
Submitted:
17 April 2023
Posted:
18 April 2023
You are already at the latest version
Abstract
Despite exhaustive studies, researchers have made little progress in the field of adoptive cellular therapies for relapsed-refractory acute myeloid leukemia (AML), unlike the notable uptake for B cell malignancies. Various single antigen targeting chimeric antigen receptor (CAR) T cell Phase I trials have been established worldwide and have recruited approximately 100 patients. The high heterogeneity at the genetic and molecular levels within and between AML patients resembles a black hole: a great gravitational field that sucks in everything, considering only around 30% of patients show a response but with consequential off-tumor effects. It is obvious that a new point of view is needed to achieve more promising results. This review first introduces the unique therapeutic challenges of not only CAR T cells but also other adoptive cellular therapies in AML. Next, recent single cell sequencing data for AML to assess somatically acquired alterations at the DNA, epigenetic, RNA and protein levels are discussed to give a perspective on cellular heterogeneity, intercellular hierarchies, and the cellular ecosystem. Finally, promising novel strategies are summarized, including more sophisticated next-generation CAR T, TCR-T and CAR-NK therapies; approaches to tailor the microenvironment and target neoantigens; and allogeneic approaches.
Keywords:
Acute Myeloid Leukemia
; cellular therapies
; chimeric antigen T cells
; T cell receptor T cells
; CAR-NK cells
; RNAseq
1. Introduction
Since early studies in the eighteenth century on the development of the first vaccines, researchers have long attempted to eliminate tumors by harnessing the immune system. One strategy, adoptive cell therapies, uses T cells that can recognize tumor antigens through tumor-specific receptors. Chimeric antigen receptor T cells (CAR-T cells) have revolutionized the treatment algorithms of patients with lymphoma, acute lymphoblastic leukemia and multiple myeloma starting in the 1980s [1]. They have shown dramatic success in the clinic, improving survival and quality of life for patients that would otherwise reach end of care with conventional therapies. Currently, six CAR-T cell products are approved by the United States Food and Drug Administration (FDA), and approvals are expanding to Europe and many other countries around the world.
There are distinct considerations in acute myeloid leukemia (AML), the most common acute leukemia in adults. AML is an aggressive blood cancer characterized by collection of immature cells of myeloid lineage that exhibit partial or complete arrest of maturation. The heterogeneity and intrinsic variability of the tumor make patient responses hard to predict, and around 75% of patients ultimately relapse. Treatment resistance (10-40%) and relapse remain the major consequences during disease follow-up, highlighting the urgent need for novel therapeutic approaches. Allogeneic hematopoietic stem cell transplantation (allo-HSCT) is the only curative option, but many patients are not suitable candidates [2].
Many in vitro and in vivo studies have shown that CAR T cells against surface proteins, such as CD33, CD123, CLL-1, CD13, CD7, NKG2D ligand, CD38, CD70 and TIM3, effectively eradicate AML cells [3]. However, clinical trials are limited, with not-very-promising response rates accompanied by high ‘on-target off-tumor’ toxicity due to frequent expression on healthy hematopoietic stem cells or progenitors as well as other tissues. Major clinical trials and case reports are shown in Table 1, and ongoing trials were summarized in a review by Marofi et al [4]. The data in the CAR T field for AML so far resembles a black hole that none of the strategies have the power to survive. This review focuses on the main questions regarding challenges for adoptive immune therapies in the setting of relapsed/refractory AML and the novel approaches to overcome them.

Table 1.
CAR T cell Clinical Trials and Case Reports in AML (ALT: Alanine aminotransferase, AST: Aspartate aminotransferase, CLL-1: C-type lectin like molecule-1, CR: Complete Response, CRS: Cytokine Release Syndrome, Cy: Cyclophosphamide, Flu: Fludarabine, ICANS: Immune-effector cell associated neurotoxicity syndrome, MRD: Minimal Residual Disease, PR: Partial Response, SD: Stable Disease).
Table 1.
CAR T cell Clinical Trials and Case Reports in AML (ALT: Alanine aminotransferase, AST: Aspartate aminotransferase, CLL-1: C-type lectin like molecule-1, CR: Complete Response, CRS: Cytokine Release Syndrome, Cy: Cyclophosphamide, Flu: Fludarabine, ICANS: Immune-effector cell associated neurotoxicity syndrome, MRD: Minimal Residual Disease, PR: Partial Response, SD: Stable Disease).
| Study | Target | CAR T cell/lymphodepletion | Number of Patients | Response | Safety | Reference |
|---|---|---|---|---|---|---|
| Phase I/II | CLL-1 | 0.35-1x10^6.kg/anti-CLL-1-CD8-41BB/ Cy+Flu |
8 | 4/8 morphological leukemia-free, MRD (-) 1 morphological leukemia-free, MRD+, 1 CR with incomplete hematologic recovery MRD(+), 1 PR, 1 SD |
CRS: 5 grade 1, 3 grade 2 | 150 |
| Phase I | CD123 | Dose escalation 50x10^6-200x10^6/anti-CD123-IgG4-CD28/Cy+Flu | 6 | 4/6 CR 2 reduced blast |
CRS: 4 grade 1, 1 grade 2; 1 adenoviral pneumonia requiring intubation; and 1 grade 3 rash due to drug hypersensitivity | 151 |
| Phase I | CD33 | CD33-4-1BB/0.3x10^6/kg | 3 | 0/3 response | 2 CRS; 1 ICANS. A grade 3 tumor lysis syndrome-acute kidney injury, grade 2 mucositis, grade 1 tachycardia for 1 patient; and a second patient experienced grade 2 intermittent orthostatic hypotension, grade 2 increased bilirubin and grade 3 increased ALT and AST | 152 |
| Case report | CD33 | CD33-4-1BB/ 1.12x10^9 | 1 | Disease progression at week 9 | Grade 4 chills and a high fever, pancytopenia | 153 |
| Phase I | CD38 | NA | 6 | 66.7% of patients (4/6) CR (including 1 with CR and 3 with CR with incomplete count recovery (CRi)) and full donor chimerism) | Five patients presented mild CRS (Grade I–II), and only one experienced grade III hepatotoxicity with elevated serum transaminase and bilirubin levels | 154 |
| Phase I | LeY | Anti-LeY-CD28/Flu-Cy | 4 | In the patient with active leukemia, a temporary reduction in peripheral blood blast cells was observed. One other patient achieved a cytogenetic remission, while the other two patients had SD | One patient (patient 2) had a transient grade 2 neutropenia | 155 |
2. Challenges in Adoptive T cell Therapy for Acute Myeloid Leukemia
2.1. AML is highly heterogenous:
Normal hematopoietic stem cells give rise to mature cells of the myeloid, lymphoid, and erythroid/megakaryocyte lineages. Single cell RNA sequencing (scRNA-seq) analyses have shown that normal hematopoietic stem cell (HSC) commitment proceeds through a series of increasingly lineage-committed progenitor states [5]. AML consists of leukemia stem cells (LSCs) and differentiated cells. LSCs sustain the disease and display self-renewal, quiescence, and therapy resistance. Differentiated AML cells that lack stem cell characteristics affect tumor biology through pathologic effects on the tumor microenvironment [6].
AML is a highly heterogeneous disease between patients due to the presence of specific chromosomal abnormalities, gene mutations or gene fusions. 17 genetic subtypes have been discovered so far in ELN risk stratification, but the number of molecular entities may increase over time [7]. Not all gene expression subtypes correlate well with the underlying disease-driven gene fusions or mutations, in contrary to acute lymphoblastic leukemia (ALL), which is characterized by distinct gene expression subtypes [8]. Recurrent somatic mutations are categorized by their biological roles, such as signaling and kinase pathway genes. Mutation in FLT3 (encodes a receptor tyrosine kinase) and KRAS/NRAS (encode a small GTPase) lead to uncontrolled cell growth and proliferation whereas mutation in JAK2 (encodes tyrosine kinase) promotes leukemogenesis. There are epigenetic modifiers encoding a DNA methyltransferase enzyme (DNMT3A), isocitrate dehydrogenase enzymes (IDH1/2) and a DNA demethylase (TET2). Mutation in ASXL1 (a polycomb repressive complex proteins implicated in chromatin) can lead to the alteration of DNA methylation patterns, dysregulation of gene expression and altered hematopoietic differentiation. Dysregulation of transcription factors (e.g. CEBPA, RUNX1, MLL, EVI1, etc.), RNA splicing factors (e.g. SRSF2 which encodes a serine and arginine-rich splicing factor, U2AF1, SF3B1, ZRSR2, etc.), tumor suppressors (TP53), nucleophosmin (NPM1), cohesin complex genes (e.g. RAD21, STAG2, SMC1A, SM3 etc.) can lead to impaired differentiation and uncontrolled cell growth [9]. The initiating leukemogenic NPM1, TET2 and SMC1A mutations emerge in self-renewing cells that Jan et al identified as highly purified HSCs, namely ‘pre-leukemic HSCs’ [10]. The same mutations may lead to highly variable preleukemic burden in different patients [11]. In a study conducted by Stengel and colleagues, 37% of 572 AML cases had fusion events, and 41% of these fusions were detected in cases with TP53 alterations or complex karyotypes [12]. A subset of germline mutations have also been shown in myeloid neoplasms: mutations in GATA 1 gene for Down syndrome patients, DNA damage repair for Fanconi anemia, telomere maintenance genes (DKC1, TERC, TIF2) for Dyskeratosis congenita, RUNX1, ANKFRD26 and ETV6 for platelet disorders, and ANKRD26 for abnormal thrombopoiesis. Epigenetic modifications such as histone modifications, DNA methylation and post-transcriptional regulation of mRNAs by noncoding RNAs have additional roles in the pathogenic heterogeneity [13].
Furthermore, a crucial problem in patients with relapse or treatment resistance is intra-tumoral heterogeneity, formed with different subclones of leukemia cells, with distinct genetic and epigenetic features coexisting within a single patient [14,15]. Highly heterogeneous LSCs have variable drug sensitivity. Some LSCs acquire quiescence, which plays a major role in drug resistance profiles [16,17]. Recent advances in genomic, transcriptomic, epigenomic and proteomic data have helped to elucidate the biological differences between pre-treatment AML cells and equivalents in relapse. During disease progression in t(8;21) positive AML patients, heterogenous cell populations show their own clonal characteristics and obtain cloned components [18]. In a single-cell analysis, Li et al found that proliferating stem/progenitor-like cells were reprogrammed to a quiescent stem-like expression pattern in primary refractory AML by upregulation of CD52 and LGALS1 expression [19]. Stratmann and colleagues used mass-spectrometry based in-depth proteomics to show that at relapse, energy metabolism is reprogrammed by enrichment of mitochondrial ribosomal proteins and subunits of respiratory chain complex as well as higher levels of granzymes and lower levels of the anti-inflammatory protein CR1/CD35 [20].
2.2. There is no ideal surface antigen to target:
CAR T cells can bind to cell surface molecules without requiring any antigen processing or HLA expression [21]. The choice of which surface antigen to target is the critical step in manufacturing. The most important feature of an ideal target is unique and high expression profile on tumor cells, above the detection and activation threshold for CAR T cells, as well as tolerable or no expression on healthy tissues to prevent toxicity. CD19 is now a widely accepted B lineage target of lymphoma and leukemia that is expressed in all tumor cells and absent in normal HSCs as well as all normal tissues [22]. Despite many discoveries in the immunopathology of AML, a single AML-specific target has still remained elusive. Various surface proteins have been reported as potential targets, such as CD123, CLL-1, CD33, CD44, CD96, CD47, CD23, TIM3, CD7 and FLT3 [23,24,25,26]. However, the considerable risk of on-target/off-tumor activity needs to be addressed. Most of the surface antigens on AML blasts are co-expressed by other healthy tissues, mature myeloid cells, and HSCs, raising concerns about prolonged myelosuppression (Table 2). In some Phase I trials, substantial toxicities and deaths have already reported [27,28].
Table 2.
Characteristics of Various AML Target Molecules (ADP: Adenosine Diphosphate, CLL-1: C-type lectin like molecule-1, CTL: Cytotoxic T Cell, DC: Dendritic Cell, HSC: Hematopoietic Stem Cell, LSC: Leukemic Stem Cell, NK: Natural Killer, TNF: Tumor Necrosis Factor).
Table 2.
Characteristics of Various AML Target Molecules (ADP: Adenosine Diphosphate, CLL-1: C-type lectin like molecule-1, CTL: Cytotoxic T Cell, DC: Dendritic Cell, HSC: Hematopoietic Stem Cell, LSC: Leukemic Stem Cell, NK: Natural Killer, TNF: Tumor Necrosis Factor).
| Target Antigen | Function | Expression on normal cells | Expression on HSCs | Expression on LSCs |
|---|---|---|---|---|
| CLL-1 | Glycoprotein, Transmembrane receptor | Myeloid, lung, epithelial cells | - | + |
| CD 33 | SIGLEC family protein, Transmembrane receptor | Progenitor, myeloid, kuppffer cells | + | + |
| CD 7 | Ig superfamily/Glycoprotein, B and T cell lymphoid development, transmembrane protein | T, NK cells and myeloid progenitor | - | + |
| FLT3 | Type III cytokine receptor, Tyrosine kinase receptor | Neurons, testis | + | + |
| CD 38 | Glycoprotein, Cyclic ADP ribose hydroxylase | B, T, NK cells | - | + |
| CD 123 | Type I cytokine receptor of IL-3, IL3 receptor a subunit | Myeloid progenitors, DC and basophils | + | + |
| CD 44v6 | Glycoprotein, Transmembrane receptor | Keratinocytes | - | + |
| LeY | Glycosphingolipid, Blood group Ag | Intestinal epithelial cells | + | + |
| NKG2D | C-type lectin-like receptor protein, Activator receptor | NK, NKT, Tαδ, Th, and CTL | - | + |
| CD 70 | Glycoprotein from the TNF family, Transmembrane receptor | T and B cells | - | + |
| CD 96 | Member of immunoglobulin superfamily, adhesion of activated T and NK cells | T cells and NK cells | - | + |
CD33 is a common transmembrane protein of the sialic acid-binding immunoglobulin-like lectin (SIGLEC) family found on normal progenitor cells, myeloid cells, monocytes, tissue-resident macrophages and more than 90% of leukemic blasts [29]. Severe pancytopenia and cytokine release syndrome (CRS) were reported to be the main adverse effects after administration in a Phase 1 trial (30). CD44V6 CAR T has shown promising preclinical results however monocytopenia was observed due to shared expression of CD44v6 in circulating monocytes [31]. CD123, which has the IL-3 receptor subunit, is overexpressed on LSCs, AML blasts, and early hematopoietic cells such as hematopoietic stem/progenitor cells that induce myeloablation [32]. A recent established target, folate receptor 1 (FOLR1), overexpressed in AML, was shown to have no impact on normal HSPCs in C/G positive pediatric acute megakaryoblastic leukemia, but it is expressed in various healthy tissues (kidney, intestine, lung, retina, placenta and choroid plexus) and severe lung toxicity has already been demonstrated in studies with a T cell bispecific antibody against FOLR1 in nonhuman primates [33].
2.3. Interactions in the Tumor Microenvironment:
LSCs reside in a specialized niche that promotes their survival and chemoresistance, through which they can alter their microenvironment [34]. LSCs secrete pro-angiogenic VEGF and interleukins to stimulate angiogenesis to provide additional nutrients, oxygen and growth factors and to promote proliferation [35,36]. AML blasts support a low-arginine microenvironment [37]. AML LSCs induce the expression of a growth-arresting protein, GAS6, in BM stromal cells [38]. AML cells promote the expression of immunomodulatory factors that impair cytotoxic T lymphocyte (CTL) activation in the tumor microenvironment [39]; these include programmed death receptor (PD-1), transforming growth factor β (TGF β), arginase II, prostaglandin E2 (PGE2), cytotoxic T-lymphocyte associated protein 4 (CTLA-4), lymphocyte activation gene 3 (LAG3), and T cell immunoglobulin and mucin-containing-3 (TIM3) on T cells [40]. Furthermore, leukemia cells modulate the NK cell receptor repertoire that inhibits NK cell activity [41].
The interactions are reciprocal: niche cells also foster LSC growth. In vitro 3D bone marrow microenvironment models have elucidated these interactions [42]. The AML microenvironment contains various cell types, including myeloid-derived suppressor cells (MDSCs), regulatory T cells (Tregs), macrophages, and dendritic cells, that suppress T-cell activity. The high expression of Indoleamine 2,3-dioxygenase (IDO) has been reported to promote Treg conversion and enhance the immunosuppressive ability [43]. Treg cells was shown to express abnormally high levels of CD39 and the increase of CD73 has been associated with poor prognosis (E1). Decrease in CXCL12 expression in BM stromal cells triggers proliferation of AML cells [44]. Osteoblasts in the BM produce WNT ligands to promote leukemia cell survival [45]. AML blasts induce monocytes to secrete pro-inflammatory cytokines including tumor necrosis factor-α (TNF-α), IL1β, and IL6, and the anti-inflammatory cytokine IL10 [46].
Assessment at scRNA-seq, cellular indexing of transcriptomes and epitopes by sequencing (CITE-seq) or single-cell assay for transposase-accessible chromatin (ATAC) sequencing has paved the way to study cellular heterogeneity, infer cellular hierarchies and obtain insights into cellular ecosystem of malignant and normal cells. Galen et al showed that primitive AML cells had dysregulated transcriptional programs with co-expression of stemness and myeloid priming genes and that differentiated monocyte-like AML cells suppressed T cell activity in vitro by immunomodulatory genes [47]. Furthermore, recent advances in syngeneic models with a fully functional immune system provided opportunity to evaluate the important aspects of cancer disease progression and interactions between tumor microenvironment [48,49].
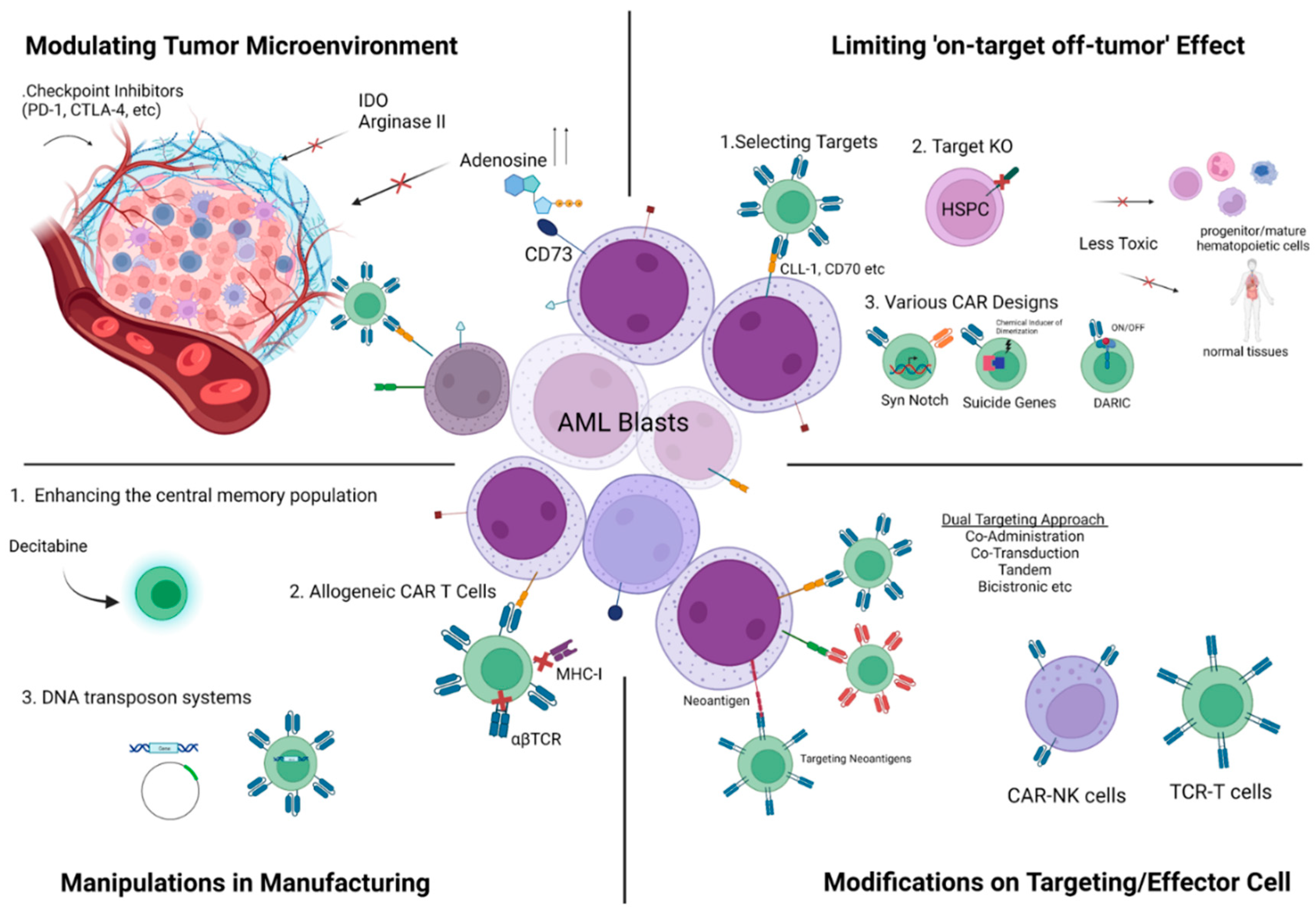
3. Promising Strategies to Overcome Challenges
3.1. Safer targets with less ‘on-target off-tumor’ effect:
Some potential targets appear more reliable in terms of off-tumor toxicity, especially on normal hematopoietic cells, but none of them have proven to be an ideal target in AML. C-type lectin like molecule-1 (CLL-1) is a type II transmembrane glycoprotein overexpressed in over 90% of AML patients on AML blasts, LSCs and differentiated myeloid cells absent in normal CD34+CD38- hematopoietic stem cells. Tashiro and colleagues reported that CLL-1 CAR T cells can eliminate mature normal myeloid cells but spare healthy HSCs in vitro [50]. The CAR T cells targeting CLL-1 were optimized and demonstrated efficient cytotoxicity in vitro and in vivo [51]. One step ahead, when CLL-1 CAR T cells were transduced with a second vector encoding soluble IL 15, a cytokine that promote the survival and proliferation of memory T cells. The expansion and the maintenance of less differentiated phenotype was detected. In xenograft models of AML, addition of IL15 induced the production of Tumor Necrosis Factor-alpha (TNFα), likely through the activation of JAK-STAT pathway. The production of TNF-a contributed to the development of cytokine release syndrome (CRS) and this controlled with the related antibody blockage [51]. In fact, the TNFα pathway resembles a double-edged sword in immune regulation. The suitable modifications in TNFα signaling may enable to enhance activity of CAR [52].
CD70, a ligand for CD27 identified as a type II transmembrane glycoprotein, was reported to be expressed on AML bulk cells and leukemic stem cells but not on normal hematopoietic stem cells. It showed promising anti-tumor effect without toxicity on healthy HSCs [53]. Later, Leick and colleagues designed a panel of CD8 hinge and transmembrane modified CD70 CAR T cells that are less prone to cleavage and have enhanced binding avidity, leading to more potent activity and expansion [54]. CD7 is a transmembrane glycoprotein that plays a co-stimulatory role in B and T cell lymphogenesis expressed by T cells, NK cells, myeloid progenitors and leukemic cells but not by healthy myeloid cells [55]. The obstacle of shared expression on T cells, which causes T cell fratricide, can be overcome by removing the CD7 gene by CRISPR/Cas9, as shown by our group [56]. Interleukin receptor accessory protein (IL-1RAP) is another promising target on the surface of LSCs of AML, myelodysplastic syndrome and chronic myeloid leukemia (CML) but not on healthy HSCs, and it was shown to be effective in vitro and in vivo [57]. PRAME is an intracellular cancer and testis antigen highly expressed in acute myeloid leukemia blasts and normal reproductive tissues. A T cell receptor (TCR) mimic antibody, Pr20, that recognizes the peptide-HLA complex can target intracellular PRAME. Kirkey and colleagues generated PRAME mTCR CAR T cells that were cytotoxic to HLA-A2 restricted AML cells in vitro and in vivo without an impact on normal hematopoiesis [58]. Jetani et al suggested that Siglec-6 could be a convenient target sparing HSC or hematopoietic progenitor cells [59].
Efforts to discover efficient targets for immunotherapeutic strategies accelerated following technical progress in proteomic and transcriptomic assays. These assays help to understand cellular behavior on the protein level instead of immunophenotyping malignant cells. Hoffman et al described mass-spectrometry based phenotyping of HL60 and NB4 cell lines [60]. Perna et al performed surface-specific proteomic and transcriptomic studies in AML patients and normal tissues to indicate a potential therapeutic target. However, none of the surface proteins showed a similar expression profile to CD19. These studies suggested a combinatorial targeting strategy, which is discussed further in other sections [26]. Kohnke and colleagues aimed to discover de novo targets using cell-surface capture technology to detect the surfaceome, a set of proteins expressed on the surface of primary AML patient samples, including surface receptor, transporters, adhesion molecules among others. They identified three promising targets: CD148, ITGA4 and Integrin beta-7. Among these, Integrin beta-7 was the most favorable due to low or absent expression in healthy hematopoietic tissues [61]. In a recent scRNA-sec approach, two antigen targets-CSF1R and CD86 revealed potential targets for CAR T-cell therapy with broad expression on AML blasts accompanied by minimal toxicities toward relevant healthy cells and tissues [62].
3.2. Limiting the ‘on target-off tumor’ effect:
Unacceptably severe or prolonged toxicities, especially cytopenias and infections, can occur more frequently than in clinical trials with CD19 CAR T cells in lymphomas due to older and therapy-resistant patient populations. It is crucial to maintain anti-cancer immune surveillance and clinical efficacy while avoiding toxicity. Safety or suicide genes are widely used to alleviate toxicity from CAR T cells [51,63,64]. Alternatively, administration of cross-reactive CAR T cells can be a bridge to allogeneic stem cell transplants. Tasian et al showed three different approaches (anti-CD123 messenger RNA electroporated CAR T cells, administration of alemtuzumab, and administration of rituximab to CD20-coexpressing CART123) to eliminate CD123 CAR T cells without effecting the antitumor activity in murine models [65].
Many cell engineering approaches have attempted to improve safety in designing CARs, such as logic gating of T cell recognition and SnyNotch receptors [66]. Mutation in the anti-CD123 CAR antigen binding domain reduced the antigen binding affinity, as reported by Archangeli et al [67]. Mild adverse events were demonstrated in the interim analysis of a phase 1 trial of rapidly switchable universal CAR-T (UniCAR) targeting CD123 [68,69]. Benmebarek and colleagues generated a controllable CAR platform-synthetic agonist receptor (SAR) T cells only activated in the presence of their CD33 or CD123 scFv construct in vitro and in AML xenograft models [70]. Dimerizing Agent Regulated ImmunoReceptor Complex (DARIC) is a split receptor design that modulates CAR T cell activation by rapamycin to dimerize units. This approach aims to aid hematopoietic recovery and mitigate toxicity. We have recently demonstrated the potential benefits of DOX-inducible CAR-T therapy, allowing the control of CAR-T using an external trigger. It is an effective and sensitive way to turn CAR-T activity ON or OFF in order to prevent unwanted side effects and reduce prolonged toxicities [71]. Cooper and colleagues developed a lentiviral DARIC construct that targets a C2 splice isoform with the membrane proximal domain of CD33, and a Phase I study using this strategy is now open for enrollment [72]. Potentially, the most feasible approach is to knock out the targeted antigen in normal marrow cells. CD33 deletion in primary HSPCs maintained their full function in terms of engraftment and differentiation, and it efficiently reduced off-tumor targeting while preserving on-tumor efficacy [73,74].
3.3. Combinatorial Antigen Targeting for Heterogeneity:
Combinations of CARs against different AML targets might be a promising solution due to the lack of a leukemia-specific target antigen [66]. Previously, promising results were reported with dual or trispecific CAR T cells against B-cell malignancies and solid tumor models to overcome the heterogeneity and antigen escape [75,76]. Indeed, the phenomenon of ‘antigen escape’ as a reason of failure in CAR T cells for AML has not been clearly demonstrated in previous pre-clinical reports [77,78]. Nonetheless, regardless of antigen expression levels, dual-targeting CAR T cells were associated with increased T-cell activation and proliferation. This effect might be due to increased interaction with the target cell, favoring immune synapse formation and subsequent T cell recruitment [79,80]. How to combine suitable pairs of antigens to enhance therapeutic efficacy without increasing off-tumor toxicity are still pieces of the puzzle yet to be solved. Perna et al suggested four possible combinatorial pairings (CD33+ADGRE, CLEC12A+CCR1, CD33+CD70 and LILRB+CLEC12A) to target AML with an algorithm integrating proteomics and transcriptomics [26].
The combinatorial antigen targeting strategy has already been applied in AML. Bicistronic CD123 and CD33 CAR T cells showed significant anti-tumor activity in artificially created cell lines (CD33+CD123-, CD33+CD123-) and in vivo [81]. In a phase I trial, CLL-1 and CD33 bicistronic CAR T cells reported remarkable results [82]. Similarly, 10 of 11 pediatric R/R AML patients infused with CLL-1 or CLL-1-CD33 dual CAR T cells had a response (5 reached CR/MRD-) without dose-limiting toxicities [83]. Atilla et al reported that dual targeting with either a CD33 CAR or a CD123 CAR and a CLL-1 CAR increased anti-tumor activity most profoundly when the target antigen expression on the tumor cells was low. Since the expression of each target antigen is highly variable, we chose to modify T cells with two separate vectors targeting CLL-1 and CD33/CD123 to get a mixed product rather than a bicistronic or tandem CAR design in which the molar ratio of each target is fixed [77]. A universal CAR T cell platform (on/off switching mechanism) successfully targeted CD33 and CD123 AML blasts in vitro and in vivo [84]. Haubner et al presented a novel combinatorial ADGRE-2 targeting CAR and CLEC12A-targeting chimeric costimulatory receptor (CCR) (IF-BETTER gate) that triggered high anti-leukemic activity in vitro and in vivo while sparing vital normal hematopoietic cells [85].
3.4. Neoantigens:
Neoantigens are limited to malignant clones that arise from somatic mutation [86,87]. Because recurrent gene alterations can be shared by AML patients, neoantigens trigger potent anti-leukemic responses [88]. Distinct from other cancers, AML presents with low mutational burden, so recognizing neoantigens arising from mutations is rare [89,90]. Neoantigens from driver gene mutations appear to be ideal targets for immunotherapy since immune evasion is unlikely [91].
The major difficulty in identifying neoantigens was resolved following MS HLA-ligand profiling with whole-exon sequencing and RNA-sequencing techniques [92]. An adequate number of T cells may not be present in AML patients following chemotherapy, or the response may be scarce due to illness-related immunodeficiency [93]. Therefore, the preferred source of neoantigen-specific T cells is usually healthy donors; cells can be isolated using MHC multimers or tetramers carrying the same neoepitopes [94]. Several studies tested the efficacy of neoantigens: CD 8 T cell clones induced by a nonameric neo-peptide (REEMEVHEL) derived from CBFβ-SMMHC fusion protein and in a HLA-A 40:1 restricted manner [93] showed cytotoxicity in in vitro and in vivo models. A neoepitope from NPM1c (CLAVEEVSL) was identified from HLA-A02:01+ AML patients, and healthy CD8 T cells showed lysis when transduced with the same TCR [95]. CD 8+ T cells from an FLT-ITD-positive patient (HLA-A 01:01-restricted) showed an anti-tumor response to a neoepitope (YVDFREYEYY) encoded by ITD protein region [96].
3.5. T cell receptor (TCR) T cells for treatment of AML:
T cell receptor (TCR) engineered T cells act through their modified TCRs and tumor-associated antigens (TAAs) presented by human leukocyte antigen (HLA) molecules on the surfaces of leukemic cells. The target protein can be expressed intracellularly or on the cell surface. TCR T cells have less stringent antigen requirements for T cell activation than CAR T cells [97]. TCR-T cell immunotherapy in AML has still barriers that need to be addressed. The major drawbacks are that TAAs might be expressed by non-malignant cells causing on-target, off-tumor toxicities, dose-related toxicity, limited persistence, and chance of immune escape [98,99]. Dose optimization of TCR-T cells, combining the treatment with exogenous cytokines (e.g., IL-21, IL-7 and IL-15), or adding genetically engineered signaling during cell expansion and demethylating agents such as decitabine might overcome the disadvantages of TCR-T cell application [100]. One other limitation of TCR transfer is the mispairing of endogenous and exogenous TCR components that impair the function; this limitation might be prevented by swapping the constant regions of mouse and human TCRs or codon-optimized cysteine modified TCRs in which TCR-α and β are linked by a T2A sequence [101,102,103]. Another approach uses TCR-like CAR T cells that contain scFv and CAR signaling mechanisms that recognize peptides in the context of MHC class I molecules [104].
TCR-T cells against WT1, PRAME and HA-1 demonstrated anti-leukemic effects in vitro and in a clinical setting in AML [105,106,107,108]. Responses from clinical trials are variable due to different patient populations, doses of administration, and targets. In the first-in-human trial of TCR-T reacting with WT1 in the context of HLA-A*24:02, only 2 patients out of 8 showed transient decreases in blast counts in bone marrow. The patients had minimal toxicities including fever, edema, arthritis, skin reactions [109]. Preferable adverse events were observed in ten patients treated with autologous WT-1 specific TCR-T cells that persisted through 12 months in another trial [110]. More clinical trials on TCR-T cells for AML are detailed in Table 3.
Table 3.
Clinical Studies of TCR-T cells against AML (CML: Chronic Myeloid Leukemia, CRS: Cytokine Release Syndrome, MDS: Myelodysplastic Syndrome, TCR-T: T-cell Receptor T cell, WT-1: Wilms’ tumor-1).
Table 3.
Clinical Studies of TCR-T cells against AML (CML: Chronic Myeloid Leukemia, CRS: Cytokine Release Syndrome, MDS: Myelodysplastic Syndrome, TCR-T: T-cell Receptor T cell, WT-1: Wilms’ tumor-1).
| Study | TCR-T Therapy | Study Phase/Number of Patients | Study Outcomes | Adverse Events |
|---|---|---|---|---|
| NCT02550535 | Autologous WT1 TCR-T cells | Phase I/II, 10 patients (6 AML, 3 MDS and 1 TKI-resistant CML) | All 6 AML patients were alive at last follow up (median 12 months; range 7-12.8 months). The 3 patients with MDS had a median survival of 3 months (range 2.1-3.96 months).2 died from progressive disease and one from other causes. 2 patients had disease progression. |
1 CRS |
| UMIN00001159 | Autologous WT1 siTCR-T cells | Unknown | 2 patients showed transient decrease in blast counts. | None |
| NCT01640301 | Allogeneic WT1 TCR-T cells | Phase I/II, 12 patients | With a median follow-up of 44 months (range 21–57 months) following infusion all 12 patients did not have evidence of disease. | None |
| NCT03503958 | Autologous PRAME TCR-T cells | Phase I | Not posted | Not posted |
| NCT01621724 | Autologous WT1 TCR-T cells | Phase I/II, 7 patients | Not posted | Not posted |
In order to promote graft-vs-leukemia (GVL) reactivity after HLA-matched allogeneic stem cell transplantation, Chapuis et al isolated WT-1 TCR (TCRC4) from HLA-A2+ normal donor repertoires (WT-1 TCR CAR T cells) following allogeneic hematopoietic stem cell transplantation (allo-HSCT), inserted TCRC4 into Epstein-Bar virus-specific donor CD8+ T cells to minimize graft-versus-host disease and infused them prophylactically post-alloHSCT. This strategy achieved 100% relapse-free survival at a median of 44 months [105]. In a similar approach, donor-derived EBV and/or CMV specific T cells were redirected by HA-1H TCR to treat HA-1H positive HLA-A* 02:01 positive patients with high-risk leukemia after alloHSCT. However, the overall feasibility and efficacy was too low to warrant further clinical development [111].
3.6. CAR NK cells:
NK cells are lymphoid cells involved in the innate immune response; they are programmed to kill virus-infected and malignant cells without causing significant graft vs host disease, CRS or neurotoxicity [112]. AML has been an attractive target for NK cell therapy as an allogeneic product [113]. Despite several manipulations for longer persistence of NK cells, the response to NK cell infusions varies without long-term remissions [113]. NK cells differentiated into cytokine induced memory-like NK cells following stimulation with IL-12, IL-15 and IL-18 and showed a distinct transcriptional and surface proteomic profile as well as enhanced functionality [114]. Cytokine induced memory-like (CIML) NK cells are generated by in vitro pre-activation with IL-12, IL-15 and IL-18 and showed promising responses in a Phase I trial in relapsed refractory AML [115]. Dong et al reported potent antileukemic activity in vitro and in vivo with peripheral-blood derived CIML NK cells with TCR-like CAR specific for NPM1c+ HLA-A*0201+ AML [116]. Genome editing of NK cells to upregulate the cytotoxicity by knocking out suppression-associated markers ADAM17 (involved in cleavage of CD19) and PD-1 is the next step [117].
The first CAR NK cell therapy was administered for B-cell relapsed and refractory lymphoma and CLL. Umbilical-derived, HLA-mismatched, anti-CAR 19 CAR NK cell therapy has been studied in relapsed and refractory lymphoma and CLL and showed promising results, with 73% of patients in complete remission [118]. CAR NK cell therapy is favorable in terms of minimal risk of toxicity and potential ‘off-the-shelf’ application. Successful application of CD33-targeted CAR-modified NK cells by transduction of blood-derived primary NK cells showed promising cytotoxicity with unimpeded proliferation in vitro and in vivo without observable side effects [119]. Transgenic expression of secretory IL-15 promoted anti-AML activity and enhanced the persistence of CAR NK cells in vitro, but it was associated with systemic toxicities in vivo with anti-CD123 CAR NK [120] Off-the-shelf cord-derived FLT3 CAR NK cells expressing soluble IL-15 enhanced cytotoxicity and IFN-γ secretion in vitro and improved survival in vivo without HSC toxicity [121]. In a first-in-human Phase I trial, 10 relapsed refractory AML patients received anti-CD33 CAR NK cells; six of them achieved minimal residual disease-negative CR at day 28 without major toxicities [122]. Other phase I trials on CAR-NK cells targeting relapsed refractory AML are still ongoing (NCT05092451, NCT02892695, NCT02944162).
3.7. Manipulations in Manufacturing:
AML is a highly aggressive disease affecting older populations. In a relapsed refractory setting, patients receive many lines of immunosuppressive therapies prior to apheresis, which affects T cell function, and the timeline of manufacturing raises serious concerns in practice. Optimal CAR construct design will preserve the naïve and central memory phenotype as well as the persistence of T cells. It has been shown that naïve and early memory T-cells have been enriched by decitabine administered with CD123 CAR T-cells [123].
Administration of off-the shelf ready-to-use products (allogeneic CAR T cells) generated from healthy donors will provide a valuable solution, since the CAR-T cells product are pre-manufactured without need for customized manufacturing for a specific patient. Two major issues of allogeneic production--graft-versus-host disease and alloreactivity--can be overcome by various strategies [124]. While the AML-off-the-shelf CAR-T cell therapies are being developed, these approaches are still in the early stages of developments, in comparison with B cell malignancies. Two patients from a CD38 targeted CAR T cell trial received a donor-derived CAR construct [125]. TCRαβ negative T cells manufactured from healthy donors by TALEN gene editing targeting CD123 (allogeneic, UCART123) eliminated AML in vitro and in vivo with modest toxicity to normal hematopoietic stem/progenitor cells [126]. A phase I clinical trial was halted following the death of first patient because of severe CRS and capillary leak syndrome with unrelated-donor derived allogeneic anti-CD123 CAR Ts. This trial resumed following the revision of the eligibility criteria and dose modifications [127].
DNA transposon systems are sophisticated systems for stable genetic modification that can deliver large genetic cargos and can be used to reduce cost [128]. Clinical-grade CAR-T cell products using Sleeping Beauty and piggyBac for multiple myeloma and leukemia are under investigation [129,130,131]. Gurney and colleagues applied a non-viral approach to primary CAR-NK cell production combining the TcBuster DNA transposon system targeting a C-type lectin-like molecule-1 (CLL-1/C-Type Lectin Domain Family 12 Member A, CLEC12) with GMP-grade Epstein-Barr virus-transformed lymphoblastoid feeder cell (EBC-LCL) for expansion. This approach knocked out a negative regulator of NK cell stimulation, cytokine-inducible SH-2 containing protein (CISH), using CRISPR/Cas9 to enhance the functionality of CLL-1 CAR NK cells without requiring IL-15 stimulation [132].
3.8. Strategies to Overcome the Negative Effects of Microenvironment:
There are several approaches described to modulate immunosuppressive microenvironment. Immune evasion such as upregulating immune checkpoint proteins has proven to be a way which can dampen the antitumor response and limit efficacy of CAR-T cell therapy. Although immune checkpoint blockade in AML has not proven beneficial, [133,134,135] there may be still additional effects in combining CAR T cells and immune checkpoint blockage (PD-1, CTLA-4, etc) that will improve T cell persistence and anti-tumor efficacy [136]. One promising approach is to use gene editing approach to eliminate the expression of immune checkpoint proteins (PD-1, CTLA-4, etc), thereby making them less susceptible tumor microenvironment.
Targeting immunosuppressive cells such as Tregs (with anti-CD25 antibodies) and MDSCs (anti-Gr-1-antibodies) in the tumor microenvironment may enhance anti-tumor immunity. CD33 is also present in MDSC, so targeting CD33 will mediate antitumor activity through direct cytotoxicity of CD33+ blasts and also through inhibition of CD33+ MDSCs [137]. In an AML murine model, the depletion of Tregs increased the proliferation and activity of adoptively transferred tumor reactive cytotoxic T cells [138]. Lymphodepleting chemotherapy prior to CAR T cell infusion suppresses Tregs and augments the expansion of adoptively transferred CAR T cells [139]. This was previously shown in CD19 CAR T cell models through the down regulation of indoleamine 2,3-dioxygenase (IDO), a protein able to deplete tryptophan and other metabolites that inhibit CAR-T cell function [140]. Therapies targeted towards the adenosinergic pathways (antibodies targeting CD73 and CD39) have proven anti-tumor efficacy in mice models [141]. A combinatorial approaches targeting CD73 and IDO could potentially enhance the AML CAR-T therapy, in this regard, our group is integrating universal off the shelf CLL-1 CAR T cells and several nanocarriers to deliver CD73 short hairpin (shRNA) and miRNA-135 as promising strategies for targeting CD73 and IDO, respectively. Combining CD73 and IDO targeted therapy with CAR-T cell approach could potentially enhance the anti-tumor immune response by blocking two separate pathways of immune suppression.
3.9. Allogeneic Hematopoietic Stem Cell Transplantation with CAR T cells:
When and how to combine allo-HSCT with adoptive immunotherapy in AML is still debated. The mechanisms of resistance to T cell-mediated antitumor effects after allo-HSCT are well-defined in sophisticated murine models of allo-HSCT [142]. Combining a novel myeloablative irradiation-based conditioning regimen with regulatory and conventional T cell immunotherapy in haploidentical transplantation was shown to eradicate AML [143]. Published studies on how and when to combine CAR T cells in the setting of allo-HSCT showed conflicting results. Pan et al reported that allo-HSCT following CD19 CAR T treatment improved event-free survival and reduced relapse risk [144,145] while other studies failed to demonstrate a benefit [146]. Summers et al reported the clearest leukemia-free survival in patients who had early loss of functionality of CAR T cells (for CD19 CART-loss of B-cell aplasia) [147]. Data on evidence of the benefit of allo-HSCT following CAR T cell therapy in AML patients is scarce. Zhang et al demonstrated that among 6 patients who received allo-HSCT following anti-CLL1 CAR T cell treatment received, 4 of them achieved CR [148]. One other approach takes advantage of the myelosuppression effect of CD123 CAR T cells and administers donor-derived CD123 CAR T cells as a part of a conditioning regimen for haplo-HSCT [149].
4. Summary and Conclusion:
The tremendous advances in understanding the molecular and cellular mechanisms of AML have made it possible to manipulate the immune system and BM niches. Treating AML with CAR T cells is still in an immature stage. Experience with allogeneic stem cell transplantation, which is the most effective immune cellular therapy for AML, is guiding other directed therapies. One of the major challenges in developing CAR-T cell therapy for AML is the lack of a suitable antigen that is expressed uniquely on AML cells. Identifying and isolating target antigens that are homogeneously and stably expressed in all leukemic blasts and leukemic stem cells with limited on-target off-tumor toxicity, investigating complex interactions in the AML microenvironment, and seeking a suitable cell source all improve the fine-tuning of CARs.
Sophisticated methods for ex vivo manufacturing are now changing the in vivo dynamics and the character of the final product (Figure). In AML, personalization should be taken a step further in directed cellular therapies with platforms that will standardize the optimal CAR design for the target antigen or antigens in line with patient-specific immunophenotyping findings, the selection of a compatible carrier cell, and the cellular subtype.
Figure.
dioxygenase (IDO), KO:Knock-out, MHC-I: Major Histocompatibility Complex-I, PD-1:Programmed Cell Death-1, TCR: T cell Receptor)Author Contributions: Conceptualization, EA; Methodology EA; Resources: EA; Writing-Review and Editing EA, KB.
Figure.
dioxygenase (IDO), KO:Knock-out, MHC-I: Major Histocompatibility Complex-I, PD-1:Programmed Cell Death-1, TCR: T cell Receptor)Author Contributions: Conceptualization, EA; Methodology EA; Resources: EA; Writing-Review and Editing EA, KB.
Acknowledgments
EA is a PhD student from the Biomedicine Program of the University of Granada (Spain). K.B; PECART-0027-2020 funded by Consejería de Salud y Familias and ProyExcel_00875 Consejería de Transformación Económica, Industria, Conocimiento y Universidades; EA and KB are active members of COST Action (CA21113), supported by COST (European Cooperation in Science and Technology)
Conflicts of Interest
None
References
- Vishwasrao P, Li G, Boucher JC, Smith DL, Hui SK. Emerging CAR T Cell Strategies for the Treatment of AML. Cancers (Basel). 2022 Feb 27;14(5):1241. [CrossRef]
- Döhner H, Weisdorf DJ, Bloomfield CD. Acute Myeloid Leukemia. New England Journal of Medicine, 2015 373(12), 1136-1152.
- Hofmann S, Schubert ML, Wang L, He B, Neuber B, Dreger P, Müller-Tidow C, Schmitt M. Chimeric Antigen Receptor (CAR) T Cell Therapy in Acute Myeloid Leukemia (AML). J Clin Med. 2019 Feb 6;8(2):200. [CrossRef]
- Marofi F, Rahman HS, Al-Obaidi ZMJ, Jalil AT, Abdelbasset WK, Suksatan W, Dorofeev AE, Shomali N, Chartrand MS, Pathak Y, Hassanzadeh A, Baradaran B, Ahmadi M, Saeedi H, Tahmasebi S, Jarahian M. Novel CAR T therapy is a ray of hope in the treatment of seriously ill AML patients. Stem Cell Res Ther. 2021 Aug 20;12(1):465. [CrossRef]
- Karamitros D, Stoilova B, Aboukhalil Z, Hamey F, Reinisch A, Samitsch M, Quek L, Otto G, Repapi E, Doondeea J, Usukhbayar B, Calvo J, Taylor S, Goardon N, Six E, Pflumio F, Porcher C, Majeti R, Göttgens B, Vyas P. Single-cell analysis reveals the continuum of human lympho-myeloid progenitor cells. Nat Immunol. 2018 Jan;19(1):85-97. [CrossRef]
- Pollyea DA, Jordan CT. Therapeutic targeting of acute myeloid leukemia stem cells. Blood. 2017 Mar 23;129(12):1627-1635. [CrossRef]
- Döhner H, Estey E, Grimwade D, Amadori S, Appelbaum FR, Büchner T, Dombret H, Ebert BL, Fenaux P, Larson RA, Levine RL, Lo-Coco F, Naoe T, Niederwieser D, Ossenkoppele GJ, Sanz M, Sierra J, Tallman MS, Tien HF, Wei AH, Löwenberg B, Bloomfield CD. Diagnosis and management of AML in adults: 2017 ELN recommendations from an international expert panel. Blood. 2017 Jan 26;129(4):424-447. [CrossRef]
- Cancer Genome Atlas Research Network; Ley TJ, Miller C, Ding L, Raphael BJ, Mungall AJ, Robertson A, Hoadley K, Triche TJ Jr, Laird PW, Baty JD, Fulton LL, Fulton R, Heath SE, Kalicki-Veizer J, Kandoth C, Klco JM, Koboldt DC, Kanchi KL, Kulkarni S, Lamprecht TL, Larson DE, Lin L, Lu C, McLellan MD, McMichael JF, Payton J, Schmidt H, Spencer DH, Tomasson MH, Wallis JW, Wartman LD, Watson MA, Welch J, Wendl MC, Ally A, Balasundaram M, Birol I, Butterfield Y, Chiu R, Chu A, Chuah E, Chun HJ, Corbett R, Dhalla N, Guin R, He A, Hirst C, Hirst M, Holt RA, Jones S, Karsan A, Lee D, Li HI, Marra MA, Mayo M, Moore RA, Mungall K, Parker J, Pleasance E, Plettner P, Schein J, Stoll D, Swanson L, Tam A, Thiessen N, Varhol R, Wye N, Zhao Y, Gabriel S, Getz G, Sougnez C, Zou L, Leiserson MD, Vandin F, Wu HT, Applebaum F, Baylin SB, Akbani R, Broom BM, Chen K, Motter TC, Nguyen K, Weinstein JN, Zhang N, Ferguson ML, Adams C, Black A, Bowen J, Gastier-Foster J, Grossman T, Lichtenberg T, Wise L, Davidsen T, Demchok JA, Shaw KR, Sheth M, Sofia HJ, Yang L, Downing JR, Eley G. Genomic and epigenomic landscapes of adult de novo acute myeloid leukemia. N Engl J Med. 2013 May 30;368(22):2059-74.
- DiNardo CD, Cortes JE. Mutations in AML: prognostic and therapeutic implications. Hematology Am Soc Hematol Educ Program. 2016;2016(1):348–55. [CrossRef]
- Jan M, Snyder TM, Corces-Zimmerman MR, Vyas P, Weissman IL, Quake SR, Majeti R. Clonal evolution of preleukemic hematopoietic stem cells precedes human acute myeloid leukemia. Sci Transl Med. 2012 Aug 29;4(149):149ra118.
- Corces MR, Buenrostro JD, Wu B, Greenside PG, Chan SM, Koenig JL, Snyder MP, Pritchard JK, Kundaje A, Greenleaf WJ, Majeti R, Chang HY. Lineage-specific and single-cell chromatin accessibility charts human hematopoiesis and leukemia evolution. Nat Genet. 2016 Oct;48(10):1193-203. [CrossRef]
- Stengel A, Shahswar R, Haferlach T, Walter W, Hutter S, Meggendorfer M, Kern W, Haferlach C. Whole transcriptome sequencing detects a large number of novel fusion transcripts in patients with AML and MDS. Blood Adv. 2020 Nov 10;4(21):5393-5401. doi: 10.1182/bloodadvances.2020003007. [CrossRef]
- Chakraborty, S., Park, C.Y. Pathogenic Mechanisms in Acute Myeloid Leukemia. Curr. Treat. Options in Oncol. 23, 1522–1534 (2022). [CrossRef]
- Li S, Garrett-Bakelman FE, Chung SS, Sanders MA, Hricik T, Rapaport F, Patel J, Dillon R, Vijay P, Brown AL, Perl AE, Cannon J, Bullinger L, Luger S, Becker M, Lewis ID, To LB, Delwel R, Löwenberg B, Döhner H, Döhner K, Guzman ML, Hassane DC, Roboz GJ, Grimwade D, Valk PJ, D'Andrea RJ, Carroll M, Park CY, Neuberg D, Levine R, Melnick AM, Mason CE. Distinct evolution and dynamics of epigenetic and genetic heterogeneity in acute myeloid leukemia. Nat Med. 2016 Jul;22(7):792-9. doi: 10.1038/nm.4125. [CrossRef]
- Kreso A, Dick JE. Evolution of the cancer stem cell model. Cell Stem Cell. 2014;14:275–91. [CrossRef]
- Thomas D, Majeti R. Biology and relevance of human acute myeloid leukemia stem cells. Blood. 2017;129:1577–85. [CrossRef]
- Zeng AGX, Bansal S, Jin L, Mitchell A, Chen WC, Abbas HA, Chan-Seng-Yue M, Voisin V, van Galen P, Tierens A, Cheok M, Preudhomme C, Dombret H, Daver N, Futreal PA, Minden MD, Kennedy JA, Wang JCY, Dick JE. A cellular hierarchy framework for understanding heterogeneity and predicting drug response in acute myeloid leukemia. Nat Med. 2022 Jun;28(6):1212-1223. [CrossRef]
- Jiang, X.-P. Li, Y.-T. Dai, B. Chen, X.-Q. Weng, S.-M. Xiong, M. Zhang, J.- Y. Huang, Z. Chen, S.-J. Chen, Multidimensional study of the heterogeneity of leukemia cells in t(8;21) acute myelogenous leukemia identifies the subtype with poor outcome, Proc. Natl. Acad. Sci. U. S. A. 117 (2020) 20117–20126. [CrossRef]
- Li K, Du Y, Cai Y, Liu W, Lv Y, Huang B, Zhang L, Wang Z, Liu P, Sun Q, Li N, Zhu M, Bosco B, Li L, Wu W, Wu L, Li J, Wang Q, Hong M, Qian S. Single-cell analysis reveals the chemotherapy-induced cellular reprogramming and novel therapeutic targets in relapsed/refractory acute myeloid leukemia. Leukemia. 2023 Feb;37(2):308-325. [CrossRef]
- Stratmann S, Vesterlund M, Umer HM, Eshtad S, Skaftason A, Herlin MK, Sundström C, Eriksson A, Höglund M, Palle J, Abrahamsson J, Jahnukainen K, Munthe-Kaas MC, Zeller B, Tamm KP, Lindskog C, Cavelier L, Lehtiö J, Holmfeldt L. Proteogenomic analysis of acute myeloid leukemia associates relapsed disease with reprogrammed energy metabolism both in adults and children. Leukemia. 2023 Mar;37(3):550-559.
- Sadelain M (2016). Chimeric antigen receptors: driving immunology towards synthetic biology. Curr Opin Immunol 41, 68–76. [CrossRef]
- LeBien TW, and Tedder TF (2008). B lymphocytes: how they develop and function. Blood 112, 1570– 1580. [CrossRef]
- Bakker AB, van den Oudenrijn S, Bakker AQ, Feller N, van Meijer M, Bia JA, Jongeneelen MA, Visser TJ, Bijl N, Geuijen CA, et al. (2004). C-type lectin-like molecule-1: a novel myeloid cell surface marker associated with acute myeloid leukemia. Cancer Res 64, 8443–8450.
- Kenderian SS, Ruella M, Shestova O, Klichinsky M, Aikawa V, Morrissette JJ, Scholler J, Song D, Porter DL, Carroll M, et al. (2015). CD33-specific chimeric antigen receptor T cells exhibit potent preclinical activity against human acute myeloid leukemia. Leukemia 29, 1637–1647. [CrossRef]
- Bendall LJ, Bradstock KF, and Gottlieb DJ (2000). Expression of CD44 variant exons in acute myeloid leukemia is more common and more complex than that observed in normal blood, bone marrow or CD34+ cells. Leukemia 14, 1239–1246. [CrossRef]
- Perna F, Berman SH, Soni RK, Mansilla-Soto J, Eyquem J, Hamieh M, Hendrickson RC, Brennan CW, Sadelain M. Integrating Proteomics and Transcriptomics for Systematic Combinatorial Chimeric Antigen Receptor Therapy of AML. Cancer Cell. 2017 Oct 9;32(4):506-519. [CrossRef]
- Melao, A. (2017). FDA Suspends UCART123 Trials after Patient Death (Immuno-Oncology News).
- Wang QS, Wang Y, Lv HY, Han QW, Fan H, Guo B, Wang LL, Han WD. Treatment of CD33-directed chimeric antigen receptor-modified T cells in one patient with relapsed and refractory acute myeloid leukemia. Mol Ther. 2015 Jan;23(1):184-91. [CrossRef]
- Laszlo GS, Estey EH, Walter RB. The past and future of CD33 as therapeutic target in acute myeloid leukemia. Blood Rev. 2014;28(4):143–53. [CrossRef]
- Wang QS, Wang Y, Lv HY, Han QW, Fan H, Guo B, Wang LL, Han WD. Treatment of CD33-directed chimeric antigen receptor-modified T cells in one patient with relapsed and refractory acute myeloid leukemia. Mol Ther. 2015 Jan;23(1):184-91. [CrossRef]
- Casucci M, Nicolis di Robilant B, Falcone L, Camisa B, Norelli M, Genovese P, Gentner B, Gullotta F, Ponzoni M, Bernardi M, Marcatti M, Saudemont A, Bordignon C, Savoldo B, Ciceri F, Naldini L, Dotti G, Bonini C, Bondanza A. CD44v6-targeted T cells mediate potent antitumor effects against acute myeloid leukemia and multiple myeloma. Blood. 2013 Nov 14;122(20):3461-72. [CrossRef]
- Jordan CT, Upchurch D, Szilvassy SJ, Guzman ML, Howard DS, Pettigrew AL, Meyerrose T, Rossi R, Grimes B, Rizzieri DA, Luger SM, Phillips GL. The interleukin-3 receptor alpha chain is a unique marker for human acute myelogenous leukemia stem cells. Leukemia. 2000 Oct;14(10):1777-84. [CrossRef]
- Geiger M, Stubenrauch KG, Sam J, Richter WF, Jordan G, Eckmann J, Hage C, Nicolini V, Freimoser-Grundschober A, Ritter M, Lauer ME, Stahlberg H, Ringler P, Patel J, Sullivan E, Grau-Richards S, Endres S, Kobold S, Umaña P, Brünker P, Klein C. Protease-activation using anti-idiotypic masks enables tumor specificity of a folate receptor 1-T cell bispecific antibody. Nat Commun. 2020 Jun 24;11(1):3196. [CrossRef]
- Huntly BJ, Gilliland DG. Leukaemia stem cells and the evolution of cancer-stem-cell research. Nat Rev Cancer. 2005;5(4):311–21. [CrossRef]
- Dias S, Hattori K, Heissig B, Zhu Z, Wu Y, Witte L, Hicklin DJ, Tateno M, Bohlen P, Moore MA, Rafii S. Inhibition of both paracrine and autocrine VEGF/ VEGFR-2 signaling pathways is essential to induce long-term remission of xenotransplanted human leukemias. Proc Natl Acad Sci U S A. 2001 Sep 11;98(19):10857-62. [CrossRef]
- Hussong JW, Rodgers GM, Shami PJ. Evidence of increased angiogenesis in patients with acute myeloid leukemia. Blood. 2000;95(1):309–13.
- Mussai F, De Santo C, Abu-Dayyeh I, Booth S, Quek L, McEwen-Smith RM, Qureshi A, Dazzi F, Vyas P, Cerundolo V. Acute myeloid leukemia creates an arginase-dependent immunosuppressive microenvironment. Blood. 2013 Aug 1;122(5):749-58. [CrossRef]
- Ben-Batalla I, Schultze A, Wroblewski M, Erdmann R, Heuser M, Waizenegger JS, Riecken K, Binder M, Schewe D, Sawall S, Witzke V, Cubas-Cordova M, Janning M, Wellbrock J, Fehse B, Hagel C, Krauter J, Ganser A, Lorens JB, Fiedler W, Carmeliet P, Pantel K, Bokemeyer C, Loges S. Axl, a prognostic and therapeutic target in acute myeloid leukemia mediates paracrine crosstalk of leukemia cells with bone marrow stroma. Blood. 2013 Oct 3;122(14):2443-52. doi: 10.1182/blood-2013-03-491431. [CrossRef]
- Austin R, Smyth MJ, Lane SW. Harnessing the immune system in acute myeloid leukaemia. Crit Rev Oncol Hematol. 2016 Jul;103:62-77. [CrossRef]
- Epperly R, Gottschalk S, Velasquez MP. A Bump in the Road: How the Hostile AML Microenvironment Affects CAR T Cell Therapy. Front Oncol. 2020 Feb 28;10:262. [CrossRef]
- Baragaño Raneros A, López-Larrea C, Suárez-Á lvarez B. Acute Myeloid Leukemia and NK Cells: Two Warriors Confront Each Other. OncoImmunology (2019) 8(2):e1539617.
- Al-Kaabneh B, Frisch B, Aljitawi OS. The Potential Role of 3D In Vitro Acute Myeloid Leukemia Culture Models in Understanding Drug Resistance in Leukemia Stem Cells. Cancers (Basel). 2022 Oct 26;14(21):5252. [CrossRef]
- Mbongue JC, Nicholas DA, Torrez TW, Kim NS, Firek AF, Langridge WH. The Role of Indoleamine 2, 3-Dioxygenase in Immune Suppression and Autoimmunity. Vaccines (Basel). 2015 Sep 10;3(3):703-29. [CrossRef]
- Hanoun M, Zhang D, Mizoguchi T, Pinho S, Pierce H, Kunisaki Y, Lacombe J, Armstrong SA, Dührsen U, Frenette PS. Acute myelogenous leukemia-induced sympathetic neuropathy promotes malignancy in an altered hematopoietic stem cell niche. Cell Stem Cell. 2014 Sep 4;15(3):365-375. [CrossRef]
- Lane SW, Wang YJ, Lo Celso C, Ragu C, Bullinger L, Sykes SM, Ferraro F, Shterental S, Lin CP, Gilliland DG, Scadden DT, Armstrong SA, Williams DA. Differential niche and Wnt requirements during acute myeloid leukemia progression. Blood. 2011 Sep 8;118(10):2849-56. [CrossRef]
- Rickmann M, Macke L, Sundarasetty BS, Stamer K, Figueiredo C, Blasczyk R, Heuser M, Krauter J, Ganser A, Stripecke R. Monitoring dendritic cell and cytokine biomarkers during remission prior to relapse in patients with FLT3-ITD acute myeloid leukemia. Ann Hematol. 2013 Aug;92(8):1079-90. [CrossRef]
- van Galen P, Hovestadt V, Wadsworth Ii MH, Hughes TK, Griffin GK, Battaglia S, Verga JA, Stephansky J, Pastika TJ, Lombardi Story J, Pinkus GS, Pozdnyakova O, Galinsky I, Stone RM, Graubert TA, Shalek AK, Aster JC, Lane AA, Bernstein BE. Single-Cell RNA-Seq Reveals AML Hierarchies Relevant to Disease Progression and Immunity. Cell. 2019 Mar 7;176(6):1265-1281.e24. [CrossRef]
- Kueberuwa, G., Zheng, W., Kalaitsidou, M., Gilham, D.E., Hawkins, R.E. A Syngeneic Mouse B-Cell Lymphoma Model for Pre-Clinical Evaluation of CD19 CAR T Cells. J. Vis. Exp. (140), e58492, doi:10.3791/58492 (2018). [CrossRef]
- Minnie SA, Waltner OG, Ensbey KS, Nemychenkov NS, Schmidt CR, Bhise SS, Legg SRW, Campoy G, Samson LD, Kuns RD, Zhou T, Huck JD, Vuckovic S, Zamora D, Yeh A, Spencer A, Koyama M, Markey KA, Lane SW, Boeckh M, Ring AM, Furlan SN, Hill GR. Depletion of exhausted alloreactive T cells enables targeting of stem-like memory T cells to generate tumor-specific immunity. Sci Immunol. 2022 Oct 21;7(76):eabo3420. [CrossRef]
- Tashiro H, Sauer T, Shum T, Parikh K, Mamonkin M, Omer B, et al. Treatment of acute myeloid leukemia with T cells expressing chimeric antigen receptors directed to c-type lectin-like molecule 1. Mol Ther (2017) 25(9):2202–13. [CrossRef]
- Ataca Atilla P, McKenna MK, Tashiro H, Srinivasan M, Mo F, Watanabe N, Simons BW, McLean Stevens A, Redell MS, Heslop HE, Mamonkin M, Brenner MK, Atilla E. Modulating TNFα activity allows transgenic IL15-Expressing CLL-1 CAR T cells to safely eliminate acute myeloid leukemia. J Immunother Cancer. 2020 Sep;8(2):e001229. [CrossRef]
- Degterev A, Ofengeim D, Yuan J. Targeting RIPK1 for the treatment of human diseases. Proc Natl Acad Sci U S A. 2019 May 14;116(20):9714-9722. [CrossRef]
- Sauer T, Parikh K, Sharma S, Omer B, Sedloev D, Chen Q, Angenendt L, Schliemann C, Schmitt M, Müller-Tidow C, Gottschalk S, Rooney CM. CD70-specific CAR T cells have potent activity against acute myeloid leukemia without HSC toxicity. Blood. 2021 Jul 29;138(4):318-330. [CrossRef]
- Leick MB, Silva H, Scarfò I, Larson R, Choi BD, Bouffard AA, Gallagher K, Schmidts A, Bailey SR, Kann MC, Jan M, Wehrli M, Grauwet K, Horick N, Frigault MJ, Maus MV. Non-cleavable hinge enhances avidity and expansion of CAR-T cells for acute myeloid leukemia. Cancer Cell. 2022 May 9;40(5):494-508.e5. [CrossRef]
- Saxena A, Sheridan DP, Card RT, McPeek AM, Mewdell CC, Skinnider LF. Biologic and clinical significance of CD7 expression in acute myeloid leukemia. Am J Hematol. 1998;58(4):278–84.
- Gomes-Silva D, Atilla E, Atilla PA, Mo F, Tashiro H, Srinivasan M, Lulla P, Rouce RH, Cabral JMS, Ramos CA, Brenner MK, Mamonkin M. CD7 CAR T Cells for the Therapy of Acute Myeloid Leukemia. Mol Ther. 2019 Jan 2;27(1):272-280. [CrossRef]
- Trad R, Warda W, Alcazer V, Neto da Rocha M, Berceanu A, Nicod C, Haderbache R, Roussel X, Desbrosses Y, Daguindau E, Renosi F, Roumier C, Bouquet L, Biichle S, Guiot M, Seffar E, Caillot D, Depil S, Robinet E, Salma Y, Deconinck E, Deschamps M, Ferrand C. Chimeric antigen receptor T-cells targeting IL-1RAP: a promising new cellular immunotherapy to treat acute myeloid leukemia. J Immunother Cancer. 2022 Jul;10(7):e004222. [CrossRef]
- Kirkey DC, Loeb AM, Castro S, McKay CN, Perkins L, Pardo L, Leonti AR, Tang TT, Loken MR, Brodersen LE, Loeb KR, Scheinberg DA, Le Q, Meshinchi S. Therapeutic targeting of PRAME with mTCRCAR T cells in acute myeloid leukemia. Blood Adv. 2023 Apr 11;7(7):1178-1189. [CrossRef]
- Jetani H, Navarro-Bailon A, Maucher M, Frenz S, Verbruggen CM, Yeguas A, et al. Siglec-6 Is a Novel Target for CAR T-Cell Therapy in Acute Myeloid Leukemia (AML). Blood (2021) 138(19):1830–42.
- Hofmann A, Gerrits B, Schmidt A, Bock T, Bausch-Fluck D, Aebersold R, Wollscheid B. Proteomic cell surface phenotyping of differentiating acute myeloid leukemia cells. Blood. 2010 Sep 30;116(13):e26-34.
- Köhnke, T., Liu, X., Haubner, S. et al. Integrated multiomic approach for identification of novel immunotherapeutic targets in AML. Biomark Res 10, 43 (2022). [CrossRef]
- Gottschlich, A., Thomas, M., Grünmeier, R. et al. Single-cell transcriptomic atlas-guided development of CAR-T cells for the treatment of acute myeloid leukemia. Nat Biotechnol (2023). [CrossRef]
- Straathof KC, Pulè MA, Yotnda P, Dotti G, Vanin EF, Brenner MK, et al. An inducible caspase 9 safety switch for T-cell therapy. Blood. 2005. Jun 1;105(11):4247–54. [CrossRef]
- Warda W, Da Rocha MN, Trad R, Haderbache R, Salma Y, Bouquet L, Roussel X, Nicod C, Deschamps M, Ferrand C. Overcoming target epitope masking resistance that can occur on low-antigen-expresser AML blasts after IL-1RAP chimeric antigen receptor T cell therapy using the inducible caspase 9 suicide gene safety switch. Cancer Gene Ther. 2021 Dec;28(12):1365-1375. [CrossRef]
- Tasian SK, Kenderian SS, Shen F, Ruella M, Shestova O, Kozlowski M, et al. Optimized depletion of chimeric antigen receptor T cells in murine xenograft models of human acute myeloid leukemia. Blood (2017) 129(17):2395–407. [CrossRef]
- Simon S, Bugos G, Salter AI, Riddell SR. Synthetic receptors for logic gated T cell recognition and function. Curr Opin Immunol. 2022 Feb;74:9-17. [CrossRef]
- Arcangeli S, Rotiroti MC, Bardelli M, Simonelli L, Magnani CF, Biondi A, et al. Balance of anti-Cd123 chimeric antigen receptor binding affinity and density for the targeting of acute myeloid leukemia. Mol Ther (2017) 25(8):1933–45. [CrossRef]
- Loff S, Dietrich J, Meyer JE, Riewaldt J, Spehr J, von Bonin M, et al. Rapidly switchable universal car-T cells for treatment of Cd123-positive leukemia. Mol Ther Oncol (2020) 17:408–20. [CrossRef]
- Wermke M, Kraus S, Ehninger A, Bargou RC, Goebeler M-E, Middeke JM, et al. Proof of concept for a rapidly switchable universal car-T platform with unicar-T-Cd123 in Relapsed/Refractory aml. Blood (2021) 137(22):3145–8.
- Benmebarek, MR., Cadilha, B.L., Herrmann, M. et al. A modular and controllable T cell therapy platform for acute myeloid leukemia. Leukemia 35, 2243–2257 (2021). [CrossRef]
- Fernández L, Fernández A, Mirones I, Escudero A, Cardoso L, Vela M, Lanzarot D, de Paz R, Leivas A, Gallardo M, Marcos A, Romero AB, Martínez-López J, Pérez-Martínez A. GMP-Compliant Manufacturing of NKG2D CAR Memory T Cells Using CliniMACS Prodigy. Front Immunol. 2019 Oct 10;10:2361. [CrossRef]
- Todd Michael Cooper, Vicky Wu, Ashley Wilson, Jacob Appelbaum, Jordan Jarjour, Josh Gustafson, Stephanie Mgebroff, Catherine Lindgren, Christopher Brown, Michael C. Jensen, Julie R. Park, and Rebecca Alice Gardner. Pediatric and young adult leukemia adoptive therapy (PLAT)-08: A phase 1 study of SC-DARIC33 in pediatric and young adults with relapsed or refractory CD33+AML. Journal of Clinical Oncology 2022 40:16_suppl, TPS7078-TPS7078. [CrossRef]
- Kim MY, Yu KR, Kenderian SS, et al. Genetic inactivation of CD33 in hematopoietic stem cells to enable CAR T cell immunotherapy for acute myeloid leukemia. Cell. 2018;173:1439-53.e19. [CrossRef]
- Liu Y, Wang S, Schubert ML, Lauk A, Yao H, Blank MF, Cui C, Janssen M, Schmidt C, Göllner S, Kleist C, Zhou F, Rahfeld JU, Sauer T, Schmitt M, Müller-Tidow C. CD33-directed immunotherapy with third-generation chimeric antigen receptor T cells and gemtuzumab ozogamicin in intact and CD33-edited acute myeloid leukemia and hematopoietic stem and progenitor cells. Int J Cancer. 2022 Apr 1;150(7):1141-1155. [CrossRef]
- Kailayangiri S, Altvater B, Wiebel M, Jamitzky S, Rossig C. Overcoming Heterogeneity of Antigen Expression for Effective CAR T Cell Targeting of Cancers. Cancers (Basel). 2020 Apr 26;12(5):1075. [CrossRef]
- Schneider D, Xiong Y, Wu D, Hu P, Alabanza L, Steimle B, Mahmud H, Anthony-Gonda K, Krueger W, Zhu Z, Dimitrov DS, Orentas RJ, Dropulić B. Trispecific CD19-CD20-CD22-targeting duoCAR-T cells eliminate antigen-heterogeneous B cell tumors in preclinical models. Sci Transl Med. 2021 Mar 24;13(586):eabc6401.
- Atilla PA, McKenna MK, Watanabe N, Mamonkin M, Brenner MK, Atilla E. Combinatorial antigen targeting strategies for acute leukemia: application in myeloid malignancy. Cytotherapy. 2022 Mar;24(3):282-290. [CrossRef]
- Damiani D, Tiribelli M. Present and Future Role of Immune Targets in Acute Myeloid Leukemia. Cancers (Basel). 2022 Dec 30;15(1):253.
- Xiong W, Chen Y, Kang X, Chen Z, Zheng P, Hsu YH, et al. Immunological Synapse Predicts Effectiveness of Chimeric Antigen Receptor Cells. Mol Ther 2018;26 (4):963–75. [CrossRef]
- Wang QS, Wang Y, Lv HY, Han QW, Fan H, Guo B, et al. Treatment of CD33- directed chimeric antigen receptor-modified T cells in one patient with relapsed and refractory acute myeloid leukemia. Mol Ther 2015;23(1):184– 91. [CrossRef]
- Petrov JC, Wada M, Pinz KG, Yan LE, Chen KH, Shuai X, et al. Compound CAR Tcells as a double-pronged approach for treating acute myeloid leukemia. Leukemia 2018;32(6):1317–26. [CrossRef]
- Liu, Fang & Cao, Yuanzhen & Pinz, Kevin & Ma, Yu & Wada, Masayuki & Chen, Kevin & Ma, Gina & Shen, Jiaqi & Tse, Charlotte & Su, Yi & Xiong, Yisong & He, Guangcui & Li, Yecheng & Ma, Yupo. (2018). First-in-Human CLL1-CD33 Compound CAR T Cell Therapy Induces Complete Remission in Patients with Refractory Acute Myeloid Leukemia: Update on Phase 1 Clinical Trial. Blood. 132. 901-901. 10.1182/blood-2018-99-110579. [CrossRef]
- Zhang H, Bu C, Pen Z, et al. The efficacy and safety of anti-CLL1 based CAR-T cells in children with relapsed or refractory acute myeloid leukemia: A multicenter interim analysis. J Clin Oncol. 2021;39(suppl 15):10000.
- Cartellieri M, Feldmann A, Koristka S, Arndt C, Loff S, Ehninger AV, et al. Switching CAR T cells on and off: a novel modular platform for retargeting of T cells to AML blasts. Blood Cancer J. 2016;6:e458. [CrossRef]
- Sascha Haubner, Jorge Mansilla-Soto, Sarah Nataraj, Friederike Kogel, Qing Chang, Elisa De Stanchina, Kathryn Fraser, Jae H Park, Xiuyan Wang, Isabelle Rivière, Michel Sadelain; Target Densities in Malignant and Normal Cells Determine CAR T Cell Efficacy and Off-Target Hematotoxicity. Blood 2022; 140 (Supplement 1): 869–870.
- Blass E, Ott PA. Advances in the development of personalized neoantigen-based therapeutic cancer vaccines. Nat Rev Clin Oncol. 2021;18:215–29. [CrossRef]
- Blankenstein T, Leisegang M, Uckert W, Schreiber H. Targeting cancerspecifc mutations by T cell receptor gene therapy. Curr Opin Immunol. 2015;33:112–9.
- Tran E, Robbins PF, Lu YC, Prickett TD, Gartner JJ, Jia L, Pasetto A, Zheng Z, Ray S, Groh EM, Kriley IR, Rosenberg SA. T-Cell transfer therapy targeting mutant KRAS in cancer. N Engl J Med. 2016;375:2255–62. [CrossRef]
- Lindsley RC, Mar BG, Mazzola E, Grauman PV, Shareef S, Allen SL, Pigneux A, Wetzler M, Stuart RK, Erba HP, Damon LE, Powell BL, Lindeman N, Steensma DP, Wadleigh M, DeAngelo DJ, Neuberg D, Stone RM, Ebert BL. Acute myeloid leukemia ontogeny is defined by distinct somatic mutations. Blood. 2015 Feb 26;125(9):1367-76. [CrossRef]
- Van Allen EM, Miao D, Schilling B, Shukla SA, Blank C, Zimmer L, Sucker A, Hillen U, Foppen MHG, Goldinger SM, Utikal J, Hassel JC, Weide B, Kaehler KC, Loquai C, Mohr P, Gutzmer R, Dummer R, Gabriel S, Wu CJ, Schadendorf D, Garraway LA. Genomic correlates of response to CTLA-4 blockade in metastatic melanoma. Science. 2015 Oct 9;350(6257):207-211. [CrossRef]
- Blankenstein T, Leisegang M, Uckert W, Schreiber H. Targeting cancerspecific mutations by T cell receptor gene therapy. Curr Opin Immunol (2015) 33:112–9. [CrossRef]
- Ehx G, Larouche JD, Durette C, Laverdure JP, Hesnard L, Vincent K, Hardy MP, Theriault C, Rulleau C, Lanoix J, Bonneil E, Feghaly A, Apavaloaei A, Noronha N, Laumont CM, Delisle JS, Vago L, Hebert J, Sauvageau G, Lemieux S, Thibault P, Perreault C. Atypical acute myeloid leukemiaspecifc transcripts generate shared and immunogenic MHC class-Iassociated epitopes. Immunity. 2021;54:737–52.
- Biernacki MA, Foster KA, Woodward KB, Coon ME, Cummings C, Cunningham TM, Dossa RG, Brault M, Stokke J, Olsen TM, Gardner K, Estey E, Meshinchi S, Rongvaux A, Bleakley M. CBFB-MYH11 fusion neoantigen enables T cell recognition and killing of acute myeloid leukemia. J Clin Invest. 2020;130:5127–41. [CrossRef]
- Cohen CJ, Gartner JJ, Horovitz-Fried M, Shamalov K, Trebska-McGowan K, Bliskovsky VV, Parkhurst MR, Ankri C, Prickett TD, Crystal JS, Li YF, El-Gamil M, Rosenberg SA, Robbins PF. Isolation of neoantigenspecifc T cells from tumor and peripheral lymphocytes. J Clin Invest. 2015;125:3981–91.
- van der Lee DI, Reijmers RM, Honders MW, Hagedoorn RS, de Jong RC, Kester MG, van der Steen DM, de Ru AH, Kweekel C, Bijen HM, Jedema I, Veelken H, van Veelen PA, Heemskerk MH, Falkenburg J, Grifoen M. Mutated nucleophosmin 1 as immunotherapy target in acute myeloid leukemia. J Clin Invest. 2019;129:774–85. [CrossRef]
- Graf C, Heidel F, Tenzer S, Radsak MP, Solem FK, Britten CM, Huber C, Fischer T, Wolfel T. A neoepitope generated by an FLT3 internal tandem duplication (FLT3-ITD) is recognized by leukemia-reactive autologous CD8+ T cells. Blood. 2007;109:2985–8. [CrossRef]
- Harris DT, Kranz DM. Adoptive T Cell Therapies: A Comparison of T Cell Receptors and Chimeric Antigen Receptors. Trends Pharmacol Sci (2016) 37:220–30. [CrossRef]
- Coulie PG, Van den Eynde BJ, van der Bruggen P, Boon T. Tumour antigens recognized by T lymphocytes: at the core of cancer immunotherapy. Nat Rev Cancer. 2014;14:135–46. [CrossRef]
- Rosenberg SA, Restifo NP. Adoptive cell transfer as personalized immunotherapy for human cancer. Science. 2015;348:62. [CrossRef]
- Kang S, Li Y, Qiao J, Meng X, He Z, Gao X, Yu L. Antigen-Specific TCR-T Cells for Acute Myeloid Leukemia: State of the Art and Challenges. Front Oncol. 2022 Mar 9;12:787108. [CrossRef]
- Xue S, Gillmore R, Downs A, et al. Exploiting T cell receptor genes for cancer immunotherapy. Clin Exp Immunol 2005;139:167–72. [CrossRef]
- Cohen CJ, Zhao Y, Zheng Z, Rosenberg SA, Morgan RA. Enhanced antitumor activity of murine-human hybrid T-cell receptor (TCR) in human lymphocytes is associated with improved pairing and TCR/CD3 stability. Cancer Res. 2006;66:8878–86. [CrossRef]
- Van Loenen MM, de Boer R, Hagedoorn RS, van Egmond EH, Falkenburg JH, Heemskerk MH. Optimization of the HA-1-specific T-cell receptor for gene therapy of hematologic malignancies. Haematologica. (2011) 96:477–81. [CrossRef]
- Poorebrahim, M.; Mohammadkhani, N.; Mahmoudi, R.; Gholizadeh, M.; Fakhr, E.; Cid-Arregui, A. TCR-like CARs and TCR-CARs targeting neoepitopes: An emerging potential. Cancer Gene Ther. 2021, 28, 581–589. [CrossRef]
- Chapuis AG, Egan DN, Bar M, Schmitt TM, McAfee MS, Paulson KG, Voillet V, Gottardo R, Ragnarsson GB, Bleakley M, Yeung CC, Muhlhauser P, Nguyen HN, Kropp LA, Castelli L, Wagener F, Hunter D, Lindberg M, Cohen K, Seese A, McElrath MJ, Duerkopp N, Gooley TA, Greenberg PD. T cell receptor gene therapy targeting WT1 prevents acute myeloid leukemia relapse post-transplant. Nat Med. 2019;25:1064–72. [CrossRef]
- Tawara I, Kageyama S, Miyahara Y, Fujiwara H, Nishida T, Akatsuka Y, Ikeda H, Tanimoto K, Terakura S, Murata M, Inaguma Y, Masuya M, Inoue N, Kidokoro T, Okamoto S, Tomura D, Chono H, Nukaya I, Mineno J, Naoe T, Emi N, Yasukawa M, Katayama N, Shiku H. Safety and persistence of WT1-specifc T-cell receptor gene-transduced lymphocytes in patients with AML and MDS. Blood. 2017;130:1985–94.
- Amir AL, van der Steen DM, van Loenen MM, Hagedoorn RS, de Boer R, Kester MD, de Ru AH, Lugthart GJ, van Kooten C, Hiemstra PS, Jedema I, Grifoen M, van Veelen PA, Falkenburg JH, Heemskerk MH. PRAMEspecifc Allo-HLA-restricted T cells with potent antitumor reactivity useful for therapeutic T-cell receptor gene transfer. Clin Cancer Res. 2011;17:5615–25.
- Marijt WA, Heemskerk MH, Kloosterboer FM, Goulmy E, Kester MG, van der Hoorn MA, van Luxemburg-Heys SA, Hoogeboom M, Mutis T, Drijfhout JW, van Rood JJ, Willemze R, Falkenburg JH. Hematopoiesisrestricted minor histocompatibility antigens HA-1- or HA-2-specifc T cells can induce complete remissions of relapsed leukemia. Proc Natl Acad Sci U S A. 2003;100:2742.
- Tawara I, Kageyama S, Miyahara Y, Fujiwara H, Nishida T, Akatsuka Y, Ikeda H, Tanimoto K, Terakura S, Murata M, Inaguma Y, Masuya M, Inoue N, Kidokoro T, Okamoto S, Tomura D, Chono H, Nukaya I, Mineno J, Naoe T, Emi N, Yasukawa M, Katayama N, Shiku H. Safety and persistence of WT1-specific T-cell receptor gene-transduced lymphocytes in patients with AML and MDS. Blood. 2017 Nov 2;130(18):1985-1994. [CrossRef]
- Morris EC, Tendeiro-Rego R, Richardson R, Fox TA, Sillito F, Holler A, et al. A Phase I Study Evaluating the Safety and Persistence of Allorestricted WT1- TCR Gene Modified Autologous T Cells in Patients With High-Risk Myeloid Malignancies Unsuitable for Allogeneic Stem Cell Transplantation. Blood (2019) 134:1367–7. Doi: 10.1182/blood-2019-128044). [CrossRef]
- van Balen P, Jedema I, van Loenen MM, de Boer R, van Egmond HM, Hagedoorn RS, Hoogstaten C, Veld SAJ, Hageman L, van Liempt PAG, Zwaginga JJ, Meij P, Veelken H, Falkenburg JHF, Heemskerk MHM. HA-1H T-Cell Receptor Gene Transfer to Redirect Virus-Specific T Cells for Treatment of Hematological Malignancies After Allogeneic Stem Cell Transplantation: A Phase 1 Clinical Study. Front Immunol. 2020 Aug 20;11:1804.
- T. Bald, M. F. Krummel, M. J. Smyth, K. C. Barry, The NK cell-cancer cycle: Advances and new challenges in NK cell-based immunotherapies. Nat. Immunol. 21, 835–847 (2020). [CrossRef]
- S. Sivori et al., NK cells and ILCs in tumor immunotherapy. Mol. Aspects Med. 80, 100870 (2021). [CrossRef]
- Jennifer A. Foltz, Melissa M. Berrien-Elliott, David A. Russler-Germain, Carly C. Neal, Jennifer Tran, Margery Gang, Pamela Wong, Matthew Mosior, Jeffrey J. Bednarski, Clare Zimmerman, Celia C. Cubitt, Nancy D. Marin, Alice Y. Zhou, Miriam T. Jacobs, Mark Foster, Timothy Schappe, Ethan McClain, Sweta Desai, Patrick Pence, Michelle Becker-Hapak, Lynne Marsala, Obi L. Griffith, Malachi Griffith, Saad M. Khan, Bryan Fisk, Amanda F. Cashen, Allegra A. Petti, Todd A. Fehniger; Cytokine-Induced Memory-like NK Cells Have a Distinct Single Cell Transcriptional Profile and Persist for Months in Adult and Pediatric Leukemia Patients after Adoptive Transfer. Blood 2021; 138 (Supplement 1): 3825.
- R. Romee et al., Cytokine-induced memory-like natural killer cells exhibit enhanced responses against myeloid leukemia. Sci. Transl. Med. 8, 357ra123 (2016). [CrossRef]
- Dong H, Ham JD, Hu G, Xie G, Vergara J, Liang Y, Ali A, Tarannum M, Donner H, Baginska J, Abdulhamid Y, Dinh K, Soiffer RJ, Ritz J, Glimcher LH, Chen J, Romee R. Memory-like NK cells armed with a neoepitope-specific CAR exhibit potent activity against NPM1 mutated acute myeloid leukemia. Proc Natl Acad Sci U S A. 2022 Jun 21;119(25):e2122379119. [CrossRef]
- Pomeroy, E.J.; Hunzeker, J.T.; Kluesner, M.G.; Lahr, W.S.; Smeester, B.A.; Crosby, M.R.; Lonetree, C.-l.; Yamamoto, K.; Bendzick, L.; Miller, J.S.; et al. A Genetically Engineered Primary Human Natural Killer Cell Platform for Cancer Immunotherapy. Mol. Ther. 2020, 28, 52–63). [CrossRef]
- Liu, E.; Marin, D.; Banerjee, P.; Macapinlac, H.A.; Thompson, P.; Basar, R.; Nassif Kerbauy, L.; Overman, B.; Thall, P.; Kaplan, M.; et al. Use of CAR-Transduced Natural Killer Cells in CD19-Positive Lymphoid Tumors. N. Engl. J. Med. 2020, 382, 545–553. [CrossRef]
- Albinger, N., Pfeifer, R., Nitsche, M. et al. Primary CD33-targeting CAR-NK cells for the treatment of acute myeloid leukemia. Blood Cancer J. 12, 61 (2022). [CrossRef]
- Christodoulou I, Ho WJ, Marple A, et al Engineering CAR-NK cells to secrete IL-15 sustains their anti-AML functionality but is associated with systemic toxicities Journal for ImmunoTherapy of Cancer 2021;9:e003894. ).
- Anthony G. Mansour, Kun-Yu Teng, Zhiyao Li, Zheng Zu, Hanyu Chen, Aliya Ali, Jianying Zhang, Ting Lu, Shoubao Ma, Michael A. Caligiuri, Jianhua Yu; Abstract LB102: Off-the-shelf cord blood FLT3 CAR-NK cells for immunotherapy of acute myeloid leukemia. Cancer Res 15 June 2022; 82 (12_Supplement): LB102.
- Ruihao Huang, Qin Wen, Xiaoqi Wang, Hongju Yan, Yingying Ma, Wang Mai-Hong, Xiao Han, Li Gao, Lei Gao, Cheng Zhang, Xi Zhang; Off-the-Shelf CD33 CAR-NK Cell Therapy for Relapse/Refractory AML: First-in-Human, Phase I Trial. Blood 2022; 140 (Supplement 1): 7450–7451.).
- You, L.; Han, Q.; Zhu, L.; Zhu, Y.; Bao, C.; Yang, C.; Lei, W.; Qian, W. Decitabine-Mediated Epigenetic Reprograming Enhances Anti-leukemia Efficacy of CD123-Targeted Chimeric Antigen Receptor T-Cells. Front. Immunol. 2020, 11, 1787. [CrossRef]
- Guzman ML, Sugita M, Zong H, Ewing-Crystal N, Trujillo-Alonso V, et al. Allogeneic Tcra/b Deficient CAR T-Cells Targeting CD123 Prolong Overall Survival of AML Patient-Derived Xenografts. Blood (2016) 128 (22):765.
- Cui, Q., Qian, C., Xu, N. et al. CD38-directed CAR-T cell therapy: a novel immunotherapy strategy for relapsed acute myeloid leukemia after allogeneic hematopoietic stem cell transplantation. J Hematol Oncol 14, 82 (2021). [CrossRef]
- Sugita, M., Galetto, R., Zong, H. et al. Allogeneic TCRαβ deficient CAR T-cells targeting CD123 in acute myeloid leukemia. Nat Commun 13, 2227 (2022). [CrossRef]
- Celectis Reports Clinical Hold of UCART 123 Studies, published on September 04, 2017, New York (N.Y.), https://www.cellectis.com/en/press/cellectis-reports-clinical-hold-of-ucart123-studies.
- Hudecek M, Ivics Z. Non-viral therapeutic cell engineering with the Sleeping Beauty transposon system. Curr Opin Genet Dev 2018;52:100–8. https://doi.org/ 10.1016/j.gde.2018.06.003. [CrossRef]
- Costello CL, Gregory TK, Ali SA, Berdeja JG, Patel KK, Shah ND, et al. Phase 2 Study of the Response and Safety of P-Bcma-101 CAR-T Cells in Patients with Relapsed/ Refractory (r/r) Multiple Myeloma (MM) (PRIME). Blood 2019;134:3184. https:// doi.org/10.1182/blood-2019-129562.
- Magnani CF, Gaipa G, Lussana F, Belotti D, Gritti G, Napolitano S, et al. Sleeping Beauty-engineered CAR T cells achieve antileukemic activity without severe toxicities. J Clin Invest 2020;130:6021–33. [CrossRef]
- Prommersberger S, Reiser M, Beckmann J, Danhof S, Amberger M, Quade-Lyssy P, et al. CARAMBA: a first-in-human clinical trial with SLAMF7 CAR-T cells prepared by virus-free Sleeping Beauty gene transfer to treat multiple myeloma. Gene Ther 2021;28:560. [CrossRef]
- Gurney M, O’Reilly E, Corcoran S, Brophy S, Krawczyk J, Otto NM, Hermanson DL, Childs RW, Szegezdi E, O’Dwyer ME. Concurrent transposon engineering and CRISPR/Cas9 genome editing of primary CLL-1 chimeric antigen receptor-natural killer cells. Cytotherapy. 2022 Nov;24(11):1087-1094. Doi: 10.1016/j.jcyt.2022.07.008. [CrossRef]
- Daver N, Basu S, Garcia-Manero G, Cortes J, Ravandi E, Jabbour E, et al. Phase IB/II study of nivolumab in combination wth azacytidine in patients with relapsed acute myeloid leukemia. Blood. (2016) 22:763. doi: 10.1182/blood.V128.22.763.763 39. [CrossRef]
- Davids M, Kim H, Bachireddy P, Costello C, Liguori R, Savell A, et al. Ipilimumab for patients with relapse after allogeneic transplantation. N Engl J Med. (2016) 375:143–53. doi: 10.1056/NEJMoa1601202. [CrossRef]
- Kadia T, Cortes J, Ghorab A, Ravandi F, Jabbour E, Daver N, et al. Nivolumab maintenance in high-risk acute myeloid leukemia patients. J Clin Oncol. (2018) 36:15. doi: 10.1200/JCO.2018.36.15_suppl.7014. [CrossRef]
- Epperly R, Gottschalk S, Velasquez MP. A Bump in the Road: How the Hostile AML Microenvironment Affects CAR T Cell Therapy. Front Oncol. 2020 Feb 28;10:262.). [CrossRef]
- Jitschin R, Saul D, Braun M, Tohumeken S, Volkl S, Kischel R, et al. CD33/CD3-bispecific T-cell engaging (BiTE R ) antibody construct targets monocytic AML myeloid-derived suppressor cells. J Immunother Cancer. (2018) 6:116. doi: 10.1186/s40425-018-0432-9). [CrossRef]
- Zhou Q, Bucher C, Munger M, Highfill S, Tolar J, Munn D, et al. Depletion of endogenous tumor-associated regulatory T cells improves the efficacy of adoptive cytotoxic T-cell immunotherapy in murine acute myeloid leukemia. Blood. (2009) 114:3793–802. doi: 10.1182/blood-2009-03-208181. [CrossRef]
- Suryadevara C, Desai R, Farber S, Choi B, Swartz A, Shen S, et al. Preventing Lck activation in CAR T cells confers Treg resistance but requires 4-1BB signaling for them to persist and treat solid tumors in nonlymphodepleted hosts. Clin Cancer Res. (2019) 25:358– 68. doi: 10.1158/1078-0432.CCR-18-1211. [CrossRef]
- Ninomiya S, Narala N, Huye L, Yagyu S, Savoldo B, Dotti G, Heslop HE, Brenner MK, Rooney CM, Ramos CA. Tumor indoleamine 2,3-dioxygenase (IDO) inhibits CD19-CAR T cells and is downregulated by lymphodepleting drugs. Blood. 2015 Jun 18;125(25):3905-16. [CrossRef]
- Perrot I, Michaud HA, Giraudon-Paoli M, Augier S, Docquier A, Gros L, Courtois R, Déjou C, Jecko D, Becquart O, Rispaud-Blanc H, Gauthier L, Rossi B, Chanteux S, Gourdin N, Amigues B, Roussel A, Bensussan A, Eliaou JF, Bastid J, Romagné F, Morel Y, Narni-Mancinelli E, Vivier E, Paturel C, Bonnefoy N. Blocking Antibodies Targeting the CD39/CD73 Immunosuppressive Pathway Unleash Immune Responses in Combination Cancer Therapies. Cell Rep. 2019 May 21;27(8):2411-2425.e9.
- Minnie SA, Waltner OG, Ensbey KS, Nemychenkov NS, Schmidt CR, Bhise SS, Legg SRW, Campoy G, Samson LD, Kuns RD, Zhou T, Huck JD, Vuckovic S, Zamora D, Yeh A, Spencer A, Koyama M, Markey KA, Lane SW, Boeckh M, Ring AM, Furlan SN, Hill GR. Depletion of exhausted alloreactive T cells enables targeting of stem-like memory T cells to generate tumor-specific immunity. Sci Immunol. 2022 Oct 21;7(76):eabo3420. doi: 10.1126/sciimmunol.abo3420. [CrossRef]
- Antonio Pierini, Loredana Ruggeri, Alessandra Carotti, Franca Falzetti, Simonetta Saldi, Adelmo Terenzi, Claudio Zucchetti, Gianluca Ingrosso, Tiziana Zei, Roberta Iacucci Ostini, Sara Piccinelli, Samanta Bonato, Sara Tricarico, Antonella Mancusi, Sara Ciardelli, Roberto Limongello, Mara Merluzzi, Mauro Di Ianni, Rita Tognellini, Olivia Minelli, Cristina Mecucci, Maria Paola Martelli, Brunangelo Falini, Massimo Fabrizio Martelli, Cynthia Aristei, Andrea Velardi; Haploidentical age-adapted myeloablative transplant and regulatory and effector T cells for acute myeloid leukemia. Blood Adv 2021; 5 (5): 1199–1208.
- Pan J, Yang JF, Deng BP, Zhao XJ, Zhang X, Lin YH, et al. High efficacy and safety of low-dose CD19-directed CAR-T cell therapy in 51 refractory or relapsed B acute lymphoblastic leukemia patients. Leukemia 2017;31:2587–93. [CrossRef]
- Hay KA, Gauthier J, Hirayama AV, Voutsinas JM, Wu Q, Li D, et al. Factors associated with durable EFS in adult B-cell ALL patients achieving MRD-negative CR after CD19 CAR T-cell therapy. Blood 2019;133:1652–63. [CrossRef]
- Carlos A. Ramos. Should CD19 CAR-T Cells for ALL be Followed by Allogeneic Stem Cell Transplant? Transplantation and Cellular Therapy, Volume 28, Issue 1, 2022, Pages 1-2, ISSN 2666-6367.
- Summers C,Wu QV, Annesley C, et al.Hematopoietic Cell Transplantation after CD19 Chimeric Antigen Receptor T Cell-Induced Acute Lymphoblastic Lymphoma Remission Confers a Leukemia-Free Survival Advantage.Transplant Cell Ther. 2021; 28: 19-27.
- Zhang, H., Bu, C., Peng, Z. et al. Characteristics of anti-CLL1 based CAR-T therapy for children with relapsed or refractory acute myeloid leukemia: the multi-center efficacy and safety interim analysis. Leukemia 36, 2596–2604 (2022). [CrossRef]
- Yao S, Jianlin C, Yarong L, Botao L, Qinghan W, Hongliang F, Lu Z, Hongmei N, Pin W, Hu C, Liangding H, Bin Z. Donor-Derived CD123-Targeted CAR T Cell Serves as a RIC Regimen for Haploidentical Transplantation in a Patient With FUS-ERG+ AML. Front Oncol. 2019 Dec 3;9:1358. [CrossRef]
- Zhang, H., Bu, C., Peng, Z. et al. Characteristics of anti-CLL1 based CAR-T therapy for children with relapsed or refractory acute myeloid leukemia: the multi-center efficacy and safety interim analysis. Leukemia 36, 2596–2604 (2022). [CrossRef]
- Budde L, Song JY, Kim Y, Blanchard S, Wagner J, Stein AS, et al. Remissions of acute myeloid leukemia and blastic plasmacytoid dendritic cell neoplasmfollowing treatmentwith Cd123- specific car T cells: A first-in-Human clinical trial. Blood (2017) 130(Suppl_1):811–. Doi: 10.1182/ blood.V130.Suppl_1.811.811. [CrossRef]
- Tambaro FP, Singh H, Jones E, Rytting M, Mahadeo KM, Thompson P, Daver N, DiNardo C, Kadia T, Garcia-Manero G, Chan T, Shah RR, Wierda WG. Autologous CD33-CAR-T cells for treatment of relapsed/refractory acute myelogenous leukemia. Leukemia. 2021 Nov;35(11):3282-3286. [CrossRef]
- Wang QS, Wang Y, Lv HY, Han QW, Fan H, Guo B, et al. Treatment of CD33-Directed Chimeric Antigen Receptor-Modified T Cells in One Patient With Relapsed and Refractory Acute Myeloid Leukemia. Mol Ther (2015) 23 (1):184–91. [CrossRef]
- Cui, Q., Qian, C., Xu, N. et al. CD38-directed CAR-T cell therapy: a novel immunotherapy strategy for relapsed acute myeloid leukemia after allogeneic hematopoietic stem cell transplantation. J Hematol Oncol 14, 82 (2021). [CrossRef]
- Ritchie DS, Neeson PJ, Khot A, Peinert S, Tai T, Tainton K, et al. Persistence and Efficacy of Second Generation CAR T Cell Against the LeY Antigen in Acute Myeloid Leukemia. Mol Ther (2013) 21(11):2122–9. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



