Submitted:
11 August 2025
Posted:
12 August 2025
You are already at the latest version
Abstract
Campylobacter hepaticus is the etiologic agent of Spotty Liver Disease (SLD), a newly emerging bacterial disease of laying hens resulting in massive mortality and production losses primarily in free-range systems. Although its economic impact continues to grow, the molecular basis of C. hepaticus pathogenesis is poorly understood. In this study, we conducted transcriptomic profiling of C. hepaticus in three host-relevant conditions, exposure to chicken bile, infection of a chicken liver hepatocellular carcinoma (LMH) cell line, and isolation from liver lesions of naturally infected chickens. Through RNA-seq analysis, we found unique gene expression signatures in each environment. In bile, C. hepaticus exhibited differential expression of 412 genes, with upregulation of genes related to motility, cell envelope remodeling, glycosylation, nitrate respiration, and multidrug efflux systems, indicating a stress-adaptive, metabolically active lifestyle. In LMH cells, on the other hand, 125 genes were differentially expressed, primarily reflecting downregulation of motility, oxidative stress response, chaperones, and core metabolic processes, which suggests of a low-activity, intracellular dormant lifestyle. Transcriptomic analysis of C. hepaticus isolated from the liver identified 26 differentially expressed genes, featuring selective upregulation of nitrate respiration, sulfur metabolism, and pyridoxal 5’ phosphate (PLP) homeostasis, along with down regulation the major outer membrane porin (mompA), stress response chaperones (dnaK, groL), and genes involved in oxidative stress defense and energy production-. Further, immune evasion-related gene cmeA and a glycosyltransferase were found to be highly upregulated. This work presents the first in-depth transcriptomic exploration of C. hepaticus in multiple host relevant niches. Our findings reveal niche-specific gene expression profiles and highlight metabolic and structural accommodations allowing C. hepaticus survival during bile exposure, persistence within host cells, and induction of liver pathology. These insights provide a basis for the identification of novel virulence determinants and may inform the development of targeted interventions, including vaccines or antimicrobial therapy, to control SLD in commercial poultry operations.
Keywords:
1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Culture Conditions
2.2. In Vitro Exposure of C. hepaticus to Chicken Bile
2.3. Chicken Liver Hepatocellular Carcinoma Epithelial Cells (LMH) and Infection
2.4. Liver Sample Collection from Spotty Liver Disease (SLD) Infected Chicken
2.5. RNA Extraction, rRNA Depletion, and mRNA Enrichment
2.6. Transcriptome Library Preparation, Sequencing, and Differential Gene Expression Analysis
2.7. Data Availability
3. Results
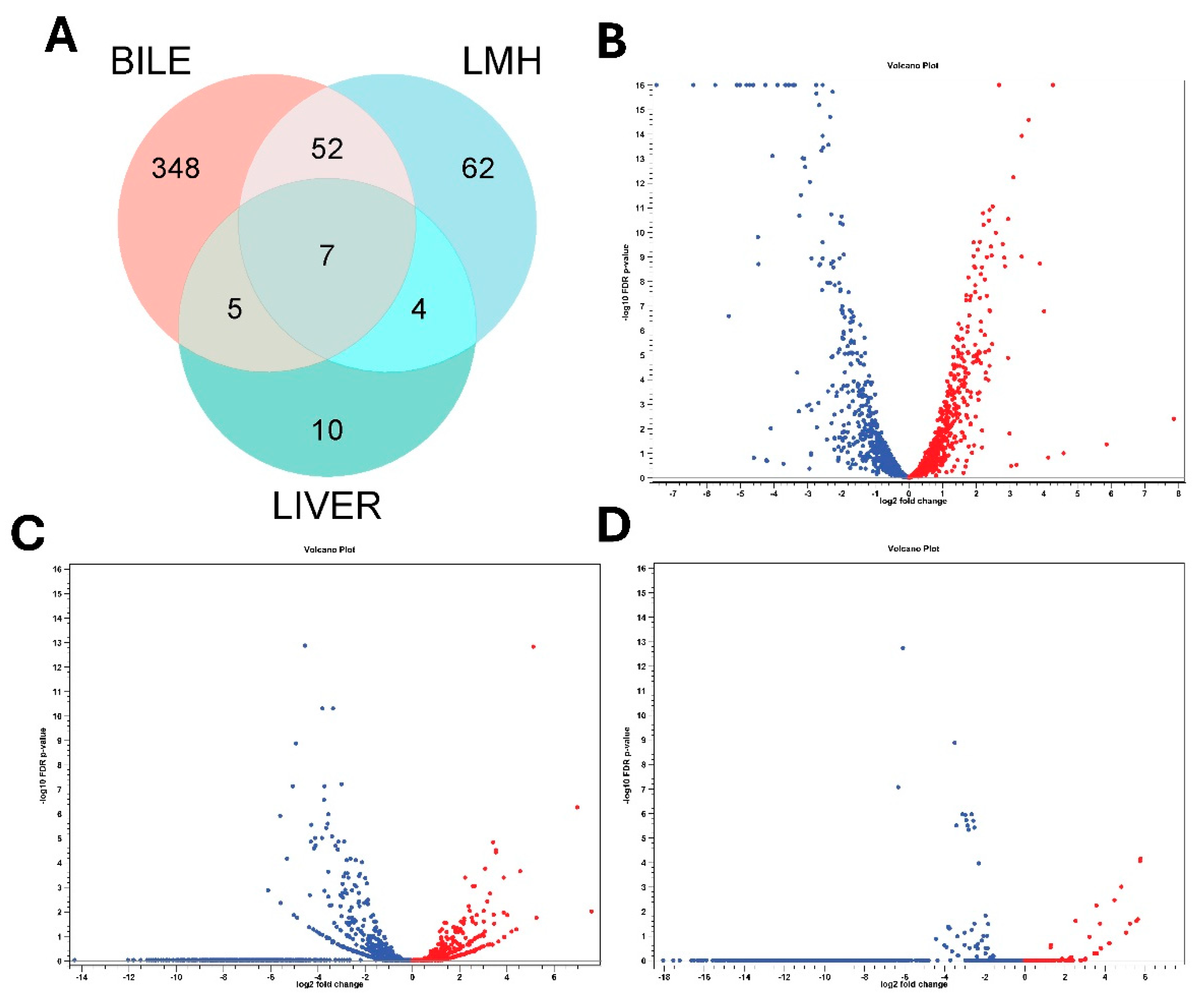
3.1. Overall Transcriptomic Response of C. hepaticus Across Different Host-Associated Environments
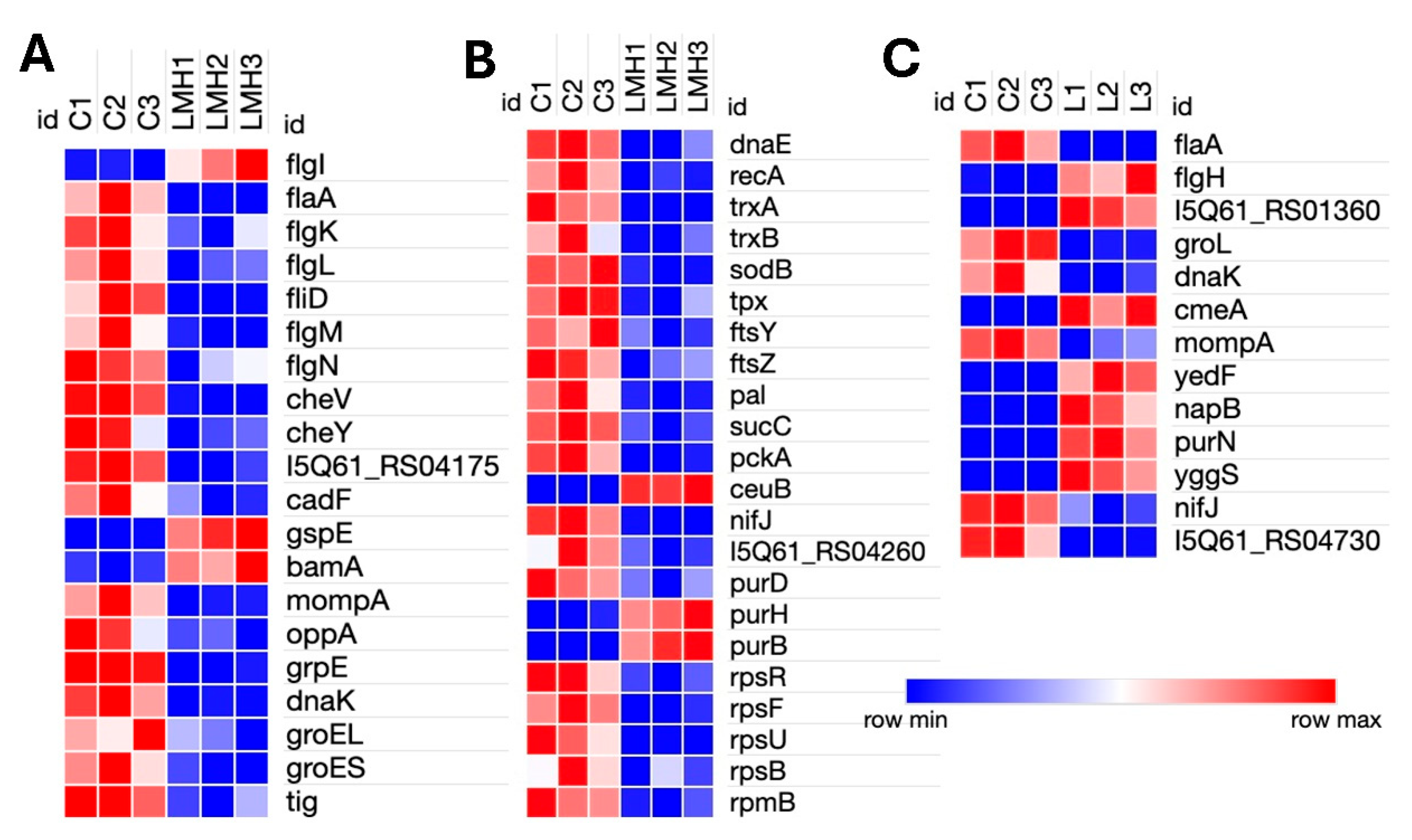
3.2. Transcriptomic Response of C. hepaticus to Bile Exposure
3.3. Transcriptomic Adaptation of C. hepaticus During In Vitro LMH Cell Infection
3.4. Transcriptomic Analysis of C. hepaticus Isolated from Infected Livers
4. Discussion
4.1. Adaptation to Bile Exposure
4.2. Adaptation Within LMH Cells
4.3. In Vivo Adaptation in the Liver
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| SLD | Spotty liver disease |
| LMH | Chicken liver hepatocellular carcinoma epithelial cells |
| FBS | Fetal bovine serum |
| FDR | false discovery rate |
| DEG | Differentially expressed gene |
| RIN | RNA integrity number |
| FC | Fold change |
| LOS | Lipooligosaccharide |
| VBNC | Viable but non-culturable |
References
- Courtice, J.M.; Mahdi, L.K.; Groves, P.J.; Kotiw, M. Spotty Liver Disease: A Review of an Ongoing Challenge in Commercial Free-Range Egg Production. Vet Microbiol 2018, 227, 112–118. [Google Scholar] [CrossRef]
- Wade, B.; Rautenschlein, S.; Hao Van, T.T.; Moore, R.J. Campylobacter Hepaticus and Spotty Liver Disease in Poultry. Avian Dis 2025, 68, 481–489. [Google Scholar] [CrossRef]
- Ienes-Lima, J.; Becerra, R.; Logue, C.M. Comparative Genomic Analysis of Campylobacter Hepaticus Genomes Associated with Spotty Liver Disease, Georgia, United States. Front Microbiol 2023, 14. [Google Scholar] [CrossRef]
- Groves, P.J.; Gao, Y.K.; Kotiw, M.; Wong, P.H.L.; Muir, W.I. Epidemiological Risk Factors and Path Models for Spotty Liver Disease in Australian Cage-Free Flocks Incorporating a Scratch Area. Poult Sci 2025, 104. [Google Scholar] [CrossRef]
- Courtice, J.M.; Ahmad, T.B.; Wei, C.; Mahdi, L.K.; Palmieri, C.; Juma, S.; Groves, P.J.; Hancock, K.; Korolik, V.; Petrovsky, N.; et al. Detection, Characterization, and Persistence of Campylobacter Hepaticus, the Cause of Spotty Liver Disease in Layer Hens. Poult Sci 2023, 102. [Google Scholar] [CrossRef]
- Crawshaw, T.R.; Chanter, J.I.; Young, S.C.L.; Cawthraw, S.; Whatmore, A.M.; Koylass, M.S.; Vidal, A.B.; Salguero, F.J.; Irvine, R.M. Isolation of a Novel Thermophilic Campylobacter from Cases of Spotty Liver Disease in Laying Hens and Experimental Reproduction of Infection and Microscopic Pathology. Vet Microbiol 2015, 179, 315–321. [Google Scholar] [CrossRef]
- Van, T.T.H.; Elshagmani, E.; Gor, M.C.; Scott, P.C.; Moore, R.J. Campylobacter Hepaticus Sp. Nov., Isolated from Chickens with Spotty Liver Disease. Int J Syst Evol Microbiol 2016, 66, 4518–4524. [Google Scholar] [CrossRef] [PubMed]
- Van, T.T.H.; Elshagmani, E.; Gor, M.C.; Anwar, A.; Scott, P.C.; Moore, R.J. Induction of Spotty Liver Disease in Layer Hens by Infection with Campylobacter Hepaticus. Vet Microbiol 2017, 199, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Petrovska, L.; Tang, Y.; van Rensburg, M.J.J.; Cawthraw, S.; Nunez, J.; Sheppard, S.K.; Ellis, R.J.; Whatmore, A.M.; Crawshaw, T.R.; Irvine, R.M. Genome Reduction for Niche Association in Campylobacter Hepaticus, a Cause of Spotty Liver Disease in Poultry. Front Cell Infect Microbiol 2017, 7. [Google Scholar] [CrossRef]
- Gregory, M.; Klein, B.; Sahin, O.; Girgis, G. Isolation and Characterization of Campylobacter Hepaticus from Layer Chickens with Spotty Liver Disease in the United States. Avian Dis 2018, 62, 79–85. [Google Scholar] [CrossRef]
- Phung, C.; Wilson, T.B.; Quinteros, J.A.; Scott, P.C.; Moore, R.J.; Van, T.T.H. Enhancement of Campylobacter Hepaticus Culturing to Facilitate Downstream Applications. Sci Rep 2021, 11. [Google Scholar] [CrossRef]
- Hao Van, T.T.; Lacey, J.A.; Vezina, B.; Phung, C.; Anwar, A.; Scott, P.C.; Moore, R.J. Survival Mechanisms of Campylobacter Hepaticus Identified by Genomic Analysis and Comparative Transcriptomic Analysis of in Vivo and in Vitro Derived Bacteria. Front Microbiol 2019, 10. [Google Scholar] [CrossRef]
- Jiarpinitnun, P.; Iwakiri, A.; Fuke, N.; Pongsawat, P.; Miyanishi, C.; Sasaki, S.; Taniguchi, T.; Matsui, Y.; Luangtongkum, T.; Yamada, K.; et al. Involvement of Campylobacter Species in Spotty Liver Disease-like Lesions in Broiler Chickens Detected at Meat Inspections in Miyazaki Prefecture, Japan. Microorganisms 2024, 12. [Google Scholar] [CrossRef]
- Gao, Y.K.; Singh, M.; Muir, W.I.; Kotiw, M.; Groves, P.J. Identification of Epidemiological Risk Factors for Spotty Liver Disease in Cage-Free Layer Flocks in Houses with Fully Slatted Flooring in Australia. Poult Sci 2023, 102. [Google Scholar] [CrossRef]
- Gottapu, C.; Sahin, O.; Edison, L.K.; Srednik, M.E.; Kariyawasam, S. Complete Genome Sequences of Campylobacter Hepaticus Strains USA1 and USA5 Isolated from a Commercial Layer Flock in the United States. Microbiol Resour Announc 2025, 14. [Google Scholar] [CrossRef] [PubMed]
- Arukha, A.; Denagamage, T.N.; Butcher, G.; Kariyawasam, S. Complete Genome Sequence of Campylobacter Hepaticus Strain UF2019SK1, Isolated from a Commercial Layer Flock in the United States. 2021. [Google Scholar] [CrossRef]
- Van, T.T.H.; Elshagmani, E.; Gor, M.C.; Anwar, A.; Scott, P.C.; Moore, R.J. Induction of Spotty Liver Disease in Layer Hens by Infection with Campylobacter Hepaticus. Vet Microbiol 2017, 199, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Edison, L.K.; Kudva, I.T.; Kariyawasam, S. Comparative Transcriptome Analysis of Shiga Toxin-Producing Escherichia Coli O157:H7 on Bovine Rectoanal Junction Cells and Human Colonic Epithelial Cells during Initial Adherence. Microorganisms 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Edison, L.K.; Kariyawasam, S. From the Gut to the Brain: Transcriptomic Insights into Neonatal Meningitis Escherichia Coli Across Diverse Host Niches. Pathogens 2025, 14. [Google Scholar] [CrossRef]
- Begley, M.; Gahan, C.G.M.; Hill, C. The Interaction between Bacteria and Bile. FEMS Microbiol Rev 2005, 29, 625–651. [Google Scholar] [CrossRef]
- Cremers, C.M.; Knoefler, D.; Vitvitsky, V.; Banerjee, R.; Jakob, U. Bile Salts Act as Effective Protein-Unfolding Agents and Instigators of Disulfide Stress in Vivo. Proc Natl Acad Sci U S A 2014, 111. [Google Scholar] [CrossRef]
- Negretti, N.M.; Gourley, C.R.; Clair, G.; Adkins, J.N.; Konkel, M.E. The Food-Borne Pathogen Campylobacter Jejuni Responds to the Bile Salt Deoxycholate with Countermeasures to Reactive Oxygen Species. Sci Rep 2017, 7. [Google Scholar] [CrossRef]
- Benoit, S.L.; Maier, R.J.; Sawers, R.G.; Greening, C. Molecular Hydrogen Metabolism: A Widespread Trait of Pathogenic Bacteria and Protists. 2020. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, J.; Jiang, Q.; Huang, N.; Ding, X.; Peng, L.; Deng, X. The Molybdate-Binding Protein ModA Is Required for Proteus Mirabilis-Induced UTI. Front Microbiol 2023, 14. [Google Scholar] [CrossRef]
- Chen, E.C.; Freel Meyers, C.L. DXP Synthase Function in a Bacterial Metabolic Adaptation and Implications for Antibacterial Strategies. Antibiotics 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Sakur, S.G.J.; Williamson, S.L.; Pavic, A.; Gao, Y.K.; Harris, T.; Kotiw, M.; Muir, W.I.; Groves, P.J. Developing a Selective Culturing Approach for Campylobacter Hepaticus. PLoS One 2024, 19. [Google Scholar] [CrossRef]
- Hajam, I.A.; Dar, P.A.; Shahnawaz, I.; Jaume, J.C.; Lee, J.H. Bacterial Flagellin-a Potent Immunomodulatory Agent. Exp Mol Med 2017, 49. [Google Scholar] [CrossRef]
- Elmi, A.; Nasher, F.; Dorrell, N.; Wren, B.; Gundogdu, O. Revisiting Campylobacter Jejuni Virulence and Fitness Factors: Role in Sensing, Adapting, and Competing. Front Cell Infect Microbiol 2021, 10. [Google Scholar] [CrossRef]
- Friman, V.P.; Lindstedt, C.; Hiltunen, T.; Laakso, J.; Mappes, J. Predation on Multiple Trophic Levels Shapes the Evolution of Pathogen Virulence. PLoS One 2009, 4. [Google Scholar] [CrossRef]
- Cox, C.A.; Bogacz, M.; El Abbar, F.M.; Browning, D.D.; Hsueh, B.Y.; Waters, C.M.; Lee, V.T.; Thompson, S.A. The Campylobacter Jejuni Response Regulator and Cyclic-Di-GMP Binding CbrR Is a Novel Regulator of Flagellar Motility. Microorganisms 2022, 10. [Google Scholar] [CrossRef]
- Korotkov, K. V.; Sandkvist, M.; Hol, W.G.J. The Type II Secretion System: Biogenesis, Molecular Architecture and Mechanism. Nat Rev Microbiol 2012, 10, 336–351. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Gao, B.; Novik, V.; Galán, J.E. Quantitative Proteomics of Intracellular Campylobacter Jejuni Reveals Metabolic Reprogramming. PLoS Pathog 2012, 8. [Google Scholar] [CrossRef] [PubMed]
- Allameh, A.; Niayesh-Mehr, R.; Aliarab, A.; Sebastiani, G.; Pantopoulos, K. Oxidative Stress in Liver Pathophysiology and Disease. Antioxidants 2023, 12. [Google Scholar] [CrossRef]
- Randaisi, V.R.; Bunch, M.L.; Beavers, W.N.; Rogers, T.; Mesler, R.; Ashurst, T.D.; Donohoe, D.R.; Monteith, A.J.; Johnson, J.G. Efficient Gastrointestinal Colonization by Campylobacter Jejuni Requires Components of the ChuABCD Heme Transport System 2025.
- Miller, C.E.; Williams, P.H.; Ketley, J.M. Pumping Iron: Mechanisms for Iron Uptake by Campylobacter. Microbiology (N Y) 2009, 155, 3157–3165. [Google Scholar] [CrossRef]
- Urdaneta, V.; Casadesús, J. Interactions between Bacteria and Bile Salts in the Gastrointestinal and Hepatobiliary Tracts. Front Med (Lausanne) 2017, 4. [Google Scholar] [CrossRef]
- Bloomfield, S.J.; Midwinter, A.C.; Biggs, P.J.; French, N.P.; Marshall, J.C.; Hayman, D.T.S.; Carter, P.E.; Mather, A.E.; Fayaz, A.; Thornley, C.; et al. Genomic Adaptations of Campylobacter Jejuni to Long-Term Human Colonization. Gut Pathog 2021, 13. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Makino, F.; Miyata, T.; Minamino, T.; Kato, T.; Namba, K. Structure of the Molecular Bushing of the Bacterial Flagellar Motor. Nat Commun 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Soutourina, O.A.; Bertin, P.N. Regulation Cascade of Flagellar Expression in Gram-Negative Bacteria. FEMS Microbiol Rev 2003, 27, 505–523. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.; Schmitt-Kopplin, P. The Role of Fecal Sulfur Metabolome in Inflammatory Bowel Diseases. International Journal of Medical Microbiology 2021, 311. [Google Scholar] [CrossRef]
- He, S.; Chen, Y.; Wang, L.; Bai, X.; Bu, T.; Zhang, J.; Lu, M.; Ha, N.C.; Quan, C.; Nam, K.H.; et al. Structural and Functional Analysis of the Pyridoxal Phosphate Homeostasis Protein YggS from Fusobacterium Nucleatum. Molecules 2022, 27. [Google Scholar] [CrossRef]
- Lin, J.; Cagliero, C.; Guo, B.; Barton, Y.W.; Maurel, M.C.; Payot, S.; Zhang, Q. Bile Salts Modulate Expression of the CmeABC Multidrug Efflux Pump in Campylobacter Jejuni. J Bacteriol 2005, 187, 7417–7424. [Google Scholar] [CrossRef] [PubMed]
- Dzieciol, M.; Wagner, M.; Hein, I. CmeR-Dependent Gene Cj0561c Is Induced More Effectively by Bile Salts than the CmeABC Efflux Pump in Both Human and Poultry Campylobacter Jejuni Strains. Res Microbiol 2011, 162, 991–998. [Google Scholar] [CrossRef]
- Fox, E.M.; Raftery, M.; Goodchild, A.; Mendz, G.L. Campylobacter Jejuni Response to Ox-Bile Stress. FEMS Immunol Med Microbiol 2007, 49, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Day, C.J.; Semchenko, E.A.; Korolik, V. Glycoconjugates Play a Key Role in Campylobacter Jejuni Infection: Interactions between Host and Pathogen. Front Cell Infect Microbiol 2012, 2, 9. [Google Scholar] [CrossRef] [PubMed]
- Henderson, B.; Allan, E.; Coates, A.R.M. Stress Wars: The Direct Role of Host and Bacterial Molecular Chaperones in Bacterial Infection. Infect Immun 2006, 74, 3693–3706. [Google Scholar] [CrossRef]
- Figaj, D. The Role of Heat Shock Protein (Hsp) Chaperones in Environmental Stress Adaptation and Virulence of Plant Pathogenic Bacteria. Int J Mol Sci 2025, 26. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



