
Submitted:
19 June 2025
Posted:
20 June 2025
You are already at the latest version
Abstract
Keywords: Endometriosis-associated infertility; Epigenetics, biomarkers, prevention; Therapeutics.
Keywords:
Endometriosis-associated infertility
; Epigenetics
; biomarkers
; prevention
; Therapeutics
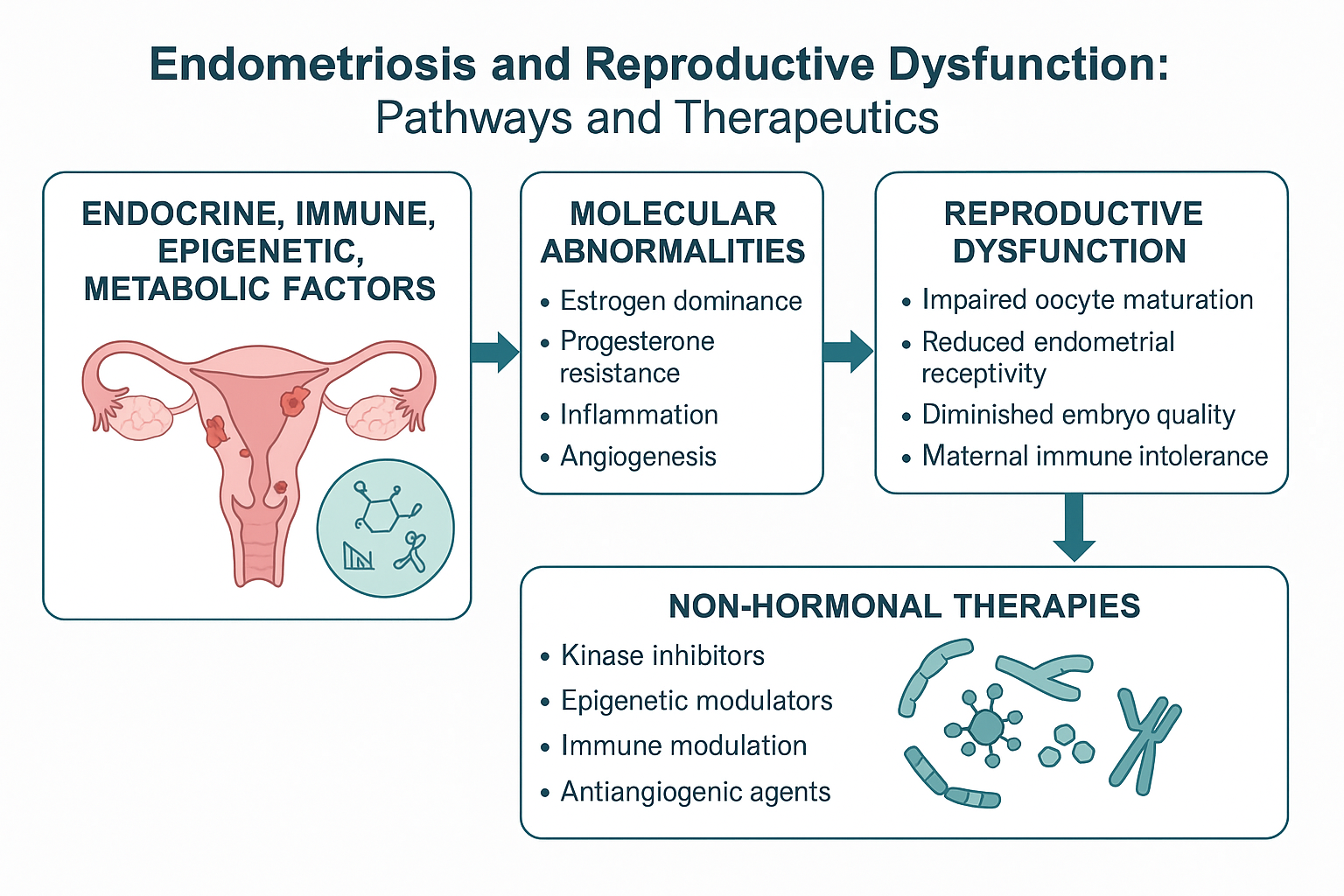
1. Introduction
Endometriosis—a chronic inflammatory disorder driven by estrogen-dependent growth of ectopic endometrial tissue—remains a formidable clinical challenge, profoundly impacting the physical, psychological, and reproductive health of 190 million women worldwide [1,2]. Despite affecting approximatively 10% of reproductive-age women and up to 50% of infertility cases, its elusive pathogenesis results in diagnostic delays averaging 7-10 years and limited therapeutic efficacy beyond symptomatic control [3]. Crucially, molecular dysregulation lies at the heart of this disease. Aberrant estrogen signaling, progesterone resistance, and immune dysfunction collaborate to create a self-sustaining microenvironment that promotes lesion survival, angiogenesis, and therapeutic evasion [4,5]. These same mechanisms directly impair key reproductive functions and contribute to infertility: not merely through anatomical distortion, but by disrupting oocyte competence, endometrial receptivity, and embryo-maternal crosstalk. Even minimal disease can inflict molecular scars that compromise pregnancy chances.
This review deciphers how endometriosis hijacks reproductive biology at the molecular level. We first unravel the pathogenic triad (hormonal, immune, and epigenetic disturbances) fueling lesion persistence. We then expose their direct fallout on fertility—from folliculogenesis to implantation. By connecting these dots, we highlight actionable therapeutic windows and advocate for precision care tailored to women’s reproductive aspirations.
2. Molecular Mechanisms of Endometriosis
The pathogenesis of endometriosis is driven by a complex interplay of hormonal, immune, oxidative, and epigenetic dysregulations. Estrogen excess and progesterone resistance sustain lesion growth, while chronic inflammation—mediated by interleukin (IL) -1β and tumor necrosis factor (TNF)-α—amplifies tissue injury. Impaired natural killer (NK) cell activity, oxidative DNA damage, angiogenic remodeling, and epithelial-to-mesenchymal transition (EMT) further promote immune evasion and invasiveness. Together, these processes form a self-reinforcing molecular loop that underpins lesion persistence and reproductive dysfunction. The following subsections dissect the key pathways contributing to this pathogenic cascade.
2.1. Hormonal Dysregulation
Endometriotic implants acquire biochemical autonomy, transforming into miniature endocrine organs that escape the timing and magnitude of ovarian cycles. This autonomy is driven by an interplay of enzyme induction, receptor reconfiguration, and epigenetic dysregulation, resulting in local estrogen excess and progesterone resistance [6]. Estrogen production is amplified on two fronts. First, prostaglandin-E₂ (PGE2) stimulates steroidogenic factor-1 (SF-1), enhancing Cytochrome P450 19A1 (CYP19A1)and aromatase expression and creating a feedback loop with estradiol and cyclo-oxygenase-2 [7,8,9]. Second, a recently documented sulfatase pathway supplements de-novo synthesis: steroid sulfatase liberates estrone from the abundant circulating reservoir of estrone-sulfate that is imported via the transporter Solute Carrier Family 10 Member 6 (SLC10A6), ensuring precursor availability even under low androgen conditions [10,11,12]. Simultaneously, reduced 17β-hydroxysteroid dehydrogenase type 2 (17β-HSD2) activity limits estradiol inactivation [13,14], amplifying local estrogenic effects.
Estrogen sensing is also altered. A dominance of Estrogen Receptor (ER) β over ERα in stromal cells promotes inflammation and reduces apoptosis, supporting lesion persistence [15,16,17,18,19]. Notably, ERβ also activates NACHT, LRR and PYD Domains-Containing Protein 3 (NLRP3) inflammasome via caspase-1, increasing interleukin-1β and linking estrogen signaling directly to sterile inflammation [20,21,22]. At the same time, cytoplasmic ERβ disrupts TNF-α-mediated apoptosis by forming complexes with Apoptosis Signal-Regulating Kinase 1 (ASK1) , Serine/Threonine Kinase Receptor-Associated Protein (STRAP), and 14-3-3 proteins [23,24,25]. Membrane-anchored G-protein-coupled estrogen receptor (GPER, also known as GPR30) accelerates estrogenic signaling by mobilizing cAMP and trans-activating SF-1, further boosting aromatase transcription [26,27,28,29]. This pathway may explain lesion responsiveness to environmental xenoestrogens like bisphenol A (BPA) and the Selective Estrogen Receptor Modulator (SERM) tamoxifen.
Hormonal dysregulation is closely tied to aberrant epigenetic regulation. Promoter hypermethylation and histone deacetylase (HDAC)-mediated repression of the progesterone receptor gene reduce progesterone receptor isoform B (PR-B) expression, a hallmark of progesterone resistance [30,31,32,33]. Estrogen enhances these changes by upregulating DNA methyltransferase 1 (DNMT1), promoting Runt-related transcription factor (RUNX3) hypermethylation and silencing [34]. BPA contributes through a WD repeat domain 5 (WDR5)/ten-eleven translocation methylcytosine dioxygenase 2 (TET2) axis that increases ERβ transcription [35]. Metabolic–epigenetic crosstalk has recently emerged as a further amplifier. The cellular prion protein (PrPC) has emerged as an estrogen-responsive gene that represses peroxisome proliferator-activated receptor-α (PPAR-α), driving intracellular cholesterol accumulation and fuelling additional aromatase activity [36]. Estradiol, in turn, up-regulates PrPC, closing a vicious metabolic loop [37,38,39].
Collectively, enhanced enzymatic activity, altered receptor signaling, and epigenetic reprogramming shape an estrogen-dominant, progesterone-resistant environment that initiates and sustains downstream cellular dysregulation.
2.2. Epigenetic Reprogramming and Non-Coding RNAs
Endometriotic chromatin is marked by stable yet dynamic epigenetic alterations that reshape transcription long before lesions become clinically apparent. Recent methylome and histone-mark surveys confirm that DNA methylation and post-translational modifications cooperate to reprogram gene expression [40]. CpG hypermethylation at the promoters of Homeobox Protein A10 (HOXA10) and PR-B silences genes crucial for endometrial receptivity, reinforcing progesterone resistance and impaired decidualization [30,40,41,42,43]. These changes are strengthened by histone deacetylation—particularly via sirtuins and class-I/II HDACs—which enforce a transcriptionally repressive chromatin conformation. Superimposed on this epigenetic framework, non-coding RNAs exert an agile layer of gene regulation. MicroRNAs (miRNAs) such as miR-135a/b and miR-135b repress HOXA10 [44,45], mirroring its epigenetic silencing, while others (miR-29c, miR-194-3p) modulate immune response, stromal proliferation, and extracellular matrix dynamics [46,47,48,49,50]. These miRNAs, often upregulated by inflammatory cytokines, act as both sensors and amplifiers of lesion inflammation. Regulation becomes still more intricate with long non-coding RNAs (lncRNAs; >200 nt) and circular RNAs (circRNAs; covalently closed transcripts that resist exonuclease degradation) [51]. The lncRNA H19 sequesters multiple miRNA, facilitating EMT and immune evasion [52,53,54]. Parallelly, circ_0007331 captures miR-200c-3p, stabilizing hypoxia-inducible factor-1α (HIF-1α) and promoting both steroid resistance and angiogenesis [55,56]. These lncRNAs and circRNAs engage in competing endogenous RNA (ceRNA) networks, modulating transcript abundance through shared miRNA interactions.
2.3. Intracellular Signaling Pathways
On top of the epigenetically altered chromatin landscape, endometriotic cells engage multiple constitutively active intracellular signaling pathways that drive their survival, invasion, and proliferation. The phosphoinositide 3-kinase/protein kinase B/mammalian target of rapamycin (PI3K/AKT/mTOR) and Wingless-related integration site (Wnt)/β-catenin pathways are central to this dysregulated network, enhancing cellular fitness while promoting hormonal and immune escape [57,58,59,60,61]. Hyperactivation of PI3K–AKT–mTOR heightens glucose uptake, stimulates aerobic glycolysis and neovascularization, and impairs immune surveillance, thus granting ectopic foci a tumor-like resilience [62,63,64]. Activated AKT further diminishes progesterone-receptor activity, tightening the hormonal resistance already established by epigenetic modifications [65]. Simultaneously, dysregulated Wnt signaling facilitates nuclear translocation of β-catenin, which complexes with T-cell factor (TCF)/Lymphoid Enhancer-binding Factor (LEF) transcription factors to drive the expression of EMT markers, motility genes, and matrix-remodeling proteases such as matrix metalloproteinases (MMP)-2 and MMP-9 [60,66]. These pathways intersect with the transforming growth factor-β (TGF-β) signaling axis, an upstream conductor of EMT and fibrogenesis. Upon activation, (S) homologues of Mothers Against Decapentaplegic (SMAD) complexes cooperate with β-catenin and phosphorylated AKT to upregulate Snail Family Transcriptional Repressor 1 (SNAIL), Snail Family Transcriptional Repressor 2 (SLUG), and Zinc Finger E-box-Binding Homeobox 1 and 2 (ZEB1/2), repress E-cadherin, and induce vimentin and N-cadherin, thereby reducing cell–cell adhesion and promoting deep tissue invasion—hallmarks of advanced endometriosis [67].
Together, PI3K/AKT/mTOR, Wnt/β-catenin, and TGF-β form a tightly linked, self-reinforcing network that integrates hormonal, inflammatory and metabolic signals. This robust molecular machinery converts endometrial cells into a semi-transformed phenotype that is plastic, invasive, and resistant to endocrine regulation—sustaining lesion persistence even in adverse peritoneal conditions.
2.4. Inflammation, Immune Cell Recruitment, and Neuro-Immune Crosstalk
Intracellular hyperactivation alone does not explain how ectopic tissue persist under mechanical stress and immune surveillance. The same signals that activate PI3K/AKT/mTOR and Wnt/β-catenin pathways also ignite chronic inflammation, which recruits innate and adaptive immune cells and wires pain circuitry into lesions. Peritoneal fluid from affected women contains abundant macrophages, dendritic cells, neutrophils, mast cells, and both CD4⁺ and CD8⁺ lymphocytes. These cells accumulate in ectopic sites and amplify cytokine production [68,69,70]. Recent single-cell RNA sequencing (scRNA-seq) has identified a macrophage subset within lesions that mimics "foamy" tumor-associated macrophages. These cells suppress cholesterol-efflux pumps ATP-binding cassette transporter A1 (ABCA1) and ATP-binding cassette transporter G1 (ABCG1), accumulate lipids, and hyperactivate IL-1β/IL-6 inflammasomes and PGE₂ synthesis [71]. This links metabolic overload to the cytokine–pain axis and highlights ABCA1/ABCG1 and their upstream Liver X Receptor–Peroxisome Proliferator-Activated Receptor (LXR–PPAR) regulators as promising therapeutic targets.
Danger signals like hypoxia and oxidized cell debris drive the release of IL-1β, IL-6, IL-8, TNF-α and IL-17A through Th17 differentiation. These cytokines are linked to immune dysfunction, angiogenesis and neuroinflammation [72,73,74]. IL-17A, in particular, promotes vascular endothelial growth factor (VEGF) transcription, enhances MMP production, and when neutralized, reduces vascularization and lesion size in models [75]. IL-1β and TNF-α sustain this inflammatory state by upregulating cyclo-oxygenase-2 (COX2) and reinforcing the PGE₂-estrogen loop [29,76,77,78]. Complement activation further exacerbates the environment. Complement C5a receptor (C5aR1)-positive macrophages and Schwann cells heighten amplify inflammasome activity and mechanical allodynia [79]. Mast-cell infiltration also contributes. Tryptase-positive mast cells accumulate in deep infiltrating endometriosis, activating Protease-Activated Receptor 2 (PAR-2), promoting fibrosis and sustaining neurogenic inflammation [80]. All these cues converge on the TGF-β axis. Activated SMAD complexes work with β-catenin and phosphorylated AKT to upregulate SNAIL, SLUG and ZEB1/2, repress E-cadherin and promote fibroblast-to-myofibroblast transition, key events in matrix stiffening and fibrosis [81,82]. TGF-β also promotes immune tolerance by expanding Forkhead Box P3-Positive (FOXP3⁺) regulatory T cells and suppressing cytotoxicity [83,84,85]. Neuro-immune crosstalk closes the circle. Pro-inflammatory cytokines (TNF-α, IL-1β, IL-6) induce nerve-growth factor (NGF) and brain-derived neurotrophic factor, which stimulate nerve fiber sprouting and lower nociceptor thresholds [86]. The resulting peritoneal niche is thus proinflammatory, neurotrophic, and angiogenic— conditions that support lesion growth, invasion, and pain.
2.5. Oxidative Stress and Peritoneal Toxicity
Retrograde menstruation and lesion hemorrhage expose the peritoneal cavity to hemolyzed erythrocytes. Macrophage-mediated clearance releases ferrous iron, overwhelming iron-binding proteins and triggering Fenton reactions that produce hydroxyl radicals. Elevated ferritin, transferrin, and free iron in peritoneal fluid coincide with oxidative damage markers, including lipid peroxides and protein carbonyls [87,88,89,90,91]. This oxidative stress disrupts mitochondrial membrane potential and electron transport, leading to cytochrome c release and impaired DNA repair [92,93,94]. Damaged macromolecules act as danger signals, attracting and activating macrophages and reinforcing inflammation [95]. Reactive oxygen species (ROS) also modulate gene expression by altering DNA methylation at antioxidant loci via Ten-Eleven Translocation dioxygenase (TET) enzymes, sustaining pro-survival transcriptional programs [96]. Systemically, ROS damage granulosa cells, impair mitochondrial distribution [97], reduce glutathione stores, and compromise meiotic spindle stability, factors that affect oocyte quality [98]. In the peritoneal niche, oxidative damage remodels the extracellular matrix by modifying structural proteins and activating redox-sensitive proteases, facilitating the invasive processes discussed in the next section.
2.6. Extracellular Matrix Remodeling, EMT, and Fibrogenesis
Oxidative and inflammatory cues initiate a cascade of structural remodeling in the extracellular-matrix (ECM). MMPs, particularly MMP-2 and MMP-9, drive ECM degradation and are elevated in serum, peritoneal fluid and lesion tissue [99,100]. Their role has now been confirmed by a 2024 meta-analysis of over 1,000 patients [101]. Spatially divergent levels of tissue inhibitors of metalloproteinases (TIMPs) create gradients of proteolytic activity that guide directional invasion [102,103,104]. The urokinase-type plasminogen activator (uPA) system further enhances matrix remodeling. Overexpression of uPA and its receptor (uPAR) facilitates plasmin generation, which activates latent MMPs and degrades fibrin and fibronectin [105,106,107]. TGF-β–induced PAI-1 signaling through uPAR has been linked to endothelial senescence and fibrosis [108]. Concurrently, epithelial cells undergo EMT, losing polarity and epithelial markers like E-cadherin while gaining mesenchymal traits including vimentin and N-cadherin. These changes, driven by TGF-β, oxidative stress, hypoxia, and cytokines, are more pronounced in advanced disease stages [109,110]. Fibrogenesis is completed through TGF-β/SMAD–mediated conversion of fibroblasts into myofibroblasts, which deposit dense collagen I, fibronectin and laminin. Enzymes such as lysyl oxidase (LOX) stiffen this matrix, correlating with pain intensity, disease severity and surgical difficulty [111]. Redox-sensitive LOX-like proteins and angiotensin II/Angiotensin II Type 1 Receptor (AT1R) signaling further potentiate fibrosis [106]. In sum, ECM degradation, EMT, and fibrogenesis form a reinforcing loop that embeds endometrial cells within a fibrotic, immune-evasive niche.
2.7. Aberrant Vascular Remodeling and Non-Canonical Angiogenic Pathways
Endometriotic angiogenesis diverges markedly from physiological endometrial vascularization. Hypoxia and oxidative stress stabilize Hypoxia-Inducible Factor-1 α (HIF-1α), which cooperates with inflammatory cytokines to drive a burst of VEGF-A. However, resulting vessels are disorganized, leaky, and poorly supported by pericytes [112,113]. Non-canonical angiogenic signals further shape the aberrant vasculature. The Slit guidance ligand 2 (Slit2)/Roundabout guidance receptor 1 (Robo1) guidance system disrupts vessel branching and stability [114,115], while prokineticin-1 (PROK1) and its receptor PROKR1 axis enhances endothelial proliferation and permeability. Importantly, PROK1 also interfaces with Nuclear Factor κ-Light-Chain-Enhancer of Activated B Cells (NF-κB)-driven transcriptional programs, reinforcing the molecular crosstalk between angiogenesis, inflammation, and estrogen signaling [116]. Additionally, suppression of Notch/Delta-like ligand 4 (Dll4) signaling—normally a regulator of tip-cell formation—exacerbates vascular disorganization and excessive sprouting [117]. Collectively, these alterations generate a structurally unstable yet functionally permissive microvascular network that facilitates stromal invasion, immune cell infiltration, neural ingrowth and pain. Recognition of these VEGF-independent mechanisms opens new avenues for precision-based anti-angiogenic therapies
Together, these interwoven pathways—hormonal, inflammatory, oxidative, and structural—create a reinforcing network that sustains endometriotic lesions and impairs reproductive function.
3. Molecular Bridges Between Endometriosis and Infertility
Infertility associated with endometriosis unfolds as a progressive, system-wide injury that shadows every milestone of the reproductive journey—from initial follicle recruitment in the ovary to the moment a competent blastocyst meets a receptive endometrium. Lesion-derived cytokines, iron-driven oxidative bursts, endocrine disequilibrium and epigenetic drift leak far beyond ectopic implants, rewriting the molecular script of tissues that can appear perfectly normal on ultrasound or at laparoscopy. Recent single-cell atlases, high-resolution metabolomics and spatial transcriptomics now allow these hidden injuries to be traced with unprecedented precision, revealing a common signature of inflammatory, oxidative and hormonal stress across the ovary, fallopian tube and uterus [118]. Below, six interconnected facets illustrate how this ripple effect steadily erodes reproductive competence (Figure 1).
3.1. Ovarian Folliculogenesis and Reserve Erosion
The ovarian environment in endometriosis is subject to persistent inflammatory and oxidative stress, which undermines folliculogenesis and progressively diminishes ovarian reserve. Key pro-inflammatory cytokines, including TNF-α and IL-1β, have been shown to impair granulosa cell function, notably by downregulating essential genes such as the FSH receptor (FSHR) and aromatase (CYP19A1)—two critical mediators of follicle growth and estrogen synthesis [119,120,121,122,123]. The presence of endometriomas—ovarian cysts lined with ectopic endometrial tissue—exerts both mechanical and biochemical insults on adjacent ovarian cortex. The cyst fluid, often enriched with free iron, proteolytic enzymes, and ROS, contributes to stromal fibrosis, follicular apoptosis, and the premature activation of primordial follicles, which over time exhausts the follicular reserve. Clinically, these effects are reflected by lower serum anti-Müllerian hormone (AMH) levels and reduced antral follicle counts, both of which serve as biomarkers of diminished ovarian reserve [124]. While surgical excision of endometriomas is often necessary—particularly for large, symptomatic lesions—it carries the risk of unintentional removal of healthy cortical tissue, further compromising ovarian reserve. This trade-off highlights the importance of careful preoperative assessment, particularly in women of reproductive age, and supports the adoption of fertility-preserving strategies, including cyst-sparing techniques, ovarian tissue cryopreservation, or adjunctive medical suppression before and after surgery [125].
3.2. Oxidative Damage to Oocytes and Early Embryos
The same oxidative siege compromises oocyte integrity. Exposure of human oocytes to pathophysiological ROS (≈150 µM H₂O₂) disrupts mitochondrial distribution, fragments actin–microtubule scaffolds, and misaligns kinetochores, reducing fertilization [126,127,128,129,130,131,132]. Down-stream, early embryos from women with endometriosis display higher rates of fragmentation and developmental arrest, consistent with the carry-over of oxidative and spindle damage [133].
3.3. Tubal Ciliary Dysfunction and Peritoneal Toxicity
In endometriosis, the chronic inflammatory milieu extends beyond the pelvic lesions and profoundly affects the fallopian tubes and peritoneal environment, disrupting key steps in fertilization and early embryo transport. Elevated levels of pro-inflammatory cytokines, particularly IL-6 and TNF-α, have been shown to impair both ciliary activity and smooth muscle contractility, two essential mechanisms for gamete capture and embryo transit [134,135,136]. Chronic exposure to cytokines and prostaglandins damages the ciliated epithelium and impairs smooth muscle contractility, disrupting coordinated gamete and embryo transport [136,137]. Simultaneously, peritoneal adhesions resulting from fibrosis and scarring can form physical barriers that hinder sperm-oocyte interaction and embryo migration. Additionally, the peritoneal fluid in endometriosis is enriched in pro-inflammatory cytokines, proteases, and ROS, which together create a toxic microenvironment detrimental to fertilization and early embryonic development [138,139,140].
3.4. Endometrial Receptivity Breakdown
Because the embryo reaches the uterus just a few days after fertilization, the brief “window of implantation” acts as the earliest decisive link between fertilization, blastocyst development, and uterine attachment. Disruption at this juncture can swiftly lead to infertility. Adequate endometrial receptivity remains indispensable for implantation [141], yet in women with endometriosis, key receptivity markers are epigenetically silenced [142,143]. Among these, HOXA10 is central to stromal differentiation and integrin expression, and its silencing results in reduced levels of integrin αvβ3, leukemia inhibitory factor (LIF), and glycodelin—all of which are critical for embryo adhesion [43,144,145]. Transcriptomic analyses show that over-expression of miR-135a/b reinforces HOXA10 silencing, tightening this epigenetic brake on receptivity [44]. Concurrently, diminished PR-B expression limits stromal decidualization and contributes to progesterone resistance [146,147]. These changes are reinforced by aberrant activation of PI3K/AKT and mitogen-activated protein kinase (MAPK) pathways, which further impair endometrial receptivity [148,149].
3.5. Maternal–Fetal Immune Imbalance and Implantation Failure
Once the blastocyst attaches to a receptive endometrium, the maternal immune system must swiftly shift from surveillance to tolerance at the maternal–fetal interface; in endometriosis, disruption of this transition forms the second critical bottleneck leading to implantation failure [150,151]. Dysfunctional uterine natural killer (uNK) cells exhibit altered cytokine secretion and reduced angiogenic potential, impairing spiral artery remodeling and early placental development [152]. Concurrently, a deficiency in regulatory T cells (Tregs) disrupts maternal immune tolerance, increasing the risk of embryo rejection [153,154]. A skew toward pro-inflammatory Th17 cells—driven by an up-regulated IL-23/Th17 axis—exacerbates oxidative stress and destabilizes the implantation micro-environment [155]. Clinical studies in women with repeated implantation failure confirm an elevated Th17/Treg ratio and heightened endometrial inflammation [156], underscoring the immunologic hurdle to successful nidation.
3.6. Genetic Susceptibility and Metabolic Rewiring
Even after fertilization and a provisional immune truce, inherited genetic variants and metabolic reprogramming can still compromise embryonic development and placentation—highlighting the substantial contribution of genetic predisposition to both endometriosis and infertility [157]. Genome-wide association studies (GWAS) have identified susceptibility loci such as Wnt family member 4 (WNT4), Vezatin Adherens junctions Transmembrane protein (VEZT), and cyclin dependent kinase inhibitor 2B antisense RNA 1 (CDKN2B-AS1) that influence uterine development, endometrial function, and cellular proliferation [158,159,160]. Metabolic reprogramming in endometriotic lesions—including altered tryptophan and arginine metabolism—perturbs immune regulation, redox balance, and local inflammation [161,162,163]. Targeted metabolomics has further uncovered dysregulated sphingolipid and ceramide pathways in follicular fluid from women with endometriosis, correlating with poorer in vitro fertilization (IVF) outcomes [164]. At the transcriptome level, compromised LIF- Signal Transducer and Activator of Transcription 3 (STAT3) signaling and progesterone response have been demonstrated in the eutopic endometrium of patients with repeated implantation failure [165], closing the loop between metabolic, inflammatory, and hormonal insults that collectively undermine reproductive success.
4. Emerging Therapeutic Targets in Endometriosis-Associated Infertility
Recent advances in the molecular understanding of endometriosis have catalyzed the development of targeted therapies aimed not only at symptom management but also at preserving and restoring fertility. This represents a shift away from traditional symptom-oriented treatments toward mechanistically driven interventions that directly address the biological underpinnings of lesion persistence and reproductive dysfunction [166]. Several emerging non-hormonal approaches now focus on key molecular pathways—such as inflammation, oxidative stress, angiogenesis, and immune imbalance—central to the disease’s pathogenesis (Figure 2).
4.1. Targeting Estrogen Receptor Beta (ERβ) Signaling.
Endometriotic lesions consistently exhibit dysregulated estrogen receptor profile, marked by a heightened ERβ-to-ERα ratio. This receptor imbalance fosters a microenvironment marked by sustained inflammation, apoptotic resistance, progesterone insensitivity, and enhanced angiogenesis. Through transcriptional reprogramming, ERβ activation promotes lesion survival and impairs endometrial receptivity. Selective ERβ antagonists have demonstrated promising results in preclinical models, reducing lesion growth and partially restoring progesterone responsiveness [167,168]. Unlike systemic estrogen suppression, these compounds allow targeted modulation of local estrogen signaling without compromising ovulation, making them particularly attractive for fertility-preserving interventions. In parallel, attention has turned to intracrine steroidogenesis and receptor subtype modulation to recalibrate the hormonal landscape of the endometrium. Selective estrogen receptor modulators (SERMs), selective androgen receptor modulators (SARMs), and selective androgen receptor degraders (SARDs) are currently being explored for their capacity to restore endocrine balance and enhance receptivity without systemic suppression. These compounds frequently converge on metabolic enzymes such as aromatase, steroid sulfatase, and Aldo-Keto Reductase family 1 member C3 (AKR1C3), which regulate local estrogen and androgen biosynthesis within stromal and epithelial compartments. By modulating cross-talk among hormonal, immune, and vascular networks, these agents aim to reestablish a receptive endometrial microenvironment and improve fertility outcomes [169].
Altogether, ERβ-targeted therapies represent a compelling alternative to traditional hormonal suppression. Their ability to selectively modulate the pathological features of endometriotic tissue while preserving systemic reproductive function places them at the forefront of emerging fertility-friendly treatments.
4.2. Inhibition of inflammatory Kinase Pathways
Recent advances in the molecular profiling of endometriosis have highlighted several intracellular kinase pathways as central orchestrators of lesion persistence, chronic inflammation, and hormonal resistance. Among these, the NF-κB pathway is one of the most consistently activated in ectopic endometrial tissue. Its sustained activation drives the transcription of pro-inflammatory cytokines, enhances anti-apoptotic signaling, and promotes an immune-tolerant lesion microenvironment [170,171]. The MAPK cascade — including extracellular signal-regulated kinases 1 and 2 (ERK1/2), c-Jun N-terminal kinase (JNK), and p38— is similarly hyperactivated in endometriotic cells. This pathway promotes epithelial proliferation, matrix degradation, and immune cell recruitment. Moreover, the PI3K/AKT/mTOR signaling axis, which has been extensively characterized in oncology, supports stromal cell growth, neovascularization, and insensitivity to progesterone-based therapies [172,173].
Pharmacological inhibition of these kinase pathways has shown encouraging results in preclinical studies. For example, Mitogen-Activated Protein Kinase kinase (MEK) inhibitors (e.g., U0126) and NF-κB pathway blockers (e.g., BAY11-7082) have been shown to significantly reduce lesion volume, epithelial proliferation, and inflammatory infiltration in rodent models. These agents also downregulate markers associated with proliferation (Proliferating Cell Nuclear Antigen (PCNA)) and invasion (MMP9), indicating a multi-layered effect on lesion pathophysiology [174]. The mTOR pathway has attracted significant interest due to its dual role in cell metabolism and immunomodulation. Inhibitors such as everolimus and rapamycin—already in clinical use for oncology and transplant medicine—are now being evaluated in endometriosis models. Beyond suppressing lesion growth, mTOR inhibition appears to improve implantation rates, reduce miscarriage risk, and preserve ovarian reserve in preclinical studies [175]. Additionally, when combined with hormonal therapies, mTOR blockade may help overcome progesterone resistance, thereby enhancing the efficacy of conventional treatments. Emerging evidence also implicates oxidative stress in the activation of kinase cascades within endometriotic lesions. The deubiquitinase cylindromatosis (CYLD), a redox-sensitive negative regulator of NF-κB, has been proposed as a key modulator of inflammation in this context [176]. This highlights a mechanistic link between redox imbalance and sustained inflammatory signaling—an avenue that will be further explored in the section on antioxidant strategies.
Taken together, these findings support the repurposing of kinase inhibitors—originally developed for cancer and autoimmune diseases—as promising non-hormonal therapies in endometriosis. Their ability to target lesion growth, modulate inflammation, and restore reproductive function makes them attractive candidates for fertility-preserving treatment strategies.
4.3. Non-Hormonal Immunomodulation and Immune Checkpoint Targets
Immune dysfunction is a hallmark of endometriosis, characterized by chronic inflammation, impaired immune clearance, and local immune tolerance that permits ectopic tissue survival. In this context, immune checkpoint molecules such as programmed cell death protein 1 (PD-1) and its ligand PD-L1 have emerged as novel targets to restore immune surveillance and enhance endometrial receptivity. Although still in early stages of investigation, modulating PD-1/PD-L1 signaling holds promise for correcting immune escape and dampening lesion-associated inflammation [177,178]. Another promising axis involves the CD47–signal-regulatory protein alpha (SIRPα) pathway, through which ectopic stromal cells evade macrophage-mediated phagocytosis. CD47 is overexpressed in endometriotic lesions and binds to SIRPα on macrophages, delivering a “don’t eat me” signal. Experimental blockade of CD47 has been shown to promote phagocytic clearance and trigger apoptosis in ectopic cells, offering a dual mechanism to reduce lesion viability [179]. Additionally, several non-hormonal immunomodulatory agents are being explored to address the immune imbalance in endometriosis. TNF-α inhibitors such as etanercept and infliximab have demonstrated efficacy in reducing lesion volume and pelvic pain in animal models. Notably, recombinant human TNFRSF1A (r-hTBP1) significantly reduced lesion burden and inflammatory markers in primate models, without suppressing estrogen signaling—an advantage for fertility preservation [180,181,182]. IL-8, a chemokine involved in leukocyte recruitment and tissue remodeling, is also a target of interest. In a Phase 1 trial, the anti-IL-8 antibody AMY109 showed a favorable safety and pharmacokinetic profile in both healthy women and patients with endometriosis. Crucially, it did not disrupt menstrual cycling, suggesting its potential compatibility with fertility goals [183]. However, not all cytokine-targeting interventions yield positive results. For example, interferon-α2b was associated with increased recurrence rates postoperatively and lacked clear immunological benefit, highlighting the need for context-specific, precisely targeted immunotherapies [184].
4.4. Epigenetic Therapies and Non-Coding RNA Modulators
Epigenetic dysregulation is increasingly recognized as a central contributor to progesterone resistance and impaired endometrial receptivity in endometriosis-associated infertility. Key transcriptional regulators such as HOXA10 and the PR-B are frequently silenced via aberrant DNA methylation and histone deacetylation mechanisms. HDACs and DNA methyltransferases have been implicated in this repression, impairing stromal decidualization and progesterone responsiveness [33,185,186,187]. Pharmacological inhibitors of HDACs have demonstrated the ability to reverse these epigenetic marks, restoring transcriptional activity of implantation-related genes and offering a potential therapeutic pathway for patients unresponsive to hormonal therapies. For instance, treatment with the DNA demethylating agent 5-azacytidine (5-Aza) successfully reactivated HOXA10 expression in endometrial stromal cells, validating the role of promoter methylation in gene silencing [188].
Beyond chromatin-modifying enzymes, non-coding RNAs, particularly miRNAs, have emerged as powerful post-transcriptional regulators of gene networks involved in inflammation, hormone signaling, and extracellular matrix remodeling. Dysregulated miRNAs in endometriosis, including let-7b, miR-135a/b, and miR-29c, modulate key pathways relevant to implantation and lesion progression [189]. Therapeutic delivery of miRNA mimics or inhibitors (antagomirs) offers a highly specific strategy to reprogram pathological signaling. For example, inhibition of miR-451a in mouse models resulted in reduced lesion size and downregulation of multiple targets including 14-3-3 protein zeta/delta (YWHAZ), MAPK1, β-catenin (CTNNB1), and IL-6, underscoring its multifaceted action [190]. Similarly, targeting the circ_0007331/miR-200c-3p/HIF-1α axis reduced angiogenesis and lesion invasiveness in vivo [191]. Fibrosis—a defining feature of deep infiltrating endometriosis—can also be modulated through miRNA-based interventions. Exosomal delivery of miR-214 suppressed connective tissue growth factor (CTGF) and collagen type I alpha 1 chain (COL1A1), reducing fibrosis in cell and xenograft models [192]. Moreover, miR-205-5p was found to inhibit stromal invasion by targeting angiopoietin-2 (ANGPT2) and repressing AKT/ERK signaling, with its expression inversely correlating with disease severity in patient samples [193].
4.5. Antioxidant-based Strategies and Redox Modulation
Oxidative stress is increasingly recognized as a key contributor to the reproductive complications observed in endometriosis, including oocyte damage, impaired fertilization, and embryo development arrest. Elevated levels of ROS disrupt mitochondrial function, impair chromosomal alignment, and activate inflammatory pathways like MAPK and NF-κB, aggravating hormonal resistance [94]. In this context, antioxidant supplementation has been explored as a strategy to counteract redox imbalance. Vitamins C and E have been associated with improved oocyte quality and reduced oxidative markers in early studies [194]. Such interventions may be particularly relevant in assisted reproductive technologies (ART), where oxidative stress is amplified. Reviews have highlighted antioxidant potential to improve fertility outcomes in endometriosis patients [94,195]. Coenzyme Q10, by supporting mitochondrial bioenergetics and scavenging ROS, has shown benefits in oocyte maturation and embryo development under oxidative stress competence [194]. Melatonin, another potent antioxidant, improves mitochondrial function and reduces inflammation in preclinical endometriosis models [196,197,198]. N-acetylcysteine (NAC), a glutathione precursor, has demonstrated efficacy in reducing lesion size and inflammation while enhancing oocyte quality and fertilization in IVF cycles [199]. Plant-derived polyphenols exhibit a broad spectrum of bioactive properties, including antioxidant, anti-inflammatory, antiproliferative, and anti-angiogenic effects across various disease contexts, including cancer and reproductive disorders [200,201,202]. Among them, resveratrol has shown promise in improving endometrial receptivity and reducing lesion vascularization, contributing to better implantation outcomes [202,203,204]. Although findings are encouraging, larger clinical trials are needed to confirm the impact of antioxidants on fertility outcomes in endometriosis [205]. Meanwhile, antioxidant therapies may serve as supportive options for patients experiencing oxidative stress–related reproductive impairment.
4.6. Anti-Angiogenic and Vascular Normalization Therapies
Pathological angiogenesis is a hallmark of endometriotic lesion development, supplying ectopic tissue with the vascular support necessary for survival, proliferation, and invasion. These lesions are marked by increased microvascular density, largely driven by overexpression of VEGF and its receptor VEGFR2, which orchestrate neovascularization in the peritoneal environment [206]. Inhibition of the VEGF/VEGFR axis has thus emerged as a promising non-hormonal strategy to suppress lesion progression. In preclinical models, VEGF inhibitors such as aflibercept—a recombinant VEGF-trap that sequesters VEGF-A—have demonstrated significant reductions in lesion size, VEGF expression, and CD31-positive microvessel density, outperforming leuprolide acetate in various histological parameters [207]. In parallel, the bradykinin B1 receptor antagonist R-954 has shown efficacy in reducing VEGF/VEGFR expression, cyst formation, cytokine production, and cellular infiltration in murine models, without disturbing estrous cyclicity, thereby supporting its compatibility with reproductive goals [208,209]. Dopaminergic compounds have also been evaluated for their anti-angiogenic effects. Dopamine receptor agonists such as cabergoline and quinagolide inhibit the VEGF/VEGFR2 cascade, limiting vascular expansion critical for ectopic lesion maintenance. Clinical evidence supports the relevance of this strategy: in women with endometriosis-associated hyperprolactinemia, quinagolide significantly reduced lesion vascularization and total lesion volume [210,211]. Collectively, these findings underscore a therapeutic paradigm shift toward vascular normalization, aimed at selectively impairing pathological angiogenesis while preserving endocrine function and endometrial receptivity.
Such approaches may provide valuable fertility-preserving alternatives to conventional hormonal suppression, particularly for patients actively seeking conception [212]. Table 1 summarizes the most promising candidates currently under investigation, detailing their mechanisms of action, stage of development, and potential impact on fertility outcomes.
While novel molecular approaches offer compelling therapeutic avenues, several obstacles continue to hinder their clinical implementation. Challenges include suboptimal pharmacokinetics, concerns about drug delivery systems, long-term safety and off-target effects [215], especially for RNA-based therapies [216,217,218]. Regulatory frameworks pose further barriers which demand extensive toxicological and immunogenicity profiling before clinical approval [219,220]. Finally, the high development costs combined with persistent underfunding of non-hormonal endometriosis research constrain the pace of innovation and hinder large-scale, fertility-focused clinical trials [221,222]. Bridging these translational gaps will require integrated efforts in research funding, regulatory strategy, and design of trials that prioritize reproductive endpoints.
5. From Risk Reduction to Fertility Preservation: Biomarker-Guided Strategies in Endometriosis Care
Although considerable molecular progress has been made in understanding endometriosis, preventive strategies—particularly those focused on fertility preservation—remain inadequately developed. Integrating biomarkers into the framework of preventive care represents a promising paradigm shift, offering the possibility to identify at-risk individuals, initiate earlier interventions, and optimize reproductive outcomes [223]. To date, no single biomarker has demonstrated sufficient accuracy or specificity to function as a standalone diagnostic or triage tool in clinical settings. Many of the candidate molecules identified thus far lack disease specificity and are also elevated in other inflammatory or physiological conditions, resulting in false positives and limiting their clinical applicability. However, recent advances in high-throughput omics technologies—including proteomics, transcriptomics, and metabolomics—have accelerated the identification of promising biomarker candidates with potential clinical relevance [224]. This section explores the emerging role of such biomarkers across three complementary levels of prevention. By supporting risk stratification before symptom onset (primary prevention), guiding ovarian preservation in early-stage disease (secondary prevention), and personalizing assisted reproductive protocols following surgical intervention (tertiary prevention), these approaches offer a path toward breaking the cycle of disease progression, recurrence, and infertility [225].
5.1. Primary Prevention: Risk Stratification and Early Identification
Primary prevention aims to reduce disease onset in asymptomatic individuals, particularly those with familial or genetic susceptibility. Well-known risk factors include early menarche, short menstrual cycles, nulliparity, and a family history of endometriosis—all contributing to prolonged estrogen exposure and retrograde menstruation [226,227]. Preventive strategies such as regular physical activity, early use of combined oral contraceptives, and minimizing chronic inflammation may attenuate hormonal and immunological drivers during adolescence.
Given the rapid evolution of the field, this section highlights a selection of recent studies—particularly those involving large patient cohorts or rigorous validation frameworks. However, we acknowledge that this is not an exhaustive review, and that numerous other promising genomic, epigenetic, proteomic, and miRNA biomarkers are currently under active investigation. A growing number of studies are seeking optimal combinations of markers with improved specificity and clinical utility, particularly for early diagnosis and patient stratification [228,229]. The coming years are expected to yield a significant expansion in the availability of validated, non-invasive biomarkers for endometriosis. Recent studies suggest that integrating molecular biomarkers into screening tools may allow for stratified risk assessment in asymptomatic individuals— especially first-degree relatives of affected women. Several GWAS have identified single-nucleotide polymorphisms (SNPs) in WNT4, Growth Regulation by Estrogen in Breast Cancer 1 (GREB1), and VEZT—genes implicated in genital tract development, hormonal signaling, cell proliferation, and tissue remodeling [230,231,232,233]. Recent large-scale, multi-ancestry GWAS, supported by transcriptome-wide (TWAS) and proteome-wide association studies (PWAS), have identified over 45 susceptibility loci for endometriosis, including ancestry-specific signals such as POLR2M in African-ancestry populations. These integrative -omics analyses revealed novel genes such as R-spondin 3 (RSPO3) (PWAS) and confirmed others like GREB1, WNT4, and Cell Division Cycle 42 (CDC42) (TWAS), implicating dysregulated Wnt signaling, immune pathways, and altered endometrial cell differentiation and migration [234] These findings provide a genetic and molecular basis for developing polygenic risk scores and early screening strategies tailored to ancestry and molecular subtype.
In parallel, epigenetic alterations—particularly DNA methylation—are under investigation as predictive indicators. HOXA10 promoter hypermethylation, a key regulator of endometrial receptivity, has been consistently found in eutopic and ectopic tissues. Although promising for diagnosis and prognosis, its sensitivity in early disease remains variable [42]. More recently, circulating cell-free DNA (cf-DNA) has emerged as a minimally invasive biomarker. Elevated serum cf-DNA —up to 3.9-fold in women with endometriosis—along with altered methylation patterns in genes such as Ribosomal RNA-Processing Protein 1 (RRP1) ), Disco-Interacting Protein 2 Homolog C (DIPC2), Ubiquitin-Specific Peptidase 1 (USP1), and DNMT1, highlights its potential for early-stage detection [235]. Beyond DNA methylation, circulating miRNAs are gaining attention as early stratification tools. A five-miRNA panel (miR-17-5p, miR-20a-5p, miR-199a-3p, miR-143-3p, and let-7b-5p) distinguished cases from controls with 96% sensitivity and 79% specificity [236], while another six-miRNA signature (miR-125b-5p, miR-150-5p, miR-342-3p, miR-451a, miR-3613-5p, and let-7b) achieved an AUC of 0.94 for early-stage disease detection [237]. Several recent studies also identified additional miRNAs that regulate inflammation, fibrosis, and hormonal signaling, reinforcing their relevance in non-invasive detection [238,239]. Additional miRNAs with diagnostic potential continue to be identified, and future research is expected to refine and expand these panels for broader clinical application.
Proteomic approaches have yielded complementary insights. A large scale study involving over 800 women identified and validated a 10-proteins plasma panel —including Selenoprotein P, Neuropilin-1, Complement C9, and Vitamin K-dependent protein S—that discriminated endometriosis from both healthy and symptomatic controls with an AUC up to 0.997 [240]. These proteins reflect immune dysregulation, oxidative stress and, coagulation anomalies. In a separate prospective cohort, blood samples collected up to nine years before clinical diagnosis showed elevated levels of innate immunity proteins such as S100 Calcium-Binding Protein A9 (S100A9), Intercellular Adhesion Molecule 2 (ICAM2), DNA Topoisomerase I (TOP1), and CD5 Molecule-Like (CD5L), suggesting that systemic immune perturbations may precede clinical manifestation [241]. Proteomics remains an expanding area, with numerous candidate biomarkers currently being validated in longitudinal and multi-cohort studies.
Although traditional inflammatory markers like IL-8 and CA-125 lack adequate sensitivity and specificity for early-stage disease [242], they may gain clinical utility when combined with omics-based signatures [243,244]. Notably, circulating endometrial cells (CECs) have recently demonstrated superior diagnostic performance [245,246]. One study reported an 89.5% detection rate in endometriosis patients—significantly outperforming CA-125 and especially effective in early-stage detection [247]. Moreover, recent findings by Guder et al. demonstrate that elevated circulating Stress-inducible heat shock protein 70 (Hsp70) levels and mHsp70-expressing CECs (which share features with tumor cells) could serve as non-invasive diagnostic markers for endometriosis, particularly in extra-uterine cases, suggesting a pathogenic role of mesenchymal stem-like cells [248].
Beyond molecular diagnostics, mitigating modifiable environmental exposures remains key. Endocrine-disrupting chemicals (EDCs)—including dioxins, phthalates, and BPA—can epigenetically reprogram endometrial tissue, compromising receptivity and implantation [30,249,250,251,252]. Reducing exposure during gestation and adolescence represents a feasible public health strategy to lower long-term disease risk [253,254,255]. Additionally, diets rich in omega-3 fatty acids and antioxidants may help modulate oxidative and inflammatory pathways implicated in disease initiation [213,256,257].
In summary, primary prevention in endometriosis is evolving from generalized health recommendations to biomarker-guided, personalized risk stratification. Combining genomic, epigenetic, and proteomic data into predictive models may enable earlier intervention and improved fertility outcomes in high-risk, asymptomatic populations.
5.2. Secondary Prevention: Halting Disease Progression and Preserving Ovarian Function
Secondary prevention in women already diagnosed with endometriosis focuses on halting disease progression and preserving ovarian reserve before irreversible damage occurs. This approach emphasizes timely interventions guided by molecular biomarkers and personalized strategies to maintain fertility and prevent long-term reproductive sequelae [258,259]. Among the earliest indicators of ovarian compromise is a decline in serum AMH, which may precede reductions in antral follicle count or menstrual irregularity. AMH decline, alongside elevated FSH, has been consistently associated with diminished ovarian reserve in endometriosis patients [259,260,261,262]. More recently, insulin-like peptide 3 (INSL3) has emerged as a complementary marker reflecting ovarian stromal aging and functionality, although clinical validation is ongoing [263].
Epigenetic alterations also provide insight into disease activity and therapeutic targets. Hypermethylation of the HOXA10 gene, pivotal for endometrial receptivity, has been associated with progesterone resistance and aberrant Wnt/β-catenin signaling. This methylation is frequently observed in eutopic but not ectopic tissue, correlating with impaired implantation potential. Therapeutic agents such as Gonadotropin-Releasing Hormone (GnRH) analogs, letrozole, and metformin have been shown to restore HOXA10 expression, positioning it as a dual diagnostic and therapeutic candidate [42].
In parallel, miRNAs offer promising avenues for diagnosis, therapy, and fertility management. They serve as valuable biomarkers for disease activity and severity, with circulating and peritoneal miRNA profiles reflecting inflammatory and angiogenic dynamics [264,265,266]. Beyond their diagnostic utility, certain miRNAs, such as miR-451a, may enable non-hormonal therapeutic strategies targeting multiple pathways [190]. Moreover, recent studies have highlighted the role of miRNAs in fertility, identifying lesion-derived miRNAs in serum extracellular vesicles that may aid in optimizing assisted reproduction timing [267]. Notably, a saliva-based miRNA signature has demonstrated high diagnostic accuracy for infertility in endometriosis, opening the door to non-invasive and personalized fertility assessment [268,269]. While these results are compelling, many other miRNAs with predictive value are currently under investigation. The rapid expansion of miRNA-based studies is expected to yield optimized diagnostic and prognostic panels in the near future.
Oxidative stress is another central factor in oocyte quality and IVF success. Elevated follicular 8-hydroxy-2′-deoxyguanosine (8-OhdG) and imbalanced Glutathione (GSH)/Glutathione disulfide (GSSG) ratios reflect oxidative DNA damage and reduced embryo viability, making antioxidant therapy a rational preventive strategy [140,260,270]. Concurrently, follicular fluid miRNA profiles correlate with oxidative markers and oocyte competence, underscoring their value as integrated prognostic indicators for ART [271].
The endometrial microbiome has emerged as a key modulator of implantation, immune tolerance, and endometrial receptivity [272]. Lactobacillus-dominant profiles (LDM) are associated with higher pregnancy and live birth rates, while non-Lactobacillus-dominant microbiomes—rich in Gardnerella, Prevotella, or Acinetobacter—have been linked to implantation failure and poor endometrial receptivity [273,274]. Dysbiosis in women with repeated implantation failure (RIF) correlates with reduced expression of receptivity markers such as LIF, HOXA11, and VEGF. Recent clinical data support microbiota-targeted interventions in endometriosis. Transvaginal Lactobacillus supplementation has been shown to improve pregnancy rates in women with prior ART failure [275], while Lactobacillus gasseri reduced lesion growth in animal models by enhancing IL-2 production and NK cell responses [276]. A broader review also highlights the contribution of urogenital dysbiosis in sustaining chronic inflammation and promoting lesion recurrence [277,278]. Although current treatments for bacterial dysbiosis remain limited in the clinical practice, emerging strategies such as the use of pro- and prebiotics or microbial transplants, hold considerable promise for modulating the uterine microbiota. Nonetheless, randomized controlled trials are still required to evaluate their clinical efficacy, identify the most beneficial probiotic strains, and clarify their role in ART outcomes and fertility preservation. Future research should also focus on characterizing the composition of the upper female reproductive tract microbiota and elucidating the mechanisms underlying its relationship with endometriosis pathophysiology.
Multi-omic approaches continue to offer critical insights for individualized care. While discussed in detail in Section 5.1, their integration at this stage allows dynamic monitoring of biological activity and early detection of therapeutic windows [279]. This supports the move toward precision-based monitoring and fertility-preserving interventions in women with active disease. In summary, secondary prevention in endometriosis is moving toward a precision-medicine framework, integrating molecular, epigenetic, and microbial biomarkers to guide early therapeutic interventions. By stratifying patients based on biological activity rather than symptoms alone, clinicians can implement fertility-preserving strategies before significant ovarian compromise occurs—shifting the paradigm from reactive to proactive care.
5.3. Tertiary Prevention: Preventing Recurrence and Individualizing Long-Term Fertility Planning
Tertiary prevention in endometriosis focuses on reducing recurrence risk and supporting long-term fertility planning following surgical or medical management. Given the chronic and often relapsing nature of the disease, maintenance therapy is critical for sustaining remission, optimizing reproductive outcomes, and preserving quality of life [258]. Hormonal maintenance, using combined oral contraceptives, progestins, or levonorgestrel-releasing intrauterine systems (LNG-IUS), remains the cornerstone post-surgery and is effective in reducing recurrence and preserving ovarian function [280,281,282,283,284]. These agents suppress ovulation and cyclic estrogen fluctuations, reducing inflammation and the potential for lesion reactivation. For patients with ovarian endometriomas, long-term hormonal suppression also mitigates the risks of inflammation-induced follicular damage and surgical insult to the ovarian cortex. Current strategies advocate for individualized regimens based on clinical phenotype, reproductive goals, and safety considerations, including the choice of hormone type and route of administration. Incorporating emerging molecular markers of disease activity into follow-up may further enhance early detection of recurrence and timely intervention [29,166]. Molecular diagnostics—such as miRNA panels, angiogenic markers like VEGF-A and miR-486-5p, and inflammatory mediators (IL-6, IL-8)—may allow for more precise risk stratification and longitudinal monitoring of residual or reactivating disease [265].
Fertility preservation remains a core component of tertiary prevention, particularly in women delaying conception. Cryopreservation of oocytes or embryos is increasingly recommended before repeat surgery, especially in cases of declining AMH, bilateral ovarian involvement, or diminished reserve [285,286,287,288]. Longitudinal assessment of AMH, Insulin-Like Peptide 3 (INSL3), and follicular fluid biomarkers (e.g., oxidative stress markers, follicular miRNAs) can help determine optimal timing for ART or preservation procedures [258,263,289]. In addition to hormonal and surgical considerations, patient education and clinician awareness are essential to maintaining reproductive health. Reducing diagnostic delay and promoting early initiation of maintenance therapy may have profound impacts on long-term outcomes. Multidisciplinary care models that incorporate reproductive endocrinologists, pain specialists, and fertility counselors are increasingly advocated.
In essence, tertiary prevention marks a transition from episodic treatment to a continuum of long-term, patient-centered care, integrating maintenance therapy, biomarker surveillance, reproductive counseling, and personalized follow-up to break the cycle of recurrence and protect reproductive health.
6. Conclusions and Perspectives
Endometriosis epitomizes a multifactorial and systemic disorder that transcends pelvic confinement, rooted in the complex interplay of hormonal dysregulation, chronic inflammation, epigenetic reprogramming, oxidative stress, immune dysfunction, and metabolic remodeling. Far beyond its physical manifestations, the disease disrupts the molecular architecture of both eutopic and ectopic tissues, compromising ovarian reserve, endometrial receptivity, embryo viability, and maternal–fetal immunotolerance. The present review underscores how estrogen dominance, progesterone resistance, and aberrant activation of signaling pathways (e.g., PI3K/AKT, MAPK) jointly sustain lesion survival, angiogenesis, and fibrotic remodeling, while also perturbing fertility-related processes at multiple levels. Importantly, infertility in endometriosis is no longer viewed as a mere consequence of anatomical distortion but is increasingly recognized as the result of deeply embedded molecular dysfunctions affecting gametogenesis, implantation, and systemic homeostasis.
Advances in high-throughput technologies have unveiled a wide spectrum of therapeutic targets—including ERβ modulation, inflammatory kinase inhibition, epigenetic therapies, immune checkpoint regulators, miRNA-based strategies, and redox modulators—that collectively pave the way for personalized, fertility-preserving interventions. Several of these approaches, although still in the preclinical or early clinical phase, demonstrate tangible potential to move beyond hormone suppression, offering non-hormonal avenues for patients desiring conception. Simultaneously, the emergence of circulating biomarkers, polygenic risk scores, cfDNA methylation patterns, and proteomic signatures has begun to redefine the landscape of early detection and disease monitoring. This molecular arsenal supports the integration of stratified prevention into clinical care—from risk reduction in asymptomatic individuals (primary prevention), through fertility-preserving interventions in early disease (secondary prevention), to recurrence surveillance and reproductive planning after treatment (tertiary prevention). Taken together, these insights advocate for a paradigm shift toward translational precision medicine in endometriosis, where diagnostic, therapeutic, and preventive strategies are molecularly guided and patient centered. To achieve this shift, clinical implementation will require the development of supportive infrastructure, including the integration of molecular diagnostics and longitudinal biomarker monitoring into assisted reproduction and gynecology care settings. Bridging fundamental mechanisms with clinical practice will not only improve reproductive outcomes but also transform endometriosis care into a proactive, multidisciplinary, and personalized continuum.
Author Contributions
All authors—Inès Limam, Mohamed Abdelkarim, Khadija Kacem, Mohamed Khrouf, Anis Feki, Marouen Braham and Nozha Chakroun—contributed to the conceptualization, methodology, investigation, resources, original draft preparation, review and editing, visualization, supervision, project administration, and funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Aziza Othmena Hospital and the Tunisian Ministry of Higher Education and Scientific Research.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
The authors would like to thank all those who provided administrative and technical support during the preparation of this manuscript. During the preparation of this manuscript, the authors used ChatGPT (OpenAI, GPT-4, 2024) for assistance with text refinement and consistency. The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript.
| 17β-HSD2 | 17β-Hydroxysteroid Dehydrogenase Type 2 |
| 8-OHdG | 8-Hydroxy-2′-Deoxyguanosine |
| ABCA1 | ATP-Binding Cassette Transporter A1 |
| ABCG1 | ATP-Binding Cassette Transporter G1 |
| AKR1C3 | Aldo-Keto Reductase Family 1 Member C3 |
| AKT | Protein Kinase B |
| AMH | Anti-Müllerian Hormone |
| ANGPT2 | Angiopoietin-2 |
| ASK1 | Apoptosis Signal-Regulating Kinase 1 |
| AT1R | Angiotensin II Type 1 Receptor |
| BPA | Bisphenol A |
| C5aR1 | Complement C5a Receptor 1 |
| CD5L | CD5 Molecule-Like |
| CDKN2B-AS1 | Cyclin-Dependent Kinase Inhibitor 2B Antisense RNA 1 |
| CDC42 | Cell Division Cycle 42 |
| ceRNA | Competing Endogenous RNA |
| CECs | Circulating Endometrial Cells |
| circRNA | Circular RNA |
| COL1A1 | Collagen Type I Alpha 1 Chain |
| COX-2 | Cyclo-Oxygenase-2 |
| CTGF | Connective Tissue Growth Factor |
| CTNNB1 | Catenin Beta 1 (β-Catenin) |
| CYLD | Cylindromatosis (Deubiquitinase) |
| CYP19A1 | Cytochrome P450 19A1 |
| DIPC2 | Disco-Interacting Protein 2 Homolog C |
| Dll4 | Delta-Like Ligand 4 |
| DNMT1 | DNA (Cytosine-5) Methyltransferase 1 |
| EDC | Endocrine-Disrupting Chemical |
| ER | Estrogen Receptor (generic) |
| Erα | Estrogen Receptor Alpha |
| ERβ | Estrogen Receptor Beta |
| ERK/ERK1/2 | Extracellular Signal-Regulated Kinase 1/2 |
| ESC | Endometrial Stromal Cell |
| FOXP3⁺ | Forkhead Box P3-Positive Regulatory T Cell |
| FSHR | Follicle-Stimulating Hormone Receptor |
| GnRH | Gonadotropin-Releasing Hormone |
| GREB1 | Growth Regulation by Estrogen in Breast Cancer 1 |
| GPER | G-Protein-Coupled Estrogen Receptor |
| GPR30 | G-Protein-Coupled Receptor 30 |
| GSH | Glutathione (reduced) |
| GSSG | Glutathione Disulfide (oxidized) |
| GWAS | Genome-Wide Association Study |
| HIF-1α | Hypoxia-Inducible Factor-1 Alpha |
| HOXA10 | Homeobox A10 |
| ICAM2 | Intercellular Adhesion Molecule 2 |
| IL | Interleukin |
| INSL3 | Insulin-Like Peptide 3 |
| IVF | In Vitro Fertilization |
| JNK | c-Jun N-Terminal Kinase |
| LIF | Leukemia Inhibitory Factor |
| LNG-IUS | Levonorgestrel-Releasing Intrauterine System |
| lncRNA | Long Non-Coding RNA |
| LOX | Lysyl Oxidase |
| LXR | Liver X Receptor |
| MAPK | Mitogen-Activated Protein Kinase |
| MAPK1 | Mitogen-Activated Protein Kinase 1 |
| MEK | Mitogen-Activated Protein Kinase Kinase |
| miRNA | MicroRNA |
| mTOR | Mammalian Target of Rapamycin |
| MMP | Matrix Metalloproteinase |
| NAC | N-Acetyl-L-Cysteine |
| NLRP3 | NACHT, LRR and PYD Domains-Containing Protein 3 |
| NGF | Nerve Growth Factor |
| NF-κB | Nuclear Factor κ-Light-Chain-Enhancer of Activated B Cells |
| NK | Natural Killer (Cell) |
| PAR-2 | Protease-Activated Receptor 2 |
| PCB | Polychlorinated Biphenyl |
| PCNA | Proliferating Cell Nuclear Antigen |
| PD-1 | Programmed Cell Death Protein 1 |
| PD-L1 | Programmed Death-Ligand 1 |
| PI3K | Phosphoinositide 3-Kinase |
| PPAR | Peroxisome Proliferator-Activated Receptor |
| PPAR-α | Peroxisome Proliferator-Activated Receptor Alpha |
| PR-B | Progesterone Receptor Isoform B |
| PROK1 | Prokineticin-1 |
| PROKR1 | Prokineticin Receptor 1 |
| PrPC | Cellular Prion Protein |
| PWAS | Proteome-Wide Association Study |
| r-hTBP1 | Recombinant Human TNFRSF1A |
| ROS | Reactive Oxygen Species |
| RRP1 | Ribosomal RNA-Processing Protein 1 |
| RSPO3 | R-Spondin 3 |
| RUNX3 | Runt-Related Transcription Factor 3 |
| S100A9 | S100 Calcium-Binding Protein A9 |
| SARM | Selective Androgen Receptor Modulator |
| SARD | Selective Androgen Receptor Degrader |
| SERM | Selective Estrogen Receptor Modulator |
| SF-1 | Steroidogenic Factor 1 |
| SIRPα | Signal-Regulatory Protein Alpha |
| Slit2 | Slit Guidance Ligand 2 |
| SMAD | (S)-Homologues of Mothers Against Decapentaplegic |
| SNAIL | Snail Family Transcriptional Repressor 1 |
| SNP | Single-Nucleotide Polymorphism |
| STAT3 | Signal Transducer and Activator of Transcription 3 |
| STRAP | Serine/Threonine Kinase Receptor-Associated Protein |
| TET | Ten-Eleven Translocation Dioxygenase |
| TET2 | Ten-Eleven Translocation Methylcytosine Dioxygenase 2 |
| TGF-β | Transforming Growth Factor Beta |
| TIMPs | Tissue Inhibitors of Metalloproteinases |
| TOP1 | DNA Topoisomerase I |
| TNF-α | Tumor Necrosis Factor Alpha |
| Tregs | Regulatory T Cells |
| TWAS | Transcriptome-Wide Association Study |
| uNK | Uterine Natural Killer Cell |
| uPA | Urokinase-Type Plasminogen Activator |
| uPAR | Urokinase-Type Plasminogen Activator Receptor |
| USP1 | Ubiquitin-Specific Peptidase 1 |
| VEGF | Vascular Endothelial Growth Factor |
| VEZT | Vezatin Adherens Junctions Transmembrane Protein |
| WDR5 | WD Repeat Domain 5 |
| Wnt | Wingless-Related Integration Site (Wnt) Family |
| WNT4 | Wnt Family Member 4 |
| YWHAZ | 14-3-3 Protein Zeta/Delta |
| ZEB1/2 | Zinc Finger E-Box-Binding Homeobox 1 & 2 |
References
- Giudice, L.C. Endometriosis. New England Journal of Medicine 2010, 362, 2389–2398. [Google Scholar] [CrossRef] [PubMed]
- Zondervan, K.T.; Becker, C.M.; Missmer, S.A. Endometriosis. New England Journal of Medicine 2020, 382, 1244–1256. [Google Scholar] [CrossRef]
- Parasar, P.; Ozcan, P.; Terry, K.L. Endometriosis: Epidemiology, Diagnosis and Clinical Management. Curr Obstet Gynecol Rep 2017, 6, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Bulun, S.E. Endometriosis. New England Journal of Medicine 2009, 360, 268–279. [Google Scholar] [CrossRef]
- Vercellini, P.; Viganò, P.; Somigliana, E.; Fedele, L. Endometriosis: Pathogenesis and Treatment. Nat Rev Endocrinol 2014, 10, 261–275. [Google Scholar] [CrossRef] [PubMed]
- Psilopatis, I.; Burghaus, S.; Au, K.; Hofbeck, L.; Windischbauer, L.; Lotz, L.; Beckmann, M.W. The Hallmarks of Endometriosis. Geburtshilfe Frauenheilkd 2024, 84, 555–563. [Google Scholar] [CrossRef]
- Amanda, C.R.; Asmarinah; Hestiantoro, A.; Tulandi, T. Febriyeni Gene Expression of Aromatase, SF-1, and HSD17B2 in Menstrual Blood as Noninvasive Diagnostic Biomarkers for Endometriosis. European Journal of Obstetrics & Gynecology and Reproductive Biology 2024, 301, 95–101. [Google Scholar] [CrossRef]
- Bulun, S.E.; Fang, Z.; Imir, G.; Gurates, B.; Tamura, M.; Yilmaz, B.; Langoi, D.; Amin, S.; Yang, S.; Deb, S. Aromatase and Endometriosis. Seminars in Reproductive Medicine 2004, 22, 45–50. [Google Scholar] [CrossRef]
- Chantalat, E.; Valera, M.-C.; Vaysse, C.; Noirrit, E.; Rusidze, M.; Weyl, A.; Vergriete, K.; Buscail, E.; Lluel, P.; Fontaine, C.; et al. Estrogen Receptors and Endometriosis. International Journal of Molecular Sciences 2020, 21, 2815. [Google Scholar] [CrossRef]
- Rižner, T.L.; Gjorgoska, M. Steroid Sulfatase and Sulfotransferases in the Estrogen and Androgen Action of Gynecological Cancers: Current Status and Perspectives. Essays in Biochemistry 2024, 68, 411–422. [Google Scholar] [CrossRef]
- Da Costa, K. de A.; Malvezzi, H.; Dobo, C.; Neme, R.M.; Filippi, R.Z.; Aloia, T.P.A.; Prado, E.R.; Meola, J.; Piccinato, C. de A. Site-Specific Regulation of Sulfatase and Aromatase Pathways for Estrogen Production in Endometriosis. Front. Mol. Biosci. 2022, 9. [Google Scholar] [CrossRef]
- Secky, L.; Svoboda, M.; Klameth, L.; Bajna, E.; Hamilton, G.; Zeillinger, R.; Jäger, W.; Thalhammer, T. The Sulfatase Pathway for Estrogen Formation: Targets for the Treatment and Diagnosis of Hormone-Associated Tumors. Journal of Drug Delivery 2013, 2013, 1–13. [Google Scholar] [CrossRef]
- Qi, Q.; Li, Y.; Chen, Z.; Luo, Z.; Zhou, T.; Zhou, J.; Zhang, Y.; Chen, S.; Wang, L. Update on the Pathogenesis of Endometriosis-Related Infertility Based on Contemporary Evidence. Front. Endocrinol. 2025, 16. [Google Scholar] [CrossRef] [PubMed]
- Stevens Brentjens, L.B.P.M.; Delvoux, B.; den Hartog, J.E.; Obukhova, D.; Xanthoulea, S.; Romano, A.; van Golde, R.J.T. Endometrial Metabolism of 17β-Estradiol during the Window of Implantation in Women with Recurrent Implantation Failure. Gynecologic and Obstetric Investigation 2025. [CrossRef] [PubMed]
- Kobayashi, H.; Kimura, M.; Maruyama, S.; Nagayasu, M.; Imanaka, S. Revisiting Estrogen-Dependent Signaling Pathways in Endometriosis: Potential Targets for Non-Hormonal Therapeutics. European Journal of Obstetrics & Gynecology and Reproductive Biology 2021, 258, 103–110. [Google Scholar] [CrossRef]
- Laganà, A.S.; Garzon, S.; Götte, M.; Viganò, P.; Franchi, M.; Ghezzi, F.; Martin, D.C. The Pathogenesis of Endometriosis: Molecular and Cell Biology Insights. International Journal of Molecular Sciences 2019, 20, 5615. [Google Scholar] [CrossRef]
- Hu, L.; Zhang, J.; Lu, Y.; Fu, B.; Hu, W. Estrogen Receptor Beta Promotes Endometriosis Progression by Upregulating CD47 Expression in Ectopic Endometrial Stromal Cells. Journal of Reproductive Immunology 2022, 151, 103513. [Google Scholar] [CrossRef] [PubMed]
- Gou, Y.; Li, X.; Li, P.; Zhang, H.; Xu, T.; Wang, H.; Wang, B.; Ma, X.; Jiang, X.; Zhang, Z. Estrogen Receptor β Upregulates CCL2 via NF-κB Signaling in Endometriotic Stromal Cells and Recruits Macrophages to Promote the Pathogenesis of Endometriosis. Hum Reprod 2019, 34, 646–658. [Google Scholar] [CrossRef]
- Szukiewicz, D. Insight into the Potential Mechanisms of Endocrine Disruption by Dietary Phytoestrogens in the Context of the Etiopathogenesis of Endometriosis. Int J Mol Sci 2023, 24, 12195. [Google Scholar] [CrossRef]
- Choi, J.; Jo, M.; Lee, E.; Kim, S.E.; Lee, D.-Y.; Choi, D. Inhibition of the NLRP3 Inflammasome by Progesterone Is Attenuated by Abnormal Autophagy Induction in Endometriotic Cyst Stromal Cells: Implications for Endometriosis. Molecular Human Reproduction 2022, 28, gaac007. [Google Scholar] [CrossRef]
- Irandoost, E.; Najibi, S.; Talebbeigi, S.; Nassiri, S. Focus on the Role of NLRP3 Inflammasome in the Pathology of Endometriosis: A Review on Molecular Mechanisms and Possible Medical Applications. Naunyn-Schmiedeberg’s Arch Pharmacol 2023, 396, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.-G.; Wu, X.-X.; Hua, T.; Xin, X.-Y.; Feng, D.-L.; Chi, S.-Q.; Wang, X.-X.; Wang, H.-B. NLRP3 Inflammasome Activation by Estrogen Promotes the Progression of Human Endometrial Cancer. Onco Targets Ther 2019, 12, 6927–6936. [Google Scholar] [CrossRef]
- Leavy, O. Evading Immunosurveillance in Endometriosis. Nat Rev Immunol 2015, 15, 729–729. [Google Scholar] [CrossRef] [PubMed]
- García-Gómez, E.; Vázquez-Martínez, E.R.; Reyes-Mayoral, C.; Cruz-Orozco, O.P.; Camacho-Arroyo, I.; Cerbón, M. Regulation of Inflammation Pathways and Inflammasome by Sex Steroid Hormones in Endometriosis. Front. Endocrinol. 2020, 10. [Google Scholar] [CrossRef]
- Simmen, R.C.M.; Kelley, A.S. Reversal of Fortune: Estrogen Receptor-β in Endometriosis. J Mol Endocrinol 2016, 57, F23–27. [Google Scholar] [CrossRef]
- Azuma, K.; Inoue, S. Genomic and Non-Genomic Actions of Estrogen: Recent Developments. BioMolecular Concepts 2012, 3, 365–370. [Google Scholar] [CrossRef]
- Greygoose, E.; Metharom, P.; Kula, H.; Seckin, T.K.; Seckin, T.A.; Ayhan, A.; Yu, Y. The Estrogen–Immune Interface in Endometriosis. Cells 2025, 14, 58. [Google Scholar] [CrossRef]
- Lin, B.C.; Suzawa, M.; Blind, R.D.; Tobias, S.C.; Bulun, S.E.; Scanlan, T.S.; Ingraham, H.A. Stimulating the GPR30 Estrogen Receptor with a Novel Tamoxifen Analogue Activates SF-1 and Promotes Endometrial Cell Proliferation. Cancer Res 2009, 69, 5415–5423. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Xu, T.; Tong, D.; Li, S.; Yu, X.; Liu, B.; Jiang, L.; Liu, K. Research Advances in Endometriosis-Related Signaling Pathways: A Review. Biomed Pharmacother 2023, 164, 114909. [Google Scholar] [CrossRef]
- Wu, Y.; Strawn, E.; Basir, Z.; Halverson, G.; Guo, S.-W. Promoter Hypermethylation of Progesterone Receptor Isoform B (PR-B) in Endometriosis. Epigenetics 2006, 1, 106–111. [Google Scholar] [CrossRef]
- Colón-Díaz, M.; Báez-Vega, P.; García, M.; Ruiz, A.; Monteiro, J.B.; Fourquet, J.; Bayona, M.; Alvarez-Garriga, C.; Achille, A.; Seto, E.; et al. HDAC1 and HDAC2 Are Differentially Expressed in Endometriosis. Reprod Sci 2012, 19, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Szukiewicz, D. Aberrant Epigenetic Regulation of Estrogen and Progesterone Signaling at the Level of Endometrial/Endometriotic Tissue in the Pathomechanism of Endometriosis. Vitam Horm 2023, 122, 193–235. [Google Scholar] [CrossRef]
- Promoter Hypermethylation of Progesterone Receptor Isoform B (PR-B) in Adenomyosis and Its Rectification by a Histone Deacetylase Inhibitor and a Demethylation Agent. Reprod Sci 2010, 17, 995–1005. [CrossRef]
- Wang, D.; Guo, C.; Li, Y.; Zhou, M.; Wang, H.; Liu, J.; Chen, P. Oestrogen Up-Regulates DNMT1 and Leads to the Hypermethylation of RUNX3 in the Malignant Transformation of Ovarian Endometriosis. Reprod Biomed Online 2022, 44, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.; Yao, X.; Ting, G.; Ling, J.; Huimin, L.; Yuan, Q.; Chun, Z.; Ming, Z.; Yuanzhen, Z. BPA Modulates the WDR5/TET2 Complex to Regulate ERβ Expression in Eutopic Endometrium and Drives the Development of Endometriosis. Environmental Pollution 2021, 268, 115748. [Google Scholar] [CrossRef] [PubMed]
- Ding, N.-Z.; Wang, X.-M.; Jiao, X.-W.; Li, R.; Zeng, C.; Li, S.-N.; Guo, H.-S.; Wang, Z.-Y.; Huang, Z.; He, C.-Q. Cellular Prion Protein Is Involved in Decidualization of Mouse Uterus. Biol Reprod 2018, 99, 319–325. [Google Scholar] [CrossRef]
- Peng, H.-Y.; Lei, S.-T.; Hou, S.-H.; Weng, L.-C.; Yuan, Q.; Li, M.-Q.; Zhao, D. PrPC Promotes Endometriosis Progression by Reprogramming Cholesterol Metabolism and Estrogen Biosynthesis of Endometrial Stromal Cells through PPARα Pathway. Int J Biol Sci 2022, 18, 1755–1772. [Google Scholar] [CrossRef]
- Psilopatis, I.; Theocharis, S.; Beckmann, M.W. The Role of Peroxisome Proliferator-Activated Receptors in Endometriosis. Front. Med. 2024, 11, 1329406. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, Y.; Gu, P.; Tuo, L.; Wang, L.; Jiang, S.-W. Transgenic Mice Applications in the Study of Endometriosis Pathogenesis. Front. Cell Dev. Biol. 2024, 12, 1376414. [Google Scholar] [CrossRef]
- Ducreux, B.; Patrat, C.; Firmin, J.; Ferreux, L.; Chapron, C.; Marcellin, L.; Parpex, G.; Bourdon, M.; Vaiman, D.; Santulli, P.; et al. Systematic Review on the DNA Methylation Role in Endometriosis: Current Evidence and Perspectives. Clin Epigenetics 2025, 17, 32. [Google Scholar] [CrossRef]
- Wu, Y.; Halverson, G.; Basir, Z.; Strawn, E.; Yan, P.; Guo, S.-W. Aberrant Methylation at HOXA10 May Be Responsible for Its Aberrant Expression in the Endometrium of Patients with Endometriosis. American Journal of Obstetrics and Gynecology 2005, 193, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Elias, M.H.; Lazim, N.; Sutaji, Z.; Abu, M.A.; Abdul Karim, A.K.; Ugusman, A.; Syafruddin, S.E.; Mokhtar, M.H.; Ahmad, M.F. HOXA10 DNA Methylation Level in the Endometrium Women with Endometriosis: A Systematic Review. Biology (Basel) 2023, 12, 474. [Google Scholar] [CrossRef]
- Pîrlog, L.-M.; Pătrășcanu, A.-A.; Ona, M.-D.; Cătană, A.; Rotar, I.C. HOXA10 and HOXA11 in Human Endometrial Benign Disorders: Unraveling Molecular Pathways and Their Impact on Reproduction. Biomolecules 2025, 15, 563. [Google Scholar] [CrossRef] [PubMed]
- Petracco, R.; Grechukhina, O.; Popkhadze, S.; Massasa, E.; Zhou, Y.; Taylor, H.S. MicroRNA 135 Regulates HOXA10 Expression in Endometriosis. The Journal of Clinical Endocrinology & Metabolism 2011, 96, E1925–E1933. [Google Scholar] [CrossRef]
- Mirabutalebi, S.H.; Karami, N.; Montazeri, F.; Fesahat, F.; Sheikhha, M.H.; Hajimaqsoodi, E.; Karimi Zarchi, M.; Kalantar, S.M. The Relationship between the Expression Levels of miR-135a and HOXA10 Gene in the Eutopic and Ectopic Endometrium. Int J Reprod Biomed 2018, 16, 501–506. [Google Scholar] [CrossRef]
- Zhou, T.; Ni, T.; Li, Y.; Zhang, Q.; Yan, J.; Chen, Z.-J. circFAM120A Participates in Repeated Implantation Failure by Regulating Decidualization via the miR-29/ABHD5 Axis. FASEB J 2021, 35, e21872. [Google Scholar] [CrossRef] [PubMed]
- Long, M.; Wan, X.; La, X.; Gong, X.; Cai, X. miR-29c Is Downregulated in the Ectopic Endometrium and Exerts Its Effects on Endometrial Cell Proliferation, Apoptosis and Invasion by Targeting c-Jun. International Journal of Molecular Medicine 2015, 35, 1119–1125. [Google Scholar] [CrossRef]
- Pei, T.; Liu, C.; Liu, T.; Xiao, L.; Luo, B.; Tan, J.; Li, X.; Zhou, G.; Duan, C.; Huang, W. miR-194-3p Represses the Progesterone Receptor and Decidualization in Eutopic Endometrium From Women With Endometriosis. Endocrinology 2018, 159, 2554–2562. [Google Scholar] [CrossRef]
- Zhao, Q.; Han, B.; Zhang, Y.; Su, K.; Wang, C.; Hai, P.; Bian, A.; Guo, R. Effect of miR-194-5p Regulating STAT1/mTOR Signaling Pathway on the Biological Characteristics of Ectopic Endometrial Cells from Mice. Am J Transl Res 2020, 12, 6136–6148. [Google Scholar]
- Wu, Y.; Lin, Y.; Li, S.; Yao, X.; Cheng, M.; Zhu, L.; Liu, H. microRNA-194 Is Increased in Polycystic Ovary Syndrome Granulosa Cell and Induce KGN Cells Apoptosis by Direct Targeting Heparin-Binding EGF-like Growth Factor. Reprod Biol Endocrinol 2021, 19, 170. [Google Scholar] [CrossRef]
- Choi, M.; Chang, H.; Heo, J.-H.; Yum, S.; Jo, E.; Kim, M.; Lee, S.-R. Expression Profiling of Coding and Noncoding RNAs in the Endometrium of Patients with Endometriosis. International Journal of Molecular Sciences 2024, 25, 10581. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Xin, W.; Lu, Q.; Tang, X.; Wang, F.; Shao, W.; Zhang, Y.; Qiu, J.; Hua, K. Knockdown of lncRNA H19 Suppresses Endometriosis in Vivo. Braz J Med Biol Res 2021, 54, e10117. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Xin, W.; Tang, X.; Qiu, J.; Zhang, Y.; Hua, K. LncRNA H19 Overexpression in Endometriosis and Its Utility as a Novel Biomarker for Predicting Recurrence. Reprod Sci 2020, 27, 1687–1697. [Google Scholar] [CrossRef]
- Hudson, Q.J.; Proestling, K.; Perricos, A.; Kuessel, L.; Husslein, H.; Wenzl, R.; Yotova, I. The Role of Long Non-Coding RNAs in Endometriosis. Int J Mol Sci 2021, 22, 11425. [Google Scholar] [CrossRef]
- Zhao, L.; Gu, C.; Ye, M.; Zhang, Z.; Li, L.; Fan, W.; Meng, Y. Integration Analysis of microRNA and mRNA Paired Expression Profiling Identifies Deregulated microRNA-Transcription Factor-Gene Regulatory Networks in Ovarian Endometriosis. Reprod Biol Endocrinol 2018, 16, 4. [Google Scholar] [CrossRef]
- Dana, P.M.; Taghavipour, M.; Mirzaei, H.; Yousefi, B.; Moazzami, B.; Chaichian, S.; Asemi, Z. Circular RNA as a Potential Diagnostic and/or Therapeutic Target for Endometriosis. Biomark Med 2020, 14, 1277–1287. [Google Scholar] [CrossRef]
- Matsuzaki, S.; Darcha, C. Co-Operation between the AKT and ERK Signaling Pathways May Support Growth of Deep Endometriosis in a Fibrotic Microenvironment in Vitro. Hum Reprod 2015, 30, 1606–1616. [Google Scholar] [CrossRef]
- Cinar, O.; Seval, Y.; Uz, Y.H.; Cakmak, H.; Ulukus, M.; Kayisli, U.A.; Arici, A. Differential Regulation of Akt Phosphorylation in Endometriosis. Reproductive BioMedicine Online 2009, 19, 864–871. [Google Scholar] [CrossRef] [PubMed]
- An, M.; Fu, X.; Meng, X.; Liu, H.; Ma, Y.; Li, Y.; Li, Q.; Chen, J. PI3K/AKT Signaling Pathway Associates with Pyroptosis and Inflammation in Patients with Endometriosis. Journal of Reproductive Immunology 2024, 162, 104213. [Google Scholar] [CrossRef]
- Matsuzaki, S.; Darcha, C. Involvement of the Wnt/β-Catenin Signaling Pathway in the Cellular and Molecular Mechanisms of Fibrosis in Endometriosis. PLoS One 2013, 8, e76808. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, X.; Li, Z.; Han, X.; Wang, W.; Xu, P.; Liu, Y.; Xue, Y.; Wang, Z.; Xu, S.; et al. Interactions between miRNAs and the Wnt/β-Catenin Signaling Pathway in Endometriosis. Biomedicine & Pharmacotherapy 2024, 171, 116182. [Google Scholar] [CrossRef]
- Driva, T.S.; Schatz, C.; Sobočan, M.; Haybaeck, J. The Role of mTOR and eIF Signaling in Benign Endometrial Diseases. Int J Mol Sci 2022, 23, 3416. [Google Scholar] [CrossRef]
- Assaf, L.; Eid, A.A.; Nassif, J. Role of AMPK/mTOR, Mitochondria, and ROS in the Pathogenesis of Endometriosis. Life Sciences 2022, 306, 120805. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Liu, Y.; Zhong, Z.; Wei, C.; Liu, Y.; Zhu, X. Peritoneal Immune Microenvironment of Endometriosis: Role and Therapeutic Perspectives. Front. Immunol. 2023, 14. [Google Scholar] [CrossRef]
- Eaton, J.L.; Unno, K.; Caraveo, M.; Lu, Z.; Kim, J.J. Increased AKT or MEK1/2 Activity Influences Progesterone Receptor Levels and Localization in Endometriosis. J Clin Endocrinol Metab 2013, 98, E1871–1879. [Google Scholar] [CrossRef]
- Ruan, J.; Tian, Q.; Li, S.; Zhou, X.; Sun, Q.; Wang, Y.; Xiao, Y.; Li, M.; Chang, K.; Yi, X. The IL-33-ST2 Axis Plays a Vital Role in Endometriosis via Promoting Epithelial–Mesenchymal Transition by Phosphorylating β-Catenin. Cell Commun Signal 2024, 22, 318. [Google Scholar] [CrossRef]
- Hui San, S.; Ching Ngai, S. E-Cadherin Re-Expression: Its Potential in Combating TRAIL Resistance and Reversing Epithelial-to-Mesenchymal Transition. Gene 2024, 909, 148293. [Google Scholar] [CrossRef] [PubMed]
- Capobianco, A.; Rovere-Querini, P. Endometriosis, a Disease of the Macrophage. Front Immunol 2013, 4, 9. [Google Scholar] [CrossRef]
- Schulke, L.; Berbic, M.; Manconi, F.; Tokushige, N.; Markham, R.; Fraser, I.S. Dendritic Cell Populations in the Eutopic and Ectopic Endometrium of Women with Endometriosis. Hum Reprod 2009, 24, 1695–1703. [Google Scholar] [CrossRef]
- Scheerer, C.; Bauer, P.; Chiantera, V.; Sehouli, J.; Kaufmann, A.; Mechsner, S. Characterization of Endometriosis-Associated Immune Cell Infiltrates (EMaICI). Arch Gynecol Obstet 2016, 294, 657–664. [Google Scholar] [CrossRef]
- Henlon, Y.; Panir, K.; McIntyre, I.; Hogg, C.; Dhami, P.; Cuff, A.O.; Senior, A.; Moolchandani-Adwani, N.; Courtois, E.T.; Horne, A.W.; et al. Single-Cell Analysis Identifies Distinct Macrophage Phenotypes Associated with Prodisease and Proresolving Functions in the Endometriotic Niche. Proc. Natl. Acad. Sci. U.S.A. 2024, 121, e2405474121. [Google Scholar] [CrossRef] [PubMed]
- Iwabe, T.; Harada, T.; Tsudo, T.; Nagano, Y.; Yoshida, S.; Tanikawa, M.; Terakawa, N. Tumor Necrosis Factor-Alpha Promotes Proliferation of Endometriotic Stromal Cells by Inducing Interleukin-8 Gene and Protein Expression. J Clin Endocrinol Metab 2000, 85, 824–829. [Google Scholar] [CrossRef] [PubMed]
- Oală, I.E.; Mitranovici, M.-I.; Chiorean, D.M.; Irimia, T.; Crișan, A.I.; Melinte, I.M.; Cotruș, T.; Tudorache, V.; Moraru, L.; Moraru, R.; et al. Endometriosis and the Role of Pro-Inflammatory and Anti-Inflammatory Cytokines in Pathophysiology: A Narrative Review of the Literature. Diagnostics (Basel) 2024, 14, 312. [Google Scholar] [CrossRef] [PubMed]
- Kacem-Berjeb, K.; Braham, M.; Massoud, C.B.; Hannachi, H.; Hamdoun, M.; Chtourou, S.; Debbabi, L.; Bouyahia, M.; Fadhlaoui, A.; Zhioua, F.; et al. Does Endometriosis Impact the Composition of Follicular Fluid in IL6 and AMH? A Case-Control Study. JCM 2023, 12, 1829. [Google Scholar] [CrossRef]
- Godin, S.K.; Wagner, J.; Huang, P.; Bree, D. The Role of Peripheral Nerve Signaling in Endometriosis. FASEB Bioadv 2021, 3, 802–813. [Google Scholar] [CrossRef]
- Ahn, S.H.; Edwards, A.K.; Singh, S.S.; Young, S.L.; Lessey, B.A.; Tayade, C. IL-17A Contributes to the Pathogenesis of Endometriosis by Triggering Proinflammatory Cytokines and Angiogenic Growth Factors. J Immunol 2015, 195, 2591–2600. [Google Scholar] [CrossRef]
- Hirata, T.; Osuga, Y.; Hamasaki, K.; Yoshino, O.; Ito, M.; Hasegawa, A.; Takemura, Y.; Hirota, Y.; Nose, E.; Morimoto, C.; et al. Interleukin (IL)-17A Stimulates IL-8 Secretion, Cyclooxygensase-2 Expression, and Cell Proliferation of Endometriotic Stromal Cells. Endocrinology 2008, 149, 1260–1267. [Google Scholar] [CrossRef]
- Yang, C.-M.; Yang, C.-C.; Hsiao, L.-D.; Yu, C.-Y.; Tseng, H.-C.; Hsu, C.-K.; Situmorang, J.H. Upregulation of COX-2 and PGE2 Induced by TNF-α Mediated Through TNFR1/MitoROS/PKCα/P38 MAPK, JNK1/2/FoxO1 Cascade in Human Cardiac Fibroblasts. J Inflamm Res 2021, 14, 2807–2824. [Google Scholar] [CrossRef]
- Titiz, M.; Landini, L.; Souza Monteiro De Araujo, D.; Marini, M.; Seravalli, V.; Chieca, M.; Pensieri, P.; Montini, M.; De Siena, G.; Pasquini, B.; et al. Schwann Cell C5aR1 Co-Opts Inflammasome NLRP1 to Sustain Pain in a Mouse Model of Endometriosis. Nat Commun 2024, 15, 10142. [Google Scholar] [CrossRef]
- Atiakshin, D.; Patsap, O.; Kostin, A.; Mikhalyova, L.; Buchwalow, I.; Tiemann, M. Mast Cell Tryptase and Carboxypeptidase A3 in the Formation of Ovarian Endometrioid Cysts. IJMS 2023, 24, 6498. [Google Scholar] [CrossRef]
- Kim, H.G.; Lim, Y.S.; Hwang, S.; Kim, H.-Y.; Moon, Y.; Song, Y.J.; Na, Y.-J.; Yoon, S. Di-(2-Ethylhexyl) Phthalate Triggers Proliferation, Migration, Stemness, and Epithelial-Mesenchymal Transition in Human Endometrial and Endometriotic Epithelial Cells via the Transforming Growth Factor-β/Smad Signaling Pathway. Int J Mol Sci 2022, 23, 3938. [Google Scholar] [CrossRef] [PubMed]
- Zubrzycka, A.; Migdalska-Sęk, M.; Jędrzejczyk, S.; Brzeziańska-Lasota, E. The Expression of TGF-Β1, SMAD3, ILK and miRNA-21 in the Ectopic and Eutopic Endometrium of Women with Endometriosis. Int J Mol Sci 2023, 24, 2453. [Google Scholar] [CrossRef] [PubMed]
- Young, V.J.; Brown, J.K.; Saunders, P.T.K.; Duncan, W.C.; Horne, A.W. The Peritoneum Is Both a Source and Target of TGF-β in Women with Endometriosis. PLoS One 2014, 9, e106773. [Google Scholar] [CrossRef]
- Hanada, T.; Tsuji, S.; Nakayama, M.; Wakinoue, S.; Kasahara, K.; Kimura, F.; Mori, T.; Ogasawara, K.; Murakami, T. Suppressive Regulatory T Cells and Latent Transforming Growth Factor-β-Expressing Macrophages Are Altered in the Peritoneal Fluid of Patients with Endometriosis. Reprod Biol Endocrinol 2018, 16, 9. [Google Scholar] [CrossRef]
- Agostinis, C.; Balduit, A.; Mangogna, A.; Zito, G.; Romano, F.; Ricci, G.; Kishore, U.; Bulla, R. Immunological Basis of the Endometriosis: The Complement System as a Potential Therapeutic Target. Front Immunol 2021, 11, 599117. [Google Scholar] [CrossRef] [PubMed]
- Zaninelli, T.H.; Fattori, V.; Heintz, O.K.; Wright, K.R.; Bennallack, P.R.; Sim, D.; Bukhari, H.; Terry, K.L.; Vitonis, A.F.; Missmer, S.A.; et al. Targeting NGF but Not VEGFR1 or BDNF Signaling Reduces Endometriosis-Associated Pain in Mice. Journal of Advanced Research 2024, S2090123224003606. [Google Scholar] [CrossRef]
- Ansariniya, H.; Yavari, A.; Javaheri, A.; Zare, F. Oxidative Stress-Related Effects on Various Aspects of Endometriosis. Am J Reprod Immunol 2022, 88, e13593. [Google Scholar] [CrossRef]
- Scutiero, G.; Iannone, P.; Bernardi, G.; Bonaccorsi, G.; Spadaro, S.; Volta, C.A.; Greco, P.; Nappi, L. Oxidative Stress and Endometriosis: A Systematic Review of the Literature. Oxid Med Cell Longev 2017, 2017, 7265238. [Google Scholar] [CrossRef]
- Li, S.; Zhou, Y.; Huang, Q.; Fu, X.; Zhang, L.; Gao, F.; Jin, Z.; Wu, L.; Shu, C.; Zhang, X.; et al. Iron Overload in Endometriosis Peritoneal Fluid Induces Early Embryo Ferroptosis Mediated by HMOX1. Cell Death Discov 2021, 7, 355. [Google Scholar] [CrossRef]
- Wyatt, J.; Fernando, S.M.; Powell, S.G.; Hill, C.J.; Arshad, I.; Probert, C.; Ahmed, S.; Hapangama, D.K. The Role of Iron in the Pathogenesis of Endometriosis: A Systematic Review. Human Reproduction Open 2023, 2023, hoad033. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhao, X.; Zhang, L.; Xia, Q.; Peng, Y.; Zhang, H.; Yan, D.; Yang, Z.; Li, J. Iron Overload Inhibits Cell Proliferation and Promotes Autophagy via PARP1/SIRT1 Signaling in Endometriosis and Adenomyosis. Toxicology 2022, 465, 153050. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, H.; Yan, H.; Han, P.; Zhang, J.; Liu, Y. Deciphering the Role of Oxidative Stress in Male Infertility: Insights from Reactive Oxygen Species to Antioxidant Therapeutics. FBL 2025, 30, 27046. [Google Scholar] [CrossRef]
- Ye, Z.; Cheng, M.; Lian, W.; Leng, Y.; Qin, X.; Wang, Y.; Zhou, P.; Liu, X.; Peng, T.; Wang, R.; et al. GPX4 Deficiency-Induced Ferroptosis Drives Endometrial Epithelial Fibrosis in Polycystic Ovary Syndrome. Redox Biology 2025, 83, 103615. [Google Scholar] [CrossRef]
- Xie, C.; Lu, C.; Lv, N.; Kong, W.; Liu, Y. Identification and Analysis of Oxidative Stress-Related Genes in Endometriosis. Front Immunol 2025, 16, 1515490. [Google Scholar] [CrossRef]
- Yun, B.H.; Chon, S.J.; Choi, Y.S.; Cho, S.; Lee, B.S.; Seo, S.K. Pathophysiology of Endometriosis: Role of High Mobility Group Box-1 and Toll-Like Receptor 4 Developing Inflammation in Endometrium. PLoS ONE 2016, 11, e0148165. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Liu, B.; Dai, Y.; Li, F.; Bellone, S.; Zhou, Y.; Mamillapalli, R.; Zhao, D.; Venkatachalapathy, M.; Hu, Y.; et al. TET3-Overexpressing Macrophages Promote Endometriosis. Journal of Clinical Investigation 2024, 134, e181839. [Google Scholar] [CrossRef] [PubMed]
- Limam, I.; Ghali, R.; Abdelkarim, M.; Ouni, A.; Araoud, M.; Abdelkarim, M.; Hedhili, A.; Ben-Aissa Fennira, F. Tunisian Artemisia Campestris L.: A Potential Therapeutic Agent against Myeloma - Phytochemical and Pharmacological Insights. Plant Methods 2024, 20, 59. [Google Scholar] [CrossRef]
- Campara, K.; Rodrigues, P.; Viero, F.T.; da Silva, B.; Trevisan, G. A Systematic Review and Meta-Analysis of Advanced Oxidative Protein Products Levels (AOPP) Levels in Endometriosis: Association with Disease Stage and Clinical Implications. European Journal of Pharmacology 2025, 996, 177434. [Google Scholar] [CrossRef]
- Arendt, W.; Kleszczyński, K.; Gagat, M.; Izdebska, M. Endometriosis and Cytoskeletal Remodeling: The Functional Role of Actin-Binding Proteins. Cells 2025, 14, 360. [Google Scholar] [CrossRef]
- Bałkowiec, M.; Maksym, R.B.; Włodarski, P.K. The Bimodal Role of Matrix Metalloproteinases and Their Inhibitors in Etiology and Pathogenesis of Endometriosis. Mol Med Rep 2018, 18, 3123–3136. [Google Scholar] [CrossRef]
- Huang, Q.; Song, Y.; Lei, X.; Huang, H.; Nong, W. MMP-9 as a Clinical Marker for Endometriosis: A Meta-Analysis and Bioinformatics Analysis. Front. Endocrinol. 2024, 15, 1475531. [Google Scholar] [CrossRef] [PubMed]
- Khalili-Tanha, G.; Radisky, E.S.; Radisky, D.C.; Shoari, A. Matrix Metalloproteinase-Driven Epithelial-Mesenchymal Transition: Implications in Health and Disease. J Transl Med 2025, 23, 436. [Google Scholar] [CrossRef]
- Muharam, R.; Bowolaksono, A.; Maidarti, M.; Febri, R.; Mutia, K.; Iffanolida, P.; Ikhsan, M.; Sumapraja, K.; Pratama, G.; Harzif, A.; et al. Elevated MMP-9, Survivin, TGB1 and Downregulated Tissue Inhibitor of TIMP-1, Caspase-3 Activities Are Independent of the Low Levels miR-183 in Endometriosis. IJWH 2024, Volume 16, 1733–1742. [Google Scholar] [CrossRef]
- Luddi, A.; Marrocco, C.; Governini, L.; Semplici, B.; Pavone, V.; Luisi, S.; Petraglia, F.; Piomboni, P. Expression of Matrix Metalloproteinases and Their Inhibitors in Endometrium: High Levels in Endometriotic Lesions. Int J Mol Sci 2020, 21, 2840. [Google Scholar] [CrossRef]
- Gilabert-Estellés, J.; Estellés, A.; Gilabert, J.; Castelló, R.; España, F.; Falcó, C.; Romeu, A.; Chirivella, M.; Zorio, E.; Aznar, J. Expression of Several Components of the Plasminogen Activator and Matrix Metalloproteinase Systems in Endometriosis. Human Reproduction 2003, 18, 1516–1522. [Google Scholar] [CrossRef] [PubMed]
- Zanon, P.; Terraciano, P.B.; Quandt, L.; Palma Kuhl, C.; Pandolfi Passos, E.; Berger, M. Angiotensin II – AT1 Receptor Signalling Regulates the Plasminogen-Plasmin System in Human Stromal Endometrial Cells Increasing Extracellular Matrix Degradation, Cell Migration and Inducing a Proinflammatory Profile. Biochemical Pharmacology 2024, 225, 116280. [Google Scholar] [CrossRef] [PubMed]
- Limam, I.; Abdelkarim, M.; El Ayeb, M.; Crepin, M.; Marrakchi, N.; Di Benedetto, M. Disintegrin-like Protein Strategy to Inhibit Aggressive Triple-Negative Breast Cancer. Int J Mol Sci 2023, 24, 12219. [Google Scholar] [CrossRef]
- Wu, J.; Wang, J.; Pei, Z.; Zhu, Y.; Zhang, X.; Zhou, Z.; Ye, C.; Song, M.; Hu, Y.; Xue, P.; et al. Endothelial Senescence Induced by PAI-1 Promotes Endometrial Fibrosis. Cell Death Discov. 2025, 11, 89. [Google Scholar] [CrossRef]
- Kazmi, I.; Alharbi, K.S.; Al-Abbasi, F.A.; Almalki, W.H.; G, S.K.; Yasmeen, A.; Khan, A.; Gupta, G. Role of Epithelial-to-Mesenchymal Transition Markers in Different Stages of Endometriosis: Expression of the Snail, Slug, ZEB1, and Twist Genes. Crit Rev Eukaryot Gene Expr 2021, 31, 89–95. [Google Scholar] [CrossRef]
- Ntzeros, K.; Mavrogianni, D.; Blontzos, N.; Soyhan, N.; Kathopoulis, N.; Papamentzelopoulou, M.-S.; Chatzipapas, I.; Protopapas, A. Expression of ZEB1 in Different Forms of Endometriosis: A Pilot Study. European Journal of Obstetrics & Gynecology and Reproductive Biology 2023, 286, 121–125. [Google Scholar] [CrossRef]
- Ruiz, L.A.; Báez-Vega, P.M.; Ruiz, A.; Peterse, D.P.; Monteiro, J.B.; Bracero, N.; Beauchamp, P.; Fazleabas, A.T.; Flores, I. Dysregulation of Lysyl Oxidase Expression in Lesions and Endometrium of Women With Endometriosis. Reprod. Sci. 2015, 22, 1496–1508. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, S.; Canis, M.; Murakami, T.; Dechelotte, P.; Bruhat, M.A.; Okamura, K. Immunohistochemical Analysis of the Role of Angiogenic Status in the Vasculature of Peritoneal Endometriosis. Fertility and Sterility 2001, 76, 712–716. [Google Scholar] [CrossRef] [PubMed]
- Don, E.E.; Middelkoop, M.-A.; Hehenkamp, W.J.K.; Mijatovic, V.; Griffioen, A.W.; Huirne, J.A.F. Endometrial Angiogenesis of Abnormal Uterine Bleeding and Infertility in Patients with Uterine Fibroids-A Systematic Review. Int J Mol Sci 2023, 24, 7011. [Google Scholar] [CrossRef]
- Guo, S.-W.; Zheng, Y.; Lu, Y.; Liu, X.; Geng, J.-G. Slit2 Overexpression Results in Increased Microvessel Density and Lesion Size in Mice with Induced Endometriosis. Reprod Sci 2013, 20, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liu, J.; Gao, S.; Qiu, Y.; Wang, Y.; Zhang, Y.; Gao, L.; Qi, G.; Wu, Y.; Lash, G.E.; et al. Role of Slit2 Upregulation in Recurrent Miscarriage through Regulation of Stromal Decidualization. Placenta 2021, 103, 1–9. [Google Scholar] [CrossRef]
- Karaer, A.; Tuncay, G.; Uysal, O.; Semerci Sevimli, T.; Sahin, N.; Karabulut, U.; Sariboyaci, A.E. The Role of Prokineticins in Recurrent Implantation Failure. Journal of Gynecology Obstetrics and Human Reproduction 2020, 49, 101835. [Google Scholar] [CrossRef]
- Körbel, C.; Gerstner, M.D.; Menger, M.D.; Laschke, M.W. Notch Signaling Controls Sprouting Angiogenesis of Endometriotic Lesions. Angiogenesis 2018, 21, 37–46. [Google Scholar] [CrossRef]
- Torres, J.P.V.; Eulálio, E.C.; Oliveira, F. da C.; Ximenes, G.F.; Feitosa Filho, H.N.; de Souza, L.B.; Araujo Júnior, E.; Cavalcante, M.B. Endometriosis and Infertility: A Bibliometric Analysis of the 100 Most-Cited Articles from 2000 to 2023. Gynecologic and Obstetric Investigation 2024, 90, 129–142. [Google Scholar] [CrossRef]
- Li, Y.; Li, R.; Ouyang, N.; Dai, K.; Yuan, P.; Zheng, L.; Wang, W. Investigating the Impact of Local Inflammation on Granulosa Cells and Follicular Development in Women with Ovarian Endometriosis. Fertility and Sterility 2019, 112, 882–891. [Google Scholar] [CrossRef]
- Silva, J.R.V.; Lima, F.E.O.; Souza, A.L.P.; Silva, A.W.B. Interleukin-1β and TNF-α Systems in Ovarian Follicles and Their Roles during Follicular Development, Oocyte Maturation and Ovulation. Zygote 2020, 28, 270–277. [Google Scholar] [CrossRef]
- Adamczak, R.; Ukleja-Sokołowska, N.; Lis, K.; Dubiel, M. Function of Follicular Cytokines: Roles Played during Maturation, Development and Implantation of Embryo. Medicina (Kaunas) 2021, 57, 1251. [Google Scholar] [CrossRef]
- Sanchez, A.M.; Somigliana, E.; Vercellini, P.; Pagliardini, L.; Candiani, M.; Vigano, P. Endometriosis as a Detrimental Condition for Granulosa Cell Steroidogenesis and Development: From Molecular Alterations to Clinical Impact. The Journal of Steroid Biochemistry and Molecular Biology 2016, 155, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Han, M.-T.; Cheng, W.; Zhu, R.; Wu, H.-H.; Ding, J.; Zhao, N.-N.; Li, H.; Wang, F.-X. The Cytokine Profiles in Follicular Fluid and Reproductive Outcomes in Women with Endometriosis. Am J Reprod Immunol 2023, 89, e13633. [Google Scholar] [CrossRef]
- Vitagliano, A.; Saccardi, C.; Noventa, M.; Di Spiezio Sardo, A.; Saccone, G.; Cicinelli, E.; Pizzi, S.; Andrisani, A.; Litta, P.S. Effects of Chronic Endometritis Therapy on in Vitro Fertilization Outcome in Women with Repeated Implantation Failure: A Systematic Review and Meta-Analysis. Fertility and Sterility 2018, 110, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Peoc’h, K.; Puy, V.; Fournier, T. Haem Oxygenases Play a Pivotal Role in Placental Physiology and Pathology. Human Reproduction Update 2020, 26, 634–649. [Google Scholar] [CrossRef]
- Didziokaite, G.; Biliute, G.; Gudaite, J.; Kvedariene, V. Oxidative Stress as a Potential Underlying Cause of Minimal and Mild Endometriosis-Related Infertility. Int J Mol Sci 2023, 24, 3809. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Wang, Z.; Lv, L.; Liu, C.; Wang, L.; Sun, Y.; Zhao, Z.; Shi, B.; Li, Q.; Hao, G. Molecular Regulation of DNA Damage and Repair in Female Infertility: A Systematic Review. Reprod Biol Endocrinol 2024, 22, 103. [Google Scholar] [CrossRef]
- Máté, G.; Bernstein, L.R.; Török, A.L. Endometriosis Is a Cause of Infertility. Does Reactive Oxygen Damage to Gametes and Embryos Play a Key Role in the Pathogenesis of Infertility Caused by Endometriosis? Front Endocrinol (Lausanne) 2018, 9, 725. [Google Scholar] [CrossRef]
- Chen, C.; Zhou, Y.; Hu, C.; Wang, Y.; Yan, Z.; Li, Z.; Wu, R. Mitochondria and Oxidative Stress in Ovarian Endometriosis. Free Radical Biology and Medicine 2019, 136, 22–34. [Google Scholar] [CrossRef]
- Kobayashi, H.; Imanaka, S.; Nakamura, H.; Tsuji, A. Understanding the Role of Epigenomic, Genomic and Genetic Alterations in the Development of Endometriosis (Review). Molecular Medicine Reports 2014, 9, 1483–1505. [Google Scholar] [CrossRef]
- Rajani, S.; Chattopadhyay, R.; Goswami, S.K.; Ghosh, S.; Sharma, S.; Chakravarty, B. Assessment of Oocyte Quality in Polycystic Ovarian Syndrome and Endometriosis by Spindle Imaging and Reactive Oxygen Species Levels in Follicular Fluid and Its Relationship with IVF-ET Outcome. Journal of Human Reproductive Sciences 2012, 5, 187. [Google Scholar] [CrossRef]
- Mendes Godinho, C.; Soares, S.R.; Nunes, S.G.; Martínez, J.M.M.; Santos-Ribeiro, S. Natural Proliferative Phase Frozen Embryo Transfer—a New Approach Which May Facilitate Scheduling without Hindering Pregnancy Outcomes. Human Reproduction 2024, 39, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Simopoulou, M.; Rapani, A.; Grigoriadis, S.; Pantou, A.; Tsioulou, P.; Maziotis, E.; Tzanakaki, D.; Triantafyllidou, O.; Kalampokas, T.; Siristatidis, C.; et al. Getting to Know Endometriosis-Related Infertility Better: A Review on How Endometriosis Affects Oocyte Quality and Embryo Development. Biomedicines 2021, 9, 273. [Google Scholar] [CrossRef] [PubMed]
- Vallvé-Juanico, J.; Houshdaran, S.; Giudice, L.C. The Endometrial Immune Environment of Women with Endometriosis. Human Reproduction Update 2019, 25, 565–592. [Google Scholar] [CrossRef]
- McGlade, E.A.; Miyamoto, A.; Winuthayanon, W. Progesterone and Inflammatory Response in the Oviduct during Physiological and Pathological Conditions. Cells 2022, 11, 1075. [Google Scholar] [CrossRef]
- He, L.; Xu, H.; Liu, M.; Tan, Y.; Huang, S.; Yin, X.; Luo, X.; Chung, H.Y.; Gao, M.; Li, Y.; et al. The Ignored Structure in Female Fertility: Cilia in the Fallopian Tubes. Reproductive BioMedicine Online 2025, 50, 104346. [Google Scholar] [CrossRef] [PubMed]
- Nutu, M. The Role of Progesterone in the Regulation of Ciliary Activity in the Fallopian Tube. 2009; ISBN 978-91-628-7744-6. [Google Scholar]
- Artimovič, P.; Badovská, Z.; Toporcerová, S.; Špaková, I.; Smolko, L.; Sabolová, G.; Kriváková, E.; Rabajdová, M. Oxidative Stress and the Nrf2/PPARγ Axis in the Endometrium: Insights into Female Fertility. Cells 2024, 13, 1081. [Google Scholar] [CrossRef]
- Huang, E.; Wang, X.; Chen, L. Regulated Cell Death in Endometriosis. Biomolecules 2024, 14, 142. [Google Scholar] [CrossRef]
- Moustakli, E.; Stavros, S.; Katopodis, P.; Skentou, C.; Potiris, A.; Panagopoulos, P.; Domali, E.; Arkoulis, I.; Karampitsakos, T.; Sarafi, E.; et al. Oxidative Stress and the NLRP3 Inflammasome: Focus on Female Fertility and Reproductive Health. Cells 2025, 14, 36. [Google Scholar] [CrossRef]
- Jia, Z.; Li, B.; Matsuo, M.; Dewar, A.; Mustafaraj, A.; Dey, S.K.; Yuan, J.; Sun, X. Foxa2-Dependent Uterine Glandular Cell Differentiation Is Essential for Successful Implantation. Nat Commun 2025, 16, 2465. [Google Scholar] [CrossRef]
- Liu, H.; Huang, X.; Mor, G.; Liao, A. Epigenetic Modifications Working in the Decidualization and Endometrial Receptivity. Cell Mol Life Sci 2019, 77, 2091–2101. [Google Scholar] [CrossRef] [PubMed]
- Adamczyk, M.; Wender-Ozegowska, E.; Kedzia, M. Epigenetic Factors in Eutopic Endometrium in Women with Endometriosis and Infertility. Int J Mol Sci 2022, 23, 3804. [Google Scholar] [CrossRef] [PubMed]
- Daftary, G.S.; Troy, P.J.; Bagot, C.N.; Young, S.L.; Taylor, H.S. Direct Regulation of Β3-Integrin Subunit Gene Expression by HOXA10 in Endometrial Cells. Molecular Endocrinology 2002, 16, 571–579. [Google Scholar] [CrossRef]
- Du, H.; Taylor, H.S. The Role of Hox Genes in Female Reproductive Tract Development, Adult Function, and Fertility. Cold Spring Harb Perspect Med 2016, 6, a023002. [Google Scholar] [CrossRef]
- Kaya, H.S.; Hantak, A.M.; Stubbs, L.J.; Taylor, R.N.; Bagchi, I.C.; Bagchi, M.K. Roles of Progesterone Receptor A and B Isoforms during Human Endometrial Decidualization. Mol Endocrinol 2015, 29, 882–895. [Google Scholar] [CrossRef]
- Liu, H.; Franken, A.; Bielfeld, A.P.; Fehm, T.; Niederacher, D.; Cheng, Z.; Neubauer, H.; Stamm, N. Progesterone-Induced Progesterone Receptor Membrane Component 1 Rise-to-Decline Changes Are Essential for Decidualization. Reprod Biol Endocrinol 2024, 22, 20. [Google Scholar] [CrossRef] [PubMed]
- Fabi, F.; Grenier, K.; Parent, S.; Adam, P.; Tardif, L.; Leblanc, V.; Asselin, E. Regulation of the PI3K/Akt Pathway during Decidualization of Endometrial Stromal Cells. PLoS One 2017, 12, e0177387. [Google Scholar] [CrossRef]
- Tran, D.N.; Hwang, Y.J.; Kim, K.C.; Li, R.; Marquardt, R.M.; Chen, C.; Young, S.L.; Lessey, B.A.; Kim, T.H.; Cheon, Y.-P.; et al. GRB2 Regulation of Essential Signaling Pathways in the Endometrium Is Critical for Implantation and Decidualization. Nat Commun 2025, 16, 2192. [Google Scholar] [CrossRef]
- Li, X.; Zhou, J.; Fang, M.; Yu, B. Pregnancy Immune Tolerance at the Maternal-Fetal Interface. Int Rev Immunol 2020, 39, 247–263. [Google Scholar] [CrossRef]
- Boucher, A.; Brichant, G.; Gridelet, V.; Nisolle, M.; Ravet, S.; Timmermans, M.; Henry, L. Implantation Failure in Endometriosis Patients: Etiopathogenesis. J Clin Med 2022, 11, 5366. [Google Scholar] [CrossRef]
- Xie, M.; Li, Y.; Meng, Y.-Z.; Xu, P.; Yang, Y.-G.; Dong, S.; He, J.; Hu, Z. Uterine Natural Killer Cells: A Rising Star in Human Pregnancy Regulation. Front Immunol 2022, 13, 918550. [Google Scholar] [CrossRef]
- Huang, N.; Chi, H.; Qiao, J. Role of Regulatory T Cells in Regulating Fetal-Maternal Immune Tolerance in Healthy Pregnancies and Reproductive Diseases. Front Immunol 2020, 11, 1023. [Google Scholar] [CrossRef]
- Li, Z.; Si, P.; Meng, T.; Zhao, X.; Zhu, C.; Zhang, D.; Meng, S.; Li, N.; Liu, R.; Ni, T.; et al. CCR8+ Decidual Regulatory T Cells Maintain Maternal-Fetal Immune Tolerance during Early Pregnancy. Sci Immunol 2025, 10, eado2463. [Google Scholar] [CrossRef]
- Sisnett, D.J.; Zutautas, K.B.; Miller, J.E.; Lingegowda, H.; Ahn, S.H.; McCallion, A.; Bougie, O.; Lessey, B.A.; Tayade, C. The Dysregulated IL-23/TH17 Axis in Endometriosis Pathophysiology. The Journal of Immunology 2024, 212, 1428–1441. [Google Scholar] [CrossRef] [PubMed]
- Berdiaki, A.; Vergadi, E.; Makrygiannakis, F.; Vrekoussis, T.; Makrigiannakis, A. Title: Repeated Implantation Failure Is Associated with Increased Th17/Treg Cell Ratio, during the Secretory Phase of the Human Endometrium. Journal of Reproductive Immunology 2024, 161, 104170. [Google Scholar] [CrossRef] [PubMed]
- Vassilopoulou, L.; Matalliotakis, M.; Zervou, M.I.; Matalliotaki, C.; Krithinakis, K.; Matalliotakis, I.; Spandidos, D.A.; Goulielmos, G.N. Defining the Genetic Profile of Endometriosis (Review). Experimental and Therapeutic Medicine 2019, 17, 3267–3281. [Google Scholar] [CrossRef] [PubMed]
- Pagliardini, L.; Gentilini, D.; Sanchez, A.M.; Candiani, M.; Viganò, P.; Di Blasio, A.M. Replication and Meta-Analysis of Previous Genome-Wide Association Studies Confirm Vezatin as the Locus with the Strongest Evidence for Association with Endometriosis. Human Reproduction 2015, 30, 987–993. [Google Scholar] [CrossRef]
- Lalami, I.; Abo, C.; Borghese, B.; Chapron, C.; Vaiman, D. Genomics of Endometriosis: From Genome Wide Association Studies to Exome Sequencing. Int J Mol Sci 2021, 22, 7297. [Google Scholar] [CrossRef]
- Sheu, J.J.-C.; Lin, W.-Y.; Liu, T.-Y.; Chang, C.Y.-Y.; Cheng, J.; Li, Y.-H.; Chen, C.-M.; Tseng, C.-C.; Ding, W.Y.; Chung, C.; et al. Ethnic-Specific Genetic Susceptibility Loci for Endometriosis in Taiwanese-Han Population: A Genome-Wide Association Study. J Hum Genet 2024, 69, 573–583. [Google Scholar] [CrossRef]
- Kasvandik, S.; Samuel, K.; Peters, M.; Eimre, M.; Peet, N.; Roost, A.M.; Padrik, L.; Paju, K.; Peil, L.; Salumets, A. Deep Quantitative Proteomics Reveals Extensive Metabolic Reprogramming and Cancer-Like Changes of Ectopic Endometriotic Stromal Cells. J. Proteome Res. 2016, 15, 572–584. [Google Scholar] [CrossRef]
- Young, V.J.; Brown, J.K.; Maybin, J.; Saunders, P.T.K.; Duncan, W.C.; Horne, A.W. Transforming Growth Factor-β Induced Warburg-Like Metabolic Reprogramming May Underpin the Development of Peritoneal Endometriosis. The Journal of Clinical Endocrinology & Metabolism 2014, 99, 3450–3459. [Google Scholar] [CrossRef]
- Lu, C.; Xu, J.; Li, K.; Wang, J.; Dai, Y.; Chen, Y.; Chai, R.; Xu, C.; Kang, Y. Chronic Stress Blocks the Endometriosis Immune Response by Metabolic Reprogramming. Int J Mol Sci 2023, 25, 29. [Google Scholar] [CrossRef] [PubMed]
- Karaer, A.; Tuncay, G.; Mumcu, A.; Dogan, B. Metabolomics Analysis of Follicular Fluid in Women with Ovarian Endometriosis Undergoing in Vitro Fertilization. Systems Biology in Reproductive Medicine 2019, 65, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Kim, H.-R.; Lim, E.J.; Park, M.; Yoon, J.A.; Kim, Y.S.; Kim, E.-K.; Shin, J.-E.; Kim, J.H.; Kwon, H.; et al. Integrative Analyses of Uterine Transcriptome and MicroRNAome Reveal Compromised LIF-STAT3 Signaling and Progesterone Response in the Endometrium of Patients with Recurrent/Repeated Implantation Failure (RIF). PLoS ONE 2016, 11, e0157696. [Google Scholar] [CrossRef] [PubMed]
- Sadłocha, M.; Toczek, J.; Major, K.; Staniczek, J.; Stojko, R. Endometriosis: Molecular Pathophysiology and Recent Treatment Strategies—Comprehensive Literature Review. Pharmaceuticals (Basel) 2024, 17, 827. [Google Scholar] [CrossRef]
- Clemenza, S.; Vannuccini, S.; Ruotolo, A.; Capezzuoli, T.; Petraglia, F. Advances in Targeting Estrogen Synthesis and Receptors in Patients with Endometriosis. Expert Opin Investig Drugs 2022, 31, 1227–1238. [Google Scholar] [CrossRef]
- Harris, H.A.; Bruner-Tran, K.L.; Zhang, X.; Osteen, K.G.; Lyttle, C.R. A Selective Estrogen Receptor-β Agonist Causes Lesion Regression in an Experimentally Induced Model of Endometriosis. Human Reproduction 2005, 20, 936–941. [Google Scholar] [CrossRef]
- Gibson, D.A.; Simitsidellis, I.; Collins, F.; Saunders, P.T.K. Androgens, Oestrogens and Endometrium: A Fine Balance between Perfection and Pathology. 2020. [Google Scholar] [CrossRef]
- Jiang, L.; Wang, S.; Xia, X.; Zhang, T.; Wang, X.; Zeng, F.; Ma, J.; Fang, X. Novel Diagnostic Biomarker BST2 Identified by Integrated Transcriptomics Promotes the Development of Endometriosis via the TNF-α/NF-κB Signaling Pathway. Biochemical Genetics 2025, 63, 354–377. [Google Scholar] [CrossRef]
- Xu, X.; Mei, J.; Zhang, B.; Jiang, X.; Wang, L.; Zhang, A.; Li, J.; Chen, S.; He, Y.; Fang, Y.; et al. Association Between Circulating Cytokines and Endometriosis: A Mendelian Randomization Study. J Cell Mol Med 2025, 29, e70532. [Google Scholar] [CrossRef]
- Bora, G.; Yaba, A. The Role of Mitogen-Activated Protein Kinase Signaling Pathway in Endometriosis. J Obstet Gynaecol Res 2021, 47, 1610–1623. [Google Scholar] [CrossRef]
- Wang, B.; Gao, M.; Yao, Y.; Li, H.; Zhang, X. Focusing on the Role of Protein Kinase mTOR in Endometrial Physiology and Pathology: Insights for Therapeutic Interventions. Molecular Biology Reports 2024, 51, 359. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Li, Y.M.; Li, R.Y.; Yang, Y.E.; Wei, M.; Ha, C. U0126 and BAY11-7082 Inhibit the Progression of Endometriosis in a Rat Model by Suppressing the MEK/ERK/NF-κB Pathway. Womens Health Rep (New Rochelle) 2023, 4, 65–77. [Google Scholar] [CrossRef]
- Nakamura, A.; Tanaka, Y.; Amano, T.; Takebayashi, A.; Takahashi, A.; Hanada, T.; Tsuji, S.; Murakami, T. mTOR Inhibitors as Potential Therapeutics for Endometriosis: A Narrative Review. Mol Hum Reprod 2024, 30, gaae041. [Google Scholar] [CrossRef]
- Huang, Z.; Tan, Y. The Potential of Cylindromatosis (CYLD) as a Therapeutic Target in Oxidative Stress-Associated Pathologies: A Comprehensive Evaluation. Int J Mol Sci 2023, 24, 8368. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K. Immunotoxicological Disruption of Pregnancy as a New Research Area in Immunotoxicology. J Immunotoxicol 2025, 22, 2475772. [Google Scholar] [CrossRef]
- Shi, J.; Xu, Q.; Yu, S.; Zhang, T. Perturbations of the Endometrial Immune Microenvironment in Endometriosis and Adenomyosis: Their Impact on Reproduction and Pregnancy. Seminars in Immunopathology 2025, 47, 16. [Google Scholar] [CrossRef]
- Li, J.; Yan, S.; Li, Q.; Huang, Y.; Ji, M.; Jiao, X.; Yuan, M.; Wang, G. Macrophage-Associated Immune Checkpoint CD47 Blocking Ameliorates Endometriosis. Molecular Human Reproduction 2022, 28, gaac010. [Google Scholar] [CrossRef] [PubMed]
- Keleş, C.D.; Vural, B.; Filiz, S.; Vural, F.; Gacar, G.; Eraldemir, F.C.; Kurnaz, S. THE EFFECTS OF ETANERCEPT AND CABERGOLINE ON ENDOMETRIOTIC IMPLANTS, UTERUS AND OVARIES IN RAT ENDOMETRIOSIS MODEL. J Reprod Immunol 2021, 146, 103340. [Google Scholar] [CrossRef]
- Lu, D.; Song, H.; Shi, G. Anti-TNF-α Treatment for Pelvic Pain Associated with Endometriosis. Cochrane Database Syst Rev 2013, 2013, CD008088. [Google Scholar] [CrossRef]
- D’Hooghe, T.M.; Nugent, N.P.; Cuneo, S.; Chai, D.C.; Deer, F.; Debrock, S.; Kyama, C.M.; Mihalyi, A.; Mwenda, J.M. Recombinant Human TNFRSF1A (r-hTBP1) Inhibits the Development of Endometriosis in Baboons: A Prospective, Randomized, Placebo- and Drug-Controlled Study. Biol Reprod 2006, 74, 131–136. [Google Scholar] [CrossRef]
- Wang, P.-H.; Hsiao, S.-M.; Matsuki, S.; Hanada, R.; Chen, C.-A.; Nishimoto-Kakiuchi, A.; Sekiya, M.; Nakai, K.; Matsushima, J.; Sawada, M. Safety and Pharmacology of AMY109, a Long-Acting Anti-Interleukin-8 Antibody, for Endometriosis: A Double-Blind, Randomized Phase 1 Trial. F&S Reports, 2025. [Google Scholar] [CrossRef]
- Acién, P.; Quereda, F.; Campos, A.; Gomez-Torres, M.-J.; Velasco, I.; Gutierrez, M. Use of Intraperitoneal Interferon α-2b Therapy after Conservative Surgery for Endometriosis and Postoperative Medical Treatment with Depot Gonadotropin-Releasing Hormone Analog: A Randomized Clinical Trial. Fertility and Sterility 2002, 78, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Yan, Q.; Zhang, M.; Zhu, Y.; Liu, J.; Jiang, Y.; Zhen, X.; Xu, M.; Yue, Q.; Zhou, J.; et al. FHL1 Mediates HOXA10 Deacetylation via SIRT2 to Enhance Blastocyst-Epithelial Adhesion. Cell Death Discov 2022, 8, 461. [Google Scholar] [CrossRef] [PubMed]
- Zubrzycka, A.; Zubrzycki, M.; Perdas, E.; Zubrzycka, M. Genetic, Epigenetic, and Steroidogenic Modulation Mechanisms in Endometriosis. J Clin Med 2020, 9, 1309. [Google Scholar] [CrossRef] [PubMed]
- Psilopatis, I.; Vrettou, K.; Fleckenstein, F.N.; Theocharis, S. The Impact of Histone Modifications in Endometriosis Highlights New Therapeutic Opportunities. Cells 2023, 12, 1227. [Google Scholar] [CrossRef]
- Lu, H.; Yang, X.; Zhang, Y.; Lu, R.; Wang, X. Epigenetic Disorder May Cause Downregulation of HOXA10 in the Eutopic Endometrium of Fertile Women with Endometriosis. Reprod Sci 2013, 20, 78–84. [Google Scholar] [CrossRef]
- Chandrakanth, A.; Firdous, S.; Vasantharekha, R.; Santosh, W.; Seetharaman, B. Exploring the Effects of Endocrine-Disrupting Chemicals and miRNA Expression in the Pathogenesis of Endometriosis by Unveiling the Pathways: A Systematic Review. Reproductive Sciences 2024, 31, 932–941. [Google Scholar] [CrossRef]
- Li, M.; Zhou, Y.; Taylor, H.S. miR-451a Inhibition Reduces Established Endometriosis Lesions in Mice. Reprod Sci 2019, 26, 1506–1511. [Google Scholar] [CrossRef]
- Dong, L.; Zhang, L.; Liu, H.; Xie, M.; Gao, J.; Zhou, X.; Zhao, Q.; Zhang, S.; Yang, J. Circ_0007331 Knock-down Suppresses the Progression of Endometriosis via miR-200c-3p/HiF-1α Axis. Journal of Cellular and Molecular Medicine 2020, 24, 12656–12666. [Google Scholar] [CrossRef]
- Wu, D.; Lu, P.; Mi, X.; Miao, J. Exosomal miR-214 from Endometrial Stromal Cells Inhibits Endometriosis Fibrosis. Molecular Human Reproduction 2018, 24, 357–365. [Google Scholar] [CrossRef]
- Zhou, C.-F.; Liu, M.-J.; Wang, W.; Wu, S.; Huang, Y.-X.; Chen, G.-B.; Liu, L.-M.; Peng, D.-X.; Wang, X.-F.; Cai, X.-Z.; et al. miR-205-5p Inhibits Human Endometriosis Progression by Targeting ANGPT2 in Endometrial Stromal Cells. Stem Cell Res Ther 2019, 10, 287. [Google Scholar] [CrossRef]
- Budani, M.C.; Tiboni, G.M. Effects of Supplementation with Natural Antioxidants on Oocytes and Preimplantation Embryos. Antioxidants (Basel) 2020, 9, 612. [Google Scholar] [CrossRef] [PubMed]
- Jara-Palacios, M.J.; Gonçalves, S.; Heredia, F.J.; Hernanz, D.; Romano, A. Extraction of Antioxidants from Winemaking Byproducts: Effect of the Solvent on Phenolic Composition, Antioxidant and Anti-Cholinesterase Activities, and Electrochemical Behaviour. Antioxidants 2020, 9, 675. [Google Scholar] [CrossRef]
- Sharifi, M.; Rajabpoor Nikoo, N.; Badehnoosh, B.; Shafabakhsh, R.; Asemi, R.; Reiter, R.J.; Asemi, Z. Therapeutic Effects of Melatonin on Endometriosis, Targeting Molecular Pathways: Current Knowledge and Future Perspective. Pathology - Research and Practice 2023, 243, 154368. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hung, S.-W.; Zhang, R.; Man, G.C.-W.; Zhang, T.; Chung, J.P.-W.; Fang, L.; Wang, C.-C. Melatonin in Endometriosis: Mechanistic Understanding and Clinical Insight. Nutrients 2022, 14, 4087. [Google Scholar] [CrossRef]
- Park, S.; Ham, J.; Yang, C.; Park, W.; Park, H.; An, G.; Song, J.; Hong, T.; Park, S.J.; Kim, H.S.; et al. Melatonin Inhibits Endometriosis Development by Disrupting Mitochondrial Function and Regulating tiRNAs. Journal of Pineal Research 2023, 74, e12842. [Google Scholar] [CrossRef]
- Anastasi, E.; Scaramuzzino, S.; Viscardi, M.F.; Viggiani, V.; Piccioni, M.G.; Cacciamani, L.; Merlino, L.; Angeloni, A.; Muzii, L.; Porpora, M.G. Efficacy of N-Acetylcysteine on Endometriosis-Related Pain, Size Reduction of Ovarian Endometriomas, and Fertility Outcomes. IJERPH 2023, 20, 4686. [Google Scholar] [CrossRef]
- Limam, I.; Ben Aissa-Fennira, F.; Essid, R.; Chahbi, A.; Kefi, S.; Mkadmini, K.; Elkahoui, S.; Abdelkarim, M. Hydromethanolic Root and Aerial Part Extracts from Echium Arenarium Guss Suppress Proliferation and Induce Apoptosis of Multiple Myeloma Cells through Mitochondrial Pathway. Environ Toxicol 2021, 36, 874–886. [Google Scholar] [CrossRef] [PubMed]
- Limam, I.; Abdelkarim, M.; Essid, R.; Chahbi, A.; Fathallah, M.; Elkahoui, S.; Ben Aissa-Fennira, F. Olea Europaea L. Cv. Chetoui Leaf and Stem Hydromethanolic Extracts Suppress Proliferation and Promote Apoptosis via Caspase Signaling on Human Multiple Myeloma Cells. European Journal of Integrative Medicine 2020, 37, 101145. [Google Scholar] [CrossRef]
- Gołąbek-Grenda, A.; Kaczmarek, M.; Juzwa, W.; Olejnik, A. Natural Resveratrol Analogs Differentially Target Endometriotic Cells into Apoptosis Pathways. Sci Rep 2023, 13, 11468. [Google Scholar] [CrossRef]
- Podgrajsek, R.; Ban Frangez, H.; Stimpfel, M. Molecular Mechanism of Resveratrol and Its Therapeutic Potential on Female Infertility. IJMS 2024, 25, 3613. [Google Scholar] [CrossRef]
- Gołąbek-Grenda, A.; Juzwa, W.; Kaczmarek, M.; Olejnik, A. Resveratrol and Its Natural Analogs Mitigate Immune Dysregulation and Oxidative Imbalance in the Endometriosis Niche Simulated in a Co-Culture System of Endometriotic Cells and Macrophages. Nutrients 2024, 16, 3483. [Google Scholar] [CrossRef]
- Fan, J.; Chen, C.; Zhong, Y. A Cohort Study on IVF Outcomes in Infertile Endometriosis Patients: The Effects of Rapamycin Treatment. Reprod Biomed Online 2024, 48, 103319. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wang, J.; Ding, X.; Zhang, Q.; Duan, P. Emerging Strategies for the Treatment of Endometriosis. Biomedical Technology 2024, 7, 46–62. [Google Scholar] [CrossRef]
- Sanamiri, K.; Mahdian, S.; Moini, A.; Shahhoseini, M. Non-Hormonal Therapy for Endometriosis Based on Angiogenesis, Oxidative Stress and Inflammation. Int J Fertil Steril 2024, 18, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Ates, C.; Dilbaz, B.; Ergani, S.Y.; Atabay, F.; Ates, C.; Dilbaz, B.; Ergani, S.Y.; Atabay, F. The Effect of Antiangiogenic Agent Aflibercept on Surgically Induced Endometriosis in a Rat Model. Cirugía y cirujanos 2024, 92, 10–19. [Google Scholar] [CrossRef]
- França, P.R. de C.; Paiva, J.P.B. de; Carvalho, R.R. de; Figueiredo, C.P.; Sirois, P.; Fernandes, P.D. R-954, a Bradykinin B1 Receptor Antagonist, as a Potential Therapy in a Preclinical Endometriosis Model. Peptides 2024, 181, 171294. [Google Scholar] [CrossRef]
- Shahgholi, N.; Noormohammadi, Z.; Moieni, A.; Karimipoor, M. Cabergoline’s Promise in Endometriosis: Restoring Molecular Balance to Improve Reproductive Potential. Gynecologic and Obstetric Investigation 2025. [CrossRef]
- Pellicer, A.; Taylor, H.S.; Alberich-Bayarri, A.; Liu, Y.; Gamborg, M.; Barletta, K.E.; Pinton, P.; Heiser, P.W.; Bagger, Y.Z. Quinagolide Vaginal Ring for Reduction of Endometriotic Lesions: Results from the QLARITY Trial. European Journal of Obstetrics & Gynecology and Reproductive Biology 2025, 310, 113946. [Google Scholar] [CrossRef]
- Estole-Casanova, L.A. A Comprehensive Review of the Efficacy and Safety of Dopamine Agonists for Women with Endometriosis-Associated Infertility from Inception to July 31, 2022. Acta Med Philipp 2024, 58, 49–64. [Google Scholar] [CrossRef]
- Dymanowska-Dyjak, I.; Frankowska, K.; Abramiuk, M.; Polak, G. Oxidative Imbalance in Endometriosis-Related Infertility—The Therapeutic Role of Antioxidants. Int J Mol Sci 2024, 25, 6298. [Google Scholar] [CrossRef]
- Meneghetti, J.K.; Pedrotti, M.T.; Coimbra, I.M.; da Cunha-Filho, J.S.L. Effect of Dietary Interventions on Endometriosis: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Reprod Sci 2024, 31, 3613–3623. [Google Scholar] [CrossRef] [PubMed]
- Acharya, B.; Behera, A.; Behera, S.; Moharana, S. Recent Advances in Nanotechnology-Based Drug Delivery Systems for the Diagnosis and Treatment of Reproductive Disorders. ACS Appl. Bio Mater. 2024, 7, 1336–1361. [Google Scholar] [CrossRef] [PubMed]
- Taghavipour, M.; Sadoughi, F.; Mirzaei, H.; Yousefi, B.; Moazzami, B.; Chaichian, S.; Mansournia, M.A.; Asemi, Z. Apoptotic Functions of microRNAs in Pathogenesis, Diagnosis, and Treatment of Endometriosis. Cell Biosci 2020, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Akkaya-Ulum, Y.Z.; Sen, B.; Akbaba, T.H.; Balci-Peynircioglu, B. InflammamiRs in Focus: Delivery Strategies and Therapeutic Approaches. The FASEB Journal 2024, 38, e23528. [Google Scholar] [CrossRef]
- Qian, H.; Maghsoudloo, M.; Kaboli, P.J.; Babaeizad, A.; Cui, Y.; Fu, J.; Wang, Q.; Imani, S. Decoding the Promise and Challenges of miRNA-Based Cancer Therapies: An Essential Update on miR-21, miR-34, and miR-155. Int. J. Med. Sci. 2024, 21, 2781–2798. [Google Scholar] [CrossRef]
- Monnaka, V.U.; Hernandes, C.; Heller, D.; Podgaec, S. Overview of miRNAs for the Non-Invasive Diagnosis of Endometriosis: Evidence, Challenges and Strategies. A Systematic Review. Einstein (São Paulo) 2021, 19, eRW5704. [Google Scholar] [CrossRef]
- Nshimiyimana, P.; Major, I.; Colbert, D.M.; Buckley, C. Progress in the Biomedical Application of Biopolymers: An Overview of the Status Quo and Outlook in Managing Intrauterine Adhesions. Macromol 2025, 5, 25. [Google Scholar] [CrossRef]
- Ellis, K.; Munro, D.; Clarke, J. Endometriosis Is Undervalued: A Call to Action. Front. Glob. Womens Health 2022, 3, 902371. [Google Scholar] [CrossRef]
- Yuspa, R. Economic Burden of Endometriosis: A Systematic Review and Meta-Analysis. a-jhr 2024, 3, 304–319. [Google Scholar] [CrossRef]
- Saunders, P.T.K.; Horne, A.W. Endometriosis: Etiology, Pathobiology, and Therapeutic Prospects. Cell 2021, 184, 2807–2824. [Google Scholar] [CrossRef]
- Brulport, A.; Bourdon, M.; Vaiman, D.; Drouet, C.; Pocate-Cheriet, K.; Bouzid, K.; Marcellin, L.; Santulli, P.; Abo, C.; Jeljeli, M.; et al. An Integrated Multi-Tissue Approach for Endometriosis Candidate Biomarkers: A Systematic Review. Reprod Biol Endocrinol 2024, 22, 21. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, S.D. Prevention of Endometriosis: Is It Possible? In Endometriosis and Adenomyosis: Global Perspectives Across the Lifespan; Oral, E., Ed.; Springer International Publishing: Cham, 2022; pp. 353–360. ISBN 978-3-030-97236-3. [Google Scholar]
- As-Sanie, S.; Mackenzie, S.C.; Morrison, L.; Schrepf, A.; Zondervan, K.T.; Horne, A.W.; Missmer, S.A. Endometriosis: A Review. JAMA 2025. [Google Scholar] [CrossRef] [PubMed]
- Smolarz, B.; Szyłło, K.; Romanowicz, H. Endometriosis: Epidemiology, Classification, Pathogenesis, Treatment and Genetics (Review of Literature). Int J Mol Sci 2021, 22, 10554. [Google Scholar] [CrossRef]
- Encalada Soto, D.; Rassier, S.; Green, I.C.; Burnett, T.; Khan, Z.; Cope, A. Endometriosis Biomarkers of the Disease: An Update. Current Opinion in Obstetrics & Gynecology 2022, 34, 210–219. [Google Scholar] [CrossRef]
- Pant, A.; Moar, K.; K. Arora, T.; Maurya, P.K. Biomarkers of Endometriosis. Clinica Chimica Acta 2023, 549, 117563. [Google Scholar] [CrossRef]
- Cardoso, J.V.; Perini, J.A.; Machado, D.E.; Pinto, R.; Medeiros, R. Systematic Review of Genome-Wide Association Studies on Susceptibility to Endometriosis. European Journal of Obstetrics & Gynecology and Reproductive Biology 2020, 255, 74–82. [Google Scholar] [CrossRef]
- Saunders, P.T.K. Insights from Genomic Studies on the Role of Sex Steroids in the Aetiology of Endometriosis. Reproduction and Fertility 2022, 3, R51–R65. [Google Scholar] [CrossRef]
- Verma, S.; Guare, L.; Humphrey, L.A.; Rush, M.; Pollie, M.; Jaworski, J.; Akerele, A.; Luo, Y.; Weng, C.; Wei, W.-Q.; et al. Enhancing Genetic Association Power in Endometriosis through Unsupervised Clustering of Clinical Subtypes Identified from Electronic Health Records 2024.
- Pujol Gualdo, N.; Džigurski, J.; Rukins, V.; Pajuste, F.-D.; Wolford, B.N.; Võsa, M.; Golob, M.; Haug, L.; Alver, M.; Läll, K.; et al. Atlas of Genetic and Phenotypic Associations across 42 Female Reproductive Health Diagnoses. Nat Med 2025, 31, 1626–1634. [Google Scholar] [CrossRef]
- Guare, L.A.; Das, J.; Caruth, L.; Rajagopalan, A.; Akerele, A.T.; Brumpton, B.M.; Chen, T.-T.; Kottyan, L.; Lin, Y.-F.; Moreno, E.; et al. Expanding the Genetic Landscape of Endometriosis: Integrative -Omics Analyses Uncover Key Pathways from a Multi-Ancestry Study of over 900,000 Women 2024.
- Benkhalifa, M.; Menoud, P.A.; Piquemal, D.; Hazout, J.Y.; Mahjoub, S.; Zarquaoui, M.; Louanjli, N.; Cabry, R.; Hazout, A. Quantification of Free Circulating DNA and Differential Methylation Profiling of Selected Genes as Novel Non-Invasive Biomarkers for Endometriosis Diagnosis. Biomolecules 2025, 15, 69. [Google Scholar] [CrossRef]
- Papari, E.; Noruzinia, M.; Kashani, L.; Foster, W.G. Identification of Candidate microRNA Markers of Endometriosis with the Use of Next-Generation Sequencing and Quantitative Real-Time Polymerase Chain Reaction. Fertility and Sterility 2020, 113, 1232–1241. [Google Scholar] [CrossRef]
- Moustafa, S.; Burn, M.; Mamillapalli, R.; Nematian, S.; Flores, V.; Taylor, H.S. Accurate Diagnosis of Endometriosis Using Serum microRNAs. American Journal of Obstetrics and Gynecology 2020, 223, 557.e1–557.e11. [Google Scholar] [CrossRef] [PubMed]
- Ravaggi, A.; Bergamaschi, C.; Galbiati, C.; Zanotti, L.; Fabricio, A.S.C.; Gion, M.; Cappelletto, E.; Leon, A.E.; Gennarelli, M.; Romagnolo, C.; et al. Circulating Serum Micro-RNA as Non-Invasive Diagnostic Biomarkers of Endometriosis. Biomedicines 2024, 12, 2393. [Google Scholar] [CrossRef]
- Mohammadi, S.D.; Moeini, A.; Rastegar, T.; Amidi, F.; Saffari, M.; Zhaeentan, S.; Akhavan, S.; Moradi, B.; Heydarikhah, F.; Takzare, N. Diagnostic Accuracy of Plasma microRNA as a Potential Biomarker for Detection of Endometriosis. Systems Biology in Reproductive Medicine 2025, 71, 61–75. [Google Scholar] [CrossRef]
- Schoeman, E.M.; Bringans, S.; Peters, K.; Casey, T.; Andronis, C.; Chen, L.; Duong, M.; Girling, J.E.; Healey, M.; Boughton, B.A.; et al. Identification of Plasma Protein Biomarkers for Endometriosis and the Development of Statistical Models for Disease Diagnosis. Human Reproduction 2025, 40, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Sasamoto, N.; Ngo, L.H.; Vitonis, A.F.; Dillon, S.T.; Aziz, M.; Shafrir, A.L.; Missmer, S.A.; Libermann, T.A.; Terry, K.L. Prospective Evaluation of Plasma Proteins in Relation to Surgical Endometriosis Diagnosis in the Nurses’ Health Study II. eBioMedicine 2025, 115, 105688. [Google Scholar] [CrossRef]
- Dominoni, M.; Pasquali, M.F.; Musacchi, V.; De Silvestri, A.; Mauri, M.; Ferretti, V.V.; Gardella, B. Neutrophil to Lymphocytes Ratio in Deep Infiltrating Endometriosis as a New Toll for Clinical Management. Sci Rep 2024, 14, 7575. [Google Scholar] [CrossRef] [PubMed]
- Wójtowicz, M.; Zdun, D.; Owczarek, A.J.; Skrzypulec-Plinta, V.; Olszanecka-Glinianowicz, M. Evaluation of Proinflammatory Cytokines Concentrations in Plasma, Peritoneal, and Endometrioma Fluids in Women Operated on for Ovarian Endometriosis—A Pilot Study. IJMS 2025, 26, 5117. [Google Scholar] [CrossRef]
- Kimber-Trojnar, Ż.; Pilszyk, A.; Niebrzydowska, M.; Pilszyk, Z.; Ruszała, M.; Leszczyńska-Gorzelak, B. The Potential of Non-Invasive Biomarkers for Early Diagnosis of Asymptomatic Patients with Endometriosis. JCM 2021, 10, 2762. [Google Scholar] [CrossRef]
- Petr Klezl; Pavla Svobodova; Eliska Pospisilova; Vilem Maly; Vladimir Bobek; Katarina Kolostova Morphology of Mitochondrial Network in Disseminated Endometriosis Cellsin Spontaneous Pneumothorax Diagnostic Process. Cell Mol Biol (Noisy-le-grand) 2024, 70, 25–29. [CrossRef]
- Wang, S.; Cheng, H.; Zhu, H.; Yu, X.; Ye, X.; Chang, X. Precise Capture of Circulating Endometrial Cells in Endometriosis. Chinese Medical Journal 2024, 137, 1715–1723. [Google Scholar] [CrossRef]
- Chen, Y.; Zhu, H.-L.; Tang, Z.-W.; Neoh, K.H.; Ouyang, D.-F.; Cui, H.; Cheng, H.-Y.; Ma, R.-Q.; Ye, X.; Han, R.P.S.; et al. Evaluation of Circulating Endometrial Cells as a Biomarker for Endometriosis. Chin Med J (Engl) 2017, 130, 2339–2345. [Google Scholar] [CrossRef] [PubMed]
- Guder, C.; Heinrich, S.; Seifert-Klauss, V.; Kiechle, M.; Bauer, L.; Öllinger, R.; Pichlmair, A.; Theodoraki, M.-N.; Ramesh, V.; Bashiri Dezfouli, A.; et al. Extracellular Hsp70 and Circulating Endometriotic Cells as Novel Biomarkers for Endometriosis. IJMS 2024, 25, 11643. [Google Scholar] [CrossRef]
- Liang, Y.; Lu, Q.; Chen, M.; Zhao, X.; Chu, C.; Zhang, C.; Yuan, J.; Liu, H.; Lash, G.E. Impact of Endocrine Disrupting Chemicals (EDCs) on Epigenetic Regulation in the Uterus: A Narrative Review. Reproductive Biology and Endocrinology 2025, 23, 80. [Google Scholar] [CrossRef]
- Bromer, J.G.; Wu, J.; Zhou, Y.; Taylor, H.S. Hypermethylation of Homeobox A10 by in Utero Diethylstilbestrol Exposure: An Epigenetic Mechanism for Altered Developmental Programming. Endocrinology 2009, 150, 3376–3382. [Google Scholar] [CrossRef] [PubMed]
- Bromer, J.G.; Zhou, Y.; Taylor, M.B.; Doherty, L.; Taylor, H.S. Bisphenol-A Exposure in Utero Leads to Epigenetic Alterations in the Developmental Programming of Uterine Estrogen Response. FASEB J 2010, 24, 2273–2280. [Google Scholar] [CrossRef]
- Vallée, A.; Ceccaldi, P.-F.; Carbonnel, M.; Feki, A.; Ayoubi, J.-M. Pollution and Endometriosis: A Deep Dive into the Environmental Impacts on Women’s Health. BJOG 2024, 131, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Di Renzo, G.C.; Conry, J.A.; Blake, J.; DeFrancesco, M.S.; DeNicola, N.; Martin, J.N.; McCue, K.A.; Richmond, D.; Shah, A.; Sutton, P.; et al. International Federation of Gynecology and Obstetrics Opinion on Reproductive Health Impacts of Exposure to Toxic Environmental Chemicals. Int J Gynaecol Obstet 2015, 131, 219–225. [Google Scholar] [CrossRef]
- Dutta, S.; Banu, S.K.; Arosh, J.A. Endocrine Disruptors and Endometriosis. Reprod Toxicol 2023, 115, 56–73. [Google Scholar] [CrossRef]
- Rumph, J.T.; Stephens, V.R.; Archibong, A.E.; Osteen, K.G.; Bruner-Tran, K.L. Environmental Endocrine Disruptors and Endometriosis. Adv Anat Embryol Cell Biol 2020, 232, 57–78. [Google Scholar] [CrossRef]
- Ruder, E.H.; Hartman, T.J.; Goldman, M.B. Impact of Oxidative Stress on Female Fertility. Curr Opin Obstet Gynecol 2009, 21, 219–222. [Google Scholar] [CrossRef]
- Gupta, S.; Agarwal, A.; Krajcir, N.; Alvarez, J.G. Role of Oxidative Stress in Endometriosis. Reprod Biomed Online 2006, 13, 126–134. [Google Scholar] [CrossRef]
- Giudice, L.C.; Oskotsky, T.T.; Falako, S.; Opoku-Anane, J.; Sirota, M. Endometriosis in the Era of Precision Medicine and Impact on Sexual and Reproductive Health across the Lifespan and in Diverse Populations. FASEB J 2023, 37, e23130. [Google Scholar] [CrossRef] [PubMed]
- Coccia, M.E.; Nardone, L.; Rizzello, F. Endometriosis and Infertility: A Long-Life Approach to Preserve Reproductive Integrity. IJERPH 2022, 19, 6162. [Google Scholar] [CrossRef] [PubMed]
- Casalechi, M.; Di Stefano, G.; Fornelli, G.; Somigliana, E.; Viganò, P. Impact of Endometriosis on the Ovarian Follicles. Best Practice & Research Clinical Obstetrics & Gynaecology 2024, 92, 102430. [Google Scholar] [CrossRef]
- Tian, Z.; Zhang, Y.; Zhang, C.; Wang, Y.; Zhu, H.-L. Antral Follicle Count Is Reduced in the Presence of Endometriosis: A Systematic Review and Meta-Analysis. Reproductive BioMedicine Online 2021, 42, 237–247. [Google Scholar] [CrossRef]
- Romanski, P.A.; Brady, P.C.; Farland, L.V.; Thomas, A.M.; Hornstein, M.D. The Effect of Endometriosis on the Antimüllerian Hormone Level in the Infertile Population. J Assist Reprod Genet 2019, 36, 1179–1184. [Google Scholar] [CrossRef]
- Ozcan, P.; Sezer, F.; Altun, A.; Yildiz, C.; Timur, H.T.; Keles, E.C.; Dural, O.; Taha, H.S.; Saridogan, E. Potential Ability of Circulating INSL3 Level for the Prediction of Ovarian Reserve and IVF Success as a Novel Theca Cell–Specific Biomarker in Women with Unexplained Infertility and Diminished Ovarian Reserve. Reprod Biol Endocrinol 2025, 23, 40. [Google Scholar] [CrossRef]
- Walasik, I.; Klicka, K.; Grzywa, T.M.; Szymusik, I.; Włodarski, P.; Wielgoś, M.; Pietrzak, B.; Ludwin, A. Circulating miR-3613-5p but Not miR-125b-5p, miR-199a-3p, and miR-451a Are Biomarkers of Endometriosis. Reproductive Biology 2023, 23, 100796. [Google Scholar] [CrossRef] [PubMed]
- Marí-Alexandre, J.; Barceló-Molina, M.; Belmonte-López, E.; García-Oms, J.; Estellés, A.; Braza-Boïls, A.; Gilabert-Estellés, J. Micro-RNA Profile and Proteins in Peritoneal Fluid from Women with Endometriosis: Their Relationship with Sterility. Fertility and Sterility 2018, 109, 675–684.e2. [Google Scholar] [CrossRef]
- Nothnick, W.B.; Peterson, R.; Minchella, P.; Falcone, T.; Graham, A.; Findley, A. The Relationship and Expression of miR-451a, miR-25-3p and PTEN in Early Peritoneal Endometriotic Lesions and Their Modulation In Vitro. IJMS 2022, 23, 5862. [Google Scholar] [CrossRef]
- Muraoka, A.; Yokoi, A.; Yoshida, K.; Kitagawa, M.; Bayasula; Murakami, M.; Miyake, N.; Sonehara, R.; Nakamura, T.; Osuka, S.; et al. Serum-Derived Small Extracellular Vesicles as Biomarkers for Predicting Pregnancy and Delivery on Assisted Reproductive Technology in Patients with Endometriosis. Front. Endocrinol. 2025, 15, 1442684. [Google Scholar] [CrossRef] [PubMed]
- Salmasi, S.; Heidar, M.S.; Khaksary Mahabady, M.; Rashidi, B.; Mirzaei, H. MicroRNAs, Endometrial Receptivity and Molecular Pathways. Reprod Biol Endocrinol 2024, 22, 139. [Google Scholar] [CrossRef] [PubMed]
- Dabi, Y.; Suisse, S.; Puchar, A.; Delbos, L.; Poilblanc, M.; Descamps, P.; Haury, J.; Golfier, F.; Jornea, L.; Bouteiller, D.; et al. Endometriosis-Associated Infertility Diagnosis Based on Saliva microRNA Signatures. Reproductive BioMedicine Online 2023, 46, 138–149. [Google Scholar] [CrossRef]
- Huang, Y.; Cheng, Y.; Zhang, M.; Xia, Y.; Chen, X.; Xian, Y.; Lin, D.; Xie, S.; Guo, X. Oxidative Stress and Inflammatory Markers in Ovarian Follicular Fluid of Women with Diminished Ovarian Reserve during in Vitro Fertilization. J Ovarian Res 2023, 16, 206. [Google Scholar] [CrossRef]
- Voros, C.; Varthaliti, A.; Athanasiou, D.; Mavrogianni, D.; Papahliou, A.-M.; Bananis, K.; Koulakmanidis, A.-M.; Athanasiou, A.; Athanasiou, A.; Zografos, C.G.; et al. The Whisper of the Follicle: A Systematic Review of Micro Ribonucleic Acids as Predictors of Oocyte Quality and In Vitro Fertilization Outcomes. Cells 2025, 14, 787. [Google Scholar] [CrossRef]
- Karadbhajne, P.; More, A.; Dzoagbe, H.Y. The Role of Endometrial Microbiota in Fertility and Reproductive Health: A Narrative Review. Cureus 2025. [Google Scholar] [CrossRef]
- Enciso, M.; Medina-Ruiz, L.; Ferrández-Rives, M.; Jurado-García, I.; Rogel-Cayetano, S.; Aizpurua, J.; Sarasa, J. P-293 Influence of the Endometrial Microbiota Composition on the Clinical Outcomes of Egg Donation PGT-A Cycles with Controlled Endometrial Receptivity. Human Reproduction 2024, 39, deae108.660. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, M.; Zhong, J.; Xue, H. Altered Endometrial Microbiota Profile Is Associated With Poor Endometrial Receptivity of Repeated Implantation Failure. American J Rep Immunol 2024, 92, e70005. [Google Scholar] [CrossRef]
- Wei, Q.; Chen, H.; Zou, H.; Zhang, H.; Liu, S.; Zheng, J.; Zhang, S.; Hu, L. Impact of Vaginal Microecological Differences on Pregnancy Outcomes and Endometrial Microbiota in Frozen Embryo Transfer Cycles. J Assist Reprod Genet 2024, 41, 929–938. [Google Scholar] [CrossRef]
- Itoh, H.; Sashihara, T.; Hosono, A.; Kaminogawa, S.; Uchida, M. Lactobacillus Gasseri OLL2809 Inhibits Development of Ectopic Endometrial Cell in Peritoneal Cavity via Activation of NK Cells in a Murine Endometriosis Model. Cytotechnology 2011, 63, 205–210. [Google Scholar] [CrossRef]
- Ser, H.-L.; Au Yong, S.-J.; Shafiee, M.N.; Mokhtar, N.M.; Ali, R.A.R. Current Updates on the Role of Microbiome in Endometriosis: A Narrative Review. Microorganisms 2023, 11, 360. [Google Scholar] [CrossRef]
- Cai, Z.; Zhou, Z.; Huang, S.; Ma, S.; Chen, Y.; Cao, Y.; Ma, Y. Gut Microbiome in Patients with Early-Stage and Late-Stage Endometriosis. BMC Women’s Health 2025, 25, 163. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Dimitriadis, E. Endometrial Receptivity–Lessons from “Omics”. Biomolecules 2025, 15, 106. [Google Scholar] [CrossRef]
- Xu, Z.; Yang, B.; Shan, W.; Liao, J.; Shao, W.; Wu, P.; Zhou, S.; Ning, C.; Luo, X.; Zhu, Q.; et al. Comparison of the Effect of Levonorgestrel-Intrauterine System with or without Oral Megestrol Acetate on Fertility-Preserving Treatment in Patients with Atypical Endometrial Hyperplasia: A Prospective, Open-Label, Randomized Controlled Phase II Study. Gynecologic Oncology 2023, 174, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Cooper, K.G.; Bhattacharya, S.; Daniels, J.P.; Horne, A.W.; Clark, T.J.; Saridogan, E.; Cheed, V.; Pirie, D.; Melyda, M.; Monahan, M.; et al. Long Acting Progestogens versus Combined Oral Contraceptive Pill for Preventing Recurrence of Endometriosis Related Pain: The PRE-EMPT Pragmatic, Parallel Group, Open Label, Randomised Controlled Trial. BMJ 2024, e079006. [Google Scholar] [CrossRef]
- Zakhari, A.; Delpero, E.; McKeown, S.; Tomlinson, G.; Bougie, O.; Murji, A. Endometriosis Recurrence Following Post-Operative Hormonal Suppression: A Systematic Review and Meta-Analysis. Human Reproduction Update 2021, 27, 96–107. [Google Scholar] [CrossRef]
- Begum, M.R.; Ehsan, M.; Ehsan, N.; Rashid, F. How to Protect Fertility Potential in Endometriosis. Journal of South Asian Federation of Obstetrics and Gynaecology 2024, 16, 134–144. [Google Scholar] [CrossRef]
- Shi, J.; Zhu, S.; Li, X.; Dai, Y.; Leng, J. The Role of Levonorgestrel-Releasing Intrauterine System for Recurrence Prevention after Conservative Surgery among Patients with Coexistent Ovarian Endometrioma and Diffuse Adenomyosis: A Retrospective Case Control Study with Long-Term Follow Up. Journal of Gynecology Obstetrics and Human Reproduction 2023, 52, 102572. [Google Scholar] [CrossRef]
- Gazzo, I.; Moffa, F.; Ferrero, S. Fertility Preservation in Women with Endometriosis: Oocyte Cryopreservation and Other Techniques. Best Practice & Research Clinical Obstetrics & Gynaecology 2024, 95, 102503. [Google Scholar] [CrossRef]
- Nicolì, P.; Viganò, P.; Saccone, G.; Di Naro, E.; Vimercati, A.; Cicinelli, E.; Vitagliano, A. Oocyte Cryopreservation for Patients with Endometriosis: Where Are We Now? A Systematic Review. Reproductive BioMedicine Online 2025, 51, 104839. [Google Scholar] [CrossRef] [PubMed]
- Rangi, S.; Hur, C.; Richards, E.; Falcone, T. Fertility Preservation in Women with Endometriosis. JCM 2023, 12, 4331. [Google Scholar] [CrossRef] [PubMed]
- Benaglia, L.; Fornelli, G.; La Vecchia, I.; Sterpi, V.; Basili, L.; Viganò, P.; Somigliana, E. Elective Oocyte Freezing for Fertility Preservation in Endometriosis: Opportunity or Resource Wastage? Journal of Endometriosis and Uterine Disorders 2023, 1, 100017. [Google Scholar] [CrossRef]
- La Marca, A.; Semprini, M.; Mastellari, E.; Donno, V.; Capuzzo, M.; Alboni, C.; Giulini, S. Fertility Preservation in Women with Endometriosis. Human Reproduction Open 2025, 2025, hoaf012. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Interconnected mechanisms driving infertility in endometriosis.
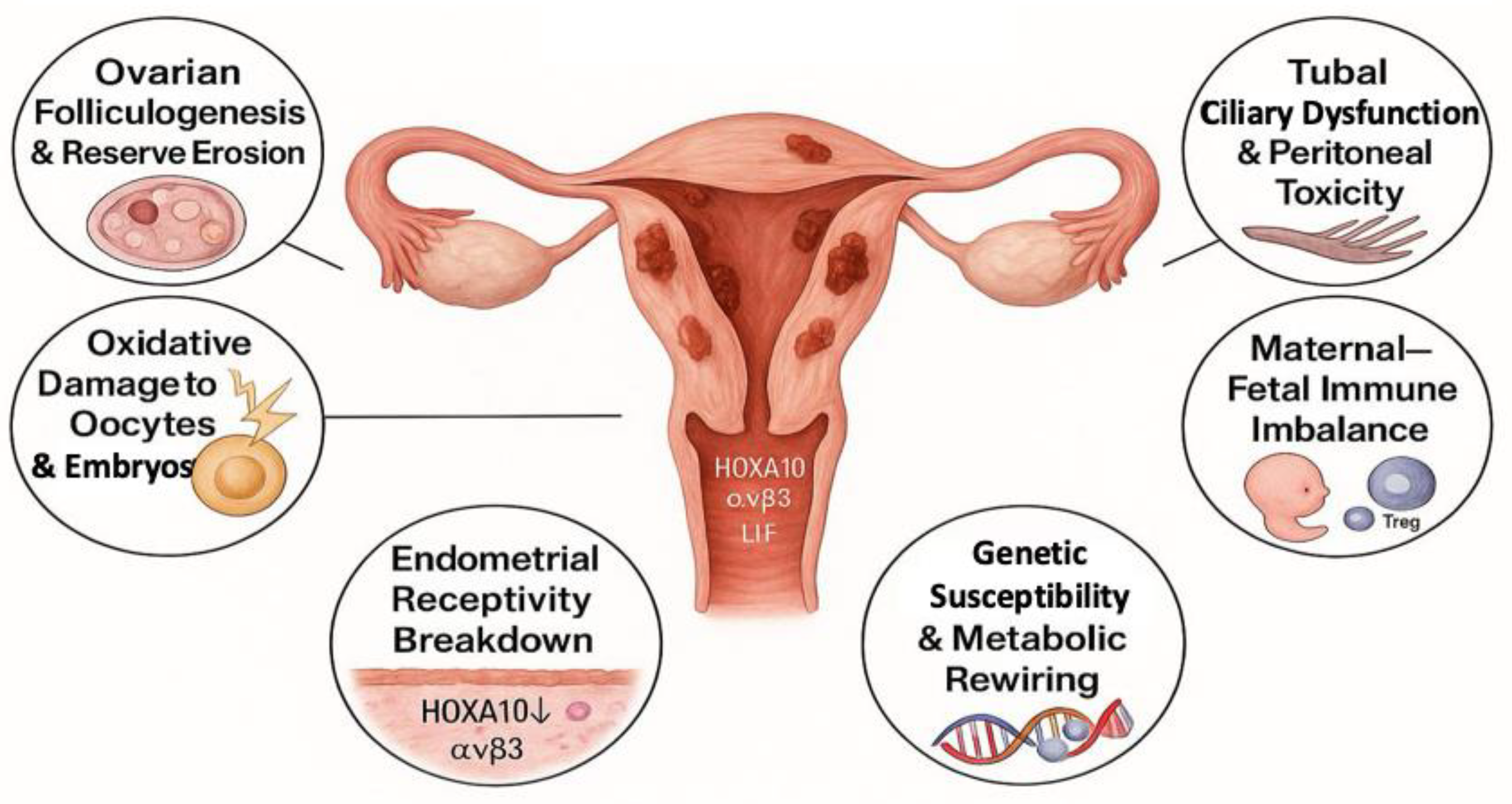
Figure 2.
Molecular landscape of non-hormonal therapies in endometriosis.


Table 1.
Promising molecular targets and their impact on infertility in endometriosis.
| Therapeutic Strategy | Molecular Target/ Mechanism |
Effects on Endometriosis/Fertility | Development Phase | References |
|---|---|---|---|---|
| Selective ERβ antagonists | ERβ-mediated inflammation and progesterone resistance | Reduce lesion survival; restore PR-B signaling; preserve ovulation | Preclinical | [16,17,167,168] |
| Kinase inhibitors (MEK, mTOR, p38, JNK) | NF-κB; MAPK/ERK1/2; PI3K/AKT/mTOR pathways | Decrease lesion growth & inflammation; overcome progesterone resistance | Preclinical | [166,171,173,174,175] |
| Immune-checkpoint blockade | PD-1/PD-L1 and CD47-SIRPα axes | Restore immune clearance; lower lesion burden; enhance tolerance | Preclinical | [23,177,178,179] |
| Anti-TNFα therapy | TNFα-driven inflammatory pathways | Reduce pain, inflammation, and lesion size | Preclinical and early clinical |
[166,180,181] |
| Anti-IL-8 therapy (AMY109) | IL-8 mediated chemotaxis and fibrogenesis | Suppress fibrosis and lesion expansion without disrupting cycles | Phase I clinical trial | [183] |
| Epigenetic modulators (HDACi, DNMTi) | HDACs & DNMTs at HOXA10 and PR-B promoters | Restore decidualization and uterine receptivity | Preclinical | [16,185,186,187,188] |
| Antioxidants (Vit C/E, CoQ10, NAC, melatonin, resveratrol) | ROS; MAPK & NF-κB activation | Improve oocyte/embryo quality; mitigate oxidative injury & inflammation | Preclinical and early clinical | [194,205,213,214] |
| Anti-angiogenic/vascular-normalisation agents | VEGF/VEGFR2 cascade; dopamine D2 agonists (cabergoline, quinagolide); bradykinin B1 | Reduce microvessel density; maintain endocrine function & fertility | Preclinical and Phase 2 clinical trial | [206,207,208,209,210,211,212] |
| miRNA-based therapy (miR-135a/b, miR-451a, miR-214, miR-205-5p) | Post-transcriptional control of HOXA10, VEGF, CTGF, ANGPT2, CTNNB1, etc. | Reprogram gene networks; suppress angiogenesis, invasion, and fibrosis | Preclinical | [44,190,191,192,193] |
| SERMs/SARMs/SARDs | ER-subtype balance; intracrine steroidogenesis (aromatase, steroid sulfatase, SLC10A6, AKR1C3) | Re-calibrate local hormones; improve endometrial receptivity | Preclinical and early-to-advanced clinical (Phase II–III) depending on compound | [169] |
Abbreviations (in alphabetical order): AKR1C3 (Aldo-Keto Reductase Family 1 Member C3); ANGPT2 (Angiopoietin-2); CD47 (Cluster of Differentiation 47); CoQ10 (Coenzyme Q10); CYLD (Cylindromatosis Deubiquitinase); CTGF (Connective Tissue Growth Factor); CTNNB1 (Catenin Beta 1); DNMTi (DNA Methyltransferase Inhibitor); ERβ (Estrogen Receptor Beta); ERK1/2 (Extracellular Signal-Regulated Kinase 1/2); HDACi (Histone Deacetylase Inhibitor); HOXA10 (Homeobox A10); IL-8 (Interleukin-8); JNK (c-Jun N-Terminal Kinase); MAPK (Mitogen-Activated Protein Kinase); MEK (Mitogen-Activated Protein Kinase Kinase); miR (microRNA); MMP9 (Matrix Metalloproteinase-9); mTOR (Mammalian Target of Rapamycin); NAC (N-Acetyl-L-Cysteine); NF-κB (Nuclear Factor κ-Light-Chain-Enhancer of Activated B Cells); PCNA (Proliferating Cell Nuclear Antigen); PD-1 (Programmed Cell Death Protein 1); PD-L1 (Programmed Death-Ligand 1); PI3K (Phosphoinositide 3-Kinase); PR-B (Progesterone Receptor Isoform B); ROS (Reactive Oxygen Species); SARM (Selective Androgen Receptor Modulator); SARD (Selective Androgen Receptor Degrader); SERM (Selective Estrogen Receptor Modulator); SIRPα (Signal-Regulatory Protein Alpha); SLC10A6 (Solute Carrier Family 10 Member 6); TNF-α (Tumor Necrosis Factor Alpha); VEGF (Vascular Endothelial Growth Factor); VEGFR2 (Vascular Endothelial Growth Factor Receptor 2); p38 (p38 Mitogen-Activated Protein Kinase).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



