
Submitted:
21 May 2025
Posted:
21 May 2025
You are already at the latest version
Abstract
A chronic positive energy balance, as observed in obesity, leads to an increase in circulating free fatty acids (FFAs), which can activate inflammatory signaling in M1 macrophages. Nuclear factor erythroid 2-related factor 2 (Nrf2) is a transcription factor that controls the expression of several genes involved in the cellular response to redox changes, xenobiotic detoxification, and adipogenesis. The signaling pathway that leads to the migration of Nrf2 to the cell nucleus is well known, but its role in regulating the adipogenesis process remains controversial. Activation of the Nrf2 signaling pathway leads to increased expression of critical enzymes involved in FFA esterification, such as triglycerides, the safest way to store FFAs. This paper reviews recent information on the role of Nrf2 in the differentiation of preadipocytes into adipocytes, which helps explain controversial information. Adipocytes specialize in storing FFAs as triglycerides and act as endocrine cells. However, in obesity, its functions are dysregulated along with the macrophage-induced inflammation of adipose tissue. Adipose tissue inflammation is the main cause of low-grade systemic inflammation observed in obese patients and the main link between adipose tissue inflammation and obesity-associated insulin resistance. Therefore, a deeper understanding of the role of Nrf2 in the process of adipogenesis is essential for the development of new agonists to prevent metabolic abnormalities associated with obesity.
Keywords:
Nrf2
; signaling pathway
; gene expression
; adipogenesis
; obesity
; insulin resistance
Introduction
Obesity is a chronic and progressive disease that affects a large proportion of the global population. In Latin America, it is increasing at a faster rate than in the rest of the world, particularly among individuals with low income [1,2]. In Argentina, the prevalence of obesity is estimated to be approximately 35% [3]. Obesity leads to various metabolic abnormalities, including metabolic syndrome (MS) (central obesity, insulin resistance (IR), dyslipidemia, hypertension, fatty liver, and a systemic state of oxidative stress and inflammation) [4]. Obesity, characterized by the excessive storage of fatty acids in adipose tissue, is strongly associated with diabetes mellitus, stroke, hyperlipidemia, cancer, cardiovascular disease, and osteoarthritis [5]. Type 2 diabetes, one of the most widespread metabolic diseases, and obesity are closely related due to their association with IR. As a state of chronic inflammation, obesity is a well-known risk factor for the onset and development of IR [6].
Inflammation is a process that is characterized by the activation of immune and nonimmune cells that protect the host, depending on the degree and extent of the inflammatory response; that is, whether it is systemic or local, metabolic and neuroendocrine changes can occur. The clinical consequences of chronic low-grade inflammation include an increased risk of metabolic syndrome [7]. Obesity-induced IR is strongly associated with white adipose tissue (WAT) and systemic chronic low-grade inflammation [8]. An extended hyperglycemic condition leads to a periodic and sustained increase in reactive oxygen species (ROS) production and elevates the levels of free fatty acids (FFAs) released by WAT from obese individuals, promoting further ROS production [9]. In turn, the overproduction of ROS mediated by hyperglycemia and the increase in ROS
generation in obese WAT leads to the secretion of metabolically adverse patterns of adipocytokines and causes inflammatory responses, which contribute to IR through the interaction between inflammatory molecules (certain adipocytokines) and insulin signaling. Collectively, they can serve as both a cause and a result of each other, thus forming a vicious cycle [9].
Nuclear factor erythroid 2-related factor 2 (Nrf2) is a transcription factor that plays a crucial role in maintaining redox and metabolic homeostasis by regulating cellular antioxidants and decreasing inflammatory stress [7]. Since obesity is closely associated with inflammation and oxidative stress, the potential protective function of Nrf2 is of great interest [7]. However, the effects of Nrf2 are controversial and even contradictory. Therefore, understanding the interactions among obesity, inflammation, and oxidative stress in the pathogenesis of IR is a critical area of study [7].
Nrf2 Structure
The structure of Nrf2 is complex and multifunctional, allowing it to effectively regulate the cellular antioxidant response. It contains 7 domains (Figure 1): Neh1 (N-terminal) contains a basic leucine-zipper region (bZIP) crucial for dimerization with small Maf factors and DNA binding in antioxidant response elements (AREs); Neh2 regulates the stability of Nrf2 by interacting with Kelch-like ECH-associated protein 1 (Keap1), which facilitates the ubiquitination and proteasomal degradation of Nrf2 under normal conditions; Neh3 participates in the transactivation of target genes by interacting with transcription coactivators; Neh4 and Neh5 cooperate in gene transactivation by interacting with coactivating proteins such as CBP/p300; and Neh6 facilitates the degradation of Nrf2 independently of Keap1, playing an additional role in regulating Nrf2 stability; and Neh7 interacts with retinoid receptor X (RXRα) and represses Nrf2 transcriptional activity. These domains allow Nrf2 to play a master regulator of antioxidant and detoxification genes, coordinating an integral cellular response to oxidative stress [10,11].
Nrf2 Signaling Pathway
Under conditions of homeostasis, Nrf2 is inhibited by its binding to Keap1, which is an adaptor subunit of the ubiquitin ligase E3 and is responsible for adding ubiquitin chains to Nrf2 as a marker for its degradation in the proteasome [13]. Under oxidative stress conditions, oxidants bind to keap1 cysteine sensors, causing a conformational change that allows Nrf2 to dissociate from the inhibitory protein, stabilize, and translocate to the nucleus [13]. Once it is present, it forms a complex via heterodimerization with musculoaponeurotic
fibrosarcoma proteins (Figure 2). These heterodimers recognize antioxidant response elements (AREs) located in the DNA-promoting region of genes encoding a number of genes, including those encoding phase II antioxidant enzymes [14,15]. Nrf2 transduces chemical signals to regulate a battery of cytoprotective genes. The target genes include superoxide dismutase (SOD), catalase, glutathione S-transferase (GST), nicotinamide adenine dinucleotide phosphate (NADPH)-oxidase, quinone oxidoreductase 1 (NQO1), γ-glutamate cysteine ligase (GCL), and heme oxygenase-1 (HO-1) [16] (Figure 3).
Nrf2 downstream genes identified thus far in humans and mice can be grouped into several categories (Figure 3), including genes encoding (i) intracellular redox-balancing proteins that maintain cellular glutathione and thioredoxin levels and reduce ROS levels, such as glutamate cysteine ligases (GCLC, GCLM), glutathione peroxidase (GPx), thioredoxin 1 (TXN1), thioredoxin reductase (TXNR), peroxiredoxin 1 (PRDX1), and sulfiredoxin 1 (SRXN1); (ii) phases I/II/III detoxifying enzymes that metabolize xenobiotics into less toxic forms or catalyze conjugation reactions to increase the solubility of xenobiotics, thereby facilitating their elimination, such as glutathione S-transferases (GSTA1, GSTM1), NAD(P)H quinone oxidoreductase-
1 (NQO1), UDP-glucuronosyltransferase (UGT), transporters (ABCB6, ABCC2), aldo-keto reductases (AKR1B10, AKR1C1, and AKR1C3); (iii) enzymes involved in heme metabolism, such as heme oxygenase-1 (HMOX-1) and ferrochelatase (FECH); and (iv) enzymes involved in lipid metabolism, such as acyl-CoA wax alcohol acyltransferase 1 (AWAT1).
The Role of Nrf2 in the Adipogenesis Process
Adipogenesis is a process by which precursor cells, preadipocytes, differentiate into mature adipocytes, which are the cells responsible for storing fat in the body [19,20]. Morphologically, mature WAT adipocytes
carry large lipid droplets but few mitochondria within the cell [21]. This process is crucial for the development of adipose tissue and has important implications for energy metabolism, homeostasis, and metabolic diseases such as obesity and diabetes [22,23].
Energy intake above expenditure leads to a chronic positive energy balance that culminates in obesity [17,24]. The unspent energy is used for the synthesis of de novo triglycerides that are stored in adipose tissue. Excess triglycerides (TGs) in the body are transferred to the liver, skeletal muscle, and other nonadipose tissues as FFAs, resulting in ectopic lipid deposition (ELD) [25]. Ectopic lipid deposition in the liver and skeletal muscle is associated with lipotoxicity, insulin resistance, and metabolic abnormalities [26]. There is adipocyte heterogeneity in individual adipose tissue deposits, with subpopulations exhibiting more proinflammatory phenotypes; this heterogeneity may influence adipose tissue health and susceptibility to metabolic diseases [27,28].
During excessive fat intake, adipose tissue expands by hyperplasia, is enlarged by hypertrophy, or both, depending on several factors, including the location of the adipose deposit [17]. The distribution of white adipose tissue is a predictor of metabolic health in obese individuals [27]. People who expand mostly intra-abdominal adipose tissue are at greater risk of developing metabolic syndrome than those who accumulate subcutaneous white adipose tissue [27].
During adipogenesis, MSCs from adipose tissue become preadipocytes under the influence of growth factors and hormonal signals, such as insulin- like growth factor 1 (IGF-1) and growth hormone, and the differentiation process consists of several phases [29] (Figure 4). Preadipocytes have many more IGF-1 receptors than insulin receptors do. Both IGF-1 and insulin bind to IGF-1 receptors to induce preadipocyte differentiation [30]. Insulin can bind to these receptors, but it only does so at high concentrations; therefore, under physiological conditions, IGF-1 is the inducer of differentiation. Growth hormones can induce the autocrine or paracrine secretion of IGF-1; the secretion of IGF-1 at concentrations lower than insulin is capable of inducing differentiation, which demonstrates its relevance in this process [31]. In addition, IGF-1 is able to counteract the inhibitory effects of the Pref-1 protein, which is a natural inhibitor of differentiation [32].
During early differentiation, preadipocytes proliferate in response to mitogenic signals [23]. During this phase, 3T3-L1 fibroblasts are still undifferentiated cells, and signals such as fetal bovine serum, insulin, glucocorticoids (such as dexamethasone) and isobutylmethylxanthine (IBMX) induce the differentiation of preadipocytes [33]. These signals activate a cascade of molecular events, including activation of the PPARγ pathway and the C/EBP pathway [23]. In terms of regulation, a key gateway to adipogenic gene expression that drives preadipocyte differentiation is the PPAR family of nuclear proteins, particularly the PPARγ isoform [21,30]. During intermediate differentiation, key transcription factors, such as PPARγ and C/EBPα, which are essential for the progression of the adipogenic process, are expressed [23]. These transcription factors regulate the expression of specific genes that are necessary for adipocyte maturation [23]. Preadipocytes begin to accumulate lipids in small droplets within the cell, and this accumulation is facilitated by lipogenic enzymes such as acetyl-CoA carboxylase (ACC) and fatty acid synthase (FAS) [27].
During late differentiation and maturation, mature adipocytes are characterized by the accumulation of large and unilocular lipid droplets. Mature fat cells also express a variety of adipocyte- specific proteins, such as adiponectin and leptin, which play critical roles in regulating metabolism and energy homeostasis [34].
Mature adipocytes specialize in storing energy in the form of triglycerides and releasing free fatty acids during periods of fasting or increased energy demand [22,23]. Adipogenesis and its regulation are governed mainly by mitochondrial homeostasis and intracellular levels of ROS, with variable effects attributed to the latter [17,36] (Figure 5). The maturation of preadipocytes is associated with increased mitochondrial mass and a doubling of oxygen consumption [36]. In conclusion, increased oxidative stress in adipocytes decreases adipogenesis and adipokine secretion, leading to an imbalance in energy homeostasis, insulin resistance, and type 2 diabetes [17].
Nrf2 in the Process of s
Fibroblast-like MSCs express growth factor receptors that lead them to become preadipocytes and eventually mature insulin-sensitive adipocytes [37]. Terminal differentiation is regulated by transcription factors such as PPARγ and C/EBPα, leading to the arrest of cell growth and lipid accumulation [17]. These processes are coordinated by intricate networks of transcription factors (Figure 6), which are responsible for establishing the mature adipocyte phenotype [17].
Nrf2 is an important player in PDGFRα signaling and mediates PDGF-A expression and adipogenesis [39]. Oxidative stress can promote the recruitment of Nrf2 to the SREBP1 promoter, inducing target gene transcription and subsequent lipogenesis [40]. Nrf2 expression significantly affects adipogenesis, as adipocyte differentiation is inhibited in Nrf2 knockout (KO) mice, with downregulation of PPARγ and C/EBPα expression induced by a high-fat diet (HFD) [41,42]. Suppressing Nrf2 activity, either genetically or chemically, leads to impaired adipogenesis in 3T3-L1 preadipocytes, primary mouse embryonic fibroblasts, and/or human subcutaneous preadipocytes [43]. A study revealed that leptin deficient (ob-/ob-) mice with whole-body or adipocyte-specific Nrf2 ablation presented reduced white adipose tissue mass but exhibited more severe metabolic syndrome characterized by hyperlipidemia, worsened insulin resistance, and hyperglycemia [44]. The deletion of Nrf2 in adipocytes led to a worsened diabetic phenotype, whereas the deletion of Nrf2 in hepatocytes modestly reduced insulinemia after HFD-induced obesity, although hepatic lipid accumulation was not affected [45]. These data suggest that Nrf2 is a key transcription factor controlling the development and function of white adipose tissue, thereby affecting insulin sensitivity, glucose tolerance, and lipid homeostasis.
Adipose Tissue: Hyperplasia and Hypertrophy
Adipose tissue plays an important role in regulating metabolism and body homeostasis [46]. Unlike other organs, it can expand to accommodate excess energy in the form of lipids. It is divided into two main types, white adipose tissue (WAT) and brown adipose tissue; WAT deposits are characterized as visceral or subcutaneous. Brown adipose tissue is a thermogenic tissue that is perfused by an extensive network of blood capillaries and highly innervated by noradrenergic fibers [47,48]. A third type of adipocyte called beige adipocytes is essential for weight control, the regulation of energy balance and the improvement of glucose and lipid metabolism [48,49].
Adipocytes respond naturally to excess nutrient intake in obese individuals by increasing in number (hyperplasia) and size (hypertrophy) [50]. In obese individuals, blood flow to adipose tissue does not increase relative to that in lean individuals. With the rapid expansion of adipose tissue in obesity, the blood supply to adipocytes can become insufficient, leading to hypoxia [51]. In fact, hypoxia has been proposed to be one of the main triggers of adipose tissue remodeling, including adipocyte death and the inflammatory response in obesity [7].
The lipids of visceral adipose tissue are composed of 90% triglycerides and only 3% phospholipids and sphingolipids [52]; changes in phospholipid composition appear to play an important role in the development of metabolic diseases [53]. Phospholipids containing 18-carbon acyl chains in visceral adipose tissue have been identified as major lipid species that distinguish insulin-sensitive individuals from obese patients with IR [46,54].
However, obesity, which is associated with impaired lipid metabolism, is characterized by a lower turnover of triglycerides in adipocytes, since it is estimated that they are only replaced 3 times during the lifespan of adipocytes [55].
The Adipocyte as an Endocrine Cell
Not only does adipose tissue function as an energy storage organ, but it also performs important endocrine functions in the body through the secretion of biologically active compounds that regulate metabolic homeostasis [56,57]. The first discovery, established by Zhang, Y. et al. [58] Leptin, a peptide hormone secreted primarily by adipocytes, is essential for body weight control and other biological functions [58]. In addition to adipocytes, adipose tissue also contains several types of cells, such as immune cells, fibroblasts, and endothelial cells, which secrete different types of hormones, such as peptide hormones (adipokines) and bioactive lipids (lipokines), to regulate appetite and glucose and lipid metabolism [59]. In a healthy state, endocrine factors secreted by adipose tissue maintain organ and metabolic homeostasis. However, in obesity, proinflammatory profiles with abnormal secretion of adipokines and lipokines from adipocytes and immune cells from adipose tissue contribute to the occurrence of chronic inflammation and oxidative stress [60].
Inflammation of Adipose Tissue and Insulin Resistance Associated with Obesity
Insulin is a hormone produced by the β cells of the pancreatic islets that regulates the metabolism of carbohydrates, fats, and proteins [61]. Blood glucose stimulates insulin synthesis and release; its effects on whole-body metabolism result from its binding to the cell membrane receptor, which is activated by autophosphorylation of specific tyrosine residues [62].
Insulin resistance is a pathological condition in which insulin-dependent cells, such as skeletal muscle, liver, and adipocytes, do not respond adequately to normal circulatory insulin levels [7]. IR is characterized by severe impairments in glucose uptake, glycogen synthesis, and, to a lesser extent, glucose oxidation [63]. In skeletal muscle, insulin promotes glucose uptake by stimulating the translocation of glucose transporter 4 (GLUT4) to the plasma membrane, and impaired insulin signaling in skeletal muscle results in decreased removal of glucose from the blood [64]. In the liver, insulin inhibits the expression of key gluconeogenic enzymes; therefore, insulin resistance leads to elevated hepatic glucose production [65]. In adipose tissue, insulin-regulated GLUT4 levels are reduced in individuals with insulin-resistant obesity [66]. In addition, insulin decreases lipase activity and inhibits the outflow of free fatty acids from adipocytes [66].
Inflammation and insulin resistance can be closely related to each other through the production of proinflammatory cytokines and chemokines [7,66]. The infiltration of inflammatory cells into adipose tissues leads to increased secretion of proinflammatory cytokines and chemokines, such as TNFα, MCP-1, C- reactive protein (CRP), and interleukins, and these same inflammatory mediators have been shown to increase insulin resistance [67,68].
Activation of the JNK and NF-κB signaling pathways increases the production of proinflammatory cytokines, endothelial adhesion molecules, and chemotactic mediators that promote monocyte infiltration into adipose tissue and differentiation into proinflammatory M1 macrophages [69]. Infiltrated macrophages produce and secrete many inflammatory mediators that promote local and systemic proinflammatory states and impair insulin signaling [62].
Activation of the Nrf2 Signaling Pathway and Metabolic Risk Associated with Obesity
An experiment was conducted with wild-type (WT) and ob-/ob-type mice fed a high-fat diet (HFD) or a normal diet (ND) for 16 weeks. The protein levels of Nrf2 and nuclear Nrf2, as well as the mRNA levels of Nrf2- related genes, such as superoxide dismutase (SOD) and glutathione peroxidase (GPX), tended to increase in the white adipose tissue (WAT) of ob/ob mice compared with those of control mice [70].
In addition, previous studies have revealed that, compared with WT mice, ob-/ob- mice exhibit increased expression of heme oxygenase 1 (HO1) mRNA and NAD(P)H: quinone oxidoreductase 1 (NQO1) in the liver and HO1 in white adipose tissue [71]. These findings suggest that obesity may increase oxidative stress and that Nrf2 may play a role in regulating the adaptive antioxidant response in obesity [72]. Disruption of Nrf2 can negatively affect this adaptive antioxidant response [45,73,74].
However, conflicting results from previous studies on the transcription factor Nrf2 and its response to obesity induced by a high-fat diet (HFD) have been reported. Some studies have shown that the mRNA expression levels of Nrf2 and its targets, such as glutathione transferase m6 (GSTm6) and NQO1, are reduced after 4 weeks of HFD feeding in wild-type (WT) mice [75].
In addition, other studies have shown that Nrf2 total and nuclear protein levels in skeletal muscle are nearly 50% lower in mice fed a HFD for 18 weeks than in those fed a normal diet (ND) [76]. In WAT, the mRNA levels and DNA-binding activities of Nrf2 decreased significantly after 12 weeks of exposure to a HFD [74].
On the other hand, a 12-week HFD did not have a substantial effect on Nrf2 mRNA levels or the levels of its target genes, NQO1 and HO1, in the livers of WT mice or specific mice with Nrf2 deficiency in hepatocytes [77,78]. In addition, in another study, HFD feeding significantly increased the levels of nuclear, but not total, Nrf2 protein in the liver [79]. The functional suppression of Nrf2 in response to a high-fat diet (HFD) could be implicated in obesity and steatohepatitis in mice [80]. However, the possibility that Nrf2 activation may reduce these effects has also been suggested [16,45,74]. In general, there is evidence of complexity in the regulation of Nrf2 in the context of HFD-induced obesity, with different studies showing contradictory results and complex interactions between Nrf2 and the development of obesity and its associated effects.
These contradictory results concerning the activation of the Nrf2 pathway could be due to the composition of the diet supplied and the duration of the diet. In response to obesity, a certain degree of Nrf2 activation exerts cellular-protective effects through antioxidant stress, but it can severely disrupt redox homeostasis and aggravate obesity if its activation exceeds the scope or duration [9]. Nrf2 activators are always protective, alleviate obesity and relatedmetabolic diseases, decrease ROS production, inhibit lipid accumulation during adipogenesis, attenuate proinflammatory cytokines, and improve glucose homeostasis [4]. In the future, developing effective and safe Nrf2 activators for obesity therapy may be possible.
Role of Nrf2 Pathway Agonists
Several compounds have been identified for their ability to activate the Nrf2 signaling pathway in adipose tissue [81]. These compounds reduce oxidative stress and inhibit lipid accumulation and de novo adipogenesis by acting on adipogenic factors [17]. The structures of some of these compounds are depicted in Table 1.
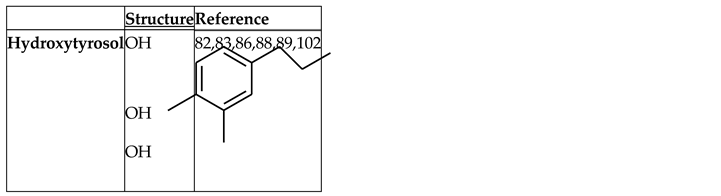
Hydroxytyrosol
Hydroxytyrosol (HT) is a phenolic phytochemical of natural origin in the olive tree and an effective antioxidant [82]. As a hydrophilic compound, it can be recovered from olive oil residues and byproducts through sustainable processes such as membrane technologies [83,847]. The presence of a phenolic group in the oil that is capable of reducing ROS [82] endows it with redox properties. It has potential therapeutic effects and exhibits anticancer, cardioprotective, antidiabetic, anti-inflammatory and neuroprotective activities [83,85], as well as a preventive effect on metabolic syndrome in obese people by reducing fat deposition in the liver and muscle, enhancing antioxidant capacity and improving mitochondrial function [86] (Figure 7). In a study conducted on mice, a diet with olive oil resulted in greater weight gain, an increase in subcutaneous fat, higher serum levels of anti-inflammatory cytokines (TGF-β), and Nrf2 expression in adipose tissue than other oils (coconut and sunflower) [87]. In contrast, Cao and colleagues demonstrated that the administration of HT inhibits
weight gain and organ enlargement; both low- and high-dose treatments improved glucose tolerance and reduced the serum levels of glucose, insulin, lipids, and inflammatory cytokines [88].
Figure 7.
Principal Cytoprotective beneficial effects of hydroxytyrosol (modified from [86]).
Figure 7.
Principal Cytoprotective beneficial effects of hydroxytyrosol (modified from [86]).
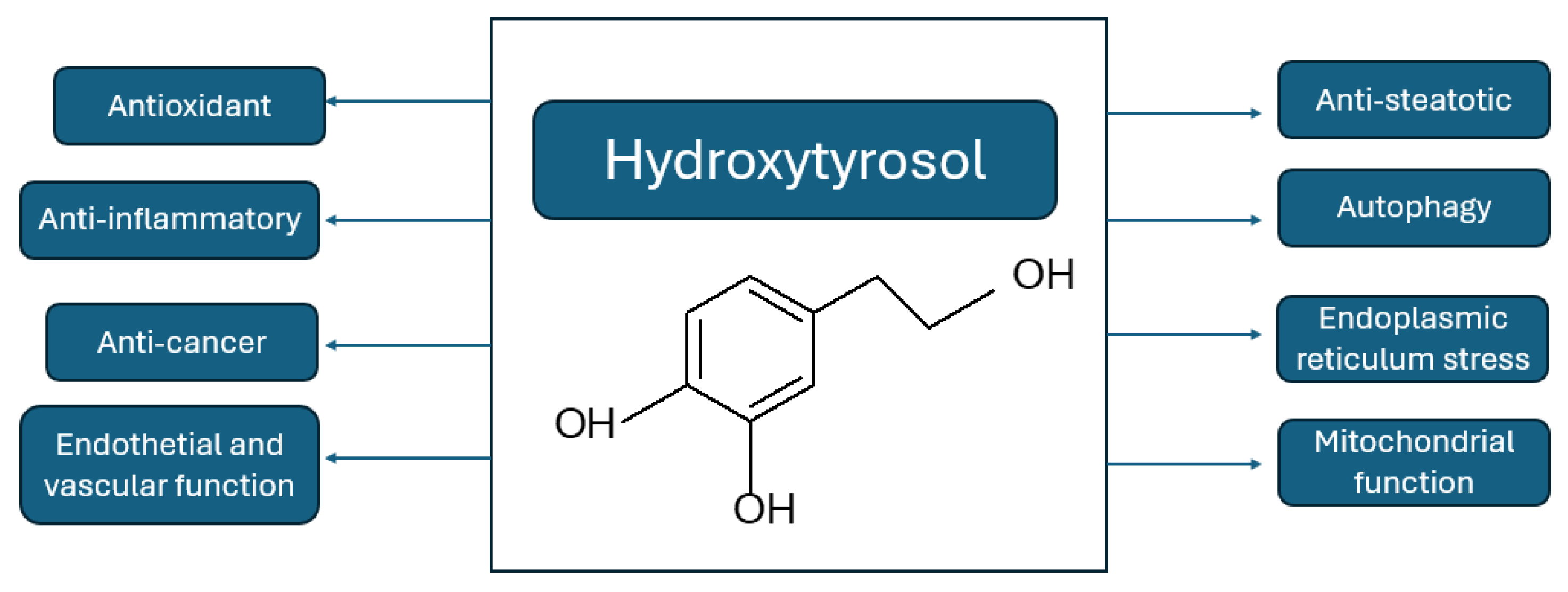
Several in vivo studies have used HT to improve metabolic conditions in obese and overweight mice [89]. The administration of HT to HFD-fed mice reduces adipose tissue dysfunction by increasing Nrf2 and PPARγ expression and insulin sensitivity, thereby improving mitochondrial function and minimizing oxidative stress, endoplasmic reticulum stress, and lipid accumulation [89]. Similar results were observed with the combined administration of HT and docosahexaenoic acid [17]. Hydroxytyrosol has antioxidant, anti-inflammatory, and anticancer effects; improves endothelial and vascular function; has pathway in response to the regulation of antioxidant and phase II detoxification enzymes and nuclear factor-kappa β (NF-κβ) in different organs, particularly in liver tissue [93,94,95].
GT may delay the development or progression of metabolic diseases such as hypertension and cardiovascular disorders [96,97]. The consumption of green and black tea reduces body weight and exerts lipolytic and antiadipogenic effects, as well as insulin-sensitizing activity [98]. In green tea, these effects have been attributed to the presence of monomeric catechins such as epicatechin, antisteatotic properties; and improves endoplasmic epigallocatechin, epicatechin gallate, and reticulum stress, autophagy, and mitochondrial function.
Green and Black Tea Extract
The health benefits ascribed to the consumption of teas are thought to be associated with their high content of bioactive ingredients such as polyphenols. The latter are secondary plant metabolites and include subclasses of flavonoids, flavones, flavonols, flavanols, isoflavones, flavanones and anthocyanidins [90]. Among the polyphenols, tea flavanols, catechins and theaflavins have been identified as the bioactive phytochemicals of green tea (GT) and black tea (BT), respectively, and have been shown to be responsible for their antioxidant activity [91,92]. Most of the biological effects of GTPs are attributed to their ability to transcriptionally upregulate the Nrf2 signaling affecting IL-6 or CRP levels. However, subgroup analysis revealed an increase in CRP levels in studies with a duration of 8 weeks or less, and meta- regression analysis revealed a significant association epigallocatechin gallate, whereas in black tea, these effects are due to arubigins and mainly to theaflavins [98,99].
In terms of insulin resistance, supplementation with a standardized extract of green and black tea (ADM® Complex Tea Extract (CTE)) significantly increased plasma adiponectin concentrations and reduced circulating insulin levels and Homeostatic Model Assessment of Insulin Resistance (HOMA-IR) [100]. In skeletal muscle, CTE- treated mice presented elevated levels of aryl hydrocarbon receptor (Ahr), Arnt, and Nrf2 mRNAs, suggesting that the insulin-sensitizing effects of CTE could be the result of activation of this pathway [100].
The analysis conducted by Souza and colleagues showed that green tea consumption significantly reduced serum TNF-α levels without between increased IL-6 concentrations and the duration of green tea treatment [101].
Concluding Remarks
This review highlights the crucial role of the Nrf2 signaling pathway in the regulation of adipogenesis, oxidative stress, and inflammation in the context of obesity. Nrf2 influences the differentiation of preadipocytes into mature adipocytes, thus controlling fat storage and metabolic function. However, the chronic low-grade inflammation induced by obesity and insulin resistance complicates this regulatory function, leading to systemic metabolic alterations. Although some evidence supports the protective role of Nrf2 against metabolic abnormalities through an enhanced antioxidant response, contradictory findings emphasize the complexity of Nrf2 activation in the management of obesity. Agonists of the Nrf2 signaling pathway induce PPARγ and C-EBPα expression, leading to the expression of critical enzymes involved in TG biosynthesis. This in turn reduces circulating inflammogenic FFAs, thus reducing the phenotypic switch of macrophages toward M1 macrophages within AT and other tissues, as well as systemic inflammation, and reducing metabolic risk in obesity, such as insulin resistance. Future research should focus on developing safe and effective synthetic Nrf2 activators, to modulate adipogenesis, reduce oxidative stress and inflammation, and mitigate insulin resistance associated with obesity.
Acknowledgments
This research was supported by the Agencia Nacional para la Promoción de la Ciencia y la Technología, FONCYT (PICT-2021-I-A-0147 to RDC), Ministerio. de Capital Humano, República Argentina.
References
- AR, J. et al. Health Effects of Overweight and Obesity in.
- 195 Countries over 25 Years. Yearbook of Pediatric Endocrinology (2018). [CrossRef]
- Halpern, B. et al. Obesity and COVID-19 in Latin America: A tragedy of two pandemics—Official document of the Latin American Federation of Obesity Societies. Obesity Reviews vol. 22 Preprint at (2021). [CrossRef]
- Filozof, C., Gonzalez, C., Sereday, M., Mazza, C. & Braguinsky, J. Obesity prevalence and trends in Latin- American countries. Obesity Reviews 2, (2001). [CrossRef]
- Vasileva, L. V., Savova, M. S., Amirova, K. M., Dinkova- Kostova, A. T. & Georgiev, M. I. Obesity and NRF2- mediated cytoprotection: Where is the missing link? Pharmacological Research vol. 156 Preprint at (2020). [CrossRef]
- Lin, X. & Li, H. Obesity: Epidemiology, Pathophysiology, and Therapeutics. Frontiers in Endocrinology vol. 12 Preprint at (2021). [CrossRef]
- Barazzoni, R., Gortan Cappellari, G., Ragni, M. & Nisoli,.
- E. Insulin resistance in obesity: an overview of fundamental alterations. Eating and Weight Disorders vol. 23 Preprint at (2018). [CrossRef]
- Li, S., Eguchi, N., Lau, H. & Ichii, H. The role of the nrf2 signaling in obesity and insulin resistance. International Journal of Molecular Sciences vol. 21 Preprint at (2020). [CrossRef]
- Wondmkun, Y. T. Obesity, insulin resistance, and type 2 diabetes: Associations and therapeutic implications. Diabetes, Metabolic Syndrome and Obesity vol. 13 Preprint at (2020). [CrossRef]
- Xia, Y., Zhai, X., Qiu, Y., Lu, X. & Jiao, Y. The Nrf2 in Obesity: A Friend or Foe? Antioxidants vol. 11 Preprint at (2022). [CrossRef]
- Kim, M.-J. & Jeon, J.-H. Recent Advances in Understanding Nrf2 Agonism and Its Potential Clinical Application to Metabolic and Inflammatory Diseases. Int J Mol Sci 23, 2846 (2022). [CrossRef]
- Ulasov, A. V., Rosenkranz, A. A., Georgiev, G. P. & Sobolev, A. S. Nrf2/Keap1/ARE signaling: Toward specific regulation. Life Sciences vol. 291 Preprint at (2022). [CrossRef]
- Namani, A., Li, Y., Wang, X. J. & Tang, X. Modulation of NRF2 signaling pathway by nuclear receptors: Implications for cancer. Biochimica et Biophysica Acta - Molecular Cell Research vol. 1843 Preprint at (2014). [CrossRef]
- Bruneska Gondim Martins, D., Walleria Aragão Santos, T., Helena Menezes Estevam Alves, M. & Ferreira Frade de Araújo, R. The Role of NRF2 Transcription Factor in Metabolic Syndrome. in The Role of NRF2 Transcription Factor [Working Title] (IntechOpen, 2024). [CrossRef]
- Romano-Lozano, V., Cruz-Avelar, A. & Peralta Pedrero,.
- M. L. Factor nuclear eritroide similar al factor 2 en el vitíligo. Actas Dermosifiliogr 113, 705–711 (2022).
- Da Costa, R. M. et al. Nrf2 as a potential mediator of cardiovascular risk in metabolic diseases. Frontiers in Pharmacology vol. 10 Preprint at (2019). [CrossRef]
- Zhang, Z. et al. The role of the Nrf2/Keap1 pathway in obesity and metabolic syndrome. Reviews in Endocrine and Metabolic Disorders vol. 16 Preprint at (2015). [CrossRef]
- Annie-Mathew, A. S. et al. The pivotal role of Nrf2 activators in adipocyte biology. Pharmacological Research vol. 173 Preprint at (2021). [CrossRef]
- Menegon, S., Columbano, A. & Giordano, S. The Dual Roles of NRF2 in Cancer. Trends in Molecular Medicine vol. 22 Preprint at (2016). [CrossRef]
- GREGOIRE, F. M., SMAS, C. M. & SUL, H. S.
- Understanding Adipocyte Differentiation. Physiol Rev 78, 783–809 (1998).
- Ambele, M. A., Dhanraj, P., Giles, R. & Pepper, M. S. Adipogenesis: A complex interplay of multiple molecular determinants and pathways. International Journal of Molecular Sciences vol. 21 Preprint at (2020). [CrossRef]
- Huang, C. J. & Choo, K. B. Circular RNA- and microRNA-Mediated Post-Transcriptional Regulation of Preadipocyte Differentiation in Adipogenesis: From Expression Profiling to Signaling Pathway. International Journal of Molecular Sciences vol. 24 Preprint at (2023). [CrossRef]
- Rosen, E. D. & Spiegelman, B. M. Molecular regulation of adipogenesis. Annual Review of Cell and Developmental Biology vol. 16 Preprint at (2000). [CrossRef]
- Moseti, D., Regassa, A. & Kim, W. K. Molecular regulation of adipogenesis and potential anti-adipogenic bioactive molecules. International Journal of Molecular Sciences vol. 17 Preprint at https://doi.org/10.3390/ijms17010124 (2016). [CrossRef]
- Romieu, I. et al. Energy balance and obesity: what are the main drivers? Cancer Causes and Control 28, (2017).
- Su, K. et al. Liraglutide attenuates renal tubular ectopic lipid deposition in rats with diabetic nephropathy by inhibiting lipid synthesis and promoting lipolysis. Pharmacol Res 156, (2020).
- Jia, Y. et al. Increased FGF-21 Improves Ectopic Lipid Deposition in the Liver and Skeletal Muscle. Nutrients 16, 1254 (2024). [CrossRef]
- Vishvanath, L. Vishvanath, L. & Gupta, R. K. Contribution of adipogenesis to healthy adipose tissue expansion in obesity. Journal of Clinical Investigation vol. 129 Preprint at (2019). [CrossRef]
- Lee, K. Y. et al. Developmental and functional heterogeneity of white adipocytes within a single fat depot. EMBO J 38, (2019).
- Cristancho, A. G. & Lazar, M. A. Forming functional fat: A growing understanding of adipocyte differentiation. Nature Reviews Molecular Cell Biology vol. 12 Preprint at (2011). [CrossRef]
- Ambele, M. A., Dhanraj, P., Giles, R. & Pepper, M. S. Adipogenesis: A Complex Interplay of Multiple Molecular Determinants and Pathways. Int J Mol Sci 21, 4283 (2020). [CrossRef]
- MacDougald, O. A. & Lane, M. D. Transcriptional regulation of gene expression during adipocyte differentiation. Annu Rev Biochem 64, (1995).
- Zhang, H. et al. Insulin-like growth factor-1/insulin bypasses Pref-1/FA1-mediated inhibition of adipocyte differentiation. Journal of Biological Chemistry 278, (2003).
- Ailhaud, G. Early adipocyte differentiation. Biochem Soc Trans 24, 400–402 (1996).
- Kumar, R. et al. Association of Leptin With Obesity and Insulin Resistance. Cureus (2020). [CrossRef]
- Al-Mansoori, L., Al-Jaber, H., Prince, M. S. & Elrayess,.
- M. A. Role of Inflammatory Cytokines, Growth Factors and Adipokines in Adipogenesis and Insulin Resistance.
- Inflammation vol. 45 Preprint at (2022). [CrossRef]
- Lee, J. H. et al. The role of adipose tissue mitochondria: Regulation of mitochondrial function for the treatment of metabolic diseases. International Journal of Molecular Sciences vol. 20 Preprint at (2019). [CrossRef]
- Matsushita, K. & Dzau, V. J. Mesenchymal stem cells in obesity: Insights for translational applications. Laboratory Investigation vol. 97 Preprint at (2017). [CrossRef]
- Guo, L., Li, X. & Tang, Q. Q. Transcriptional regulation of adipocyte differentiation: A central role for CCAAT/enhancer-binding protein (C/EBP) β. Journal of Biological Chemistry vol. 290 Preprint at (2015). [CrossRef]
- Haider, N. & Larose, L. Activation of the PDGFRα-Nrf2 pathway mediates impaired adipocyte differentiation in bone marrow mesenchymal stem cells lacking Nck1. Cell Communication and Signaling 18, (2020). [CrossRef]
- Sun, X. et al. Nuclear Factor E2-Related Factor 2 Mediates Oxidative Stress-Induced Lipid Accumulation in Adipocytes by Increasing Adipogenesis and Decreasing Lipolysis. Antioxid Redox Signal 32, (2020).
- Pi, J. et al. Deficiency in the nuclear factor E2-related factor-2 transcription factor results in impaired adipogenesis and protects against diet-induced obesity. Journal of Biological Chemistry 285, (2010).
- Kim, B. R. et al. Suppression of Nrf2 attenuates adipogenesis and decreases FGF21 expression through PPAR gamma in 3T3-L1 cells. Biochem Biophys Res Commun 497, (2018).
- Chen, Y. et al. Isoniazid suppresses antioxidant response element activities and impairs adipogenesis in mouse and human preadipocytes. Toxicol Appl Pharmacol 273, (2013).
- Xue, P. et al. Adipose deficiency of Nrf2 in ob/ob mice results in severe metabolic syndrome. Diabetes 62, (2013).
- Chartoumpekis, D. V. et al. Nrf2 deletion from adipocytes, but not hepatocytes, potentiates systemic metabolic dysfunction after long-term high-fat diet-induced obesity in mice. Am J Physiol Endocrinol Metab 315, (2018).
- Cho, Y. K. et al. Lipid remodeling of adipose tissue in metabolic health and disease. Experimental and Molecular Medicine vol. 55 Preprint at (2023). [CrossRef]
- Khandekar, M. et al. Beige Adipocytes are a Distinct Type of Thermogenic Fat Cell in Mouse and Human. Cell 150, (2013).
- Petito, G. et al. Adipose Tissue Remodeling in Obesity: An Overview of the Actions of Thyroid Hormones and Their Derivatives. Pharmaceuticals vol. 16 Preprint at (2023). [CrossRef]
- Cheng, L. et al. Brown and beige adipose tissue: a novel therapeutic strategy for obesity and type 2 diabetes mellitus. Adipocyte vol. 10 Preprint at (2021). [CrossRef]
- Horwitz, A. & Birk, R. Adipose Tissue Hyperplasia and Hypertrophy in Common and Syndromic Obesity—The Case of BBS Obesity. Nutrients vol. 15 Preprint at (2023). [CrossRef]
- Engin, A. Adipose tissue hypoxia in obesity and its impact on preadipocytes and macrophages: Hypoxia hypothesis. in Advances in Experimental Medicine and Biology vol. 960 (2017).
- Al-Sari, N. et al. Lipidomics of human adipose tissue reveals diversity between body areas. PLoS One 15, (2020).
- Meikle, P. J. & Summers, S. A. Sphingolipids and phospholipids in insulin resistance and related metabolic disorders. Nature Reviews Endocrinology vol. 13 Preprint at (2017). [CrossRef]
- Palau-Rodriguez, M. et al. Visceral Adipose Tissue Phospholipid Signature of Insulin Sensitivity and Obesity. J Proteome Res 20, (2021).
- White, U. Adipose tissue expansion in obesity, health, and disease. Frontiers in Cell and Developmental Biology vol. 11 Preprint at (2023). [CrossRef]
- Gu, X., Wang, X., Wang, S., Shen, Y. & Lu, L. Nonlinear association between visceral adipose tissue area and remnant cholesterol in US adults: a cross-sectional study. Lipids Health Dis 23, 228 (2024).
- Cypess, A. M. Reassessing Human Adipose Tissue. New England Journal of Medicine 386, (2022).
- Zhang, Y. et al. Positional cloning of the mouse obese gene and its human homolog. Nature 372, (1994).
- Scheja, L. & Heeren, J. The endocrine function of adipose tissues in health and cardiometabolic disease. Nature Reviews Endocrinology vol. 15 Preprint at (2019). [CrossRef]
- Marseglia, L. et al. Oxidative stress in obesity: A critical component in human diseases. International Journal of Molecular Sciences vol. 16 Preprint at (2015). [CrossRef]
- Małkowska, P. Positive Effects of Physical Activity on Insulin Signaling. Curr Issues Mol Biol 46, 5467–5487 (2024).
- Zatterale, F. et al. Chronic Adipose Tissue Inflammation Linking Obesity to Insulin Resistance and Type 2 Diabetes. Frontiers in Physiology vol. 10 Preprint at (2020). [CrossRef]
- Ormazabal, V. et al. Association between insulin resistance and the development of cardiovascular disease. Cardiovascular Diabetology vol. 17 Preprint at (2018). [CrossRef]
- Karlsson, H. K. R. & Zierath, J. R. Insulin signaling and glucose transport in insulin resistant human skeletal muscle. Cell Biochemistry and Biophysics vol. 48 Preprint at (2007). [CrossRef]
- Samuel, V. T. & Shulman, G. I. The pathogenesis of insulin resistance: integrating signaling pathways and substrate flux. Journal of Clinical Investigation 126, 12– 22 (2016).
- Santoro, A., McGraw, T. E. & Kahn, B. B. Insulin action in adipocytes, adipose remodeling, and systemic effects. Cell Metabolism vol. 33 Preprint at (2021). [CrossRef]
- Patel, R., Varghese, J. F. & Yadav, U. C. S. Crosstalk between adipose tissue, macrophages, and other immune cells: Development of obesity and inflammation-induced metabolic diseases. in Obesity and Diabetes: Scientific Advances and Best Practice (2020).
- Malenica, M. & Meseldžić, N. Oxidative stress and obesity. Arhiv za Farmaciju vol. 72 Preprint at (2022). [CrossRef]
- Capece, D. et al. NF-κB: blending metabolism, immunity, and inflammation. Trends in Immunology vol. 43 Preprint at (2022). [CrossRef]
- Milhem, F., Skates, E., Wilson, M. & Komarnytsky, S. Obesity-Resistant Mice on a High-Fat Diet Display a Distinct Phenotype Linked to Enhanced Lipid Metabolism. Nutrients 16, (2024).
- Di Francesco, A. et al. NQO1 protects obese mice through improvements in glucose and lipid metabolism. NPJ Aging Mech Dis 6, (2020).
- Zhan, X., Li, J. & Zhou, T. Targeting Nrf2-Mediated Oxidative Stress Response Signaling Pathways as New Therapeutic Strategy for Pituitary Adenomas. Front Pharmacol 12, (2021).
- Gutiérrez-Cuevas, J. et al. The Role of NRF2 in Obesity- Associated Cardiovascular Risk Factors. Antioxidants vol.
- 11 Preprint at (2022). [CrossRef]
- Liu, Z. et al. Deletion of Nrf2 leads to hepatic insulin resistance via the activation of NF-κB in mice fed a high- fat diet. Mol Med Rep 14, (2016).
- Chanas, S. A. et al. Loss of the Nrf2 transcription factor causes a marked reduction in constitutive and inducible expression of the glutathione S-transferase Gsta1, Gsta2, Gstm1, Gstm2, Gstm3 and Gstm4 genes in the livers of male and female mice. Biochemical Journal 365, (2002).
- Schneider, K. et al. Increased energy expenditure, ucp1 expression, and resistance to diet-induced obesity in mice lacking nuclear factor-erythroid-2-related transcription factor-2 (nrf2). Journal of Biological Chemistry 291, (2016).
- Sugimoto, H. et al. Deletion of nuclear factor-E2-related factor-2 leads to rapid onset and progression of nutritional steatohepatitis in mice. Am J Physiol Gastrointest Liver Physiol 298, (2010).
- Wakabayashi, N., Yagishita, Y., Joshi, T. & Kensler, T. W. Forced Hepatic Expression of NRF2 or NQO1 Impedes Hepatocyte Lipid Accumulation in a Lipodystrophy Mouse Model. Int J Mol Sci 24, (2023).
- Shin, S. M., Yang, J. H. & Ki, S. H. Role of the Nrf2-are pathway in liver diseases. Oxidative Medicine and Cellular Longevity Preprint at (2013). [CrossRef]
- Li, L. et al. Hepatocyte-specific Nrf2 deficiency mitigates high-fat diet-induced hepatic steatosis: Involvement of reduced PPARγ expression. Redox Biol 30, (2020).
- Yi, M. et al. Nrf2 Pathway and Oxidative Stress as a Common Target for Treatment of Diabetes and Its Comorbidities. International Journal of Molecular Sciences vol. 25 Preprint at (2024). [CrossRef]
- Martínez, L., Ros, G. & Nieto, G. Hydroxytyrosol: Health Benefits and Use as Functional Ingredient in Meat. Medicines 5, (2018).
- Micheli, L. et al. Role of Hydroxytyrosol and Oleuropein in the Prevention of Aging and Related Disorders: Focus on Neurodegeneration, Skeletal Muscle Dysfunction and Gut Microbiota. Nutrients vol. 15 Preprint at (2023). [CrossRef]
- Sabatini, N. Recent Patents in Olive Oil Industry: New Technologies for the Recovery of Phenols Compounds from Olive Oil, Olive Oil Industrial by-Products and Waste Waters. Recent Patents on Food, Nutrition & Agriculturee 2, (2012).
- Franconi, F., Campesi, I. & Romani, A. Is Extra Virgin Olive Oil an Ally for Women’s and Men’s Cardiovascular Health? Cardiovascular Therapeutics vol. 2020 Preprint at (2020). [CrossRef]
- Echeverría, F., Ortiz, M., Valenzuela, R. & Videla, L. A. Hydroxytyrosol and cytoprotection: A projection for clinical interventions. International Journal of Molecular Sciences vol. 18 Preprint at (2017). [CrossRef]
- Rodríguez-García, C., Sánchez-Quesada, C., Algarra, I. & Gaforio, J. J. Differential Immunometabolic Effects of High-Fat Diets Containing Coconut, Sunflower, and Extra Virgin Olive Oils in Female Mice. Mol Nutr Food Res 66, (2022).
- Cao, K. et al. Hydroxytyrosol prevents diet-induced metabolic syndrome and attenuates mitochondrial.
- abnormalities in obese mice. Free Radic Biol Med 67, (2014).
- Liu, Z., Wang, N., Ma2, Y. & Wen, D. Hydroxytyrosol improves obesity and insulin resistance by modulating gut microbiota in high-fat diet-induced obese mice. Front Microbiol 10, (2019).
- Del Rio, D. et al. Dietary (poly)phenolics in human health: Structures, bioavailability, and evidence of protective effects against chronic diseases. Antioxidants and Redox Signaling vol. 18 Preprint at (2013). [CrossRef]
- Benzie, I. F. F. & Wachtel-Galor, S. Herbal Medicine: Biomolecular and Clinical Aspects: Second Edition. Herbal Medicine: Biomolecular and Clinical Aspects: Second Edition (2011).
- Peluso, I. & Serafini, M. Antioxidants from black and green tea: from dietary modulation of oxidative stress to pharmacological mechanisms. British Journal of Pharmacology vol. 174 Preprint at (2017). [CrossRef]
- Na, H. K. & Surh, Y. J. Modulation of Nrf2-mediated antioxidant and detoxifying enzyme induction by the green tea polyphenol EGCG. Food and Chemical Toxicology 46, (2008).
- Oz, H. S. Chronic inflammatory diseases and green tea polyphenols. Nutrients vol. 9 Preprint at (2017). [CrossRef]
- Wang, D. et al. Green tea polyphenols mitigate the plant lectins-induced liver inflammation and immunological reaction in C57BL/6 mice via NLRP3 and Nrf2 signaling pathways. Food and Chemical Toxicology 144, (2020).
- Surma, S., Sahebkar, A. & Banach, M. Coffee or tea: Anti- inflammatory properties in the context of atherosclerotic cardiovascular disease prevention. Pharmacological Research vol. 187 Preprint at (2023). [CrossRef]
- James, A., Wang, K. & Wang, Y. Therapeutic Activity of Green Tea Epigallocatechin-3-Gallate on Metabolic Diseases and Non-Alcoholic Fatty Liver Diseases: The Current Updates. Nutrients vol. 15 Preprint at (2023). [CrossRef]
- Liu, C. Y. et al. Effects of green tea extract on insulin resistance and glucagon-like peptide 1 in patients with type 2 diabetes and lipid abnormalities: A randomized, double-blinded, and placebo-controlled trial. PLoS One 9, (2014).
- Yu, J., Song, P., Perry, R., Penfold, C. & Cooper, A. R. The effectiveness of green tea or green tea extract on insulin resistance and glycemic control in type 2 diabetes mellitus: A meta-analysis. Diabetes and Metabolism Journal vol. 41 Preprint at (2017). [CrossRef]
- De la Fuente-Muñoz, M. et al. Supplementation with a New Standardized Extract of Green and Black Tea Exerts Antiadipogenic Effects and Prevents Insulin Resistance in Mice with Metabolic Syndrome. Int J Mol Sci 24, (2023).
- de Oliveira Assis, F. S., Vasconcellos, G. L., Lopes, D. J. P., de Macedo, L. R. & Silva, M. Effect of Green Tea Supplementation on Inflammatory Markers among Patients with Metabolic Syndrome and Related Disorders: A Systematic Review and Meta-Analysis. Prev Nutr Food Sci 29, 106–117 (2024).
- Martínez-Zamora, L., Peñalver, R., Ros, G. & Nieto, G. Olive tree derivatives and hydroxytyrosol: Their potential effects on human health and its use as functional ingredient in meat. Foods vol. 10 Preprint at (2021). [CrossRef]
- Wang, D., Wang, T., Li, Z., Guo, Y. & Granato, D. Green Tea Polyphenols Upregulate the Nrf2 Signaling Pathway and Suppress Oxidative Stress and Inflammation Markers in D-Galactose-Induced Liver Aging in Mice. Front Nutr 9, (2022).
Figure 1.
Nrf2 structural domains. Nrf2 contains seven domains known as Neh1–Neh7. The N-terminal domain of Neh2 contains two motifs, DLG and ETGE, which are responsible for binding KEAP1. The Neh3, Neh4, and Neh5 domains are important for the transactivation activity of Nrf2. The Neh7 domain binds RXRα and inhibits ARE gene activity. Neh6 is a serine-rich domain required for β-TrCP binding. The C-terminal domain of Neh1 is a leucine zipper motif responsible for DNA binding and dimerization with Maf (Modified from [12]).
Figure 1.
Nrf2 structural domains. Nrf2 contains seven domains known as Neh1–Neh7. The N-terminal domain of Neh2 contains two motifs, DLG and ETGE, which are responsible for binding KEAP1. The Neh3, Neh4, and Neh5 domains are important for the transactivation activity of Nrf2. The Neh7 domain binds RXRα and inhibits ARE gene activity. Neh6 is a serine-rich domain required for β-TrCP binding. The C-terminal domain of Neh1 is a leucine zipper motif responsible for DNA binding and dimerization with Maf (Modified from [12]).
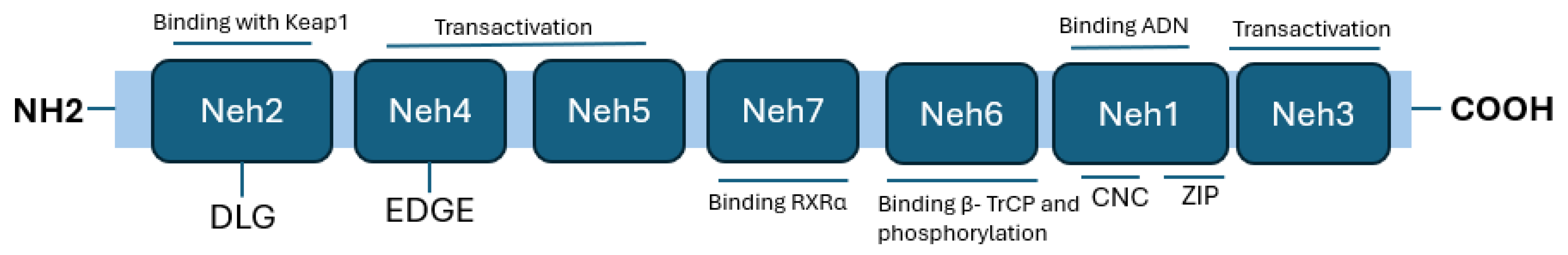
Figure 2.
Nrf2 signaling pathway. Under homeostatic conditions, Nrf2 binds to its main inhibitor, Keap1, which has two specific coupling sites: a cyclic E3-ubiquitin ligase and the Cullin-3/Rbx1 complex. Therefore, Nrf2 enters the ubiquitin proteasome to be degraded. In the process of oxidative stress, oxidative factors can act and alter the chemical conformation of the Keap1 cysteine, thus inhibiting ubiquitination and leading to excessive activation of Nrf2. Nrf2 can be phosphorylated by several enzymes, such as protein kinase C (PKC), which phosphorylates it at the Ser40 position, disrupting its interaction with Keap1. ROS can also increase Nrf2 translocation to the nucleus through promotion of the oxidation of cysteine residues in Keap1, allowing the release of Nrf2, which initiates the antioxidant transcription process (modified from [17]).
Figure 2.
Nrf2 signaling pathway. Under homeostatic conditions, Nrf2 binds to its main inhibitor, Keap1, which has two specific coupling sites: a cyclic E3-ubiquitin ligase and the Cullin-3/Rbx1 complex. Therefore, Nrf2 enters the ubiquitin proteasome to be degraded. In the process of oxidative stress, oxidative factors can act and alter the chemical conformation of the Keap1 cysteine, thus inhibiting ubiquitination and leading to excessive activation of Nrf2. Nrf2 can be phosphorylated by several enzymes, such as protein kinase C (PKC), which phosphorylates it at the Ser40 position, disrupting its interaction with Keap1. ROS can also increase Nrf2 translocation to the nucleus through promotion of the oxidation of cysteine residues in Keap1, allowing the release of Nrf2, which initiates the antioxidant transcription process (modified from [17]).
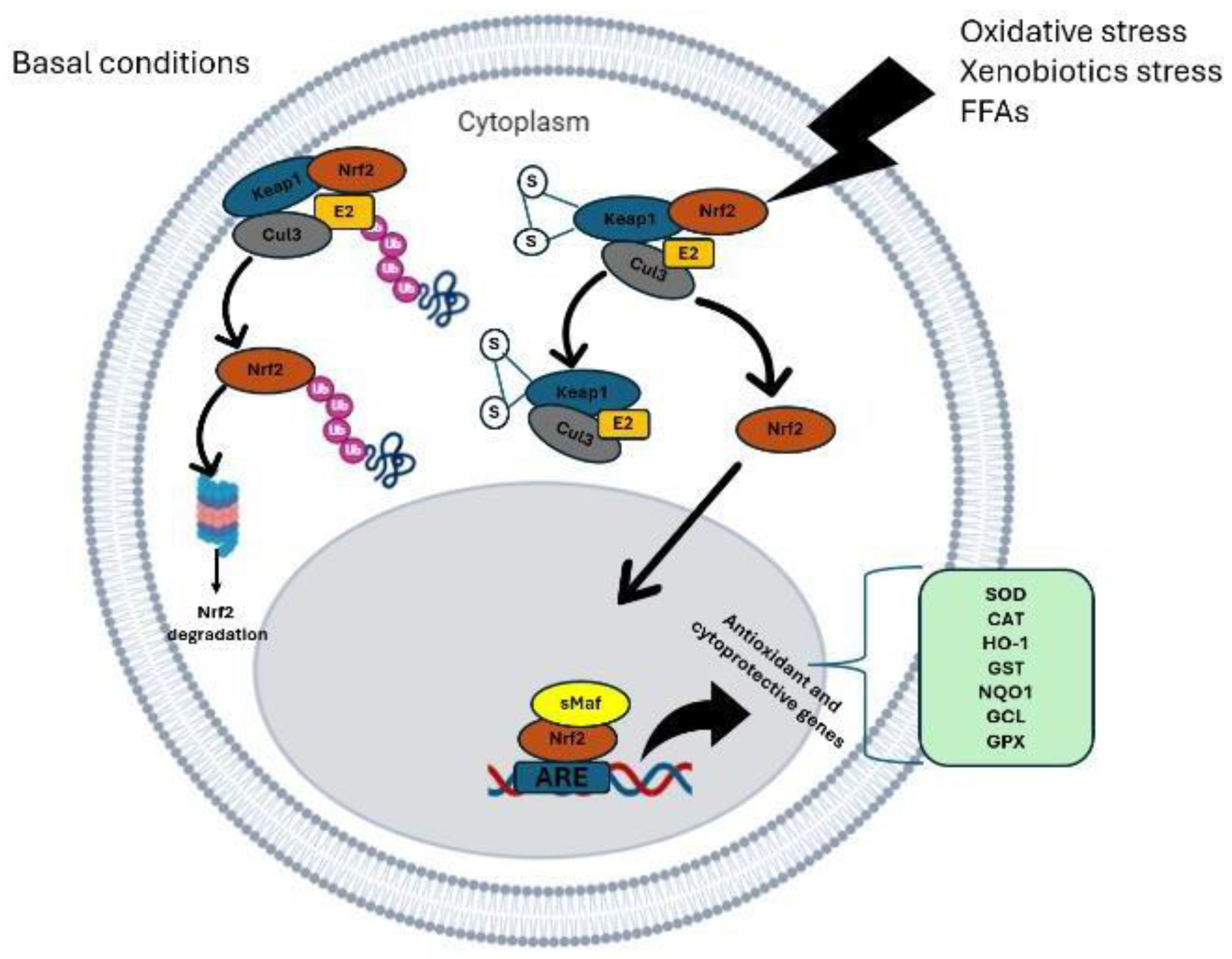
Figure 3.
Genes under the transcriptional control of Nrf2. Modified from [18].
Figure 3.
Genes under the transcriptional control of Nrf2. Modified from [18].
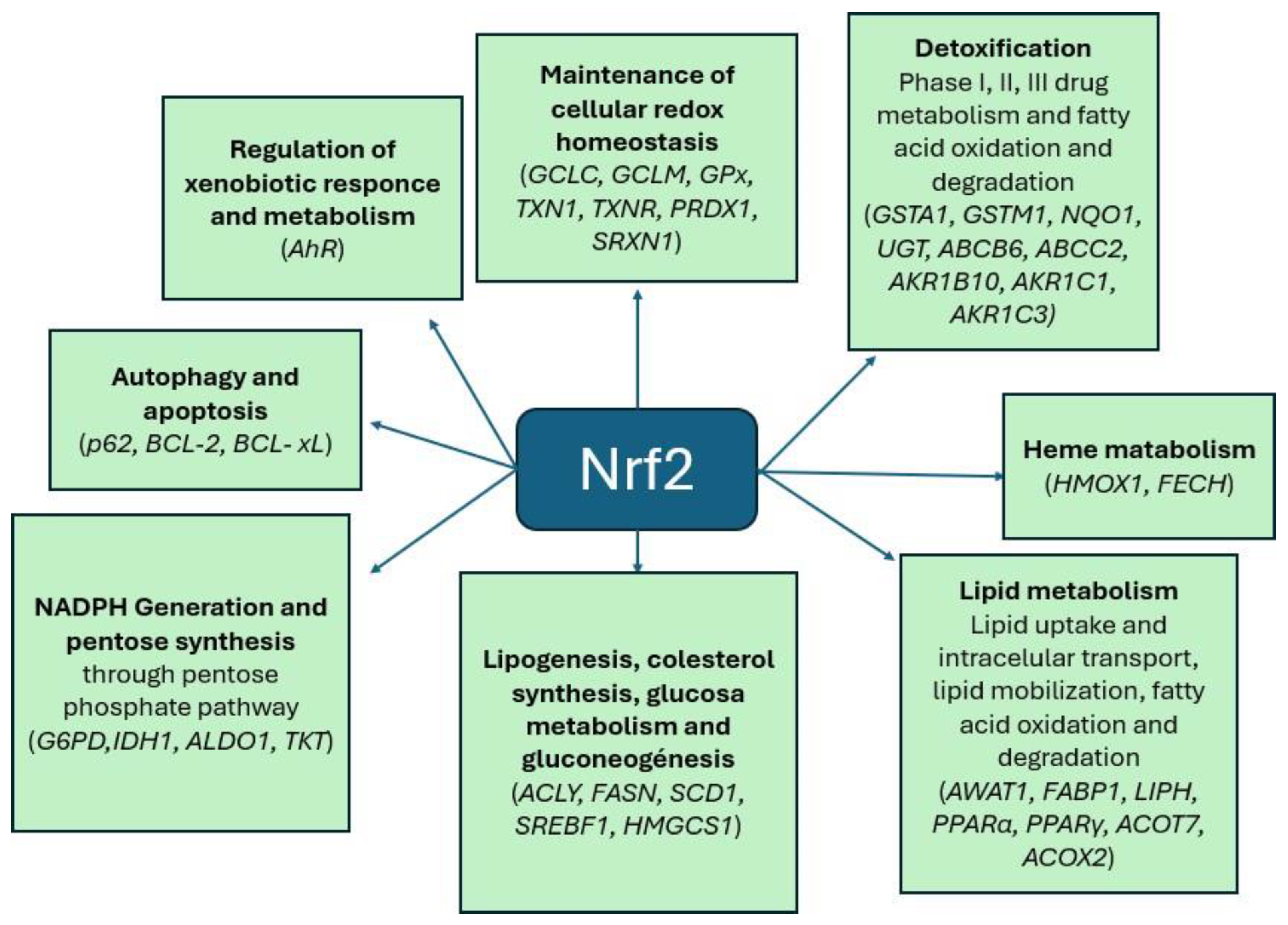
Figure 4.
Adipogenesis process. Mesenchymal stem cells give rise to adipoblasts that further differentiate into preadipocytes under the influence of multiple transcription factors, such as preadipocyte factor-1 (Pref-1), sterol regulatory element-binding protein 1 (SREBP-1), and peroxisome proliferator-activated receptor gamma (PPARγ), and changes in the extracellular matrix (ECM) and cytoskeleton. Preadipocytes are then further differentiated into immature adipocytes and mature adipocytes under the influence of CCAAT/enhancer-binding protein alpha (C/EBPα), adipocyte protein 2 (aP2), leptin, lipoprotein lipase (LPL), leukocyte differentiation antigen (CD36), and glucose transporter number 4 (GLUT4). The two arrows indicate the expression of the specific factors throughout the transition period (modified from [35]).
Figure 4.
Adipogenesis process. Mesenchymal stem cells give rise to adipoblasts that further differentiate into preadipocytes under the influence of multiple transcription factors, such as preadipocyte factor-1 (Pref-1), sterol regulatory element-binding protein 1 (SREBP-1), and peroxisome proliferator-activated receptor gamma (PPARγ), and changes in the extracellular matrix (ECM) and cytoskeleton. Preadipocytes are then further differentiated into immature adipocytes and mature adipocytes under the influence of CCAAT/enhancer-binding protein alpha (C/EBPα), adipocyte protein 2 (aP2), leptin, lipoprotein lipase (LPL), leukocyte differentiation antigen (CD36), and glucose transporter number 4 (GLUT4). The two arrows indicate the expression of the specific factors throughout the transition period (modified from [35]).
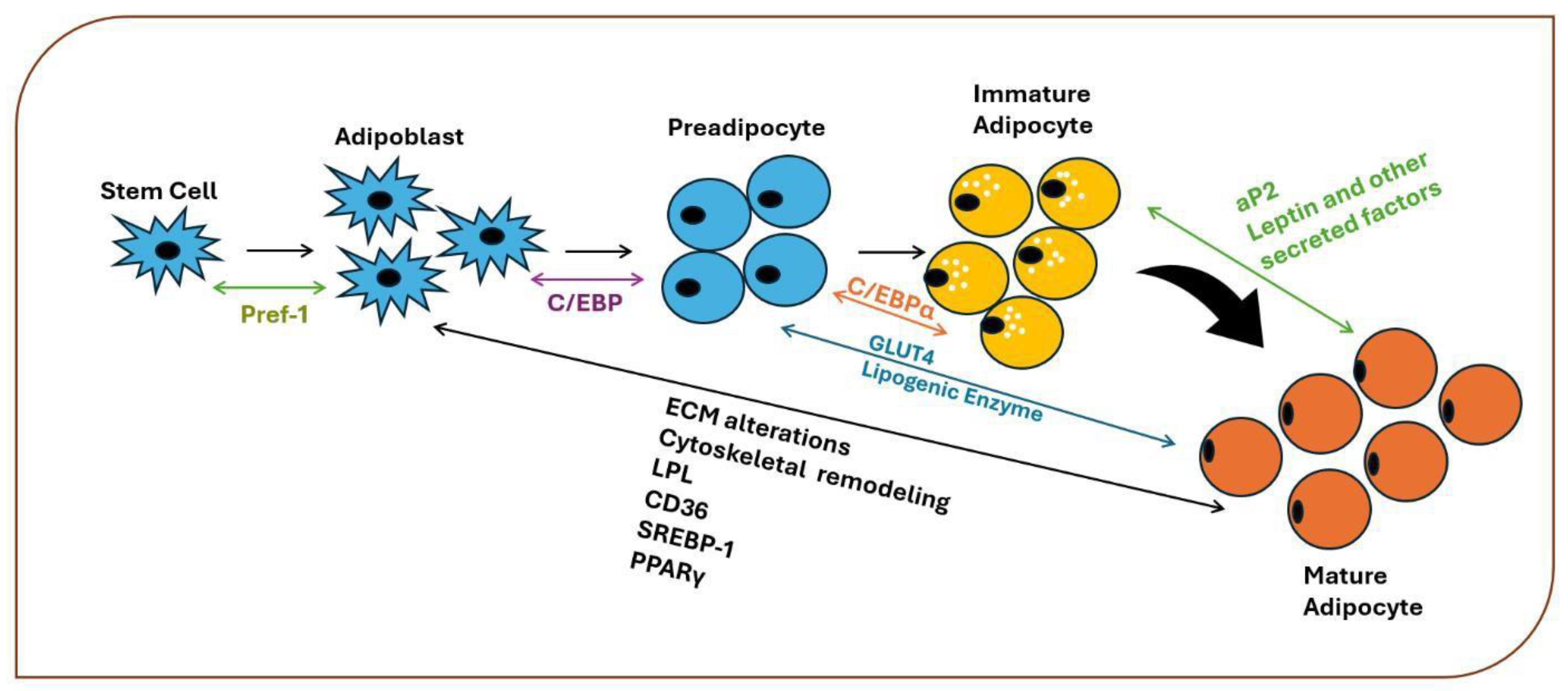
Figure 5.
The role of oxidative stress in adipocyte differentiation (modified from [17]). Increased levels of glucose and free fatty acids, along with the depletion of antioxidants, contribute to increased oxidative stress by increasing the production of reactive oxygen species (ROS). In addition, increased oxidative stress arises through endoplasmic reticulum stress and mitochondrial stress. Increased ROS activate AMPK and induce disulfide bond formation between cysteine residues and C/EBPβ dimerization, leading to further increases in DNA-binding activity. C/EBP-β activates PPAR-γ expression during the early stages of adipogenesis.
Figure 5.
The role of oxidative stress in adipocyte differentiation (modified from [17]). Increased levels of glucose and free fatty acids, along with the depletion of antioxidants, contribute to increased oxidative stress by increasing the production of reactive oxygen species (ROS). In addition, increased oxidative stress arises through endoplasmic reticulum stress and mitochondrial stress. Increased ROS activate AMPK and induce disulfide bond formation between cysteine residues and C/EBPβ dimerization, leading to further increases in DNA-binding activity. C/EBP-β activates PPAR-γ expression during the early stages of adipogenesis.
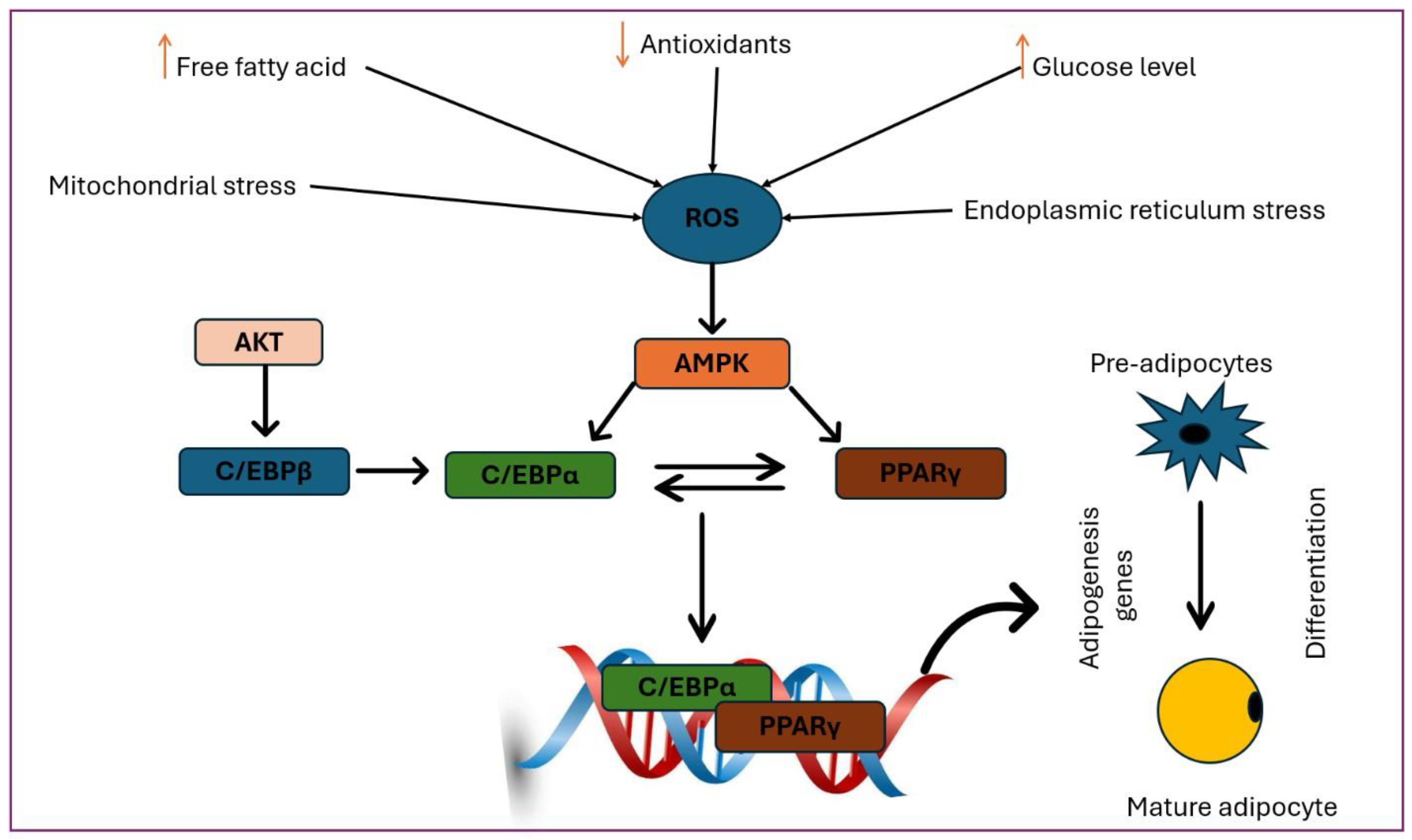
Figure 6.
Role of Nrf2 in adipogenesis. Adipocyte progenitors, which are characterized by platelet-derived growth factor receptor-α (PDGFRα) and PDGFRβ expression, are restricted to the adipocyte lineage without obvious morphological changes and then form preadipocytes. This cellular involvement is subsequently followed by terminal differentiation, during which selected preadipocytes undergo growth arrest, accumulate lipid droplets, and form mature, insulin-sensitive, functional adipocytes. The receptor γ is activated by the peroxisome proliferator (PPARγ) and the enhancer/CCAAT-binding α protein (C/EBPα), which together oversee the entire process of terminal differentiation. Intricate networks of the above transcription factors coordinate the expression of hundreds of proteins responsible for establishing the mature adipocyte phenotype. Modified from [17].
Figure 6.
Role of Nrf2 in adipogenesis. Adipocyte progenitors, which are characterized by platelet-derived growth factor receptor-α (PDGFRα) and PDGFRβ expression, are restricted to the adipocyte lineage without obvious morphological changes and then form preadipocytes. This cellular involvement is subsequently followed by terminal differentiation, during which selected preadipocytes undergo growth arrest, accumulate lipid droplets, and form mature, insulin-sensitive, functional adipocytes. The receptor γ is activated by the peroxisome proliferator (PPARγ) and the enhancer/CCAAT-binding α protein (C/EBPα), which together oversee the entire process of terminal differentiation. Intricate networks of the above transcription factors coordinate the expression of hundreds of proteins responsible for establishing the mature adipocyte phenotype. Modified from [17].
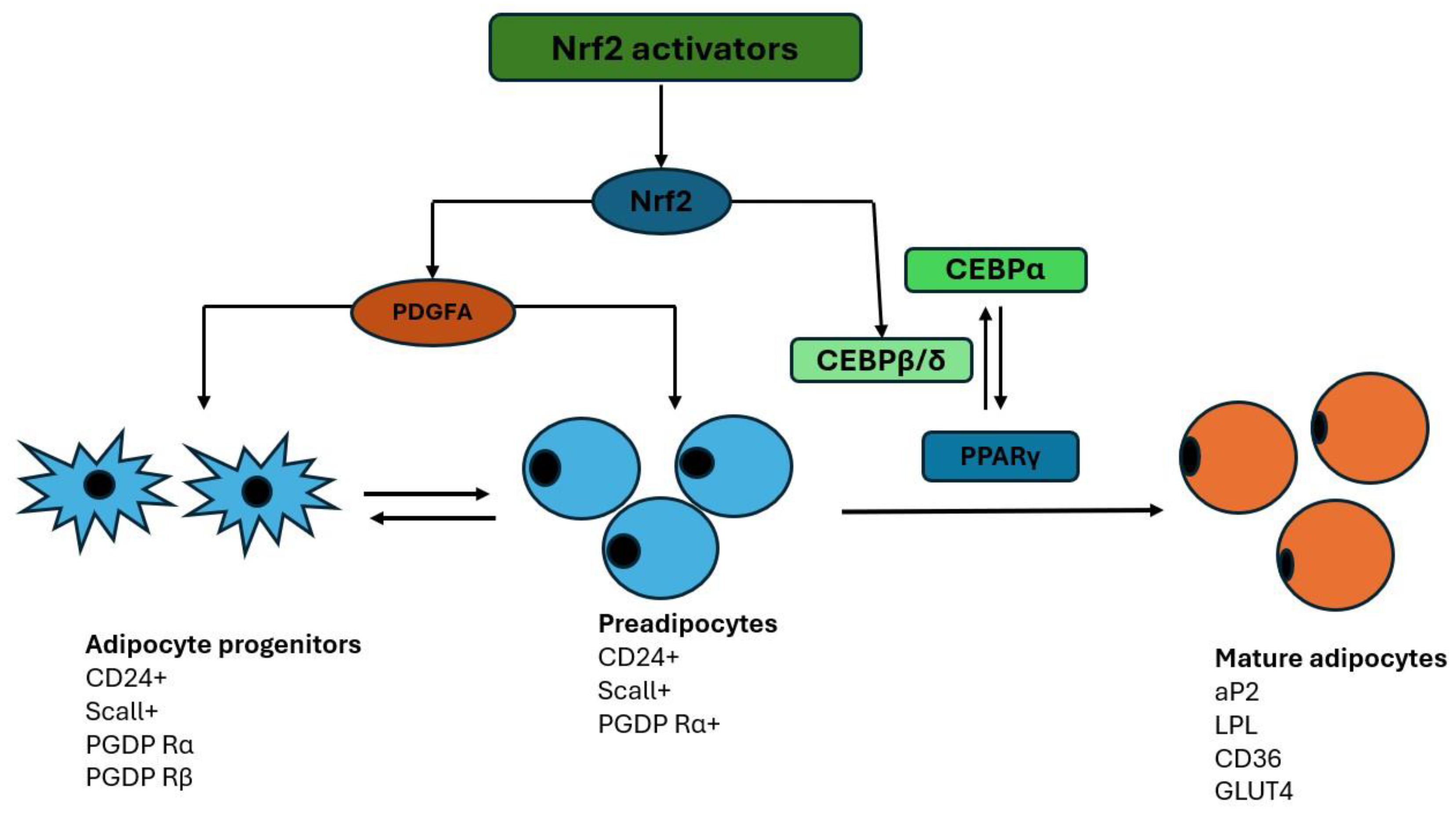

Table 1.
Some proven Nrf2 agonists.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



