Submitted:
13 April 2025
Posted:
14 April 2025
You are already at the latest version
Abstract
The widespread dissemination of the blaNDM gene, which encodes New Delhi metallo-β-lactamase, in animal-derived settings poses a threat to public health security. Live poultry markets represent critical nodes in public health surveillance. However, there is currently limited reporting on the spread of the blaNDM gene within these markets under One Health approach. This study investigated the prevalence of the blaNDM gene in live poultry markets and performed an in-depth analysis of its association networks with other genetic elements across species, by integrating newly sequenced genomes with publicly available database entries. The samples for this study were collected from two live poultry markets in Jiangsu, China. Among the blaNDM-positive strains identified, we detected multiple variants, primarily blaNDM-5, followed by blaNDM-1, blaNDM-13, blaNDM-27, and blaNDM-39. We detected the coexistence of blaNDM-5 and mcr-1 in five Escherichia coli strains. Additionally, we found one E. coli strain in which blaNDM-5 coexisted with estT and tet(X4), and another E. coli strain where blaNDM-5 coexisted with estT. Network analysis of publicly available genomes revealed that the genetic element preferences of blaNDM variants vary significantly across species. The genetic element preferences of Escherichia coli carrying blaNDM-5 are similar to those of Klebsiella pneumoniae harboring blaNDM-1. In Klebsiella aerogenes, Enterobacter cloacae, and Proteus mirabilis, strains carrying blaNDM-1 have opposite genetic element preferences compared to strains harboring blaNDM-5 or blaNDM-7. Notably, we report the first evidence of the blaNDM-1 gene transfer mediated by ISKpn13, ISSpu2, and MITEKpn1. The findings highlight live poultry markets were important transmission hotspot of AMR, which requires continuous surveillance.
Keywords:
Live poultry
; carbapenem resistance
; blaNDM
; genomic analysis
; transmission
Introduction
Antibiotics are the primary weapons for humans to combat various infectious diseases and have made significant contributions to human and animal health in the fields of medicine, animal husbandry, and food safety. However, with the widespread use of antibiotics, antimicrobial resistance (AMR) has become a major threat to global public health, and the increasing multidrug resistance (MDR) in clinical pathogens has further exacerbated the problem. Horizontal gene transfer of antibiotic resistance genes (ARGs) across ecological niches amplifies the risk of clinical resistance. Globally, live poultry markets are high-risk interfaces for human-animal contact. These markets aggregate poultry from diverse regions, facilitating ARGs transfer and pathogen dissemination[1]. Live poultry markets have been proven to be reservoirs and dissemination centers for ARGs[2]. The ARGs detected in people, poultry, and the environment within the markets are more diverse than those detected in poultry farms[3]. This indicates that the risk of ARGs spreading through food animals is high, and they can easily be further disseminated through pathways such as water and air[4], posing a threat to the entire public health security.
Carbapenems, broad-spectrum antibiotics reserved for human MDR infections (and prohibited in veterinary use), are a last-line defense. Nevertheless, the increasing prevalence of carbapenem-resistant Gram-negative bacteria in recent years has raised significant concerns in the global public health community. The blaNDM gene, which encodes New Delhi metallo-β-lactamase (NDM), is a clinically significant determinant of carbapenem resistance. Its product can degrade the majority of β-lactam antibiotics, thereby compromising the effectiveness of these agents against pathogens that harbor this gene. To date, the spread of the blaNDM gene across different ecological niches has been extensively documented[5,6,7,8,9]. However, information regarding the prevalence of blaNDM in live poultry markets remains limited.
In this study, we isolated and identified multiple blaNDM-positive strains from various ecological niches within live poultry markets. Through whole-genome sequencing (WGS), we elucidated the genomic characteristics of these blaNDM-harboring strains. By integrating the sequenced genomes with those available in databases, we conducted a comprehensive analysis of the association networks between various blaNDM gene variants and other genetic elements across different species.
Materials and Methods
Sample Collection and Strain Identification
In July 2022, a total of 388 non-duplicate samples were collected from two large-scale live poultry markets in Yangzhou to investigate the epidemiology of blaNDM-positive strains in both animals and the environment. The poultry traded in these markets originated from Anhui Province and several cities in Jiangsu Province, including Huai’an, Nanjing, Nantong, Taizhou, Yangzhou, and Yancheng. The samples comprised animal feces (chicken, n = 159; duck, n = 29; goose, n = 66; pigeon, n = 21) and other samples (soil, n = 17; water, n = 36; environment, n = 57; plant, n = 3) (Table S1). All samples were transported to the laboratory in cool boxes with ice packs (4 ℃) for bacterial cultivation and DNA extraction. The collected samples were transferred into 2 ml Brain Heart Infusion (BHI) liquid broth and incubated at 37 ℃ for 6 h for pre-bacterial growth. Preculture samples were then spread onto MacConkey plates supplemented with 2 mg/L meropenem and incubated for 18 h at 37 ℃. Different colored colonies were selected from each plate to identify carbapenem-resistant isolates. All confirmed carbapenem-resistant strains were tested for the presence of blaNDM genes (Table S2). All blaNDM-positive bacteria were identified using MALDI-TOF MS AximaTM and 16S rRNA gene sequencing (Table S2).
Antimicrobial Susceptibility Testing
Antimicrobial susceptibility was tested using the broth dilution method. The susceptibility of carbapenem-resistant isolates was evaluated for a range of antimicrobial drugs commonly used in both human medicine and veterinary practice, including meropenem (MEM), imipenem (IMP), ampicillin (AMP), ceftazidime (CAZ), kanamycin (KAN), gentamicin (GEN), ciprofloxacin (CIP) and colistin (CL). Minimum inhibitory concentrations (MICs) were interpreted in accordance with the guidelines provided by the Clinical and Laboratory Standards Institute (2021) [10] and the breakpoint tables specified in the European Committee on Antimicrobial Susceptibility Testing v.12.0. E. coli ATCC 25922 was used as a quality control strain.
Plasmid Conjugation Assay
To explore the transferability of genetic elements carrying the blaNDM gene, we conducted a conjugation assay using rifampicin-resistant E. coli C600 as the recipient strain. The liquid mating method was utilized for this purpose. Initially, overnight cultures of the original isolates and recipient strains were prepared in Luria-Bertani (LB) liquid broth. These cultures were subsequently adjusted to an optical density of 0.6 at 600 nm. A volume of 50 μl of the mixed bacterial cultures was then pipetted and evenly spread onto LB solid media containing 100 μg/mL rifampicin and 2.0 μg/mL meropenem. Following an overnight incubation at 37℃, single bacterial colonies were selected for PCR analysis to confirm the successful transfer of the blaNDM gene.
Whole genome Sequencing of blaNDM-Positive Strains
The genomes of 38 blaNDM-positive strains were extracted using the FastPure Bacteria DNA Isolation Mini Kit (Vazyme, Nanjing, China). The concentration and purity of the extracted DNA were evaluated using NanoDrop 2000 and gel electrophoresis, with the final concentration determined precisely by the QubitTM 4.0 fluorometer (Invitrogen, CA, USA). Subsequently, short-read sequencing was performed on the extracted DNA using DNBseq, producing paired-end reads of 2×150 bp. The collected raw reads, with a minimum coverage of 100-fold, were then processed for trimming using SOAPnuke v.2.17[11]. De novo assembly was subsequently carried out using SPAdes v.3.13.1 [12].
Bioinformatics Analysis of Assembled Genomes
Mlst v.2.23.0 (https://github.com/tseemann/mlst) was used to determine multi-locus sequence type (MLST) of all assembled genomes. Resfinder[13], ISfinder[14], Plasmidfinder[15], VFDB core dataset[16] and ICEberg[17] were run with 80% coverage and 80% identity in Abricate (https://github.com/tseemann/abricate) to identify ARGs, insertion sequences (ISs), plasmid replicons, integrating conjugative elements (ICEs). ECTyper[18] was used to identify serotypes of all E. coli genomes. Prokka v.1.14.6[19] was used to conduct genome annotation. Phylogenetic trees were constructed using Roary v.3.13.0[20] and FastTree v.2.1.11[21] and visualized using Chiplot (https://www.chiplot.online). Heatmap was drawn using Chiplot. Genetic environment of plasmids was visualized using BRIG v.0.95 [22].
Genetic Environment Analysis of blaNDM-Positive Strains
In order to analyze the differences in the genetic environment among different blaNDM gene variants, we used Abricate (https://github.com/tseemann/abricate) to identify 4,072 blaNDM-positive strains (Table S3) from the Carbapenem-resistant Escherichia coli (CREC) dataset of a previous study[23]. We also downloaded 66,609 genomes from Klebsiella genus, 10,762 genomes from Enterobacter genus and 3,446 genomes from Proteus genus from the NCBI database (as of Dec 10, 2023). CheckM2 [24]was used to identify genomes with over 95% completeness and less than 5% contamination. Feature information including collection date, host, country, species and isolation source of these genomes were collected using a homemade python script. Linear genomic comparison and bar plot were visualized using ChiPlot. The network graph depicting the coexistence patterns of different blaNDM gene variants with other ARGs, ISs and plasmid replicons was constructed using Gephi [25].
Statistical Analysis
Statistical analysis and plotting were performed using R v.4.3.1 (R Foundation for Statistical Computing, Vienna, Austria). Spearman correlation analysis was used to determine the correlation among blaNDM gene, other ARGs, ISs and plasmid replicons.
Results
blaNDM-Positive Strains Profile
A total of 388 original samples were collected from two live poultry markets in Yangzhou, China. A total of 351 meropenem-resistant strains were isolated from these samples, among which 233 strains were blaNDM-positive (isolated from 144 original samples). The blaNDM detection rates were 37.11% (144/388) among samples and 66.38% (233/351) among meropenem-resistant isolates. Among the 233 blaNDM-positive strains, there were 218 Escherichia coli strains (93.56%), 4 Enterobacter cloacae strains (1.72%), 7 Klebsiella pneumoniae (3.00%), 2 Klebsiella aerogenes (0.86%), 1 Providencia rettgeri (0.43%), and 1 Proteus mirabilis (0.43%). Conjugation assays were conducted on the 233 blaNDM-positive strains, and ultimately 91 E. coli C600 transconjugants were obtained, with a conjugation success rate of 39.10%. Among the strains that successfully transferred the blaNDM gene through conjugation, all were E. coli except for 3 E. cloacae strains and 1 Proteus mirabilis strain.
A total of 233 blaNDM-positive strains were tested for susceptibility to a variety of antibiotics (Table 1). The tested strains exhibited extremely high resistance to meropenem, imipenem, ampicillin, and ceftazidime, with resistance rates approaching 100%. Among the aminoglycoside antibiotics, resistance rates to kanamycin and gentamicin were also high, reaching 87.12% and 82.40%, respectively. Additionally, the tested strains showed a resistance rate of 74.25% to ciprofloxacin and 20.12% to colistin. Only 11.19% (15/134) of strains isolated from chickens were resistant to colistin, while 50% (17/34) of strains from environmental sources were resistant to colistin. Although colistin demonstrated relatively good antimicrobial activity against blaNDM-positive strains, the presence of resistance must be taken seriously and monitored more closely.
Genomic Analysis of blaNDM-Positive Strains
To investigate the genetic characteristics of blaNDM-positive strains, we selected 38 representative blaNDM-positive strains for whole-genome sequencing and analysis, including 29 E. coli strains, 4 E. cloacae strains, 2 K. pneumoniae strains, 2 K. aerogenes strains, and 1 P. mirabilis strain. Based on core genome SNPs, we constructed a phylogenetic tree of 29 blaNDM-positive E. coli strains (Figure 1). The 29 E. coli strains from this study presented 18 distinct sequence types, with ST226 (13.79%, 4/29), ST6858 (13.79%, 4/29) and ST1630 (10.34%, 3/29) being the most prominent. A total of 19 serotypes were identified, mainly including O1:H45 (13.79%, 4/29), O8:H4 (13.79%, 4/29), and O16:H48 (10.34%, 3/29). We counted the number of virulence genes of all the E. coli strains based on the VFDB core datasets. It is worth noting that one strain of serotype O153:H2 E. coli carries 122 virulence genes, and one strain of serotype O8:H16 E. coli carries 108 virulence genes (Figure 1).
Among the 29 blaNDM-positive E. coli strains, 24 harbored the blaNDM-5 gene. The remaining strains included three with blaNDM-39, one with blaNDM-13, and one with blaNDM-27. Co-occurring β-lactamase genes included blaOXA-10 (19/29, 65.52%), with multiple blaTEM and blaCTX-M variants also present. Moreover, the floR gene was carried by almost all strains (96.55%, 28/29), and the majority of strains also harbored the qnrS1 gene (75.86%, 22/29). It is noteworthy that the coexistence of the colistin resistance gene mcr-1 and blaNDM-5 was found in five strains, and the coexistence of the tigecycline resistance gene tet(X4) and blaNDM-5 was detected in one strain. Furthermore, the resistance gene estT encoding macrolide hydrolase was identified in two strains (Figure 1).
ARGs harbored by E. cloacae, K. pneumoniae, K. aerogenes, and P. mirabilis differed from that harbored by E. coli (Figure S1). Except for P. mirabilis, which harbored blaNDM-1, all other strains carry blaNDM-5. Additionally, blaOXA-10 was detected in two K. aerogenes strains and one P. mirabilis strain. Moreover, blaTEM-176 and blaTEM-1B were identified in two K. pneumoniae strains. Except for two E. cloacae strains, all other strains harbored the floR gene. Furthermore, strains from different genera carried different variants of the fosA gene: E. cloacae carried fosA2, P. mirabilis carried fosA3, K. aerogenes carried fosA5 and fosA7, and K. pneumoniae carried fosA6.
Genetic Environment Analysis of Various blaNDM Gene Variants
Multiple plasmid replicon types were detected in all the blaNDM-positive strains, but we only observed that the blaNDM-5 gene is directly located on the IncX3-type plasmids in 3 E. cloacae and 1 E. coli (Figure 2). The transfer of blaNDM-5 was mediated by the upstream IS5 or ISAba125. In addition, we observed that in two strains of E. coli, the mcr-1 gene was located on a 60kb Incl2-type plasmid and a 105kb IncHI2A-type plasmid, respectively (Figure S2).
Genetic environment analysis revealed the diversity of blaNDM variants-bearing genetic contexts. ISAba125-IS5-blaNDM-5-bleMBL was the most common transposable structure found in E. coli, E. cloacae, and K. aerogenes (Figure 3A). In another K. aerogenes, we discovered the genetic structure of bleMBL-blaNDM-5-IS5-IS1A-aph(3 '')-Ib-aph(6)-Id-aph(3 '')-Ia. This genetic structure may have been formed by the insertion of IS1A-aph(3 '')-Ib-aph(6)-Id-aph(3 '')-Ia mediated by IS1A, which replaced the previous ISAba125. Additionally, the ISAba125-blaNDM-1-bleMBL transposon structure was identified in one P. mirabilis strain.
Correlation Analysis of blaNDM with Other ARGs, ISs and Plasmid Replicons
To thoroughly investigate the genetic background of the blaNDM gene, we collected the CREC samples used in the previous study[23] and downloaded all the genomes of the genera Klebsiella, Enterobacter, and Proteus from the NCBI database. Through sequence alignment, we identified a total of 4,072 blaNDM-positive CREC strains (Table S3), 8,465 blaNDM-positive K. pneumoniae strains (Table S5), 84 blaNDM-positive K. aerogenes strains (Table S6), 139 blaNDM-positive P. mirabilis strains (Table S7), and 105 blaNDM-positive E. cloacae strains (Table S8).
Distinct distributions of blaNDM variants were observed across species (Figure 4A). Upon analysis of the assembled genomes from this study and downloaded genomes, it was observed that 76.88% (3,153/4,101) of blaNDM-positive CREC strains harbored the blaNDM-5 gene, 15.51% (636/4,101) possessed the blaNDM-1 gene, 3.71% (152/4,101) carried the blaNDM-7 gene, and 1.95% (80/4,101) contained the blaNDM-4 gene (Table S9). Notably, two CREC strains were found to concurrently harbor blaNDM-1, blaNDM-4, blaNDM-5, and blaNDM-24. In K. pneumoniae strains, the distribution was as follows: 66.78% (5,653/8,477) carried the blaNDM-1 gene, 27.23% (2,305/8,477) possessed the blaNDM-5 gene, 3.61% (306/8,477) harbored the blaNDM-7 gene, and 1.44% (122/8,477) contained the blaNDM-4 gene (Table S10). For K. aerogenes strains, the proportions are 44.18% (38/86) for the blaNDM-1 gene, 31.40% (27/86) for the blaNDM-5 gene, and 23.26% (20/86) for the blaNDM-7 gene (Table S11). In blaNDM-positive P. mirabilis strains, 70.71% (99/140) carried the blaNDM-1 gene, 22.86% (32/140) possessed the blaNDM-7 gene, and 6.43% (9/140) harbored the blaNDM-5 gene (Table S12). As for blaNDM-positive E. cloacae strains, 75.23% (82/109) carried the blaNDM-1 gene, while 20.18% (22/109) possessed the blaNDM-5 gene (able S13).
Network graph analysis revealed that different blaNDM gene variants in different species exhibit distinct preferences for genetic elements (Figure 4B). When the absolute value of R is greater than 0.3 and p is less than 0.05, we consider that there is a correlation between different genetic elements. In CREC strains, we found that blaNDM-5 was strongly correlated with blaTEM-1B, blaCTX-M-15, and blaOXA-1, while blaNDM-1 was strongly correlated with blaSHV-12 (R > 0.3, p < 0.05). In addition, blaNDM-5 was strongly correlated with ARGs such as sul1, aadA2, mph(A), and insertion sequence IS6100, whereas blaNDM-1 was strongly correlated with rmtC and aph(3')-VI, and blaNDM-7 was strongly correlated with ISCfr27 (R > 0.3, p < 0.05). However, unlike CREC strains, in K. pneumoniae strains, blaNDM-5 only showed positive associations with ARGs such as rmtB, erm(B), oqxA, oqxB, and mph(A), as well as the plasmid replicon IncX3, while blaNDM-1 was strongly correlated with blaCTX-M-15, blaTEM-1B, blaOXA-1, and blaOXA-9 (R > 0.3, p < 0.05). Additionally, blaNDM-1 was also strongly correlated with ARGs such as oqxB, oqxA, and sul1 (R > 0.3, p < 0.05). In K. aerogenes strains, a distinct correlation pattern was observed. Genetic elements such as blaSHV-12, ISSen4, ISCfr4, and ISKpn26 were found to be strongly positively correlated with blaNDM-1 (R > 0.3, p < 0.05), while floR, ISAba125, and IS5 exhibited negative correlations with blaNDM-1 (R< -0.3, p < 0.05). Notably, plasmid replicons including IncN2, IncHI1B, and IncFIB were identified as being strongly positively correlated with blaNDM-1 (R > 0.3, p < 0.05), whereas IncX3 showed a negative correlation with blaNDM-1 (R< -0.3, p < 0.05). However, IncX3 and IS5 were positively correlated with blaNDM-5 (R > 0.3, p < 0.05). Similar correlation patterns were also observed in P. mirabilis strains and E. cloacae strains. In both P. mirabilis strains and E. cloacae strains, blaNDM-1 was strongly negatively correlated with ISAba125, IncX3, and IS5 (R< -0.3, p < 0.05). In P. mirabilis strains, blaNDM-7 was positively correlated with IS5 and ISAba125 (R > 0.3, p < 0.05), meanwhile in E. cloacae strains, blaNDM-5 was positively correlated with IncX3, IS5, and ISAba125 (R > 0.3, p < 0.05). Additionally, in P. mirabilis strains, blaNDM-1 was positively correlated with blaOXA-10, sul1, arr-3, aph(3')-Ia, and Col3M (R > 0.3, p < 0.05), and negatively correlated with IncC and qnrS1 (R< -0.3, p < 0.05). In contrast, blaNDM-7 was positively correlated with qnrS1, IncC, and floR (R > 0.3, p < 0.05), and negatively correlated with Col3M, sul1, arr-3, and aph(3')-Ia (R< -0.3, p < 0.05). In E. cloacae strains, blaNDM-1 was positively correlated with blaCMH-3 (R > 0.3, p < 0.05), and negatively correlated with ISKox3 and floR (R< -0.3, p < 0.05), while blaNDM-5 was positively correlated with ISKox3 and floR (R > 0.3, p < 0.05).
Unexpectedly, blaNDM-1 exhibited negative associations with ISAba125 and IS5 in K. aerogenes, P. mirabilis, and E. cloacae—despite these IS elements being canonical mediators of blaNDM-1 transfer. We further investigated the genomic characteristics of K. aerogenes strains, P. mirabilis strains and E. cloacae strains and found that ISAba125 was often interrupted by various insertion sequences other than IS5 (Figure 3B). In K. aerogenes strains, ISAba125 was interrupted by ISEc33 and ISSpu2. In P. mirabilis strains, ISAba125 was interrupted by IS26, ISKpn26, and ISKpn13. In E. cloacae strains, ISAba125 was interrupted by ISEc33, IS903B, ISSpu2, MITEKpn1, ISKpn14, and ISKpn19. This may suggest that different species capture the heterologous ISAba125-blaNDM-1 transposon via different types of insertion sequences and integrate it into their own genomes to better adapt the blaNDM-1 gene to different genetic environments.
Discussion
Carbapenem-resistant Enterobacteriaceae of animal origin represent a critical group of antimicrobial-resistant pathogens. The increasing number of carbapenem-resistant isolates identified poses a severe threat to global public health security. The blaNDM gene, which encodes NDM, is an important ARG associated with human clinical medicine. It was first identified in a clinical isolate of K. pneumoniae from a hospitalized patient[26]. Although it is only prevalent in Gammaproteobacteria [27], it has had a significant impact on human clinical medicine [28,29,30,31,32], markedly reducing the efficacy of clinical treatments.
Live poultry markets serve as reservoirs and dissemination centers for ARGs [2]. The convergence of live poultry from various regions significantly amplifies the risk of ARG spread. Given the close contact between humans, animals, and the environment in live poultry markets, establishing a “One Health” AMR monitoring system in these settings is crucial for preventing the transmission of multidrug-resistant pathogens and for devising effective containment strategies [33]. In this study, we investigated blaNDM -positive strains in two live poultry markets in Jiangsu Province, China. We found that over 90% of blaNDM-positive strains were E. coli, indicating the widespread presence of CREC strains in poultry. This may be because the blaNDM-bearing plasmids have a high fitness cost in other Enterobacteriaceae bacteria, but there is still a risk of further dissemination. Additionally, conjugation assays revealed that nearly 40% of blaNDM-positive strains harbored transferable blaNDM genes, suggesting that the blaNDM gene can be widely disseminated in live poultry markets.
We obtained assembled genomes of 38 blaNDM-positive strains through whole-genome sequencing. In five E. coli isolates, we detected the coexistence of blaNDM-5 and mcr-1. This once again demonstrates that, despite China’s ban on the use of colistin in animal husbandry, animal sources still harbor stable populations of E. coli that are resistant to both carbapenems and colistin [34]. Notably, in one strain of O8:H16 serotype E. coli isolate coharboring blaNDM-5 and mcr-1, we identified 108 virulence genes, indicating the potential for the spread of highly pathogenic multidrug-resistant bacteria in live poultry markets. Additionally, we identified 122 virulence genes in an O153:H2, ST648-type blaNDM-5-positive E. coli strain. ST648-type E. coli is considered a high-risk, globally epidemic clone that can cause human infections [35]. This finding serves as a warning for the sanitation efforts in live poultry markets.
Genetic environment analysis of assembled genomes from this study revealed that the blaNDM gene was commonly transferred via ISAba125 or IS5. However, surprisingly, through network analysis of downloaded blaNDM-positive strains from the database, we found that in K. aerogenes strains, P. mirabilis strains, and E. cloacae strains, the blaNDM-1 gene was negatively correlated with ISAba125 and IS5, which is contrary to the common situation. Upon further investigation, we discovered that in blaNDM-1-positive K. aerogenes strains, P. mirabilis strains, and E. cloacae strains that lack ISAba125 (actually harboring truncated sequences), different insertion sequences interrupt the ISAba125. Among these insertion sequences, the transfer of the blaNDM-1 gene mediated by ISEc33 [36,37], IS6100 [38], IS903B [39], ISKpn14 [40], ISKpn19 [41], ISKpn26 [39], and MITESen1 [39] has been reported. However, to our knowledge, this study is the first to report the transfer of the blaNDM-1 gene mediated by ISKpn13, ISSpu2, and MITEKpn1. In addition, regarding the fact that ISAba125 is frequently truncated by various types of insertion sequences across different species, we hypothesize that this phenomenon may result from the adaptation of certain insertion sequences to the genomes of these species. This adaptation allows insertion sequences for the capture of the ISAba125-blaNDM-1 transposon and its integration into the genetic environment of the respective strains.
Overall, our findings indicate that the prevalence of carbapenem-resistant strains in live poultry markets is a cause for concern. The potential spread of highly virulent, multidrug-resistant pathogens underscores the importance of comprehensive surveillance efforts. Moreover, the molecular mechanisms by which strains of different species capture the blaNDM-1 gene warrant further investigation. Herein, we call for enhanced sanitation management in live poultry markets, the implementation of appropriate measures to curb the dissemination of blaNDM-positive strains, and safeguarding food safety in animal husbandry through a One Health approach [42].
Funding
This work was supported by the National Key Research and Development Program of China (2023YFD1800500), the Outstanding Youth Foundation of Jiangsu Province of China (BK20231524), the National Natural Science Foundation of China (32373061, 32161133005 and 12411530085), National Key Laboratory of Veterinary Public Health and Safety Open Project Fund (2024SKLVPHS04) and the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Data Availability Statement
WGS data generated from this study are openly available at the China National GeneBank Database (CNGBdb) with accession number of CNP0007032.
Acknowledgements
We acknowledge the genomic data submitters on which this research is based. This work was supported by the High-Performance Computing Cluster of College of Veterinary Medicine, Yangzhou University.
Conflicts of Interest
The authors declare no competing interests.
References
- ALAM M U, RAHMAN M, ABDULLAH AL M, et al. Human exposure to antimicrobial resistance from poultry production: Assessing hygiene and waste-disposal practices in Bangladesh [J]. International journal of hygiene and environmental health, 2019, 222(8): 1068-76. [CrossRef]
- WANG Y, HU Y, CAO J, et al. Antibiotic resistance gene reservoir in live poultry markets [J]. J Infect, 2019, 78(6): 445-53. [CrossRef]
- WANG Y, LYU N, LIU F, et al. More diversified antibiotic resistance genes in chickens and workers of the live poultry markets [J]. Environ Int, 2021, 153: 106534. [CrossRef]
- GAO X L, SHAO M F, LUO Y, et al. Airborne bacterial contaminations in typical Chinese wet market with live poultry trade [J]. Sci Total Environ, 2016, 572: 681-7. [CrossRef]
- TOO R J, KARIUKI S M, GITAO G C, et al. Carbapenemase-producing bacteria recovered from Nairobi River, Kenya surface water and from nearby anthropogenic and zoonotic sources [J]. PLoS One, 2024, 19(11): e0310026. [CrossRef]
- SU Y, XIN L, ZHANG F, et al. Drug resistance analysis of three types of avian-origin carbapenem-resistant Enterobacteriaceae in Shandong Province, China [J]. Poult Sci, 2023, 102(3): 102483. [CrossRef]
- YANG H, XIONG Z, CAO K, et al. Risk factors and molecular epidemiology of colonizing carbapenem-resistant Enterobacterales in pediatric inpatient in Shenzhen, China [J]. Journal of infection and public health, 2025, 18(1): 102614. [CrossRef]
- AL-MUSTAPHA A I, TIWARI A, LAUKKANEN-NINIOS R, et al. Wastewater based genomic surveillance key to population level monitoring of AmpC/ESBL producing Escherichia coli [J]. Sci Rep, 2025, 15(1): 7400. [CrossRef]
- LI C A, GUO C H, YANG T Y, et al. Whole-Genome Analysis of bla(NDM)-Bearing Proteus mirabilis Isolates and mcr-1-Positive Escherichia coli Isolates Carrying bla(NDM) from the Same Fresh Vegetables in China [J]. Foods (Basel, Switzerland), 2023, 12(3). [CrossRef]
- HUMPHRIES R, BOBENCHIK A M, HINDLER J A, et al. Overview of Changes to the Clinical and Laboratory Standards Institute Performance Standards for Antimicrobial Susceptibility Testing, M100, 31st Edition [J]. J Clin Microbiol, 2021, 59(12): e0021321. [CrossRef]
- CHEN Y, CHEN Y, SHI C, et al. SOAPnuke: a MapReduce acceleration-supported software for integrated quality control and preprocessing of high-throughput sequencing data [J]. Gigascience, 2018, 7(1): 1-6. [CrossRef]
- BANKEVICH A, NURK S, ANTIPOV D, et al. SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing [J]. Journal of computational biology : a journal of computational molecular cell biology, 2012, 19(5): 455-77. [CrossRef]
- ZANKARI E, HASMAN H, COSENTINO S, et al. Identification of acquired antimicrobial resistance genes [J]. J Antimicrob Chemother, 2012, 67(11): 2640-4. [CrossRef]
- SIGUIER P, PEROCHON J, LESTRADE L, et al. ISfinder: the reference centre for bacterial insertion sequences [J]. Nucleic Acids Res, 2006, 34(Database issue): D32-6. [CrossRef]
- CARATTOLI A, ZANKARI E, GARCíA-FERNáNDEZ A, et al. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing [J]. Antimicrob Agents Chemother, 2014, 58(7): 3895-903. [CrossRef]
- CHEN L, YANG J, YU J, et al. VFDB: a reference database for bacterial virulence factors [J]. Nucleic Acids Res, 2005, 33(Database issue): D325-8. [CrossRef]
- LIU M, LI X, XIE Y, et al. ICEberg 2.0: an updated database of bacterial integrative and conjugative elements [J]. Nucleic Acids Res, 2019, 47(D1): D660-d5. [CrossRef]
- BESSONOV K, LAING C, ROBERTSON J, et al. ECTyper: in silico Escherichia coli serotype and species prediction from raw and assembled whole-genome sequence data [J]. Microb Genom, 2021, 7(12). [CrossRef]
- SEEMANN T. Prokka: rapid prokaryotic genome annotation [J]. Bioinformatics (Oxford, England), 2014, 30(14): 2068-9. [CrossRef]
- PAGE A J, CUMMINS C A, HUNT M, et al. Roary: rapid large-scale prokaryote pan genome analysis [J]. Bioinformatics (Oxford, England), 2015, 31(22): 3691-3. [CrossRef]
- PRICE M N, DEHAL P S, ARKIN A P. FastTree: computing large minimum evolution trees with profiles instead of a distance matrix [J]. Molecular biology and evolution, 2009, 26(7): 1641-50. [CrossRef]
- ALIKHAN N F, PETTY N K, BEN ZAKOUR N L, et al. BLAST Ring Image Generator (BRIG): simple prokaryote genome comparisons [J]. BMC genomics, 2011, 12: 402. [CrossRef]
- LI Y, SUN X, DONG N, et al. Global distribution and genomic characteristics of carbapenemase-producing Escherichia coli among humans, 2005-2023 [J]. Drug resistance updates : reviews and commentaries in antimicrobial and anticancer chemotherapy, 2024, 72: 101031. [CrossRef]
- CHKLOVSKI A, PARKS D H, WOODCROFT B J, et al. CheckM2: a rapid, scalable and accurate tool for assessing microbial genome quality using machine learning [J]. Nature methods, 2023, 20(8): 1203-12. [CrossRef]
- BASTIAN M, HEYMANN S, JACOMY M. Gephi: An Open Source Software for Exploring and Manipulating Networks [M]. 2009.
- YONG D, TOLEMAN M A, GISKE C G, et al. Characterization of a new metallo-beta-lactamase gene, bla(NDM-1), and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India [J]. Antimicrob Agents Chemother, 2009, 53(12): 5046-54. [CrossRef]
- DIEBOLD P J, RHEE M W, SHI Q, et al. Clinically relevant antibiotic resistance genes are linked to a limited set of taxa within gut microbiome worldwide [J]. Nat Commun, 2023, 14(1): 7366. [CrossRef]
- YAO J, HU Y, WANG X, et al. Carbapenem-resistant Morganella morganii carrying bla(KPC-2) or bla(NDM-1) in the clinic: one-decade genomic epidemiology analysis [J]. Microbiol Spectr, 2025: e0247624. [CrossRef]
- KE Y, ZHU Z, LU W, et al. Emerging bla(NDM)-positive Salmonella enterica in Chinese pediatric infections [J]. Microbiol Spectr, 2024, 12(12): e0148524. [CrossRef]
- ZHAO Q, GUO L, YE K, et al. Epidemiology, Phylogeny and Genetic Characterization of Carbapenem-resistant Citrobacter spp. from 5 hospitals in China [J]. J Glob Antimicrob Resist, 2025. [CrossRef]
- MARANO R B M, OSTER Y, BENENSON S, et al. An Omics-Guided Investigation of a Hospital Outbreak Caused by blaNDM-1-Producing Pseudocitrobacter faecalis [J]. The Journal of infectious diseases, 2025. [CrossRef]
- LIAN S, LIU C, CAI M, et al. Risk factors and molecular characterization of carbapenem resistant Escherichia coli recovered from a tertiary hospital in Fujian, China from 2021 to 2023 [J]. BMC Microbiol, 2024, 24(1): 374. [CrossRef]
- MORRIS R, WANG S. Building a pathway to One Health surveillance and response in Asian countries [J]. Science in One Health, 2024, 3: 100067. [CrossRef]
- GUAN Y, WANG Z, SHANG Z, et al. Steady existence of Escherichia coli co-resistant to carbapenem and colistin in an animal breeding area even after the colistin forbidden [J]. J Environ Manage, 2024, 371: 123084. [CrossRef]
- SHAFIQ M, ZENG M, PERMANA B, et al. Coexistence of bla (NDM-5) and tet(X4) in international high-risk Escherichia coli clone ST648 of human origin in China [J]. Front Microbiol, 2022, 13: 1031688. [CrossRef]
- WANG Y, WU C, ZHANG Q, et al. Identification of New Delhi metallo-β-lactamase 1 in Acinetobacter lwoffii of food animal origin [J]. PLoS One, 2012, 7(5): e37152. [CrossRef]
- MEDUGU N, TICKLER I A, DURU C, et al. Phenotypic and molecular characterization of beta-lactam resistant Multidrug-resistant Enterobacterales isolated from patients attending six hospitals in Northern Nigeria [J]. Sci Rep, 2023, 13(1): 10306. [CrossRef]
- ZHANG F, LI Z, LIU X, et al. Carbapenem-resistant Citrobacter freundii harboring bla(KPC-2) and bla(NDM-1): a study on their transferability and potential dissemination via generating a transferrable hybrid plasmid mediated by IS6100 [J]. Front Microbiol, 2023, 14: 1239538. [CrossRef]
- PAJAND O, RAHIMI H, BADMASTI F, et al. Various arrangements of mobile genetic elements among CC147 subpopulations of Klebsiella pneumoniae harboring bla(NDM-1): a comparative genomic analysis of carbapenem resistant strains [J]. Journal of biomedical science, 2023, 30(1): 73. [CrossRef]
- OYELADE A A, IKHIMIUKOR O O, NWADIKE B I, et al. Assessing the risk of exposure to antimicrobial resistance at public beaches: Genome-based insights into the resistomes, mobilomes and virulomes of beta-lactams resistant Enterobacteriaceae from recreational beaches in Lagos, Nigeria [J]. International journal of hygiene and environmental health, 2024, 258: 114347. [CrossRef]
- ZI P, FANG M, YANG H, et al. Characterization of an NDM-1-Producing Citrobacter koseri Isolate from China [J]. Infect Drug Resist, 2024, 17: 61-7. [CrossRef]
- QIAN J, WU Z, ZHU Y, et al. One Health: a holistic approach for food safety in livestock [J]. Sci One Health, 2022, 1: 100015. [CrossRef]
Figure 1.
The phylogenetic tree and ARG heatmap of 29 E. coli isolates. The phylogenetic tree was generated by FastTree based on core genes alignment using Roary and was visualized using Chiplot. Isolates from different sources are highlighted in different colors. The three columns of information marked next to the strain names are isolation source, ST type (identified by MLST), and Serotype (identified by ECTyper). The three sets of heatmaps show the presence of ARGs in the strains. The outermost bar chart shows the number of virulence genes in the isolates based on the VFDB core dataset.
Figure 1.
The phylogenetic tree and ARG heatmap of 29 E. coli isolates. The phylogenetic tree was generated by FastTree based on core genes alignment using Roary and was visualized using Chiplot. Isolates from different sources are highlighted in different colors. The three columns of information marked next to the strain names are isolation source, ST type (identified by MLST), and Serotype (identified by ECTyper). The three sets of heatmaps show the presence of ARGs in the strains. The outermost bar chart shows the number of virulence genes in the isolates based on the VFDB core dataset.
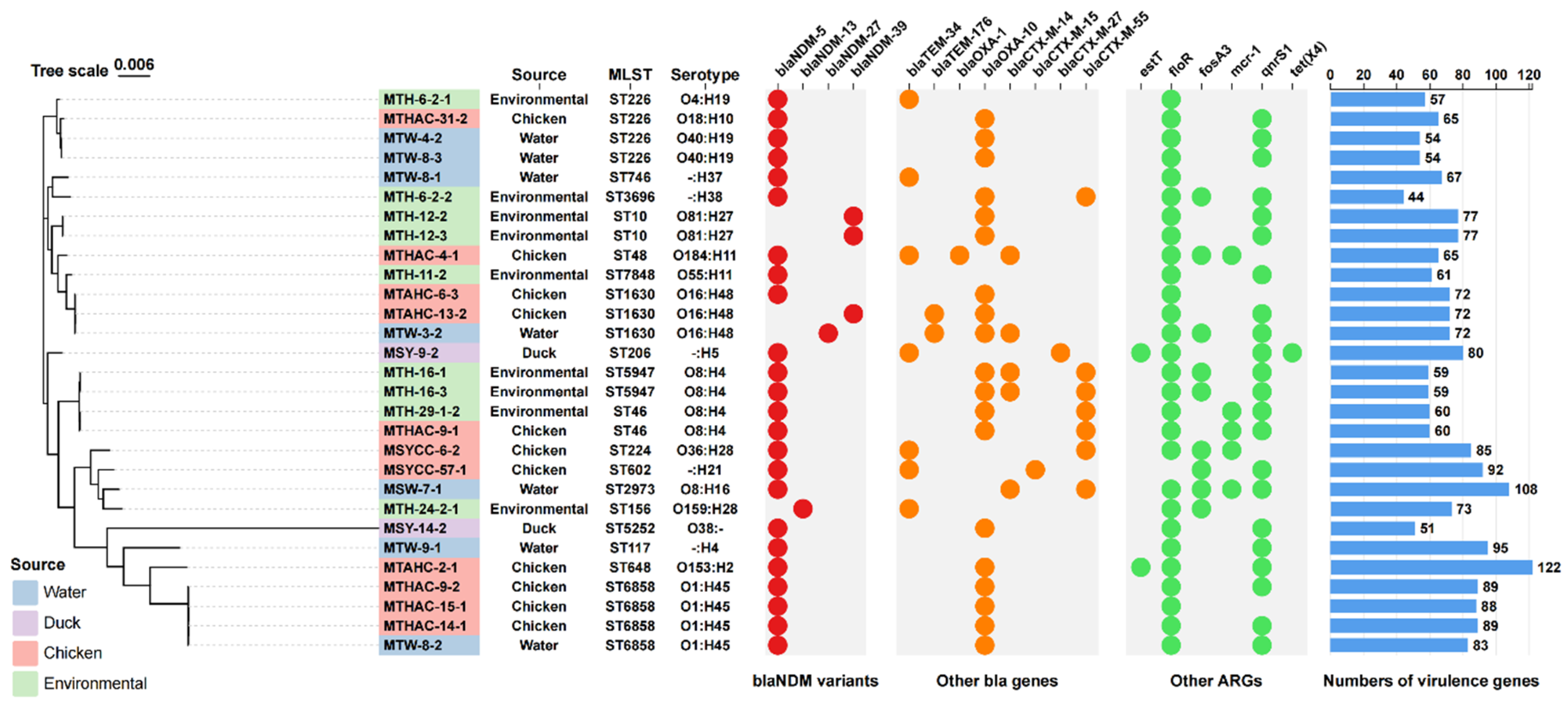
Figure 2.
Plasmid profile of the blaNDM-5-containing IncX3-type plasmid. Plasmid slices (assembled contigs, not complete plasmids) from this study were compared with a plasmid (CP033399.1) derived from E. coli. The GC skew and GC content are depicted in an inward-to-outward sequence. The outermost arrows indicate the positions and transcriptional orientations of the open reading frames.
Figure 2.
Plasmid profile of the blaNDM-5-containing IncX3-type plasmid. Plasmid slices (assembled contigs, not complete plasmids) from this study were compared with a plasmid (CP033399.1) derived from E. coli. The GC skew and GC content are depicted in an inward-to-outward sequence. The outermost arrows indicate the positions and transcriptional orientations of the open reading frames.
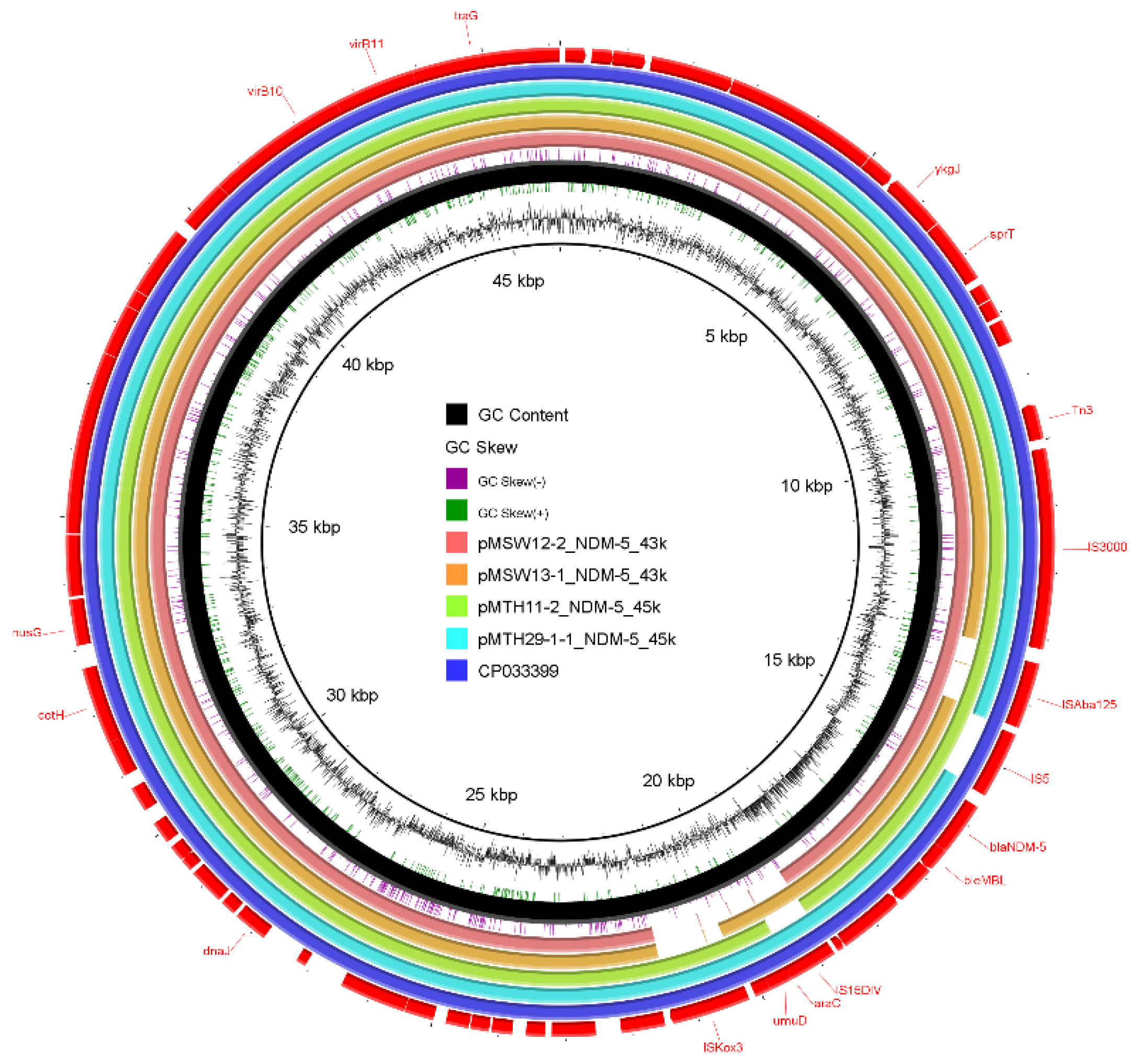
Figure 3.
Genetic environments of several blaNDM variants in assembled genomes and genomes from database. (A) Different types of genetic environments of blaNDM variants in assembled genomes. Different genetic elements are highlighted in different colors. (B) Primary genetic contexts of blaNDM-1 across different species. In different species, the transfer of blaNDM-1 is mediated by various IS elements.
Figure 3.
Genetic environments of several blaNDM variants in assembled genomes and genomes from database. (A) Different types of genetic environments of blaNDM variants in assembled genomes. Different genetic elements are highlighted in different colors. (B) Primary genetic contexts of blaNDM-1 across different species. In different species, the transfer of blaNDM-1 is mediated by various IS elements.

Figure 4.
The proportion of blaNDM variants across different species and the network graph depicting the coexistence patterns of different blaNDM gene variants with other ARGs, ISs and plasmid replicons harbored in different bacteria. (A) The bar chart shows the percentage of blaNDM variants in blaNDM-positive strains of different species. (B-E) The network graph illustrates the correlations between blaNDM variants and other genetic elements in blaNDM -positive strains of different species. The nodes represent ARGs, ISs and plasmid replicons identified in all blaNDM-positive strains of from different species. The connections between nodes signify their interrelatedness. Blue hues and increased line thickness denote stronger positive correlations. The intensity of the yellow color on the lines indicates the strength of negative correlations, with darker shades of yellow corresponding to stronger negative correlations. Additionally, the thickness of the lines is directly proportional to the correlation strength, where a thicker line signifies a more pronounced relationship between the variables. All associated genes depicted in the figure exhibited p values less than 0.05.
Figure 4.
The proportion of blaNDM variants across different species and the network graph depicting the coexistence patterns of different blaNDM gene variants with other ARGs, ISs and plasmid replicons harbored in different bacteria. (A) The bar chart shows the percentage of blaNDM variants in blaNDM-positive strains of different species. (B-E) The network graph illustrates the correlations between blaNDM variants and other genetic elements in blaNDM -positive strains of different species. The nodes represent ARGs, ISs and plasmid replicons identified in all blaNDM-positive strains of from different species. The connections between nodes signify their interrelatedness. Blue hues and increased line thickness denote stronger positive correlations. The intensity of the yellow color on the lines indicates the strength of negative correlations, with darker shades of yellow corresponding to stronger negative correlations. Additionally, the thickness of the lines is directly proportional to the correlation strength, where a thicker line signifies a more pronounced relationship between the variables. All associated genes depicted in the figure exhibited p values less than 0.05.
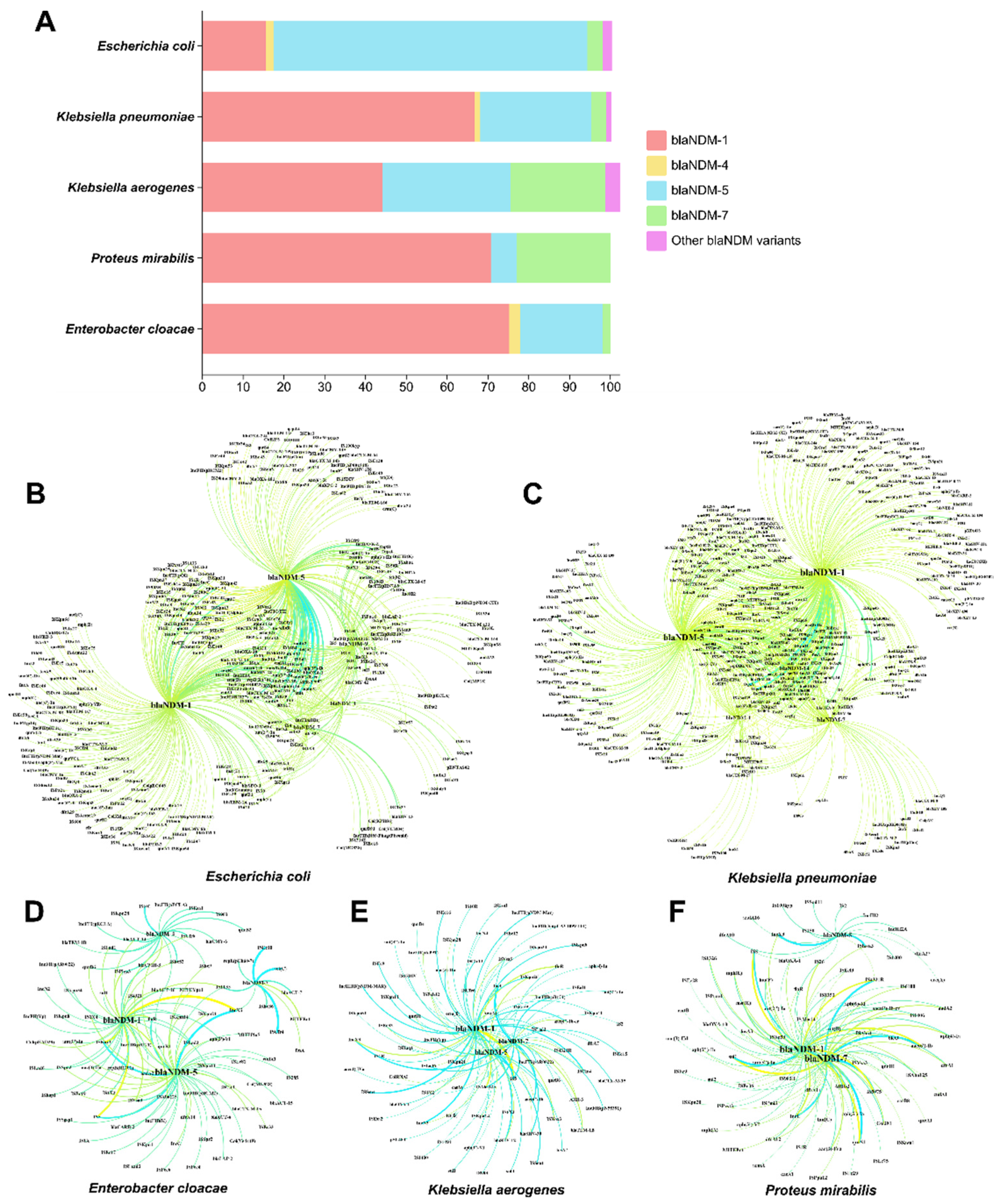

Table 1.
Antimicrobial susceptibility profiles of 233 blaNDM-positive strains. MEM: Meropenem; IMP: Imipenem; AMP: Ampicillin; CAZ: Ceftazidime; KAN: Kanamycin; GEN: Gentamicin; CIP: Ciprofloxacin; CL: Colistin.
Table 1.
Antimicrobial susceptibility profiles of 233 blaNDM-positive strains. MEM: Meropenem; IMP: Imipenem; AMP: Ampicillin; CAZ: Ceftazidime; KAN: Kanamycin; GEN: Gentamicin; CIP: Ciprofloxacin; CL: Colistin.
| Strains | Source | Species | Conjugation recipient | MIC | |||||||
| MEM | IMP | AMP | CAZ | KAN | GEN | CIP | CL | ||||
| MTHAC-1-1 | Chicken | Escherichia coli | - | >128 | 8 | >128 | >128 | >128 | >128 | ≤0.25 | ≤0.25 |
| MTHAC-1-2 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 8 | 1 |
| MTHAC-2-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | 2 | 0.5 | 0.5 |
| MTHAC-2-2 | Chicken | Escherichia coli | - | 128 | >128 | >128 | >128 | >128 | 4 | 1 | ≤0.25 |
| MTHAC-3-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | 128 | 0.5 | ≤0.25 |
| MTHAC-3-3 | Chicken | Escherichia coli | - | 128 | >128 | >128 | >128 | >128 | >128 | 16 | ≤0.25 |
| MTHAC-4-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 64 | 8 |
| MTHAC-4-2 | Chicken | Escherichia coli | - | 128 | >128 | >128 | >128 | >128 | >128 | 2 | ≤0.25 |
| MTHAC-5-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 0.5 | ≤0.25 |
| MTHAC-6-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | >128 | ≤0.25 |
| MTHAC-6-2 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | 128 | 1 | 1 |
| MTHAC-8-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | >128 | ≤0.25 |
| MTHAC-8-2 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | 32 | 16 | 0.5 | 1 |
| MTHAC-9-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 1 | 8 |
| MTHAC-9-2 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 0.5 | 1 |
| MTHAC-10-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | >128 | 1 |
| MTHAC-10-2 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 1 | 0.5 |
| MTHAC-10-3 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 1 | ≤0.25 |
| MTHAC-11-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 4 | 1 |
| MTHAC-11-2 | Chicken | Escherichia coli | - | 64 | >128 | >128 | >128 | >128 | >128 | ≤0.25 | ≤0.25 |
| MTHAC-12-1 | Chicken | Escherichia coli | - | 64 | >128 | >128 | >128 | >128 | >128 | 8 | ≤0.25 |
| MTHAC-12-2 | Chicken | Escherichia coli | - | 128 | >128 | >128 | >128 | >128 | >128 | 2 | 0.5 |
| MTHAC-13-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 16 | 2 |
| MTHAC-13-2 | Chicken | Escherichia coli | - | 128 | >128 | >128 | >128 | >128 | >128 | 1 | ≤0.25 |
| MTHAC-14-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | >128 | 0.5 |
| MTHAC-15-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | >128 | 0.5 |
| MTHAC-15-2 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 2 | 0.5 |
| MTHAC-16-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 64 | 1 |
| MTHAC-16-2 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 16 | 0.5 |
| MTHAC-17-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 8 | 2 |
| MTHAC-17-2 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 8 | 0.5 |
| MTHAC-17-3 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 8 | 1 |
| MTHAC-18-1 | Chicken | Escherichia coli | - | 64 | 128 | >128 | >128 | >128 | 128 | 8 | ≤0.25 |
| MTHAC-18-2 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 16 | ≤0.25 |
| MTHAC-24-1 | Chicken | Escherichia coli | - | 64 | 128 | >128 | >128 | >128 | 64 | 0.5 | ≤0.25 |
| MTHAC-25-4 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | 128 | 32 | 4 |
| MTHAC-27-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 8 | 2 |
| MTHAC-27-2 | Chicken | Escherichia coli | - | >128 | 4 | >128 | >128 | >128 | 128 | 8 | ≤0.25 |
| MTHAC-28-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 64 | 2 |
| MTHAC-31-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 128 | 16 |
| MTHAC-31-2 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 128 | 16 |
| MTAHC-1-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | 128 | 2 | 4 |
| MTAHC-1-2 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | 8 | 128 | 1 |
| MTAHC-2-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 4 | 8 |
| MTAHC-3-2 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 4 | 1 |
| MTAHC-4-1 | Chicken | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | 128 | >128 | 2 |
| MTAHC-4-3 | Chicken | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | >128 | 64 | 4 |
| MTAHC-4-4 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | >128 | 2 |
| MTAHC-5-1 | Chicken | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | >128 | 32 | 1 |
| MTAHC-5-2 | Chicken | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | >128 | 64 | 2 |
| MTAHC-6-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 128 | 1 |
| MTAHC-6-2 | Chicken | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | 2 | 64 | ≤0.25 |
| MTAHC-6-3 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | >128 | 4 |
| MTAHC-8-1 | Chicken | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | >128 | 128 | 4 |
| MTAHC-12-1 | Chicken | Escherichia coli | - | 64 | >128 | >128 | >128 | >128 | >128 | 64 | 2 |
| MTAHC-12-2 | Chicken | Escherichia coli | - | 128 | >128 | >128 | >128 | >128 | >128 | 64 | 0.5 |
| MTAHC-13-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 128 | 0.5 |
| MTAHC-13-2 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | >128 | 4 |
| MTAHC-13-3 | Chicken | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | >128 | >128 | 4 |
| MTAHC-14-3-1 | Chicken | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | >128 | 128 | - |
| MTAHC-14-3-2 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | >128 | - |
| MTAHC-15-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 64 | - |
| MTAHC-15-2 | Chicken | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | >128 | 8 | - |
| MTAHC-16-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 32 | - |
| MTAHC-16-2 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | >128 | - |
| MTAHC-16-3 | Chicken | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | >128 | 64 | - |
| MTH-1-3-2 | Environmental | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | ≤0.25 | ≤0.25 |
| MTH-3-1 | Environmental | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 16 | ≤0.25 |
| MTH-4-1 | Environmental | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | 64 | 64 | ≤0.25 |
| MTH-4-2 | Environmental | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | 64 | 64 | ≤0.25 |
| MTH-6-1-1 | Environmental | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | >128 | 32 | ≤0.25 |
| MTH-6-1-2 | Environmental | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | >128 | 32 | ≤0.25 |
| MTH-6-2-1 | Environmental | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | >128 | 32 | >128 |
| MTH-6-2-2 | Environmental | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | 32 | 2 | >128 |
| MTH-6-3-1 | Environmental | Klebsiella aerogenes | - | >128 | >128 | >128 | >128 | >128 | >128 | 0.5 | >128 |
| MTH-6-3-2 | Environmental | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 32 | ≤0.25 |
| MTH-7-2-1 | Environmental | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 16 | ≤0.25 |
| MTH-7-2-2 | Environmental | Klebsiella aerogenes | - | >128 | >128 | >128 | >128 | >128 | >128 | 32 | 128 |
| MTH-10-1 | Environmental | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | 128 | ≤0.25 | ≤0.25 |
| MTH-11-1 | Environmental | Escherichia coli | C600 | >128 | >128 | >128 | >128 | 8 | 4 | 4 | 4 |
| MTH-11-2 | Environmental | Escherichia coli | C600 | >128 | >128 | >128 | >128 | 8 | 1 | 8 | 8 |
| MTH-12-1 | Environmental | Escherichia coli | - | >128 | >128 | >128 | >128 | 8 | 2 | 1 | 0.5 |
| MTH-12-2 | Environmental | Escherichia coli | - | >128 | >128 | >128 | >128 | 16 | 2 | 1 | >128 |
| MTH-12-3 | Environmental | Escherichia coli | - | >128 | >128 | >128 | >128 | 8 | 2 | 1 | >128 |
| MTH-13-2 | Environmental | Klebsiella pneumoniae | - | 128 | >128 | >128 | >128 | >128 | >128 | >128 | >128 |
| MTH-16-1 | Environmental | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 16 | >128 |
| MTH-16-2 | Environmental | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 16 | 64 |
| MTH-16-3 | Environmental | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 32 | >128 |
| MTH-19-2 | Environmental | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | >128 | 64 |
| MTH-24-2-1 | Environmental | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | >128 | >128 |
| MTH-26-1 | Environmental | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | 4 | 2 | ≤0.25 |
| MTH-29-1-1 | Environmental | Enterobacter cloacae | C600 | >128 | >128 | >128 | >128 | 4 | 1 | ≤0.25 | >128 |
| MTH-29-1-2 | Environmental | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | 128 | 1 | >128 |
| MTH-30-1-2 | Environmental | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 64 | ≤0.25 |
| MTW-1-1 | Water | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | >128 | ≤0.25 |
| MTW-1-2 | Water | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | >128 | 8 | ≤0.25 |
| MTW-2-1 | Water | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | >128 | 32 | ≤0.25 |
| MTW-3-1 | Water | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | 128 | 128 | ≤0.25 |
| MTW-3-2 | Water | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | >128 | >128 |
| MTW-4-1 | Water | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 4 | >128 |
| MTW-4-2 | Water | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 4 | >128 |
| MTW-7-1 | Water | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | >128 | 128 | >128 |
| MTW-8-1 | Water | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 2 | >128 |
| MTW-8-2 | Water | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 0.5 | >128 |
| MTW-8-3 | Water | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | 64 | 4 | >128 |
| MTW-9-1 | Water | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | 128 | 0.5 | >128 |
| MTW-18-1 | Water | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | 128 | 64 | ≤0.25 |
| MTW-18-2 | Water | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | 64 | 64 | 4 |
| MTW-18-3 | Water | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | 128 | ≤0.25 | ≤0.25 |
| MTAHC18-1 | Chicken | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | 64 | 64 | 2 |
| MTAHC18-2 | Chicken | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | 128 | 64 | ≤0.25 |
| MTNJC-3-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 64 | ≤0.25 |
| MTNJC-3-2 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | 128 | 0.5 | ≤0.25 |
| MTNTC-2-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 1 | ≤0.25 |
| MTNTC-5-1 | Chicken | Klebsiella pneumoniae | - | >128 | >128 | >128 | >128 | >128 | 32 | 1 | ≤0.25 |
| MTNTC-5-2 | Chicken | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | >128 | 1 | ≤0.25 |
| MTNTC-6-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | 128 | 0.5 | ≤0.25 |
| MTTZC-2-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | 64 | 64 | ≤0.25 |
| MTTZC-2-2 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 64 | ≤0.25 |
| MTTZC-3-1 | Chicken | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | >128 | ≤0.25 | ≤0.25 |
| MTAHG-11-1 | Pigeon | Providencia rettgeri | - | 32 | >128 | >128 | >128 | 16 | 32 | 8 | >128 |
| MTAHY-2-1 | Duck | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 32 | ≤0.25 |
| MTAHY-4-1 | Duck | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 8 | ≤0.25 |
| MTAHY-4-2 | Duck | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | 128 | ≤0.25 | ≤0.25 |
| MTAHY-12-1 | Duck | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 8 | 8 |
| MTAHY-12-2 | Duck | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | 4 | ≤0.25 | ≤0.25 |
| MTAHY-13-1 | Duck | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | 32 | 1 | >128 |
| MTAHY-13-2 | Duck | Escherichia coli | - | >128 | >128 | >128 | >128 | 16 | 4 | ≤0.25 | ≤0.25 |
| MTYZG-24-3 | Pigeon | Klebsiella pneumoniae | - | ≤0.25 | ≤0.25 | ≤0.25 | ≤0.25 | ≤0.25 | ≤0.25 | ≤0.25 | ≤0.25 |
| MTYZG-33-1 | Pigeon | Klebsiella pneumoniae | - | 32 | 128 | >128 | 16 | 1 | ≤0.25 | ≤0.25 | ≤0.25 |
| MSYCC-4-2 | Chicken | Escherichia coli | C600 | 2 | 128 | >128 | >128 | >128 | 128 | ≤0.25 | ≤0.25 |
| MSYCC-5-1 | Chicken | Escherichia coli | C600 | 64 | >128 | >128 | >128 | 8 | 1 | ≤0.25 | ≤0.25 |
| MSYCC-6-1 | Chicken | Escherichia coli | C600 | 128 | >128 | >128 | >128 | 16 | 4 | 32 | ≤0.25 |
| MSYCC-6-2 | Chicken | Escherichia coli | - | 32 | 128 | >128 | >128 | >128 | >128 | 8 | 4 |
| MSYCC-11-1 | Chicken | Escherichia coli | - | 8 | >128 | >128 | >128 | >128 | >128 | 128 | ≤0.25 |
| MSYCC-11-2 | Chicken | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | >128 | 2 | ≤0.25 |
| MSYCC-13-1 | Chicken | Escherichia coli | C600 | 64 | >128 | >128 | >128 | >128 | 128 | ≤0.25 | ≤0.25 |
| MSYCC-13-2 | Chicken | Escherichia coli | C600 | 2 | >128 | >128 | >128 | >128 | 32 | ≤0.25 | ≤0.25 |
| MSYCC-19-1 | Chicken | Escherichia coli | C600 | 128 | >128 | >128 | >128 | >128 | >128 | 1 | ≤0.25 |
| MSYCC-20-1 | Chicken | Escherichia coli | - | 128 | >128 | >128 | >128 | >128 | 128 | ≤0.25 | ≤0.25 |
| MSYCC-20-2 | Chicken | Escherichia coli | - | 128 | >128 | >128 | >128 | >128 | >128 | ≤0.25 | ≤0.25 |
| MSYCC-21-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 0.5 | ≤0.25 |
| MSYCC-21-2 | Chicken | Escherichia coli | C600 | 32 | >128 | >128 | >128 | >128 | >128 | 0.5 | ≤0.25 |
| MSYCC-24-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | 2 | ≤0.25 |
| MSYCC-25-1 | Chicken | Escherichia coli | C600 | 32 | >128 | >128 | >128 | >128 | >128 | 64 | ≤0.25 |
| MSYCC-25-2 | Chicken | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | >128 | 16 | 0.5 |
| MSYCC-28-2 | Chicken | Escherichia coli | - | 128 | >128 | >128 | >128 | >128 | >128 | 2 | ≤0.25 |
| MSYCC-31-1 | Chicken | Escherichia coli | - | 32 | 128 | >128 | >128 | >128 | 128 | 32 | ≤0.25 |
| MSYCC-32-1 | Chicken | Escherichia coli | C600 | 16 | >128 | >128 | >128 | >128 | 32 | ≤0.25 | 1 |
| MSYCC-34-1 | Chicken | Escherichia coli | - | 16 | >128 | >128 | >128 | >128 | >128 | ≤0.25 | 2 |
| MSYCC-34-2 | Chicken | Escherichia coli | C600 | 4 | 32 | >128 | >128 | >128 | 64 | 8 | ≤0.25 |
| MSYCC-35-1 | Chicken | Escherichia coli | C600 | 32 | >128 | >128 | >128 | >128 | >128 | 0.5 | 0.5 |
| MSYCC-37-1 | Chicken | Escherichia coli | C600 | 64 | >128 | >128 | >128 | >128 | 64 | 32 | ≤0.25 |
| MSYCC-40-1 | Chicken | Escherichia coli | C600 | 128 | >128 | >128 | >128 | >128 | >128 | 32 | ≤0.25 |
| MSYCC-42-1 | Chicken | Escherichia coli | C600 | 16 | 16 | >128 | >128 | >128 | 1 | ≤0.25 | ≤0.25 |
| MSYCC-43-1 | Chicken | Escherichia coli | C600 | 128 | 128 | >128 | >128 | >128 | >128 | 32 | ≤0.25 |
| MSYCC-45-1 | Chicken | Escherichia coli | C600 | 64 | >128 | >128 | >128 | >128 | 32 | 32 | ≤0.25 |
| MSYCC-45-2 | Chicken | Escherichia coli | C600 | 64 | >128 | >128 | >128 | >128 | 64 | ≤0.25 | ≤0.25 |
| MSYCC-46-1 | Chicken | Escherichia coli | - | 64 | >128 | >128 | >128 | >128 | >128 | 32 | ≤0.25 |
| MSYCC-47-1 | Chicken | Escherichia coli | C600 | 64 | >128 | >128 | >128 | >128 | >128 | 2 | ≤0.25 |
| MSYCC-48-1 | Chicken | Escherichia coli | - | 64 | >128 | >128 | >128 | >128 | 2 | 2 | ≤0.25 |
| MSYCC-49-1 | Chicken | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | 128 | 16 | ≤0.25 |
| MSYCC-51-1 | Chicken | Escherichia coli | - | 128 | >128 | >128 | >128 | >128 | >128 | 2 | ≤0.25 |
| MSYCC-51-2 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | >128 | >128 | ≤0.25 |
| MSYCC-51-3 | Chicken | Escherichia coli | C600 | 32 | >128 | >128 | >128 | >128 | >128 | 64 | ≤0.25 |
| MSYCC-52-1 | Chicken | Escherichia coli | C600 | 64 | >128 | >128 | >128 | >128 | 128 | 32 | 0.5 |
| MSYCC-52-2 | Chicken | Escherichia coli | C600 | 128 | >128 | >128 | >128 | >128 | >128 | 16 | 1 |
| MSYCC-54-1 | Chicken | Escherichia coli | C600 | 32 | >128 | >128 | >128 | >128 | 2 | 0.5 | ≤0.25 |
| MSYCC-54-2 | Chicken | Proteus mirabilis | C600 | >128 | >128 | >128 | >128 | >128 | >128 | 16 | >128 |
| MSYCC-55-1 | Chicken | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | >128 | 64 | 1 |
| MSYCC-57-1 | Chicken | Escherichia coli | - | >128 | >128 | >128 | >128 | >128 | 64 | ≤0.25 | 8 |
| MSNTC-3-1 | Chicken | Escherichia coli | C600 | 64 | 128 | >128 | >128 | 4 | 2 | 16 | ≤0.25 |
| MSNTC-4-1 | Chicken | Escherichia coli | C600 | 8 | 64 | >128 | >128 | >128 | 128 | 8 | ≤0.25 |
| MSNTC-7-1 | Chicken | Escherichia coli | C600 | 64 | 64 | >128 | >128 | 4 | 1 | >128 | ≤0.25 |
| MSNTC-9-1 | Chicken | Escherichia coli | C600 | 32 | 64 | >128 | >128 | >128 | 128 | >128 | ≤0.25 |
| MSNTC-10-1 | Chicken | Escherichia coli | - | 32 | 64 | >128 | >128 | >128 | >128 | ≤0.25 | ≤0.25 |
| MSNTC-10-2 | Chicken | Escherichia coli | C600 | 64 | 128 | >128 | >128 | >128 | >128 | ≤0.25 | ≤0.25 |
| MSNTC-12-1 | Chicken | Escherichia coli | C600 | 64 | 128 | >128 | >128 | >128 | 128 | 64 | ≤0.25 |
| MSNTC-12-2 | Chicken | Escherichia coli | - | 64 | >128 | >128 | >128 | >128 | >128 | >128 | ≤0.25 |
| MSNTC-13-1 | Chicken | Escherichia coli | C600 | 2 | >128 | >128 | >128 | 4 | 2 | 32 | ≤0.25 |
| MSNTC-13-2 | Chicken | Escherichia coli | C600 | 32 | 64 | >128 | >128 | 4 | 2 | 16 | ≤0.25 |
| MSNTC-14-1 | Chicken | Escherichia coli | C600 | 16 | 64 | >128 | >128 | >128 | 1 | 8 | ≤0.25 |
| MSNTC-14-2 | Chicken | Escherichia coli | C600 | 32 | 64 | >128 | >128 | >128 | 2 | 8 | ≤0.25 |
| MSNTC-17-1 | Chicken | Escherichia coli | C600 | 32 | 64 | >128 | >128 | >128 | 1 | ≤0.25 | ≤0.25 |
| MSNTC-21-1 | Chicken | Escherichia coli | C600 | 4 | 8 | >128 | >128 | >128 | >128 | 8 | ≤0.25 |
| MSNTC-21-2 | Chicken | Escherichia coli | C600 | 4 | 8 | >128 | >128 | >128 | >128 | 4 | ≤0.25 |
| MSNTC-29-1 | Chicken | Escherichia coli | C600 | 64 | 128 | >128 | >128 | 4 | 1 | 16 | ≤0.25 |
| MSY-2-1 | Duck | Escherichia coli | - | 64 | 64 | >128 | >128 | 8 | 32 | 64 | ≤0.25 |
| MSY-2-2 | Duck | Escherichia coli | - | 64 | 128 | >128 | >128 | >128 | 128 | 0.5 | ≤0.25 |
| MSY-3-1 | Duck | Escherichia coli | - | 64 | 64 | >128 | >128 | 16 | 32 | 32 | ≤0.25 |
| MSY-3-2 | Duck | Escherichia coli | C600 | 8 | 16 | >128 | >128 | >128 | 128 | ≤0.25 | ≤0.25 |
| MSY-4-1 | Duck | Escherichia coli | - | 32 | 64 | >128 | >128 | 8 | 32 | 64 | ≤0.25 |
| MSY-4-2 | Duck | Escherichia coli | - | ≤0.25 | 8 | >128 | >128 | >128 | 16 | ≤0.25 | ≤0.25 |
| MSY-6-2 | Duck | Escherichia coli | - | 2 | 8 | >128 | >128 | >128 | 64 | ≤0.25 | ≤0.25 |
| MSY-7-1 | Duck | Escherichia coli | - | ≤0.25 | 1 | >128 | >128 | >128 | 8 | ≤0.25 | ≤0.25 |
| MSY-8-1 | Duck | Escherichia coli | C600 | 32 | 128 | >128 | >128 | >128 | 64 | ≤0.25 | ≤0.25 |
| MSY-9-2 | Duck | Escherichia coli | C600 | 64 | 64 | >128 | >128 | 128 | 64 | 1 | >128 |
| MSY-10-1 | Duck | Escherichia coli | - | 32 | 128 | >128 | >128 | 4 | 1 | 16 | ≤0.25 |
| MSY-10-2 | Duck | Escherichia coli | C600 | 16 | 32 | >128 | >128 | >128 | 64 | ≤0.25 | ≤0.25 |
| MSY-11-1 | Duck | Escherichia coli | C600 | 32 | 32 | >128 | >128 | >128 | 16 | 4 | ≤0.25 |
| MSY-13-1 | Duck | Escherichia coli | C600 | 128 | 128 | >128 | >128 | >128 | 64 | 8 | ≤0.25 |
| MSY-13-2 | Duck | Escherichia coli | C600 | 2 | 8 | >128 | >128 | >128 | 64 | ≤0.25 | ≤0.25 |
| MSY-14-1 | Duck | Escherichia coli | - | 32 | 64 | >128 | >128 | 16 | 32 | 64 | ≤0.25 |
| MSY-14-2 | Duck | Escherichia coli | - | 128 | 128 | >128 | >128 | >128 | >128 | 1 | >128 |
| MSY-15-1 | Duck | Escherichia coli | C600 | 128 | 128 | >128 | >128 | >128 | 128 | ≤0.25 | ≤0.25 |
| MSG-2-1 | Pigeon | Escherichia coli | - | 64 | 64 | >128 | >128 | >128 | 1 | 8 | ≤0.25 |
| MSG-4-1 | Pigeon | Escherichia coli | - | 32 | 128 | >128 | >128 | >128 | >128 | 64 | ≤0.25 |
| MSG-5-1 | Pigeon | Escherichia coli | C600 | 128 | 128 | >128 | >128 | >128 | 64 | 64 | ≤0.25 |
| MSE-1-1 | Goose | Escherichia coli | - | 1 | 8 | >128 | >128 | >128 | 16 | ≤0.25 | ≤0.25 |
| MSE-2-1 | Goose | Escherichia coli | - | 64 | 128 | >128 | >128 | >128 | 16 | 2 | ≤0.25 |
| MSW-2-1 | Water | Escherichia coli | C600 | 16 | 8 | >128 | >128 | >128 | >128 | 8 | 2 |
| MSW-4-1 | Water | Klebsiella pneumoniae | - | 32 | 16 | >128 | >128 | >128 | 0.5 | 0.5 | ≤0.25 |
| MSW-4-2 | Water | Escherichia coli | - | 16 | 16 | >128 | >128 | 8 | 2 | 0.5 | ≤0.25 |
| MSW-5-1 | Water | Escherichia coli | C600 | 16 | 8 | >128 | >128 | 8 | 1 | ≤0.25 | ≤0.25 |
| MSW-5-2 | Water | Escherichia coli | - | 8 | 4 | >128 | >128 | >128 | 32 | 16 | ≤0.25 |
| MSW-6-1 | Water | Escherichia coli | C600 | 64 | 8 | >128 | >128 | >128 | 128 | ≤0.25 | ≤0.25 |
| MSW-7-1 | Water | Escherichia coli | C600 | 64 | 8 | >128 | >128 | >128 | 32 | 128 | 4 |
| MSW-7-2 | Water | Klebsiella pneumoniae | - | 32 | 16 | >128 | >128 | >128 | 64 | 0.5 | ≤0.25 |
| MSW-9-1 | Water | Escherichia coli | C600 | 32 | 8 | >128 | >128 | >128 | 16 | 1 | ≤0.25 |
| MSW-11-1 | Water | Escherichia coli | - | 32 | 16 | >128 | >128 | >128 | 2 | ≤0.25 | ≤0.25 |
| MSW-11-2 | Water | Escherichia coli | C600 | 8 | 16 | >128 | >128 | >128 | 128 | 0.5 | ≤0.25 |
| MSW-12-1 | Water | Escherichia coli | C600 | 32 | 16 | >128 | >128 | 8 | 2 | 8 | ≤0.25 |
| MSW-12-2 | Water | Enterobacter cloacae | C600 | 32 | 16 | >128 | >128 | 4 | 1 | ≤0.25 | ≤0.25 |
| MSW-13-1 | Water | Enterobacter cloacae | C600 | >128 | >128 | >128 | >128 | 1 | 0.5 | ≤0.25 | ≤0.25 |
| MSW-13-2 | Water | Escherichia coli | C600 | 64 | 16 | >128 | >128 | 4 | 0.5 | 16 | ≤0.25 |
| MSW-14-1 | Water | Escherichia coli | - | 32 | 8 | >128 | >128 | 8 | ≤0.25 | 8 | ≤0.25 |
| MSW-15-1 | Water | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | >128 | 128 | ≤0.25 |
| MSH-11-1 | Environmental | Escherichia coli | C600 | >128 | >128 | >128 | >128 | >128 | 128 | 1 | ≤0.25 |
| MSH-14-1 | Environmental | Enterobacter cloacae | - | >128 | >128 | >128 | >128 | >128 | >128 | 128 | >128 |
| MSH-17-1 | Environmental | Escherichia coli | C600 | >128 | >128 | >128 | >128 | 64 | 128 | 64 | ≤0.25 |
| MSH-19-1 | Environmental | Klebsiella pneumoniae | - | >128 | >128 | >128 | >128 | >128 | >128 | 8 | ≤0.25 |
| MSH-19-2 | Environmental | Escherichia coli | - | 64 | 16 | >128 | >128 | >128 | 8 | 1 | ≤0.25 |
| MSH-21-1 | Environmental | Escherichia coli | C600 | 8 | 4 | >128 | >128 | >128 | 16 | 2 | ≤0.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



