; Lao, G.J.; Liu, D.; et al. MicroRNA-129 and -335 Promote Diabetic Wound Healing by Inhibiting Sp1-Mediated MMP-9 Expression. Diabetes. 2018, 67, 1627–38. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wu, W.; Zhang, L.; Dorset-Martin, W.; Morris, M.W.; Mitchell, M.E.; et al. The role of microRNA-146a in the pathogenesis of the diabetic wound-healing impairment: correction with mesenchymal stem cell treatment. Diabetes. 2012, 61, 2906–12. [Google Scholar] [CrossRef]
- Xu, M.; Li, Y.; Tang, Y.; Zhao, X.; Xie, D.; Chen, M. Increased Expression of miR-155 in Peripheral Blood and Wound Margin Tissue of Type 2 Diabetes Mellitus Patients Associated with Diabetic Foot Ulcer. Diabetes Metab Syndr Obes. 2022, 15, 3415–28. [Google Scholar] [CrossRef]
- Liu, J.; Xu, Y.; Shu, B.; Wang, P.; Tang, J.; Chen, L.; et al. Quantification of the differential expression levels of microRNA-203 in different degrees of diabetic foot. Int J Clin Exp Pathol. 2015, 8, 13416–20. [Google Scholar] [PubMed]
- Wu, T.; Xie, D.; Zhao, X.; Xu, M.; Luo, L.; Deng, D.; et al. Enhanced Expression of miR-34c in Peripheral Plasma Associated with Diabetic Foot Ulcer in Type 2 Diabetes Patients. Diabetes Metab Syndr Obes. 2021, 14, 4263–73. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat Rev Mol Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Kuai, L.; Jiang, J.S.; Li, W.; Li, B.; Yin, S.Y. Long non-coding RNAs in diabetic wound healing: Current research and clinical relevance. Int Wound J. 2022, 19, 583–600. [Google Scholar] [CrossRef]
- Shen, J.; Zhao, X.; Zhong, Y.; Yang, P.; Gao, P.; Wu, X.; et al. Exosomal ncRNAs: The pivotal players in diabetic wound healing. Front Immunol. 2022, 13, 1005307. [Google Scholar] [CrossRef]
- Pirola, L. Epigenetics of Diabetic Microvascular Disease. Microvascular Disease in Diabetes2020. p. 45-57.
- Legiawati, L. The Role of Oxidative Stress, Inflammation, and Advanced Glycation End Product in Skin Manifestations of Diabetes Mellitus. Current Diabetes Reviews. 2022, 18, 87–92. [Google Scholar] [CrossRef]
- Rai, V.; Moellmer, R.; Agrawal, D. Role of fibroblast plasticity and heterogeneity in modulating angiogenesis and healing in the diabetic foot ulcer. Molecular Biology Reports. 2023, 50, 1913–29. [Google Scholar] [CrossRef] [PubMed]
- Liechty, C.; Hu, J.; Zhang, L.; Liechty, K.W.; Xu, J. Role of microRNA-21 and Its Underlying Mechanisms in Inflammatory Responses in Diabetic Wounds. International Journal of Molecular Sciences. 2020, 21, 3328. [Google Scholar] [CrossRef]
- Choi, L.-S.; Ahmed, K.; Kim, Y.-S.; Yim, J.-E. Skin accumulation of advanced glycation end products and cardiovascular risk in Korean patients with type 2 diabetes mellitus. Heliyon. 2022, 8, e09571. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Li, H.; Go, Y.; Chan, X.H.; Huang, Q.; Wu, J. Research Advances on the Damage Mechanism of Skin Glycation and Related Inhibitors. Nutrients. 2022, 14, 4588. [Google Scholar] [CrossRef]
- Deng, L.; Du, C.; Song, P.; Chen, T.; Rui, S.; Armstrong, D.G.; et al. The Role of Oxidative Stress and Antioxidants in Diabetic Wound Healing. Oxidative Medicine and Cellular Longevity. 2021, 2021, 8852759. [Google Scholar] [CrossRef] [PubMed]
- Al-Rikabi, A.H.A.; Tobin, D.J.; Riches-Suman, K.; Thornton, M.J. Dermal fibroblasts cultured from donors with type 2 diabetes mellitus retain an epigenetic memory associated with poor wound healing responses. Sci Rep. 2021, 11, 1474. [Google Scholar] [CrossRef] [PubMed]
- Miwa, S.; Kashyap, S.; Chini, E.; Zglinicki, T. Mitochondrial dysfunction in cell senescence and aging. Journal of Clinical Investigation 2022, 132. [Google Scholar] [CrossRef] [PubMed]
- Pinti, M.V.; Fink, G.K.; Hathaway, Q.A.; Durr, A.J.; Kunovac, A.; Hollander, J.M. Mitochondrial dysfunction in type 2 diabetes mellitus: an organ-based analysis. American Journal of Physiology-Endocrinology and Metabolism. 2019, 316, E268–E85. [Google Scholar] [CrossRef]
- Sreedhar, A.; Aguilera-Aguirre, L.; Singh, K.K. Mitochondria in skin health, aging, and disease. Cell Death & Disease. 2020, 11, 444. [Google Scholar]
- Kashpur, O.; Smith, A.; Gerami-Naini, B.; Maione, A.G.; Calabrese, R.; Tellechea, A.; et al. Differentiation of diabetic foot ulcer-derived induced pluripotent stem cells reveals distinct cellular and tissue phenotypes. FASEB J. 2019, 33, 1262–77. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Li, S.; Bjorklund, M.; Xu, S. Mitochondrial fragmentation and ROS signaling in wound response and repair. Cell Regen. 2022, 11, 38. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Lu, X.; Yang, Y.; Yang, Y.; Li, Y.; Kuai, L.; et al. Microenvironment-Based Diabetic Foot Ulcer Nanomedicine. Adv Sci (Weinh). 2023, 10, e2203308. [Google Scholar] [CrossRef] [PubMed]
- Cabal Mirabal, C.; González Dalmau, E.; Berlanga-Acosta, J.; Zayas, D.; Herrera, L.; Lopez-Saura, P.; et al. Quantitative Studies of the Evolution of Diabetic Foot Lesions under EGF Treatment by Magnetic Resonance Imaging. Journal of Radiology Research and Practice 2014, 2014. [Google Scholar]
- Khalid, M.; Petroianu, G.; Adem, A. Advanced Glycation End Products and Diabetes Mellitus: Mechanisms and Perspectives. Biomolecules. 2022, 12. [Google Scholar] [CrossRef]
- Tatmatsu-Rocha, J.C.; Tim, C.R.; Avo, L.; Bernardes-Filho, R.; Brassolatti, P.; Kido, H.W.; et al. Mitochondrial dynamics (fission and fusion) and collagen production in a rat model of diabetic wound healing treated by photobiomodulation: comparison of 904 nm laser and 850 nm light-emitting diode (LED). J Photochem Photobiol B. 2018, 187, 41–7. [Google Scholar] [CrossRef]
- Sacco, A.M.; Belviso, I.; Romano, V.; Carfora, A.; Schonauer, F.; Nurzynska, D.; et al. Diversity of dermal fibroblasts as major determinant of variability in cell reprogramming. J Cell Mol Med. 2019, 23, 4256–68. [Google Scholar] [CrossRef]
- Kirk, T.; Ahmed, A.; Rognoni, E. Fibroblast Memory in Development, Homeostasis and Disease. Cells. 2021, 10. [Google Scholar] [CrossRef]
- Li, B.; Bian, X.; Hu, W.; Wang, X.; Li, Q.; Wang, F.; et al. Regenerative and protective effects of calcium silicate on senescent fibroblasts induced by high glucose. Wound Repair Regen. 2020, 28, 315–25. [Google Scholar] [CrossRef]
- Phang, S.J.; Arumugam, B.; Kuppusamy, U.R.; Fauzi, M.B.; Looi, M.L. A review of diabetic wound models-Novel insights into diabetic foot ulcer. J Tissue Eng Regen Med. 2021, 15, 1051–68. [Google Scholar] [CrossRef]
- Maitra, S.; Dutta, D. Downregulation of hexose sugar metabolism in diabetes decreases the rate of wound healing. 2020. p. 259-70.
- Soydas, T.; Sayitoglu, M.; Sarac, E.Y.; Cinar, S.; Solakoglu, S.; Tiryaki, T.; et al. Metformin reverses the effects of high glucose on human dermal fibroblasts of aged skin via downregulating RELA/p65 expression. J Physiol Biochem. 2021, 77, 443–50. [Google Scholar] [CrossRef] [PubMed]
- Vujcic, S.; Kotur-Stevuljevic, J.; Vekic, J.; Perovic-Blagojevic, I.; Stefanovic, T.; Ilic-Mijailovic, S.; et al. Oxidative Stress and Inflammatory Biomarkers in Patients with Diabetic Foot. Medicina (Kaunas). 2022, 58. [Google Scholar]
- Theocharidis, G.; Baltzis, D.; Roustit, M.; Tellechea, A.; Dangwal, S.; Khetani, R.S.; et al. Integrated Skin Transcriptomics and Serum Multiplex Assays Reveal Novel Mechanisms of Wound Healing in Diabetic Foot Ulcers. Diabetes. 2020, 69, 2157–69. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.C.; Lan, C.E. The Epidermal Keratinocyte as a Therapeutic Target for Management of Diabetic Wounds. Int J Mol Sci. 2023, 24, 4290. [Google Scholar] [CrossRef] [PubMed]
- Dhariwala, M.O.; Scharschmidt, T.C. Baby's skin bacteria: first impressions are long-lasting. Trends Immunol. 2021, 42, 1088–99. [Google Scholar] [CrossRef]
- Haftek, M. 'Memory' of the stratum corneum: exploration of the epidermis' past. Br J Dermatol. 2014, 171 Suppl 3, 6–9. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Huang, S.M.; Cheng, Y.W.; Yen, M.C.; Hsu, Y.L.; Lan, C.E. Investigation of the keratinocyte transcriptome altered in high-glucose environment: An in-vitro model system for precision medicine. J Dermatol Sci. 2023, 109, 37–46. [Google Scholar] [CrossRef]
- Cibrian, D.; de la Fuente, H.; Sanchez-Madrid, F. Metabolic Pathways That Control Skin Homeostasis and Inflammation. Trends Mol Med. 2020, 26, 975–86. [Google Scholar] [CrossRef]
- Hosseini Mansoub, N. The role of keratinocyte function on the defected diabetic wound healing. Int J Burns Trauma. 2021, 11, 430–41. [Google Scholar] [CrossRef]
- Lan, C.C.; Huang, S.M.; Wu, C.S.; Wu, C.H.; Chen, G.S. High-glucose environment increased thrombospondin-1 expression in keratinocytes via DNA hypomethylation. Transl Res. 2016, 169, 91–101e1-3. [Google Scholar] [CrossRef]
- Li, D.; Kular, L.; Vij, M.; Herter, E.K.; Li, X.; Wang, A.; et al. Human skin long noncoding RNA WAKMAR1 regulates wound healing by enhancing keratinocyte migration. Proc Natl Acad Sci U S A. 2019, 116, 9443–52. [Google Scholar] [CrossRef]
- Zhu, P.; Chen, C.; Wu, D.; Chen, G.; Tan, R.; Ran, J. AGEs-induced MMP-9 activation mediated by Notch1 signaling is involved in impaired wound healing in diabetic rats. Diabetes Res Clin Pract. 2022, 186, 109831. [Google Scholar] [CrossRef]
- Tan, Q.; Wang, W.; Yang, C.; Zhang, J.; Sun, K.; Luo, H.C.; et al. alpha-ketoglutarate is associated with delayed wound healing in diabetes. Clin Endocrinol (Oxf). 2016, 85, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yang, P.; Liu, D.; Gao, M.; Wang, J.; Wang, X.; et al. c-Myc Upregulated by High Glucose Inhibits HaCaT Differentiation by S100A6 Transcriptional Activation. Front Endocrinol (Lausanne). 2021, 12, 676403. [Google Scholar] [CrossRef]
- Stojadinovic, O.; Brem, H.; Vouthounis, C.; Lee, B.; Fallon, J.; Stallcup, M.; et al. Molecular pathogenesis of chronic wounds: the role of beta-catenin and c-myc in the inhibition of epithelialization and wound healing. Am J Pathol. 2005, 167, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Graves, D.T. Keratinocyte Function in Normal and Diabetic Wounds and Modulation by FOXO1. J Diabetes Res. 2020, 2020, 3714704. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.L.; Ding, H.; Miao, W.W.; Mao, C.X.; Zhan, M.Q.; Chen, H.L. Global recurrence rates in diabetic foot ulcers: A systematic review and meta-analysis. Diabetes Metab Res Rev. 2019, 35, e3160. [Google Scholar] [CrossRef]
- Bus, S.A.; van Netten, J.J.; Lavery, L.A.; Monteiro-Soares, M.; Rasmussen, A.; Jubiz, Y.; et al. IWGDF guidance on the prevention of foot ulcers in at-risk patients with diabetes. Diabetes Metab Res Rev. 2016, 32 Suppl 1, 16–24. [Google Scholar] [CrossRef]
- Mahdipour, E.; Sahebkar, A. The Role of Recombinant Proteins and Growth Factors in the Management of Diabetic Foot Ulcers: A Systematic Review of Randomized Controlled Trials. J Diabetes Res. 2020, 2020, 6320514. [Google Scholar] [CrossRef]
- Tsang, M.W.; Wong, W.K.; Hung, C.S.; Lai, K.M.; Tang, W.; Cheung, E.Y.; et al. Human epidermal growth factor enhances healing of diabetic foot ulcers. Diabetes Care. 2003, 26, 1856–61. [Google Scholar] [CrossRef]
- Hong, J.P.; Jung, H.D.; Kim, Y.W. Recombinant human epidermal growth factor (EGF) to enhance healing for diabetic foot ulcers. Ann Plast Surg. 2006, 56, 394–8, discussion 9-400. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Han, S.H.; Hong, J.P.; Han, S.K.; Lee, D.H.; Kim, B.S.; et al. Topical epidermal growth factor spray for the treatment of chronic diabetic foot ulcers: A phase III multicenter, double-blind, randomized, placebo-controlled trial. Diabetes Res Clin Pract. 2018, 142, 335–44. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, V.; Pendsey, S.; Sekar, N.; Murthy, G.S.R. A phase III study to evaluate the safety and efficacy of recombinant human epidermal growth factor (REGEN-D™ 150) in healing diabetic foot ulcers. Wounds. 2006, 18, 186–96. [Google Scholar]
- Tuyet, H.L.; Nguyen Quynh, T.T.; Vo Hoang Minh, H.; Thi Bich, D.N.; Do Dinh, T.; Le Tan, D.; et al. The efficacy and safety of epidermal growth factor in treatment of diabetic foot ulcers: the preliminary results. Int Wound J. 2009, 6, 159–66. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Montequin, J.I.; Valenzuela-Silva, C.M.; Diaz, O.G.; Savigne, W.; Sancho-Soutelo, N.; Rivero-Fernandez, F.; et al. Intra-lesional injections of recombinant human epidermal growth factor promote granulation and healing in advanced diabetic foot ulcers: multicenter, randomised, placebo-controlled, double-blind study. Int Wound J. 2009, 6, 432–43. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Villa, R.; Aguilar-Rebolledo, F.; Lozano-Platonoff, A.; Teran-Soto, J.M.; Fabian-Victoriano, M.R.; Kresch-Tronik, N.S.; et al. Response to Uzun G et al. Wound Repair Regen. 2014, 22, 767. [Google Scholar] [CrossRef]
- Bartın, M.; Okut, G. The effect of human recombinant epidermal growth factor in the treatment of diabetic foot ulcers. Journal of Clinical and Investigative Surgery. 2022, 7, 38–42. [Google Scholar] [CrossRef]
- Yera-Alos, I.B.; Alonso-Carbonell, L.; Valenzuela-Silva, C.M.; Tuero-Iglesias, A.D.; Moreira-Martinez, M.; Marrero-Rodriguez, I.; et al. Active post-marketing surveillance of the intralesional administration of human recombinant epidermal growth factor in diabetic foot ulcers. BMC Pharmacol Toxicol. 2013, 14, 44. [Google Scholar] [CrossRef]
- López-Saura, P.A.; Yera-Alos, I.B.; Valenzuela-Silva, C.M.; González-Díaz, O.; del Río-Martín, A.; Berlanga-Acosta, J.; et al. Medical Practice Confirms Clinical Trial Results of the Use of Intralesional Human Recombinant Epidermal Growth Factor in Advanced Diabetic Foot Ulcers. Advances in Pharmacoepidemiology & Drug Safety 2013, 25. [Google Scholar]
- Kahraman, M.; Misir, A.; Kizkapan, T.B.; Ozcamdalli, M.; Uzun, E.; Mutlu, M. The Long-Term Outcomes Following the Application of Intralesional Epidermal Growth Factor in Patients With Diabetic Foot Ulcers. J Foot Ankle Surg. 2019, 58, 282–7. [Google Scholar] [CrossRef] [PubMed]
- Embil, J.M.; Papp, K.; Sibbald, G.; Tousignant, J.; Smiell, J.M.; Wong, B.; et al. Recombinant human platelet-derived growth factor-BB (becaplermin) for healing chronic lower extremity diabetic ulcers: an open-label clinical evaluation of efficacy. Wound Repair Regen. 2000, 8, 162–8. [Google Scholar] [CrossRef] [PubMed]
- Smiell, J.M.; Wieman, T.J.; Steed, D.L.; Perry, B.H.; Sampson, A.R.; Schwab, B.H. Efficacy and safety of becaplermin (recombinant human platelet-derived growth factor-BB) in patients with nonhealing, lower extremity diabetic ulcers: a combined analysis of four randomized studies. Wound Repair Regen. 1999, 7, 335–46. [Google Scholar] [CrossRef]
- Wieman, T.J.; Smiell, J.M.; Su, Y. Efficacy and safety of a topical gel formulation of recombinant human platelet-derived growth factor-BB (becaplermin) in patients with chronic neuropathic diabetic ulcers. A phase III randomized placebo-controlled double-blind study. Diabetes Care. 1998, 21, 822–7. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Hernandez, M.A.; Kirkpatrick, V.E.; Liang, L.J.; Nouvong, A.L.; Gordon, I.I. Topical platelet-derived growth factor vs placebo therapy of diabetic foot ulcers offloaded with windowed casts: a randomized, controlled trial. Wounds. 2015, 27, 83–91. [Google Scholar]
- Veves, A.; Falanga, V.; Armstrong, D.G.; Sabolinski, M.L.; Apligraf Diabetic Foot Ulcer, S. Graftskin, a human skin equivalent, is effective in the management of noninfected neuropathic diabetic foot ulcers: a prospective randomized multicenter clinical trial. Diabetes Care. 2001, 24, 290–5. [Google Scholar] [CrossRef]
- Marston, W.A.; Hanft, J.; Norwood, P.; Pollak, R.; Dermagraft Diabetic Foot Ulcer Study, G. The efficacy and safety of Dermagraft in improving the healing of chronic diabetic foot ulcers: results of a prospective randomized trial. Diabetes Care. 2003, 26, 1701–5. [Google Scholar] [CrossRef]
- Zelen, C.M.; Serena, T.E.; Gould, L.; Le, L.; Carter, M.J.; Keller, J.; et al. Treatment of chronic diabetic lower extremity ulcers with advanced therapies: a prospective, randomised, controlled, multi-centre comparative study examining clinical efficacy and cost. Int Wound J. 2016, 13, 272–82. [Google Scholar] [CrossRef]
- Zelen, C.M.; Serena, T.E.; Fetterolf, D.E. Dehydrated human amnion/chorion membrane allografts in patients with chronic diabetic foot ulcers: A long-term follow-up study. Wound Medicine. 2014, 4, 1–4. [Google Scholar] [CrossRef]
- Blume, P.A.; Walters, J.; Payne, W.; Ayala, J.; Lantis, J. Comparison of negative pressure wound therapy using vacuum-assisted closure with advanced moist wound therapy in the treatment of diabetic foot ulcers: a multicenter randomized controlled trial. Diabetes Care. 2008, 31, 631–6. [Google Scholar] [CrossRef]
- Armstrong, D.G.; Lavery, L.A.; Diabetic Foot Study, C. Negative pressure wound therapy after partial diabetic foot amputation: a multicentre, randomised controlled trial. Lancet. 2005, 366, 1704–10. [Google Scholar] [CrossRef] [PubMed]
- Bui, T.Q.; Bui, Q.V.P.; Nemeth, D.; Hegyi, P.; Szakacs, Z.; Rumbus, Z.; et al. Epidermal Growth Factor is Effective in the Treatment of Diabetic Foot Ulcers: Meta-Analysis and Systematic Review. Int J Environ Res Public Health. 2019, 16. [Google Scholar] [CrossRef] [PubMed]
- Berlanga-Acosta, J.; fernandez montequin, J.; Santana, H.; Pérez, C.; Savigne, W.; Mendoza-Mari, Y.; et al. A narrative review on Epidermal Growth Factor (EGF) intralesional infiltrations for diabetic complex wounds: The rational of an innovative delivery route. Vascular Diseases and Therapeutics. 2022, 7, 1–9. [Google Scholar] [CrossRef]
- Al-Jalodi, O.; Kupcella, M.; Breisinger, K.; Serena, T.E. A multicenter clinical trial evaluating the durability of diabetic foot ulcer healing in ulcers treated with topical oxygen and standard of care versus standard of care alone 1 year post healing. Int Wound J. 2022, 19, 1838–42. [Google Scholar] [CrossRef] [PubMed]

Table 1.
Post-healing recurrence rates reported in major clinical studies of biologics, cell and/or tissue-based products, and VAC.
Table 1.
Post-healing recurrence rates reported in major clinical studies of biologics, cell and/or tissue-based products, and VAC.
| Reference | Major outcome | Administration route | Follow up period | Recurrences |
|---|---|---|---|---|
| Recombinant Epidermal Growth Factor (EGF) | ||||
| Tsang MW et al. 2003 (93) | 20 of 21 diabetic foot ulcers healed with daily application of 0.04% (wt/wt) hEGF for 12 weeks | Topical | 6 months | ND |
| Hong JP et al. 2006 (94) | The study suggests that topical treatment with EGF combined with advanced dressing may have positive effects in promoting healing | Topical | 6 months | No recurrences were observed in EGF group |
| Park KH et al. 2018 (95) | The phase III study supports the efficacy and safety of spray-applied EGF treatment for DFUs, by significantly increasing healing velocity and decreasing time to complete healing | Topical | ND | ND |
| Viswanathan V et al. 2006 (96) | It took 13 weeks for ulcers to heal in the control group versus 9 weeks in the test group. In the test group, 90% of ulcers healed in 15 weeks compared with 22 weeks in the control group | Topical | ND | ND |
| Tuyet HL et al. 2009 (97) | Easyef (topical EGF spray) has positive effects on healing of moderate-to-severe foot ulcers and demonstrated being safe to diabetic patients | Topical | ND | ND |
| Fernández-Montequin J et al. 2009 (98) | Locally infiltrated EGF at 75 µg enhanced granulation tissue growth and wound closure | Intralesional injection | 12 months | No recurrences reported for EGF groups |
| Gomez-Villa R et al. 2014 (99) | Patients with DFU who received intralesional rhEGF application resulted in complete healing, larger epithelialization of the wound bed, and reduction of ulcer area | Intralesional injection | ND | ND |
| Bartın M & Okut G 2022 (100) | The study shows that intralesional administration of EGF in T2DM can prevent amputations in DFU and also accelerate wound healing | Intralesional injection | 6 months | Two cases in the group receiving EGF |
| Yera-Alos IB et al. 2013 (101) | Post-marketing study including 1788 patients treated with intraulcer injected EGF. 1835 DFU (81% Wagner’s grades 3 or 4; 43% ischemic) were treated. Re-epithelization was documented in 61% of the 1659 followed-up cases | Intralesional injection | 14 months | 5% / year |
| López-Saura PA et al., 2013 (102) | A review summarizing the clinical information about intralesional use of EGF for high grade DFU, in more than 2000 subjects. It confirms the results of the clinical trials, with 75% probability of complete granulation response, 61% healing, and a 16% absolute, and 71% relative reduction of amputation risk | Intralesional injection | 12 months | The frequency of relapses at any momentwas significantly lower (p<0.001) in patients that received rhEGF: 2.0% person-years of follow-up |
| Kahraman M et al. 2019 (103) | Study aimed to investigate the long-term outcomes after intralesional epidermal growth factor injections in the treatment of 34 diabetic patients with foot ulcers. | Intralesional injection | 60 months | Of 29 patients involved in the 5-years follow up, 27 were ulcer free |
| Regranex or Becaplermin (rh-PDGF-BB) | ||||
| Embil JM et al. 2000 (104) | Results of the study further confirm the efficacy and safety of becaplermin gel for the treatment of lower extremity diabetic ulcers | Topical | 6 months | 21% of recurrence in Becaplermin- treated patients |
| Smiell JM et al. 1999 (105) | Treatment with becaplermin gel at a dose of 100 μg/g once daily, in conjunction with good ulcer care, is effective in patients with full thickness lower extremity diabetic ulcers | Topical | 3 months | ND |
| Wieman TJ et al. 1998 (106) | Becaplermin gel 100 μg/g significantly increased the incidence of complete wound closure by 43% (50 vs. 35%, P=0.007) and decreased the time to achieve complete wound closure by 32% | Topical | 3 months | The incidence of ulcer recurrence was ≈30% in all treatment groups |
| Ma C et al. 2015 (107) | Topical platelet derived growth factor does not appear to significantly improve healing of Wagner grade I diabetic foot ulcers | Topical | 6 months | No difference was observed between groups in recurrence followed by amputation |
| Cellular and tissue-based products | ||||
| Veves A et al. 2001 (108) | At the 12-week follow-up visit, 63 (56%) Graftskin-treated patients achieved complete wound healing compared with 36 (38%) in the control group (P=0.0042). | Topical –bioengineered skin substitutes | 6 months | The incidence of ulcer recurrence was similar for Graftskin and control groups |
| Marston WA et al. 2003 (109) | Patients with chronic diabetic foot ulcers of >6 weeks duration experienced a significant clinical benefit when treated with Dermagraft versus patients treated with conventional therapy alone. | Topical-bioengineered skin substitutes | ND | ND |
| Zelen CM et al. 2016 (110) | EpiFix® (dehydrated human amnion/chorion membrane) is superior to standard wound care SWC and Apligraf®, in achieving complete wound closure within 4–6 weeks. | Topical –bioengineered skin substitutes | ND | ND |
| Zelen CM et al. 2014 (111) | Study addressed to evaluate recurrence rates of DFU healed with use of dehydrated human amnion/chorion membrane (EpiFix) in 18 available subjects with healed DFU. Wound median size of 1.7 cm2. | Topical –bioengineered skin substitutes | 9-12 months | 17 wounds remained healed |
| Vacuum Assisted Closure (VAC)/ Negative Pressure Wound Therapy (NPWT) | ||||
| Blume P et al. 2008 (112) | A greater proportion of foot ulcers achieved complete ulcer closure with NPWT (73 of 169, 43.2%) than with advanced moist therapy within the 112-day active treatment | Topical- sub atmospheric pressure over the wound area | ND | ND |
| Armstrong DG et al. 2005 (113) | More patients healed in the NPWT group than in the control group (43 [56%] vs 33 [39%], p=0·040) | Topical- sub atmospheric pressure over the wound area | ND | ND |
ND- not defined.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



