
Submitted:
08 December 2025
Posted:
09 December 2025
You are already at the latest version
Abstract
Dystocia represents a multifactorial and clinically significant reproductive disorder af-fecting a broad spectrum of reptilian species. Commonly resulting from prolonged vitellogenesis, endocrine disruption, or hepatic lipidosis, dystocia is often exacerbated by suboptimal husbandry or concurrent disease. This review critically evaluates the etiology, diagnostic criteria, and therapeutic interventions associated with this condition. Emphasis is placed on the interplay between metabolic exhaustion and hepatic compromise, which may lower the threshold for surgical intervention. The efficacy and limitations of oxytocin-based protocols are discussed in the context of hormonal receptor variability and response attenuation. Advanced diagnostic modalities, including ultrasonography, radiography/CT, and biochemical profiling, are reviewed for their utility in case stratification. Finally, surgical management options are considered for cases refractory to medical treatment, with attention to timing, anaesthetic risk, and postoperative care. Collectively, this synthesis aims to inform evidence-based clinical decision-making and promote improved standards of care in reptile reproductive medicine.
Keywords:
reptiles
; dystocia
; egg-binding
; reproductive disorder
; hepatic lipidosis
; treatment
; diagnostic imaging
; surgery
1. Introduction
Reproductive disorders are a significant clinical concern in captive reptiles, particularly as exotic species gain popularity in zoological collections, private ownership, and conservation programs. Dystocia is ranked as one of the most common and potentially life-threatening conditions found in this taxon [1]. Unlike domestic mammals, reptiles possess diverse reproductive strategies, influenced by phylogeny, environment, and ecology. These variations directly impact the presentation, progression, and management of reproductive diseases, such as dystocia. Despite its frequent occurrence, the pathophysiology of dystocia remains incompletely understood. Moreover, clinical management often varies significantly depending on the species, reproductive stage, and clinical and metabolic status of the animal. To address this disparity, a search was conducted for peer-reviewed articles in academic literature databases, such as PubMed, Scopus, and Web of Science, by using words and word combinations, such as dystocia, reptile, reproduction, reproductive disease, follicular stasis, hepatic lipidosis, diagnosis, surgery, and oxytocin, to identify potentially relevant publications. We then evaluated reports that we considered as clinically relevant and articles that were deemed irrelevant to the review’s objectives were discarded. The scope of this review is to provide an integrated perspective on reptilian dystocia by examining the interplay between hormonal resistance, hepatic metabolic function, diagnostic strategies, and clinical management decisions, a multidimensional framework not previously synthesized in the published literature.
1.1. Reproductive Physiology Across Reptilian Taxa
Reptiles reproduce primarily through oviparity, although both viviparity and oviparous have evolved independently multiple times among squamates. Ovoviviparity is common in boas (Boidae), whereas true viviparity, with placental nutrient transfer, has been documented in several skinks (Scincidae) and in certain geckos such as Naultinus spp. (Gekkonidae) [2,3]. This reproductive diversity results in fundamental reproductive anatomical and physiological differences, which in turn influence the incidence and clinical presentation of reproductive disorders. Oviparous reptiles produce eggs that are retained within paired oviducts until deposition and may be either soft-shelled or hard-shelled, depending on the species’ environmental adaptations. In general, hard-shelled eggs are produced by species that nest in arid or semi-arid environments, while soft-shelled eggs are laid by those in moist habitats. These structural differences can affect the risk of specific complications, such as egg fracture or retention. In contrast, viviparous and oviparous species develop internal embryos, with the former exhibiting varying degrees of placentation and maternal nutrient transfer, and the latter relying primarily on yolk nutrients for embryonic development [4].
Reproductive endocrinology in reptiles is regulated by a tightly coordinated interaction of hypothalamic-pituitary-gonadal (HPG) axis hormones. The onset of vitellogenesis is driven by circulating oestrogen, primarily estradiol-17β, which stimulates the hepatic production of yolk precursors and follicular development [5,6]. As follicles mature, progesterone concentrations increase. This rise initiates oviductal preparation for ovulation and shell formation. After ovulation, oviductal motility is primarily controlled by neurohypophyseal hormones, especially arginine vasotocin (AVT). Prostaglandins, including prostaglandin F2α (PGF2α), provide additional regulation. These mediators trigger calcium-dependent rhythmic contractions of the oviductal smooth muscle, enabling effective egg transport and oviposition [7].
Disruption in this hormonal cascade at any point, as well as lack of calcium, whether from environmental, nutritional, or pathological causes, can lead to follicular arrest, ovulatory failure, or oviductal dystocia. Hormonal resistance or dysfunction, particularly involving AVT receptors, underlie most cases of dystocia, especially in iguanas, chelonians, and skinks [8,9].
Anatomical and physiological differences among reptile taxa influence both the clinical presentation and diagnostic challenges of reproductive disorders. Chelonians possess a rigid pelvic canal that can obstruct oviposition, particularly in cases of pelvic deformities or oversized eggs [10]. Lizards, such as iguanas and agamids (Agamidae), have paired ovaries and oviducts that may ovulate asynchronously, sometimes resulting in the accumulation of yolk-filled follicles. While this has traditionally been referred to as pre-ovulatory follicular stasis (POFS), recent work suggests this condition may be overdiagnosed or misunderstood, with some cases representing normal physiological variation rather than true pathology [11]. Snakes have elongated coelomic cavities and linear reproductive tracts, which can obscure clinical detection of retained eggs until late in the disease process [12]. Crocodilians, though less commonly treated in clinical settings, display seasonal reproductive cycles and complex maternal behaviours, which may predispose them to oviposition failure under suboptimal captive conditions [13,14].
1.2. Dystocia and Related Reproductive Disorders
Dystocia in reptiles, also referred as egg-binding or ovostasis, is defined as a failure to successfully oviposit or give birth to viable offspring within a biologically appropriate timeframe. This may manifest either as POFS, where follicles fail to ovulate and persist within the coelomic cavity, or as post-ovulatory dystocia (POD), involving the retention of shelled eggs or fetuses within the oviduct. This condition can have functional, mechanical, environmental, or metabolic causes [15,16].
Reproductive disorders in reptiles are broadly categorized into pre-ovulatory and post-ovulatory pathologies, each with distinct pathophysiological mechanisms and clinical implications. POFS is characterized by the failure of mature ovarian follicles to ovulate, often resulting from chronic reproductive stimulation in the absence of appropriate environmental or social cues, or due to endocrine dysfunction. These retained follicles may persist, undergo fibrosis, degenerate into tumors, or become secondarily infected, potentially leading to systemic illness or yolk coelomitis if ruptured [15,17]. Follicular atresia, a normal physiological regression of non-ovulated follicles, must be distinguished from POFS, particularly in species like chameleons where ultrasonographic appearances may resemble pathological stasis. Misinterpretation of imaging may lead to unnecessary surgical or medical interventions, underscoring the need for context-specific clinical evaluation and a thorough understanding of species-specific reproductive physiology [18,19]. Discerning between these conditions often requires integrating imaging modalities (e.g., ultrasonography, radiography and computed tomography (CT)), hormone assays, and consideration of environmental and husbandry factors.
In contrast, POD involves the retention of ovulated and often fully shelled eggs or developed fetuses within the oviducts. Causes include physical obstruction, oviductal inertia, or inadequate environmental parameters necessary for nesting behavior, such as temperature, humidity, or substrate [20].
2. Integrative Scope: Hormonal Resistance, Hepatic Function, and Diagnostic Framework
Reproductive tract motility in reptiles is primarily regulated by AVT and prostaglandins, particularly PGF2α, with oxytocin often used empirically despite variable species sensitivity and poorly characterized receptor pharmacodynamics [6]. In cases of dystocia, particularly in iguanas, bearded dragons (Pogona vitticeps), and chelonians, prolonged retention of follicles or eggs may result in reduced responsiveness to AVT or oxytocin. This resistance is thought to arise from receptor downregulation, chronic smooth muscle distension, and/or local inflammation. This hormone-resistance renders medical therapy ineffective beyond a narrow therapeutic window [8,10].
Simultaneously, the liver plays a central role in reproductive physiology by synthesizing vitellogenin under oestrogenic stimulation, but chronic vitellogenesis or reproductive stasis can induce hepatic lipidosis, hypoalbuminemia, and elevated liver enzymes, which are frequently reported in severe or chronic cases of POFS [15,21]. However, recent clinical reviews emphasize that these biochemical changes are often absent or only mild in early or functional cases; thus, normal blood chemistry does not reliably exclude POFS, and should not be used as a primary diagnostic criterion [11]. This is believed to result from a negative energy balance wherein adipose stores are mobilized to meet sustained metabolic demands, leading to hepatic accumulation of triglycerides [22]. Hypoalbuminemia may also develop in affected individuals, potentially due to decreased hepatic protein synthesis or the preferential diversion of amino acids toward vitellogenin production rather than albumin [23]. In contrast, hepatic changes appear to be significantly less prominent or relevant in POD. While POFS is often associated with prolonged oestrogenic stimulation and sustained vitellogenesis, factors that predispose to hepatic lipidosis, hypoalbuminemia, and elevated liver enzymes, POD occurs after ovulation, when vitellogenic activity has ceased or markedly declined [24,25]. Consequently, the hepatic metabolic burden is presumed to be lower in POD. Available literature primarily describes POD in terms of mechanical obstruction, oviductal inertia, or egg-associated coelomitis, with little emphasis on hepatic involvement [1,26]. To date, no peer-reviewed studies have reported consistent alterations in liver enzymes or serum proteins specific to POD, suggesting that hepatic pathology, if present, is likely secondary to systemic inflammation or sepsis rather than a direct consequence of reproductive physiology. Further comparative studies would be beneficial to confirm this distinction. These alterations, along with elevated liver enzymes, are frequently observed in clinical cases of reproductive disorders, such as follicular stasis. Consequently, the presence of elevated liver enzymes or hepatic lipidosis may be a significant diagnostic indicator in POFS, but are rarely observed in other reproductive conditions such as POD [27,28]. This hepatic dysfunction not only compromises anaesthetic safety but may also impair reproductive hormone metabolism, further perpetuating follicular arrest. These hepatic alterations, including elevated liver enzymes, are most frequently observed in reptiles with POFS, due to prolonged vitellogenesis and estrogenic stimulation [15]. Histopathological and epidemiological studies in bearded dragons confirm that reproductively active females are significantly predisposed to hepatic lipid accumulation, with adult females showing substantially higher odds of moderate to severe hepatic lipidosis compared to males [29].
In hepatic lipidosis, fat-laden hepatocytes impair normal liver function, including the metabolism and clearance of estradiol-17β, normally processed through hydroxylation and conjugation pathways [30]. As a result, persistently elevated estradiol-17b levels drive continued hepatic vitellogenin synthesis and follicular development. In the absence of ovulation, this creates a self-reinforcing feedback loop: sustained oestrogenic stimulation exacerbates hepatic lipidosis, which in turn further impairs hormone clearance and perpetuates follicular arrest [29,30].
As previously discussed, reproductive disorders such as follicular stasis and dystocia often result from a complex interplay between hepatic dysfunction and hormonal dysregulation. Understanding these mechanisms is essential for guiding diagnostic and therapeutic decision-making.
Diagnostic imaging plays a pivotal role in differentiating pre- from post-ovulatory reproductive disease. Ultrasonography is particularly useful for evaluating follicular integrity, echogenicity, inner architecture and vascularization, especially in lizards and snakes, while radiography allows for the evaluation of egg calcification. CT is the more powerful imaging tool in chelonians, as it can display follicles, eggs and salpinx as well as the skeleton. Position, number, size, shape, density and inner architecture can be differentiated as well as causes of soft tissue distension, mechanical obstruction or other comorbidities [31,32]. Furthermore, the density of the liver may be indicative for hepatic lipidosis in all reptiles [33,34,35] (Figure 1).
Reptile reproductive disorders are frequently complicated by concurrent hepatic dysfunction, yet standardized protocols for assessing hormonal responsiveness or determining optimal surgical timing remain lacking. Clinicians must therefore rely on a combination of biochemical, imaging, and clinical indicators, each with its own limitations. Traditional liver biochemical markers, including aspartate aminotransferase (AST), bile acids, and albumin, provide useful context but often remain within normal limits even in animals with histologically confirmed hepatic lipidosis, limiting their diagnostic reliability [28,36]. Imaging has emerged as a more sensitive tool in this setting. Unenhanced CT allows quantitative assessment of hepatic attenuation, which correlates strongly with lipid infiltration and clearly outperforms ultrasonography and serum biochemistry in sensitivity. These findings underscore the need for an integrated diagnostic framework. Incorporating CT-based liver density evaluation alongside reproductive imaging and interpretation of clinical timing offers a more accurate basis for risk stratification and therapeutic planning. This chapter therefore focuses on bridging this diagnostic gap by outlining a multifaceted, evidence-based approach to evaluating reproductive disorders in reptiles with potential hepatic involvement.
3. Hormonal, Environmental, and Species-Specific Drivers
Dystocia in reptiles arises from a complex interplay of physiological, environmental, nutritional, and anatomical factors. Despite the frequent presentation of dystocia in captive reptile medicine, most proposed risk factors remain extrapolated from case experience rather than experimental confirmation. This section reframes the aetiology of dystocia under three key clinical dimensions: endocrine-metabolic dysfunction, environmental and husbandry mismatch, and species-specific anatomical and physiological predispositions.
3.1. Endocrine-Metabolic Dysfunction and Reproductive Arrest
Reproductive arrest in reptiles, encompassing both POFS and POD, is frequently the result of underlying endocrine or metabolic dysfunctions. These disruptions are often subclinical until reproductive failure occurs, and they play a particularly critical role in oviparous lizards, snakes, and chelonians maintained in captivity. While reproductive endocrinology in reptiles shares hormonal analogues with other vertebrates, the nuances of oestrogen-driven vitellogenesis, progesterone-dependent oviductal preparation, and AVT-mediated oviposition are finely tuned by environmental and physiological cues—disruptions of which can precipitate ovulatory failure or impaired egg passage [7,37].
Estradiol-17β is the primary estrogen responsible for inducing hepatic vitellogenin synthesis in reptiles, as demonstrated in Kemp’s ridley sea turtles (Lepidochelys kempii), where experimental administration of estradiol-17β resulted in a marked increase in plasma vitellogenin concentrations within one week [38]. Chronic elevation, often due to prolonged exposure to reproductive triggers in the absence of mating or oviposition cues, can cause excessive hepatic lipogenesis. This may manifest clinically as hepatic lipidosis, which can interfere with hepatic detoxification, albumin synthesis, and hormone clearance [17,39]. For example, in Fiji island banded iguanas (Brachylophus fasciatus), repeated reproductive cycling without oviposition, which commonly occurs in single-housed females, leads to coelomic accumulation of yolk-laden follicles and progressive hepatic degeneration. In the majority of cases, yolk leakage was linked to the presence of large vitellogenic follicles undergoing atresia, which subsequently induced yolk coelomitis, characterized by pronounced mesothelial proliferation [39]. A histopathological study in bearded dragons showed that reproductive females had a significantly higher prevalence of hepatic lipid infiltration, with advanced cases correlating with inactive ovaries and non-responsive dystocia [29]. This hepatic compromise often coexists with hypoalbuminemia and impaired coagulation, posing anaesthetic risks if surgical intervention is later required. Additionally, hepatic impairment alters oestrogen and progesterone metabolism, potentially contributing to a cycle of continued follicular development without ovulation, particularly in species with high hepatic lipid loads. For instance, in the North African spiny-tailed lizard (Uromastyx acanthinura), the oestrogen receptor beta (ERβ) is predominantly involved in this regulatory mechanism and is associated with the inhibition of vitellogenin synthesis during certain reproductive phases [40]. Although oestrogen, particularly estradiol-17β, generally promotes vitellogenesis via oestrogen receptor alpha (ERα), ERβ may play an inhibitory or modulatory role, potentially acting to fine-tune or suppress estrogenic activity under specific physiological conditions. Therefore, disruption in liver function could impair this oestrogen-mediated pathway, resulting in reduced vitellogenin production and compromised reproductive capacity. Progesterone often acts to counterbalance oestrogen’s effects on vitellogenesis. The presence of hepatic progesterone receptors (PRA and PRB) in Uromastyx acanthinura, and their association with inhibition of vitellogenin synthesis during specific reproductive phases, presents an intriguing avenue for potential therapeutic intervention [40]. This mechanism suggests that progesterone or selective progesterone receptor modulators could theoretically be explored to downregulate vitellogenesis in cases of chronic reproductive stimulation, such as POFS, where prolonged estrogenic stimulation and failure of ovulation result in persistent vitellogenesis and follicular retention [30,38]. However, this approach would require caution and species-specific validation, as the hormonal feedback loops governing reproduction are complex and poorly understood in many reptilian taxa. Further research into receptor distribution, ligand specificity, and endocrine timing across species is needed before clinical application could be considered.
Reproductive success in reptiles is calcium-dependent, particularly during egg calcification and oviductal contraction. Hypocalcemia may result from inadequate dietary intake, poor gastrointestinal absorption, or insufficient ultraviolet B (UVB) exposure leading to reduced vitamin D₃ (cholecalciferol) synthesis. Additionally, repeated reproductive cycling, such as in females housed with males year-round, can lead to chronically increased calcium demand and depletion of physiological calcium stores, further predisposing individuals to hypocalcemia [41]. Calcium deficiency contributes to uterine inertia, especially in chelonians and gravid lizards, where sustained oviductal contraction is essential for successful oviposition [42]. In red-eared sliders (Trachemys scripta elegans), egg retention has been directly associated with ionized calcium levels below physiological thresholds, necessitating injectable calcium and oxytocin to restore oviductal motility [6].
Finally, yolk coelomitis, a severe inflammatory sequela of reproductive dysfunction, may develop secondary to chronic follicular atresia and vitellogenic failure. This condition, while often classified under reproductive or infectious pathology, is closely tied to endocrine-metabolic dysfunction, particularly when associated with hepatic impairment, prolonged vitellogenesis, or immune suppression. In one reported case, a captive white-throated monitor lizard (Varanus albigularis) exhibited nonspecific clinical signs including lethargy, anorexia, and coelomic distension before succumbing to severe coelomitis. Post-mortem findings revealed florid mesothelial hyperplasia attributed to yolk leakage from atretic follicles, underscoring the systemic impact of prolonged endocrine and metabolic dysregulation [43].
While hepatic lipidosis and hypocalcemia are widely recognized contributors to reproductive arrest in reptiles, there remains a significant gap in understanding the underlying endocrine and metabolic mechanisms. Notably, a limited number of studies have examined the expression of AVT receptors in oviductal smooth muscle, a factor that could influence oviductal contractility and contribute to dystocia. Additionally, comprehensive histopathological analyses of the liver in reptiles with idiopathic POFS or recurrent reproductive failure are lacking, despite evidence suggesting a link between hepatic lipid accumulation and reproductive disorders. Furthermore, comparative studies on calcium metabolism under varying UVB exposure and dietary regimes across different reptile taxa are sparse, yet such factors are crucial for proper reproductive function. To address these gaps, future research should adopt a systems biology approach, integrating assessments of hormonal levels, receptor functionality, liver function, and immune responses. This holistic perspective will enhance our understanding of the thresholds at which physiological variations transition into pathological conditions, ultimately improving diagnostic and therapeutic strategies for reproductive disorders in reptiles.
3.2. Risk Factors
3.2.1. Exogenous Factors
Environmental parameters play a critical role in reproductive stasis in reptiles; inadequate temperature gradients and humidity levels can disrupt normal oviductal contractility and nesting behavior, leading to egg retention. For instance, in chelonians, poor temperature regulation and improper humidity have been recognized as key contributors to reproductive disorders, including dystocia and follicular stasis [44]. The absence of appropriate nesting substrates or sites can cause females to retain eggs, as the lack of suitable laying environments impedes the natural oviposition process [45]. Nutritional deficiencies, particularly hypocalcaemia resulting from inadequate dietary calcium or vitamin D3, are significant contributors to dystocia. Hypocalcaemia impairs muscle contractions necessary for egg laying, leading to uterine inertia. In lizards, hypocalcaemia has been associated with clinical signs such as muscle tremors and lethargy, which are commonly observed in cases of dystocia [45]. Additionally, metabolic bone disease, meaning nutritional secondary hyperparathyroidism, resulting from chronic calcium and vitamin D3 deficiencies, has been reported in up to 84.4% of captive lizards in certain studies, indicating a widespread issue that can predispose reptiles to reproductive complications [46].
3.2.2. Endogenous Factors
Endocrine dysfunctions also play a role; disruptions in the hypothalamic-pituitary-gonadal axis can lead to persistent follicular growth without ovulation, culminating in POFS [47]. While specific studies quantifying this in reptiles are limited, the association between hormonal imbalances and reproductive issues is well-documented in veterinary literature [48]. Any space-occupying process within the coelomic cavity, as well as infections such as salpingitis and oophoritis, neoplasia, or urolithiasis, directly obstruct egg passage, or may lead to inflammation and adhesions that obstruct egg passage, particularly in species like tortoises and iguanids [26]. Physiological factors, including age and reproductive history, influence the risk of dystocia. Primiparous females and older reptiles are more susceptible, potentially due to inexperience or reduced muscular efficiency [49]. In addition, the effects of nutritional secondary hyperparathyroidism may compound with age, particularly in animals housed in suboptimal environments, where cumulative depletion of calcium and other resources further impairs the ability to pass eggs effectively [50]. Obesity, often resulting from overfeeding in captivity, can lead to excessive fat deposits in the coelomic cavity, physically impeding egg passage and increasing the risk of dystocia [45]. Moreover, once such animals enter a negative energy balance, during anorexia, reproductive stasis, or illness, they mobilize these fat stores, predisposing them to hepatic lipidosis [29].
3.3. Species-Specific Tendencies
While general pathophysiological mechanisms underlying dystocia in reptiles are often shared, the clinical presentation, progression, and optimal management of these reproductive disorders vary significantly between taxonomic groups. This variation is rooted in species-specific anatomical structures, reproductive physiology, environmental needs, and behavioural traits. For this reason, identifying and understanding taxon-specific predispositions is crucial for accurate diagnosis, timely intervention, and effective treatment planning. The most common types of dystocia, relevant species, risk factors, and supporting literature are shown for chelonians in Table 1, for snakes in Table 2, and for lizards in Table 3. By presenting these distinctions in a structured format, this review emphasizes the importance of tailored clinical approaches based on species, rather than applying uniform protocols across all reptiles, a practice that may result in misdiagnosis or suboptimal outcomes.
4. Diagnostic Workup
Dystocia often presents with nonspecific clinical signs such as anorexia, lethargy, coelomic distension, and restlessness. In some cases, straining behaviour or cloacal prolapse may be observed. These signs can persist for weeks or even months, making early detection challenging [24].
4.1. Radiographic Examination
Radiography remains one of the most accessible and widely used diagnostic tools in the assessment of dystocia in reptiles. It allows for non-invasive visualization of mineralized reproductive material, such as shelled eggs, dystrophic calcifications, as well as gas-filled coelomic structures, that may be suggestive for necrosis or secondary infection.
Standard lateral and dorsoventral radiographic views are typically sufficient for most lizard and chelonian species. In snakes, coiling often complicates interpretation, so careful straightening and multiple views may be needed [59]. For chelonians, horizontal beam radiography is highly valuable, particularly when using cranio-caudal or oblique projections to assess the pelvic inlet and caudal oviduct [60]. In gravid reptiles, a normal radiograph shows uniformly round to ovoid, more or less mineralized (depending on the species) eggs arranged linearly or clustered in the coelomic cavity. The number of eggs can be counted and the size measured, if sufficiently mineralized, which assists in determining normal versus pathological retention [24]. In lizards and snakes with obstructive dystocia, eggs may be crowded cranial to a narrowed pelvic region or appear deformed due to prolonged retention and compression. For example, a report in a bearded dragon describes the presence of three retained eggs, which caused caudal oviductal dilation and cloacal distension [56].
Radiographic examination can reveal several abnormalities indicative of dystocia in reptiles (Figure 2). One notable finding is the presence of overlapping or collapsed eggs, which typically suggest mechanical obstruction or repeated failed oviposition attempts (Figure 3). These eggs may appear irregular or crumpled on radiographs and are frequently observed in calcium-deficient individuals, particularly in green iguanas (Iguana iguana) [56]. Another diagnostic feature includes the detection of mineralized follicular shadows lacking definitive shell structure. While these may be misinterpreted as early-stage eggs, they more likely represent dystrophic calcifications associated with chronic follicular stasis or coelomitis [26]. In oviparous snakes such as ball pythons (Python regius), egg retention beyond the expected gestation period, typically 45 to 60 days post-ovulation, combined with radiographic evidence of persistent, calcified eggs, has been considered diagnostic for dystocia [61]. The presence of free intra-coelomic gas or gas within an egg on radiographs is another critical finding, the former being often indicative of oviductal rupture, secondary infection, or necrosis [62]. Additionally, skeletal abnormalities, particularly affecting the spine or pelvis, can predispose animals to obstructive dystocia. In chelonian species, such as spur-thighed tortoises (Testudo graeca), congenital deformities or narrowed pelvic outlets may be radiographically detectable and should be considered during reproductive evaluations [26].
4.2. Ultrasonography
Ultrasonography is the modality of choice for assessing POFS, soft tissue changes in the oviducts, and associated coelomic pathology, especially in lizards and snakes. Ultrasonography can be performed in dorsal or lateral recumbency or even upright position using a high-frequency linear transducer. For chelonians, the prefemoral fossa provides an acoustic window to the reproductive tract, but effective visualization depends on the size of animal and the shape of the transducer. Usually, adults have well-developed fossae, while in smaller or younger individuals, limited soft tissue access may hinder probe placement and diagnostic clarity [10,63]. Developing follicles show a homogenous, round, anechoic appearance, and may be arranged in clusters. Only in chelonians do already small follicles appear hyperechoic. Normal oviducts should not be distended or surrounded by hyperechoic fat or fluid. In gravid females, normal oviductal eggs appear as oval anechoic to mildly hypoechoic structures with a thin hyperechoic wall. As calcification progresses, distal acoustic shadowing may begin to appear [60,64] (Figure 4).
Recent work on female veiled chameleons (Chamaeleo calyptratus) has demonstrated that follicular atresia is a more dynamic and diagnostically complex process than previously assumed. In a longitudinal imaging study using ultrasonography and CT, four distinct stages of follicular development, previtellogenesis, vitellogenesis, gravidity, and atresia, were identified, and subsequently confirmed by histology. Atretic follicles were frequently present in multiple generations within the same animal, often without apparent disruption of normal folliculogenesis, suggesting that atresia can be a physiological process across several cycles. However, this diagnostic nuance presents clinical challenges: echogenic changes seen during ultrasonography (e.g., heterogeneous content and irregular follicular shape) can mimic POFS, potentially leading to overtreatment, including unnecessary surgical or hormonal interventions [49].
In cases of POFS, retained follicles remain within the ovary without evidence of ovulation. Ultrasonographically, these structures appear as enlarged, rounded, anechoic or heterogeneously hypoechoic areas, often devoid of peripheral vascular flow on Doppler examination [64] (Figure 5). In veiled chameleons, follicles exceeding 10 mm in diameter and persisting for several weeks without regression are considered diagnostic for POFS [55]. In more chronic cases of POFS or reproductive failure, follicular regression or atresia may result in internal echoes, septations, or hyperechoic rims, which can mimic abscesses or neoplastic masses. Mineralization of retained follicles has also been reported in chronic or advanced stasis [29]. Finally, Doppler ultrasonography may aid in differentiating pathologies: the absence of peripheral vascular flow supports a diagnosis of inactive or atretic follicles, while hypervascularization may suggest inflammation, infection, or neoplasia [64].
POD is characterized ultrasonographically by oviductal eggs that may appear irregularly shaped, collapsed, or surrounded by echogenic coelomic fluid, among other features. In a reported case in a bearded dragon, ultrasonography revealed free coelomic fluid and distorted oviductal structures encasing abnormally positioned eggs, consistent with dystocia complicated by yolk coelomitis [56]. In more severe presentations, oviductal distension or impaction may be evident, often manifesting as tubular, fluid-filled structures with echogenic debris. The presence of gas suggests secondary bacterial infection or necrosis, while mural thickening and loss of the normal oviductal wall layering may also be visualized [65,66].
4.3. Computed Tomography
CT has emerged as a powerful diagnostic modality in the assessment of reptile dystocia, offering high resolution and three-dimensional visualization of mineralized and soft tissue structures. In chelonians, even very small ovarian follicles can usually be identified on non-contrast CT, whereas in smaller lizards intravenous (IV) contrast medium is often required to reliably delineate ovarian and oviductal soft tissues. For further evaluation of vascular changes in the oviductal or ovarian tissues contrast medium application is needed [60]. CT allows precise evaluation of the number, shape, and positioning of eggs or follicles, which is particularly useful in species with complex internal anatomy such as snakes or in chelonians being enclosed in a bony shell. It also overcomes the limitations posed by complex external anatomy, like the bony shells of chelonians and osteodermal plates of crocodilians, that often obscure or distort conventional imaging modalities [60].
For example, in a Greek tortoise, CT imaging was used to identify a congenital pelvic narrowing that contributed to recurrent dystocia [44]. CT also allows for clear differentiation between shelled eggs, calcified follicles, and inflammatory or necrotic debris based on tissue density, which is essential in chronic or complicated cases [60]. CT was successfully used in ball python (Python regius) to identify malformed and obstructively positioned post-ovulatory eggs causing oviductal dilation [10]. Furthermore, CT aids in detecting secondary complications, such as coelomic gas, fluid, or oviductal rupture. It also plays a critical role in pre-surgical planning by delineating anatomical landmarks. Although the technique may require anaesthesia in for example agile individuals and requires access to specialized equipment, its diagnostic precision makes it an invaluable tool in the evaluation of suspected dystocia cases in reptiles. Although MRI has not yet been widely studied in reptilian reproductive medicine, it holds theoretical promise for superior soft tissue contrast, especially in visualizing vascularization, oviductal edema, or early-stage follicular structures, suggesting a potential future role alongside ultrasound and CT [67]. However, it usually requires general anesthesia and takes much longer than CT.
4.4. Endoscopy
Endoscopic evaluation, particularly cloacoscopy and coelioscopy, constitutes a minimally invasive and highly informative diagnostic approach in cases of reptile dystocia. Cloacoscopy is especially useful in cases of POD to directly visualize retained eggs in the caudal oviduct or cloaca, and to assess mucosal health, obstructions, or signs of infection. This technique allows for targeted flushing, biopsy, or even assisted egg removal without the need for full surgical intervention [68].
Coelioscopy is typically performed via the prefemoral approach in lizards and chelonians, and allows for direct inspection of ovarian follicles, oviducts, and associated structures within the coelomic cavity [69]. For example, in a case series of female bearded dragons with suspected POFS, coelioscopy enabled clear visualization of enlarged, regressing follicles and oviductal abnormalities, which guided decision-making for salpingectomy [70]. Endoscopy also allows dynamic assessment (e.g., identifying adhesions, inflammation, or coelomic effusion) and can be used therapeutically for follicular aspiration or lavage. While endoscope-assisted coeliotomy requires specialized training and equipment, it can enhance diagnostic accuracy and reduce invasiveness in select cases compared to coelioscopic plastrotomy. However, in chelonians, particularly tortoises, the endoscopic approach often demands longer operative time with no clear wound-healing advantage. Moreover, anatomical constraints, e.g. shell morphology, frequently limit access [71]. In chelonians, an important additional consideration is the occurrence of ectopic eggs in the urinary bladder. Such misdirected eggs can arise acutely during dystocia or present later as mineralized uroliths, with potential for chronic irritation, hematuria, or lower urinary tract dysfunction. Radiography (sometimes with retrograde contrast media application), CT and even sonography may be used to detect these mineralized structures. However, cystoscopy may be the preferred tool, enabling direct visualization and minimally invasive retrieval with snares or baskets [72,73]. Recognition of this phenomenon is vital: retention of ectopic eggs can complicate medical treatment of dystocia, obscure interpretation of imaging and delaying required intervention, thus bridging the diagnostic pathways of dystocia and urolithiasis.
4.5. Hematology
Comprehensive blood analyses, including hematology and biochemistry, are vital for diagnosis and management. Common hematologic findings in reptiles with dystocia include heterophilia, anemia, and monocytosis, consistent with chronic inflammation or stress responses. For instance, in Greek tortoises, follicular stasis has been associated with anemia and leukopenia. Heterophilia and monocytosis are observed in cases of ovarian teratomas and chronic egg retention, reflecting inflammatory responses [59]. A retrospective study in green iguanas with POFS reported mild non-regenerative anemia and an elevated total white blood cell count with a predominance of heterophils and mild non-regenerative anemia (Pack cell volume (PCV) 18–24%, reference range: 25–45%) [24]. However, interpreting anemia in reptiles requires caution, as venipuncture techniques often dilute samples with lymphatic fluid, particularly when using tail or subcarapacial sinuses, resulting in artifactually low PCV, RBC, and protein values [74]. Dystocic chameleons showed increased monocyte percentages and elevated AST levels, indicating tissue trauma associated with egg retention [75]. Finally, in African rock pythons (Python sebae), studies have reported variations in white blood cell counts, with elevated heterophils and lymphocytes in individuals with reproductive disorders, although specific values vary [76].
4.6. Blood Biochemistry
Metabolic disturbances, including alterations in blood glucose, are not uncommon in reproductive disease in reptiles. Hypoglycemia may be observed in pathological states such as egg yolk coelomitis [75]. However, it is important to recognise that some species, such as the common chameleon (Chamaeleo chamaeleon), exhibit higher baseline glucose levels, with mean values around 362 mg/dL, compared to the typical reptilian reference range of 60–100 mg/dL [75]. Increased AST activity, a marker of hepatic or muscular injury, was significantly elevated in dystocic chameleons compared to healthy controls. The frequent concurrent observation of hepatic lipidosis in these cases provides further support for a potential link between hepatic dysfunction and reproductive complications such as dystocia. Elevated creatine phosphokinase (CPK) levels may indicate skeletal muscle damage associated with prolonged egg retention. It is important to emphasize that it remains unclear whether these values exceeded established reference ranges, which are often broad or derived from limited sample sizes in exotic species. This underscores the challenge of interpreting clinicopathologic changes in reptiles, particularly when population-based reference intervals are poorly defined. When assessed alongside AST, CPK can assist clinicians in distinguishing between muscular and hepatic sources of AST elevation, an important diagnostic consideration in reptiles presenting with dystocia. A disproportionately elevated AST with normal or only mildly increased CPK may suggest hepatic involvement, whereas concurrent increases in both enzymes may reflect muscle injury, which could result from coelomic straining due to dystocia, but may also arise from other causes such as trauma, handling, prolonged recumbency, or systemic illness [77]. Hypercalcemia and hyperphosphatemia are common during vitellogenesis. However, in cases of chronic egg retention, total calcium and ionized calcium levels may decrease due to continuous deposition on retained eggs [44].
Hepatic dysfunction is a key concern in reptiles presenting with reproductive arrest, particularly those experiencing prolonged follicular stasis or anorexia. Among the hepatic pathologies associated with reproductive disorders, hepatic lipidosis is most frequently reported and can substantially alter plasma biochemical profiles. Elevated AST is a common indicator of hepatocellular injury. In dystocic bearded dragons, AST concentrations have been documented between 180 and 520 international units (IU)/Liter (reference range: 50–250 IU/L), with some individuals presenting values still within the upper end of the normal range. While elevated AST may serve as an indicator of underlying hepatic or muscular compromise, values between 180 and 250 IU/L overlap with reference intervals and thus should not be interpreted in isolation. Given the variability and non-specificity of AST in reptiles, it is best regarded as a supportive finding rather than a definitive diagnostic marker [29,78]. Plasma bile acids have also proven to be sensitive markers of liver dysfunction in reptiles. In green iguanas diagnosed with chronic hepatopathies, histologically confirmed as hepatic lipidosis, cirrhosis, hepatocellular carcinoma, fibrosis, and hepatocellular degeneration, plasma bile acid levels averaged 84.85 ± 22.29 μmol/L after 48 hours of fasting, compared with 9.56 ± 8.52 μmol/L in healthy, fasted controls [79]. This finding confirms that various chronic liver diseases, including hepatic lipidosis, are associated with markedly elevated bile acids in green iguanas. However, it is important to note that such elevations were documented in end-stage liver disease; in subclinical cases such as vitellogenesis-induced hepatolipidosis, bile acid levels may remain within normal limits or even fall below detectable thresholds, limiting their diagnostic reliability in early reproductive hepatopathies. Hypoalbuminemia, defined as serum albumin levels below 15 g/L in green iguanas (reference range: 18–35 g/L), has been associated with chronic hepatic compromise and inadequate protein intake [24]. However, other differentials, such asprotein-losing nephropathy (PLN), should also be considered [80]. Accurate assessment of albumin concentrations in reptiles is method-dependent; photometric analyzers such as VetScan often underestimate true albumin levels due to protein-binding artifacts, and reliable quantification ideally requires gel electrophoresis, an approach that may be impractical in smaller species where sample volumes are limited. Additionally, while elevated uric acid levels are not specific to hepatic dysfunction, they may increase in cases of systemic illness or renal compromise. However, specific uric acid trends in dystocic individuals remain insufficiently characterized in the current literature.
Recent advances in reptile metabolomics have led to the identification of promising plasma biomarkers that can aid in the early diagnosis and monitoring of hepatic lipidosis. [81].
B-hydroxybutyric acid (bHBA) is a ketone body involved in lipid metabolism and energy regulation during periods of anorexia or increased metabolic demand, such as folliculogenesis. In bearded dragons, significantly reduced levels of bHBA have been strongly associated with hepatic lipidosis. A plasma bHBA concentration below 272 µmol/L demonstrated a sensitivity of 86% and specificity of 100% for detecting moderate to severe hepatic lipidosis [28]. The suppression of ketogenesis, reflected in low bHBA, may indicate an overwhelmed or dysfunctional liver unable to compensate metabolically. Thus, bHBA serves as an early warning sign of metabolic exhaustion and hepatic burden in dystocic reptiles [82]. A recent study established reference intervals for βHBA in healthy adult central bearded dragons and evaluated a point-of-care ketone meter against a reference laboratory analyzer. The point-of-care device consistently underestimated BHBA concentrations compared with the laboratory method, showing a proportional negative bias. However, the total observed error remained within clinically acceptable limits, indicating that the device is still appropriate for routine clinical monitoring. The authors proposed a simple correction factor (multiplying point-of-care results by 0.9) to improve agreement between methods. Importantly, no significant interference was detected from hematocrit, lipemia, hemolysis, or blood glucose. This study also provided the first species-specific βHBA reference intervals for Pogona vitticeps, supporting the use of βHBA as an accessible metabolic biomarker in the assessment of hepatic lipid metabolism in lizards [83].
Succinic acid, a key intermediate in the tricarboxylic acid cycle, has been identified as a promising biomarker for hepatic lipidosis in bearded dragons. In a recent metabolomics study, plasma succinic acid concentrations exceeding 13.7 µmol/L were significantly associated with moderate to severe hepatic lipid infiltration. At this threshold, succinic acid demonstrated a sensitivity of 86% and a specificity of 100% for detecting moderate to severe lipidosis, with an area under the receiver operating characteristic curve of 0.98 [82]. In reproductive females, increased succinic acid may indicate mitochondrial dysfunction or altered energy metabolism, which could further compromise oocyte maturation, vitellogenesis, or uterine contractions. This makes succinate not only a marker of hepatic stress, but also a potential marker for impaired reproductive physiology [84].
Differentiating between metabolic changes due to fasting and true hepatic lipidosis presents a key diagnostic challenge in dystocic reptiles. Prolonged anorexia is common in females with follicular stasis or egg retention, leading to fat mobilization and altered levels of plasma metabolites such as bHBA and succinic acid. However, these changes may occur in both adaptive fasting states and pathological hepatic lipidosis, complicating interpretation [28]. However, these changes may occur in both adaptive fasting states and pathological hepatic lipidosis, complicating interpretation. In such cases, liver cytology via fine-needle aspiration may aid in diagnosis, as hepatocellular vacuolation, lipid-laden hepatocytes, and the presence of inflammatory cells can help distinguish true hepatic lipidosis from fasting-induced hepatic metabolic shift [29,85]. Nevertheless, hepatic biochemical markers, despite their diagnostic limitations, may still serve as a useful tool in pre-anaesthetic risk assessment, particularly in reproductively active females where hepatic compromise may go undetected until challenged by anaesthesia or surgery. Non-esterified fatty acids (NEFAs), widely used in mammals to assess lipid mobilization, have not yet been validated in reptiles but hold promise as future biomarkers to distinguish physiological from pathological fat metabolism. If proven reliable, NEFA monitoring, especially in combination with bHBA, could offer valuable insight into energy balance and hepatic stress in females at risk of dystocia [86].
5. Hormonal Treatment
5.1. General Considerations
Hormonal therapy is commonly employed in POD once eggs have progressed into the oviduct, typically after shell deposition, when the reproductive tract can respond appropriately to contractile hormones. Before pharmacological stimulation is attempted, basic supportive and husbandry measures should be optimised, as many cases are precipitated or exacerbated by management errors; restoring an appropriate thermal gradient within the species’ preferred optimal temperature zone, minimising stress, and providing a quiet, suitable nesting area with an appropriate substrate are recommended and may be sufficient to allow normal oviposition in some non-obstructive cases [45,47]. Patients should also be stabilised with fluid therapy and correction of electrolyte and mineral imbalances, particularly hypocalcaemia, together with adequate analgesia and treatment of concurrent disease before any hormonal induction is considered [42,87]. Several authors advocate including parenteral calcium gluconate in medical protocols, frequently administered shortly before oxytocin or AVT in chelonians to enhance oviductal smooth-muscle contractility and improve the response to hormonal treatment, and as part of supportive management in lizards with dystocia [44,68]. Once the animal is adequately supported and obstructive causes have been excluded, oxytocin or can be administered to stimulate effective oviductal contractions via smooth-muscle receptors in both the oviduct and cloacal sphincter, facilitating egg passage. Premature administration, before shell calcification, risks serious complications such as oviduct rupture, hemorrhage, and yolk coelomitis [44,60].
5.2. Chelonians
Prior research using 253 female red-eared sliders demonstrated that intramuscular (IM) doses of 10 or 20 IU/kg oxytocin, given up to twice 60 minutes apart, resulted in egg passage in 100% of cases. The success of lower doses (4 or 5 IU/kg) was 88.9% after two injections [88]. Even at a reduced dose of 4 IU/kg, red-eared sliders, common musk turtles (Sternotherus odoratus), and painted turtles (Chrysemys picta) exhibited equally high oviposition success after two injections, suggesting broad species applicability [88]. A study involving hawksbill sea turtles (Eretmochelys imbricata) found that 0.6–0.8 IU/kg IM oxytocin triggered egg-laying in 8 out of 9 individuals, with oviposition beginning between 17–43 minutes after administration and completed within 24 hours [89]. In contrast, mediterranean tortoises appear to respond to substantially lower doses: in free-ranging Hermann’s tortoises, IM oxytocin at approximately 1.5–3 IU/kg reliably induced oviposition, with first eggs laid 13–68 minutes after injection and most clutches completed within about 20 minutes of the first egg [90].
5.3. Lizards
Hormonal therapy for POD in lizards typically involves a combined protocol of calcium supplementation followed by oxytocin administration. A well-documented case report in a bearded dragon describes successful oviposition of 14 eggs within 20 hours: the protocol included two IM injections of calcium borogluconate (35 mg/kg followed by 50 mg/kg, 30 minutes apart) followed by a single dose of oxytocin (5 IU, IM) shortly thereafter, with complete resolution and no further retained eggs on radiographs 24 hours later. The same individual successfully laid all 38 subsequent eggs uneventfully in the next cycle, suggesting both efficacy and safety of this approach. In contrast, a case series involving leopard geckos (Eublepharis macularius) and crested geckos (Correlophus ciliatus) demonstrated that medical management with calcium and oxytocin alone was insufficient. In that report, all affected geckos required surgical salpingotomy, followed by supportive therapy, highlighting that while lizards may be candidates for hormonal therapy, success is variable and some species exhibit poor responsiveness to medical protocols [ [20]. These estradiol-stimulating protocols align with broader clinical recommendations that assert oxytocin should be used within 48 hours of shelling, maintained at species-appropriate temperatures, and ideally following calcium correction. Timing is critical to avoid oviductal rupture or ineffective dosing. Though lizard success rates vary, usually cited between 30% and 70%, these peer-reviewed cases confirm that calcium-primed oxytocin therapy can effectively resolve dystocia in small lizards when administered promptly and with supportive care [20,56]. Nonetheless, larger species and delayed cases still frequently require surgical resolution.
5.4. Snakes
In snakes, hormonal therapy for dystocia has limited success, with reported effectiveness under 50%, even when administered at the appropriate post-ovulatory stage. The particularly elongated and tortuous oviducts combined with a lower density of oxytocin and AVT receptors likely lowers the effectiveness of oxytocin-based therapies in snakes compared to chelonians or lizards [8]. Intervention must occur within 48–72 hours after the onset of oviposition attempts. Delayed administration often results in obstructive dystocia, which carries a high risk of oviduct rupture if oxytocin is used. The typical oxytocin dosing protocol involves administering 5 IU/kg IM or intracoelomically (ICe), with a second dose of up to 20 IU/kg given 6–12 hours later if no response is observed [91]. If there is no improvement after two doses, medical therapy should be considered unsuccessful. AVT may be more effective due to its higher receptor affinity, with suggested dosages ranging from 0.01–1.0 µg/kg given IM, subcutaneous (SC), or ICe. However, AVT is rarely available outside of research settings, which limits its clinical application [91,92]. Adjunctive methods can include sedated egg manipulation using propofol (5–10 mg/kg IV) to facilitate cloacal relaxation, followed by manual or endoscopic removal of accessible eggs. In cases where retained eggs are visible but unresponsive to oxytocin, percutaneous ovocentesis may be considered only as a last-resort option, as it carries significant risks of yolk leakage, coelomitis, and oviductal injury and is generally not recommended when surgical management is feasible. This involves aspirating the contents of the egg using a 20-gauge needle under anesthesia to reduce volume and facilitate expulsion. This technique should only be used early in the dystocia process and always with readiness for surgical conversion, given the risk of yolk leakage and subsequent yolk coelomitis [91]. If the snake’s clinical condition deteriorates, evidenced by anorexia, increasing coelomic distension, or signs of sepsis, or if no response is seen after two hormonal treatments, surgical intervention is indicated. Several detailed case reports document successful non-surgical resolution of POD in snakes using hormonal and supportive interventions. A notable study described a 15 kg female albino Burmese python (Python bivittatus) with confirmed egg retention; following coelomic fluid therapy, ICe calcium gluconate (50 mg/kg SC, diluted), and 5 IU/kg IM oxytocin, the snake oviposited all 25 eggs within 12 hours of a second identical oxytocin dose given after 12 hours [93]. This shows effectiveness when medical therapy is initiated within a critical post-ovulatory window and combined with calcium and environmental support. Older veterinary literature also notes successful oxytocin use in oviparous snakes, with dosages from 0.4 to 10 IU/kg, achieving egg passage under appropriate conditions [8]. These cases reinforce the principle that oxytocin can be effective in early, non-obstructive dystocia in snakes.
5.5. Various
Beyond oxytocin and AVT, treatment with prostaglandins, calcium supplementation, and combination therapies have shown clinical value in treating dystocia. In a case report involving leopard geckos and crested geckos, Di Giuseppe et al. administered IM calcium gluconate at 35 mg/kg, followed by ICe oxytocin, and noted that in some cases additional PGF₂α was experimented with, facilitating uterine contraction and completion of egg passage [20]. The authors emphasized that prostaglandins must be used cautiously due to limited reptile data, yet when combined with calcium and oxytocin, they helped avoid surgery in small lizards.
Although literature is scarce for snakes, hybrid approaches have been documented. A Burmese python case incorporated calcium before 5 IU/kg oxytocin IM, followed by a second dose, to facilitate passage of 25 eggs, illustrating that combined protocol principles apply beyond lizards [93,94]. Experimental studies in oviparous lizards have demonstrated that ICe administration of prostaglandin F₂α at doses between 50–200 µg/kg effectively induces uterine contractions and oviposition when oxytocin alone fails [95].
These cases demonstrate that combined protocols, calcium priming, oxytocin, and prostaglandin administration, can significantly improve outcomes and reduce surgical necessity when applied early in POD. However, if two to three cycles over 48–72 hours with combination therapy yield no response or if systemic deterioration ensues, surgical intervention remains the indicated next step [20,91].
6. Surgical Intervention Thresholds
6.1. General Considerations
Indications for surgery include non-response to oxytocin or AVT therapy after 24–48 hours of appropriate dosing, especially in the absence of uterine contractions or egg movement on imaging [47].
Worsening clinical condition, manifested as persistent anorexia, weight loss, lethargy, regurgitation, or weakness despite supportive therapy, should prompt surgical reassessment. In these cases, waiting may increase the risk of egg calcification, oviductal rupture, or yolk peritonitis.
Imaging findings play a pivotal role. Failure of eggs to progress or reposition on serial radiographs, CT, or ultrasonographic examinations despite appropriate husbandry and medical therapy, prolonged egg retention over several weeks accompanied by clinical deterioration, or excessive oviductal distension with mechanically obstructive eggs (e.g., markedly oversized, fused, or severely deformed eggs wedged within the pelvic canal) are widely regarded as indications to consider surgical management, rather than egg shape alone [44,47,60,63]. However, in chelonians, mildly irregular or malshaped eggs may occur in otherwise uncomplicated clutches and do not, by themselves, constitute dystocia; surgical decision-making should therefore be based on the overall clinical picture and imaging evidence of obstruction or secondary coelomic disease, not morphology in isolation [44,60,63]. Egg retention into late summer or early autumn may reflect physiological preparation for hibernation rather than dystocia. In such cases, otherwise static eggs may still respond to oxytocin or AVT therapy, and surgical intervention is not always indicated [63]. Ultrasound or radiographic signs of dystrophic or malformed eggs, coupled with coelomic effusion or gas shadows, indicate possible oviductal rupture or infection and warrant urgent intervention [47].
Laboratory abnormalities, such as marked heterophilia, hyperuricemia, elevated AST or CK, or hypoalbuminemia, may support the presence of secondary infection, hepatic compromise, or metabolic exhaustion, all of which lower the threshold for surgery [29]. Leukocytosis, especially when accompanied by a left shift or toxic heterophils, is a well-established indicator of systemic inflammation or sepsis in reptiles [96].
Finally, failure to lay despite multiple ovulatory cycles or visible retained follicles across seasons may represent chronic follicular stasis, which often requires ovariosalpingectomy due to fibrosis, adhesions, and risk of neoplasia. Prompt surgical resolution, ideally before systemic decompensation, has been shown to improve survival, recovery, and future reproductive success [44].
Surgical management of dystocia in reptiles varies significantly depending on species anatomy and the reproductive structures involved. In snakes, a ventrolateral coeliotomy, with the skin incision made between the first and second rows of lateral scales, is the standard approach to access the oviducts, avoiding disruption of the ventral scales and allowing improved post-operative ambulation [61]. A salpingotomy, involving singular or several direct incision(s) into the oviduct to remove retained ova, may be performed if fertility preservation is desired and the contralateral tract appears unaffected. More severe or recurrent cases may warrant a salpingectomy or complete ovariosalpingectomy [97,98].
In lizards with POD, the recommended surgical approach is a ventral paramedian coeliotomy followed by salpingotomy, allowing direct removal of retained eggs while preserving ovarian tissue when fertility is to be maintained [61]. In cases of POFS, a bilateral or unilateral ovariectomy or ovariosalpingectomy is considered the treatment of choice, effectively resolving the underlying hormonal imbalance and preventing recurrence [99]. In breeding individuals, where fertility preservation is desirable and disease is unilateral, a hemi-ovariosalpingectomy may be performed to remove only the affected tract while maintaining reproductive potential [17]. In cases of advanced reproductive disease with concurrent systemic compromise (e.g., hepatic lipidosis or coelomitis), open coeliotomy is preferred to allow full exploration, removal of diseased tissues, and thorough lavage [61].
6.2. Considerations for Hepatic and Metabolic Derangements
Reptiles with hepatic lipidosis or broader hepatic dysfunction present unique challenges under anaesthesia due to alterations in pharmacokinetics, drug clearance, and physiological stability. The liver plays a central role in metabolizing most injectable anaesthetic agents, including ketamine, medetomidine, alphaxalone, and midazolam. In hepatic lipidosis, hepatocellular lipid infiltration impairs enzymatic function, leading to delayed drug clearance, prolonged sedation, and increased sensitivity to standard anaesthetic doses [24,100]. For example, studies in reptiles have demonstrated prolonged recovery times following alphaxalone or ketamine–medetomidine protocols in individuals with suspected hepatic dysfunction, attributed to reduced hepatic blood flow and enzyme activity [100,101].
Additionally, hypoalbuminemia, a common consequence of liver disease, may increase the free fraction of protein-bound anaesthetic drugs, increasing effective plasma levels, with subsequent risks of toxicity [102]. This is of particular concern with agents like midazolam and propofol, which are highly protein-bound in circulation [103]. Anemic or hypovolemic patients, which are frequently seen in hepatic lipidosis due to anorexia or chronic systemic inflammation, are also less tolerant of vasodilation and hypotension under inhalant anaesthesia [15]. Therefore, caution is warranted when selecting both injectable and inhalant agents in liver-compromised reptiles.
Where possible, inhalant anaesthesia, particularly sevoflurane or isoflurane, is preferred over injectable protocols in reptiles with compromised hepatic function, as these volatile agents are primarily eliminated via the lungs with minimal hepatic metabolism [104,105]. While these agents can cause peripheral vasodilation and dose-dependent hypotension, this effect is predictable and manageable, and the anaesthetic depth can be rapidly titrated or reversed by adjusting inspired concentrations [105]. However, achieving surgical depth with inhalants alone may result in marked respiratory depression or apnea in many reptiles, limiting titration unless controlled ventilation is available. In such cases, a multimodal anaesthetic approach, combining low doses of injectable agents with inhalants, can reduce the dose-dependent side effects of individual drugs while improving anaesthetic safety and cardiovascular stability. In contrast, injectable agents often rely on hepatic metabolism and may accumulate in reptiles with impaired clearance, leading to prolonged and potentially deeper hypotension [15]. If opioids are desireds, use agents such as butorphanol (1–2 mg/kg IM q12h) that are less reliant on hepatic clearance [92]. However, even with inhalants, hepatic perfusion may still be reduced under anaesthesia, especially in dehydrated or cold-stressed patients [106].
Adequate hydration is critical to improve hepatic perfusion and drug clearance in lipidosis-affected reptiles. Isotonic fluids like Lactated Ringer’s at 20–30 mL/kg/day SC or IV/intraosseous (IO) are recommended after assessing dehydration level . In cases of hypoalbuminemia or coelomic effusion, colloid administration (e.g. hetastarch 1–2 mL/kg IV) or plasma (10 mL/kg) may be used to support oncotic pressure, though data in reptiles is limited [27]. Reptiles with hepatic lipidosis frequently present anorectic and in a negative energy balance, exacerbating fat accumulation in the liver. Early nutritional support via syringe feeding or assisted tube feeding with high-protein, low-fat reptile diets (e.g., Carnivore, Omnivore or Herbivore Critical Care) helps reverse catabolism [29]. In parallel, supplementation with L-carnitine, widely use in small animal hepatic lipidosis, may offer a promising adjunctive treatment in reptiles, though clinical trials are still needed. A dosage of 250 mg/kg orally daily in reptiles may enhance mitochondrial β-oxidation and support hepatic recovery, but further investigation is warranted [107]. Chronic illness in hepatic lipidosis can result in anemia, hypoalbuminemia, and coagulopathies. Hematologic profiles should be obtained; values like packed cell volume <15% or albumin <1.5 g/dL indicate the need for stabilization before anaesthesia [27].
To aid clinicians in applying the surgical thresholds described above, we include a flowchart that integrates patient status, diagnostic findings, and response to medical therapy to guide when surgical management becomes appropriate (Figure 6).
7. Future Directions and Clinical Gaps
Despite growing clinical experience and an increase in case documentation, the management of dystocia in reptiles is still limited by several fundamental evidence gaps. One of the most pressing needs is the development of species-specific hormonal therapy protocols. The effectiveness of oxytocin, PGF₂α, and AVT varies significantly between taxonomic groups, in part due to differences in oviductal receptor expression and endocrine sensitivity [108]. However, most therapeutic protocols remain extrapolated from mammalian practice or isolated case reports, underlining the urgent need for controlled trials in commonly affected reptile species.
Another critical gap lies in the lack of validated liver biomarkers to assess the anaesthetic and surgical risk in dystocic reptiles. While hepatic lipidosis and hepatic compromise are common comorbidities, especially in cases of prolonged follicular retention, traditional liver enzymes such as AST, alanine aminotransferase, and bile acids demonstrate high interspecific variability and poor correlation with functional reserve. This makes preoperative risk assessment challenging and inconsistent. Further research is needed to establish species-specific reference intervals and explore the use of additional biomarkers such as NEFA and sorbitol dehydrogenase, which may offer improved diagnostic sensitivity.
Preventative medicine in reproductive care is also underdeveloped. There is currently limited research on pre-emptive support strategies for high-risk breeding females. Evidence from herpetoculture and field ecology suggests that thermal gradients, nesting site quality, and pre-breeding nutritional conditioning all impact reproductive outcomes [13,14]. However, structured studies evaluating prophylactic calcium, vitamin D₃, or micronutrient supplementation protocols, particularly in captivity, are still lacking.
Another area of inconsistency is the criteria used to determine the need for surgical intervention. Without a standardized scoring system, the decision to move from medical to surgical management is highly variable between clinicians. Similarly, the interpretation of imaging data, such as ultrasound evidence of uterine wall thinning, egg deformation, or pelvic obstruction, remains subjective. The development of validated diagnostic thresholds could guide clinical decision-making and reduce delays that contribute to morbidity and mortality.
In addition to biochemical and hepatic contributors to reproductive failure in reptiles, emerging research suggests that factors such as endocrine-disrupting chemicals (EDCs), reproductive tract microbiota, and trace element imbalances (e.g., selenium, zinc) may also play a role. While not yet well studied in reptiles, EDCs have been shown to impair reproductive endocrinology in alligators and amphibians [109,110]. Moreover, alterations in the reproductive microbiome have been linked to implantation failure in mammals [111]. Similarly, trace minerals influence hepatic antioxidant capacity and reproductive hormone regulation in other vertebrates [112].These findings support future investigations into their relevance in reptilian dystocia.
8. Conclusion
This review reinforces that dystocia in reptiles is not merely a mechanical obstruction but often the clinical manifestation of a complex and self-perpetuating interplay between endocrine dysregulation, hepatic dysfunction, and inappropriate environmental conditions. Hepatic lipidosis, hypoalbuminemia, and altered oestrogen metabolism emerge as key mediators of reproductive arrest, particularly in species subject to prolonged vitellogenesis without oviposition. Although calcium-primed oxytocin remains the most widely applied medical protocol, therapeutic success varies considerably across taxa, reflecting underlying differences in receptor pharmacodynamics, metabolic resilience, and reproductive anatomy. Diagnostic advancements, including serial liver function profiling, βHBA and succinate quantification, and high-resolution imaging, present promising avenues for early case stratification and perioperative planning. However, several key gaps remain, including how chronic hepatic lipidosis affects reproductive hormone clearance and receptor responsiveness; whether βHBA and succinic acid can serve as reliable, non-invasive predictors of dystocia severity and anesthetic risk; and how oxytocin pharmacokinetics and receptor expression differ across reptilian lineages. Additionally, the potential for endocrine or nutritional modulation during vitellogenesis to reduce follicular arrest has yet to be fully explored. Addressing these questions will not only refine clinical protocols, but also advance our broader understanding of reproductive energetics and endocrine-immune crosstalk in ectothermic vertebrates.
Author Contributions
Conceptualization, M.M.; writing—original draft preparation, M.M.; writing—review and editing, M.M., M.G., H.K, R.A.N., and J.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| AST | Aspartate aminotransferase |
| AVT | Arginine vasotocin |
| βHBA | Beta-hydroxybutyric acid |
| CK | Creatine phosphokinase |
| CT | Computed tomography |
| EDC | Endocrine-disrupting chemicals |
| ERα | Oestrogen receptor alpha |
| ERβ | Oestrogen receptor beta |
| ICe | Intracoelomic |
| IU | International units |
| IO | Intraosseous |
| IM | Intramuscular |
| IV | Intravenous |
| NEFA | Non-esterified fatty acids |
| PGF₂α | Prostaglandin F2 alpha |
| POFS | Pre-ovulatory follicular stasis |
| POD | Post-ovulatory dystocia |
| SC | Subcutaneous |
| UVB | Ultraviolet B radiation |
References
- Portas, T. Disorders of the Reproductive System; 2017; pp. 307–321. [Google Scholar]
- Stewart, J.R.; Thompson, M.B. Evolution of placentation among squamate reptiles: recent research and future directions. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2000, 127, 411–431. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, D.G. Evolution of viviparity in squamate reptiles: Reversibility reconsidered. J. Exp. Zool. B Mol. Dev. Evol. 2015, 324, 473–486. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, D.G. Structure, function, and evolution of the oviducts of squamate reptiles, with special reference to viviparity and placentation. J. Exp. Zool. 1998, 282, 560–617. [Google Scholar] [CrossRef]
- Guillette, L.J., Jr.; Cree, A.; Rooney, A.A.; et al. Effects of arachidonic acid, prostaglandin F2α, prostaglandin E2, and arginine vasotocin on induction of birth in a viviparous lizard (Sceloporus jarrovi). Gen. Comp. Endocrinol. 1992, 85, 477–485. [Google Scholar] [CrossRef]
- Jones, S. Hormonal Regulation of Ovarian Function in Reptiles; 2024; pp. 89–114. [Google Scholar]
- Mahmoud, I.Y.; DeSanti, A.; Tucker, R.K. The role of arginine vasotocin and prostaglandin F2α on oviposition and luteolysis in the common snapping turtle (Chelydra serpentina). Gen. Comp. Endocrinol. 1988, 69, 56–64. [Google Scholar] [CrossRef]
- DeNardo, D. Dystocias. In Reptile Medicine and Surgery; Mader, D.R., Ed.; W.B. Saunders: Philadelphia, PA, USA, 1996; pp. 370–374. [Google Scholar]
- Krohmer, R.; Lutterschmidt, D. Environmental and Neuroendocrine Control of Reproduction in Snakes. In Reproductive Biology and Phylogeny of Snakes; 2011; Volume 9, pp. 289–346. [Google Scholar]
- Stahl, S.J. Reptile Obstetrics. The North American Veterinary Conference, 2006; p. 4. [Google Scholar]
- Kummrow, M.; Cigler, P.; Mastromonaco, G. Revisiting “Preovulatory Follicular Stasis” in Reptiles. J. Zoo Wildl. Med. 2025, 56. [Google Scholar] [CrossRef]
- Kottwitz, J.; Bohlsen, E.; Kane, L.; Sladky, K. Evaluation of the follicular cycle in ball pythons (Python regius). J. Herpetol. Med. Surg. 2016. [Google Scholar] [CrossRef]
- Huchzermeyer, F.; van Wyk, W. Crocodiles—Biology, husbandry and diseases. J. S. Afr. Vet. Assoc. 2003, 74. [Google Scholar] [CrossRef]
- Grigg, G.; Kirshner, D. Biology and Evolution of Crocodylians; 2015. [Google Scholar]
- Mader, D.; Divers, S. Current Therapy in Reptile Medicine and Surgery; 2014. [Google Scholar]
- Doneley, B.; Monks, D.; Johnson, R.; Carmel, B. Reptile Medicine and Surgery in Clinical Practice; Wiley Blackwell, 2017; pp. 1–500. [Google Scholar]
- Gartrell, B.D.; Mellow, D.; McInnes, K.; Nelson, N. Folliculectomy for the treatment of pre-ovulatory follicular stasis in three illegally captured West Coast green geckos (Naultinus tuberculatus). N. Z. Vet. J. 2024, 72, 334–340. [Google Scholar] [CrossRef]
- Guraya, S.S. Ovarian Follicles in Reptiles and Birds. In Zoophysiology; Springer: Berlin/Heidelberg, Germany, 1989; Volume 24. [Google Scholar]
- Dervas, E.; Duchesne, J.M.; van Beurden, S.J.; et al. Morphological evidence for the physiological nature of follicular atresia in veiled chameleons (Chamaeleo calyptratus). Anim. Reprod. Sci. 2024, 261, 107409. [Google Scholar] [CrossRef]
- Di Giuseppe, M.; Silvestre, A.M.; Luparello, M.; Faraci, L. Post-ovulatory dystocia in two small lizards: Leopard gecko (Eublepharis macularius) and crested gecko (Correlophus ciliatus). Russ. J. Herpetol. 2017, 24, 128–132. [Google Scholar] [CrossRef]
- Martinez Silvestre, A. Hepatic lipidosis in reptiles. Southern Europe Veterinary Conference SEVC–AVEPA 2013, 48, 1–4. [Google Scholar]
- Divers, S.J.; Stahl, S.J. Mader’s Reptile and Amphibian Medicine and Surgery, 3rd ed.; Elsevier: St. Louis, MO, USA, 2019. [Google Scholar]
- Divers, S.; Lock, B.; Camus, M. Comparison of protein electrophoresis and biochemical analysis for the quantification of plasma albumin in healthy bearded dragons (Pogona vitticeps). J. Zoo Wildl. Med. 2021, 52. [Google Scholar] [CrossRef] [PubMed]
- Mayer, J. Reptile Medicine and Surgery, 2nd Edition. In J. Exot. Pet Med.; Mader, D.R., Ed.; Elsevier Saunders, 2006; Volume 15, pp. 238–239. [Google Scholar]
- Donoghue, S.; McKeown, S. Nutrition of captive reptiles. Vet. Clin. N. Am. Exot. Anim. Pract. 1999, 2, 69–91. [Google Scholar] [CrossRef] [PubMed]
- Burgess, M.E. Common reproductive pathologies in reptiles. Clin. Theriogenol. 2010, 2, 1–6. [Google Scholar]
- Divers, S.J.; Cooper, J.E. Reptile hepatic lipidosis. Semin. Avian Exot. Pet Med. 2000, 9, 153–164. [Google Scholar] [CrossRef]
- Barboza, T.K.; Susta, L.; Zur Linden, A.; Gardhouse, S.; Beaufrère, H. Association of plasma metabolites and diagnostic imaging findings with hepatic lipidosis in bearded dragons (Pogona vitticeps) and effects of gemfibrozil therapy. PLoS One 2023, 18, e0274060. [Google Scholar] [CrossRef]
- Barboza, T.; Susta, L.; Reavill, D.; Beaufrère, H. Prevalence and risk factors of hepatic lipid changes in bearded dragons (Pogona vitticeps). Vet. Pathol. 2023, 60, 133–138. [Google Scholar] [CrossRef]
- Barboza, T.; Beaufrère, H.; Reavill, D.; Susta, L. Morphological features of hepatic lipid changes in bearded dragons (Pogona vitticeps), and a proposed grading system. Vet. Pathol. 2023, 60, 123–132. [Google Scholar] [CrossRef]
- Gumpenberger, M.; Filip, T.; Grabensteiner, E.; Nowotny, N. Diagnostic imaging of various forms of dystocia in chelonians. Wien. Tierärztl. Monschr. 2001, 88, 70–78. [Google Scholar]
- Gumpenberger, M. Veterinary computed tomography. In Veterinary Computed Tomography; Schwarz, T., Saunders, J., Eds.; Wiley-Blackwell: UK, 2011; pp. 533–544. [Google Scholar]
- Colon, V.; Gumpenberger, M. Diagnosis of hepatic lipidosis in a tiger salamander (Ambystoma tigrinum) by computed tomography. J. Exot. Pet Med. 2020, 33, 12–22. [Google Scholar] [CrossRef]
- Marchiori, A.; da Silva, I.C.; de Albuquerque Bonelli, M.; de Albuquerque Zanotti, L.C.; Siqueira, D.B.; Zanotti, A.P.; et al. Use of computed tomography for investigation of hepatic lipidosis in captive Chelonoidis carbonaria (Spix, 1824). J. Zoo Wildl. Med. 2015, 46, 320–324. [Google Scholar] [CrossRef] [PubMed]
- Gumpenberger, M.; Kuebber-Heiss, A.; Richter, B. CT as a quick non-invasive imaging tool for diagnosing hepatic lipidosis in reptiles. Proceedings of WSAVA FECAVA, 2013. [Google Scholar]
- Sosa-Higareda, M.; Beaufrère, H. Diet type, fasting duration, and computed tomography hepatic attenuation influence postprandial plasma lipids, β-hydroxybutyric acid, glucose, and uric acid in bearded dragons (Pogona vitticeps). Am. J. Vet. Res. 2025, 86. (in press). [Google Scholar] [CrossRef] [PubMed]
- X., M. A synthetic review: natural history of amniote reproductive modes in light of comparative evolutionary genomics. Biol. Rev. 2025, 100, 362–406. [Google Scholar] [CrossRef]
- Heck, J.; MacKenzie, D.S.; Rostal, D.; Medler, K.; Owens, D. Estrogen induction of plasma vitellogenin in the Kemp’s ridley sea turtle (Lepidochelys kempi). Gen. Comp. Endocrinol. 1997, 107, 280–288. [Google Scholar] [CrossRef]
- Stacy, B.A.; Howard, L.; Kinkaid, J.; Vidal, J.D.; Papendick, R. Yolk coelomitis in Fiji Island banded iguanas (Brachylophus fasciatus). J. Zoo Wildl. Med. 2008, 39, 161–169. [Google Scholar] [CrossRef]
- Hammouche, S.B.; Remana, S.; Exbrayat, J.-M. Immunolocalization of hepatic estrogen and progesterone receptors in the female lizard Uromastyx acanthinura. C. R. Biol. 2012, 335, 445–453. [Google Scholar] [CrossRef]
- Stanford, M.; Keen, J.; Campbell-Ward, M. Hypocalcemia in other species. Clinical Endocrinology of Companion Animals 2013, 335–355. [Google Scholar]
- Rivera, S. Health assessment of the reptilian reproductive tract. J. Exot. Pet Med. 2008, 17, 259–266. [Google Scholar] [CrossRef]
- Gardner, B.; Barrows, M. Yolk coelomitis in a white-throated monitor lizard (Varanus albigularis). J. S. Afr. Vet. Assoc. 2010, 81, 121–122. [Google Scholar] [CrossRef]
- Lubian, E.; Palotti, G.; Di Ianni, F.; Vetere, A. Disorders of the female reproductive tract in chelonians: A review. Animals 2025, 15, 1275. [Google Scholar] [CrossRef] [PubMed]
- Hedley, J. Reproductive diseases of reptiles. In Pract. 2016, 38, 457–462. [Google Scholar] [CrossRef]
- McWilliams, D.A.; Leeson, S. Metabolic bone disease in lizards: Prevalence and potential for monitoring bone health. In Metabolic Bone Disease in Lizards; 2001. [Google Scholar]
- Sykes, J.M. Updates and practical approaches to reproductive disorders in reptiles. Vet. Clin. N. Am. Exot. Anim. Pract. 2010, 13, 349–373. [Google Scholar] [CrossRef] [PubMed]
- Cerreta, A.J.; McEntire, M.S. Hypothalamic and pituitary physiology in birds and reptiles. Vet. Clin. N. Am. Exot. Anim. Pract. 2025, 28, 51–68. [Google Scholar] [CrossRef]
- Cigler, P.; Dervas, E.; Richter, H.; Hatt, J.-M.; Kummrow, M. Ultrasonographic and computed tomographic characteristics of the reproductive cycle in female veiled chameleons (Chamaeleo calyptratus). J. Zoo Wildl. Med. 2023, 54, 231–243. [Google Scholar] [CrossRef]
- Efendić, M.; Samardžija, M.; Prvanović Babić, N.; Bacić, G.; Karadjole, T.; Lojkić, M.; et al. Postovulatory egg retention (dystocia) in lizards—Diagnostic and therapeutic options. Kleintierpraxis 2017, 62, 754–764. [Google Scholar]
- Work, T.M.; Balazs, G.H. Pathology and distribution of sea turtles landed as bycatch in the Hawaii-based North Pacific pelagic longline fishery. J. Wildl. Dis. 2010, 46(2), 422–432. [Google Scholar] [CrossRef]
- Di Girolamo, N.; Selleri, P. Reproductive disorders in snakes. Vet. Clin. N. Am. Exot. Anim. Pract. 2017, 20(2), 391–409. [Google Scholar] [CrossRef]
- Anderson, G.; Currylow, A.; Ridgley, F. Egg retention in wild-caught Python bivittatus in the Greater Everglades Ecosystem, Florida, USA. Herpetol. J. 2022, 32, 109–113. [Google Scholar] [CrossRef]
- Williams, S.J.; Davies, D. Case report—dystocia in an African rock python. Br. Herpetol. Soc. Bull. 1993, 45. [Google Scholar]
- Hall, A.J.; Lewbart, G.A. Treatment of dystocia in a leopard gecko (Eublepharis macularius) by percutaneous ovocentesis. Vet. Rec. 2006, 158(21), 737–739. [Google Scholar] [CrossRef] [PubMed]
- Efendić, M.; Samardžija, M.; Capak, H.; Bacić, G.; Žura Žaja, I.; Magaš, V.; et al. Induction of oviposition in bearded dragon (Pogona vitticeps) with postovulatory egg retention (dystocia)—a case report. Vet. Arhiv. 2019, 89, 131–142. [Google Scholar] [CrossRef]
- Kummrow, M.S.; Mastromonaco, G.F.; Crawshaw, G.; Smith, D.A. Fecal hormone patterns during non-ovulatory reproductive cycles in female veiled chameleons (Chamaeleo calyptratus). Gen. Comp. Endocrinol. 2010, 168(3), 349–355. [Google Scholar] [CrossRef] [PubMed]
- Pimm, R.H.; Dutton, C.J.; O’Handley, S.; Mastromonaco, G.F. Assessment of the reproductive status of female veiled chameleons (Chamaeleo calyptratus) using hormonal, behavioural and physical traits. Zoo Biol. 2015, 34(1), 20–32. [Google Scholar] [CrossRef] [PubMed]
- Amiri, B.S.; Soroori, S.; Rostami, A.; Molazem, M.; Bahonar, A. Ultrasonography and CT examination of ovarian follicular development in Testudo graeca during 1 year in captivity. Vet. Med. Sci. 2023, 9(6), 2606–2616. [Google Scholar] [CrossRef]
- Gumpenberger, M. Diagnostic imaging of reproductive tract disorders in reptiles. Vet. Clin. N. Am. Exot. Anim. Pract. 2017, 20(2), 327–343. [Google Scholar] [CrossRef]
- Alworth, L.C.; Hernandez, S.M.; Divers, S.J. Laboratory reptile surgery: principles and techniques. J. Am. Assoc. Lab. Anim. Sci. 2011, 50(1), 11–26. [Google Scholar]
- Jacobson, E.; Garner, M. Infectious Diseases and Pathology of Reptiles: Color Atlas and Text, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2020. [Google Scholar]
- Innis, C.J.; Boyer, T.H. Chelonian reproductive disorders. Vet. Clin. N. Am. Exot. Anim. Pract. 2002, 5(3), 555–578. [Google Scholar] [CrossRef]
- Oliveri, M.; Sandfoss, M.R.; Reichling, S.B.; Richter, M.M.; Cantrell, J.R.; Knotek, Z.; et al. Ultrasound description of follicular development in the Louisiana pinesnake (Pituophis ruthveni). Animals 2022, 12(21), 2983. [Google Scholar] [CrossRef]
- Funk, R.S. Lizard reproductive medicine and surgery. Vet. Clin. N. Am. Exot. Anim. Pract. 2002, 5(3), 579–613. [Google Scholar] [CrossRef]
- Elliott, D.; Ed, *!!! REPLACE !!!*. Reptile reproductive problems. In Proceedings of the WSAVA World Congress, 2014. [Google Scholar]
- Urbanová, D.; Halán, M. The use of ultrasonography in diagnostic imaging of reptiles. Folia Vet. 2016, 60, 1–10. [Google Scholar] [CrossRef]
- Vetere, A.; Bigliardi, E.; Masi, M.; Rizzi, M.; Leandrin, E.; Di Ianni, F. Egg removal via cloacoscopy in three dystocic leopard geckos (Eublepharis macularius). Animals 2023, 13(5), 924. [Google Scholar] [CrossRef] [PubMed]
- Innis, C.J.; Hernandez-Divers, S.; Martinez-Jimenez, D. Coelioscopic-assisted prefemoral oophorectomy in chelonians. J. Am. Vet. Med. Assoc. 2007, 230(7), 1049–1052. [Google Scholar] [CrossRef] [PubMed]
- Frei, S.; Sanchez-Migallon Guzman, D.; Kass, P.H.; Giuffrida, M.A.; Mayhew, P.D. Evaluation of ventral and left lateral approaches to coelioscopy in bearded dragons (Pogona vitticeps). Am. J. Vet. Res. 2020, 81(3), 267–275. [Google Scholar] [CrossRef]
- Bardi, E.; Antolini, G.; Lubian, E.; Bronzo, V.; Romussi, S. Comparison of lateral and dorsal recumbency during endoscope-assisted oophorectomy in mature pond sliders (Trachemys scripta). Animals 2020, 10(9), 1662. [Google Scholar] [CrossRef]
- Mans, C.; Foster, J.D. Endoscopy-guided ectopic egg removal from the urinary bladder in a leopard tortoise (Stigmochelys pardalis). Can. Vet. J. 2014, 55(6), 569–572. [Google Scholar]
- Di Girolamo, N.; Selleri, P. Clinical applications of cystoscopy in chelonians. Vet. Clin. N. Am. Exot. Anim. Pract. 2015, 18(3), 507–526. [Google Scholar] [CrossRef]
- Campbell, T.W. Clinical pathology of reptiles. In Reptile Medicine and Surgery; Mader, D.R., Ed.; Elsevier: St. Louis, MO, USA, 2006; pp. 453–470. [Google Scholar]
- Cuadrado, M.; Díaz-Paniagua, C.; Quevedo, M.A.; Aguilar, J.M.; Prescott, I.M. Hematology and clinical chemistry in dystocic and healthy post-reproductive female chameleons. J. Wildl. Dis. 2002, 38(2), 395–401. [Google Scholar] [CrossRef]
- Dessauer, H.C. Blood chemistry of reptiles: physiology and evolutionary aspects. In Biology of the Reptilia; Gans, C., Parsons, T.S., Eds.; Academic Press: London, UK, 1970; Volume 3, pp. 1–72. [Google Scholar]
- Dutra, G.H. Diagnostic value of hepatic enzymes, triglycerides and serum proteins for detecting hepatic lipidosis in Chelonoidis carbonaria in captivity. J. Life Sci. 2014, 8, 633–639. [Google Scholar]
- Campbell, T.W. Exotic Animal Hematology and Cytology, 4th ed.; Wiley-Blackwell: Ames, IA, USA, 2015. [Google Scholar]
- Knotek, Z.; Knotková, Z.; Čermáková, E.; Dorrestein, G.M.; Heckers, K.O.; Komenda, D. Plasma bile acids in healthy green iguanas and iguanas with chronic liver diseases. Vet. Med. (Praha) 2023, 68(9), 368–374. [Google Scholar] [CrossRef]
- Paul-M. Gibbons, Ed. Reptile and amphibian urinary tract medicine: diagnosis and therapy. In Proceedings of the Association of Reptilian and Amphibian Veterinarians, 2007.
- Anthonioz, C.; Abadie, Y.; Reversat, E.; Lafargue, A.; Delalande, M.; Renaudineau, T.; et al. Heterogenous within-herd seroprevalence against epizootic hemorrhagic disease virus type 8 (EHDV-8) after massive virus circulation in cattle in France, 2023. Front. Vet. Sci. 2025, 12, 1562883. [Google Scholar] [CrossRef]
- Pacumio, L.P.T.; Tarbert, D.K.; Keel, K.; Ammersbach, M.; Beaufrère, H. Assessment of β-hydroxybutyric acid (BHBA) as a plasma biomarker for hepatic lipidosis in bearded dragons (Pogona vitticeps). UC Davis STAR Program Poster, 2022. [Google Scholar]
- Pacumio, L.; Tarbert, D.; Ammersbach, M.; Beaufrère, H. Method comparison and reference intervals of β-hydroxybutyric acid measurements using a veterinary point-of-care ketone meter and a reference laboratory analyzer in central bearded dragons (Pogona vitticeps). J. Herpetol. Med. Surg. 2025, 35. [Google Scholar] [CrossRef]
- Bennett, A.F. A comparison of activities of metabolic enzymes in lizards and rats. Comp. Biochem. Physiol. B 1972, 42(4), 637–647. [Google Scholar] [CrossRef] [PubMed]
- Roth, L. Comparison of liver cytology and biopsy diagnoses in dogs and cats: 56 cases. Vet. Clin. Pathol. 2001, 30(1), 35–38. [Google Scholar] [CrossRef] [PubMed]
- Palmquist, D.L.; Jenkins, T.C. Fat in lactation rations: review. J. Dairy Sci. 1980, 63(1), 1–14. [Google Scholar] [CrossRef] [PubMed]
- Music, M.K.; Strunk, A. Reptile critical care and common emergencies. Vet. Clin. N. Am. Exot. Anim. Pract. 2016, 19(2), 591–612. [Google Scholar] [CrossRef]
- Tucker, J.K.; Thomas, D.L.; Rose, J. Oxytocin dosage in turtles. Chelonian Conserv. Biol. 2007, 6(2), 321–324. [Google Scholar] [CrossRef]
- Feldman, M.L. Some options to induce oviposition in turtles. Chelonian Conserv. Biol. 2007, 6(2), 313–320. [Google Scholar] [CrossRef]
- Hailey, A.; Loumbourdis, N. Population ecology and conservation of tortoises: demographic aspects of reproduction in Testudo hermanni. Herpetol. J. 1990, 1, 425–434. [Google Scholar]
- Stahl, S.J.; Ed. Managing dystocia in snakes. In ExoticsCon; 2015. [Google Scholar]
- Carpenter, J.W.; Marion, C.J. Exotic Animal Formulary, 5th ed.; Elsevier: St. Louis, MO, USA, 2017. [Google Scholar]
- Govendan, P.N.; Kurniawan, L.; Raharjo, S. Non-invasive treatment in a case of post-ovulatory egg stasis in a Burmese python (Python bivittatus). 2019. pISSN : 2301-7848; eISSN : 2477-6637.
- Millichamp, N.; Lawrence, K.; Jacobson, E.; Jackson, O.; Bell, D. Egg retention in snakes. J. Am. Vet. Med. Assoc. 1984, 183, 1213–1218. [Google Scholar] [CrossRef]
- Guillette, L.J.; Masson, G.R.; DeMarco, V. Effects of prostaglandin F2α, prostaglandin E2, and arachidonic acid on induction of oviposition in oviparous lizards. Prostaglandins 1991, 42(6), 533–540. [Google Scholar] [CrossRef] [PubMed]
- Stacy, N.I.; Alleman, A.R.; Sayler, K.A. Diagnostic hematology of reptiles. Clin. Lab. Med. 2011, 31(1), 87–108. [Google Scholar] [CrossRef] [PubMed]
- Bel, L.; Mihalca, A.; Peștean, C.; Ober, C.; Oana, L. Surgical management of dystocia in snakes and lizards. Bull. Univ. Agric. Sci. Vet. Med. Cluj-Napoca Vet. Med. 2015, 72(1), 205–206. [Google Scholar]
- Patterson, R.W.; Smith, A. Surgical intervention to relieve dystocia in a python. Vet. Rec. 1979, 104(24), 551–552. [Google Scholar] [CrossRef]
- Knotek, Z.; Čermáková, E.; Oliveri, M. Reproductive medicine in lizards. Vet. Clin. N. Am. Exot. Anim. Pract. 2017, 20(2), 411–438. [Google Scholar] [CrossRef]
- Knotek, Z.; Musilová, A.; Knotková, Z.; Trnková, Š.; Babák, V. Alfaxalone anaesthesia in the green iguana (Iguana iguana). Acta Vet. Brno 2013, 82, 109–114. [Google Scholar] [CrossRef]
- Rhim, H.; Godke, A.M.; Aguilar, M.G.; Mitchell, M.A. Evaluating the physiologic effects of alfaxalone, dexmedetomidine, and midazolam in common blue-tongued skinks (Tiliqua scincoides). Animals 2024, 14(18), 2636. [Google Scholar] [CrossRef]
- Helmer, P.; Whiteside, D.; Lewington, J.; Solorza, L.M. Clinical Anatomy and Physiology of Exotic Species: Structure and Function of Mammals, Birds, Reptiles and Amphibians; Elsevier, 2005. [Google Scholar]
- Larouche, C.B.; Johnson, R.; Beaudry, F.; Mosley, C.; Gu, Y.; Zaman, K.A.; et al. Pharmacokinetics of midazolam and its major metabolite 1-hydroxymidazolam in the ball python (Python regius). J. Vet. Pharmacol. Ther. 2019, 42(6), 722–731. [Google Scholar] [CrossRef]
- Hernandez-Divers, S.M.; Schumacher, J.; Stahl, S.; Hernandez-Divers, S.J. Comparison of isoflurane and sevoflurane anesthesia after premedication with butorphanol in the green iguana (Iguana iguana). J. Zoo Wildl. Med. 2005, 36(2), 169–175. [Google Scholar] [CrossRef]
- Mosley, C.A.; Dyson, D.; Smith, D.A. Cardiovascular dose-response effects of isoflurane alone and with butorphanol in the green iguana (Iguana iguana). Vet. Anaesth. Analg. 2004, 31(1), 64–72. [Google Scholar] [CrossRef]
- Greer, L.L.; Jenne, K.J.; Diggs, H.E. Medetomidine–ketamine anesthesia in red-eared slider turtles (Trachemys scripta elegans). Contemp. Top. Lab. Anim. Sci. 2001, 40(3), 9–11. [Google Scholar]
- Johnson-Delaney, C.A.; Ed. Use of nutraceuticals to manage liver disease in bearded dragons (Pogona vitticeps). In Proceedings of the ARAV Conference, 2012. [Google Scholar]
- Lance, V. Reptile reproduction and endocrinology; 2002; pp. 338–358. [Google Scholar]
- Guillette, L.J., Jr.; Gross, T.S.; Masson, G.R.; Matter, J.M.; Percival, H.F.; Woodward, A.R. Developmental abnormalities of the gonad and altered sex hormone concentrations in juvenile alligators from contaminated lakes in Florida. Environ. Health Perspect. 1994, 102(8), 680–688. [Google Scholar] [CrossRef]
- Marlatt, V.L.; Bayen, S.; Castaneda-Cortés, D.; Delbès, G.; Grigorova, P.; Langlois, V.S.; et al. Impacts of endocrine-disrupting chemicals on reproduction in wildlife and humans. Environ. Res. 2022, 208, 112584. [Google Scholar] [CrossRef]
- Moreno, I.; Codoñer, F.M.; Vilella, F.; Valbuena, D.; Martinez-Blanch, J.F.; Jimenez-Almazán, J.; et al. Evidence that the endometrial microbiota has an effect on implantation success or failure. Am. J. Obstet. Gynecol. 2016, 215(6), 684–703. [Google Scholar] [CrossRef]
- Omu, A.E.; Al-Azemi, M.K.; Al-Maghrebi, M.; Mathew, C.T.; Omu, F.E.; Kehinde, E.O.; et al. Molecular basis for the effects of zinc deficiency on spermatogenesis: an experimental study in Sprague–Dawley rats. Indian J. Urol. 2015, 31(1), 57–64. [Google Scholar] [CrossRef]
Figure 1.
Diagnostic imaging of the liver in Hermann’s tortoises (Testudo hermanni) with and without hepatic lipidosis. (A–C) Ultrasonographic images obtained in a water bath in three different individuals. (A) Normal liver with homogeneous, finely granular parenchyma and moderate echogenicity. (B,C) Livers with hepatic lipidosis, showing diffusely increased echogenicity; in (C) the liver is also markedly enlarged with rounded margins. (D–F) CT images acquired in soft-tissue window. (D,E) Transverse images of the same female before the breeding season (D) and during folliculogenesis (E). The liver (L) is initially hyperdense to isodense relative to skeletal muscle and myocardium (h) with hypodense intrahepatic vessels, but becomes mildly to moderately hypodense during follicular development. A hyperdense, normal follicle (f) is visible in the right lateral coelomic cavity in (E), and the stomach (st) is on the left. (F) Dorsal plane reconstruction of a different tortoise with severe fatty liver disease and marked reduction in hepatic attenuation. The hepatic parenchyma contains multiple small fat-attenuating foci, and several small follicles (arrows) show a hypodense peripheral rim, suggestive of atresia.
Figure 1.
Diagnostic imaging of the liver in Hermann’s tortoises (Testudo hermanni) with and without hepatic lipidosis. (A–C) Ultrasonographic images obtained in a water bath in three different individuals. (A) Normal liver with homogeneous, finely granular parenchyma and moderate echogenicity. (B,C) Livers with hepatic lipidosis, showing diffusely increased echogenicity; in (C) the liver is also markedly enlarged with rounded margins. (D–F) CT images acquired in soft-tissue window. (D,E) Transverse images of the same female before the breeding season (D) and during folliculogenesis (E). The liver (L) is initially hyperdense to isodense relative to skeletal muscle and myocardium (h) with hypodense intrahepatic vessels, but becomes mildly to moderately hypodense during follicular development. A hyperdense, normal follicle (f) is visible in the right lateral coelomic cavity in (E), and the stomach (st) is on the left. (F) Dorsal plane reconstruction of a different tortoise with severe fatty liver disease and marked reduction in hepatic attenuation. The hepatic parenchyma contains multiple small fat-attenuating foci, and several small follicles (arrows) show a hypodense peripheral rim, suggestive of atresia.
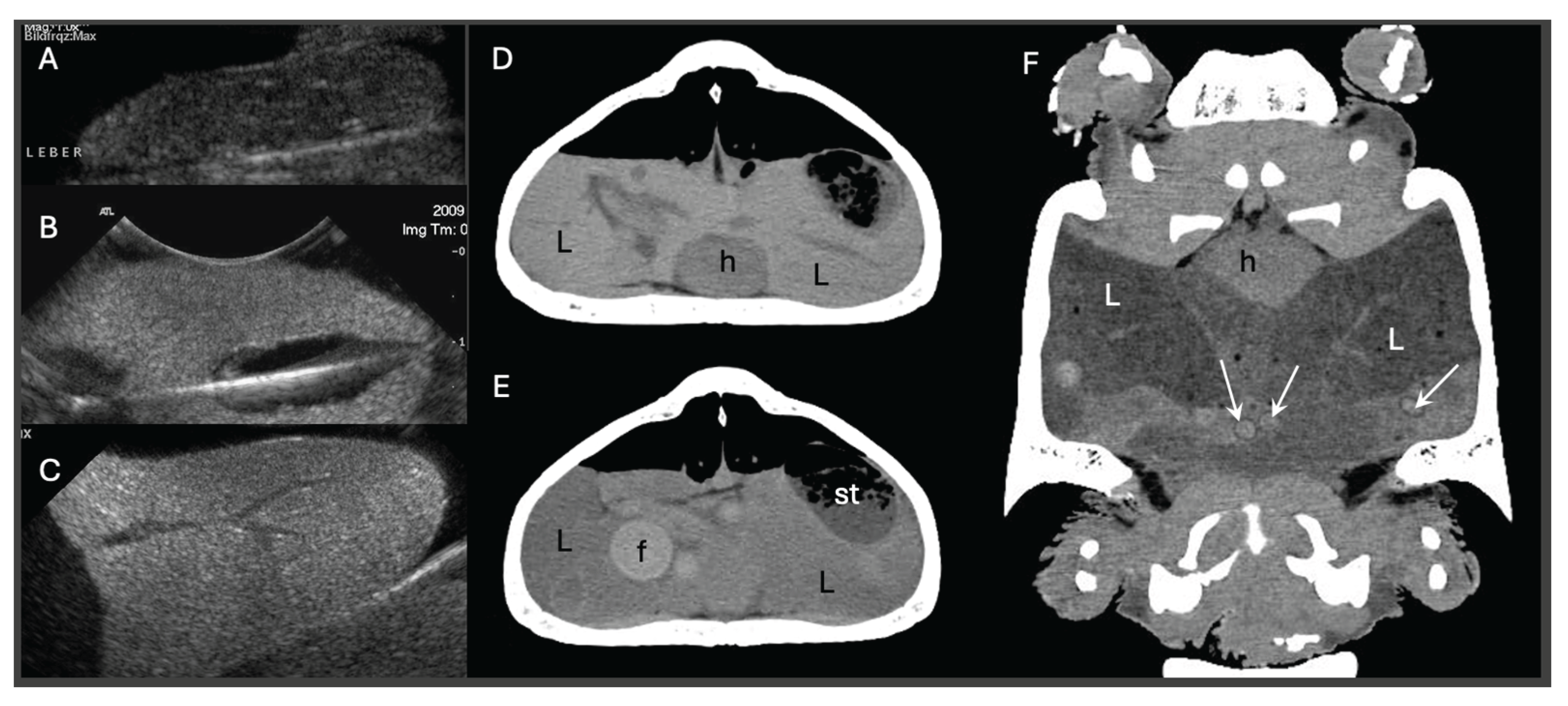
Figure 2.
(A) Dorsoventral radiographic view of five retained eggs in a female Moroccan tortoise (Testudo graeca marokkensis). (B) Oviposition in a captive female Testudo graeca marokensis housed in an indoor enclosure with a prepared sand substrate, after medical management with injections of calcium gluconate, oxytocin and morphine.
Figure 2.
(A) Dorsoventral radiographic view of five retained eggs in a female Moroccan tortoise (Testudo graeca marokkensis). (B) Oviposition in a captive female Testudo graeca marokensis housed in an indoor enclosure with a prepared sand substrate, after medical management with injections of calcium gluconate, oxytocin and morphine.
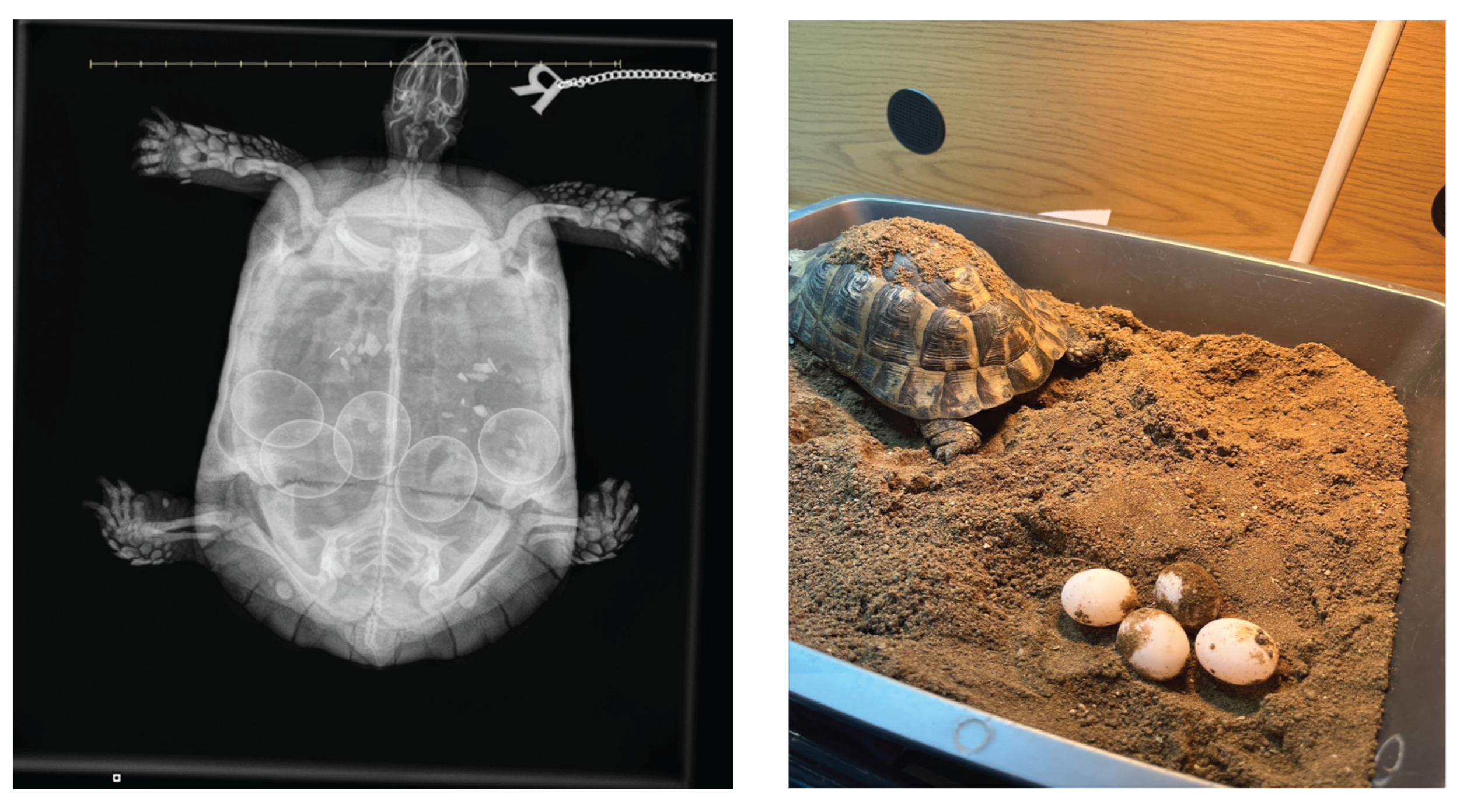
Figure 3.
Diagnostic imaging of reproductive pathology in reptiles. (A) Ultrasonographic image of multiple mildly irregular, heterogeneous follicles with a small amount of surrounding free fluid in a bearded dragon; no interval change was noted over ten days despite clinical deterioration, consistent with follicular stasis. (B) Ultrasonographic evaluation of a gravid garter snake (Thamnophis sirtalis); the skeleton of a fetus (*) is visible, but no cardiac activity was detected, confirming fetal death. (C) Serial ultrasonography of a boa showing progressive degeneration of several eggs over weeks, including loss of internal architecture and non-viable fetal structures. (D) Lateral radiograph of a veiled chameleon (Chamaeleo calyptratus) with severe superovulation; the lungs are markedly compressed and there is profound skeletal demineralisation associated with chronic reproductive disease. (E) Dorsoventral radiograph of a corn snake (Pantherophis guttatus) with an enlarged, obstructed egg at the level of the cloaca (arrows), tightly apposed to a second egg.
Figure 3.
Diagnostic imaging of reproductive pathology in reptiles. (A) Ultrasonographic image of multiple mildly irregular, heterogeneous follicles with a small amount of surrounding free fluid in a bearded dragon; no interval change was noted over ten days despite clinical deterioration, consistent with follicular stasis. (B) Ultrasonographic evaluation of a gravid garter snake (Thamnophis sirtalis); the skeleton of a fetus (*) is visible, but no cardiac activity was detected, confirming fetal death. (C) Serial ultrasonography of a boa showing progressive degeneration of several eggs over weeks, including loss of internal architecture and non-viable fetal structures. (D) Lateral radiograph of a veiled chameleon (Chamaeleo calyptratus) with severe superovulation; the lungs are markedly compressed and there is profound skeletal demineralisation associated with chronic reproductive disease. (E) Dorsoventral radiograph of a corn snake (Pantherophis guttatus) with an enlarged, obstructed egg at the level of the cloaca (arrows), tightly apposed to a second egg.
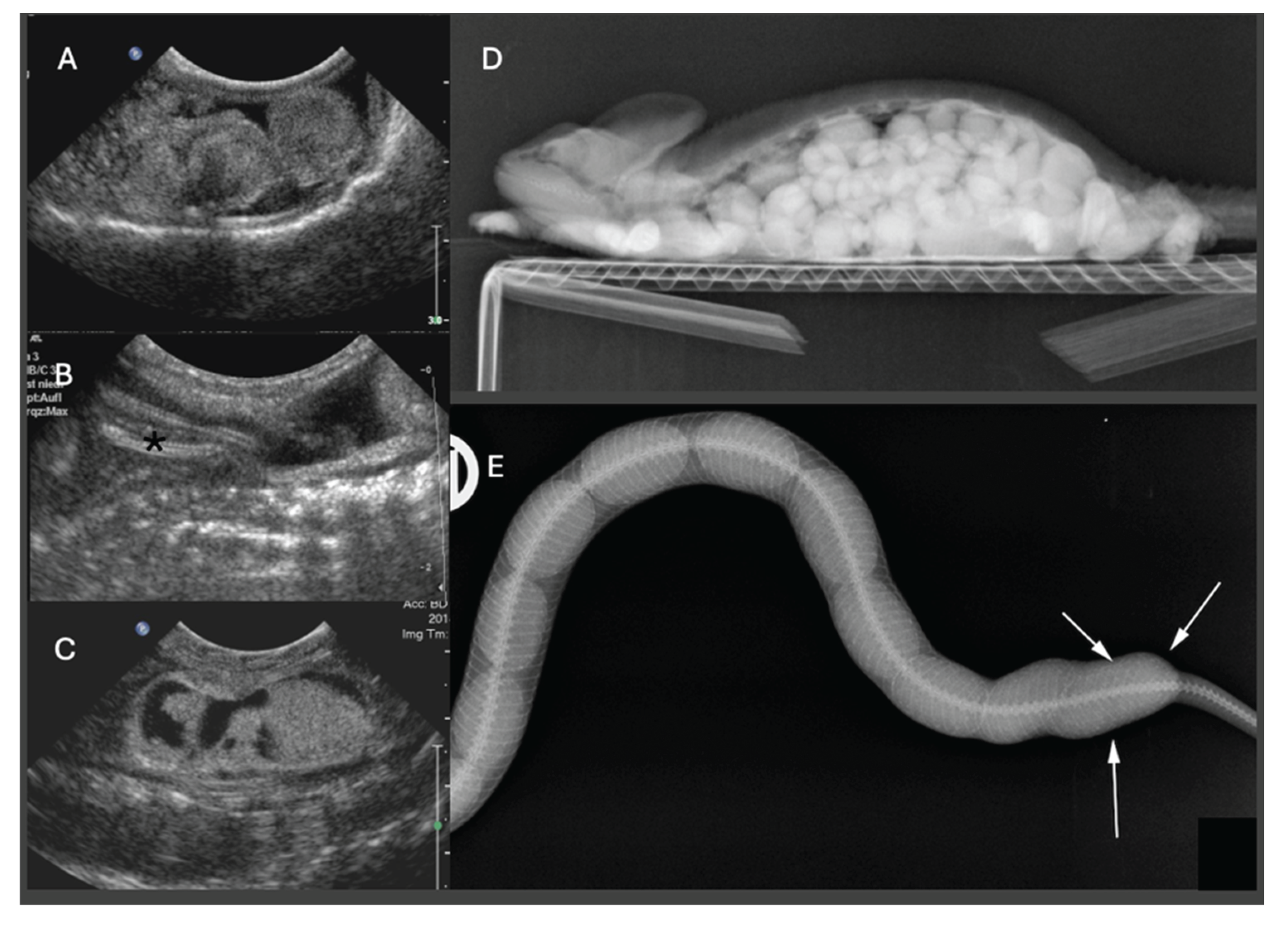
Figure 4.
Diagnostic imaging of normal follicles and eggs in chelonians and squamates. (A–C, E–F, H) Ultrasonographic appearances of normal ovarian follicles and oviductal eggs in Hermann’s tortoises (A–D), a box turtle (G), a python (E), a water dragon (Intellagama lesueurii) (F), and a green iguana (H). (A) Primary follicles with a homogeneous, mildly hyperechoic appearance typical of chelonians. (B) Mature, moderately sized hyperechoic follicles with characteristic distal acoustic shadowing. (C) Heterogeneous echotexture within a small follicle, consistent with early atresia. (D) Ovulated follicle containing yolk with a peripheral hypoechoic rim (albumen); a faint outer hyperechoic line suggests early eggshell mineralization. (E) Primary follicles in squamates (python) are comparatively hypoechoic. (F) Normal elongated oval eggs with soft shells in a water dragon. (G) Dorsoventral radiograph of a box turtle showing a normal clutch of large, uniformly sized eggs without evidence of dystocia. (H) Lateral radiograph of a green iguana displaying multiple ovoid soft-tissue–dense eggs deposited two days earlier.
Figure 4.
Diagnostic imaging of normal follicles and eggs in chelonians and squamates. (A–C, E–F, H) Ultrasonographic appearances of normal ovarian follicles and oviductal eggs in Hermann’s tortoises (A–D), a box turtle (G), a python (E), a water dragon (Intellagama lesueurii) (F), and a green iguana (H). (A) Primary follicles with a homogeneous, mildly hyperechoic appearance typical of chelonians. (B) Mature, moderately sized hyperechoic follicles with characteristic distal acoustic shadowing. (C) Heterogeneous echotexture within a small follicle, consistent with early atresia. (D) Ovulated follicle containing yolk with a peripheral hypoechoic rim (albumen); a faint outer hyperechoic line suggests early eggshell mineralization. (E) Primary follicles in squamates (python) are comparatively hypoechoic. (F) Normal elongated oval eggs with soft shells in a water dragon. (G) Dorsoventral radiograph of a box turtle showing a normal clutch of large, uniformly sized eggs without evidence of dystocia. (H) Lateral radiograph of a green iguana displaying multiple ovoid soft-tissue–dense eggs deposited two days earlier.
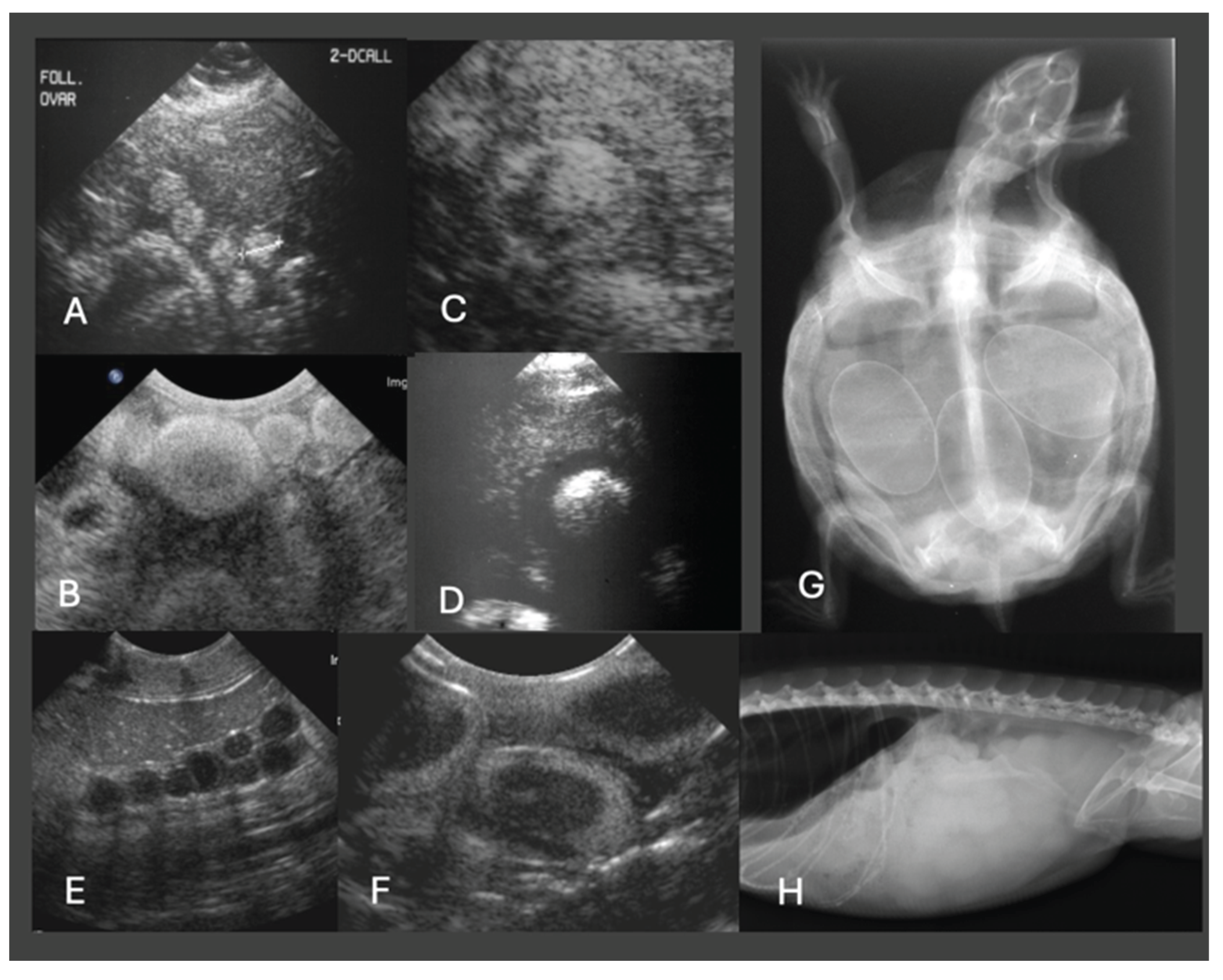
Figure 5.
Diagnostic imaging of dystocia in chelonians. (A) Dorsoventral radiograph of a Hermann’s tortoise with chronic dystocia; one oversized egg is obstructed at the pelvic canal, with marked secondary shell mineralisation. Additional smaller eggs with irregular shells are present. (B) Lateral radiograph of a spider tortoise showing an egg trapped at the cloacal opening due to a caudal shell aperture that is too narrow for passage; the cranial pole of the shell is notably thicker than the caudal pole already positioned near the vent. (C1) Dorsoventral radiograph of a red-eared slider turtle after cloacal administration of contrast medium, demonstrating an egg located within the urinary bladder; (C2) ultrasound confirmed its intravesical position. (D) Dorsoventral radiograph of a Hermann’s tortoise with metabolic bone disease and multiple fractured eggs. (E) Three-dimensional CT model of a Hermann’s tortoise with the carapace and eggs virtually removed; one egg near the right inguinal window (arrow) shows an impression fracture, likely caused by vigorous palpation. (F,G) Transverse and coronal CT images (soft-tissue window) of two Hermann’s tortoises. Several eggs contain gas (*) and exhibit loss of internal architecture (e), consistent with decomposition. Follicles of varying size are visible, some with abnormal contour or a peripheral hypodense rim suggestive of degeneration or stasis. In (G), the liver (L) displays normal hyperdense parenchyma.
Figure 5.
Diagnostic imaging of dystocia in chelonians. (A) Dorsoventral radiograph of a Hermann’s tortoise with chronic dystocia; one oversized egg is obstructed at the pelvic canal, with marked secondary shell mineralisation. Additional smaller eggs with irregular shells are present. (B) Lateral radiograph of a spider tortoise showing an egg trapped at the cloacal opening due to a caudal shell aperture that is too narrow for passage; the cranial pole of the shell is notably thicker than the caudal pole already positioned near the vent. (C1) Dorsoventral radiograph of a red-eared slider turtle after cloacal administration of contrast medium, demonstrating an egg located within the urinary bladder; (C2) ultrasound confirmed its intravesical position. (D) Dorsoventral radiograph of a Hermann’s tortoise with metabolic bone disease and multiple fractured eggs. (E) Three-dimensional CT model of a Hermann’s tortoise with the carapace and eggs virtually removed; one egg near the right inguinal window (arrow) shows an impression fracture, likely caused by vigorous palpation. (F,G) Transverse and coronal CT images (soft-tissue window) of two Hermann’s tortoises. Several eggs contain gas (*) and exhibit loss of internal architecture (e), consistent with decomposition. Follicles of varying size are visible, some with abnormal contour or a peripheral hypodense rim suggestive of degeneration or stasis. In (G), the liver (L) displays normal hyperdense parenchyma.
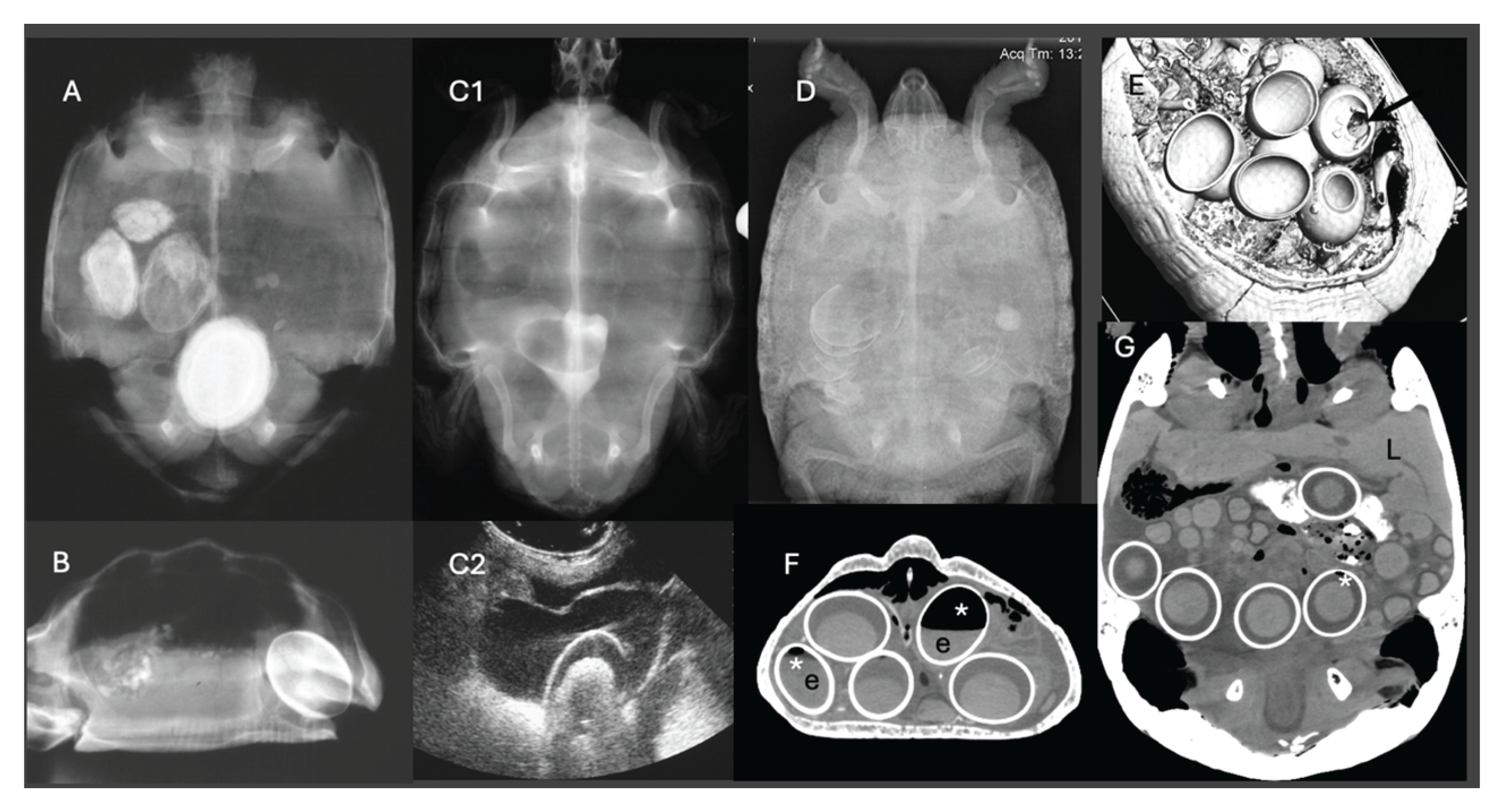
Figure 6.
Clinical decision support flowchart for reptile dystocia.
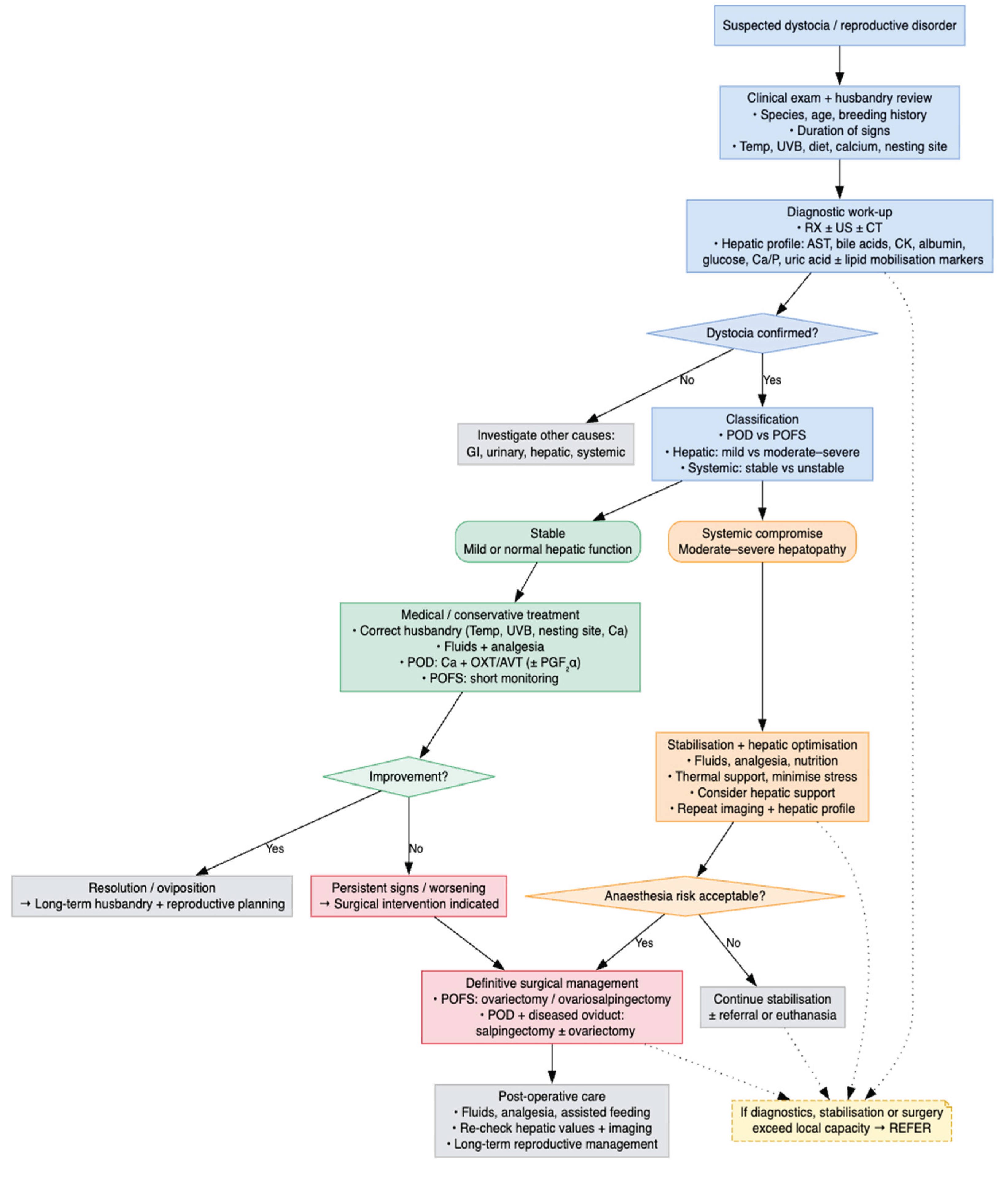

Table 1.
Species-specific most common presentations of dystocia in chelonians, associated risk factors, and relevant literature (UVB = ultraviolet B, POFS = pre-ovulatory follicular stasis, POD = post-ovulatory dystocia).
Table 1.
Species-specific most common presentations of dystocia in chelonians, associated risk factors, and relevant literature (UVB = ultraviolet B, POFS = pre-ovulatory follicular stasis, POD = post-ovulatory dystocia).
| Chelonian species | Common name | Type of dystocia | Key risk factors | References |
|---|---|---|---|---|
| Testudo graeca | Greek tortoise | POD due to pelvic obstruction | Narrow pelvis, lack of nesting site, dehydration | [10,44] |
| Testudo hermanni | Hermann’s tortoise | POFS and coelomitis | Persistent follicular development, inflammation, hypocalcaemia | [44,50] |
| Trachemys scripta elegans | Red-eared slider | Cloacal prolapse and egg impaction | Water quality issues, inadequate UVB radiation, cloacal muscle weakness | [44,45,49] |
| Chelonia mydas | Green sea turtle | Exhaustion-related egg retention during nesting | Long oviposition, dehydration, human disturbance | [44,51] |
Table 2.
Species-specific most common presentations of dystocia in snakes, associated risk factors, and relevant literature (POD = post-ovulatory dystocia).
Table 2.
Species-specific most common presentations of dystocia in snakes, associated risk factors, and relevant literature (POD = post-ovulatory dystocia).
| Snake Species | Common Name | Type of Dystocia / Reproductive Issue | Key Risk Factors | References |
|---|---|---|---|---|
| Python regius | Ball python | Follicular stasis, retained unfertilized ova, coelomitis | Prolonged follicular phase, improper pairing, low temperature | [10,52] |
| Python molurus bivittatus | Burmese python | Obstructive dystocia, egg retention | Obesity, large clutch size, low calcium, oviposition failure | [53] |
| Python sebae | African rock python | Egg-binding with cloacal prolapse | Egg retention, lack of appropriate nesting, dehydration | [54] |
| Lampropeltis getula | Common kingsnake | POD | Inadequate humidity, inappropriate nesting environment | [45] |
Table 3.
Species-specific most common presentations of dystocia in lizards, associated risk factors, and relevant literature (UVB = ultraviolet B, POFS = pre-ovulatory follicular stasis, POD = post-ovulatory dystocia).
Table 3.
Species-specific most common presentations of dystocia in lizards, associated risk factors, and relevant literature (UVB = ultraviolet B, POFS = pre-ovulatory follicular stasis, POD = post-ovulatory dystocia).
| Lizard Species | Common Name | Type of Dystocia / Reproductive Issue | Key Risk Factors | References |
|---|---|---|---|---|
| Eublepharis macularius | Leopard gecko | POD | Lack of UVB radiation, calcium deficiency, | [20,55] |
| Correlophus ciliatus | Crested gecko | POD | Poor husbandry, lack of nesting substrate | [20] |
| Pogona vitticeps | Bearded dragon | POD with large clutch | Hypocalcaemia, dehydration, improper thermal and UVB gradients | [56] |
| Chamaeleo calyptratus | Veiled chameleon | POFS | Calcium deficiency, chronic reproductive cycling, photoperiod issues | [57,58] |
| Varanus albigularis | White-throated monitor | Yolk coelomitis from retained follicles | Failure to oviposit, prolonged retention | [43] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



