Submitted:
05 May 2025
Posted:
08 May 2025
You are already at the latest version
Abstract
This study addresses the issue of whether orally administered zinc (gluconate) (26 mg BID) can induce remodeling of gastrointestinal barrier function and reduce passive leak across the human intestinal mucosal barrier in situ. It represents the first patient-based study to examine the effect of zinc on GI epithelial tight junctions and GI barrier leak in otherwise healthy test subjects. Using independent test subject groups for each endpoint, three separate molecular analyses indicated that zinc can achieve a positive outcome: 1) RNA-seq analyses of intestinal biopsies showed patterns of gene transcription changes dealing with not only transcripts of junctional proteins, but also transcripts mitigating the proinflammatory state as well as dedifferentiation (both modulators of tight junction permeability); 2) Western immunoblot analyses of intestinal tissue indicated tight junctional protein expression was being modified by administered zinc, most notably, Claudin-2 and Tricellulin; 3) zinc treatment induced a reduction of serum levels of a functional marker of passive intestinal leak, namely the GI microbiome metabolite, D-Lactate. The data together suggest that orally administered zinc can induce remodeling of the intestinal epithelial barrier resulting in reduction of GI barrier leak. The overall safety and economy of supplement levels of zinc suggest that this micronutrient could be efficacious as an adjuvant therapy to reduce the condition known as leaky gut, and possibly therefore protective regarding diseases postulated to involve leaky gut.
Keywords:
Tight Junction
; Intestine
; Claudin
; Micronutrient
; Leaky Gut
; Zinc
; D-Lactate
Introduction
The importance of epithelial barrier function to human physiology cannot be overstated. Our anatomy shows us to largely be a parallel array of sacs and tubes, each defining an organ-specific luminal compartment, separated from the vascular compartment by an epithelial cell layer interface, very often just one cell layer thick. The ability of that interface to serve as a functional barrier separating wildly different luminal versus abluminal milieus is critical to all higher animal life. The compromise of that barrier function is a lynchpin throughout a very wide array of disease etiology and is at the core of a great deal of disease-related morbidity and mortality. Although not the sole arbiter of epithelial barrier function, the apically situated, circumferential tight junctional (TJ) complex (Zonula Occludens), plays a cardinal role as the dynamic, highly regulated molecular sieve that determines size and charge selectivity along the paracellular transepithelial pathway. Dysregulation or outright failure of this key barrier element is a prominent feature in a very wide array of human disease [1,2].
The gastrointestinal epithelial barrier holds two important hallmarks of its own among our epithelial tissue barriers. Even surpassing the epidermis, it is first, the largest epithelial barrier in terms of overall surface area. Secondly, it separates the vasculature from the greatest prokaryotic population in our body, the GI microbiome. Given that the vasculature is home to our immune system, this is a barrier separation that is highly pivotal and whose compromise would a priori result in significant inflammation. Disease-related compromise of GI barrier function is obvious in conditions such as Crohn’s Disease and Ulcerative Colitis [3,4]. However, overt GI barrier compromise often begins very subtly with a focused dysregulation of the TJ complex itself. This occurs very early in diseases as disparate as colon neoplasia [5] and certain intestinal microbial infections [6].
The condition or syndrome known as “Leaky Gut” (LG) is not a recognized disease or even a diagnosis. A cursory examination of it in the literature will be sure to include words and phrases such as “hypothetical,” “may,” “difficult,” and “unrecognized.” Although controversy surrounds it, curiously no one seems to dispute those conditions of “increased intestinal permeability” are quite real [7,8]. This is certainly not surprising given that pathophysiologically increased permeability or “leak” across renal tubular barriers [9], oral epithelial barriers [10], airway epithelial barriers [11] and retinal epithelial barriers [12] among many, are fully accepted. No one disputes that GI inflammation can induce increased GI permeability [13]. The controversy may lie in whether GI leak is a necessary and sufficient cause for GI inflammation [14]. And from there one can eventually progress on to whether GI leak can be playing a role in seemingly highly unrelated (and physically remote) conditions such as cognitive disease [15].
A consensus appears to be building that at the very least, a hard look into the origins of GI barrier leak, its ramifications, and what one might possibly do to alleviate it, is fully warranted. This current investigation deals with the third endeavor - are there therapeutic options by which such leak can be reduced? Any epithelial barrier relies on its TJ complexes as the final gatekeeper of transepithelial paracellular movement, but the GI barrier also involves a wide array of components ranging from mucus layers to the rate (and manner) of epithelial cell turnover within the barrier by necrotic/apoptotic/autophagic means [5,16]. When one considers the complexity of the TJ (26+ claudins, occludin, tricellulin, 30+ TJ-related proteins) AND the above-mentioned (non TJ) contributors to the barrier as well, the demands placed on a de-novo drug to achieve barrier improvement are very high. Such a drug must decrease TJ permeability but also not adversely affect the other barrier contributors as well. A structurally highly diverse set of micronutrients has however succeeded in this endeavor [17,18,19]. This may not be entirely surprising given that animal life – and its need for barrier function and protection – has coevolved with these micronutrients in the animal diet. Natural selection may well have vetted these specific micronutrients to succeed here. The ability of Quercetin, Calcitriol and Retinoic Acid to improve barrier function in the airway epithelium [20,21], and of zinc (Zn) to improve and protect the GI barrier [22] have been well documented examples in many cell culture and animal model studies. –
The success of Zn in specifically GI barrier function studies has however not been well validated in specifically clinical, patient-based studies. In fact, there is only one patient-based study reported in the published literature that shows orally administered Zn decreasing barrier leak in humans. Sturniolo et al (2001) showed that administration of a similar oral dose (25 mg Zn, 3 times daily) to that used in our current study, reduced the GI barrier leak that accompanies Crohn’s Disease [23]. However, this pathophysiological situation obviated addressing whether Zn could improve normal GI barrier function and whether such improvement could trace itself to induced remodeling of the TJ complex. Answering those questions becomes highly important in determining if supplemental Zn can serve a prophylactic, preventative clinical role in a variety of diseases that could trace themselves to aberrantly increased GI leak.
Methods
Patient Demographics and Enrollment
mRNA and Western Immunoblot Studies
All enrolled subjects provided written informed consent, and the study was approved through the Lankenau Medical Center Institutional Review Board (E-18-3817). Patients presenting for routine upper endoscopy surveillance (Barrett’s esophagus, GERD, gastritis but no history of or active small bowel disease) were contacted for possible recruitment 6-8 weeks prior to their scheduled procedure. Prior medical history exclusions were Inflammatory Bowel Disease, Irritable Bowel Syndrome, prior GI surgery, history of gastric or duodenal ulceration, history of H. pylori infection, Celiac Disease, or insulin-dependent diabetes. Patients were also excluded if they were taking a proton pump inhibitor (PPI) as these drugs can inhibit Zn uptake by the small bowel [24]. Patients on an aspirin regimen or taking any NSAID for analgesic purposes were excluded since these can aberrantly increase GI permeability. After a medication history to exclude those taking medications that would be inhibited (or their uptake affected) by Zn (amiloride-class diuretics, fluoroquinolone antibiotics, anticoagulants [except for aspirin], hormone-replacement therapy, and cholestyramine), informed written consent was obtained. Patients 18-80 years of age were recruited without regard to gender or ethnicity.
Zinc Leaky Gut Study
As above, healthy test subjects 18-80 years of age with no history of (or active) gastrointestinal disease were enrolled and recruited without regard to gender or ethnicity. The same medical and medication exclusions used above were observed in this functional study. All enrolled subjects provided written informed consent, and the study was separately approved through the Lankenau Medical Center Institutional Review Board (E-21-5153). Test subjects were not allowed to begin the study if a current GI infection was suspected and were withdrawn from the study if a suspected GI infection occurred during the study.
Zinc Administration
- 1)
- mRNA and Western Immunoblot Study
Test subjects were instructed to take two 13.3 mg (Zn) Zn gluconate lozenges twice a day (53 mg Zn daily dose) for 14 days prior to their procedure, the final dose being taken the evening prior to the procedure. Patients were instructed to avoid foods containing phytates (seeds, nuts, etc.) as well as citrus 2 hours before and 1 hour after taking the Zn gluconate to avoid inhibition of Zn uptake. A matched set of patients were administered a chemically identical (sodium gluconate, molar equivalent) placebo. Patients were blinded as to which medication they were taking until after their procedure. No Zn-related adverse events were observed.
Normal (dietary) Zn intake is considered in the range of 5–10 mg/adult/day, and toxic limits are generally viewed to be above 150 mg/adult/day. Lower doses (< 100 mg/adult/day) have generally been considered safe and without effect on systemic copper levels [25,26]. One hundred milligrams of Zn/adult/day had no significant effect on plasma copper levels over 3 months in an elderly population [27]. Zn levels more than 150 mg/adult/day were required to cause changes in copper status, immune function, and HDL levels [28]. The transit time/turnover time of epithelial mucosa for normal stratified squamous esophagus is 7.5 days, whereas intestine is only 3 days [29]. Our 14-day treatment time allows for several refoliations of duodenal tissue in the presence of Zn. Therefore, we are allowing for not only Zn effects on existing, differentiated epithelia, but also Zn effects on undifferentiated epithelia (and stem cells) in the crypt regions of the duodenum.
- 2)
- Zinc Leaky Gut Study
Test subjects were instructed to take one 30 mg (Zn) Zn gluconate tablet twice a day (daily dose of 60 mg) for 7 days prior to their procedure, the final dose being taken the evening prior to the procedure. Patients were instructed to avoid foods containing phytates (seeds, nuts, etc.) as well as citrus 2 hrs before and 1 hr after taking the Zn gluconate to avoid inhibition of Zn uptake. A matched set of patients were administered a chemically identical (sodium gluconate) placebo. Patients were randomly assigned to the Zn or placebo subgroups by the sealed envelope method. Patients were blinded as to which medication they were taking until after they completed the entire protocol. No Zn-related adverse events occurred during the study.
Duodenal Biopsy Collection and Processing
During the upper endoscopy (after 14 days of Zn or placebo medication), four standard (1 mm3) biopsies were taken from the bulb portion of the duodenum and pooled for Western immunoblot analyses. Two biopsies were taken and pooled for mRNA analyses. Biopsies were flash frozen on dry ice in the endoscopy procedures room and stored at -196oC (liquid nitrogen) until processed for molecular analyses.
RNA-seq Data Analysis
For RNA-seq analyses, RNA was extracted by homogenizing the tissue in TRI Reagent (Sigma, USA) followed by RNA extraction using Direct-zol RNA Mini kit (Zymo Research, USA) with an in-column DNAse treatment. Total RNA was used to generate indexed libraries using QuantSeq 3’ mRNA-Seq FWD library preparation kit (Lexogen GmbH, Austria) following manufacturer’s instructions. Final libraries were subjected to quality check on the Tape Station (Agilent, USA) and quantified using KAPA Library Quant Kit (for Illumina) (KAPA Biosystems, South Africa). The libraries were pooled in equimolar concentration and sequenced on Nextseq 500 (Illumina, USA) using high output sequencing kit v2 generating 75 bp single reads. Data was aligned using STAR [30] algorithm against hg19 human genome version. RSEM v1.2.12 software [31] was used to estimate read counts and FPKM values using gene information from Ensemble transcriptome version GRCh37.p12. Raw counts were used to estimate significance of differential expression difference between two experimental groups using DESeq2 [32]. Gene Set Enrichment Analysis was done using GSEA [33] estimated by the DESeq2 gene significance for gene ranking. Results that pass the P value (0.05) threshold were considered significant.
Western Immunoblot Analyses
Biopsy tissue was thawed and resuspended in 4oC lysis buffer containing sodium dodecyl sulfate with protease and phosphatase inhibitors, and then homogenized by hand using glass mortars/pestles (Wheaton, USA), thereby generating a total cell lysate. Suspensions were then sonicated, extracted for 75-90 mins at 4oC on a rotator, and ultracentrifuged. Cellular proteins were separated on Novex tris-glycine minigels. Primary antibodies to the tight junction proteins, tricellulin, and claudins -3,-4,-5 and -7 were products of Thermo Fisher (Invitrogen) (USA). Antisera to claudin-2 was a product of Abcam (USA). Antisera to protein kinase C-α was a product of Santa Cruz Biotechnology (USA). Secondary antibodies were purchased from Southern Biotech (USA). Quantification of Western blot results was performed by densitometry of specific protein bands using a BioRad Chemidoc MP Imaging System. Densitometry conducted on Memcode stains of total protein served as loading controls for all Western blot data shown [34].
Blood Sampling
Within three days before beginning their Zn regimen, the test subject contributed two 5cc whole blood samples. This was repeated at the end of their 7-day Zn regimen, specifically the morning after their final Zn dose, thereby generating “pre-Zn” and “post-Zn” samples from the same individual. Blood samples were drawn by a phlebotomist in the Lankenau Medical Center outpatient testing lab. The samples were drawn in standard red top (serum) tubes. One tube was then sent for measurement of Hemoglobin A1C and CRP levels (Quest Diagnostics). The other was transported by a member of the research team to the research lab to centrifuge out blood cells and harvest serum for storage at -20oC until testing for D-Lactate.
D-Lactate Analyses
D-Lactate was assayed in serum samples by a fluorometric assay kit (EnzyFluoTM [BioAssay Systems, Inc.]). Fluorescence was determined by a Cytation3 imaging reader (BioTek, Inc.) against a range of D-Lactate standards. Serum samples were first purified by passing through a 10k molecular weight spin column (PierceTM [Thermo Fisher, Inc.), then the filtrate was diluted 1:2 with glass distilled water, before being assayed. An L-Lactate standard (1mM) was used to confirm specificity of the assay for the D-stereoisomer.
Results
Demographics and Zinc Administration
There were three separate patient-based studies conducted within this overall project. Each involved treating gastroenterology patients presenting for esophageal or gastric screening endoscopy or healthy test subjects with orally administered Zn for 7-14 days followed by collection of either duodenal biopsy tissue samples or blood samples. These studies had as their endpoints: 1) RNA sequencing (microarrays); 2) protein Western immunoblots; 3) serum D-Lactate analyses. Each used a separate, unique set of patients/test subjects. In the first group, patients with a history of Barrett’s Esophagus or GERD (but no intestinal disease or abnormality) and scheduled for surveillance endoscopy, were placed on a Zn gluconate (or sodium gluconate [placebo]) regimen for 14 days prior to their procedure (as described in Materials and Methods). Duodenal mucosal biopsies were then obtained from the bulb portion of the duodenum, flash frozen in the Endoscopy Procedures Room then later thawed, RNA extracted and RNAseq analyses performed. The demographic characteristics of the patients used in this RNA-seq study are shown in Table 1. The demographics of a second, independent group of similar patients whose biopsies were analyzed by Western immunoblot analyses of specific TJ proteins are also shown. Finally, a third independent group, this time of healthy control subjects with no gastrointestinal or autoimmune disease, were also administered Zn gluconate for 7 days prior to having blood withdrawn for analyses of a biomarker of passive GI permeability/leak, namely the D (dextrorotary) form of lactate (D-Lactate).
In all cases, recruitment extended to males or females of ages 18-80, without regard to race/ethnicity and with no active or recorded clinical history of intestinal disease. In each study, care was taken in enrollment to exclude persons whose GI pathophysiology could affect the parameters under study. Therefore, persons with prior or current GI disease that could affect intestinal barrier function (e.g., Crohn’s Disease, Ulcerative Colitis, Celiac Disease as well as current or recent GI viral infection or consumption of medications that are known to affect intestinal permeability such as NSAIDS, steroids, etc) were all excluded. Alcohol use was similarly restricted. Enrollees were also carefully instructed to refrain from foods (e.g. spices) that are known to affect GI barrier function at least 2 days before duodenal biopsy sampling or blood sampling. In addition, enrollees were instructed to refrain during the period of Zn administration from foods known to interfere with Zn uptake, such as foods containing phytates (e.g., whole grains, nuts, seeds), as well as citrus. Likewise, enrollees were told to discontinue any over-the-counter Zn supplements a week before starting the study, and not to take any Zn supplements (such as Zn included with many multi vitamins) other than the study-provided Zn medication.
In the mRNA and Western immunoblot studies, patients were administered Zn gluconate in lozenge form (26.6 mg BID). In the D-Lactate study, test subjects were administered Zn gluconate in tablet form (30 mg BID). The total daily dose of approximately 60 mg was chosen because of its demonstrated efficacy in our earlier patient-based study involving Zn administration to Barrett’s Esophagus patients, and its effect on Barrett’s epithelia [35]. There were no adverse effects observed that would be attributable to the Zn administration in any of the three sub studies reported here.
Age ranges and gender distributions were similar between placebo and Zn-treated groups. In general, the study population was preponderantly Caucasian due to both the catchment area of the medical center and the frequency of Barrett’s Esophagus in the participating (endoscoped) patients.
mRNA Expression
RNA was purified from duodenal biopsies, and messenger RNA-seq experiments were performed as described in Materials and Methods. Expression levels of genes in duodenal mucosal biopsy samples from 6 patients treated with Zn gluconate versus 5 patients treated with a sodium gluconate placebo were compared. Our initial focus was whether a pharmacologically effective dose of Zn had been administered, regarding the target tissue - the duodenal epithelia. We observed significant upregulation of the metallothionein genes 1A, 1B, 1G and 1M - with upregulated transcriptions ranging from 2.8 to 13.7-fold (Figure 1A, upregulated genes) - a good indication that the administered Zn dosage was sufficient to affect the targeted epithelial population. There was some patient-to-patient variability in the response to Zn (shown in the individual blocks of the heat map), resulting in varied statistical significance. Of general biological interest was the 2-fold increased transcription of Trefoil Factor 3 and the 1.6-fold increase for gastrin in the Zn-treatment group. Concerning an early indication of a Zn effect on barrier function of the duodenal epithelium, it was noted that the gene transcription of tetraspanin 1 and occludin were both increased by approximately 1.5-fold, with the occludin result having a P value = 0.008. Assayed by RNA-seq gene expression, data generated in this study for over 5,000 mRNA transcripts is included as an Excel file in Supplementary Material.
Many transcripts were also downregulated in the Zn-treated group. These tended to include indicators and mediators of inflammation (Tumor Necrosis Factor and Lymphotoxin beta) as well as signaling intermediates that are known to be involved in TJ leakiness (Mitogen Activated Protein Kinase, Transforming Growth Factor beta, and the NUAK family (Figure 1A, downregulated genes).
Arguably the best case for Zn having affected barrier function in this study, as evidenced at the transcriptional level, came from gene set enrichment analysis (GSEA) of genes affected by Zn treatment. We analyzed the mRNA changes using GSEA software [33] to find categories that demonstrate significant activation or inhibition patterns based on mRNA expression changes of their member genes in Figure 1B. This analytical method focuses on gene sets or groups that share common biological function or regulation, and showed significant activation in not only the expected categories (Response to Zinc Ion and Zinc Ion Homeostasis), but also indication of the Zn-treatment having induced an effect on duodenal genes associated with the TJ complex (KEGG Tight Junction) or gene ontology cellular components (Tight Junction and Apical Junction Complex), as well as the biological process category, Cell Cell Junction Assembly, among others. GSEA also revealed noteworthy patterns of inhibition of several categories (Figure 1B) such as biological processes Interleukin 1 and 2 Production, Hallmark [gene set] TNFA Signaling Via NFKB, and the Interferon Gamma Production biological process, among others. The general pattern would indicate downregulation of proinflammatory status, a state that is known to associate with epithelial barrier leak, and in agreement with results seen with specific individual genes in Figure 1A.
Western Immunoblot Analyses
Duodenal biopsy samples (from a separate cohort of upper endoscopy patients) were also analyzed at the translational level for Zn-induced changes in specific TJ proteins. Duodenal bulb biopsy tissue samples from placebo- and Zn-treated patients were homogenized in lysis buffer, followed by ultracentrifugation, PAGE and immunoblot as described in Materials and Methods. Figure 2 shows the results of densitometry analysis of Western immunoblot protein bands of 7 specific TJ proteins analyzed in biopsy samples from 12 placebo-treated patients and 11 Zn-treated patients. In these vertical point plots, each data point corresponds to a specific patient’s TJ protein band density. Statistically significant increases (P < 0.05) were observed for Claudin-2 (40% average increase) and Tricellulin (45% average increase) in the Zn-treatment group. Near significant upregulations (P < 0.15) were observed for Claudin-3 (17% average increase), Claudin-5 (44% average increase) and Claudin-7 (10% average decrease). Claudins-1 and -4 showed no significant change or trend as a result of Zn treatment. Considering the demographic variability existing within and between each treatment group (age, gender, genetics, medications, etc), along with the relatively small sample size, achieving statistical significance is very challenging in smaller pilot studies such as this.
The duodenal biopsy samples used in these immunoblot analyses were taken from a separate set of placebo- and Zn-treated patients than those used in the transcriptional analyses shown in Figure 1A and 1B. It thus becomes noteworthy that the protein data alludes to a similar outcome as the mRNA data, namely remodeled and potentially improved TJ complexes as a result of Zn treatment.
Serum D-Lactate Analyses
In a third, fully separate patient-based study with a unique complement of test subjects, the effect of a 7-day, 30 mg BID Zn treatment was tested for its effect on a functional indicator of the GI barrier (Figure 3). This sub study was undertaken to ask if the Zn-induced changes to structural elements of GI barrier function in Figure 1 and Figure 2 correlated with a change in passive GI permeability (“leak”). A third independent group of healthy control, test subjects (n = 20), in this case with no active gastrointestinal disease or history of GI surgery were treated with Zn as described above. Peripheral blood samples were taken before and at the end of the Zn treatments. A placebo group was not used or needed here because of each test subject’s ability to serve as his/her own control in “before” vs “after” measurements. Serum was collected and analyses of D-Lactate were performed as described, along with analyses of Hemoglobin A1C and High Sensitivity CRP as reference markers that would be unexpected to change with a 7-day Zn treatment. D-Lactate was chosen as a GI permeability biomarker because of its relative ease of analysis and it being a well-documented marker of passive GI permeability [36].
Initial analyses of the entire data set (n = 20) showed no discernible trend or statistical significance regarding pre- versus post-Zn treatment. However, when results for those test subjects that showed above average pre-Zn D-lactate leak (those test subjects whose pre-Zn D-lactate levels were above the group median) were observed and analyzed separately, the results shown in Figure 3A were observed. For those 9 test subjects, a clear trend toward lower post-Zn D-Lactate levels could be seen. Using a paired Wilcoxon signed rank test for comparison between pre and post, the median D-Lactate decreased significantly from 26.6 (IQR 23.2-32.4) to 21.2 (IQR 19.4-22.0) (P = 0.0008). Of those 9 test subjects, only one showed no decrease in their serum D-Lactate level (no change) and this test subject was found to be pre-diabetic (determined clinically for the first time, with an A1C level over 12). The average decrease of serum D-Lactate concentrations for all 9 test subjects was 21 % (+ 5% SEM, n = 9, P = 0.008), post-Zn. The maximally decreased D-Lactate serum level (post-Zn) was 43% and the lowest was 0% (no change). It should be noted that of the original 10 test subjects whose serum D-Lactate levels (pre-Zn) were above the median, one test subject was removed from consideration because a post-study survey revealed that they were taking an iron supplement along with the Zn being taken for this study, and that interference of Zn absorption by the iron was possible [37,38].
In addition to the serum level of the leak biomarker, D-Lactate, the serum components, Hemoglobin A1C and High Sensitivity CRP (HS CRP) were also measured in the same blood samples. These were chosen as being serum components that were viewed as being unlikely to be affected by Zn administration. As shown in Figure 3B, as anticipated, levels of Hemoglobin A1C were virtually identical pre- and post-Zn, showing no change as a result of Zn administration. The results shown here were specifically for the half of test subjects whose initial A1C levels were above the median (like the above-median group analyzed for D-Lactate in Figure 3A). Analysis of High Sensitivity CRP was however different and surprisingly similar to the D-Lactate results. Again, selecting the half of test subjects whose pre-Zn HS CRP values were above the median, one can see that the post-Zn values trended lower (Figure 3C). Using a paired Wilcoxon signed rank test for comparison between pre and post, the median HS CRP decreased significantly from 4.0 (IQR 2.2-4.8) to 2.3(IQR 1.4-3.5) (P = 0.047). Of those test subjects whose pre-Zn HS CRP results were above the median, only one test subject showed an increased HS CRP (post-Zn), that increase being +14%.
Based upon the Pearson Correlation Coefficient, there was not a meaningful association between the magnitude of the D-Lactate leak and the magnitude of the HS CRP (0.012; P = 0.965) for specific individual test subjects. Test subjects with higher D-Lactate leak did not correlate to having higher HS CRP levels. However, as stated, the above-median subgroups of both the D-Lactate group and the HS-CRP group did both individually respond to Zn treatment with significantly decreased blood values.
Discussion
This study has shown the following outcomes: 1) an orally administered daily dose of 53 mg Zn is sufficient to affect human duodenal gene expression as revealed by sharply upregulated expression of specific metallothioneins in transcriptome analyses of duodenal mucosal biopsies; 2) changes in expression of individual genes related to barrier function included upregulation of key junctional proteins such as occludin and tetraspanin, while downregulating negative modulators of TJ barrier function such as proinflammatory mediators and signaling proteins known to induce leak; 3) overall patterns of gene transcription changes – as revealed by Gene Set Enrichment Analysis (GSEA) – suggested improved TJ barrier function and a decreased proinflammatory milieu; 4) protein expression levels of at least two – and possibly four - specific TJ barrier proteins were increased as a result of Zn administration; and 5) the above indicators of TJ remodeling were accompanied by a clinical indicator of functionally improved GI barrier function, namely decreased blood level of the bacterial metabolite, D-Lactate, after Zn treatment. Collectively, the data suggest that orally administered Zn can reduce GI barrier leak in humans, which builds upon years of cell culture and animal model findings predicting this outcome.
A wide range of GI morbidities ranging from cancer and IBD to microbial infectious diseases result in increased GI barrier leak, typically involving TJ leak specifically [2]. This appears to be true of disease generally, no matter the specific epithelial tissue in question [1,39,40,41,42]. To “push the pendulum” in the other direction, that is to induce tightening of the TJ and the barrier made leaky by disease, is however a difficult accomplishment given the extreme structural complexity of the TJ, not to mention the interplay of other factors which can influence barrier function [43]. The octapeptide, Larazotide, by targeting the zonulin receptor, is one of the only, if not very few, “drugs” that have shown effectiveness at improving barrier function [44].
As opposed to classic drugs, micronutrients are often clinically disregarded by a “What can these really accomplish of actual use?” pattern of thinking. That thinking often derives however from the observed actions of these molecules at dietary or RDA levels. However, micronutrients can act very differently when they are presented at dosages 5-10X normal dietary levels, where they function very much like drugs, turning on signaling pathways that are normally quiescent at lower (dietary) micronutrient concentrations [17]. The action of Zn on Protein Kinase C isoforms is an excellent example [43]. The fact that humans “evolved” with these micronutrients in their diets – often at higher (supplemental) levels – may have allowed for the development of salutary effects such as barrier tightening, while also minimizing undesired side effects – the essence of natural selection [17]. The attention paid to Vitamin D in reducing fluid leak across airway epithelial barriers in COVID (“lung water”) may be a noteworthy recent example [45].Given the wide array of seemingly unrelated diseases – including cognitive diseases – that may trace their etiology in part to GI barrier leak [46,47,48], finding a micronutrient that could reduce this specific barrier leak could be clinically quite salutary and far reaching. Consider too that the operative concept here is to reduce – not necessarily eliminate - barrier leak. If a micronutrient such as Zn is only partially effective in reducing this leak, it still deserves our attention. Again, using COVID as an example, ask oneself if Vitamin D achieved only a 20% reduction of respiratory morbidity, would that have been worthwhile? If that 20% reduction of morbidity translated to a 20% decreased ICU census, or 20% less need for ventilators at the height of the SARS-COV-2 epidemic, what would that have been worth on the level of a mass population in the middle of a pandemic?
Zn has a very prodigious published literature concerning its ability to induce remodeling of TJs and improve barrier function in cell culture and animal models. The GI epithelial barrier figures prominently in this literature, which is not surprising given Zn’s utility as a clinical treatment in diarrhea going back very many years [49]. A numerous amount of review articles have been written on Zn and GI barrier function. Hering (2009) and Amasheh et al. (2009) discuss Zn’s barrier actions in a context of salutary micronutrient actions generally [19,50].
Zhang (2009) discuss Zn’s barrier enhancing action in the overall context of alcoholic liver disease compromising the GI barrier [22]. Skrovanek et al. (2014) discuss this ability of Zn in the more general context of a variety of GI diseases [51].
In terms of research publications, the studies conducted on human GI epithelial cell cultures have yielded the most controlled findings. Wang et al. (2013) and Valenzano et al. (2015) both report Zn efficacy in improving TER across CACO-2 cell layers at a 50-100 µM concentration range, though without an accompanying decrease in the leak of the paracellular probe, [14]C-D-mannitol [52,53]. A change in localization of the TJ proteins, Claudins -2 and -7, was also observed. Shao et al. (2017) also reported increased TER across CACO-2 cell layers [54]. Buddington et al. (2021) did observe Zn-induced decrease in leak of the paracellular probe, FITC-dextran, across CACO-2 cell layers [55].
Studies conducted on animal models as well as human tissue have also provided evidence of Zn improving GI barrier function. In piglet intestine, Zn improved barrier function as evidenced by the lactulose/mannitol test, while also increasing occludin and ZO-1 content [22]. Hu et al (2013) observed a Zn-induced decrease of FITC-dextran diffusion across pig jejunum [56]. Zn-induced decrease of lactulose/mannitol ratio along with occludin upregulation has been seen in mouse and rat intestine as well [57,58].
Whereas the above examples are of Zn inducing barrier improvement of normal GI cell layers and tissue, Zn also has shown efficacy in countering the barrier leak induced chemically or by pathogens. This has been shown for Caco-2 and T84 human intestinal epithelial cell culture models challenged by Ochratoxin A, enteropathogenic E. coli, Cryptosporidium, Shigella and Salmonella [54,59,60,61,62]. In animal tissue, acetic acid- and dinitro-benzene-sulfonic acid-induced GI leak was likewise countered by Zn treatment [23,63].
Our study addresses whether orally administered Zn could improve (normal) GI barrier function in healthy humans. Although the above cell culture and animal model studies certainly suggest that Zn would be successful in humans, the unique complexities of the human model would leave uncertainty. There are obfuscating effects of varied diets, medications, and genetic backgrounds intrinsic to any study on humans. Camilleri in 2021, citing the scarcity of actual human in vivo data, considered this issue of Zn efficacy (and that of other micronutrients) in the human GI tract to be quite open [64]. However, Davison et al. (2016) did show in humans that Zn partially counters the increased GI permeability brought on by heavy exercise [65]. Roy et al. (1992) and Alam et al. (1994) showed Zn reduction of barrier leak in patients compromised by microbial-generated diarrhea [66,67]. Finally, there is the hallmark paper by Sturniolo et al. (2001) showing reduced lactulose/mannitol ratios after Zn treatment of Crohn’s Disease patients [23]. So, whereas the issue of Zn and GI permeability in humans is still marked by a scarcity of reports, those that do exist, do so in the context of Zn opposing some pathophysiological condition inducing leak. Our current study on the other hand looks at Zn improvement of GI barrier leak in healthy controls, a factor that would be essential for Zn being efficacious in a prophylactic/preventative role.
To summarize, we attempted in these studies to demonstrate that orally administered Zn (in the 60 mg/adult/day range) can successfully induce TJ remodeling in human small intestine resulting in a less leaky GI barrier. More studies are clearly needed to not only substantiate this finding but also to determine whether this orally administered Zn is active specifically from within the GI lumen (as opposed to acting from the bloodstream). There is also the question of whether similar activity can be seen in the large bowel and caecum, and whether Zn effects on the microbiota are involved here. A salutary effect of this safe, inexpensive ubiquitous micronutrient on something as clinically fundamental as GI barrier leak can potentially have far-ranging medical application.
Acknowledgements:
This study was supported by grants from the Sharpe Strumia Foundation to J.M.M. The authors are very grateful for the editorial assistance of Ms. Elene Mironidis and Ms. Terri Olshefski in the preparation of the manuscript for publication. The assistance of endoscopy procedures room personnel – including gastroenterology fellows - was invaluable in duodenal biopsy procurement. We are equally grateful to Ms. Laureen Nearey, Director of the Lankenau Medical Center Phlebotomy Lab, and her team of phlebotomists who secured our study's blood samples for analyses. Finally, the selfless participation of numerous gastroenterology patients as well as healthy control test subjects made this study possible.
List of Abbreviations
| CRP | C-Reactive Protein |
| GERD | Gastroesophageal Reflux Disease |
| GI | Gastrointestinal |
| GSEA | Gene Set Enrichment Analysis |
| HS CRP | High Sensitivity C-Reactive Protein |
| LG | Leaky Gut |
| NSAID | Non-Steroidal Anti-Inflammatory Drug |
| RDA | Recommended Daily Allowance |
| TER | Transepithelial Electrical Resistance |
| TJ | Tight Junction |
| Zn | Zinc |
References
- Guttman JA, Finlay BB. Tight junctions as targets of infectious agents. Biochim Biophys Acta. 2009 Apr;1788(4):832-41. Epub 2008 Nov 14. [CrossRef] [PubMed]
- Mullin JM, Agostino N, Rendon-Huerta E, Thornton JJ. Keynote review: epithelial and endothelial barriers in human disease. Drug Discov Today. 2005 Mar 15;10(6):395-408. [CrossRef] [PubMed]
- Davidson G, Kritas S, Butler R. Stressed mucosa. Nestle Nutr Workshop Ser Pediatr Program. 2007;59:133-42; discussion 143-6. [CrossRef] [PubMed]
- Capaldo, CT. Claudin Barriers on the Brink: How Conflicting Tissue and Cellular Priorities Drive IBD Pathogenesis. Int J Mol Sci. 2023 ;24(10):8562. 10 May. [CrossRef] [PubMed] [PubMed Central]
- Soler AP, Miller RD, Laughlin KV, Carp NZ, Klurfeld DM, Mullin JM. Increased tight junctional permeability is associated with the development of colon cancer. Carcinogenesis. 1999 Aug;20(8):1425-31. [CrossRef] [PubMed]
- Shrestha A, Uzal FA, McClane BA. The interaction of Clostridium perfringens enterotoxin with receptor claudins. Anaerobe. 2016 Oct;41:18-26. Epub 2016 Apr 16. [CrossRef] [PubMed] [PubMed Central]
- Hollander, D. Intestinal permeability, leaky gut, and intestinal disorders. Curr Gastroenterol Rep. 1999 Oct;1(5):410-6. [CrossRef] [PubMed]
- Compare D, Sgamato C, Rocco A, Coccoli P, Ambrosio C, Nardone G. The Leaky Gut and Human Diseases: "Can't Fill the Cup if You Don't Plug the Holes First". Dig Dis. 2024 Jul 24:1-19. Epub ahead of print. [CrossRef] [PubMed]
- Balkovetz, DF. Tight junction claudins and the kidney in sickness and in health. Biochim Biophys Acta. 2009 Apr;1788(4):858-63. Epub 2008 Jul 16. [CrossRef] [PubMed]
- Groeger SE, Meyle J. Epithelial barrier and oral bacterial infection. Periodontol 2000. 2015 Oct;69(1):46-67. [CrossRef] [PubMed]
- Wittekindt, OH. Tight junctions in pulmonary epithelia during lung inflammation. Pflugers Arch. 2017 Jan;469(1):135-147. Epub 2016 Dec 5. [CrossRef] [PubMed] [PubMed Central]
- Wang M, Li H, Wang F. Roles of Transepithelial Electrical Resistance in Mechanisms of Retinal Pigment Epithelial Barrier and Retinal Disorders. Discov Med. 2022 Jul- Aug;34(171):19-24. [PubMed]
- John LJ, Fromm M, Schulzke JD. Epithelial barriers in intestinal inflammation. Antioxid Redox Signal. 2011 Sep 1;15(5):1255-70. Epub 2011 May 11. [CrossRef] [PubMed]
- Suzuki, T. Regulation of intestinal epithelial permeability by tight junctions. Cell Mol Life Sci. 2013 Feb;70(4):631-59. Epub 2012 Jul 11. [CrossRef] [PubMed] [PubMed Central]
- Köhler CA, Maes M, Slyepchenko A, Berk M, Solmi M, Lanctôt KL, Carvalho AF. The Gut- Brain Axis, Including the Microbiome, Leaky Gut and Bacterial Translocation: Mechanisms and Pathophysiological Role in Alzheimer's Disease. Curr Pharm Des. 2016;22(40):6152- 6166. [CrossRef] [PubMed]
- Hu CA, Hou Y, Yi D, Qiu Y, Wu G, Kong X, Yin Y. Autophagy and tight junction proteins in the intestine and intestinal diseases. Anim Nutr. 2015 Sep;1(3):123-127. Epub 2015 Sep 1. [CrossRef] [PubMed] [PubMed Central]
- DiGuilio KM, Del Rio EA, Harty RN, Mullin JM. Micronutrients at Supplemental Levels, Tight Junctions and Epithelial Barrier Function: A Narrative Review. Int J Mol Sci. 2024 Mar 19;25(6):3452. [CrossRef] [PubMed] [PubMed Central]
- Vargas-Robles H, Castro-Ochoa KF, Citalán-Madrid AF, Schnoor M. Beneficial effects of nutritional supplements on intestinal epithelial barrier functions in experimental colitis models in vivo. World J Gastroenterol. 2019 Aug 14;25(30):4181-4198. [CrossRef] [PubMed] [PubMed Central]
- Hering NA, Schulzke JD. Therapeutic options to modulate barrier defects in inflammatory bowel disease. Dig Dis. 2009;27(4):450-4. Epub 2009 Nov 4. [CrossRef] [PubMed]
- Callaghan PJ, Rybakovsky E, Ferrick B, Thomas S, Mullin JM. Retinoic acid improves baseline barrier function and attenuates TNF-α-induced barrier leak in human bronchial epithelial cell culture model, 16HBE 14o. PLoS One. 2020 Dec 10;15(12):e0242536. [CrossRef] [PubMed] [PubMed Central]
- Rybakovsky E, DiGuilio KM, Valenzano MC, Geagan S, Pham K, Harty RN, Mullin JM. Calcitriol modifies tight junctions, improves barrier function, and reduces TNF-α-induced barrier leak in the human lung-derived epithelial cell culture model, 16HBE 14o. Physiol Rep. 2023 Apr;11(7):e15592. [CrossRef] [PubMed] [PubMed Central]
- Zhang B, Guo Y. Supplemental zinc reduced intestinal permeability by enhancing occludin and zonula occludens protein-1 (ZO-1) expression in weaning piglets. Br J Nutr. 2009 Sep;102(5):687-93. Epub 2009 Mar 9. [CrossRef] [PubMed]
- Sturniolo GC, Di Leo V, Ferronato A, D'Odorico A, D'Incà R. Zinc supplementation tightens "leaky gut" in Crohn's disease. Inflamm Bowel Dis. 2001 May;7(2):94-8. [CrossRef] [PubMed]
- Farrell CP, Morgan M, Rudolph DS, Hwang A, Albert NE, Valenzano MC, Wang X, Mercogliano G, Mullin JM. Proton Pump Inhibitors Interfere With Zinc Absorption and Zinc Body Stores. Gastroenterology Res. 2011 Dec;4(6):243-251. Epub 2011 Nov 20. [CrossRef] [PubMed] [PubMed Central]
- Fosmire, GJ. Zinc toxicity. Am J Clin Nutr. 1990 Feb;51(2):225-7. [CrossRef]
- Samman S, Roberts DC. The effect of zinc supplements on plasma zinc and copper levels and the reported symptoms in healthy volunteers. Med J Aust. 1987 Mar 2;146(5):246-9. [CrossRef] [PubMed]
- Bogden JD, Oleske JM, Lavenhar MA, Munves EM, Kemp FW, Bruening KS, Holding KJ, Denny TN, Guarino MA, Krieger LM, et al. Zinc and immunocompetence in elderly people: effects of zinc supplementation for 3 months. Am J Clin Nutr. 1988 Sep;48(3):655-63. [CrossRef] [PubMed]
- Fraga CG, Oteiza PI, Keen CL. Trace elements and human health. Mol Aspects Med. 2005 Aug-Oct;26(4-5):233-4. [CrossRef] [PubMed]
- Karam, SM. Lineage commitment and maturation of epithelial cells in the gut. Front Biosci. 1999 Mar 15;4:D286-98. [CrossRef] [PubMed]
- Dobin A, Davis CA, Schlesinger F, Drenkow J, Zaleski C, Jha S, Batut P, Chaisson M, Gingeras TR. STAR: ultrafast universal RNA-seq aligner. Bioinformatics. 2013 Jan 1;29(1):15-21. Epub 2012 Oct 25. [CrossRef] [PubMed] [PubMed Central]
- Li B, Dewey CN. RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinformatics. 2011 Aug 4;12:323. [CrossRef] [PubMed] [PubMed Central]
- Love MI, Huber W, Anders S. Moderated estimation of fold change and dispersion for RNA- seq data with DESeq2. Genome Biol. 2014;15(12):550. [CrossRef] [PubMed] [PubMed Central]
- Subramanian A, Tamayo P, Mootha VK, Mukherjee S, Ebert BL, Gillette MA, Paulovich A, Pomeroy SL, Golub TR, Lander ES, Mesirov JP. Gene set enrichment analysis: a knowledge- based approach for interpreting genome-wide expression profiles. Proc Natl Acad Sci U S A. 2005 Oct 25;102(43):15545-50. Epub 2005 Sep 30. [CrossRef] [PubMed] [PubMed Central]
- Aldridge GM, Podrebarac DM, Greenough WT, Weiler IJ. The use of total protein stains as loading controls: an alternative to high-abundance single-protein controls in semi-quantitative immunoblotting. J Neurosci Methods. 2008 Jul 30;172(2):250-4. Epub 2008 May 15. [CrossRef] [PubMed] [PubMed Central]
- Valenzano MC, Rybakovsky E, Chen V, Leroy K, Lander J, Richardson E, Yalamanchili S, McShane S, Mathew A, Mayilvaganan B, Connor L, Urbas R, Huntington W, Corcoran A, Trembeth S, McDonnell E, Wong P, Newman G, Mercogliano G, Zitin M, Etemad B, Thornton J, Daum G, Raines J, Kossenkov A, Fong LY, Mullin JM. Zinc Gluconate Induces Potentially Cancer Chemopreventive Activity in Barrett's Esophagus: A Phase 1 Pilot Study. Dig Dis Sci. 2021 Apr;66(4):1195-1211. Epub 2020 May 15. [CrossRef] [PubMed] [PubMed Central]
- Murray MJ, Barbose JJ, Cobb CF. Serum D(-)-lactate levels as a predictor of acute intestinal ischemia in a rat model. J Surg Res. 1993 May;54(5):507-9. [CrossRef] [PubMed]
- Abdel-Mageed AB, Oehme FW. The effect of various dietary zinc concentrations on the biological interactions of zinc, copper, and iron in rats. Biol Trace Elem Res. 1991 Jun;29(3):239-56. [CrossRef] [PubMed]
- Segawa S, Shibamoto M, Ogawa M, Miyake S, Mizumoto K, Ohishi A, Nishida K, Nagasawa, K. The effect of divalent metal cations on zinc uptake by mouse Zrt/Irt-like protein 1 (ZIP1). Life Sci. 2014 Sep 15;113(1-2):40-4. Epub 2014 Jul 31. [CrossRef] [PubMed]
- Robles-Osorio ML, Sabath E. Tight junction disruption and the pathogenesis of the chronic complications of diabetes mellitus: A narrative review. World J Diabetes. 2023 Jul 15;14(7):1013-1026. [CrossRef] [PubMed] [PubMed Central]
- Chegini Z, Noei M, Hemmati J, Arabestani MR, Shariati A. The destruction of mucosal barriers, epithelial remodeling, and impaired mucociliary clearance: possible pathogenic mechanisms of Pseudomonas aeruginosa and Staphylococcus aureus in chronic rhinosinusitis. Cell Commun Signal. 2023 Oct 30;21(1):306. [CrossRef] [PubMed] [PubMed Central]
- Yang YF, Shen YD. Choroid plexus and its relations with age-related diseases. Yi Chuan. 2024 Feb 20;46(2):109-125. [CrossRef] [PubMed]
- Berni Canani R, Caminati M, Carucci L, Eguiluz-Gracia I. Skin, gut, and lung barrier: Physiological interface and target of intervention for preventing and treating allergic diseases. Allergy. 2024 Jun;79(6):1485-1500. Epub 2024 Mar 4. [CrossRef] [PubMed]
- DiGuilio KM, Rybakovsky E, Abdavies R, Chamoun R, Flounders CA, Shepley-McTaggart A, Harty RN, Mullin JM. Micronutrient Improvement of Epithelial Barrier Function in Various Disease States: A Case for Adjuvant Therapy. Int J Mol Sci. 2022 Mar 10;23(6):2995. [CrossRef] [PubMed] [PubMed Central]
- Paterson BM, Lammers KM, Arrieta MC, Fasano A, Meddings JB. The safety, tolerance, pharmacokinetic and pharmacodynamic effects of single doses of AT-1001 in coeliac disease subjects: a proof of concept study. Aliment PharmacolTher. 2007 Sep 1;26(5):757-66. [CrossRef] [PubMed]
- Brenner H, Holleczek B, Schöttker B. Vitamin D Insufficiency and Deficiency and Mortality from Respiratory Diseases in a Cohort of Older Adults: Potential for Limiting the Death Toll during and beyond the COVID-19 Pandemic? Nutrients. 2020 Aug 18;12(8):2488. [CrossRef] [PubMed] [PubMed Central]
- Obrenovich MEM. Leaky Gut, Leaky Brain? Microorganisms. 2018 Oct 18;6(4):107. [CrossRef] [PubMed] [PubMed Central]
- Wasiak J, Gawlik-Kotelnicka O. Intestinal permeability and its significance in psychiatric disorders - A narrative review and future perspectives. Behav Brain Res. 2023 Jun 25;448:114459. Epub 2023 Apr 29. [CrossRef] [PubMed]
- Brown GC, Heneka MT. The endotoxin hypothesis of Alzheimer's disease. Mol Neurodegener. 2024 Apr 1;19(1):30. [CrossRef] [PubMed] [PubMed Central]
- Lamberti LM, Walker CL, Chan KY, Jian WY, Black RE. Oral zinc supplementation for the treatment of acute diarrhea in children: a systematic review and meta-analysis. Nutrients. 2013 Nov 21;5(11):4715-40. [CrossRef] [PubMed] [PubMed Central]
- Amasheh M, Andres S, Amasheh S, Fromm M, Schulzke JD. Barrier effects of nutritional factors. Ann N Y Acad Sci. 2009 May;1165:267-73. [CrossRef] [PubMed]
- Skrovanek S, DiGuilio K, Bailey R, Huntington W, Urbas R, Mayilvaganan B, Mercogliano G, Mullin JM. Zinc and gastrointestinal disease. World J Gastrointest Pathophysiol. 2014 Nov 15;5(4):496-513. [CrossRef] [PubMed] [PubMed Central]
- Wang X, Valenzano MC, Mercado JM, Zurbach EP, Mullin JM. Zinc supplementation modifies tight junctions and alters barrier function of CACO-2 human intestinal epithelial layers. Dig Dis Sci. 2013 Jan;58(1):77-87. Epub 2012 Aug 19. [CrossRef] [PubMed]
- Valenzano MC, DiGuilio K, Mercado J, Teter M, To J, Ferraro B, Mixson B, Manley I, Baker V, Moore BA, Wertheimer J, Mullin JM. Remodeling of Tight Junctions and Enhancement of Barrier Integrity of the CACO-2 Intestinal Epithelial Cell Layer by Micronutrients. PLoS One. 2015 Jul 30;10(7):e0133926. [CrossRef] [PubMed] [PubMed Central]
- Shao Y, Wolf PG, Guo S, Guo Y, Gaskins HR, Zhang B. Zinc enhances intestinal epithelial barrier function through the PI3K/AKT/mTOR signaling pathway in Caco-2 cells. J Nutr Biochem. 2017 May;43:18-26. Epub 2017 Jan 31. [CrossRef] [PubMed]
- Buddington RK, Wong T, Howard SC. Paracellular Filtration Secretion Driven by Mechanical Force Contributes to Small Intestinal Fluid Dynamics. Med Sci (Basel). 2021 Feb 9;9(1):9. [CrossRef] [PubMed] [PubMed Central]
- Hu C, Song J, Li Y, Luan Z, Zhu K. Diosmectite-zinc oxide composite improves intestinal barrier function, modulates expression of pro-inflammatory cytokines and tight junction protein in early weaned pigs. Br J Nutr. 2013 Aug;110(4):681-8. Epub 2013 Jan 11. [CrossRef] [PubMed]
- Guthrie GJ, Aydemir TB, Troche C, Martin AB, Chang SM, Cousins RJ. Influence of ZIP14 (slc39A14) on intestinal zinc processing and barrier function. Am J Physiol Gastrointest Liver Physiol. 2015 Feb 1;308(3):G171-8. Epub 2014 Nov 26. [CrossRef] [PubMed] [PubMed Central]
- Fan P, Tan Y, Jin K, Lin C, Xia S, Han B, Zhang F, Wu L, Ma X. Supplemental lipoic acid relieves post-weaning diarrhoea by decreasing intestinal permeability in rats. J Anim Physiol Anim Nutr (Berl). 2017 Feb;101(1):136-146. Epub 2015 Dec 31. [CrossRef] [PubMed]
- Ranaldi G, Caprini V, Sambuy Y, Perozzi G, Murgia C. Intracellular zinc stores protect the intestinal epithelium from Ochratoxin A toxicity. Toxicol In Vitro. 2009 Dec;23(8):1516-21. Epub 2009 Aug 29. [CrossRef] [PubMed]
- Roselli M, Finamore A, Garaguso I, Britti MS, Mengheri E. Zinc oxide protects cultured enterocytes from the damage induced by Escherichia coli. J Nutr. 2003 Dec;133(12):4077-82. [CrossRef] [PubMed]
- Choudhry N, Scott F, Edgar M, Sanger GJ, Kelly P. Reversal of Pathogen-Induced Barrier Defects in Intestinal Epithelial Cells by Contra-pathogenicity Agents. Dig Dis Sci. 2021 Jan;66(1):88-104. Epub 2020 Feb 7. [CrossRef] [PubMed]
- Sarkar P, Saha T, Sheikh IA, Chakraborty S, Aoun J, Chakrabarti MK, Rajendran VM, Ameen NA, Dutta S, Hoque KM. Zinc ameliorates intestinal barrier dysfunctions in shigellosis by reinstating claudin-2 and -4 on the membranes. Am J Physiol Gastrointest Liver Physiol. 2019 Feb 1;316(2):G229-G246. Epub 2018 Nov 8. [CrossRef] [PubMed] [PubMed Central]
- Song ZH, Ke YL, Xiao K, Jiao LF, Hong QH, Hu CH. Diosmectite-zinc oxide composite improves intestinal barrier restoration and modulates TGF-β1, ERK1/2, and Akt in piglets after acetic acid challenge. J Anim Sci. 2015 Apr;93(4):1599-607. [CrossRef] [PubMed]
- Camilleri, M. Human Intestinal Barrier: Effects of Stressors, Diet, Prebiotics, and Probiotics. Clin Transl Gastroenterol. 2021 Jan 25;12(1):e00308. [CrossRef] [PubMed] [PubMed Central]
- Davison G, Marchbank T, March DS, Thatcher R, Playford RJ. Zinc carnosine works with bovine colostrum in truncating heavy exercise-induced increase in gut permeability in healthy volunteers. Am J aClin Nutr. 2016 Aug;104(2):526-36. Epub 2016 Jun 29. [CrossRef] [PubMed]
- Roy SK, Behrens RH, Haider R, Akramuzzaman SM, Mahalanabis D, Wahed MA, Tomkins AM. Impact of zinc supplementation on intestinal permeability in Bangladeshi children with acute diarrhoea and persistent diarrhoea syndrome. J Pediatr Gastroenterol Nutr. 1992 Oct;15(3):289-96. [CrossRef] [PubMed]
- Alam J, Nuzhat S, Billal SM, Ahmed T, Khan AI, Hossain MI. Nutritional Profiles and Zinc Supplementation among Children with Diarrhea in Bangladesh. Am J Trop Med Hyg. 2023 Feb 27;108(4):837-843. [CrossRef] [PubMed] [PubMed Central]
Figure 1.
Transcriptional Changes in Duodenal Epithelial Biopsy Tissue in Response to Zinc-Gluconate Treatment. Six patients (“zinc gluconate”) were treated with orally administered Zn gluconate (26 mg Zn BID) for 14 days prior to upper endoscopy and duodenal mucosa biopsy as described in Materials and Methods. Five patients (“placebo”) were treated with a molar equivalent amount of sodium gluconate BID. RNA was extracted from the biopsy samples and RNA RNA-seq analyses were performed also as described. Gene set enrichment analysis was performed by GSEA [33]. A. Expression heatmap of select individual genes related to Zn binding and TJ regulation. Fold; fold increase or decrease relative to transcription level in placebo-treated tissue; pv = DESeq2 P value; color intensity represents relative expression levels (vs placebo average) across individual patients: red – relative upregulation; blue – relative downregulation. B. Select Categories of Genes Enriched in Response to Zn Gluconate Treatment As Identified by Gene Set Enrichment Analysis (GSEA). These are common functional categories enriched among genes affected by Zn gluconate treatment. Among other Zn-induced patterns unrelated to epithelial barrier function (unreported here; see Supplemental Material), a pattern of upregulations of gene transcriptions relating to epithelial junction integrity was observed in the duodenal tissue samples. A pattern of associated downregulations in modifiers of the proinflammatory state, known junctional signaling pathways such as ERK 1/2, and general modifiers of dedifferentiation, was also in evidence. Those downregulations could be consistent with improved junctional barrier function. n = number of leading-edge genes; NES = normalized enrichment score; pv = P value of the enrichment. Category types: GOBP = Gene Ontology Biological Process; GOCC = Gene Ontology Cellular Component; REAC = REACTOME pathways; KEGG = KEGG pathways; HALL = HALLMARK gene sets.
Figure 1.
Transcriptional Changes in Duodenal Epithelial Biopsy Tissue in Response to Zinc-Gluconate Treatment. Six patients (“zinc gluconate”) were treated with orally administered Zn gluconate (26 mg Zn BID) for 14 days prior to upper endoscopy and duodenal mucosa biopsy as described in Materials and Methods. Five patients (“placebo”) were treated with a molar equivalent amount of sodium gluconate BID. RNA was extracted from the biopsy samples and RNA RNA-seq analyses were performed also as described. Gene set enrichment analysis was performed by GSEA [33]. A. Expression heatmap of select individual genes related to Zn binding and TJ regulation. Fold; fold increase or decrease relative to transcription level in placebo-treated tissue; pv = DESeq2 P value; color intensity represents relative expression levels (vs placebo average) across individual patients: red – relative upregulation; blue – relative downregulation. B. Select Categories of Genes Enriched in Response to Zn Gluconate Treatment As Identified by Gene Set Enrichment Analysis (GSEA). These are common functional categories enriched among genes affected by Zn gluconate treatment. Among other Zn-induced patterns unrelated to epithelial barrier function (unreported here; see Supplemental Material), a pattern of upregulations of gene transcriptions relating to epithelial junction integrity was observed in the duodenal tissue samples. A pattern of associated downregulations in modifiers of the proinflammatory state, known junctional signaling pathways such as ERK 1/2, and general modifiers of dedifferentiation, was also in evidence. Those downregulations could be consistent with improved junctional barrier function. n = number of leading-edge genes; NES = normalized enrichment score; pv = P value of the enrichment. Category types: GOBP = Gene Ontology Biological Process; GOCC = Gene Ontology Cellular Component; REAC = REACTOME pathways; KEGG = KEGG pathways; HALL = HALLMARK gene sets.
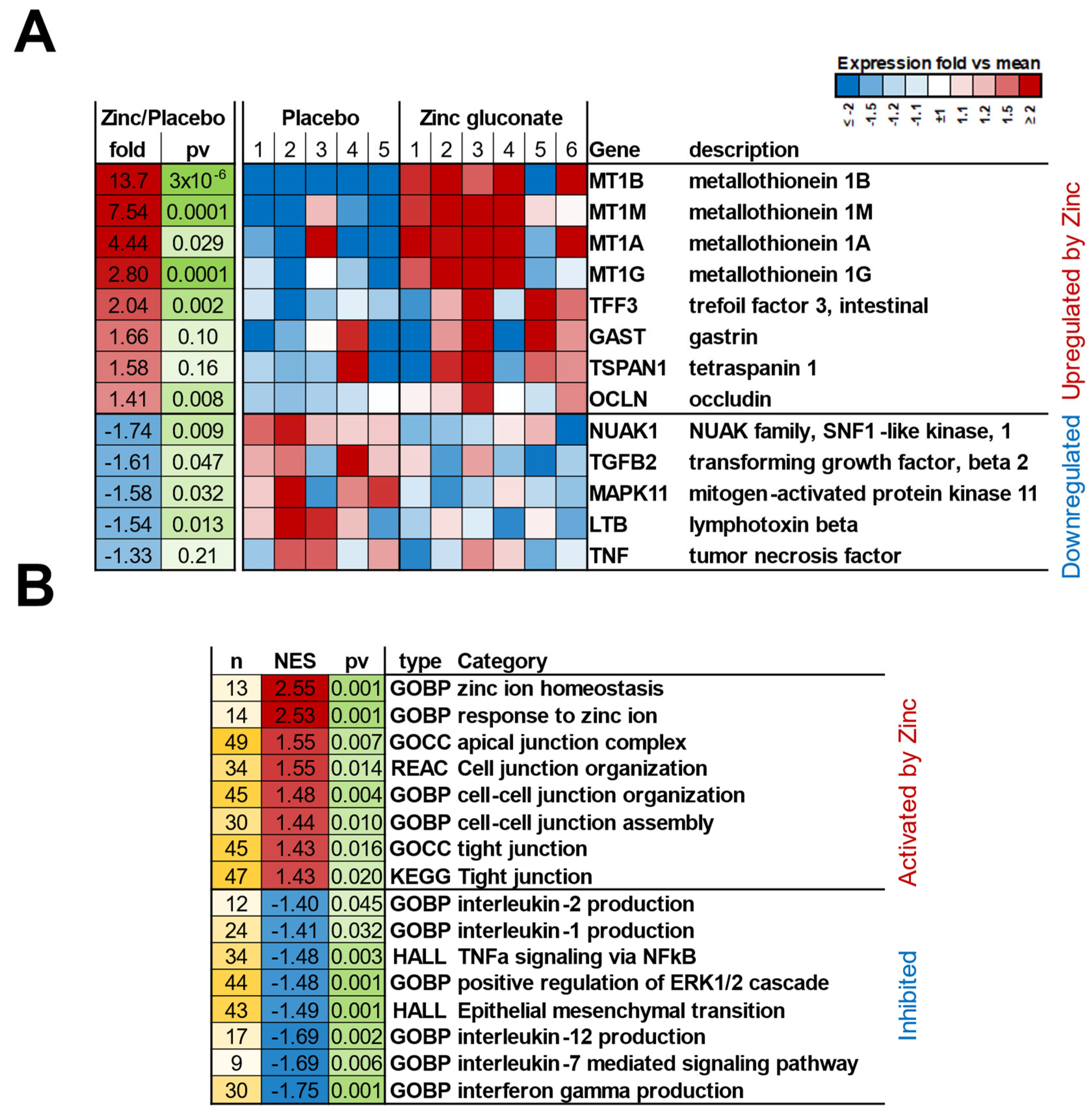
Figure 2.
Western Immunoblot Analyses of Tight Junctional Proteins in Duodenal Mucosal Biopsies As a Result of Orally Administered Zinc Treatment. Eleven patients were treated with orally administered Zn gluconate (26 mg Zn BID) for 14 days prior to upper endoscopy and duodenal mucosa biopsy as described in Materials and Methods. Twelve patients were treated with a molar equivalent sodium gluconate placebo. Whole cell lysates of the biopsy tissue were performed as described in Materials and Methods, followed by PAGE and Western immunoblot for seven specific TJ proteins. Densitometry was performed as described to quantify results. All results were normalized to total protein based on MEMCODE assays. Each data point shown represents the result for a single patient. Significance was assessed by two-sided Student’s t-tests. .
Figure 2.
Western Immunoblot Analyses of Tight Junctional Proteins in Duodenal Mucosal Biopsies As a Result of Orally Administered Zinc Treatment. Eleven patients were treated with orally administered Zn gluconate (26 mg Zn BID) for 14 days prior to upper endoscopy and duodenal mucosa biopsy as described in Materials and Methods. Twelve patients were treated with a molar equivalent sodium gluconate placebo. Whole cell lysates of the biopsy tissue were performed as described in Materials and Methods, followed by PAGE and Western immunoblot for seven specific TJ proteins. Densitometry was performed as described to quantify results. All results were normalized to total protein based on MEMCODE assays. Each data point shown represents the result for a single patient. Significance was assessed by two-sided Student’s t-tests. .
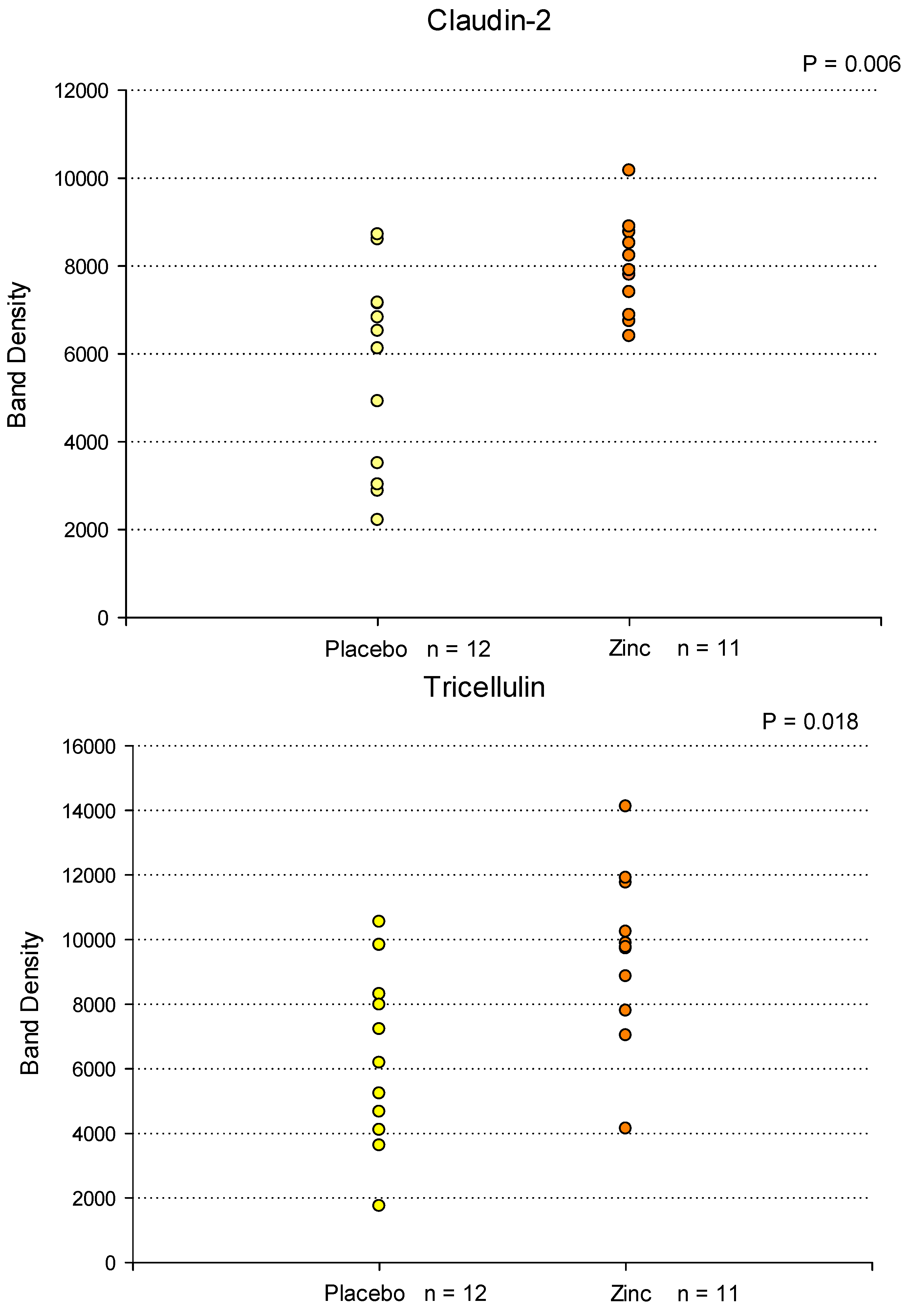
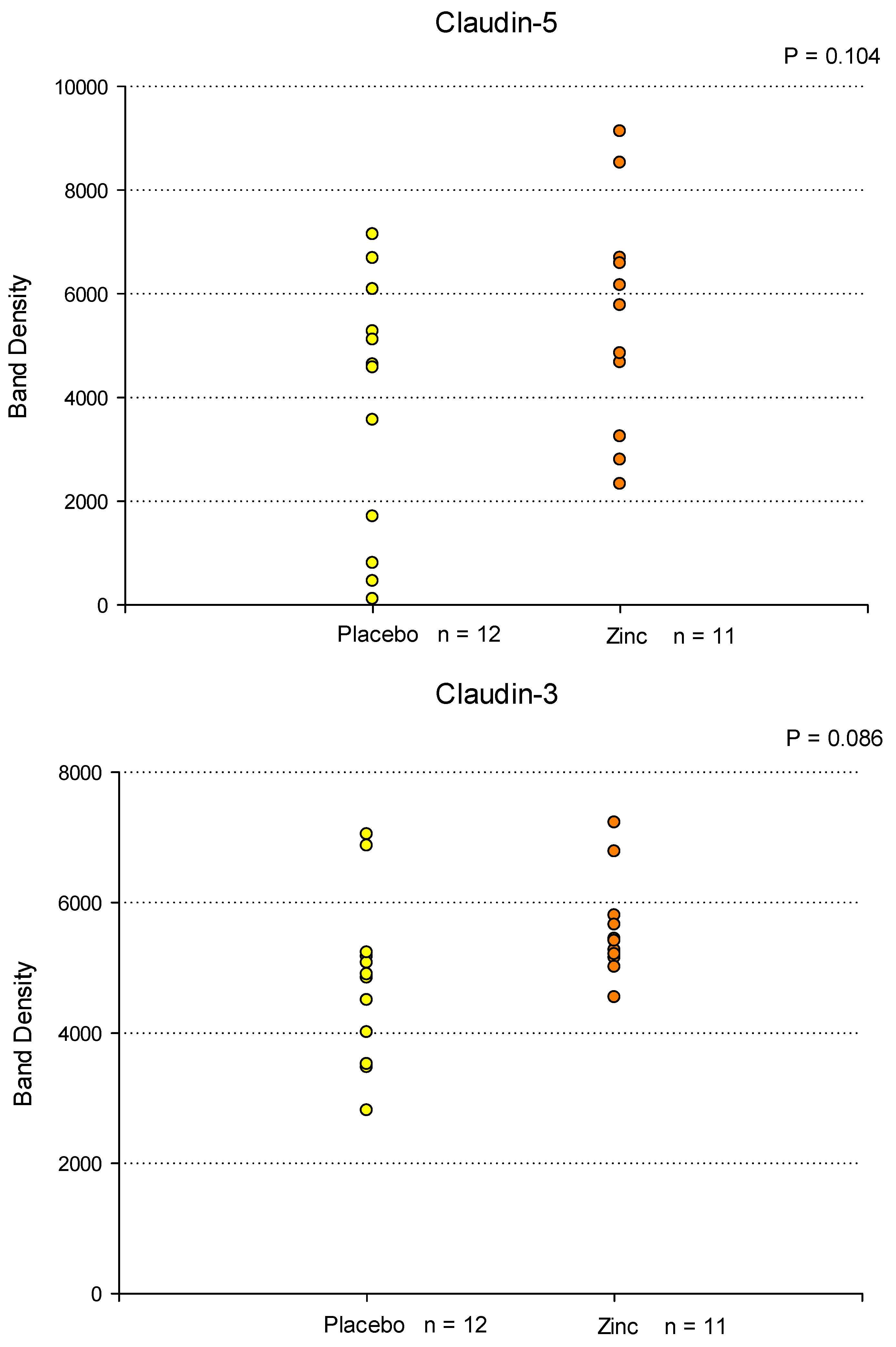
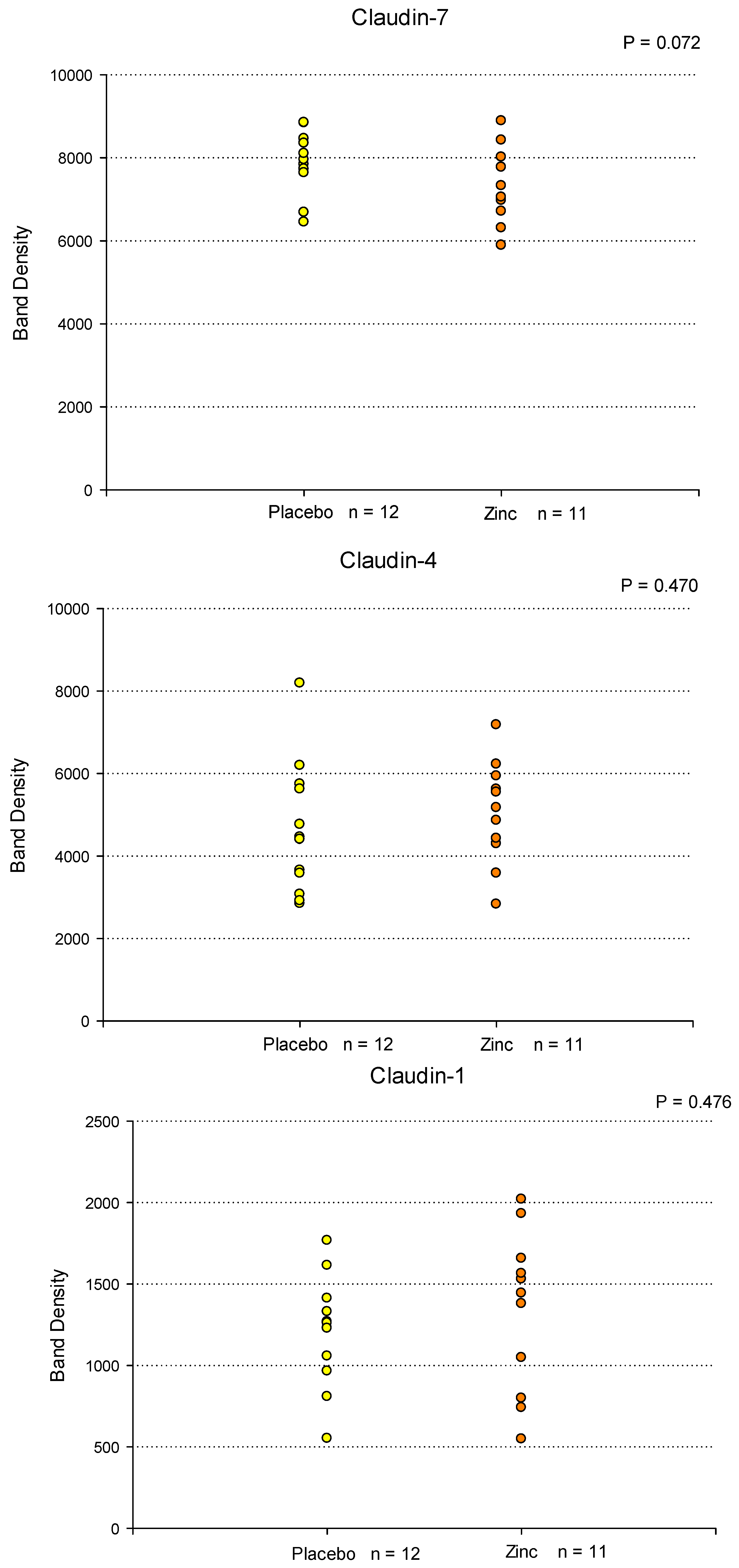
Figure 3.
Blood Levels of D-Lactate, Hemoglobin A1C and High Sensitivity CRP Before and After a 7 Day Regimen of Orally Administered Zinc. Healthy control test subjects (n = 9) were treated with Zn gluconate as described in Materials and Methods. Peripheral blood samples were taken before and after the Zn regimen. Results are shown for serum levels of the GI permeability marker D-Lactate (Panel A), along with two unrelated serum parameters, Hemoglobin A1C (Panel B) and High Sensitivity CRP (Panel C), determined spectrophotometrically as described. Whereas no obvious trend was evident for Hemoglobin A1C as a result of Zn treatment, post Zn levels of D-Lactate were significantly reduced (P = 0.008 [paired Student’s t-test] ). A significant reduction was observed for High Sensitivity CRP as well (P = 0.047).
Figure 3.
Blood Levels of D-Lactate, Hemoglobin A1C and High Sensitivity CRP Before and After a 7 Day Regimen of Orally Administered Zinc. Healthy control test subjects (n = 9) were treated with Zn gluconate as described in Materials and Methods. Peripheral blood samples were taken before and after the Zn regimen. Results are shown for serum levels of the GI permeability marker D-Lactate (Panel A), along with two unrelated serum parameters, Hemoglobin A1C (Panel B) and High Sensitivity CRP (Panel C), determined spectrophotometrically as described. Whereas no obvious trend was evident for Hemoglobin A1C as a result of Zn treatment, post Zn levels of D-Lactate were significantly reduced (P = 0.008 [paired Student’s t-test] ). A significant reduction was observed for High Sensitivity CRP as well (P = 0.047).
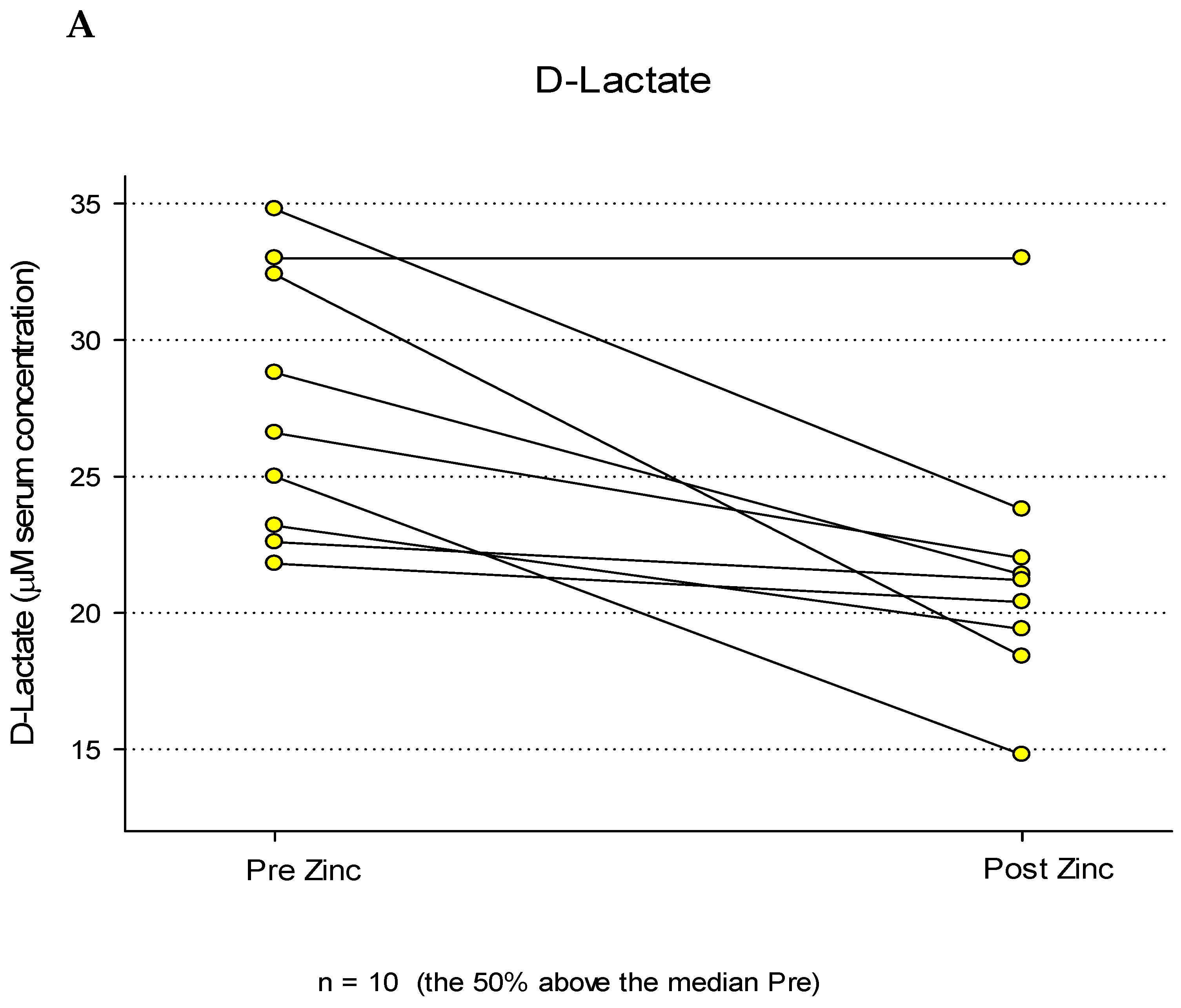
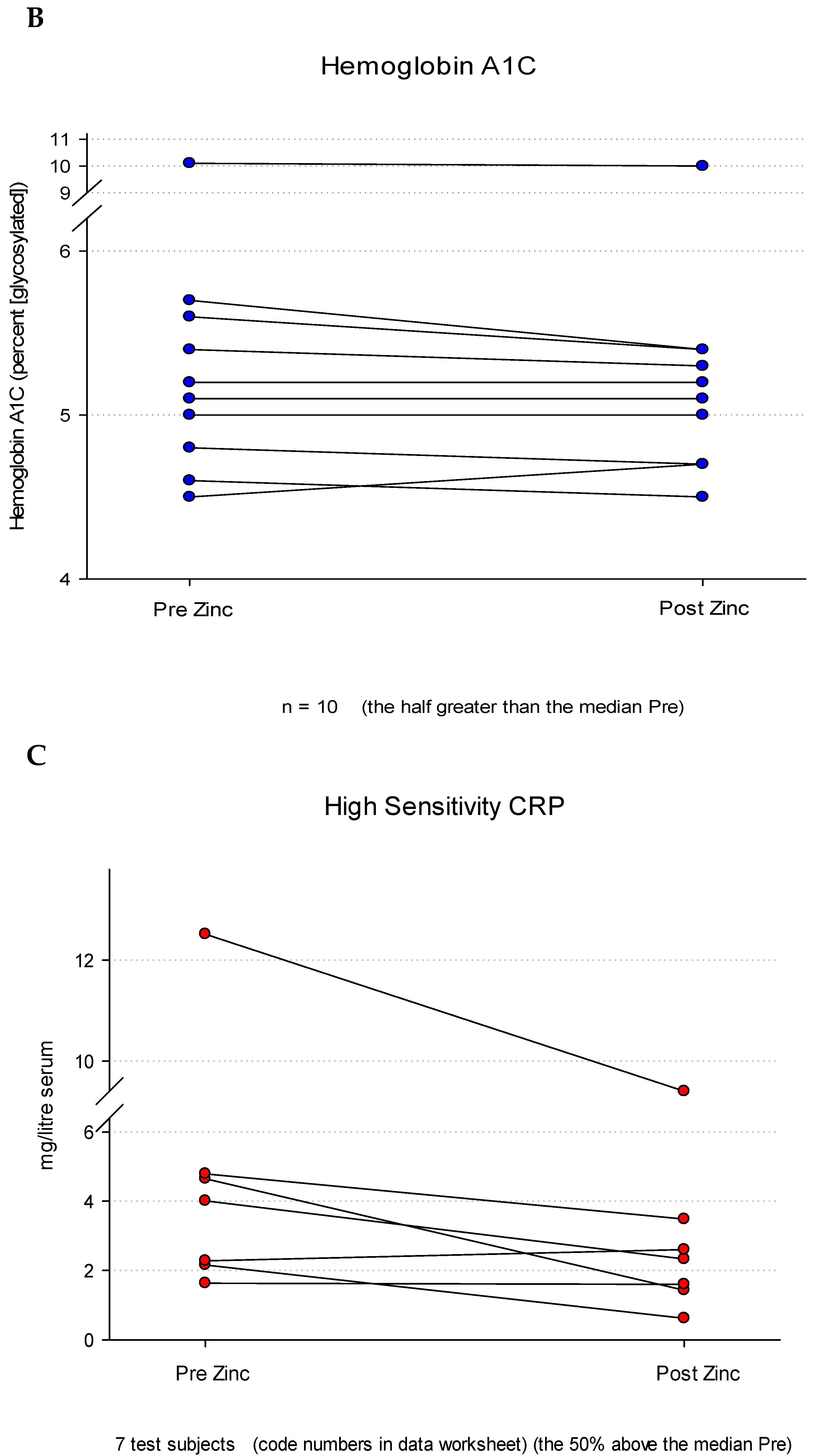

Table 1.
Patient and Test Subject Demographics Relating to mRNA Microarray, Western Immunoblot and Serum D-Lactate Analyses.
Table 1.
Patient and Test Subject Demographics Relating to mRNA Microarray, Western Immunoblot and Serum D-Lactate Analyses.
| A. RNA Microarray Study | ||
| 1. Placebo Group (n = 5) | ||
| a. Mean age: 71.6 (range: 64 - 79) | ||
| b. Gender distribution: 2 male, 3 female | ||
| c. Racial composition: 5 Caucasian | ||
| 2. Zn-Treated Group (n = 6) | ||
| a. Mean age: 71.0 (range: 61 - 76) | ||
| b. Gender distribution: 1 male, 5 female | ||
| c. Racial composition: 5 Caucasian, 1 Afr. American | ||
| B. Western Immunoblot Study | ||
| 1. Placebo Group (n = 12) | ||
| a. Mean age: 68 (range: 55 - 78) | ||
| b. Gender distribution: 7 males, 5 females | ||
| c. Racial composition: 12 Caucasian | ||
| 2. Zn-Treated Group (n = 11) | ||
| a. Mean age: 68 (range: 54 - 75) | ||
| b. Gender distribution: 5 males, 6 females | ||
| c. Racial composition: 10 Caucasian, 1 Afr. American | ||
| C. Serum D-Lactate Study (n = 19) | ||
| 1. Mean age: 44.0 (range: 21 - 71) | ||
| 2. Gender distribution: 11 male, 8 female | ||
| 3. Racial composition: 16 Caucasian, 2 African American, 1 Asian |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



