Submitted:
22 June 2025
Posted:
24 June 2025
You are already at the latest version
Abstract
A mathematical model incorporating intermittent preventive treatment of malaria using sulfadoxine-pyrimethamine in pregnant women (IPTp-SP) as an intervention strategy is developed and analyzed. The model incorporates both the human host and mosquito vector populations. The stability properties of the equilibrium points are analytically assessed, and numerical simulations are carried out to support and illustrate the theoretical results. The basic reproduction number, $R_0$, is shown to fall below unity under the intervention, suggesting the potential for malaria elimination within the pregnant population. A comparative analysis reveals that the intervention significantly enhances disease suppression, supporting its role as an effective strategy for reducing malaria burden among pregnant women.

Keywords:
IPTp-SP
; malaria
; pregnant women
; Reproduction number
1. Introduction
Malaria is a significant public health concern in many regions across the globe, particularly in tropical and sub-tropical regions where Anopheles mosquitoes are prevalent [1]. In 2023, the global estimate of malaria cases reached approximately 263 million, with an incidence rate of 60.4 cases per 1,000 individuals at risk [2]. Infants, children under the age of 5, pregnant women and girls, travellers, and people living with HIV or AIDS are at higher risk of severe infection [1]. The disease is caused by Plasmodium parasites (P. falciparum, P. vivax, P. malariae, P. ovale and P. knowlesi) transmitted to humans by female Anopheles mosquitoes [3]. The most deadly parasite among Plasmodium species is P. falciparum, which is most prevalent on the African continent [1,4]. Not all female Anopheles mosquitoes have malaria [3], but they become infectious when they bite humans carrying the parasite during a blood meal. They then transmit the parasite to other humans, continuing the malaria transmission cycle. The life cycle of malaria is depicted in Figure 1.
Despite efforts to reduce the incidence of malaria, the disease remains a major public health concern [5]. The interventions used for malaria prevention for all groups include vector control, vaccination, and preventive chemotherapies. For pregnant women, intermittent preventive treatment with sulfadoxine-pyrimethamine (IPTp-SP) is administered exclusively as a prophylactic measure during pregnancy [2,5]. When prevention fails, infected individuals are prescribed treatment based on the severity of their condition and their general health status to treatments such as artesunate and artemisinin-based combination therapy [2].
Protecting pregnant women from malarial infections has been a significant challenge in Malawi [5]. Therefore, investigating the effectiveness of sulphadoxine-pyrimethamine as an intermittent preventative treatment for malaria in pregnant women in Malawi is a plausible approach to addressing this issue. According to [5], while everyone is at risk of contracting malaria, pregnant women face a particularly high risk of its adverse consequences due to reduced immunity [6,7]. Their increased vulnerability is attributed to the immunological and physiological changes that occur during pregnancy, which can increase malaria severity and lead to complications such as maternal anaemia, preterm birth, and low birth weight [1,3,6,7]. Pregnant women are three times more likely to develop severe malaria compared to non-pregnant women in the same geographic area [4,8]. In Malawi, IPTp-SP remains a trusted intervention for preventing malaria during pregnancy.
Some studies have suggested that IPTp-SP doses effectively suppress the adverse effects of malaria during pregnancy, and their effectiveness appears to vary depending on the number of doses taken by pregnant women [9,10]. This remains true even in the presence of widespread resistance of Plasmodium species to sulphadoxine-pyrimethamine (SP) [5,11]. Three or more doses of IPTp-SP are the most effective in averting the adverse effects of malaria in pregnant women [10,11,12].
Compartmental mathematical models of malaria are widely documented in the literature [13]. The pioneering mathematical models of malaria that describe vector-host dynamics, upon which other models of malaria are based, include the Ross-Macdonald models, and the Anderson and May model [13,14,15,16]. [17] and [18] adapted these frameworks to variable population size models while trying to describe the disease dynamics of malaria. However, their models did not incorporate any interventions, but provided a framework for studying control strategies for the containment of malaria. [17] developed an SEIRS model that incorporates reinfection due to the loss of immunity in the recovered population. Through numerical simulations, they confirmed that abrupt reductions in the mosquito population may temporarily decrease malaria prevalence but do not lead to elimination in endemic regions. Using the same SEIRS framework, [18] emphasized that lowering below 1 may not always guarantee malaria eradication due to possible subcritical bifurcations and concluded that targeted interventions must be carefully structured to achieve effective malaria control. [19] suggested that combining effective antimalarial drugs with comprehensive control measures is essential for reducing transmission and the pool of infectious individuals. [20] developed a mathematical model that analyzed the dynamics of malaria disease transmission and evaluated control strategies for its management. [21] emphasized the importance of applying mathematical models to improve public health strategies against malaria. [22] considered a combination of intervention strategies such as effective mass drug administration and vector control (LLITNs and IRS) to combat and eventually eliminate the malaria. Although various transmission and control compartmental mathematical models of malaria are widely documented in the literature, our proposed model is seemingly new as it investigates the effectiveness of IPTp-SP as a prophylactic treatment for pregnant women.
The proposed compartmental model is not exhaustive. One limitation is that it does not specify the number of doses pregnant women are receiving. The recommended number of IPTp-SP doses during pregnancy is at least three, with the first dose administered as early as possible at the beginning of the second trimester, but not before week 13 of pregnancy [2].
In the following sections, we formulate and analyze a deterministic model incorporating IPTp-SP as an intervention strategy. Key parameters influencing transmission are identified through sensitivity analysis of the model. Finally, some parameter values are assumed within realistic ranges to support the analytical results, with the caveat that model outcomes are not compared with real data.
2. Methods: Model Formulation and Description
The population of pregnant women is divided based on malaria and dosing status. When malaria invades, it categorizes them into non-intersecting classes: susceptible (), exposed (), infected (), and recovered (), with representing susceptible pregnant women who receive any number of IPTp-SP doses (one, two, or at least three). Similarly, vectors are classified based on their malaria status. When malaria spreads among vectors, it divides them into non-intersecting classes: susceptible (), exposed (), and infected (). The total population of pregnant women at any time is given by:
And the total population of vectors at any given time is given by:
The recruitment rate of pregnant women through conception is , while represents the recruitment rate of the mosquitoes through birth. The natural death rates of pregnant women and mosquito populations are and , respectively, with these rates being proportional to the number of individuals or mosquitoes in each class. The biting rate of mosquitoes is . The fraction of susceptible pregnant women taking IPTp-SP prophylaxis is denoted as , while the fraction not taking IPTp-SP prophylaxis is given by . The fraction of pregnant women protected from malaria due to IPTp-SP effectiveness, , transitions directly to the recovered class at the treatment success rate . Conversely, represents the fraction of pregnant women who contract malaria at a treatment failure rate . Male (microgametocytes) and female (macrogametocytes) gametocytes are ingested by Anopheles mosquitoes during a blood meal from an infected pregnant woman at the rate:
where is the success rate of gametocyte transmission from an infected pregnant woman to susceptible mosquitoes. Thereafter, susceptible mosquitoes enter the exposed class . During this stage, gametocytes fuse, generating zygotes that later develop into oocytes, which eventually rupture to release sporozoites. At this point, mosquitoes transition into the infected class at the rate . Mosquitoes may die from the disease at the rate in the infected class. During a subsequent blood meal, malaria-infected female Anopheles mosquitoes inoculate sporozoites into pregnant women at the rates:
and
where and represent the success rates of sporozoite transmission from infected mosquitoes to susceptible pregnant women taking IPTp-SP prophylaxis and those not taking IPTp-SP prophylaxis , respectively, during a blood meal. This transitions pregnant women into the latent stage, . Upon infection, sporozoites invade liver cells, where they mature and multiply before rupturing and releasing merozoites. These merozoites then infect red blood cells, continuing a cycle of replication. The blood-stage parasites cause the disease’s symptoms and clinical manifestations, transitioning pregnant women into the infected class at the rate . Infected pregnant women may die from the disease at the rate or move to the recovered class, , at the rate . Once immunity wanes, they return to the susceptible class, , at the rate . Figure 2 provides a graphical interpretation of the malaria compartmental model
Based on our model description, assumptions, definitions of the state variables and parameters in Table 1, the proposed malaria model satisfies the following system of non-linear ordinary differential equations:
where:
and
3. Results
3.1. Invariant Region
Both the model state variables and parameters are assumed non-negative for all time . Let be any solution of the system with non-negative initial conditions. Applying Birkhoff and Rota’s Theorem [23] on differential inequality, from Equation (1), we have as , and thus, . Hence the feasible solutions on the human population enter the region
Similarly, it can be shown that the feasible solutions on the mosquito population given by Equation (2) enter the region
Therefore, from (4) and (5), the possible solutions of model (3) will enter the the positively invariant region .
3.2. Positivity of the State Variables
Since is a positively invariant set under the flow induced by model (3), we now show that every solution with initial condition in remains in that region for .
Theorem 1.
The solution set of the malaria model (3) with the initial condition is positive for all .
3.3. Existence and Stability of Steady-State Solutions
The disease-free equilibrium (DFE) of the malaria model (3) denoted by is given by
At equilibrium we consider the compartments , and of the system (3) which as non-infected compartments. Therefore from equations 1,2, and 6 of the model (3) we have
From (6) is given as:
where
Substituting in (7), we have:
where
Therefore:
Equation (9) is the general form for finding the disease-free equilibrium points for the IPTp-SP intervention model.
From (8) we can solve for to have:
Consequently, the disease-free equilibrium for the model (3) will be given by:
The effective reproduction number is obtained by using the next generation matrix [24]. Let
and
The effective reproduction number is the spectral radius and the resulting expression is given by
The effective reproduction number is defined as the number of secondary malaria infections caused by one infectious pregnant woman or mosquito during the infectious period in a completely susceptible population. This number is not only important for describing how quickly the disease could spread, but it can also provide valuable information for controlling and preventing the spread of the disease [22].
3.4. Local Stability of the Disease-Free Equilibrium
Corollary 1.
Corollary of Gershgorin Circle Theorem: Let A be an n × n matrix with real entries. If the diagonal elements of A satisfy
where
for i = 1...n, then the eigenvalues of A are negative or have negative real parts.
Lemma 1.
The DFE for the malaria model (3) is locally asymptotically stable if and unstable when .
Proof.
The Jacobian matrix for (3) at disease-free equilibrium is given as
where and . According to the corollary, the Jacobian matrix will have negative eigenvalues if the following inequalities are satisfied:
Combining (13),(14),(17) and (18) we have:
This shows that the disease disease-free equilibrium is locally asymptotically stable if and unstable if . □
3.5. Existence of Endemic Equilibria
3.6. Global Stability of the Endemic Equilibrium
The global stability of the endemic equilibrium for system(3) is analysed.
Theorem 2.
If , then the endemic equilibrium point of the model is globally asymptotically stable on ℵ for
Proof.
We define a Lyapunov function:
Getting the time derivative we have:
After some algebraic manipulations, we have:
Clearly, and And since the arithmetic mean is greater than or equal to the arithmetic mean we have:
and
Thus, for all . Thus, by the asymptotic stability theorem [27], the positive endemic equilibrium state E1 is globally asymptotically stable in ℵ. □
4. Model simulations and Discussions
Numerical simulations of the model system (3) are carried out using python to illustrate some of the analytical results. Parameters values for the model simulation are provided in Table 2, some of these parameters were obtained from literature [5,28,29,30,31] while others were assumed (within realistic range) for the purpose of simulations.
The dynamics of the population of pregnant women without treatment are depicted in Figure 3, while those with treatment are shown in Figure 4. The dynamics of the mosquito population are presented in Figure 5.
The impact of administering IPTp-SP dosage within a population of pregnant women is investigated under the assumption that infection occurs exclusively via bites from infectious mosquitoes. The initial condition introduces 100,000 individuals in the latent compartment. Parameter values are drawn from Table 2. Figure 3 and Figure 5 illustrate the dynamics of the pregnant women and mosquito populations, respectively, in the absence of IPTp-SP treatment (). The corresponding basic reproduction number is estimated at 0.91, indicating that malaria transmission would subside over time even without intervention. For comparison, Figure 4 presents the system dynamics under the implementation of IPTp-SP. A comparative analysis reveals that the intervention significantly enhances disease suppression, supporting its role as an effective strategy for reducing malaria burden among pregnant women.
Introducing a treated proportion of pregnant women, defined by , clearly changes the system dynamics. Notably, there is a substantial increase in transitions to the recovered class among pregnant women, surpassing all other demographic compartments. This effect is likely driven by the reduced incidence of malaria infections under the IPTp-SP intervention, facilitating a more direct pathway to recovery. Moreover, the susceptible population declines to zero in the presence of treatment—contrasting with the no-intervention scenario, where susceptibility remained persistently high.
A mesh plot was employed to generate a three-dimensional representation of the influence of two critical parameters: the proportion of pregnant women receiving IPTp-SP treatment () and the treatment success rate (). As shown in Figure 6, the color gradient highlights a marked decline in the basic reproduction number, , as both and approach 1. This trend emphasizes the combined effect of high treatment coverage and efficacy, with values falling below unity when both parameters attain near-optimal levels—indicating the potential for disease elimination under effective intervention strategies.
5. Discussion and Conclusions
A compartmental model describing malaria transmission dynamics among pregnant women was developed, integrating both the human host and mosquito vector populations. The stability properties of the equilibrium points were analytically assessed, and numerical simulations were carried out to support and illustrate the theoretical results. Control of infection was examined through the implementation of an IPTp-SP intervention strategy. Analytical investigation demonstrated global stability of the disease-free equilibrium, and the basic reproduction number, , was shown to fall below unity under the intervention, suggesting the potential for malaria elimination within the pregnant population. While the model provides valuable insights, it remains a simplification of real-world dynamics and offers several avenues for refinement. For example, the current framework treats the intervention as one, yet stratification into discrete treatment levels—such as single-dose, double-dose, and three or more doses—could provide a more accurate understanding. Ultimately, an integrated model encompassing all treatment levels may offer a more robust depiction of the intervention. Furthermore, the assumption of constant treatment efficacy could be relaxed by incorporating the pharmacokinetics and pharmacodynamics (PK/PD) of IPTp-SP to reflect varying drug concentrations and biological response over time. These improvements could be explored through targeted numerical simulations to evaluate the impact of differential dosing and treatment dynamics on malaria control outcomes.
References
- WHO. Malaria, 2024.
- WHO. WHO guidelines for malaria; World Health Organization: Geneva, 2024; pp. 33–146. [Google Scholar] [CrossRef]
- CDC. About Malaria, 2024.
- Schantz-Dunn, J.; Nour, N. Malaria and Pregnancy: A Global Health Perspective. REVIEWS IN OBSTETRICS & GYNECOLOGY 2009, 2, 186–192. [Google Scholar] [CrossRef]
- WHO. World malaria report 2024, 2024.
- Guilbert, L.J.; Abbasi, M.; Mosmann, T.R. The immunology of pregnancy: Maternal defenses against infectious diseases; Malaria in Pregnancy Deadly Parasite, Susceptible Host; Taylor & Francis: London, 2001; pp. 56–69. [Google Scholar] [CrossRef]
- Doritchamou, J.Y.A.; Aitken, E.H.; Luty, A.J.F. Editorial: Immunity to Parasitic Infections in Pregnancy. Frontiers in Immunology 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Bauserman, M.; Conroy, A.L.; North, K.; Patterson, J.; Bose, C.; Meshnick, S. An overview of malaria in pregnancy. Seminars in Perinatology 2019, 43, 282–290. [Google Scholar] [CrossRef]
- Apinjoh, T.O.; Ntui, V.N.; Chi, H.F.; Moyeh, M.N.; Toussi, C.T.; Mayaba, J.M.; Tangi, L.N.; Kwi, P.N.; Anchang-Kimbi, J.K.; Dionne-Odom, J.; et al. Intermittent preventive treatment with Sulphadoxine-Pyrimethamine (IPTp-SP) is associated with protection against sub-microscopic P. falciparum infection in pregnant women during the low transmission dry season in southwestern Cameroon: A Semi - longitudinal study. PLoS ONE 2022, 17, e0275370–e0275370. [Google Scholar] [CrossRef]
- Agyeman, Y.N.; Newton, S.K.; Annor, R.B.; Owusu-Dabo, E. The Effectiveness of the Revised Intermittent Preventive Treatment with Sulphadoxine Pyrimethamine (IPTp-SP) in the Prevention of Malaria among Pregnant Women in Northern Ghana. Journal of Tropical Medicine 2020, 2020, 1–9. [Google Scholar] [CrossRef]
- Gutman, J.; Kalilani, L.; Taylor, S.; Zhou, Z.; Wiegand, R.E.; Thwai, K.L.; Mwandama, D.; Khairallah, C.; Madanitsa, M.; Chaluluka, E.; et al. The A581G Mutation in the Gene Encoding Plasmodium falciparum Dihydropteroate Synthetase Reduces the Effectiveness of Sulfadoxine-Pyrimethamine Preventive Therapy in Malawian Pregnant Women. Journal of Infectious Diseases 2015, 211, 1997–2005. [Google Scholar] [CrossRef]
- Anchang-Kimbi, J.K.; Kalaji, L.N.; Mbacham, H.F.; Wepnje, G.B.; Apinjoh, T.O.; Ngole Sumbele, I.U.; Dionne-Odom, J.; Tita, A.T.N.; Achidi, E.A. Coverage and effectiveness of intermittent preventive treatment in pregnancy with sulfadoxine–pyrimethamine (IPTp-SP) on adverse pregnancy outcomes in the Mount Cameroon area, South West Cameroon. Malaria Journal 2020, 19. [Google Scholar] [CrossRef]
- Mandal, S.; Sarkar, R.R.; Sinha, S. Mathematical models of malaria - a review. Malaria Journal 2011, 10. [Google Scholar] [CrossRef]
- Smith, D.L.; Battle, K.E.; Hay, S.I.; Barker, C.M.; Scott, T.W.; McKenzie, F.E. Ross, Macdonald, and a Theory for the Dynamics and Control of Mosquito-Transmitted Pathogens. PLoS Pathogens 2012, 8, e1002588–e1002588. [Google Scholar] [CrossRef]
- Wallace, D.I.; Southworth, B.S.; Shi, X.; Chipman, J.W.; Githeko, A.K. A comparison of five malaria transmission models: benchmark tests and implications for disease control. Malaria Journal 2014, 13. [Google Scholar] [CrossRef] [PubMed]
- Pande, V.; Bahal, M.; Dua, J.; Gupta, A. Ronald Ross: Pioneer of Malaria Research and Nobel Laureate. Cureus 2024, 16. [Google Scholar] [CrossRef]
- Ngwa, G.A.; Shu, W.S. A mathematical model for endemic malaria with variable human and mosquito populations. Mathematical and Computer Modelling 2000, 32, 747–763. [Google Scholar] [CrossRef]
- Chitnis, N.; Cushing, J.M.; Hyman, J.M. Bifurcation Analysis of a Mathematical Model for Malaria Transmission. SIAM Journal on Applied Mathematics 2006, 67, 24–45. [Google Scholar] [CrossRef]
- Danquah, B.A.; Chirove, F.; Banasiak, J. Effective and ineffective treatment in a malaria model for humans in an endemic region. Afrika Matematika 2019, 30, 1181–1204. [Google Scholar] [CrossRef]
- Ayalew, A.; Molla, Y.; Woldegbreal, A. Modeling and Stability Analysis of the Dynamics of Malaria Disease Transmission with Some Control Strategies. Abstract and Applied Analysis 2024, 2024. [Google Scholar] [CrossRef]
- Ghosh, M.; Lashari, A.A.; Li, X.Z. Biological control of malaria: A mathematical model. Applied Mathematics and Computation 2025, 2013, 7923–7939. [Google Scholar] [CrossRef]
- Mwamtobe, P.M.; Abelman, S.; Tchuenche, M.J.; Kasambara, A. Optimal (Control of) Intervention Strategies for Malaria Epidemic in Karonga District, Malawi. Abstract and Applied Analysis 2014, 2014, 1–20. [Google Scholar] [CrossRef]
- Birkhoff, G.; Rota, G.C. Ordinary Differential Equations, 4 ed.; John Wiley & Sons: New York, 1978. [Google Scholar]
- Driessche, P.v.d.; Watmough, J. Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission. Mathematical Biosciences 2002, 180, 29–48. [Google Scholar] [CrossRef] [PubMed]
- Adom-Konadu, A.; Lanor Sackitey, A.; Anokye, M. Local Stability Analysis Of Epidemic Models Using A Corollary Of Gershgorin’s Circle Theorem. Applied Mathematics E-Notes 2023, 23, 159–174. [Google Scholar]
- Allen, L.J. An Introduction to Mathematical Biology; Pearson Prentice Hall: Upper Saddle River, NJ, 2007; pp. 162–163. [Google Scholar]
- LaSalle, J. Some Extensions of Liapunov’s Second Method. IRE Transactions on Circuit Theory 1960, 7, 520–527. [Google Scholar] [CrossRef]
- UNFPA. Malawi, 2022.
- UNFPA. The State of the World’s Midwifery 2014 | United Nations Population Fund, 2022.
- Konlan, M. Modeling the Inflow of Exposed and Infected Migrants on the Dynamics of Malaria. European Journal of Mathematical Analysis 2024, 4, 7–7. [Google Scholar] [CrossRef]
- Niger, A.M.; Gumel, A.B. Mathematical analysis of the role of repeated exposure on malaria transmission dynamics. Differential Equations and Dynamical Systems 2008, 16, 251–287. [Google Scholar] [CrossRef]
Figure 1.
Illustration of the malaria life cycle, showing the stages of parasite development within the mosquito vector and human host ([3]).
Figure 1.
Illustration of the malaria life cycle, showing the stages of parasite development within the mosquito vector and human host ([3]).
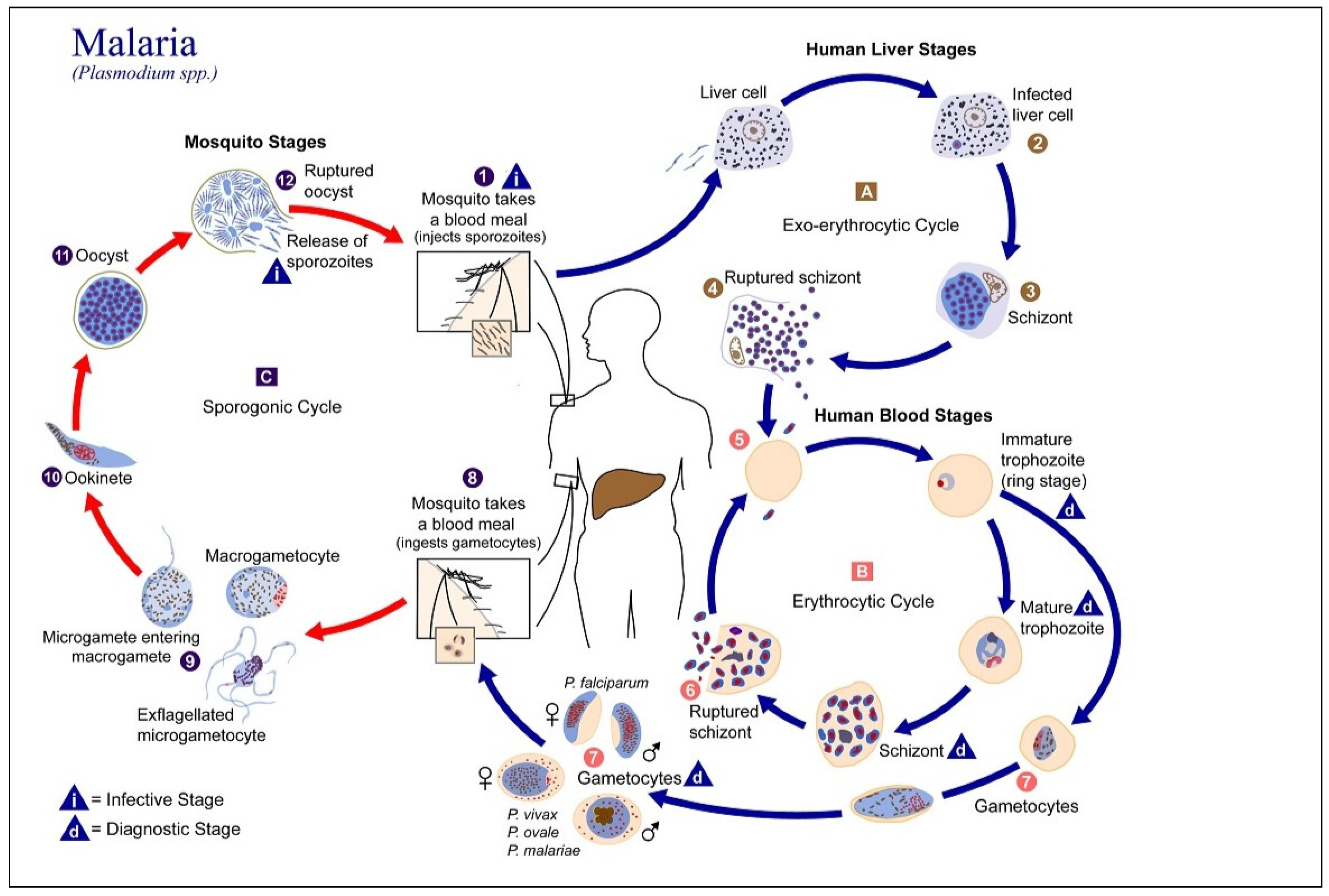
Figure 2.
Flowchart for malaria model with IPTp-SP as an intervention. The dash lines show that the infected mosquitoes infect the susceptible individuals and , the infected pregnant women infect the susceptible mosquitoes .
Figure 2.
Flowchart for malaria model with IPTp-SP as an intervention. The dash lines show that the infected mosquitoes infect the susceptible individuals and , the infected pregnant women infect the susceptible mosquitoes .
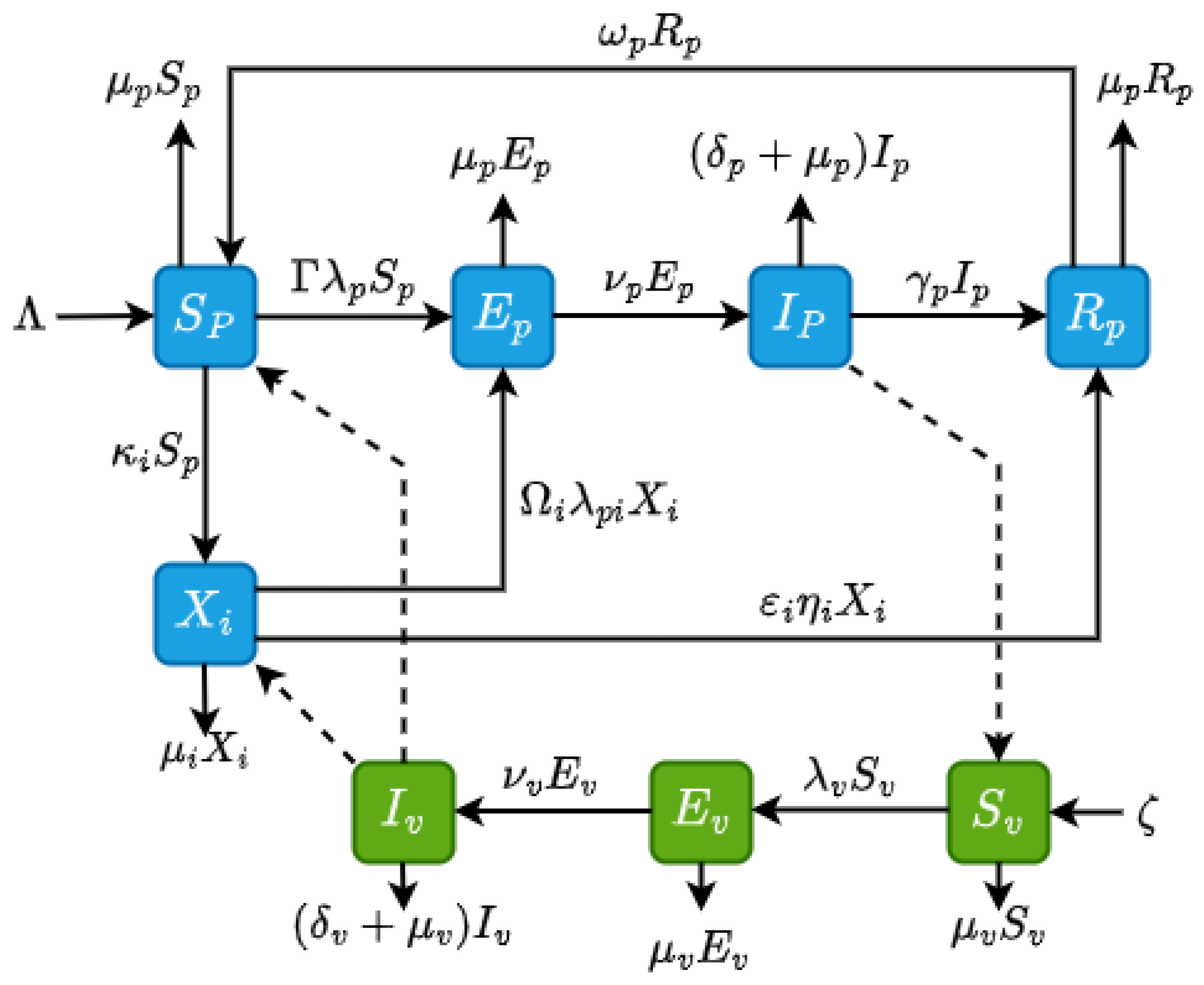
Figure 3.
Pregnant women dynamics showing infection and recovery patterns when IPtp-SP is not in use.
Figure 3.
Pregnant women dynamics showing infection and recovery patterns when IPtp-SP is not in use.
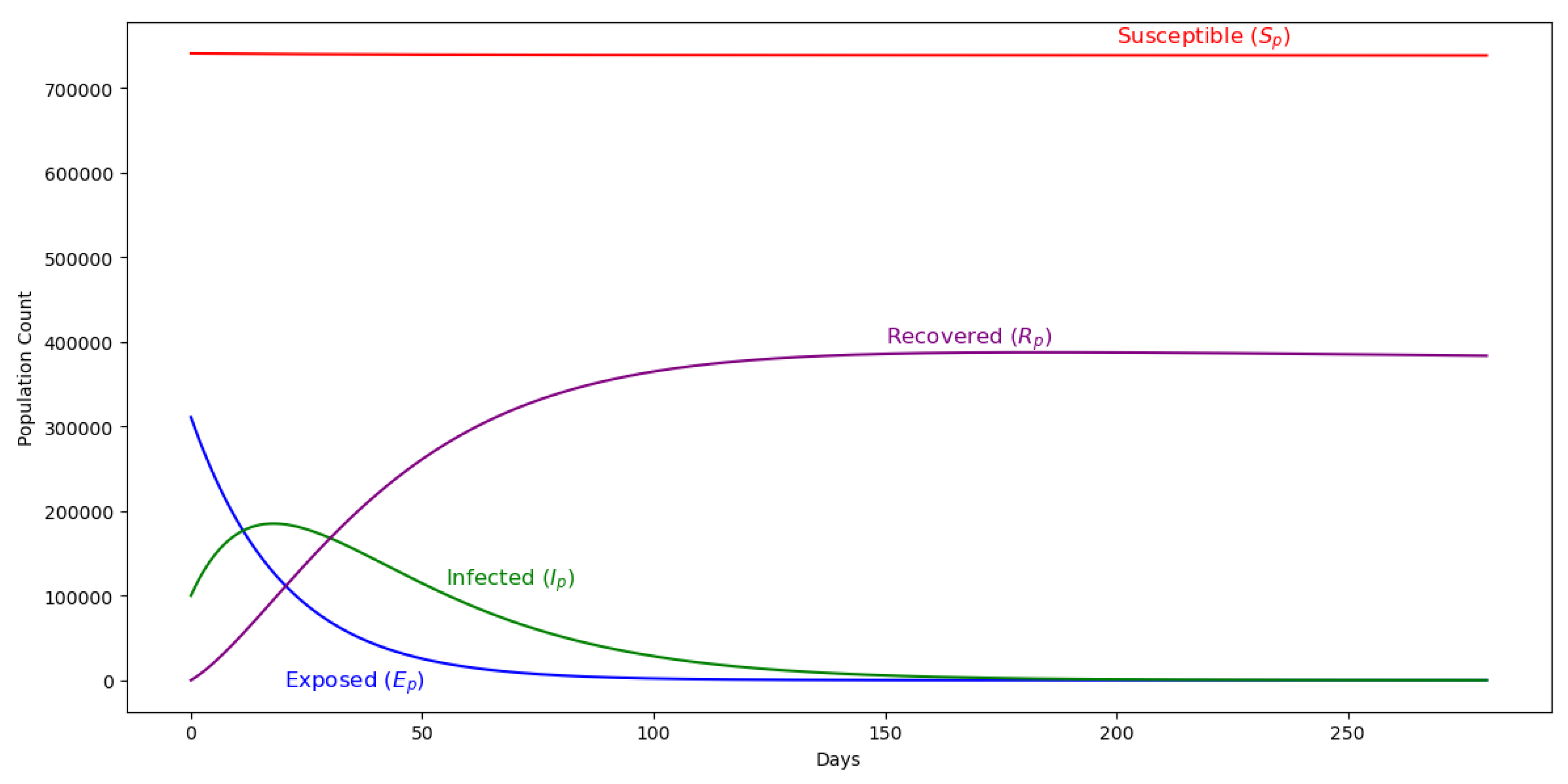
Figure 4.
Dynamics of the malaria model with IPTp-SP in use, illustrating its impact on infection rates, transmission reduction, and recovery among pregnant women over time.
Figure 4.
Dynamics of the malaria model with IPTp-SP in use, illustrating its impact on infection rates, transmission reduction, and recovery among pregnant women over time.
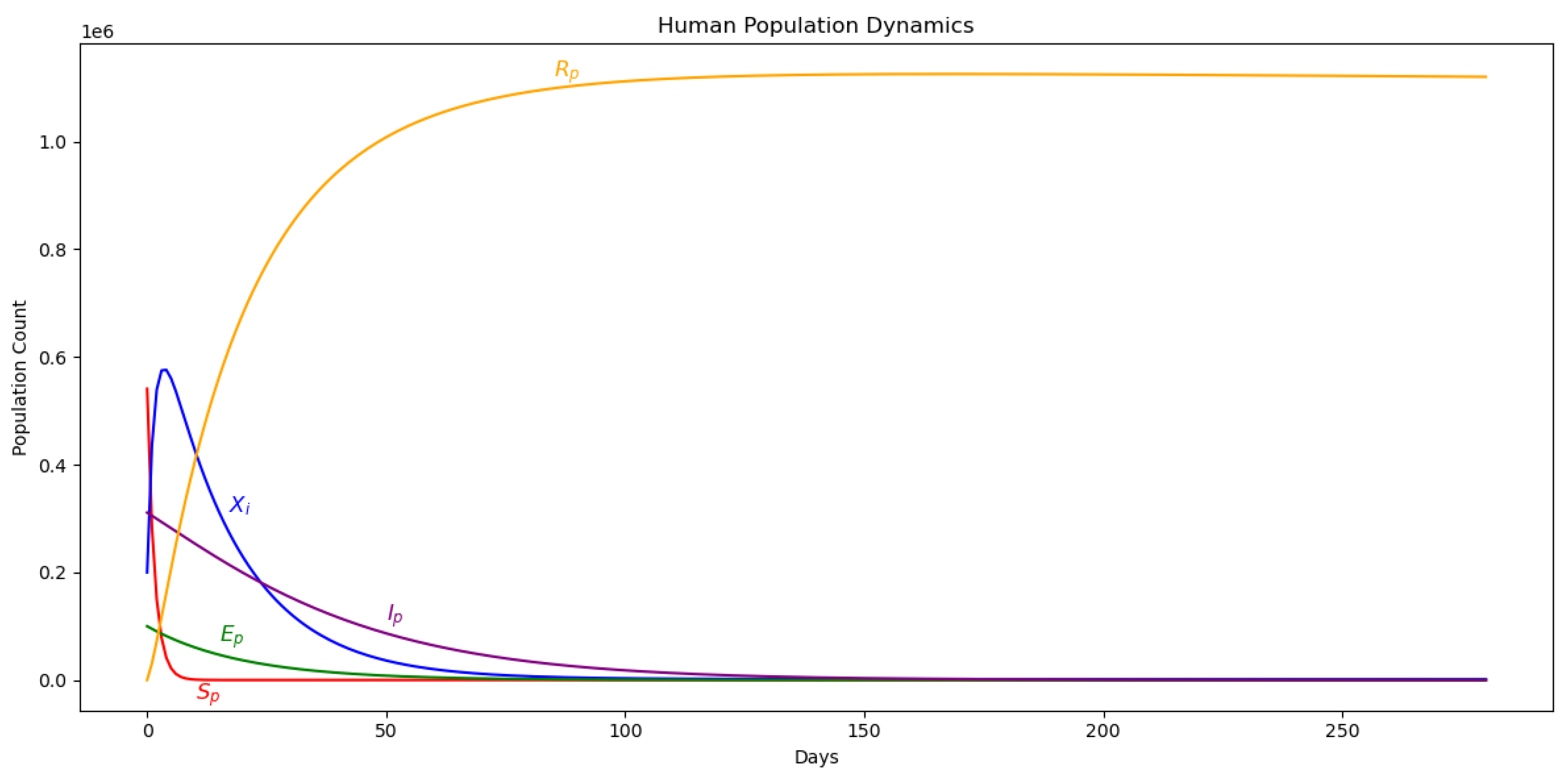
Figure 5.
Vector dynamics in the basic malaria model, showing transmission patterns.
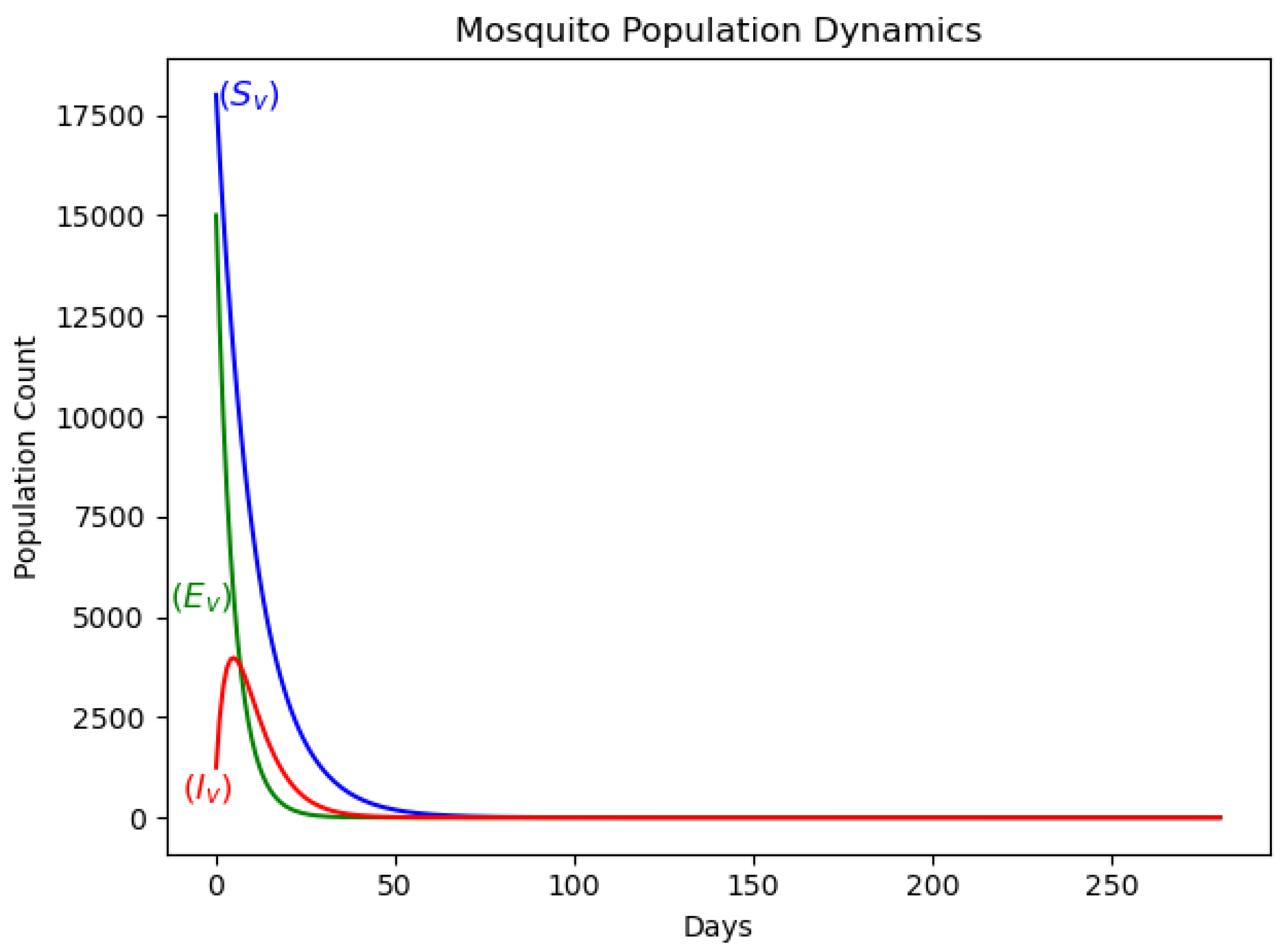
Figure 6.
Mesh plot illustrating the relationship between the fraction of pregnant women receiving IPTp-SP, treatment success rate, and the basic reproduction number , showing their combined impact on malaria transmission dynamics.
Figure 6.
Mesh plot illustrating the relationship between the fraction of pregnant women receiving IPTp-SP, treatment success rate, and the basic reproduction number , showing their combined impact on malaria transmission dynamics.
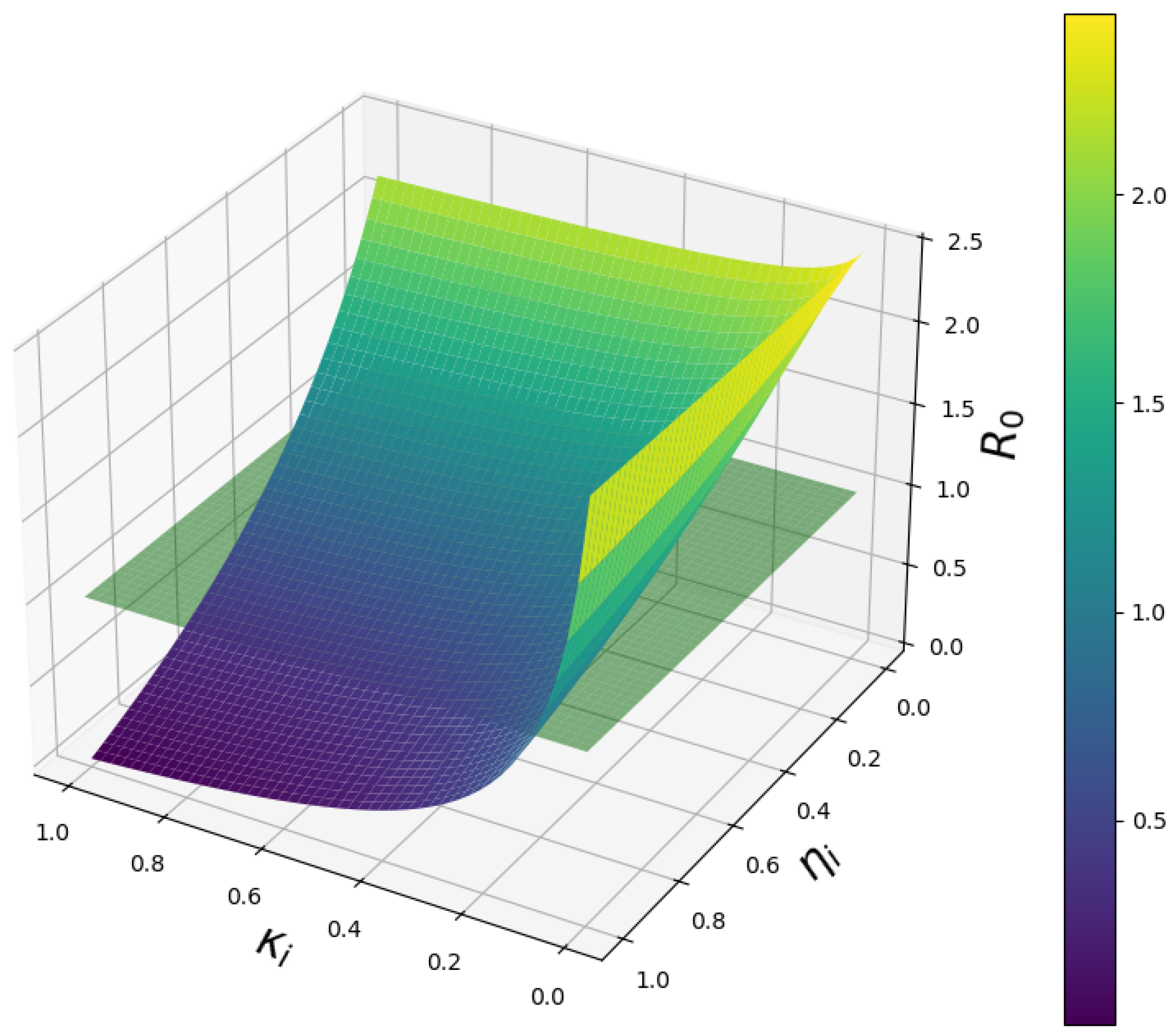

Table 1.
The parameters and description for the malaria model.
| Parameter | Description |
|---|---|
| The rate of conception of pregnant women | |
| The rate of recruitment of mosquitoes through natural birth | |
| Natural death rate of pregnant women per capita | |
| The natural death rate of mosquitoes per capita | |
| Transfer rate of pregnant women from the exposed state to the infectious state | |
| The rate of transfer of mosquitoes from the exposed state to the infectious state | |
| The infectivity of mosquitoes | |
| The infectivity of pregnant women | |
| The man-biting rate of mosquitoes. | |
| The disease induced death rate per capita for pregnant women | |
| The disease induced death rate per capita for mosquitoes | |
| Recovery rate of pregnant women with partial immunity | |
| The rate of losing immunity and going back to the susceptible | |
| Treatment success rate of doses of IPTp-SP | |
| Fraction of pregnant women protected from malaria by IPTp-SP | |
| Fraction of pregnant women taking IPTp-SP |
Table 2.
Parameter values for the malaria model with interventions.
| Parameter | Base Value | Source | Estimated Value |
|---|---|---|---|
| 0.0005948 | [28] | 0.188 | |
| 0.071 | [31] | 0.335 | |
| 0.07 | Assumed | 0.111 | |
| 0.94 | [30] | 0.977 | |
| [29] | 0.0984 | ||
| 0.00021 | [30] | 0.634 | |
| [30] | 0.490 | ||
| 0.00021 | [30] | 0.789 | |
| 0.001 | [30] | 0.223 | |
| 0.11346 | [30] | 0.257 | |
| [30] | 0.0555 | ||
| 0.091 | [30] | 0.223 | |
| [30] | 0.5000 | ||
| 0.64 | [5] | 0.159 | |
| 0.25 | Assumed | 0.133 | |
| 0.25 | Assumed | 0.633 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



