
Submitted:
16 December 2025
Posted:
17 December 2025
You are already at the latest version
Abstract
Background/objective: The intensive use of pesticides in modern agriculture has raised growing concerns about their potential adverse effects on human health. Exposure to these compounds has been linked to a variety of negative outcomes. This study aims to evaluate and synthesize the available scientific evidence on the impact of pesticide exposure in agricultural production on human health. It will integrate results from experimental and observational studies conducted on ani-mals and humans. Results: Findings show that exposure, particularly during the prenatal period, is associated with multisystemic alterations affecting fetal de-velopment, metabolism, the immune system, the nervous system, and the gut mi-crobiome. Evidence from animal model studies complements human data, providing insight into the underlying biological mechanisms, such as oxidative stress, liver dysfunction, alterations in hormonal signaling and activation of the inflammatory response. Conclusions: Public health strategies must be imple-mented to reduce pesticide exposure, strengthen environmental and mater-nal-child surveillance, and promote research into the modulation of the gut micro-biome and systemic inflammation. This will help to prevent and mitigate the met-abolic, immunological, and neurodevelopmental effects associated with these compounds.
Keywords:
pesticide exposure
; human health
; gut microbiome
; metabolic disorders
; inflammation
; neurodevelopmental effects
; immune disorders
1. Introduction
Pesticides are chemical substances, either natural or synthetic, that are used to eradicate pests and insects. They are essential for improving crop production in agricultural processes. They promote crop yields and protect crops from disease and damage. Pesticides include herbicides, insecticides, fungicides, molluscicides, ovicides, acaricides, rodenticides and nematicides. The most used are fungicides, insecticides and herbicides [1].
The extensive use of pesticides is one of the main sources of toxic compound exposure in the general population, posing a significant environmental toxicology problem [2,3].
These chemical agents are designed to eliminate organisms that are considered harmful. However, they can interfere with multiple biological processes in humans, particularly if exposure is chronic or involves a mixture of different products [4].
Various studies have demonstrated that the action of pesticides occurs via mechanisms such as oxidative stress, altered cholinergic neurotransmission, endocrine disruption, and genotoxic damage [5,6,7].
The consequences of this exposure are wide-ranging. They can include acute manifestations, such as respiratory or neuromuscular poisoning, as well as chronic effects associated with neurodegenerative pathologies and reproductive disorders [4,8].
Studies have also documented the cumulative toxic effects of pesticides on health and on non-target organisms. They have also shown that pesticide bioaccumulate and are associated with endocrine disruption and chronic diseases in exposed human populations [8,9].
Advances in molecular biology and microbiology have led to the recognition that intestinal permeability and the gut microbiota play a key role in systemic health and the development of various diseases. Gut microbiota play a fundamental role in maintaining homeostasis and human health by performing protective, structural and metabolic functions that are essential for physiological balance. These include the production of bioactive metabolites, immune regulation and the maintenance of epithelial barrier integrity [10,11].
The intestinal mucosal layer acts as the body's first line of defense, balancing mucus secretion and bacterial degradation to limit exposure to antigens and pro-inflammatory molecules. Additionally, certain bacterial communities reinforce the tight junctions between epithelial cells, thereby preventing macromolecules and endotoxins from entering the bloodstream [12].
Its main contributions include modulating the immune system, regulating the gut-brain axis, synthesizing vitamins and bioactive metabolites, protecting the intestinal barrier, and facilitating peristaltic transit [13,14].
Through these mechanisms, the gut microbiota influences the immunoinflammatory response, energy metabolism, and neuroendocrine communication, establishing itself as a determining factor in the overall health of the individual [15].
Alteration of the gut microbiota (dysbiosis) has been associated with increased susceptibility to gastrointestinal, metabolic, cardiovascular and neurodegenerative diseases, as well as autoimmune and neoplastic diseases. Microbial imbalance can contribute to both systemic inflammation and immune dysfunction [16,17] and pathologies of toxic origin [18].
In this regard, microbiota not only acts as a modulator of digestive health, but also as a mediator of the toxic response to environmental xenobiotics, by participating in their biotransformation and partial detoxification, which is an emerging area of great interest for contemporary toxicology [19,20,21,22,23,24].
Various studies have documented the disruptive effects of organophosphate, carbamate and neonicotinoid pesticides on the intestinal epithelium and microbiota [25,26,27].
These agents can alter the structure of bacterial communities, change intestinal permeability and create a pro-inflammatory state linked to oxidative stress and mitochondrial dysfunction [28,29].
These alterations are linked not only to an increased risk of chronic diseases in adults, but also to transgenerational effects, given that exposure during pregnancy can negatively impact fetal development through epigenetic and microbiological mechanisms [30,31,32].
Although regulations have been implemented to reduce exposure levels, the persistence of certain organochlorine compounds and the use of pesticides in poorly controlled environments continue to pose significant risks to public health [33].
In this context, this article systematically reviews the available scientific evidence on the effects of pesticides on human and animal health, analyzing the main toxicological mechanisms involved and their clinical repercussions.
The objective of this systematic review is to evaluate and synthesize the available scientific evidence on the effects of exposure to pesticides used in agricultural production on human health, integrating results from experimental and observational studies—both in humans and in animal models with biological and physiological similarities—with special attention to physiological, neurological and metabolic alterations and disruption of the gut microbiota. It also aims to identify the main toxicological mechanisms involved and their possible relationship with the development of chronic diseases, to provide an updated and well-founded overview to guide future research and public health policies.
The PICO format was used to establish the research question.
P (Population/Participants): Humans exposed to pesticides used in agricultural production, as well as animal models with genetic, biological or behavioral similarities to humans.
I (Intervention/Exposure): Acute or chronic exposure to pesticides, whether through occupational, environmental or dietary contact.
C (Comparator): Populations or animal models not exposed or with minimal levels of exposure to pesticides.
O (Outcomes): Physiological, neurological and metabolic alterations and changes in the gut microbiota, including modifications in the permeability of the intestinal barrier and its relationship with the development of chronic diseases.
Thus, the research question is as follows: What are the physiological, neurological, metabolic, and microbiological effects associated with exposure to agricultural pesticides in humans and comparable animal models, according to the available scientific evidence?
In order to delimit and systematize the scientific evidence available regarding this question, the use of systematic reviews is proposed. These types of studies constitute a rigorous and objective summary of the existing scientific literature on a given topic, integrating multiple individual studies into a single document that has been critically evaluated by experts. Systematic reviews also provide the best available evidence to inform decisions in clinical practice.
2. Materials and Methods
2.1. Research Design
A systematic review of the available scientific literature was conducted, following the guidelines of the PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-Analyses) statement, with the aim of ensuring transparency, reproducibility, and methodological rigor in the identification, selection, and analysis of studies.
The process included several clearly defined phases: initially, an exhaustive search was carried out in relevant scientific databases, using keywords and MeSH terms related to pesticides, human exposure and health effects. Subsequently, predefined inclusion and exclusion criteria were applied, selecting studies that provided direct evidence on the relationship between exposure to pesticides and physiological, metabolic or neurological alterations in humans and animal models.
Once the eligible studies were identified, relevant data were extracted, including information on the pesticide, study design, population, health alterations, outcome variables, main findings, and conclusions. All studies underwent a critical quality assessment, considering factors such as risk of bias, internal and external validity, and consistency of reported results.
Finally, the results were synthesized in a structured manner, integrating quantitative and qualitative evidence to provide a comprehensive and objective view of the impact of pesticides on human and animal health, and to inform recommendations for public health prevention and risk mitigation of developing diseases related to the consumption and exposure to these substances.
2.2. Search Strategy
A comprehensive search of scientific literature published between 2018, and the present day was conducted in the Medline/PubMed, Embase and Web of Science databases. The aim was to compile the most relevant information available on the relationship between exposure to pesticides and health. To ensure consistency in the evaluation and analysis of the evidence, only articles published in English were considered.
Initially, the search was conducted by combining key concepts with the Boolean operators AND and OR. The initial search algorithm was established with the terms “pesticides”, “disease” and “illness”, resulting in the following syntax: (“pesticides”) AND (“disease” OR “illness”).
As no sufficiently relevant results were found, the search was directed towards literature related to gut microbiota, using the syntax: (“pesticides”) AND (“disease” OR “illness” OR “gut microbiota”). This strategy generated many results, although they were very heterogeneous in relation to the topic of interest. Finally, the search was refined to focus specifically on the relationship between pesticides and gut microbiota, using the final syntax: (“pesticides”) AND (“gut microbiota”).
To ensure the highest quality and level of scientific evidence, only systematic reviews corresponding to Level I evidence were included. These reviews integrate the findings of all relevant randomized clinical trials and have been critically reviewed by experts in the field.
This review was conducted in two phases, from late June to late August 2024 and from late January to mid-May 2025. A systematic and structured procedure was followed to ensure the results obtained were comprehensive and valid.
2.3. Eligibility Criteria
The inclusion criteria were as follows:
- Articles that were systematic reviews (Level I)
- Systematic reviews written in English
- Systematic reviews with full text available
- Systematic reviews relating to health alterations associated with pesticides use
- Systematic reviews of experimental studies conducted with rats and mice
The exclusion criteria were as follows:
- Systematic reviews containing evidence relating to persistent organic pollutants
- Systematic reviews containing evidence relating to metals
- Systematic reviews containing hormone-disrupting substances
- Systematic reviews containing plastic substances
- Systematic reviews containing other substances not found among the group of pesticides used in traditional agriculture
- Systematic reviews conducted on birds or animals that differ greatly from the characteristics of the human organism
2.4. Article Selection
First, an initial search was conducted using the algorithms described in the 'Search strategy' section above. The aim was to estimate the volume of existing publications and compile the most relevant information to answer the research question.
Next, the articles relevant to this study were identified using the established inclusion and exclusion criteria. This phase involved reviewing titles and abstracts to determine the relevance of each publication.
Subsequently, a more detailed screening was carried out by accessing the full text of the preselected articles.
Each document was read to assess its suitability, with those that did not fit the central theme of the review or lacked relevant conclusions being discarded.
Finally, the studies included in the review were selected in accordance with the methodological recommendations of Galarza and Cruz (2024) [34] to ensure rigor and consistency in the selection process.
2.5. Data Extraction
The following variables were extracted from the selected articles as relevant: authors, year of publication, pesticide, sample and/or type of study, results and conclusions. Only relevant information was considered, and duplication of findings was avoided.
3. Results
3.1. Results of the Study Selection
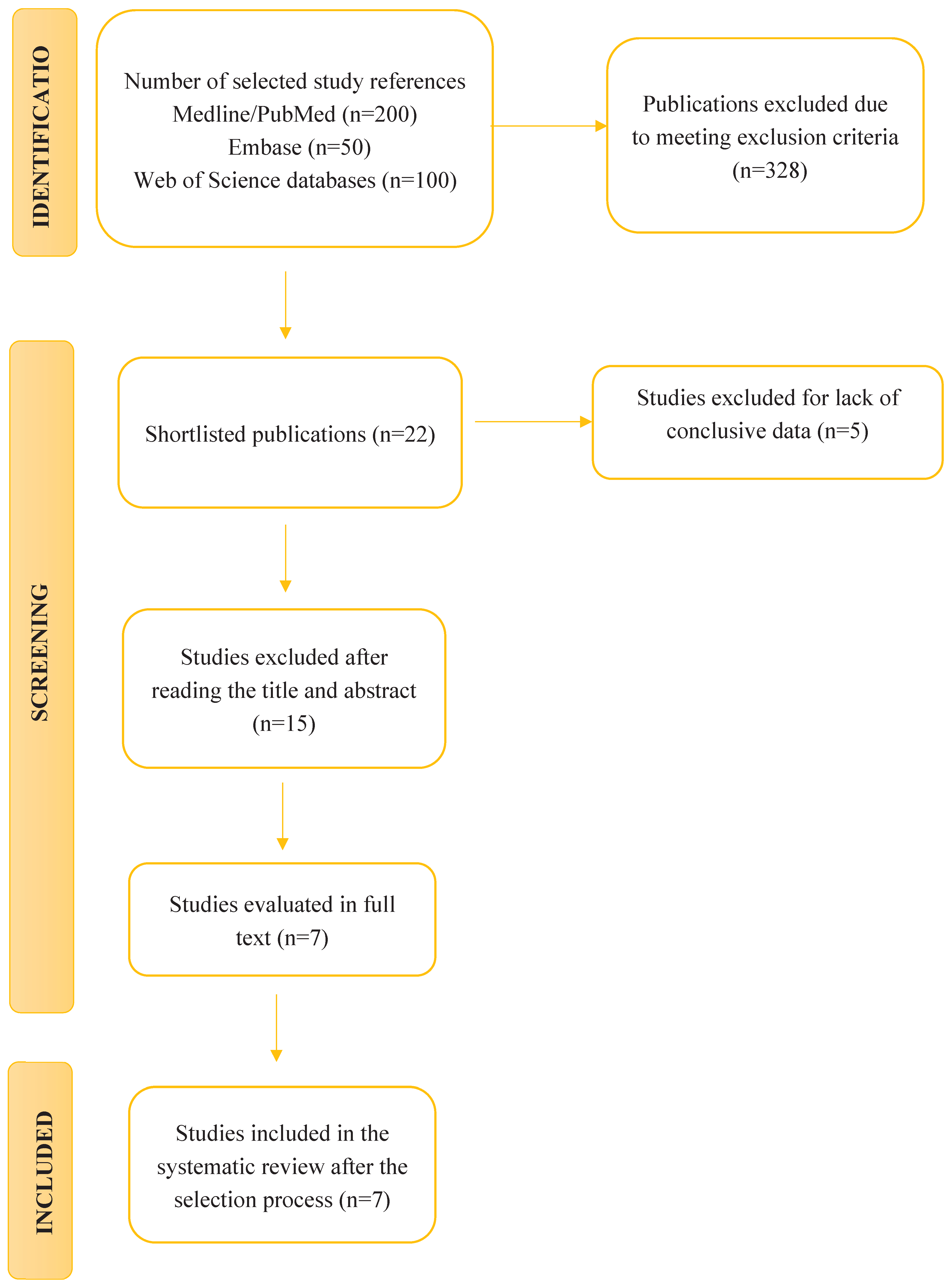
The initial search using the algorithm (“pesticides”) AND (“disease” OR “illness” OR “gut microbiota”) produced 10.850 results. This was then refined using the syntax “pesticides” AND “gut microbiota”, reducing the number of studies identified to 350 articles. After applying the previously defined eligibility criteria, 22 studies were selected for review.
These articles were then screened in detail, with 10 being excluded for not precisely matching the theme of this study and 5 for lacking conclusive results. Consequently, seven articles were finally included in the systematic review. The entire selection process is represented in the corresponding flow chart (Figure 1).
Of the seven studies selected, three were conducted in the United States and four in Europe. The main characteristics and most relevant data from these studies are presented in Table 1.
3.2. Results of the Effects of Pesticides Exposure on Humans
Research conducted on human populations has shown that exposure to pesticides during pregnancy can have adverse effects on fetal development, including, among other things, a decrease in birth weight. A significant association has also been identified between pregnant women living less than 2.000 meters from areas where these compounds are used and an increased risk of their offspring developing autism spectrum disorders. This risk intensifies depending on the degree of exposure, being particularly notable during the second and third trimesters of pregnancy, with an estimated increase of 30%. Similarly, an inverse relationship has been described between prenatal exposure to pesticides and child neuropsychological development, manifested in the appearance of autistic traits in 11-year-old children who were exposed in utero.
Furthermore, scientific evidence indicates that exposure to these chemicals is associated with various metabolic disorders, such as insulin resistance, type 2 diabetes mellitus, dyslipidemia, dysfunction in the biotransformation of chemical compounds, atherosclerosis, obesity and hepatic lipid metabolism disorders. Likewise, the existence of intestinal dysbiosis linked to the accumulation of adipose tissue has been documented. Added to this is the activation of the inflammatory response, mediated by the release of pro-inflammatory cytokines, together with alterations in the functionality of the immune system.
In relation to the intestinal microbiome, exposure to pesticides has been observed to cause microbial imbalances and dysbiosis, leading to increased intestinal barrier permeability and generating multiple adverse effects on individual health.

Table 1.
Extraction of relevant data and main results from systematic review articles.
| AUTHOR YEAR |
PESTICIDE | SAMPLE AND/OR TYPE OF STUDY | CHANGES IN THE INTESTINAL MICROBIOME/BRAIN DISORDERS | RESULTS | CONCLUSIONS |
|---|---|---|---|---|---|
|
Yue, Y. et al. 2024 |
Insecticide (β-HCH) | Human mothers | Weight loss in offspring. | This study concludes that prenatal exposure to pesticides induces changes in the gut microbiome that lead to intestinal dysbiosis, impacting health and behavior, affecting key bacterial strains and contributing to conditions such as weight fluctuations, energy homeostasis and neurobehavioral symptoms like autism. Low body weight and modifications in bacterial genes associated with carbohydrate and lipid metabolism have been identified, as well as behavioral abnormalities in offspring. Obesity, metabolic disorders, alterations in immunity and inflammation have also been observed. These findings underscore the importance of investigating the effects of pesticides on the microbiome and their role in the development of metabolic and neurological diseases. | |
| Insecticide (Mecarbam) | Human mothers | Weight loss in offspring. | |||
| Herbicide (Ammonium glufosinate) | Father mice | Behavioral abnormalities. | |||
| Insecticide (Combination of boscalid, captan, chlorpyrifos, thiacloprid, thiophanate, and ziram) |
Father mice | Obesity and metabolic disorders. | |||
| Insecticide (Chlorpyrifos) | Rat offspring | Hyperlipidemia and hyperglycemia in female offspring. Changes in rat behavior when faced with new situations. Changes in glutamine function and GABA signaling in the amygdala. |
|||
| Mouse offspring | Interference with developing neurones. | ||||
| Insecticide (Nitenpyram | Parent mice | Decrease in serum glucose in female offspring. | |||
| Fungicide (Procymidone) | Progenitor rats | Metabolic disorders. Neurological impairments throughout life. | |||
| Parent mice | Metabolic disorders. Neurodevelopmental disorders in sex-dependent offspring. | ||||
| Insecticide (Fenvalerate) | Parent mice | Increased intrauterine wet weight. Increased height of luminal epithelial cells. Increased LH. | |||
| Insecticide (Dichlorodiphenyltrichloroethane) | Parent mice | Neurodevelopmental defects in male mice. | |||
| Insecticide (Cypermethrin) | Parent mice | Hearing impairments that develop slowly over time. | |||
| Fungicide (Triticonazole) | Progenitor rats | Alteration of endocrine effects. Changes in the genome transcription of the external genitalia of the male fetus. |
|||
| Fungicide (Flusilazole) | Progenitor rats | Alteration of endocrine effects. | |||
| Insecticide (Chlordecone) | Parent mice | Defects and reduction in sperm count. | |||
| Herbicide (Glyphosate) | Progenitor rats | Prostate disease, obesity, kidney disease, ovarian disease, and birth abnormalities. Changes related to inflammation and oxidative stress genes in the cortex and cerebellum of offspring. |
|||
| Herbicide (paraquat) | Baby mice | Increased weight in adults among descendants. | |||
| Herbicide (ammonium glufosinate) | Parent mice | Abnormal behaviour. | |||
| Insecticide (permethrin) | Baby mouse | Negative impact on gut microbiota | |||
| Insecticide (endosulfan) | Parent mice | Metabolic disorders. Obesity. | |||
| Insecticide (chlorpyrifos) | Parent rats | Negative impact on gut microbiota Profound changes in the microbiome of the caecum |
Bacterial translocation in the liver and spleen. Lower birth rate. | ||
| Mouse offspring | egative impact on gut microbiota | Dysbiosis at an early age in the intestinal membrane. | |||
| Rat offspring | Microbiome modifications | Hyperlipidemia in female offspring. Hypoglycemic alterations in female offspring. Alterations in rat behavior in new situations. Reduced immune response in females. Asthma. | |||
| Insecticide (Nitenpyram) | Parent mice | Decrease in blood glucose in female offspring. | |||
|
Yang, Y. et al. 2023 |
Glyphosate | Case-control study of pregnant women | Increased risk of autism spectrum disorder in offspring of pregnant women living less than 2000 meters from places where herbicides are used. | This study establishes a link between alterations in the gut microbiome and autism spectrum disorder. Exposure to pesticides causes dysbiosis in gut microbiota and neurodevelopment, contributing to neurological defects and behavioral alterations. In summary, dysbiosis of the gut microbiome is a crucial factor in the symptoms of autism spectrum disorder related to pesticides exposure. |
|
| Sprague-Dawley rats | Changes in maternal behavior. Changes in neural plasticity. | Perinatal exposure leads to changes in the behavior of rat offspring. | |||
| ddY mice | Cognitive impairment. Social interaction impairment | Behavioral abnormalities like autism spectrum disorder in the offspring of male mice. | |||
| Swiss mice | Deficit in social interaction. Repetitive stereotypical behavior. Morphological changes in glial cells residing in the brain. Reduction in blood-brain barrier permeability. Alteration in acetylcholinesterase activity. | ||||
| Chlorpyrifos | Case-control study of pregnant women | Exposure in mothers positively correlates with autism spectrum disorder in offspring. | |||
| Cohort study pregnant women |
Inverse association between prenatal exposure and domain-specific neuropsychological development in children at 12 months. | ||||
| Cohort study pregnant women |
Increased autistic traits in 11-year-old children with prenatal exposure. | ||||
| Case-control study of pregnant women | Increased risk of autism spectrum disorder in offspring of mothers exposed during the second or third trimester of pregnancy. | ||||
| Case-control study of pregnant women | Higher risk of autism spectrum disorder with greater exposure during pregnancy. | ||||
| Sprague-Dawley rats | Existence of behaviors typical of autism spectrum disorder phenotypes (impaired social communication and confined and repetitive behavior). | ||||
| BTBR mice | Exposure during prenatal development promotes the existence of behavioral traits typical of autism spectrum disorder, including impairments in social and communication domains (alterations in ultrasonic vocalization and high levels of repetitive behaviors). | ||||
| Wistar rats | Hypermobility and stress-related hypermobility. Hypo- or hypersensitisation of the cholinergic and GABAergic systems. Increased transcription of the GABA-A-A2 subunit and M2 receptor genes. Inhibition of acetylcholinesterase activity. Stimulation of pituitary hormone release. Systemic inflammation (TNFR). Decreased responsiveness to social novelty in adulthood. | Communication deficits similar to those seen in autism spectrum disorder. | |||
| C57BL/6 mice | Exposure during prenatal development is associated with long-term negative effects on social behavior and decreased exploration of unfamiliar items. | ||||
| C57BL/6 mice | Exposure during prenatal development caused impairments in social behavior and excitatory-inhibitory balance. | ||||
| Fmr1-KO rats | Exposure during development led to exacerbation of a phenotype like autism spectrum disorder. | ||||
| Pyrethroids | Case-control study of pregnant women | Exposure during the third trimester was associated with an increased likelihood of presenting symptoms of autism spectrum disorder. | |||
| Cross-sectional study of pregnant women | Higher levels of pyrethroids in urine were associated with an increased risk of autism spectrum disorder in offspring. | ||||
| Case-control study pregnant women |
Prenatal exposure was associated with an increased risk of autism spectrum disorder. | ||||
| Wistar rats | Loss of dopaminergic neurons in the substantia nigra. | ||||
| C57BL/6 mice | Neuronal inflammation. | ||||
| Imidacloprid |
Case-control study pregnant women |
Increased risk of autism spectrum disorder in offspring in 30% of cases. | |||
| Diazinon | C57BL/6 mice | Decreased regulation of neurotransmitters. | |||
| Glufosinate ammonium |
ICR mice | Impaired motor activity. Behaviors similar to autism spectrum disorder. Impaired short-term memory formation. | |||
|
Gambarte, P. C. K. & Wolansky, M. J. 2022 |
Organophosphate insecticides (Chlorpyrifos) |
Kunming male mice | Alteration of the composition of the microbiota and metabolic pathways | This study has confirmed the existence of alterations in the gut microbiome that lead to a reduction in beneficial bacteria and an increase in harmful bacteria, causing intestinal dysbiosis. Obesity, diabetes, alterations in the inflammatory response, changes in the genome, and morphological and functional changes in the intestine have been identified as a consequence of pesticides use. Alterations in gene expression related to metabolic pathways, glucose intolerance, and pesticide biotransformation have also been demonstrated. | |
| Male Wistar rats | Alteration of the composition of the microbiota. Increase in the abundance of opportunistic pathogens. |
Microbiota alterations are associated with obesity, diabetes phenotypes, and alterations in pancreatic islet cells. Alteration of the mechanism responsible for controlling the inflammatory response. Micro- and macrostructural alterations in the right intestine. | |||
| Organophosphate insecticides (Diazinon) | C57BL/6 mice | Changes in the composition of the gut microbiome. | Changes in the functional metagenome and metabolic pathways. Differences are observed depending on the sex of the mouse. | ||
| Organophosphate insecticides (Monochrotophos) | Rats, CFT-Wistar | Changes in the composition of the gut microbiome. | Functional and morphological changes in the intestine. | ||
| Mice, BALB/c | Changes in the composition of the gut microbiome. | Changes in the expression of genes related to metabolic pathways, glucose intolerance and pesticide biotransformation. | |||
| Carbamate insecticide (Aldicarb) | C57BL/6 mice | Alteration of the composition of the microbiota. | Specific changes throughout exposure in the microbiome for each genus of bacteria. | ||
| Pyrethroid insecticide (Permethrin) | Male Wistar rats and lactating offspring | Alteration of the composition of the microbiota. | Reduction in beneficial bacterial genera and increase in non-beneficial bacterial genera compared to the control. | ||
| Systemic fungicide (Propamocarb) |
ICR male mice | Changes in the composition of the gut microbiome 7 days after the start of oral exposure. | |||
| Fungicide carbamate benzimidazole (Carbendazim) |
ICR male mice | Alteration of the microbiome composition after 7 days. | Increase in harmful bacteria and decrease in beneficial bacteria. | ||
| Male C57BL/6 mice | Alteration of the microbiome composition after 7 days. | Increases and decreases in relative abundance depending on bacterial gender (beneficial and harmful). | |||
| Fungicide triazole compounds (Epoxiconazole) |
Female Sprague-Dawley rats | Alteration of the microbiome composition. | Increases and decreases in relative abundance depending on bacterial genus. | ||
| Herbicide Organophosphate (Glyphosate) |
Sprague-Dawley rats | Gut microbiota dysbiosis and sex-dependent effects at all doses examined. | |||
| Herbicide phenoxyacetic acid (2,4 D) | Male C57BL/6 mice | Increase and decrease in relative microbial abundance based on bacterial genus. | |||
|
Djekkoun, N. et al. 2021. |
Organophosphates (Chlorpyrifos) | Rats | Higher number of harmful bacteria. Lower number of beneficial bacteria. Increase in potentially pathogenic flora. Decrease in beneficial flora | No impact or increase in body weight in adults. Low body mass and short body length at birth. Changes in plasma glucose levels and lipid profile. Significant difference in body weight. | This study has shown that exposure to pesticides modulates bacterial populations, impacting the health of the host. Intestinal dysbiosis induced by these compounds is associated with alterations similar to those observed in metabolic syndrome, where bacterial translocation, increased intestinal permeability and microbial dysmetabolism generate low-grade inflammation. These mechanisms contribute to imbalances in energy homeostasis and increase the risk of developing chronic inflammatory diseases. It has been demonstrated that various pesticides can induce endotoxemia, mucosal permeability, and proinflammatory activation, leading to systemic inflammatory responses. Alterations in microbial substrates in the intestine have also been observed, leading to changes in short-chain fatty acid profiles and altering energy collection through targeted dysbiosis, reinforcing their role in metabolic disruption and the development of chronic inflammatory pathologies. Pesticides alter the gut microbiome, influence energy metabolism and promote low-grade inflammation. They also impact on metabolic health and the development of chronic diseases. |
| Mice | Increase in potentially pathogenic flora. Decrease in beneficial flora. | Abnormal permeability. | |||
| Organophosphates (Diazinon) | Mice | Altered microbiome composition. |
Impaired energy metabolism. Male animals are more susceptible to abnormal translocation. Reduced body weight gain. |
||
| C57BL/6 mice | Increase in potentially pathogenic flora. Decrease in beneficial flora. | Significant increase in body, liver and epididymal fat. Increase in serum TG and glucose levels. | |||
| Organophosphates (MCP) | Mice | Increase in potentially pathogenic flora. |
Increased blood sugar levels. Glucose intolerance. | ||
| Organochlorines (TCDF) | Rats | Decrease in beneficial flora. | It causes inflammation and alters hepatic lipogenesis, gluconeogenesis, and glycogenolysis. | ||
| Organochlorines (DDT) | Rats | Increase in potentially pathogenic flora. Decrease in beneficial flora | Increased weight gain. Increased fasting glucose and insulin. Disturbed lipid metabolism. | ||
| Organochlorines (PCP) | Female mice | Increase in potentially pathogenic flora. Decrease in beneficial flora. | Reduction in body weight. | ||
| Benzimidazoles (CBZ) | Mice | Increase in potentially pathogenic flora. Decrease in beneficial flora. | Accumulation of liver lipids. Increase in TG, cholesterol, HDL, and LDL. |
||
|
Utembe, W. & Kamng’ona, A. W. 2021 |
Data collected in other reviews. | Case-control studies in humans. Cohort studies in humans. Rats. Mice. |
Glyphosate. Chlorpyrifos. Pyrethroids. Imidacloprid. Diazinon. Ammonium glufosinate. |
This study confirms the existence of cognitive deficits, motor dysfunction, stress-related hypermobility, social interaction dysfunction, reduced responsiveness to social novelty in adults, short-term memory impairment, changes in maternal behavior, and neural plasticity along with repetitive stereotypical behavior. Behaviors similar to those of autism spectrum disorder are observed. In addition, morphological changes in glial cells residing in the brain and reduced blood-brain barrier permeability are confirmed. There are also changes in the levels of short-chain fatty acids in the brain, along with negative regulation of neurotransmitters. Alteration of acetylcholinesterase activity, implying its inhibition. Hypo- or hypersensitization of cholinergic and GABAergic systems. Loss of dopaminergic neurons in the substantia nigra and neuronal inflammation. There is also evidence of increased transcription of the GABA-A-A2 subunit and M2 receptor genes, along with stimulation of pituitary hormone release (TNF-α, etc.). There is also increased oxidative stress, which promotes ageing and chronic diseases. | |
|
Meng, Z. et al. 2020 |
Organophosphate insecticide (Malathion) | Mice | Disorders of the composition of the gut microbiota. | Altered genes involved in quorum sensing, increased motility, pathogenicity, and genes related to cell wall components. | This work leads us to understand that exposure to pesticides can modify the composition of the gut microbiome, which, in turn, alters the production and function of its key metabolites. Metabolic profile disorders, insulin resistance, and obesity have been identified. Exposure to these substances can negatively affect health, especially the digestive system and metabolism. It has been proven that an initial impact on the delayed maturation of the digestive tract influences nutrient absorption and promotes weight gain and lipid accumulation. This imbalance in lipid metabolism generates inflammation, which interferes with bile acid metabolism and can cause enterohepatic disorders, increasing the risk of cardiovascular disease. In addition, chronic colon inflammation and liver toxicity aggravate overall health, reflecting metabolic disorders and alterations in digestive function. These effects underscore the importance of understanding the interactions between the gut microbiome and metabolism and environmental factors in the development of chronic diseases. |
| Organophosphate insecticide (Diazinon) | Mice | Alterations in the gut microbiota. | Metabolic profile disorders. | ||
| Organophosphate insecticide (Chlorpyrifos) | Mice | Intestinal inflammation and abnormal permeability. | Insulin resistance and obesity. | ||
| Rats |
Delayed maturation of the digestive tract. | ||||
| Organophosphate insecticide (Dichlorodiphenyltrichloroethane) | Rats |
Weight gain and lipid accumulation. | |||
| Pyrethroid insecticide (Permethrin) | Baby rats | Dyskinesia and intestinal disease. | |||
| Benzimidazole fungicide (Carbendazim) | Mice | Lipid metabolism disorder and inflammation. | |||
| Lipid metabolism disorder and inflammation. | |||||
| Systemic fungicide (Propamocarb) |
Mice | Disorders of lipid and bile acid metabolism. | |||
| Disorders of enterohepatic metabolism and possible cardiovascular disease. | |||||
| Systemic fungicide that inhibits ergosterol (Imazalil) |
Mice | Colon inflammation. | |||
| Systemic fungicide containing triazole compounds (Epoxiconazole) |
Rats | Hepatic toxicity. | |||
| Systemic fungicide composed of Triazole (Penconazole and its enantiomers) | Mice | Intestinal microbiota disorders. | Metabolic profile disorders. | ||
|
Yuan, X. et al. 2019 |
Data collected in other reviews. | Human body fluids. Mice. NOD mice. Adult male C57BL/6 mice. Sprague-Dawley rats. Randomized study. Pregnant women. Prospective randomized study, mother-baby pairs. Double-blind, placebo-controlled, randomized study in patients with type 2 diabetes. Case-control study in humans. |
. | This work leads us to conclude that pesticides can alter the composition of the gut microbiome and its metabolites. It also suggests that changes in the gut microbiome and metabolites can cause adverse effects in the host, in the transduction of extracellular signals from the plasma membrane to the cell and along the intracellular chain to stimulate the cellular response. Different bile acids can bind to different receptors, which can lead to atherosclerosis, hepatic lipid metabolism disorders, and dysbiosis due to fat accumulation. Short-chain fatty acids derived from microbial fermentation of fibre can inhibit histone deacetylases and serve as energy substrates by directly activating G protein-coupled receptors. The action of short-chain fatty acids on the GPR receptor in fat cells can cause dysbiosis due to fat accumulation. In addition, innate immune cells activated by endotoxic bacteria can release various pro-inflammatory cytokines, inducing low-grade inflammation and even neuronal inflammation. The alteration of the intestinal microbiome balance and increased intestinal permeability due to pesticide absorption are potential risk factors for increasing the entry of molecules with higher molecular weight, leading to an inflammatory response and, ultimately, low-grade inflammation. Furthermore, the action of short-chain fatty acids in the brain can affect appetite and is therefore associated with obesity and diabetes. In short, pesticides can act on intestinal microbes, affect their metabolites, and destroy the mucosa and intestinal cells. These changes cause pathological changes by acting on receptor sites in different tissues and organs. |
Figure 1.
Flowchart for study selection.
3.3. Results of the Effects of Pesticides Exposure in Animal Models
3.3.1. Effects of Exposure in Rats
Experimental studies conducted on rats have revealed a wide variety of physiological alterations resulting from exposure to these products. In the nervous system, there has been an increase in the activity of excitatory pathways, accompanied by a decrease in inhibitory activity, as well as the loss of dopaminergic neurons in the substantia nigra. At the endocrine and metabolic level, an increase in basal insulin levels has been documented, along with liver dysfunction affecting essential processes such as lipogenesis, gluconeogenesis and glycogenolysis, leading to manifestations of liver toxicity. Likewise, increases in body weight, obesity, a higher prevalence of phenotypes compatible with type 2 diabetes mellitus, and alterations in the pancreatic islets, characterized by lipid accumulation, have been observed.
From an immunological perspective, exposure to these compounds is associated with a decrease in immune response and alterations in the mechanisms regulating inflammation. At the intestinal level, both structural and functional changes have been described, including micro- and macro-anatomical alterations located in the right intestine, a reduction in beneficial bacterial genera, the proliferation of potentially pathogenic species, and the establishment of a state of dysbiosis in the intestinal microbiota. In addition, the results show differences in the response to exposure based on sex, present at all doses evaluated.
Taken together, these findings suggest that exposure to pesticides could compromise the homeostasis of the nervous, endocrine, immune, and gastrointestinal systems, underscoring the importance of further evaluating their effects on human health and extending the findings to humans.
3.3.2. Effects of Gestational Exposure to Pesticides on Rat Offspring
Research conducted on rat pups whose mothers were exposed to pesticides during pregnancy has revealed multiple metabolic, neurological and developmental abnormalities. Metabolically, hyperlipidemia and hyperglycemia have been observed in female offspring, while neurologically, persistent dysfunction has been observed throughout their lives. In male fetuses, changes in gene transcription in the external genitalia have been reported, along with an increased susceptibility to prostate disease, obesity and kidney disease.
The development of the digestive system is also compromised, with delayed maturation of the gastrointestinal tract, dyskinesia and the onset of various intestinal diseases. Regarding the reproductive system, ovarian alterations and abnormalities during the birthing process have been documented. Likewise, at the neuroinflammatory and cellular level, prenatal exposure is associated with changes in the inflammatory response and in the expression of genes involved in oxidative stress, particularly in brain regions such as the cortex and cerebellum.
Similarly, behavioral alterations have been identified in offspring, especially in their ability to respond to novel situations, manifesting exacerbated phenotypes similar to those observed in autism spectrum disorder. Taken together, these results consolidate the evidence that prenatal exposure to pesticides can have persistent adverse effects on the development and physiological functionality of offspring.
3.3.3. Effects of Exposure on Mice
Research conducted on mice has shown that exposure to pesticides induces significant alterations in the composition and functionality of the gut microbiome, characterized by states of dysbiosis and increased intestinal barrier permeability. In the metabolic sphere, there has been evidence of impaired energy metabolism, accompanied by a decrease in weight gain in both sexes, reduced body weight in females, and abnormal bacterial translocation phenomena. Likewise, increases in blood glucose, glucose intolerance, hepatic lipid accumulation and alterations in the lipid profile have been observed, including elevations in triglyceride, total cholesterol, HDL and LDL levels.
From a genetic and microbiological perspective, exposure to these compounds affects quorum sensing regulation, promoting an increase in bacterial motility and pathogenicity, as well as modifications in the expression of genes linked to cell wall structure and pesticide biotransformation. Similarly, alterations in enterohepatic metabolism, variations in bile acids, insulin resistance and an intensification of systemic inflammatory processes have been documented, all of which are associated with an increased risk of developing cardiovascular disease and colitis.
At the level of the central nervous system, deregulation in the synthesis and release of neurotransmitters has been observed, along with neuronal inflammation, deterioration of locomotor activity, alterations in short-term memory, and the manifestation of behaviors like those observed in autism spectrum disorder.
In terms of body composition, males show greater susceptibility to abnormal translocation, as well as a significant increase in the accumulation of body, hepatic and epididymal fat. Increases in serum triglyceride and glucose levels have also been reported, as well as an accentuation of glucose intolerance. At the same time, changes in gene expression associated with metabolic pathways and glucose processing have been identified.
Taken together, these findings indicate that pesticides exposure has a multisystemic impact, compromising energy metabolism, immune homeostasis and neurological function, with effects that differ according to sex and duration of exposure.
4. Discussion
The main objective of this study was to analyze the existing scientific evidence on the effects that pesticides can have on humans. To this end, a review was conducted of empirical research carried out on human populations and experimental studies on animal models that share genetic, biological and behavioral characteristics with humans. This methodological approach allows us to anticipate the possible effects of pesticides on humans, considering environmental conditions comparable to those used in experimental studies with animals.
Based on the analysis of the scientific literature reviewed in this study, it can be concluded that exposure to pesticides in humans, particularly during pregnancy, is associated with adverse effects on fetal development, metabolism and immune system function. These effects include significant alterations in the neurological and metabolic systems, as well as in the body's immune defense mechanisms.
Likewise, the decrease in birth weight, considered a risk marker for neonatal development and health, is particularly relevant. This finding is of great importance, given that low birth weight has been linked to increased susceptibility to chronic cardiovascular, metabolic, and neurological diseases throughout the life cycle, a phenomenon that has also been documented in animal model studies.
The evidence suggests that the relationship between exposure to pesticides and reduced birth weight could be explained by the interference of these chemical compounds in the normal biological mechanisms of pregnancy, affecting maternal-fetal nutrition and the processes of fetal growth and maturation.
On the other hand, it is worth highlighting the association between pesticides exposure and an increased likelihood of developing autism spectrum disorders, especially when such exposure occurs during the second and third trimesters of pregnancy. The fact that the risk increases by approximately 30% during these periods suggests that the observed effects of prenatal exposure are not a mere coincidence but rather respond to biological mechanisms consistent with the findings obtained in experimental research with animal models.
Furthermore, the identification of autism spectrum traits in 11-year-old children exposed to pesticides in utero shows that the consequences of such exposure transcend the short term. These effects, which become evident in later stages of child development, have significant implications for long-term neuropsychological and behavioral functioning.
Evidence from animal model studies reinforces this relationship, providing experimental support that increases the severity of the phenomenon. Thus, not only is the immediate well-being of the fetus compromised during gestation, but the individual's cognitive, emotional and adaptive functions are also substantially affected during development and in adulthood.
Evidence linking pesticides exposure to insulin resistance, type 2 diabetes mellitus, obesity, and dyslipidemia reveal a characteristic pattern of metabolic disorders affecting a significant proportion of the global population, particularly in Western societies. Insulin resistance is one of the central pathophysiological mechanisms in the development of type 2 diabetes and has been associated with low-grade chronic inflammation and increased oxidative stress, both of which have been identified as consequences of the metabolic disorders described in experimental studies with rats and mice. These findings suggest that metabolic disorders do not result exclusively from excess calories, but also from a persistent inflammatory context that interferes with insulin action and promotes the accumulation of adipose tissue in various organs.
In relation to liver alterations and lipid accumulation in the liver, the results indicate the possible presence of non-alcoholic fatty liver disease, a condition that is increasing globally and is closely related to obesity and metabolic syndrome. The accumulation of liver fat, in addition to being a marker of metabolic dysfunction, can predispose individuals to the development of more serious lesions, such as fibrosis and liver cirrhosis.
The effects of this dysfunction on systemic metabolism should be analyzed in the context of dyslipidemia, given that the liver plays an essential role in regulating lipid metabolism and releasing fats into the bloodstream. Taken together, these findings reinforce the need to consider pesticides exposure as a possible contributing factor in etiology of chronic metabolic disorders.
Atherosclerosis, characterized by the accumulation of cholesterol plaques in the arterial walls, is closely linked to the metabolic disorders described above and is one of the leading causes of cardiovascular morbidity and mortality worldwide. The interrelationship between dyslipidemia, insulin resistance and atherosclerosis shows how alterations in lipid and glucose metabolism can jointly contribute to the development of cardiovascular diseases, which remain the leading cause of death in many parts of the world. Studies conducted in animal models have corroborated this association, reinforcing the evidence that exposure to pesticides may play a significant role in the genesis of these metabolic and vascular alterations.
Furthermore, the activation of the inflammatory response and the release of pro-inflammatory cytokines are key to understanding the systemic impact of exposure to these compounds. Low-grade chronic inflammation is recognized as a common feature in many metabolic diseases and plays a decisive role in the progression of insulin resistance and obesity. Cytokines such as interleukin-6 (IL-6) and tumor necrosis factor alpha (TNF-α) have been widely identified for their ability to interfere with insulin signaling and promote tissue damage.
This persistent inflammatory response not only alters metabolism but also exerts adverse effects on various organs and tissues, promoting endothelial dysfunction and aggravating cardiovascular pathologies. Taken together, these processes reflect a complex interaction between exposure to chemical agents, systemic inflammation, and metabolic alteration, which contributes significantly to the deterioration of cardiovascular health.
Intestinal dysbiosis, defined as an imbalance in the composition and function of the gut microbiome, has been shown to be associated with various metabolic disorders, including type 2 diabetes mellitus, obesity, and liver disease, findings that have also been corroborated in animal models. Dysbiosis promotes increased intestinal permeability, facilitating the translocation of bacterial endotoxins into the systemic circulation. These endotoxins activate the immune system, amplifying the inflammatory response, and contribute to both insulin resistance and adipose tissue accumulation.
In addition, dysbiosis can cause structural damage to the intestine, compromising nutrient absorption and the protective barrier against pathogens. This microbial imbalance has been linked to a wide range of disorders, including digestive and metabolic diseases, infections, and autoimmune pathologies, reinforcing the hypothesis that exposure to pesticides may have systemic effects that transcend intestinal health.
Increased intestinal permeability is being increasingly investigated for its involvement in metabolic and autoimmune diseases. Evidence suggests that the gut microbiome plays a central role in regulating metabolism and systemic inflammation. Its alteration can perpetuate a cycle of chronic inflammation that interferes with normal metabolic functions, increasing susceptibility to disorders such as obesity, diabetes, and various autoimmune diseases.
Liver damage, together with an increased risk of inflammatory diseases such as colitis and cardiovascular pathologies, highlights the negative effect of pesticides on overall health, with a particular impact on essential organs such as the liver and cardiovascular system.
In this context, studies conducted on animal models are essential for understanding the biological and physiological effects of pesticides exposure, as they provide relevant information on the underlying mechanisms that could have implications for human health. The evaluation of effects in rats and mice has revealed a series of physiological alterations affecting multiple biological systems, including the nervous, immune and gastrointestinal systems. These studies not only identify the direct impacts of pesticides but also anticipate the possible long-term consequences on the health and well-being of organisms, including humans.
The findings summarized in Table 1 corroborate the results obtained in empirical research with human populations and highlight the adverse effects of pesticides on adult rats and mice, as well as on their offspring when exposure occurs during gestation.
This study has several notable strengths. Notably, it integrates evidence from human studies and animal models, enabling a more comprehensive and robust analysis of the effects of pesticides. The study also takes a multidisciplinary approach, addressing the metabolic, immune, neurological and gastrointestinal systems to provide a comprehensive view of the alterations induced by these compounds. The study's clinical and public health relevance is significant, particularly with regard to prenatal exposure and its potential long-term consequences. Furthermore, comparing experimental and epidemiological findings reinforces the biological plausibility of the observed effects.
However, the study also has some limitations. Firstly, the available evidence in humans is limited, which makes it difficult to establish definitive causal relationships. Secondly, there is methodological heterogeneity among the reviewed studies. Furthermore, long-term follow-up is lacking in many studies, which limits our understanding of chronic effects. All of these limitations must be considered when interpreting the results.
5. Conclusions
In summary, the results observed in human studies and supported by trials conducted in animal models show that exposure to pesticides is associated with interrelated metabolic, systemic, and immunological effects that compromise overall health. Insulin resistance, dyslipidemia, low-grade chronic inflammation, and intestinal dysbiosis are interconnected risk factors that require a multidisciplinary approach for their prevention and management. The analysis of these pathological processes not only provides valuable information on metabolic diseases but also opens research opportunities aimed at intervention on the intestinal microbiome and modulation of the inflammatory response as key therapeutic strategies.
Furthermore, pesticides have been documented to produce adverse effects on the immune system, the nervous system, and the gut microbiome, contributing to the development of inflammatory, metabolic, and degenerative diseases of the central nervous system. Exposure during the prenatal period can predispose individuals to chronic diseases in adulthood, as well as to neurodevelopmental disorders, early intestinal dysbiosis, obesity, and other metabolic and endocrine disorders.
The results highlight the need for public health strategies that reduce pesticide exposure in vulnerable populations and strengthen environmental and maternal-child surveillance. Interventions aimed at modulating the gut microbiome and systemic inflammation, together with preventive and translational research approaches, could mitigate metabolic, immunological and neurodevelopmental effects, thus protecting human health from the risks associated with these compounds.
Interventions are also being considered to reduce exposure to pesticides, especially among vulnerable populations such as pregnant women and children. This includes promoting sustainable agricultural practices, regulating the use of chemical compounds, and educational programs on safe handling. It is also essential to strengthen environmental surveillance and biomonitoring, as well as monitoring maternal and child development, with the aim of early detection and mitigation of adverse effects on metabolism, the immune system and neurodevelopment.
Taken together, these actions highlight the importance of a multidisciplinary approach integrating prevention, monitoring and translational research to protect human health from the risks associated with pesticides exposure.
Author Contributions
Conception and study design, M.J.D.-D. Methodology, M.J.D.-D. and I.O.-S. Data collection, M.J.D.-D and I.O.-S. Data analysis, M.J.D.-D and I.O.-S. Writing—original draft preparation, M.J.D.-D and I.O.-S. Writing—review and editing, M.J.D.-D, I.O.-S. and T.R.-R. Approval of final manuscript, M.J.D.-D, I.O.-S. and T.R.-R. All authors have read and agreed to the published version of the manuscript.
Funding
This work received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kaur, R.; Choudhary, D.; Bali, S.; Singh Bandral, S.; Singh, V.; Ahmad, Md. A.; Rani, N.; Gurjeet Singh, T.; Chandrasekaran, B. Pesticides: An alarming detrimental to health and environment. Sci Total Environ 2024, 915:170113. [CrossRef]
- Kim, K.H.; Kabir, E.; Jahan, S.A. Exposure to pesticides and the associated human health effects. Sci Total Environ 2017, 575:525-535. [CrossRef]
- Damalas, C.A.; Eleftherohorinos, I.G. Pesticide exposure, safety issues, and risk assessment indicators. Int J Environ Res Public Health 2011, 8(5):1402-1419. [CrossRef]
- Mostafalou, S.; Abdollahi, M. Pesticides: an update of human exposure and toxicity. Arch Toxicol 2017, 91(2):549-599. [CrossRef]
- Vehovszky, Á; Farkas, A.; Ács, A.; Stoliar, O.; Székács, U.; Mörtl, M.; Gyóri, J. Neonicotinoid insecticides inhibit cholinergic neurotransmission in a molluscan (Lymnaea stagnalis) nervous system. Aquat Toxico 2015, 167:172-179. [CrossRef]
- Czarnywojtek, A.; Jaz, K.; Ochmańska, A.; Zgorzalewicz-Stachowiak, M.; Czarnocka, B.; Sawicka-Gutaj, N.; Ziółkowska, P.; Krela-Kaźmierczak, Y.; Intestino, P.; Florek, E.; Ruchala, F. The effect of endocrine disruptors on the reproductive system - current knowledge. Eur Rev Med Pharmacol Sci 2021, 25(15):4930-4940. [CrossRef]
- Wan, M.L.Y.; Co, V.A.; El-Nezami, H. Endocrine disrupting chemicals and breast cancer: a systematic review of epidemiological studies. Crit Rev Food Sci Nutr 2022, 62(24):6549-6576. [CrossRef]
- Damalas, C.A.; Koutroubas, S.D. Farmers' Exposure to Pesticides: Toxicity Types and Ways of Prevention. Toxics 2016, 4(1):1. [CrossRef]
- Fuller, R.; Landrigan, P.J.; Balakrishnan, K.; Bathan, G.; Bose-O'Reilly, S.; Brauer, S.; Caravanos, J.; Chiles, T.; Cohen, A.; Corra, L.; et al. Pollution and health: a progress update. Lancet Planet Health 2022, 6(6):e535-e547. [CrossRef]
- Rajilić-Stojanović, M.; de Vos, W.M. The first 1000 cultured species of the human gastrointestinal microbiota. FEMS Microbiol Rev 2014, 38(5):996-1047. [CrossRef]
- Sommer F.; Bäckhed, F. The gut microbiota--masters of host development and physiology. Nat Rev Microbiol 2013, 11(4):227-238. [CrossRef]
- Álvarez,J.; Fernández Real, J.M.; Guarner, F.; Gueimonde, M.; Rodríguez, J.M.; Saenz de Pipaon, M.; Sanz, Y. Gut microbes and health. Gastroenterol Hepatol 2021, 44, 7, 519-535. doi.org/10.1016/j.gastrohep.2021.01.009.
- Wang, J.; Hou, Y.; Mu, L.; Yang, M.; Ai, X. Gut microbiota contributes to the intestinal and extraintestinal immune homeostasis by balancing Th17/Treg cells. Int Immunopharmacol 2024, 143(Pt 3):113570. [CrossRef]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem J 2017, 474(11):1823-1836. [CrossRef]
- Marchesi, J.R.; Adams, D.H.; Fava, F.; Hermes, G.D.A.; Hirschfield, G.M.; Hold, G.; Quraishi, M.N.; Kinross, J.; Smidt, H.; Tuohy, K.M.; Thomas, L.V.; Zoetendal, E.G.; Hart, H. The gut microbiota and host health: a new clinical frontier. Gut 2016, 65(2):330-339. [CrossRef]
- Cani, P.D.; Jordan, B.F. Gut microbiota-mediated inflammation in obesity: a link with gastrointestinal cancer. Nat Rev Gastroenterol Hepatol 2018, 15(11):671-682. [CrossRef]
- Quigley, E.M.M. Microbiota-Brain-Gut Axis and Neurodegenerative Diseases. Curr Neurol Neurosci Rep 2017, 17(12):94. [CrossRef]
- Gomaa, E.Z. Human gut microbiota/microbiome in health and diseases: a review. Antonie Van Leeuwenhoek 2020, 113(12):2019-2040. [CrossRef]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat Rev Microbiol 2021, 19(1):55-71. [CrossRef]
- Wang, Z.; Zhao, Y. Gut microbiota derived metabolites in cardiovascular health and disease. Protein Cell 2018, 9(5):416-431. [CrossRef]
- Qiu, P.; Ishimoto, T.; Fu, L.; Zhang, J.; Zhang, Z.; Liu, Y. The Gut Microbiota in Inflammatory Bowel Disease. Front Cell Infect Microbiol 2022, 12:733992. [CrossRef]
- Nishida, A.; Inoue, R.; Inatomi, O.; Bamba, S.; Naito, Y.; Andoh, A. Gut microbiota in the pathogenesis of inflammatory bowel disease. Clin J Gastroenterol 2018, 11(1):1-10. [CrossRef]
- Goldbaum, A.A.; Bowers, L.W.; Cox, A.D.; Gillig, M.; Clapp Organski, A.; Cross, T.L. The Role of Diet and the Gut Microbiota in the Obesity-Colorectal Cancer Link. Nutr Cancer 2025, 77(6):626-639. [CrossRef]
- Wong, C.C.; Yu, J. Gut microbiota in colorectal cancer development and therapy. Nat Rev Clin Oncol 2023, 20(7):429-452. [CrossRef]
- Khan, M.N; Bilal, H.; Khan, S.; Shafiq, M.; Xiaoyang, J. Collateral damage of neonicotinoid insecticides: Unintended effects on gut microbiota of non-target organisms. Comp Biochem Physiol C Toxicol Pharmacol 2025, 298, 110330. doi.org/10.1016/j.cbpc.2025.110330.
- Rivi, V.; Pele, G.; Yakubets, K.; Batabyal, A.; Dominici, R.; Blom, J.M.C.; Tascedda, F.; Benatti, C.; Lukowiak, K. Effects of neonicotinoid and diamide-contaminated agricultural runoff on Lymnaea stagnalis: Insights into stress, neurotoxicity, and antioxidant response. Aquat Toxicol 2025, 287:107535. [CrossRef]
- Roman, P.; Cardona, D.; Sempere, L.; Carvajal, F. Neurotoxicology 2019, 75:200-208. [CrossRef]
- Xu, S.; Li, X.; Zhang, S.; Qi, C.; Zhang, Z.; Ma, R.; Xiang; L.; Chen, L.; Zhu, Y.; Tang, C.; et al. et al. Oxidative stress gene expression, DNA methylation, and gut microbiota interaction trigger Crohn's disease: a multi-omics Mendelian randomization study. BMC Med 2023 ,21(1):179. [CrossRef]
- Alula, K.M.; Dowdell, A.S.; LeBere, B.; Lee, J.S.; Levens, C.L.; Kuhn, K.A.; Kaipparettu, B.A.; Thompson, W.E.; Blunberg, R.S.; Colgan, S.P.; Theiss, A.L. Interplay of gut microbiota and host epithelial mitochondrial dysfunction is necessary for the development of spontaneous intestinal inflammation in mice. Microbiome 2023, 11(1):256. [CrossRef]
- Liang, Y.; Zhan, J.; Liu, D.; Luo, M.; Han, J.; Liu, X.; Liu, C.; Cheng, Z.; Zhou, Z.; Wang, P. Organophosphorus pesticide chlorpyrifos intake promotes obesity and insulin resistance through impacting gut and gut microbiota. Microbiome 2019, 7(1):19. [CrossRef]
- Abou Diwan, M.; Djekkoun, N.; Boucau, M.C.; Dehouck, L.; Biendo, M.; Gosselet, F.; Bach, V.; Cnadela, P.; Khorsi-Cauet, H. Maternal exposure to pesticides induces perturbations in the gut microbiota and blood-brain barrier of dams and the progeny, prevented by a prebiotic. Environ Sci Pollut Res Int 2024, 31(49):58957-58972. [CrossRef]
- Bennett, D.; Bellinger, D.C.; Birnbaum, L.S.; Bradman, A.; Chen, A.; Cory-Slechta, D.A.; Engel, S.M.; Fallin, S.D.; Halladay, A.; Hauser, R.; et al. Project TENDR: Targeting Environmental Neuro-Developmental Risks The TENDR Consensus Statement. Environ Health Perspect 2016, 124(7):A118-A122. [CrossRef]
- World Health Organization. Public health impact of pesticides used in agriculture. 2020. Available online: https://www.who.int.
- Galarza, C.R.; Cruz, P.G. Guía para realizar estudios de revisión sistemática cuantitativa. CienciAmérica 2024, 13(1), 1-6.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



